
Alterations of ADAMTSs and TIMP-3 in human nucleus pulposus cells subjected to compressive load:...
7
Alterations of ADAMTSs and TIMP-3 in Human Nucleus Pulposus Cells Subjected to Compressive Load: Implications in the Pathogenesis of Human Intervertebral Disc Degeneration Ming Huang, 1 Hai-Qiang Wang, 1 Qiang Zhang, 2 Xiao-Dong Yan, 1 Meng Hao, 1 Zhuo-Jing Luo 1 1 Institute of Orthopaedics, Xijing Hospital, Fourth Military Medical University, 15 Changle Western Road, Xi’an 710032, P.R. China, 2 Department of Orthopaedics, General Hospital of the People’s Liberation Army, Beijing, 100853, P.R. China Received 9 February 2010; accepted 30 June 2011 Published online 1 August 2011 in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/jor.21507 ABSTRACT: Intervertebral disc degeneration (IDD) pertains to the loss of extracellular matrix (ECM), particularly the early loss of aggrecan, the turnover of which is regulated by ADAMTSs. Amongst the etiological factors of IDD, mechanical stress plays an impor- tant role in the physiological and pathological processes of nucleus pulposus (NP) cells. However, the role of ADAMTSs and their inhibitor in human NP cells under mechanical stress has not been elucidated to date. The purpose of this study was to investigate the role of ADAMTSs and TIMP-3 in NP cells under mechanical stress. Human NP cells isolated from non-degenerative and degenerative discs were subjected to dynamic compressive load. The expression of ADAMTSs, aggrecan, and TIMP-3 was detected by quantitative real-time PCR and/or Western blot. Consequently, the gene expression of ADAMTS-1, 4, and 5 increased significantly in loaded NP cells compared with not-loaded cells from either non-degenerative or degenerative discs, whereas the gene expression of aggrecan decreased significantly. Moreover, Western blot indicated increased protein levels of ADAMTSs-1, 4, and 5. However, the expression of TIMP-3 altered insignificantly. Together, this study is the first addressing the underlying mechanisms of compressive load as a contributing factor to IDD in terms of ADAMTSs. Our results suggest that compressive load leads to the increase in ADAMTS-1, 4, and 5 that contributes to the decrease of aggrecan and IDD via TIMP-3 independent machinery. ß 2011 Orthopaedic Research Society. Published by Wiley Periodicals, Inc. J Orthop Res 30:267–273, 2012 Keywords: disc degeneration; nucleus pulposus; ADAMTS; aggrecan; compressive load Low back pain is a common disorder which is clinically and experimentally concerned by much. The etiology of low back pain remains largely unclear, amongst which is intervertebral disc degeneration (IDD). Therefore, many treatment strategies aimed at reliev- ing back pain have been directed towards the degenerative intervertebral disc (IVD). However, little evidence was available on the processes of degenera- tion in the literature. 1–4 Current available evidence implicates the loss of IVD extracellular matrix (ECM) upon IDD as a major cause of low back pain. 5 During degeneration, the matrix of IVD undergoes structural, mechanical and molecular changes, which result in a loss of demarca- tion between the outer annulus fibrosus and the inner nucleus pulposus (NP) tissues. Additionally, altera- tions in collagen type and a decrease in proteoglycan content lead to the loss of tissue integrity, decreased hydration, and inability to withstand load. Moreover, the loss of proteoglycans, predominantly aggrecan, is considered as an early indicator of IDD. 6 Aggrecan monomers consist of a 250 kDa protein core with chondroitin sulfate and keratan sulfate gly- cosaminoglycan (GAG) side chains attached. 6 These GAG side chains are polyanionic due to the high con- tent of carboxyl and sulfate groups, and thus attract and bind water molecules, therefore hydrate the tissue. 7 Degradation of the proteoglycans, especially aggrecan, results in dehydration of the disc, which leads to a reduced ability to resist compressive load. 8 It is well established that the biologic hallmarks of human adults NP cells are similar to chondrocytes. 9 In the cartilage, two classes of enzymes may be involved in the breakdown of aggrecan. The first class is the matrix metalloproteinases (MMPs). The second class consists of a group of proteases, ADAMTSs (a disinte- grin and metalloproteinase with thrombospondin motifs) 8,10 which are newly defined multidomain enzymes. Aggrecan fragments in inflammatory osteo- arthritis (OA) synovial fluid appear to be generated by ADAMTSs. In IDD there is a net increase in matrix degrading proteinase activity compared with protein- ase inhibitors, which disrupts the normal balance and leads to breakdown of ECM. 8,11 Although MMPs are thought to play a role in this process, 12 it has been suggested that aggrecanases may also be involved. The ADAMTSs incorporate 19 genes in humans, form- ing a branch of the M12B subfamily of metalloendo- peptidases. 13,14 ADAMTS-1, 4, 5, 8, and 15 are capable of cleaving aggrecan at the unique aggrecanase cleav- age site in IGD (arginine–glycine–aspartic acid). 15–17 Although TIMPs are, with a few exceptions, broadly effective inhibitors of MMPs, it is clear that they dis- play much greater selectivity towards both the ADAMs and ADAMTSs. ADAMTS-4 and 5 are both potently inhibited by TIMP-3, with K i values in the subnano- molar range, although they are essentially insensitive to TIMP-1, -2, and -4. 18,19 Other ADAMTSs may have a different inhibition profile by TIMPs: ADAMTS-1 is partially inhibited by TIMP-3 at 500 nM, whereas TIMP-1 and -4 have no inhibitory effect at the same concentration. 20 TIMP-3 is thus likely to be the major Ming Huang, Hai-Qiang Wang, Qiang Zhang, and Xiao-Dong Yan contributed equally to this work. Correspondence to: Zhuo-Jing Luo (T: 86-29-84775285; F: 86-29- 84775285; E-mail: [email protected]). ß 2011 Orthopaedic Research Society. Published by Wiley Periodicals, Inc. JOURNAL OF ORTHOPAEDIC RESEARCH FEBRUARY 2012 267
-
Upload
ming-huang -
Category
Documents
-
view
213 -
download
1
Transcript of Alterations of ADAMTSs and TIMP-3 in human nucleus pulposus cells subjected to compressive load:...

Alterations of ADAMTSs and TIMP-3 in Human Nucleus Pulposus CellsSubjected to Compressive Load: Implications in the Pathogenesis ofHuman Intervertebral Disc Degeneration
Ming Huang,1 Hai-Qiang Wang,1 Qiang Zhang,2 Xiao-Dong Yan,1 Meng Hao,1 Zhuo-Jing Luo1
1Institute of Orthopaedics, Xijing Hospital, Fourth Military Medical University, 15 Changle Western Road, Xi’an 710032, P.R. China, 2Departmentof Orthopaedics, General Hospital of the People’s Liberation Army, Beijing, 100853, P.R. China
Received 9 February 2010; accepted 30 June 2011
Published online 1 August 2011 in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/jor.21507
ABSTRACT: Intervertebral disc degeneration (IDD) pertains to the loss of extracellular matrix (ECM), particularly the early loss ofaggrecan, the turnover of which is regulated by ADAMTSs. Amongst the etiological factors of IDD, mechanical stress plays an impor-tant role in the physiological and pathological processes of nucleus pulposus (NP) cells. However, the role of ADAMTSs and theirinhibitor in human NP cells under mechanical stress has not been elucidated to date. The purpose of this study was to investigate therole of ADAMTSs and TIMP-3 in NP cells under mechanical stress. Human NP cells isolated from non-degenerative and degenerativediscs were subjected to dynamic compressive load. The expression of ADAMTSs, aggrecan, and TIMP-3 was detected by quantitativereal-time PCR and/or Western blot. Consequently, the gene expression of ADAMTS-1, 4, and 5 increased significantly in loaded NP cellscompared with not-loaded cells from either non-degenerative or degenerative discs, whereas the gene expression of aggrecan decreasedsignificantly. Moreover, Western blot indicated increased protein levels of ADAMTSs-1, 4, and 5. However, the expression of TIMP-3altered insignificantly. Together, this study is the first addressing the underlying mechanisms of compressive load as a contributingfactor to IDD in terms of ADAMTSs. Our results suggest that compressive load leads to the increase in ADAMTS-1, 4, and 5 thatcontributes to the decrease of aggrecan and IDD via TIMP-3 independent machinery. � 2011 Orthopaedic Research Society. Publishedby Wiley Periodicals, Inc. J Orthop Res 30:267–273, 2012
Keywords: disc degeneration; nucleus pulposus; ADAMTS; aggrecan; compressive load
Low back pain is a common disorder which is clinicallyand experimentally concerned by much. The etiologyof low back pain remains largely unclear, amongstwhich is intervertebral disc degeneration (IDD).Therefore, many treatment strategies aimed at reliev-ing back pain have been directed towards thedegenerative intervertebral disc (IVD). However, littleevidence was available on the processes of degenera-tion in the literature.1–4
Current available evidence implicates the loss ofIVD extracellular matrix (ECM) upon IDD as a majorcause of low back pain.5 During degeneration, thematrix of IVD undergoes structural, mechanical andmolecular changes, which result in a loss of demarca-tion between the outer annulus fibrosus and the innernucleus pulposus (NP) tissues. Additionally, altera-tions in collagen type and a decrease in proteoglycancontent lead to the loss of tissue integrity, decreasedhydration, and inability to withstand load. Moreover,the loss of proteoglycans, predominantly aggrecan, isconsidered as an early indicator of IDD.6
Aggrecan monomers consist of a 250 kDa proteincore with chondroitin sulfate and keratan sulfate gly-cosaminoglycan (GAG) side chains attached.6 TheseGAG side chains are polyanionic due to the high con-tent of carboxyl and sulfate groups, and thus attractand bind water molecules, therefore hydrate thetissue.7 Degradation of the proteoglycans, especially
aggrecan, results in dehydration of the disc, whichleads to a reduced ability to resist compressive load.8
It is well established that the biologic hallmarks ofhuman adults NP cells are similar to chondrocytes.9 Inthe cartilage, two classes of enzymes may be involvedin the breakdown of aggrecan. The first class is thematrix metalloproteinases (MMPs). The second classconsists of a group of proteases, ADAMTSs (a disinte-grin and metalloproteinase with thrombospondinmotifs)8,10 which are newly defined multidomainenzymes. Aggrecan fragments in inflammatory osteo-arthritis (OA) synovial fluid appear to be generated byADAMTSs. In IDD there is a net increase in matrixdegrading proteinase activity compared with protein-ase inhibitors, which disrupts the normal balance andleads to breakdown of ECM.8,11 Although MMPs arethought to play a role in this process,12 it has beensuggested that aggrecanases may also be involved.The ADAMTSs incorporate 19 genes in humans, form-ing a branch of the M12B subfamily of metalloendo-peptidases.13,14 ADAMTS-1, 4, 5, 8, and 15 are capableof cleaving aggrecan at the unique aggrecanase cleav-age site in IGD (arginine–glycine–aspartic acid).15–17
Although TIMPs are, with a few exceptions, broadlyeffective inhibitors of MMPs, it is clear that they dis-play much greater selectivity towards both the ADAMsand ADAMTSs. ADAMTS-4 and 5 are both potentlyinhibited by TIMP-3, with Ki values in the subnano-molar range, although they are essentially insensitiveto TIMP-1, -2, and -4.18,19 Other ADAMTSs may havea different inhibition profile by TIMPs: ADAMTS-1 ispartially inhibited by TIMP-3 at 500 nM, whereasTIMP-1 and -4 have no inhibitory effect at the sameconcentration.20 TIMP-3 is thus likely to be the major
Ming Huang, Hai-Qiang Wang, Qiang Zhang, and Xiao-DongYan contributed equally to this work.Correspondence to: Zhuo-Jing Luo (T: 86-29-84775285; F: 86-29-84775285; E-mail: [email protected]).
� 2011 Orthopaedic Research Society. Published by Wiley Periodicals, Inc.
JOURNAL OF ORTHOPAEDIC RESEARCH FEBRUARY 2012 267

natural inhibitor of aggrecanase activity in IVD. How-ever, literature addressing the impact of mechanicalforce on TIMP-3 is sparse. Pockert et al.8 found thatthe gene expression of TIMP-3 had altered insignif-icantly in human degenerative IVDs compared withnon-degenerative samples.
Hitherto, some studies have investigated the ex-pression or activity of ADAMTS in IVD and the poten-tial roles in matrix degradation. Hatano et al.21 notedthe expression of ADAMTS-4 messenger RNA (mRNA)and protein in herniated discs. Le Maitre et al.22
demonstrated that native disc cells expressedADAMTS-4, which increased during IDD. However,few studies have addressed the role of ADAMTSs inNP cells under mechanical stress.
To address these issues, we collected human NPcells from non-degenerative and degenerative discs,which were subjected to dynamic compressive load.Furthermore, we detected the expression of ADAMTSsand TIMP-3 by real-time PCR and Western blot.
MATERIALS AND METHODSTissue CollectionThe study was approved by our institutional ethics reviewboard (20090611-3) and human NP tissues were collected aswe described previously.23 Briefly, written informed consentsfrom each patient were obtained. Human NP specimens werecollected from patients with idiopathic scoliosis and IDD ascontrol [n ¼ 15; average age 18.7 (range 16–24) years] anddegenerative [n ¼ 15; average age 30.2 (range 27–38) years]NP samples. Disc specimens were classified as grade II (idio-pathic scoliosis discs) and grade IV (IDD discs) according toMRI.24
Tissue CulturesNP tissues were cut into fragments with 1 mm in diameter,placed into individual compressive wells in 3 ml of culturemedium including Dulbecco’s modified Eagle’s medium(DMEM)-Ham’s F-12, 10% fetal bovine serum, 100 U/ml pen-icillin, and 100 mg/ml streptomycin.
Compressive LoadEach specimen was allowed to equilibrate for 1 h andsubjected to a compressive stress in BioPress compressionculture plates using Flexercell Compression System25
(FX-4000C; Flexercell International, Hillsborough, NC). Thecompressive stress was 0.35–0.95 MPa at 1 Hz for 2 h twice
a day for 7 days. The Flexercell FX-4000C functions via theapplication of compressed gas to the base of the BioPresscompression plates with flexible membranes, resulting incompression of the sample (contained within the foam circle)against the fixed platen at controlled rates of pressure(the load is transferred through a fluid media. Therefore, themechanical load confronting the NP tissues within the com-pressive well is actually 0.35–0.95 MPa).25 The specimenswithout compression load (control group) were cultured inBioPress compression culture plates at a similar condition.Seven days later, tissues were released by 0.25% type IIcollagenase digestion at 378C for 1 h and re-suspended incomplete medium. Cell numbers were approximately1 � 106 cells/ml. Then cells were collected for analysis of cellviability, aggrecan and ADAMTS gene expression andWestern blot.
Cell ViabilityThe assessment of cell viability was carried out as describedpreviously.2 Briefly, carboxyfluorescein diacetate succini-midyl ester (CFSE-DA) and propidium iodide (PI) stainingwere used, where CFSE-DA stains viable cells green, and PIstains nonviable cells red. Cells were visualized with aninverted microscope equipped with a dual-wavelength bypassfilter (450 nm for CFSE-DA/520 nm for PI). Emission ofgreen fluorescence and red fluorescence indicated viable cellsand nonviable cells, respectively. Total cell numbers werecounted manually and tallied as viable and nonviable cells toallow for calculation of the percentage of viability.
RNA Extraction and Real-Time Quantitative PCR for Aggrecanand ADAMTSs Gene ExpressionRNA was extracted using TRIzol reagent (Gibco) accordingto the manufacturer’s instructions. Quantitative real-timePCR was used to investigate the effect of dynamic com-pressive load on aggrecan, ADAMTSs, and TIMP-3 geneexpression. The total gene specificity of the nucleotidesequences chosen for the primers and probes was confirmedby BLAST searches (GenBank database sequences) as listedin Table 1. Primers and probes were purchased from AppliedBiosystems, Warrington, UK. For each set of primers andprobes, the efficiency of the amplification was assessed usingtemplate titrations as recommended by Applied Biosystems.Data were normalized to GAPDH and then analyzed usingthe 2�DDCT method.
TIMP-3 Treatment as Inhibitory FactorOne hour prior to load, NP samples were treated with orwithout 500 nM TIMP-3. Subsequently, aggrecan gene
Table 1. Probes and Primers for Real-Time PCR
Target Forward Primer, 50–30 Probe, 50–30 Reverse Primer, 50–30
GAPDH PDAR PDAR PDAR
Aggrecan TCGAGGACAGCGGCC ATGGAACACGATGCCTTTCACCACGA TCGAGGGTGTAGCGTGTAGAGA
ADAMTS-1 GGACAGGTGCAAGCTCATCTG CAAGCCAAAGGCATTGGCTACTTCTTCG TCTACAACCTTGGGCTGCAAA
ADAMTS-4 ACTGGTGGTGGCAGATGACA ATGGCCGCATTCCACGGTG TCACTGTTAGCAGGTAGCGCTTT
ADAMTS-5 GGACCTACCACGAAAGCAGATC CCCAGGACAGACCTACGATGCCACC GCCGGGACACACGGAGTAC
ADAMTS-8 GAGGTGGAGACGGGAGAG CTTGGCTCTCCTCCTCGCTGTCCTCC TAGCGCCTTCTGCCTCCT
ADAMTS-9 GCATTAACTCTGCCACTGACC CTTCGCCTCCTCCTCTTCCTCCTCTACCT ATAGAAACTGCTGGCCGAAGG
ADAMTS-15 ATGTGCTGGCACCCAAGGT CCTGACTCCACCTCCGTCTGTGTCCA CAGCCAGCCTTGATGCACTT
TIMP-3 GCAGATAGACTCAAGGTGTGTGAAA CCACTGCATGTCCCAACCAGACTGTGT TCCCTCACTCTTACATGCAGACA
268 HUANG ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH FEBRUARY 2012

expression was detected using real-time PCR as aforemen-tioned. All experiments were performed in triplicate.
Statistical AnalysisData were determined to be nonparametric using theShapiro–Wilke test. The Kruskal–Wallis test with Mann–Whitney U post hoc tests were used to assess the effect ofdynamic compressive load on cell viability and the expressionof aggrecan, ADAMTSs, and TIMP-3 on non-degenerativeand degenerative samples. ANOVA with Tukey’s post hocmultiple comparisons were applied for comparison of the ratiosamong groups for Western blot analysis. Difference with ap-value <0.05 was considered as significant.
RESULTS
Mechanical Stress Has an Impact on Cell Viability andAggrecan Gene Expression of NP CellsThe viability of NP cells was over 85%, with no differ-ence observed between samples subjected to dynamiccompressive loaded and not-loaded controls which de-rived from non-degenerative IVDs. A small decrease incell viability was noted in loaded NP cells derived fromdegenerative IVDs compared with not-loaded NP cells(p < 0.05) (Fig. 1). Application of compressive load at0.35–0.95 MPa and a frequency of 1 Hz on NP cellsderived from non-degenerative or degenerative IVDsled to significantly decreased aggrecan gene expression(p < 0.05) (Fig. 2).
Gene Expression of ADAMTSs and TIMP-3 Altered WithVarious Manners Under Compressive Load in HumanNP CellsADAMTS-1, 4, 5, 9, and 15 mRNA and TIMP-3 mRNAwere expressed in both not-loaded and loaded NP cells.
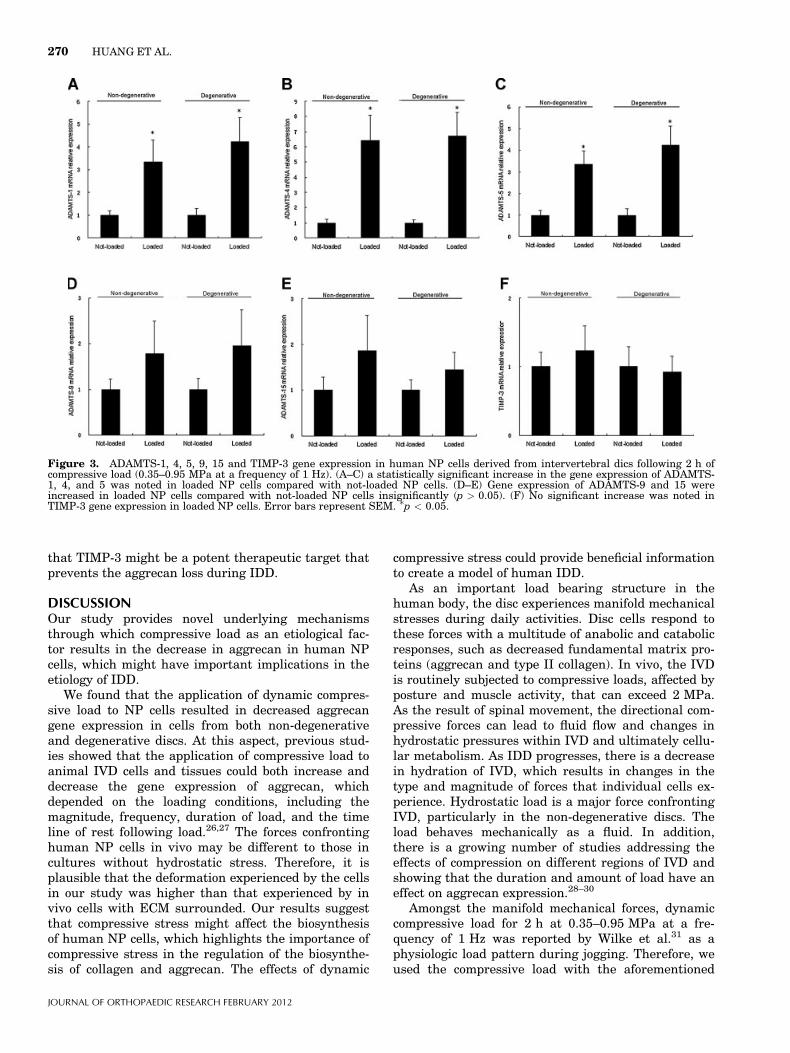
However, ADAMTS-8 gene expression was notdetected in NP tissues investigated. When we com-pared the expression levels of the target genes, astatistically significant increase in the gene expressionof ADAMTS-1, 4, and 5 was noted in loaded NP cellscompared with not-loaded NP cells derived from eithernon-degenerative or degenerative discs (Fig. 3A–C)(p < 0.05). Gene expression of ADAMTS-9 and 15 wereincreased in loaded NP cells compared with not-loadedNP cells insignificantly (Fig. 3D–E) (p > 0.05). Nosignificant increase was noted in TIMP-3 gene expres-sion in loaded NP cells (Fig. 3F).
ADAMTSs Increase Significantly and TIMP-3 AltersInsignificantly at Protein LevelADAMTS-1, 4, 5 and TIMP-3 were subjected to West-ern blot analysis to further confirm aforementionedgene expression. A statistically significant increase inthe protein levels of ADAMTS-1, 4, and 5 was noted inloaded NP cells compared with not-loaded NP cells(Fig. 4A–C). As expected, the protein level of TIMP-3altered insignificantly (Fig. 4D).
TIMP-3 Is Potent Therapeutic Target Aiming to Prevent theAggrecan Loss During IDDGiven that ADAMTS-1, 4, and 5 are highly expressedin loaded NP samples, we further investigated the roleof TIMP-3 in IDD. As shown in Figure 5, treatment ofTIMP-3 resulted in significantly increased aggrecangene expression in loaded NP cells (p < 0.05). A combi-nation of TIMP-3 treatment and load led to insignifi-cant decreased aggrecan expression compared withnot-loaded NP cells (p > 0.05). These results indicate
Figure 1. Compressive load has an impact on the percentageof viable cells following 2 h of compressive load with 0.35–0.95 MPa at a frequency of 1 Hz. A small decrease in cell viabili-ty was noted in loaded NP cells derived from degenerative IVDscompared with not-loaded NP cells. Error bars represent SEM.�p < 0.05.
Figure 2. Aggrecan gene expression in human NP cells de-rived from intervertebral dics following 2 h of compressive load(0.35–0.95 MPa at a frequency of 1 Hz). Significantly decreasedaggrecan gene expression was noted in loaded NP cells. Errorbars represent SEM. �p < 0.05.
ALTERATIONS OF ADAMTSS AND TIMP-3 269
JOURNAL OF ORTHOPAEDIC RESEARCH FEBRUARY 2012
that TIMP-3 might be a potent therapeutic target thatprevents the aggrecan loss during IDD.
DISCUSSIONOur study provides novel underlying mechanismsthrough which compressive load as an etiological fac-tor results in the decrease in aggrecan in human NPcells, which might have important implications in theetiology of IDD.
We found that the application of dynamic compres-sive load to NP cells resulted in decreased aggrecangene expression in cells from both non-degenerativeand degenerative discs. At this aspect, previous stud-ies showed that the application of compressive load toanimal IVD cells and tissues could both increase anddecrease the gene expression of aggrecan, whichdepended on the loading conditions, including themagnitude, frequency, duration of load, and the timeline of rest following load.26,27 The forces confrontinghuman NP cells in vivo may be different to those incultures without hydrostatic stress. Therefore, it isplausible that the deformation experienced by the cellsin our study was higher than that experienced by invivo cells with ECM surrounded. Our results suggestthat compressive stress might affect the biosynthesisof human NP cells, which highlights the importance ofcompressive stress in the regulation of the biosynthe-sis of collagen and aggrecan. The effects of dynamic
compressive stress could provide beneficial informationto create a model of human IDD.
As an important load bearing structure in thehuman body, the disc experiences manifold mechanicalstresses during daily activities. Disc cells respond tothese forces with a multitude of anabolic and catabolicresponses, such as decreased fundamental matrix pro-teins (aggrecan and type II collagen). In vivo, the IVDis routinely subjected to compressive loads, affected byposture and muscle activity, that can exceed 2 MPa.As the result of spinal movement, the directional com-pressive forces can lead to fluid flow and changes inhydrostatic pressures within IVD and ultimately cellu-lar metabolism. As IDD progresses, there is a decreasein hydration of IVD, which results in changes in thetype and magnitude of forces that individual cells ex-perience. Hydrostatic load is a major force confrontingIVD, particularly in the non-degenerative discs. Theload behaves mechanically as a fluid. In addition,there is a growing number of studies addressing theeffects of compression on different regions of IVD andshowing that the duration and amount of load have aneffect on aggrecan expression.28–30
Amongst the manifold mechanical forces, dynamiccompressive load for 2 h at 0.35–0.95 MPa at a fre-quency of 1 Hz was reported by Wilke et al.31 as aphysiologic load pattern during jogging. Therefore, weused the compressive load with the aforementioned
Figure 3. ADAMTS-1, 4, 5, 9, 15 and TIMP-3 gene expression in human NP cells derived from intervertebral dics following 2 h ofcompressive load (0.35–0.95 MPa at a frequency of 1 Hz). (A–C) a statistically significant increase in the gene expression of ADAMTS-1, 4, and 5 was noted in loaded NP cells compared with not-loaded NP cells. (D–E) Gene expression of ADAMTS-9 and 15 wereincreased in loaded NP cells compared with not-loaded NP cells insignificantly (p > 0.05). (F) No significant increase was noted inTIMP-3 gene expression in loaded NP cells. Error bars represent SEM. �p < 0.05.
270 HUANG ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH FEBRUARY 2012

parameters to address the biological effects of suchphysiologic load pattern.
Some studies have demonstrated the importance ofboth ADAMTS-4 and 5 in the breakdown of articularcartilage in arthritis diseases.32,33 A number ofADAMTSs expressions have been demonstrated inherniated discs with increased expression of the sameenzyme in degenerative discs.8,14–16,21,32,34 However,no studies to date have addressed the role ofADAMTSs in IVD cells during mechanical stimulation.Our results demonstrated that ADAMTS-1, 4, 9, 15and TIMP-3 mRNA and protein were present in bothnot-loaded and loaded NP cells derived from both non-degenerative and degenerative human discs. Theexpressions in non-degenerative discs indicate a
possible role for the ADAMTSs in the normal turnover of aggrecan and other matrix molecules in thehealthy disc matrix. Consistent with the findings inarticular cartilage,15 ADAMTS-8 was the only aggreca-nolytic ADAMTS that was not detected in our study.Demircan et al.35 noted that ADAMTS-8 was undetect-able in the chondrosarcoma cell line and human chon-drocytes, even when the cells were treated withinterleukin-1 or tumor necrosis factor (TNF). Anotherplausible factor might be that ADAMTS-8 is the soleweakly expressed aggrecanolytic one among theADAMTSs. It is noteworthy that a statistically signifi-cant increase in the gene expression of ADAMTS-1, 4,and 5 was noted in loaded NP cells compared with not-loaded NP cells with an increase at the protein level.
Figure 4. ADAMTS-1, 4, 5 and TIMP-3 protein expression in human NP cells derived from intervertebral dics following 2 h of com-pressive load (0.35–0.95 MPa at a frequency of 1 Hz). (A–C) A statistically significant increase in the protein levels of ADAMTS-1, 4,and 5 was noted in loaded NP cells compared with not-loaded NP cells. (D) The protein level of TIMP-3 altered insignificantly. Errorbars represent SEM. �p < 0.05.
ALTERATIONS OF ADAMTSS AND TIMP-3 271
JOURNAL OF ORTHOPAEDIC RESEARCH FEBRUARY 2012

These results suggest that ADAMTS-1, 4, and 5 mayplay a more dominant role in IDD among allADAMTSs.
TIMP-3 can inhibit a multitude of matrix protei-nases, such as MMPs and ADAMTSs36,37 by bindingnoncovalently to the active sites of the target enzymesin a 1:1 stoichiometry.37 It can also be sequestered intothe matrix via interaction with GAGs. The level ofTIMP-3 in healthy tissues may maintain the homeo-static balance of matrix turnover by inhibiting a widerange of catabolic enzymes. However, it remainsunknown whether the static level of TIMP-3 in thedegenerative state would be able to cope with thesubstantial increases in ADAMTSs, also, the knownincrease of MMPs noted in IDD.22,38 There is accumu-lating evidence demonstrating that in degraded carti-lage, aggrecan is primarily cleaved by aggrecanasesand later by MMPs. This theory is supported byMercuri et al.39 that aggrecanase-generated G1fragments are substrates for MMPs, but the MMPgenerated 342FFGVG fragments are resistant to aggre-canases’ degradation. Pockert et al.8 found that therewas no significant change in the numbers of cells stainpositive for TIMP-3 in the different grades of degener-ation in NP samples, this may explain why TIMP-3does not have the corresponding increase, the loadcondition cannot activated the mechanotransductionpathways which regulate TIMP-3 will be anotherreason. The imbalance between ADAMTSs and theirinhibitor (TIMP-3) would play a role in the pathogene-sis of IDD and therefore be a potential therapeutictarget for preventing IDD.
In addition to the increased mRNA and proteinlevel in ADAMTS-1, 4, and 5, we demonstrate thatTIMP-3 clearly increase the expression of aggrecanthe early marker gene for IDD, in other words, TIMP-3 have effective inhibitory effect on ADAMTSs orMMPs. Pockert et al.8 reported that MMPs and aggre-canases may be working simultaneously to breakdown aggrecan. TIMP-3 is thus likely to be the majorinhibitor of aggrecanase activity in IVD. TIMP-3 ispotent therapeutic target aiming to prevent the aggre-can loss during IDD.
While the results presented here do not give a defin-itive answer as to which enzyme or group of enzymesare the most important for IDD. We demonstrate thata wide range of aggrecanases (ADAMTS-1, 4, and 5)show significant increases in IDD and therefore sug-gest a role for the ADAMTSs enzymes in IVD tissuebreakdown and degradation of aggrecan. Further stud-ies are needed to establish whether MMPs or aggreca-nases play dominant roles in IDD. Our results suggestADAMTSs play an important role in the breakdown ofaggrecan in human NP during IDD. Loss of aggrecanfrom the NP is an early and reversible process indegeneration which precedes the irreversible break-down of the collagen network and eventual loss of discheight and function. Identification of a key ADAMTSin this process could implicate the development of atherapeutic target aiming to prevent the aggrecan lossduring IDD as novel treatment strategies.
ACKNOWLEDGMENTSThis study is supported by the Chinese National NaturalScience Foundation (grant number 30901509). We thankDr. Zhe Wang for technical suggestion. We are grateful forthe excellent technical assistance of our technician Li-FengLan.
REFERENCES1. Kasra M, Goel V, Martin J, et al. 2003. Effect of dynamic
hydrostatic pressure on rabbit intervertebral disc cells.J Orthop Res 21:597–603.
2. Le Maitre CL, Frain J, Millward-Sadler J, et al. 2009. Al-tered integrin mechanotransduction in human nucleus pul-posus cells derived from degenerated discs. Arthritis Rheum60:460–469.
3. Le Maitre CL, Freemont AJ, Hoyland JA. 2005. The role ofinterleukin-1 in the pathogenesis of human intervertebraldisc degeneration. Arthritis Res Ther 7:R732–R745.
4. Le Maitre CL, Hoyland JA, Freemont AJ. 2007. Interleukin-1 receptor antagonist delivered directly and by gene therapyinhibits matrix degradation in the intact degenerate humanintervertebral disc: an in situ zymographic and gene therapystudy. Arthritis Res Ther 9:R83.
5. Luoma K, Riihimaki H, Luukkonen R, et al. 2000. Low backpain in relation to lumbar disc degeneration. Spine (Phila Pa1976) 25:487–492.
6. Roughley PJ, Alini M, Antoniou J. 2002. The role of proteo-glycans in aging, degeneration and repair of the interverte-bral disc. Biochem Soc Trans 30:869–874.
7. Pratta MA, Yao W, Decicco C, et al. 2003. Aggrecan protectscartilage collagen from proteolytic cleavage. J Biol Chem278:45539–45545.
Figure 5. Aggrecan gene expression in untreated and treatedhuman NP cells derived from non-degenerative IVDs following2 h of compressive load (0.35–0.95 MPa at a frequency of 1 Hz).Treatment of TIMP-3 resulted in significantly increased aggrecangene expression in loaded NP cells (p < 0.05). A combination ofTIMP-3 treatment and load led to insignificant decreased aggre-can expression compared with not-loaded NP cells (p > 0.05). Er-ror bars represent SEM. �p < 0.05.
272 HUANG ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH FEBRUARY 2012

8. Pockert AJ, Richardson SM, Le Maitre CL, et al. 2009. Modi-fied expression of the ADAMTS enzymes and tissue inhibitorof metalloproteinases 3 during human intervertebral disc de-generation. Arthritis Rheum 60:482–491.
9. Sive JI, Baird P, Jeziorsk M, et al. 2002. Expression of chon-drocyte markers by cells of normal and degenerate interver-tebral discs. Mol Pathol 55:91–97.
10. Malfait AM, Liu RQ, Ijiri K, et al. 2002. Inhibition ofADAM-TS4 and ADAM-TS5 prevents aggrecan degradationin osteoarthritic cartilage. J Biol Chem 277:22201–22208.
11. Kanemoto M, Hukuda S, Komiya Y, et al. 1996. Immunohis-tochemical study of matrix metalloproteinase-3 and tissueinhibitor of metalloproteinase-1 human intervertebral discs.Spine (Phila Pa 1976) 21:1–8.
12. Roberts S, Caterson B, Menage J, et al. 2000. Matrix metal-loproteinases and aggrecanase: their role in disorders of thehuman intervertebral disc. Spine (Phila Pa 1976) 25:3005–3013.
13. Porter S, Clark IM, Kevorkian L, et al. 2005. The ADAMTSmetalloproteinases. Biochem J 386:15–27.
14. Kaushal GP, Shah SV. 2000. The new kids on the block:ADAMTSs, potentially multifunctional metalloproteinases ofthe ADAM family. J Clin Invest 105:1335–1337.
15. Collins-Racie LA, Flannery CR, Zeng W, et al. 2004.ADAMTS-8 exhibits aggrecanase activity and is expressed inhuman articular cartilage. Matrix Biol 23:219–230.
16. Somerville RP, Longpre JM, Jungers KA, et al. 2003. Char-acterization of ADAMTS-9 and ADAMTS-20 as a distinctADAMTS subfamily related to Caenorhabditis elegans GON-1. J Biol Chem 278:9503–9513.
17. Cal S, Obaya AJ, Llamazares M, et al. 2002. Cloning,expression analysis, and structural characterization ofseven novel human ADAMTSs, a family of metalloprotei-nases with disintegrin and thrombospondin-1 domains. Gene283:49–62.
18. Hashimoto T, Wen G, Lawton MT, et al. 2003. Abnormal ex-pression of matrix metalloproteinases and tissue inhibitorsof metalloproteinases in brain arteriovenous malformations.Stroke 34:925–931.
19. Kashiwagi M, Tortorella M, Nagase H, et al. 2001. TIMP-3is a potent inhibitor of aggrecanase 1 (ADAM-TS4) andaggrecanase 2 (ADAM-TS5). J Biol Chem 276:12501–12504.
20. Rodriguez-Manzaneque JC, Westling J, Thai SN, et al. 2002.ADAMTS1 cleaves aggrecan at multiple sites and is differen-tially inhibited by metalloproteinase inhibitors. BiochemBiophys Res Commun 293:501–508.
21. Hatano E, Fujita T, Ueda Y, et al. 2006. Expression ofADAMTS-4 (aggrecanase-1) and possible involvement in re-gression of lumbar disc herniation. Spine (Phila Pa 1976)31:1426–1432.
22. Le Maitre CL, Freemont AJ, Hoyland JA. 2004. Localizationof degradative enzymes and their inhibitors in the degener-ate human intervertebral disc. J Pathol 204:47–54.
23. Wang HQ, Yu XD, Liu ZH, et al. 2011. Deregulated miR-155promotes Fas-mediated apoptosis in human intervertebral
disc degeneration by targeting FADD and caspase-3. JPathol (in press).
24. Pfirrmann CW, Metzdorf A, Zanetti M, et al. 2001. Magneticresonance classification of lumbar intervertebral disc degen-eration. Spine (Phila Pa 1976) 26:1873–1878.
25. International F. Flexcell culture system. URL: http://www.flexcellint.com/gallery.htm.
26. Ohshima H, Urban JP, Bergel DH. 1995. Effect of static loadon matrix synthesis rates in the intervertebral discmeasured in vitro by a new perfusion technique. J OrthopRes 13:22–29.
27. Lotz JC, Chin JR. 2000. Intervertebral disc cell death is de-pendent on the magnitude and duration of spinal loading.Spine (Phila Pa 1976) 25:1477–1483.
28. MacLean JJ, Lee CR, Alini M, et al. 2005. The effects ofshort-term load duration on anabolic and catabolic gene ex-pression in the rat tail intervertebral disc. J Orthop Res23:1120–1127.
29. Korecki CL, MacLean JJ, Iatridis JC. 2008. Dynamic com-pression effects on intervertebral disc mechanics and biolo-gy. Spine (Phila Pa 1976) 33:1403–1409.
30. Walsh AJ, Lotz JC. 2004. Biological response of the interver-tebral disc to dynamic loading. J Biomech 37:329–337.
31. Wilke HJ, Neef P, Caimi M, et al. 1999. New in vivo meas-urements of pressures in the intervertebral disc in daily life.Spine (Phila Pa 1976) 24:755–762.
32. Echtermeyer F, Bertrand J, Dreier R, et al. 2009. Syndecan-4 regulates ADAMTS-5 activation and cartilage breakdownin osteoarthritis. Nat Med 15:1072–1076.
33. Song RH, Tortorella MD, Malfait AM, et al. 2007. Aggrecandegradation in human articular cartilage explants is mediat-ed by both ADAMTS-4 and ADAMTS-5. Arthritis Rheum56:575–585.
34. Rutges JP, Kummer JA, Oner FC, et al. 2008. IncreasedMMP-2 activity during intervertebral disc degeneration iscorrelated to MMP-14 levels. J Pathol 214:523–530.
35. Demircan K, Hirohata S, Nishida K, et al. 2005. ADAMTS-9is synergistically induced by interleukin-1beta and tumornecrosis factor alpha in OUMS-27 chondrosarcoma cells andin human chondrocytes. Arthritis Rheum 52:1451–1460.
36. Brew K, Dinakarpandian D, Nagase H. 2000. Tissue inhibi-tors of metalloproteinases: evolution, structure and function.Biochim Biophys Acta 1477:267–283.
37. Yu WH, Yu S, Meng Q, et al. 2000. TIMP-3 binds to sulfatedglycosaminoglycans of the extracellular matrix. J Biol Chem275:31226–31232.
38. Weiler C, Nerlich AG, Zipperer J, et al. 2002. 2002 SSEAward Competition in Basic Science: expression of majormatrix metalloproteinases is associated with intervertebraldisc degradation and resorption. Eur Spine J 11:308–320.
39. Mercuri FA, Maciewicz RA, Tart J, et al. 2000. Mutations inthe interglobular domain of aggrecan alter matrix metallo-proteinase and aggrecanase cleavage patterns. Evidencethat matrix metalloproteinase cleavage interferes withaggrecanase activity. J Biol Chem 275:33038–33045.
ALTERATIONS OF ADAMTSS AND TIMP-3 273
JOURNAL OF ORTHOPAEDIC RESEARCH FEBRUARY 2012







![r n al of S o u pi J ne Sahoo et al, Spine 216, 5:2 Journal of Spine … · of Herniated nucleus pulposus is 1-3% [2]. Intervertebral disc being aneural is a predominant site for](https://static.fdocuments.in/doc/165x107/5fcf426acb758459f013f8cd/r-n-al-of-s-o-u-pi-j-ne-sahoo-et-al-spine-216-52-journal-of-spine-of-herniated.jpg)






![r n al of S o u pi J ne Sahoo et al, Spine 216, 5:2 ... · of Herniated nucleus pulposus is 1-3% [2]. Intervertebral disc being aneural is a predominant site for low back pain [3].](https://static.fdocuments.in/doc/165x107/5f01e55e7e708231d40190e9/r-n-al-of-s-o-u-pi-j-ne-sahoo-et-al-spine-216-52-of-herniated-nucleus-pulposus.jpg)
![The protective effects of PI3K/Akt pathway on human nucleus pulposus … · 2020. 1. 28. · nucleus pulposus cells and nucleus pulposus progenitor cells [14]. Previous studies have](https://static.fdocuments.in/doc/165x107/60b265dd0d8b8040e758b496/the-protective-effects-of-pi3kakt-pathway-on-human-nucleus-pulposus-2020-1-28.jpg)


