
Age and Long-term Growth of Trees in an Old-growth … la madera de ocho especies muestreada de...
12
306 BIOTROPICA 35(3): 306–317 2003 Age and Long-term Growth of Trees in an Old-growth Tropical Rain Forest, Based on Analyses of Tree Rings and 14 C 1 Esther Fichtler Institut fu ¨ r Forstbotanik, Universitaet Goettingen, Buesgenweg 2, D-37077, Goettingen, Germany Deborah A. Clark Department of Biology, University of Missouri–St. Louis, 8001 Natural Bridge Road, St. Louis, Missouri 63121-4499, U.S.A. and Martin Worbes Institut fu ¨ r tropischen und subtropischen Pflanzenbau, Universitaet Goettingen, Grisebachstrasse 6, D-37077, Goettingen, Germany ABSTRACT In an old-growth tropical wet forest at La Selva, Costa Rica, we combined radiocarbon ( 14 C) dating and tree-ring analysis to estimate the ages of large trees of canopy and emergent species spanning a broad range of wood densities and growth rates. We collected samples from the trunks of 29 fallen, dead individuals. We found that all eight sampled species formed visible growth rings, which varied considerably in distinctiveness. For five of the six species for which we combined wood anatomical studies with 14 C-dates (ring ages), the analyses demonstrated that growth rings were of annual formation. The oldest tree we found by direct ring counting was a Hymenolobium mesoamericanum Lima (Papilionaceae) specimen, with an age of ca. 530 years at the time of death. All other sampled individuals, including very large trees of slow-growing species, had died at ages between 200 and 300 years. These results show that, even in an everwet tropical rain forest, tree growth of many species can be rhythmic, with an annual periodicity. This study thus raises the possibility of extending tree-ring analyses throughout the tropical forest types lacking a strong dry season or annual flooding. Our findings and similar measurements from other tropical forests indicate that the max- imum ages of tropical emergent trees are unlikely to be much greater than 600 years, and that these trees often die earlier from various natural causes. RESUMEN En una area de bosque muy hu ´medo tropical localizado en La Selva, Costa Rica, usamos una combinacio ´n de ana ´lisis de radiocarbono ( 14 C) y ana ´lisis de anillos de crecimiento, para estimar las edad de a ´rboles del dosel con un amplio espectro de tasas de crecimiento y de densidades de la madera. Analizamos la madera de ocho especies muestreada de troncos de 29 individuos muertos (a ´rboles caı ´dos). Encontramos que las ocho especies estudiadas forman anillos de crecimiento visibles, pero que estos varı ´an en la distincio ´n del lı ´mite de sus anillos. Para cinco de las seis especies para las cuales combinamos un estudio anato ´mico de la madera con fechado de 14 C (edades de los anillos), nuestro ana ´lisis demostro ´ que los anillos de crecimiento fueron anuales. El a ´rbol ma ´s viejo en nuestra muestra fue un ejemplar de Hymenolobium mesoamericanum Lima (Papilionaceae), que murio ´ a la edad de aproximadamente 530 an ˜os. Las edades a las que murieron los dema ´s indivı ´duos analizados, incluyendo a ´rboles de gran taman ˜o y de lento crecimiento, fueron entre 200 y 300 an ˜os. Estos resultados demuestran que, au ´n en un bosque tropical muy hu ´medo, el crecimiento de muchas especies de a ´rboles puede presentar un crecimiento annual rı ´tmico. Este estudio demuestra que es posible extender el ana ´lisis de anillos de crecimiento por todo el tro ´pico, incluyendo las zonas de bosque que no experimentan ni una estacio ´n seca fuerte ni inundacio ´n anual. Nuestros resultados, y los de otros estudios en bosques tropicales, indican que es poco probable que las edades ma ´ximas de a ´rboles tropicales emergentes lleguen a ser mucho mayores a los 600 an ˜os, y que tales a ´rboles frecuentemente mueran a edades menores por causas naturales. Key words: emergent trees; La Selva; Costa Rica; old canopy; radiocarbon dating; tree ring analysis; tropical wet forest. UNDERSTANDING THE PAST AND CURRENT DYNAMICS OF THE WORLD’S TROPICAL rain forests is a pressing challenge for ecologists. Tropical evergreen forests are disproportionately important in the global car- bon cycle. They have been estimated to account 1 Received 28 February 2003; revision accepted 3 July 2003. for 32 percent of terrestrial net primary productiv- ity (Field et al. 1998), and combined with tropical dry forests, they comprise an estimated 59 percent of forest biomass globally (Dixon et al. 1994). In this context, the nature of tropical tree growth rhythms, the maximum age attained by the low- land rain forest trees, and their response to climate variability are still questions under discussion.
Transcript of Age and Long-term Growth of Trees in an Old-growth … la madera de ocho especies muestreada de...

306
BIOTROPICA 35(3): 306–317 2003
Age and Long-term Growth of Trees in an Old-growth Tropical RainForest, Based on Analyses of Tree Rings and 14C1
Esther FichtlerInstitut fur Forstbotanik, Universitaet Goettingen, Buesgenweg 2, D-37077, Goettingen, GermanyDeborah A. ClarkDepartment of Biology, University of Missouri–St. Louis, 8001 Natural Bridge Road, St. Louis, Missouri63121-4499, U.S.A.
and
Martin WorbesInstitut fur tropischen und subtropischen Pflanzenbau, Universitaet Goettingen, Grisebachstrasse 6, D-37077,Goettingen, Germany
ABSTRACTIn an old-growth tropical wet forest at La Selva, Costa Rica, we combined radiocarbon (14C) dating and tree-ringanalysis to estimate the ages of large trees of canopy and emergent species spanning a broad range of wood densitiesand growth rates. We collected samples from the trunks of 29 fallen, dead individuals. We found that all eight sampledspecies formed visible growth rings, which varied considerably in distinctiveness. For five of the six species for whichwe combined wood anatomical studies with 14C-dates (ring ages), the analyses demonstrated that growth rings wereof annual formation. The oldest tree we found by direct ring counting was a Hymenolobium mesoamericanum Lima(Papilionaceae) specimen, with an age of ca. 530 years at the time of death. All other sampled individuals, includingvery large trees of slow-growing species, had died at ages between 200 and 300 years. These results show that, evenin an everwet tropical rain forest, tree growth of many species can be rhythmic, with an annual periodicity. This studythus raises the possibility of extending tree-ring analyses throughout the tropical forest types lacking a strong dryseason or annual flooding. Our findings and similar measurements from other tropical forests indicate that the max-imum ages of tropical emergent trees are unlikely to be much greater than 600 years, and that these trees often dieearlier from various natural causes.
RESUMENEn una area de bosque muy humedo tropical localizado en La Selva, Costa Rica, usamos una combinacion de analisisde radiocarbono (14C) y analisis de anillos de crecimiento, para estimar las edad de arboles del dosel con un amplioespectro de tasas de crecimiento y de densidades de la madera. Analizamos la madera de ocho especies muestreada detroncos de 29 individuos muertos (arboles caıdos). Encontramos que las ocho especies estudiadas forman anillos decrecimiento visibles, pero que estos varıan en la distincion del lımite de sus anillos. Para cinco de las seis especies paralas cuales combinamos un estudio anatomico de la madera con fechado de 14C (edades de los anillos), nuestro analisisdemostro que los anillos de crecimiento fueron anuales. El arbol mas viejo en nuestra muestra fue un ejemplar deHymenolobium mesoamericanum Lima (Papilionaceae), que murio a la edad de aproximadamente 530 anos. Las edadesa las que murieron los demas indivıduos analizados, incluyendo arboles de gran tamano y de lento crecimiento, fueronentre 200 y 300 anos. Estos resultados demuestran que, aun en un bosque tropical muy humedo, el crecimiento demuchas especies de arboles puede presentar un crecimiento annual rıtmico. Este estudio demuestra que es posibleextender el analisis de anillos de crecimiento por todo el tropico, incluyendo las zonas de bosque que no experimentanni una estacion seca fuerte ni inundacion anual. Nuestros resultados, y los de otros estudios en bosques tropicales,indican que es poco probable que las edades maximas de arboles tropicales emergentes lleguen a ser mucho mayoresa los 600 anos, y que tales arboles frecuentemente mueran a edades menores por causas naturales.
Key words: emergent trees; La Selva; Costa Rica; old canopy; radiocarbon dating; tree ring analysis; tropical wet forest.
UNDERSTANDING THE PAST AND CURRENT DYNAMICS
OF THE WORLD’S TROPICAL rain forests is a pressingchallenge for ecologists. Tropical evergreen forestsare disproportionately important in the global car-bon cycle. They have been estimated to account
1 Received 28 February 2003; revision accepted 3 July2003.
for 32 percent of terrestrial net primary productiv-ity (Field et al. 1998), and combined with tropicaldry forests, they comprise an estimated 59 percentof forest biomass globally (Dixon et al. 1994). Inthis context, the nature of tropical tree growthrhythms, the maximum age attained by the low-land rain forest trees, and their response to climatevariability are still questions under discussion.

Age of Tropical Rain Forest Trees 307
The very large trees in tropical rain forests playimportant roles in ecosystem structure and func-tion. In spite of their low densities (usually ,3%of stems $10 cm diam), they can comprise 25–40percent of the total aboveground biomass (Brown& Lugo 1992, Brown et al. 1995, Clark & Clark1996). Likely to be among the oldest trees present,these individuals could serve as valuable indicatorsof stand history. Very little is currently knownabout the growth histories and maximum ages ofthese large, tropical rain forest trees. In two recentstudies, individual large trees from Amazonian for-ests were aged by 14C-dating at 190 to 1380 years(1 tree, de Camargo et al. 1994; 20 trees, Cham-bers et al. 1998; cf. Worbes & Junk 1999). Therelatively few ongoing studies involving repeatedmeasurements of large samples of such trees intropical rain forests have spanned only 20 years orless (Condit et al. 1995, Clark & Clark 2000).
Given the importance of obtaining a better un-derstanding of how tropical forests have grown inthe past, methods are needed for extending the ex-isting knowledge base. One approach is throughthe study of tree rings. When trees form annualgrowth rings, the ring patterns can be used in var-ious ways to gain information on historical growthrates, tree age, and past environmental conditions(Cain & Suess 1976, Worbes 1999). Although nu-merous studies have demonstrated annual ring for-mation in the tropics (Coster 1927, 1928; Berlage1931; Mariaux 1981; Devall et al. 1995; Drew1998; Worbes 1999; Dunisch et al. 2002; Worbeset al. 2003), these cases have been restricted to for-ests with a strong annual dry season or with annualflooding. Until now, it has been concluded (cf. Bor-mann & Berlyn 1981, Swaine 1994, Kurokawa etal. 2003) that reliable annual growth rings are notproduced in forests of the everwet tropics.
In this study, we investigated the feasibility oftree-ring analysis in the lowland tropical wet forestat the La Selva Biological Station, Costa Rica. Withdata from our combined analyses of wood anatomyand 14C-dates (ring ages), we demonstrated the ex-istence of reliable annual growth rings in five eco-logically diverse tree species, and we used the ringsto estimate the age and long-term mean growthrates of trees from these species. For three addi-tional species, we demonstrated the existence ofrecognizable growth zones in the wood.
STUDY AREA AND SITECONDITIONSThe 1550 ha La Selva Biological Station of theOrganization for Tropical Studies (OTS) is located
in the Atlantic lowlands of Costa Rica at 108269N,838599W (35–150 m elev.). La Selva’s soils rangefrom relatively fertile entisols and inceptisols to in-fertile ultisols (Sollins et al. 1994). The flora in-cludes 339 tree species (Hartshorn & Hammel1994; D. A. Clark & D. B. Clark, pers. obs.). Acomprehensive site description is given in McDadeet al. (1994).
Mean annual rainfall (42 yr) is 4230 mm andall months average at least 100 mm; the highestprecipitation falls during June–August, and thelowest average rainfall occurs during February–April (OTS records). Every year during those driermonths, there are periods without appreciable rain-fall (with daily rainfall ,5mm, which does notcontribute substantially to soil moisture because oflosses through interception and evapotranspiration(McDade & Hartshorn 1994); these rainless peri-ods average 12 days but can last up to 30 days inexceptional cases (Sanford et al. 1994). A four-yearrecord of soil moisture (1998–2002; Schwenden-mann et al. 2003; L. Schwendenmann, pers.comm.) shows a brief but marked drop in soilmoisture each year during the drier season.
MATERIALS AND METHODS
We took samples from 29 trees of eight species(Table 1) that were part of long-term, ongoingstudies of either tree demography (TREES Project;Clark & Clark 2000) or ecosystem carbon cycling(CARBONO Project; cf. Clark et al. 2002). Be-cause we could not use destructive methods on liv-ing or standing dead trees, we used fallen dead treesof known species and death year. Using dead ma-terial brought some difficulties because many of thesamples showed heartrot or were partly decom-posed (especially species with lower wood density).Further, many of these trees likely died from se-nescence and showed extremely small incrementsin the outer stem regions. All eight species analyzedin this study (Table 1) were non-pioneer speciesfound in old-growth forest; three of them reachedthe canopy, three were emergent, and two specieswere subcanopy trees.
We took cross-sectional wood discs or segmentswith a chainsaw above any buttresses but otherwiseas near as possible to the base of the bole. Afterdrying the discs in an air-conditioned laboratory,we polished them with sandpaper using a grit upto 400–600. We examined the wood anatomicalstructure macroscopically for growth zones andring boundaries. To improve the contrast of thering boundaries, we sometimes carefully moistened
308 Fichtler, Clark, and Worbes
TABLE 1. The eight tree species investigated for tree rings and 14C content at the La Selva Biological Station, Costa Rica.For each species, a voucher specimen is indicated in brackets (vouchers are at the Herbario Nacional de CostaRica).
Species FamilyStratumattained
No. ofindividuals
Guatteria aeruginosa Standl. [Celso Alvarado 50]Protium pittieri (Rose) Engl. [James Folsom 10078]Balizia elegans (Ducke) Barneby & J. W. Grimes [B. Hammel 17319]Pentaclethra macroloba (Willd.) Kuntze [B. Hammel & J. Trainer 12795]Dipteryx panamensis (Pittier) Record & Mell [R. Robles 1199]Hymenolobium mesoamericanum H. C. Lima [R. Aguilar 19]Minquartia guianensis Aubl. [G. Herrera 2250]Simarouba amara Aubl. [R. Robles 1670]
AnnonaceaeBurseraceaeMimosaceaeMimosaceaePapilionaceaePapilionaceaeOlacaeaeSimaroubaceae
SubcanopySubcanopyEmergentCanopyEmergentEmergentCanopyCanopy
242
113232
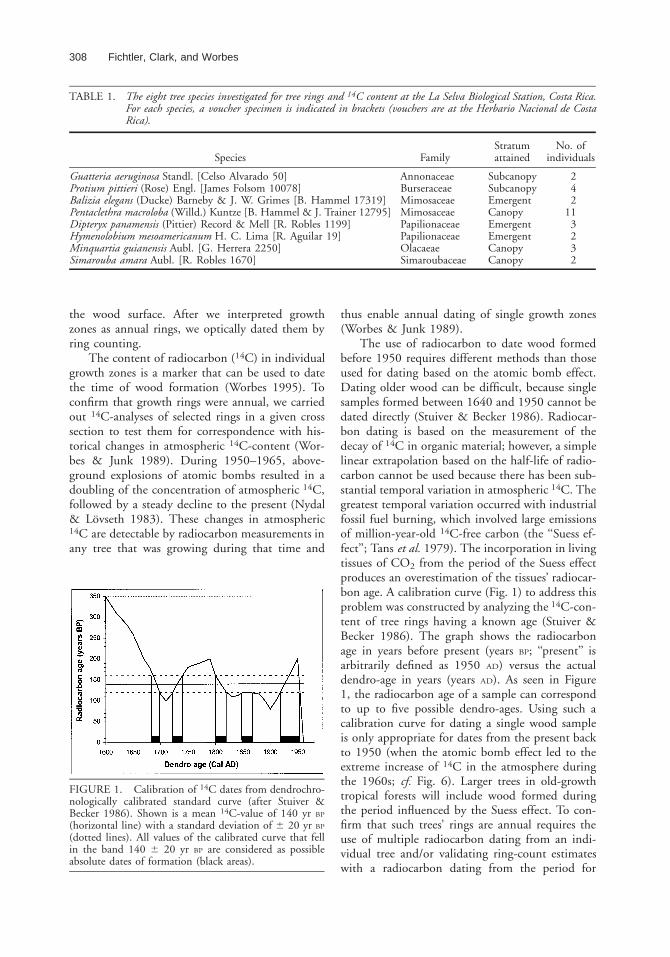
FIGURE 1. Calibration of 14C dates from dendrochro-nologically calibrated standard curve (after Stuiver &Becker 1986). Shown is a mean 14C-value of 140 yr BP
(horizontal line) with a standard deviation of 6 20 yr BP
(dotted lines). All values of the calibrated curve that fellin the band 140 6 20 yr BP are considered as possibleabsolute dates of formation (black areas).
the wood surface. After we interpreted growthzones as annual rings, we optically dated them byring counting.
The content of radiocarbon (14C) in individualgrowth zones is a marker that can be used to datethe time of wood formation (Worbes 1995). Toconfirm that growth rings were annual, we carriedout 14C-analyses of selected rings in a given crosssection to test them for correspondence with his-torical changes in atmospheric 14C-content (Wor-bes & Junk 1989). During 1950–1965, above-ground explosions of atomic bombs resulted in adoubling of the concentration of atmospheric 14C,followed by a steady decline to the present (Nydal& Lovseth 1983). These changes in atmospheric14C are detectable by radiocarbon measurements inany tree that was growing during that time and
thus enable annual dating of single growth zones(Worbes & Junk 1989).
The use of radiocarbon to date wood formedbefore 1950 requires different methods than thoseused for dating based on the atomic bomb effect.Dating older wood can be difficult, because singlesamples formed between 1640 and 1950 cannot bedated directly (Stuiver & Becker 1986). Radiocar-bon dating is based on the measurement of thedecay of 14C in organic material; however, a simplelinear extrapolation based on the half-life of radio-carbon cannot be used because there has been sub-stantial temporal variation in atmospheric 14C. Thegreatest temporal variation occurred with industrialfossil fuel burning, which involved large emissionsof million-year-old 14C-free carbon (the ‘‘Suess ef-fect’’; Tans et al. 1979). The incorporation in livingtissues of CO2 from the period of the Suess effectproduces an overestimation of the tissues’ radiocar-bon age. A calibration curve (Fig. 1) to address thisproblem was constructed by analyzing the 14C-con-tent of tree rings having a known age (Stuiver &Becker 1986). The graph shows the radiocarbonage in years before present (years BP; ‘‘present’’ isarbitrarily defined as 1950 AD) versus the actualdendro-age in years (years AD). As seen in Figure1, the radiocarbon age of a sample can correspondto up to five possible dendro-ages. Using such acalibration curve for dating a single wood sampleis only appropriate for dates from the present backto 1950 (when the atomic bomb effect led to theextreme increase of 14C in the atmosphere duringthe 1960s; cf. Fig. 6). Larger trees in old-growthtropical forests will include wood formed duringthe period influenced by the Suess effect. To con-firm that such trees’ rings are annual requires theuse of multiple radiocarbon dating from an indi-vidual tree and/or validating ring-count estimateswith a radiocarbon dating from the period for

Age of Tropical Rain Forest Trees 309
FIGURE 2. Growth zone type 1 (density variations): Guatteria aeruginosa (a; 50x), Protium pittieri (b; 66x), Pen-taclethra macroloba (c; 50x), Simarouba amara (d; 50x), Balizia elegans (e, 66x), Dipteryx panamensis (f; 16x).
which 14C-dates are not affected by the Suess ef-fect.
The samples for radiocarbon dating were iso-lated with an electric band saw. To prevent anycontamination of the wood sample with chainsawoil, we removed all parts of the samples that werein contact with the chain. For a given tree, we triedto get a series of three to five growth zones thatwere probably formed after 1950, to compare theinferred growth periodicity to dates around the
1964 ‘‘bomb peak.’’ When we counted more than300 rings in a tree’s cross section, we isolated theinnermost growth ring and additional rings. In thecase of one tree (Hymenolobium mesoamericanum),we isolated only the innermost ring. Although wetried in all cases to isolate single rings, in some treesthe very narrow growth zones forced us to includeseveral rings in a sample. The radiocarbon analysesof the samples were performed by means of an ac-celerator mass spectrometer at the University of Er-

310 Fichtler, Clark, and Worbes
TABLE 2. Phenology, distinctiveness of the growth zones, mean increments (calculated by dividing the diameter by thenumber of rings) of the analyzed individuals, and growth zone types (1 5 density variations; 2 5 marginalparenchyma bands; 3 5 patterns of parenchyma bands; 4 5 variation in vessel distribution) of the studiedspecies.
Species
Phenology
Deciduous Evergreen
Growth zone
Distinct Indistinct
Meanincrement
(mm)
Growthzonetype
Guatteria aeruginosaProtium pittieriBalizia elegansDipteryx panamensis
XX
XX
XXXX
3.251.973.973.46
111
1, 2, 4Hymenolobium mesoamericanumPentaclethra macrolobaMinquartia guianensisSimarouba amara
XXXX
XX
XX
2.812.571.676.96
31, 2
41, 3
FIGURE 3. Growth zone type 2 (marginal parenchymabands): Pentaclethra macroloba (a; 66x), Dipteryx pana-mensis (b; 32x).
FIGURE 4. Growth zone type 3 (patterns of parenchy-ma bands): Hymenolobium mesoamericanum (a; 6x), Sim-arouba amara (b; 6x).
langen–Nurnberg (Germany) and at Beta AnalyticInc. (Miami, Florida). The pretreatment of thesamples followed usual procedures to isolate cellu-lose.
RESULTS
WOOD STRUCTURE AND GROWTH ZONES. All inves-tigated species formed visible rings that varied con-siderably in distinctiveness (Figs. 2–5). Balizia ele-gans, Dipteryx panamensis, H. mesoamericanum,Pentaclethra macroloba, and the two subcanopy spe-cies, Guatteria aeruginosa and Protium pittieri, allshowed distinct rings. Minquartia guianensis andSimarouba amara formed indistinct rings. All threedeciduous species showed distinct rings (Table 2).The distinctiveness of the growth rings variedamong the evergreen species and was not clearlyrelated to tree growth rate; even the indistinctgrowth zones, however, could be identified andwere therefore suitable for tree ring analysis.
We classified the ring structure of all studiedspecies (Table 2) according to growth zone type(Coster 1927, 1928; Worbes 1995): (1) densityvariations (Fig. 2); (2) marginal parenchyma bands(Fig. 3); (3) patterns of parenchyma bands (Fig. 4);
and (4) variations in vessel distribution and/or ves-sel size (Fig. 5). Three species (D. panamensis, P.macroloba, and S. amara) combined multiplegrowth zone types (Table 2). Dipteryx panamensisalso showed ontogenetic changes in growth zonestructure: in juvenile wood, density variations werecombined with variation in vessel distribution;most mature wood showed marginal parenchymabands combined with variation in vessel distribu-tion, but wood in the outermost stem region onlyshowed marginal parenchyma bands (a similar on-togeny of wood structure occurs in the tropical treespecies Carapa guianensis; Bauch & Dunisch2000).
ANNUAL GROWTH PERIODICITY. The macroscopicage estimation of the wood samples from one in-dividual of each of four species (S. amara, B. ele-gans, D. panamensis, and P. macroloba) was per-formed by counting the rings between a 14C-datedring sample and the outermost ring, which was dat-ed according to the year of death. Because most ofthe sampled trees had been lying on the forest floorfor several years before sampling, some discs hadlost the bark and we could not exclude the possi-bility that some of the sapwood had been lost

Age of Tropical Rain Forest Trees 311
FIGURE 5. Growth zone type 4 (variation in vessel dis-tribution): Dipteryx panamensis (a; 8x), Minquartia guia-nensis (b; 20x).
FIGURE 6. Results of radiocarbon dating from woodsamples of Simarouba amara. Shown are the 14C valuesof the atmosphere (lines) in the Northern Hemisphere atvarious latitudes (from Hua et al. 1999, adopted) and the14C values of the predated growth rings (gray circles).After correcting the macroscopic dating, the samples weredated 12 yr older (white circles). To confirm the correc-tion of the macroscopic dating, the growth ring from1964 was analyzed (black circle).
FIGURE 7. Results of radiocarbon dating test rings (seetext) sampled from one individual of Balizia elegans (tree3345, 67 cm diam; triangles) and one individual of Dip-teryx panamensis (tree CES 350, 103 cm diam; squares)compared with the atmospheric 14C in the NorthernHemisphere at various latitudes (lines; from Hua et al.1999, adopted).
through decomposition. For these trees, we radio-carbon-dated three to five different growth rings,so that we could subsequently correct the macro-scopic age estimate with a high degree of certainty.
Such a correction was illustrated by the S. ama-ra individual (tree 234, 54.7 cm diam, 25.4 cmdisc radius; Fig. 6). The disc from this tree lackedbark, and the disc’s reduced radius (compared toits last measured diameter while alive) indicatedthat some of the outermost growth rings had alsobeen lost. For the macroscopic ring-count age es-timate, we treated the outermost ring on the discas if it were the last ring formed by the tree beforeits death in 1998. As expected, the results of the14C-analyses did not confirm this dating sequence.After interpreting the 14C-values of the analyzedgrowth rings (Fig. 6), we corrected our ring datingby 12 years under the prediction that 12 years ofsapwood had been lost through decomposition. Toconfirm this interpretation, we isolated the growthring that was, after the corrected dating, identifiedas formed in 1964. The 14C-content subsequentlydetermined for that growth ring was equivalent tothe atmospheric 14C-content in 1964; this agree-ment validated our ring interpretation for the en-tire disc and demonstrated the existence of annualgrowth rings in S. amara.
For the wood samples from the analyzed indi-viduals of B. elegans and D. panamensis, in all casesthe 14C-content of the tested rings correspondedwith the 14C-values of the atmosphere at the timeof their predicted formation (Fig. 7), thus confirm-ing that the identified growth rings were annual.
For P. macroloba, however, the results fromsuch test radiocarbon dating of the wood samplesdid not confirm our ring-count predictions for thetime of formation. One possible explanation is thegreat prevalence of wedging rings in this species.Further studies will be needed to determine if P.macroloba can be reliably aged by ring counts.
AGE DATING. For two additional species, we con-firmed the existence of annual rings by two differ-
ent approaches, depending on whether the sampledindividual’s lifetime exceeded 350 years (the periodover which radiocarbon dating of wood is affectedby the Suess effect; see Methods).
According to our count of the growth rings forone individual of H. mesoamericanum (530 growthrings; tree 8058, 1983 diam: 128.0 cm, death yr:1992), this tree was old enough to be aged by ra-diocarbon dating the center of the disc. The radio-carbon date of the disc center (570 6 40yr BP)confirmed our ring counting.

312 Fichtler, Clark, and Worbes
FIGURE 8. Sketch of half cross section of Minquartiaguianensis on which 291 rings were counted (wavy linesare for orientation and do not indicate individual rings)Wood samples predicted to various ages for radiocarbonanalyses were isolated from this disc (black squares).
FIGURE 9. Calibration of the 14C dating of wood sam-ples dendrochronologically predicted to 1700 AD and1780 AD from Minquartia guianensis. Shown are the cal-ibrated standard curve and the radiocarbon age for thesample predated to 1700 AD (sample A: horizontal line at70 yr BP) and to 1780 AD (sample B: horizontal line at290 yr BP), with the corresponding standard deviationsfor both results (dashed line: sample A; dotted line: sam-ple B). The corresponding possible dates of formation arefor the sample 1700 AD, the periods 1690–1730 AD, and1805–1930 AD (white areas), while for the sample 1780AD, the periods 1600–1670 AD and 1770–1790 AD (grayareas) are considered possible times of formation. Forsamples C, D, and E, only the period ca. 1950 AD canbe considered as the possible time of formation (blackarea).
We aged the one M. guianensis individual (tree50268, diam: 38.2 cm, death yr: 1998) throughinterpretation of radiocarbon-dates for the inner-most and several other rings, in combination withdendrochronological analysis of the growth zones.The calibration curve of Stuiver and Becker (1986)shows that a given 14C-concentration in a woodsample corresponds to several real (dendro-) ages(Fig. 1). We used this calibration curve to comparethe ages we predicted for the Minquartia ringsbased on optical counts, to subsequent radiocarbondates for these rings. We isolated five wood sections(Fig. 8): two individual rings with the predictedtimes of formation of 1700 AD and 1780 AD, re-spectively; and three multiple-ring samples fromthe outer stem region, which we expected to showcontrasting 14C-values from before and after the1964 bomb peak based on our ring counts. Thesample B (predicted to 1780 AD by tree ring anal-ysis) has three theoretically possible dendro-ages(Fig. 9; year ranges indicated by shaded bars). Thetime between 1940–1950 AD could be tentativelyexcluded, however, because the sample was taken170 mm in from the stem periphery, and 170 mmof radial growth in 60 years (5.6 mm diam growth/yr) was unlikely based on the very low median di-ameter increments measured for Minquartia at LaSelva (0.7–2.8 mm/yr, depending on size class;Clark & Clark 1992). The dendro-age 1600–1670AD could also be excluded for sample B becausethis sample was formed later than sample A. Theperiod 1805–1930 AD for sample A could be ex-cluded because A was formed earlier than sampleB (Figs. 8 and 9). The samples C, D, and E did
not hit the curve of the 14C-concentration in theatmosphere before and after the bomb peak in1964. Sample C was calibrated to 1680–1730 AD,1810–1930 AD, and 1950 AD. The samples D andE were both calibrated to all the following dateranges: 1670–1710 AD, 1720–1770 AD, 1800–1880 AD, 1910–1940 AD, and 1950 AD. All periodsprior to 1900 could be excluded because of thesethree samples’ peripheral position in the disc; evenconsidering extremely low increments, samples tak-en 5–20 mm from the bark could not be hundredsof years old, when the stem center, which is 210mm from the bark, was only 300 years old. Thesimilar calibrated dating ca. 1930–1950 of thethree samples was influenced by the fact that it wasnot possible to isolate single rings from before andafter the bomb peak, but also indicates that theoptical predating deviated ca. 10–20 years. Theseresults do, however, demonstrate how reduced theincrements in the outer stem regions were in thelast four decades before the tree died.
TREE AGES. The ring counts provide a basis todetermine the (minimum) ages of the sampled treesfor the five species found to produce annual rings

Age of Tropical Rain Forest Trees 313
TABLE 3. Minimum tree ages and lifetime mean diameter growth rates, based on ring counting of the studied trees, andestimated maximum ages for these species. Mean annual increments were calculated by dividing the tree’sdiameter by the total number of rings counted; when parts of the disc were missing, the increments werecalculated on the base of the analyzed radius. The total number of rings gives an estimated age in years;however, for trees in which the disc showed loss of outer sapwood or the core due to decomposition, the age isunderestimated. For each species, we divided the largest observed diameter at La Selva (Clark & Clark 1999)by mean annual diameter increment for the slowest-growing individual of the analyzed trees of that species, toestimate a theoretical maximum age.
Species
Diameter ofsampled disc atdeath year (cm)
No. of ringscounted
(5 age, yr)
Mean annualdiameter
increment(mm/yr)
Largestobserved
diameter (cm)
Theoreticalmaximumage (yr)
Balizia elegans 96.2108.0
211a,b
182b4.273.68
137 372
Dipteryx panamensis 116.0103.0
91.8
229a,b
252270a,b
2.904.093.40
187 645
Hymenolobium mesoamericanum 90.0128.0
301a
5303.202.42
156 645
Minquartia guianensis 38.242.651.5
291a
223a
274a
1.261.722.05
82 651
Simarouba amara 51.654.7
63a
76a7.246.68
81 121
a Some outer sapwood appeared to have been lost from the disc through decomposition.b Center missing from disc.
(Table 3). For the cases in which the analyzed discshad lost either heartwood or outer sapwood by de-composition, these ages are underestimates. At LaSelva, young trees of these species show very slowdiameter-growth rates (Clark & Clark 1992,1999); such processes in the very young stagescould be missed from the discs with missing cores.Similarly, adult trees can show very slow diameterincrements prior to death (D. A. Clark & D. B.Clark, pers. obs.); discs missing part of the outersapwood could miss periods of such slowed growth.
DISCUSSION
The growth and dynamics of tropical trees and for-ests remain poorly understood. Little is knownabout the age and the age structure of old-growthtropical forests. In a review of recent literature onforest dynamics and carbon fluxes, the maximumage reported for the oldest tropical forest trees wasfound to increase rapidly among successive studies(Worbes & Junk 1999). These studies calculate ormodel the maximum age of tropical trees and tendto overestimate considerably (Condit et al. 1995),with the focus on the oldest theoretically possibleage.
In the temperate zone, the easiest way to de-termine a tree’s age is ring counting. Annual growth
rings represent a reliable basis for age determina-tion and interpretation of tree growth under dif-ferent ecological conditions or management re-gimes (Stahle et al. 1999). It is a continuing par-adigm in the literature (cf. Studhalter 1955, Roth1981, Whitmore 1984, Richards 1996) that trop-ical trees do not show annual rings because of thelack of seasonality in the tropics; however, in manytropical regions, drought periods exist. That theseannual drought periods are reflected by annualrings in the wood has been shown now for tropicalmoist and tropical dry forests in all tropical regions(Coster 1927, 1928; Berlage 1931; Mariaux 1981;Worbes 1984, 1995, 1997, 1999; Dunisch et al.2002).
The climate in La Selva is classified as everwet.Under such conditions, continuous growth of thetrees has been often assumed (cf. Raven et al.1999). Usually, however, the everwet tropics actu-ally experience distinct seasonal variations in rain-fall each year (e.g., long-term averages of monthlyrainfall totals for La Selva [OTS records] of 156–271 mm/mo for the four months of the drier sea-son; 353–524 mm/mo for the eight months of thewetter season).
Phenological observations of the tree and shrubcommunities in La Selva (Frankie et al. 1974) in-dicate a distinct seasonality in phenology. A large

314 Fichtler, Clark, and Worbes
annual peak in leaf litter fall occurs during March–April, the latter part of the drier season (D. A.Clark, pers. obs.). Lieberman and Lieberman(1987) found the highest seedling mortality duringthe drier season. Tree growth studies at La Selva,however, have produced contradictory results. Haz-lett (1987) found the highest diameter incrementfor the evergreen tree species P. macroloba in thedrier period between January and April, whileBreitsprecher and Bethel (1990) found cambialdormancy in observations of small samples of Pen-taclethra macroloba and other species in the sameperiod of the year. In a recent two-year study onleaf phenology and monthly growth of canopy andemergent tree species at La Selva, O’Brien (2002)found diverse patterns for several of our study spe-cies. Dipteryx panamensis showed annual decidu-ousness in the first half of the year, although in-dividuals were not closely synchronous; these treesshowed significantly reduced growth rates imme-diately following the drier season. Simarouba ama-ra, an evergreen species, showed a synchronizedflushing of new leaves in the early part of the drierseason. For two other species for which we foundannual growth rings (B. elegans and H. mesoame-ricanum), however, O’Brien (2002) found individ-uals to be strongly asynchronous in their leaf phe-nology, with individuals being deciduous duringdiverse parts of the year (some Hymenolobium treeswere deciduous more than once in a year).
Although the underlying mechanisms are thusstill unclear, our results have demonstrated that de-ciduous and evergreen tree species show growth pe-riodicity at La Selva. All eight investigated speciesformed visible growth rings in the wood. Weshowed with complementary radiocarbon datingthat these rings were annual, in five of the six testedspecies. These annual rings indicate a yearly periodof slower growth or cambial dormancy. Triggers forsuch growth periodicity in trees can be flooding,variations in temperature, precipitation, or daylength (Worbes 1995, Borchert & Rivera 2001,Schongart et al. 2002), possibly supplemented byan unidentified internal rhythm (Alvim & Alvim1978, Roth 1981). The trees investigated in thisstudy were from upland forest at La Selva, thus notinfluenced by flooding. We believe that the slightvariation in mean monthly temperatures of 1 to28C is unlikely to trigger growth periodicity, be-cause (1) diurnal variation is much higher and (2)temperature never falls to growth-limiting values.Internal rhythms that could produce such growthperiodicities have never been demonstrated fortropical trees. There are some indications that at
least stem–succulent tropical tree species (Bursera-ceae, Bombacaceae) react to changes in photope-riod (Borchert & Rivera 2001), but growth peri-odicity of deciduous and evergreen species has beenlinked to seasonal stress factors (i.e., drought,flooding) clearly independent of photoperiod(Schongart et al. 2002). The trigger of growth ringformation in La Selva trees cannot be attributed tothe extreme stress situations (i.e., at least two aridmonths, or annual floods) that have been assumedto be required for the formation of reliable annualrings in the tropics (Stahle et al. 1999, Worbes1999). Although the climate at La Selva is classifiedas everwet, the forest may experience majordrought stress during short periods without appre-ciable rainfall (i.e., ,5 mm) in the drier seasonbetween February and April (Sanford et al. 1994);such periods lead to a brief but marked drop insoil moisture content (Schwendenmann et al.2003). Limitation in soil water can inhibit rootgrowth and affects root metabolism (Mulkey &Wright 1996, Sanford & Cuevas 1996). Such shortdry periods (10–21 d) may be effective triggers forgrowth periodicity in trees (cf. Medway 1972). Theexistence of an annual growth periodicity was dem-onstrated by our dendrochronological analysis ofthe canopy and emergent tree species we investi-gated at La Selva. The seasonality of their growthmay reflect the effects of predictable, moderatelylong rainless periods in the annual ‘‘drier season’’in this tropical wet forest.
Our results show that dendrochronologicalstudies can be extended to all parts of the tropics.This will lead to great progress in understandingthe dynamics and history of tropical forests. Theexistence of annual rings in tropical wet forest treesopens wide prospects for age and growth determi-nation. The problems of age dating with indirectmethods have been discussed elsewhere (Worbes &Junk 1999). Our findings illustrate why single ra-diocarbon-dates cannot be used to determine theage of a tree that is between 50 and 350 years old,although such findings are reported in the literature(Ogden 1981; Chambers et al. 1998, 2001). Agingtrees of this age range requires either multiple ra-diocarbon dating from the same disc, or the com-bination of 14C-dating with wood anatomical anddendrochronological methods, as performed in ourstudy.
The oldest tree among the 12 individuals weaged in this study was an H. mesoamericanum witha diameter of 128 cm and an age of 530 years. Thefour other investigated very large trees (.70 cmdiam) from La Selva were ca. 200–300 years old

Age of Tropical Rain Forest Trees 315
when they died. Lieberman et al. (1985) estimateda mean life span of 190 years (range: 42–442 yr)for trees 10 cm diameter or greater based on sim-ulated growth trajectories of trees at La Selva. Ourpreliminary findings, although based on small sam-ples, suggest that large trees at La Selva often dieat ages considerably below the maximum possibleage. The maximum theoretical life expectancies wecalculated for the La Selva trees (Table 3) parallelthose found for broadleaf tree species in the tem-perate zone (Loehle 1988). There, the definitelyoldest tree species (Quercus spp.) have a maximumlife expectancy of ca. 600 years, and the typical ageat mortality (5 rotation period of the long livingspecies) is 200–300 years less than that maximum.Similar estimates have been reported for seasonaldry and flooded tropical forests based on direct ringcounting. In an old-growth forest in Cameroon themean age of trees 10 cm diameter or greater wasestimated to be only 60 years, although trees witha theoretical maximum age of 220 years were found(Worbes et al. 2002). In Amazonian floodplain for-ests, the species Piranhea trifoliata reaches ages upto 500 years in stands where all other trees aremuch younger (Worbes 1997). In a Central Ama-zonian forest, however, three individual trees werereported to be between 900 and 1400 years old(Chambers et al. 1998). Those findings are clearoutliers to all other findings to date from tropicalforests, and they were based on single samples from
each tree and single radiocarbon estimations, with-out any corroborating data or replicated analyses;thus, to confirm such widely greater longevities fortropical trees will require more data than currentlyexist. Together with our results from La Selva, theexisting data present a picture of old-growth trop-ical forests in which relatively low average tree lon-gevity (Korning & Balslev 1994) combines withmuch greater maximum tree ages. It is very reason-able that in a highly diverse broadleaf forest, veryfew species with very few individuals can survivefor centuries when the overall stand is very dynam-ic. This is probably true for the La Selva forest andshould be confirmed in the future by more exten-sive tree sampling and direct ring counting.
ACKNOWLEDGMENTS
We thank the Organization for Tropical Studies and thestaff of the La Selva Biological Station for logistical sup-port during the fieldwork. We also thank two anonymousreviewers for valuable criticism and comments that im-proved the manuscript. We are indebted to the AndrewW. Mellon Foundation, which supported the fieldworkand the radiocarbon analyses (grant to D. A. Clark andD. B. Clark), Beta Analytic Laboratories for granting dis-counted analysis rates, and both the U.S. National Sci-ence Foundation (grants DEB 9629245 and LTREBgrants DEB 9407581, 9981591) and the U.S. Depart-ment of Energy (grant no. DE-FG02-96ER62289) forsupporting our mapping and long-term censuses of LaSelva trees.
LITERATURE CITEDALVIM, P. T., AND R. ALVIM. 1978. Relation of climate to growth periodicity in tropical trees. In P. B. Tomlinson and
M. H. Zimmermann (Eds.). Tropical trees as living systems, pp. 445–464. Cambridge University Press,London, England.
BAUCH, J., AND O. DUNISCH. 2000. Comparison of growth dynamics and wood characteristics of plantation-grownand primary forest Carapa guianensis in Central Amazonia. IAWA J. 21: 321–333.
BERLAGE, H. P. 1931. Over het verband tusschen de dikte der jaarringen van djatiboomen (Tectona grandis L.f.) en deregenval op Java. Tectona 24: 939–953.
BREITSPRECHER, A. AND J. S. BETHEL. 1990. Stem-growth periodicity of trees in a tropical wet forest of Costa Rica.Ecology 71: 1156–1164.
BORCHERT, R., AND G. RIVERA. 2001. Photoperiodic control of seasonal development and dormancy in tropical stem–succulent trees. Tree Physiol. 21: 201–212.
BORMANN, F. H., AND G. BERLYN. 1981. Age and growth rate of tropical trees: new directions for research. YaleUniversity School of Forestry and Environmental Studies, New Haven, Connecticut.
BROWN, S., AND A. E. LUGO. 1992. Aboveground biomass estimates for tropical moist forests of the Brazilian Amazon.Interciencia 17: 8–18.
BROWN, I. F., L. A. MARTINELLI, W. W. THOMAS, M. Z. MOREIRA, C. A. C. FERREIRA, AND R. A. VICTORIA. 1995.Uncertainty in the biomass of Amazonian forests: an example from Rondonia, Brazil. Forest Ecology andManagement 75: 175–189.
CAIN, W. F., AND H. E. SUESS. 1976. Carbon 14 in tree rings. J. of Geophys. Res. 81/21: 3688–3694.CHAMBERS, J. Q., N. HIGUCHI, AND J. P. SCHIMMEL. 1998. Ancient trees in Amazonia. Nature 391: 135–136.———, ———, E. S. TRIBUZY, AND S. E. TRUMBORE. 2001. Carbon sink for a century. Nature 410: 429.CLARK, D. A., AND D. B. CLARK. 1992. Life history diversity of canopy and emergent trees in a neotropical rainforest.
Ecol. Monogr. 62: 315–344.———, AND ———. 1999. Assessing the growth of the tropical rain forest trees: issues for forest modeling and
management. Ecol. Appl. 9: 981–997.

316 Fichtler, Clark, and Worbes
CLARK, D. B., AND D. A. CLARK. 1996. Abundance, growth and mortality of very large trees in Neotropical lowlandrain forest. For. Ecol. and Manage. 80: 235–244.
———, AND ———. 2000. Tree growth, mortality, physical condition, and microsite in old-growth lowland tropicalrain forest. Ecology 81: 294 and Ecol. Arch.: http://www.esapubs.org/archive/ecol/E081/003/default.htm
———, ———., S. BROWN, S. F. OBERBAUER, AND E. VELDKAMP. 2002. Stocks and flows of coarse woody debrisacross a tropical rain forest nutrient and topography gradient. For. Ecol. and Manage. 164: 237–248.
CONDIT, R., S. P. HUBBELL, AND R. B. FOSTER. 1995. Mortality rates of 205 Neotropical tree and shrub species andthe impact of a severe drought. Ecol. Monogr. 65: 418–439.
COSTER, C. 1927. Zur Anatomie und Physiologie der Zuwachszonen- und Jahresringbildung in den Tropen. Ann.Jard. Bot. Buitenzorg 37: 49–160.
———. 1928. Zur Anatomie und Physiologie der Zuwachszonen und Jahresringbildung in den Tropen. Ann. Jard.Bot. Buitenzorg 38: 1–114.
DE CAMARGO, P. B., R. P. SALOMAO, S. TRUMBORE, AND L. A. MARTINELLI. 1994. How old are large brazil-nut trees(Bertholletia excelsa) in the Amazon? Sci. Agr. 51: 389–391.
DEVALL, M. S., B. R. PARRESOL, AND S. J. WRIGHT. 1995. Dendroecological analysis of Cordia alliodora, Pseudobombaxseptenatum and Annona spraguei in central Panama. IAWA J. 16: 411–424.
DIXON, R. K., S. BROWN, R. A. HOUGHTON, A. M. SOLOMON, M. C. TREXLER, AND J. WISNIEWSKI. 1994. Carbonpools and flux of global forest ecosystems. Science 263: 185–190.
DREW, A. P. 1998. Growth rings, phenology, hurricane disturbance, and climate in Cyrilla racemiflora L., a rain foresttree of the Luquillo Mountains, Puerto Rico. Biotropica 30: 35–49.
DUNISCH, O., J. BAUCH, AND L. GASPAROTTO. 2002. Formation of increment zones and intraannual growth dynamicsin the xylem of Swietenia macrophylla, Carapa guianensis and Cedrela odorata (Meliaceae). IAWA J. 23: 101–119.
FIELD, C. B., M. J. BEHRENFELD, J. T. RANDERSON, AND P. FALKOWSKI. 1998. Primary production of the biosphere:integrating terrestrial and oceanic components. Science 281: 237–240.
FRANKIE, G. W., H. G. BAKER, AND P. A. OPLER. 1974. Comparative phenological studies of trees in tropical wet anddry forests in the lowlands of Costa Rica. J. Ecol. 62: 881–913.
HARTSHORN, G. S., AND B. E. HAMMEL. 1994. Vegetation types and floristic patterns. In L. A. McDade, K. S. Bawa,H. A. Hespenheide, and G. S. Hartshorn (Eds). La Selva: ecology and natural history of a neotropical rainforest, pp. 73–89. University of Chicago Press, Chicago, Illinois.
HAZLETT, D. L. 1987. Seasonal cambial activity for Pentaclethra, Goelthalsia, and Carapa trees in a Costa Rica lowlandforest. Biotropica 19: 357–360.
HUA, Q., M. BARBETTI, M. WORBES, J. HEAD, AND V. A. LEVCHENKO. 1999. Review of radiocarbon data from atmo-spheric and tree ring samples for the period 1945–1997 AD. IAWA J. 20: 261–283.
KORNING, J., AND H. BALSLEV. 1994. Growth rates and mortality patterns of tropical lowland tree species and therelation to forest structure in Amazonian Ecuador. J. Trop. Ecol. 10: 151–166.
KUROKAWA, H., T. YOSHIDA, T. NAKAMURA, J. LAI, AND T. NAKASHIZUKA. 2003. The age of tropical rain-forest canopyspecies Borneo ironwood (Eusideroxylon zwageri), determined by 14C dating. J. Trop. Ecol. 19: 1–7.
LIEBERMAN, D., AND M. LIEBERMAN. 1987. Forest tree growth and dynamics at La Selva, Costa Rica (1969–1982). J.Trop. Ecol. 3: 347–358.
———, ———, G. HARTSHORN, AND R. PERALTA. 1985. Growth rates and age size relationships of tropical wet foresttrees in Costa Rica. J. Trop. Ecol. 1: 97–109.
LOEHLE, C. 1988. Tree life history strategies: The role of defenses. Can. J. For. Res. 18: 209–222.MARIAUX, A. 1981. Past efforts in measuring age and annual growth in tropical trees. In F.H. Bohrmann and G.
Berlyn (Eds.). Age and growth rate of tropical trees: New directions for research, pp. 20–30. Yale UniversityPress, New Haven, Connecticut.
MCDADE, L. A., K. S. BAWA, H. A. HESPENHEIDE, AND G. S. HARTSHORN (EDS.). 1994. La Selva: Ecology and naturalhistory of a Neotropical rain forest. University of Chicago Press, Chicago, Illinois.
———, AND G. S. HARTSHORN. 1994. La Selva Biological Station. In L. A. McDade, K. S. Bawa, H. A. Hespenheide,and G. S. Hartshorn (Eds.). La Selva: Ecology and natural history of a Neotropical rain forest, pp. 6–14.University of Chicago Press, Chicago, Illinois.
MEDWAY, L. 1972. Phenology of a tropical rain forest in Malaysia. Bot. J. Linn. Soc. 4: 117–146.MULKEY, S. S., AND S. J. WRIGHT. 1996. Influence of seasonal drought on the carbon balance of tropical forest plants.
In S. S. Mulkey, R. L. Chazdon, and A. P. Smith (Eds.). Tropical forest plant ecophysiology, pp. 187–216.Chapman and Hall, New York, New York.
NYDAL, R., AND K. LOVSETH. 1983. Tracing bomb 14C in the atmosphere 1962–1980. J. Geophys. Res. 88: 3621–3642.
O’BRIEN, J. J. 2002. The effects of climate on the growth and physiology of tropical rainforest trees. Ph.D. dissertation,Duke University, Durham, North Carolina.
OGDEN, J. 1981. Dendrochronological studies and determination of tree ages in the Australian tropics. J. Biogeogr.8: 405–420.
RAVEN, P. H., R. E. EVERT, AND S. E. EICHORN. 1999. Biology of plants. W. H. Freemann and Company, New York,New York.
RICHARDS, P. W. 1996. The tropical rain forest, 2nd edition. Cambridge University Press, Cambridge, England.ROTH, I. 1981. Structural patterns of tropical barks. Handbuch Pflanzenanatomie IX/3. Borntraege, Berlin, West
Germany.

Age of Tropical Rain Forest Trees 317
SANFORD, R. L., AND E. CUEVAS. 1996. Root growth and rhizosphere interactions in tropical forests. In S. S. Mulkey,R. L. Chazdon, and A. P. Smith (Eds.). Tropical forest plant ecophysiology, pp. 268–300. Chapman and Hall,New York, New York.
———, P. PAABY, J. C. LUVALL, AND E. PHILLIPS. 1994. Climate, geomorphology and aquatic systems. In L. A. McDade,K. S. Bawa, H. A. Hespenheide and G. S. Hartshorn (Eds.). La Selva: Ecology and natural history of aNeotropical rain forest, pp. 19–33. University of Chicago Press, Chicago, Illinois.
SCHONGART, J., M. T. F. PIEDADE, S. LUDWIGSHAUSEN, V. HORNA, AND M. WORBES. 2002. Phenology and stem-growthperiodicity of tree species in Amazonian floodplain forests. J. Trop. Ecol. 18: 581–597.
SCHWENDENMANN, L., E. VELDKAMP, T. BRENES, J. J. O’BRIEN, AND J. MACKENSEN. 2003. Spatial and temporal variationin soil CO2 efflux in an old-growth Neotropical rain forest, La Selva, Costa Rica. Biogeochemistry 64: 111–128.
SOLLINS, P., F. M. SANCHO, R. CH. MATA, AND R. L. SANFORD JR. 1994. Soils and soil process research. In L. A.McDade, K. S. Bawa, H. A. Hespenheide, and G. S. Hartshorn (Eds.). La Selva: Ecology and natural historyof a Neotropical rain forest, pp. 34–53. University of Chicago Press, Chicago, Illinois, USA.
STAHLE, D. W., P. T. MUSHOVE, M. K. CLEAVELAND, F. ROIG, AND G. A. HAYNES. 1999. Management implications ofannual growth rings in Pterocarpus angolensis from Zimbabwe. For. Ecol. and Manage. 124: 217–229.
STUDHALTER, R. A. 1955. Tree growth. Bot. Rev. 21: 1–71.STUIVER, M., AND B. BECKER. 1986. High-precision decadal calibration of the radiocarbon time scale, AD 1950–2500
BC. Radiocarbon 28: 863–910.SWAINE, M. D. 1994. Long-term studies of tropical forest dynamics. In R. A. Leigh and A. E. Johnston (Eds.). Long-
term experiments in agricultural and ecological sciences, pp. 305–319. CAB International, Wallingford, Eng-land.
TANS, P. P., A. F. M. DE JONG, AND W. G. MOOK. 1979. Natural atmospheric 14C variation and the SUESS-effect.Nature 280: 826–828.
WHITMORE, T. C. 1984. Tropical rain forests of the Far East. Clarendon Press, Oxford, England.WORBES M. 1984. Periodische Zuwachszonen an Baumen zentralamazonischer Uberschwemmungswalder. Naturwis-
senshaften 71: 157.———. 1995. How to measure growth dynamics in tropical trees. A review. IAWA J. 16: 337–351.———. 1997. The forest ecosystems of the Amazonian floodplains. In W. J. Junk (Ed.). The Amazonian floodplains:
Ecology of a pulsing system, pp. 223–265. Ecological studies, vol. 126. Springer Verlag, Berlin.———. 1999. Annual growth rings, rainfall-dependent growth and long-term growth patterns of tropical trees from
the Caparo Forest Reserve in Venezuela. J. Ecol. 87: 391–403.———, AND W. J. JUNK. 1989. Dating tropical trees by means of the 14C from bomb tests. Ecology 70: 503–507.———, AND ———. 1999. How old are tropical trees? The persistence of a myth. IAWA J. 20: 255–260.———, R. STASCHEL, A. ROLOFF, AND W. J. JUNK. 2003. Tree ring analysis reveals age structure, dynamics and wood
production of a natural forest stand in Cameroon. For. Ecol. and Manage. 173: 105–123.


















