




Photo by Karl Steinbrenner Charge Card Supervisor/Reviewer Training 2014.

ACCEPTED
September 2006AEM Accepts, published online ahead of print on 29
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Transformation of the green alga Haematococcus pluvialis with a phytoene
desaturase for accelerated astaxanthin biosynthesis
Jens Steinbrenner1,* and Gerhard Sandmann2
1 Department of Plant Physiology and Biochemistry, Universität Konstanz, D-78434
Konstanz, Germany;
2 Institute for Molecular Biosciences 213, J.W. Goethe Universität, P.O. Box 111932, D-
60054 Frankfurt/M., Germany
Jens Steinbrenner
Department of Plant Physiology and Biochemistry
Universität Konstanz
D-78434 Konstanz
Germany
Tel +49-7531-883668
Fax +49-7531-883042
E-mail: [email protected]
*corresponding author, Fax +49-7531-883042, e-mail: Jens.Steinbrenner@uni-
konstanz.de
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

1
ABSTRACT 1
Astaxanthin is a high-value carotenoid, which is used as a pigmentation source in 2
fish aquaculture. Additionally, a beneficial role of astaxanthin as a food supplement for 3
humans is suggested. The unicellular alga Haematococcus pluvialis is a suitable 4
biological source for astaxanthin production. In the context of the strong biotechnological 5
relevance of H. pluvialis, we developed a genetic transformation protocol for metabolic 6
engineering of this green alga. First, the gene coding for the carotenoid biosynthesis 7
enzyme phytoene desaturase was isolated from H. pluvialis and modified by site directed 8
mutagenesis changing leucin 504 to arginine. In an in vitro assay, the modified phytoene 9
desaturase was still active in the conversion of phytoene to ζ-carotene and revealed a 43 10
fold higher resistance towards the bleaching herbicide norflurazon. Upon biolistic 11
transformation using the modified phytoene desaturase as reporter gene and selection 12
with norflurazon, the integration in the nuclear genome of H. pluvialis and gene and 13
protein expression of phytoene desaturase were shown by Southern, Northern and 14
Western blots, respectively, in eleven transformants. Some of the transformants exhibited 15
a higher carotenoid content in the green state which correlated with increased non-16
photochemical quenching. This chlorophyll fluorescence measurement can be used as a 17
screening procedure for stable transformants. Stress induction of astaxanthin biosynthesis 18
by high light showed an accelerated accumulation of astaxanthin in one of the 19
transformants as compared to the wild type. Our results strongly indicate that the 20
modified phytoene desaturase gene is a useful tool for the genetic engineering of 21
carotenoid biosynthesis in H. pluvialis. 22
23
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
Key Words: astaxanthin, carotenoid biosynthesis, genetic engineering, green algae, 24
Haematococcus pluvialis, phytoene desaturase, transformation. 25
26
Abbreviations: Bhy, β-carotenoid hydroxylase; CrtP, cyanobacterial phytoene 27
desaturase; NPQ, non-photochemical quenching; OHA, optimal Haematococcus medium 28
supplemented with sodium acetate; OHM, optimal Haematococcus medium; Pds, 29
phytoene desaturase ; Pds-L504R phytoene desaturase with codon exchange at position 30
504 from leucine to arginine; RF, resistance factor; WT, wild type. 31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
47
48
49
INTRODUCTION 50
51
Among all carotenoids used as food supplement for humans, for food coloration or as 52
pigmentation source for egg yolk the ketocarotenoid astaxanthin (3,3’-dihydroxy-diketo-53
β,β-carotene-4,4’-dione) is the most important from the biotechnological viewpoint (4, 54
22). Besides the use as colorant it was discussed that astaxanthin supplementation might 55
be a practical and beneficial strategy in human health management due to its 56
neuroprotective, its immunomodulating and its antioxidant potential (13). Although most 57
of the astaxanthin is produced by total chemical synthesis today and sold at a price of 58
$2500 kg-1 (16), the high price and the increasing demand as a food supplement provide a 59
good opportunity for naturally produced astaxanthin. 60
Biosynthesis of astaxanthin has been observed only in a limited number of 61
organisms, e.g. in some marine bacteria, in the yeast Xanthophyllomyces dendrorhous 62
and in some green algae (16). The unicellular green alga Haematococcus pluvialis reveals 63
the highest astaxanthin accumulation (up to 4% /g [dry weight]) and seems to be a very 64
promising source for natural astaxanthin (3). A promising strategy to further improve the 65
astaxanthin yield of H. pluvialis is the genetic engineering of the carotenoid biosynthesis 66
pathway. 67
There are two reports of successful transformation of H. pluvialis by particle 68
bombardment using the β-galactosidase gene (lacZ) (26, 41). The reporter gene was 69
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
transiently expressed under the control of the SV40 promoter or the β-carotene ketolase 70
promoter. Unfortunately, a stable transformation was not obtained. Moreover, to date, 71
there are no reports of a dominant selective transformation system for H. pluvialis. For 72
the establishment of a stable transformation system several genetic tools are needed. This 73
includes the choice of a reporter gene, a strong promoter and an adapted termination 74
signal and a suitable method for stable DNA transfer into the cells. For H. pluvialis no 75
strong promoters were isolated, yet. A further problem known from the transformation of 76
other green algae is, that for foreign genes introduced e.g. into Chlamydomonas 77
reinhardtii display low transcript levels despite being fused to strong endogeneous 78
promoters (24). It has been suggested that such failures of exogenous gene expression 79
might be due to mismatches in codon usage patterns (34), epigenic silencing by DNA 80
methylation or changes in chromatin domain structure (1, 15), and/or rapid mRNA 81
degradation (7). These hurdles might be overcome by selecting a modified endogenous 82
gene from the organism to be transformed (19, 29, 31). 83
Sequence information for genes that could be adapted as dominant selective 84
markers after appropriate genetic modifications are not available from H. pluvialis to 85
date. The limited sequence information that is available includes most of the carotenoid 86
biosynthesis genes (12, 17, 21, 23, 37, 38, 40). Among these the phytoene desaturase 87
(Pds) is the target for the bleaching herbicide norflurazon. (6, 8, 25, 33). For the phytoene 88
desaturase (CrtP) from Synechococcus sp. PCC 7942 it is known that four known amino 89
acid exchanges lead to norflurazon resistance as shown in figure 1A. Since H. pluvialis is 90
highly sensitive against this herbicide and high sequence similarities exist between the 91
Synechococcus sp. and the H. pluvialis protein, an exchange of one of the amino acids in 92
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
Fig. 1A should be a promising strategy to engineer norflurazon resistance to the 93
Haematococcus Pds and use this modified pds gene as a dominant selectable marker and 94
a reporter gene for the transformation of H. pluvialis. 95
The present investigation describes the isolation of the gene coding for the 96
carotenoid biosynthesis enzyme Pds from H. pluvialis. After mutation of the pds gene 97
resulting in a norflurazone-resitant phytoene desaturase, this gene was used for successful 98
nuclear transformation and the genetic engineering of the carotenoid biosynthesis 99
pathway for accelerated astaxanthin biosynthesis. 100
101
102
103
104
MATERIALS AND METHODS 105
106
Haematococcus pluvialis strain, growth conditions, media. H. pluvialis Flotow 107
NIES-144 was obtained from the National Institute for Environmental Studies (NIES), 108
Tsukuba, Japan and was grown in basal medium containing acetate (18). Growth was 109
performed at 22°C in Erlenmeyer flasks without aeration under a dark/light cycle of 12 h 110
low light (20 µmol photons m-2 s-1, provided by universal-white lamps Osram L65W/25S) 111
and 12 h dark for 3 d (final cell density approximately 3.5 × 105 cells/ml). Cultures were 112
shaken manually once a day. For high light treatments, H. pluvialis was grown at 175 113
µmol photons m-2 s-1 at continuous light as indicated. For strong light treatments, H. 114
pluvialis cells were set at 700 µmol photons m-2 s-1. For transformation protocols, optimal 115
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
H. pluvialis medium (OHM) (11), supplemented with 2.42 g Tris-Acetate pH 7.2 (OHA) 116
was used. After transformation, cells were plated on solid OHA medium, containing 5 117
µM norflurazon. 118
Construction of genomic DNA library, screening and DNA sequencing. For 119
genomic DNA isolation, H. pluvialis cells were collected by centrifugation after a growth 120
period of 3 d. The cells were frozen and subsequently powdered under liquid nitrogen 121
using a mortar and pestle. Genomic DNA was isolated by using the Plant DNA Isolation 122
Kit (Roche) following the manufacturers instructions. The genomic DNA library was 123
constructed using the SuperCos I Vector Kit (Stratagene) according to the manufacturers 124
instructions. The screening of the genomic DNA library for the isolation of the pds gene 125
was carried out by colony hybridization using a specific pds cDNA probe according to 126
the instructions in the DIG Nonradioactive Nucleic Acid Labeling and Detection System 127
(Roche). Probe labeling and hybridization were carried out as described earlier (38). 128
Positive clones were purified and cosmid DNA was isolated using High Pure Plasmid 129
Isolation Kit (Roche) and analyzed by Southern blotting. After restriction of DNA by 130
enzyme digestion, a 5.78 kb XbaI/XhoI fragment harbouring a pds gene fragment was 131
isolated and cloned in the pBluescriptSK- vector (Stratagene). The resulting plasmid was 132
named pBluescript-pds. The nucleotide sequence of H. pluvialis pds gene was determined 133
for both strands using the Abi Prism Dye Terminator Cycle Sequencing Ready Reaction 134
Kit (Perkin Elmer). The analysis of nucleotide and derived amino acid sequences was 135
carried out using the Lasergene program (DNASTAR, Inc.). The intron exon organization 136
of the pds gene is shown in Fig. 1B. 137
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
Construction of the transformation vector Plat-pds-L504R. The mutated form 138
of the pds gene from H. pluvialis (with codon exchange 504 leucine to arginine) was 139
constructed by site-directed mutagenesis using the vector pBluescript-pds and the primers 140
5´-cc aag cag aag tac cgc gcc tcc atg gag gg-3´ and 5´-ccc tcc atg gag gcg cgg tac ttc tgc 141
ttg g-3´ (the codon for the exchange from leucine to arginine is marked in bold). For 142
mutagenesis the Quickchange Mutagenesis Kit (Stratagene) was used. The plasmid with 143
the modified pds was named pBluescript-pds-L504R. To verify the nucleotide exchange, 144
the H. pluvialis pds gene was partially sequenced using the Abi Prism Dye Terminator 145
Cycle Sequencing Ready Reaction Kit (Perkin Elmer). To maintain the multiple cloning 146
site for the final transformation vector, the mutated pds gene was amplified by PCR using 147
the primers 5´-cgc gat atc ggt gag ggg ttc aag tgc c-3´ and 5´-ccg ata tcg ttt gaa ttt tgg ctt 148
gtt tgc-3´. In this step, the end standing XhoI and XbaI restriction sites of the pds gene 149
were eliminated and two EcoRV sites were introduced into the PCR product. By 150
restriction digest with EcoRV the PCR product was cloned into the NaeI site of a 151
pBluescriptSK(-) vector. The resulting transformation vector was named pPlat-pds-152
L504R (Fig. 1 C). 153
cDNA synthesis. To recover the mutation within the cDNA pool of pds WT and 154
the mutated pds (Pds-L504R) transcripts, total RNA was isolated and cDNA synthesis 155
was performed using iScript cDNA synthesis Kit (Biorad) according to the 156
manufacturer’s instructions. For the specific PCR amplification of pds cDNAs, the 157
primers 5´-act cta gaa atg cag aca aca atg cgt ggc-3´ together with 5´-ccc gga ttc aat agt 158
ata cac cac aag c-3´ were used. The resulting PCR products were sequenced to verify the 159
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
nucleotide exchange at the codon position 504 within the cDNA pool of the H. pluvialis 160
pds transcripts. 161
Transformation and growth of Escherichia coli. Escherichia coli strain BL21 162
(DE3) (Novagene) was transformed with individual plasmids for the expression of Pds 163
from H. puvialis in its wild type (Pds WT) or mutated form (Pds-L504R). For the 164
production of phytoene as substrate in the enzyme reaction, E. coli JM101 cells 165
harbouring the plasmid pACCRT-EB were used (28). Growth was performed in LB 166
medium supplied with appropriate antibiotics in concentrations of 100 µg/mL ampicillin 167
and 50 µg/mL chloramphenicol. To the enzyme expressing strains, 168
isopropylthiogalactose (1 mM) was added to the culture after an optical cell density 169
(OD600, 1 cm light path) of 0.6 was reached. Growth was performed at 25 °C overnight. 170
Plasmid pBlue-pdsWT, which mediates the overexpression of the Pds WT of H. pluvialis, 171
was constructed by RT-PCR reaction of the corresponding cDNA using the primers 5´-172
act cta gaa atg cag aca aca atg cgt ggc-3´ together with 5´-ccc gga ttc aat agt ata cac cac 173
aag c-3´ and cloning in frame into the XbaI/EcoRI sites of the vector pBluescriptSK- 174
(Stratagene). 175
The Pds-L504R from H. pluvialis was constructed by site-directed mutagenesis 176
using the vector pBlue-pdsWT and the primers 5´-cc aag cag aag tac cgc gcc tcc atg gag 177
gg-3´ and 5´-ccc tcc atg gag gcg cgg tac ttc tgc ttg g-3´ (codon 504 for the exchange from 178
leucine to arginine is marked in bold). The resulting plasmid was named pBlue-pds-179
L504R For the overexpression of the Pds WT and Pds-L504R the pET system 180
(Novagene) was used. Therefore the plasmids pBlue-pdsWT and pBlue-pds-L504R were 181
restriction digested and the corresponding cDNAs were cloned in frame into the 182
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
SacI/XhoI sites of the expression vector pET24b. The resulting plasmids were pET24b-183
pdsWT and pET24b-pds-L504R. 184
PDS assay including substrate and enzyme preparation. Extraction of 185
phytoene, the preparation of the enzyme and the Pds enzyme assay were performed 186
according to Breitenbach and coworker (5). To the PDS assay, norflurazon was applied in 187
increasing concentrations from 0.001 µM to 0.5 µM to the Pds WT enzyme and from 0.5 188
µM to 20 µM to the engineered Pds-L504R 189
Transformation protocol. Haematococcus cells were grown in basal medium for 190
3 d under standard conditions (see above). Cells were concentrated by centrifugation and 191
re-suspended to a density of approximately 1 × 108 cells/ml OHA medium. 1 ml of 192
concentrated cells was used for each bombardment, plated on nylon filters on OHA 193
plates. For transformation, the Biolistic PDS-1000/He system was used (Biorad). 194
Fifty µl of M 17 tungsten particles solution (60 mg/ml in H2O) were mixed with 2 195
µl of DNA solution (1 µg/µl), 50 µl 2.5 M CaCl2 and 20 µl of 0.1 M spermidine-base. 196
The mixture was incubated for 10 min at RT, centrifuged for 10 sec and the pellet was re-197
suspended in 250 µl ethanol. After an additional centrifugation for 10 sec the pellet was 198
re-suspended in 50 µl ethanol. Twenty µl of DNA-coated particles were layered on a 199
macrocarrier. 200
Plates were bombarded at a distance of 7.5 cm under vacuum of 25 mmHg using 201
1350 psi rupture disks. After transformation, the cell-containing nylon disks were placed 202
in 50 ml OHA medium for a regeneration period of 24 h under a dark/light cycle of 12 h 203
low light (20 µmol photons m-2 s-1). After this period the cells were concentrated by 204
centrifugation and plated on 10 8-cm-diameter Petri dishes containing selective OHA 205
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
medium. Colonies appearing after one month were picked and re-streaked at least three 206
times on selective OHA medium. 207
Northern and Southern blot analysis. Probe labeling and hybridization were 208
carried out according to the instructions in the DIG Nonradioactive Nucleic Acid 209
Labeling and Detection System (Roche). The H. pluvialis cDNAs pds (EMBL GenBank 210
accession no. X86783) and bhy (EMBL GenBank accession no. AF162276) were applied 211
as probes. For the analysis of vector integration into the nuclear genome of H. pluvialis, 212
an additional hybridization probe against the ampicillin resistance cassette was used. 213
For Northern Blot analysis the H. pluvialis cells were collected by centrifugation 214
after 3 d of growth. The cells were frozen and subsequently powdered under liquid 215
nitrogen using a mortar and pestle. RNA was then isolated according to the miniprep 216
RNA extraction procedure as described (36). The Northern blot analysis was performed 217
as described earlier (38). 218
For Southern blot analysis, genomic DNA was isolated by using the Plant DNA 219
Isolation Kit (Roche) following the manufacturers instructions. The genomic DNA (5 µg) 220
was XbaI/XhoI digested, the DNA fragments were separated by electrophoresis on a 0.8% 221
agarose gel, transferred to a positively charged nylon membrane (Roche) and hybridized 222
with pds cDNA probe or the ampicilin resistance gene probe in the presence of 50% 223
formamide at 42°C. 224
Isolation of proteins and Western blot analysis. The polyclonal antibody 225
against the Pds was kindly provided by Dr. C. Hagen (12). For protein isolation, H. 226
pluvialis transformants were collected by centrifugation after a growth period of 3 d. The 227
cells were frozen and subsequently powdered under liquid nitrogen using a mortar and 228
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
pestle. The proteins were precipitated and 75 µg of each transformant protein extract was 229
separated on a 12% SDS-polyacrylamide gel, electroblotted to a PVDF membrane (0.45 230
µm pore-size) (Amersham Biosiences Europe GmbH, Freiburg, Germany), probed with 231
anti-Pds antibody at 1:5000 dilution and a horseradish peroxidase-linked anti-rabbit-232
immunoglobin antibody at 1:20000 dilution. Signals were visualized by using the 233
enhanced chemiluminescence (ECL) Western blotting detection system and Hyperfilm 234
ECL (Amersham, Biosiences). A second, identically loaded gel was run as control and 235
stained with Coomassie Brilliant Blue 250R (not shown). 236
Carotenoid and chlorophyll analysis. For carotenoid analysis in the green state, 237
cells under strong light were instantly frozen in liquid nitrogen and freeze-dried. 238
Extraction was with methanol containing 6% (w/v) KOH at 60°C for 20 min. The 239
extracts were partitioned against 10% (v/v) diethyl ether in petrol and the carotenoid 240
content determined by HPLC on a 25 cm C18 Vydac 218TP54 column with methanol as 241
mobile phase at 20°C (10). Reference carotenoids were isolated from cyanobacteria (39) 242
or pepper leaves (35). Carotenoid extraction, HPLC analysis and the quantification of 243
astaxanthin and astaxanthin esters from H. pluvialis was carried as reported earlier (38). 244
Chlorophyll was extracted in a similar way using methanol without the addition of KOH 245
and quantitated as described by Lichtenthaler and Wellburn (20). Carotenoid and 246
chlorophyll values are given as the means of four independent determinations together 247
with the standard deviation. 248
Chlorophyll fluorescent measurements. Screening of transformants for changed 249
fluorescence parameters was performed using the Walz pulse-amplitude modulation 250
(PAM) Fluorescent Imaging System, model IMAG (Walz, Effeltrich, Germany). 251
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
Transformants and WT cells were grown on OHA solid medium without norflurazon for 252
3 weeks. Before measurement the cells were adapted for 4 hours in the dark. A saturating 253
pulse (800 ms) of blue light (2400 µmol photons m-2 s-1) was provided via LED lamps for 254
determination of Fm (maximum fluorescence in the dark-adapted state). Actinic light (180 255
µmol photons m-2 s-1) was provided via blue light LED lamps. The measuring beam was 256
switched to 2 Hz during saturating pulses and during actinic illumination. NPQ was 257
calculated as (Fm- Fm´)/ Fm´ using the provided imaging software. 258
Nucleotide sequence and accession number. The nucleotide sequence of the 259
transformation vector pPlat-pds-L504R has been deposited in the GeneBank database 260
under the accession number DQ404589. 261
262
263
264
RESULTS 265
266
Isolation of the Haematococcus pluvialis nuclear gene coding for Pds. 267
Norflrazon resistance of the mutated pds gene was chosen as a suitable selection marker. 268
Furthermore, transformation with this gene should modify carotenogenesis in H. 269
pluvialis. For the isolation of the pds gene, a genomic DNA library of H. pluvialis was 270
screened using a specific hybridization probe against the pds cDNA. Eight different 271
clones with genomic fragments sizes between 30 and 45 kbp harboring the putative pds 272
gene were collected. By XbaI/XhoI digestion a 5.78 kb fragment was isolated and cloned 273
in the corresponding restriction sites of the pBluescriptSK(-) vector (Stratagene). The 274
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
DNA fragment was sequenced and an alignment with the known pds cDNA of H. 275
pluvialis revealed a 2 kb promoter region, 7 introns and a 423 bps 3´-untranslated region. 276
The intron and exon organization is shown in Fig. 1B. 277
The Pds-L504R shows higher resistance towards the bleaching herbicide 278
norflurazon in the in vitro assay compared to the Pds WT. The desaturase activity of 279
an E. coli recombinant Pds WT and Pds-L504R of H. pluvialis (with codon exchange 504 280
leucine to arginine) was measured by the conversion of phytoene to ζ-carotene after the 281
addition of different concentrations of norflurazon using an in vitro assay. The Dixon plot 282
of the reciprocal product formation versus increasing concentrations of the inhibitor 283
norflurazon resulted in a straight line with r = 90% for the Pds WT and r = 91% for the 284
Pds-L504R, respectively (Fig. 2) (9). From the intercepts with the x-axis the ki values for 285
norflurazon as inhibitor were calculated to be 0.14 µM and 5.98 µM for the heterologous 286
expressed Pds WT and Pds-L504R, respectively. In comparison to the wild type enzyme 287
the mutated form revealed a 43 fold higher resistance. 288
Analysis of Haematococcus transformants. After transformation of H. pluvialis 289
with the vector pPlat-pds-L504R and pBluescript-Pds by particle bombardment and a 290
regeneration period the cells were plated on norflurazon containing OHA plates. After 291
one month of growth, colonies appeared in samples that were transformed with the vector 292
pPlat-pds-L504R. No growth was visible when the vector pBluescript-pds, harbouring the 293
pds WT gene, was used for transformation. It is difficult to estimate the efficiency of 294
transformation by particle bombardment on a cell basis because many of the cells are 295
outside the target zone. In these experiments using ∼108 cells and 1 µg of DNA, generally 296
100 transformants were recovered. Therefore, transformation efficiency was estimated in 297
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
the range of 10-6 cells/µg DNA. Although the transformation rate is rather low when 298
compared to C. reinhardtii, transformations with similar rates were reproducible and the 299
obtained transformants grow well under selective and nonselective conditions. 300
Transformants show additional copies of pds-L504R within the nuclear 301
genome. After a growth period of one month, the appearing colonies were re-streaked for 302
at least three rounds on norflurazon containing plates and then incubated in liquid media. 303
To determine if the resistance against norflurazon was due to stable nuclear integration of 304
the engineered pds gene, Southern blot analysis of the transformants was performed with 305
hybridization probes against the pds and the ampicillin resistance cassette of the 306
transformation vector. DNA was isolated from cultures of eleven individual 307
transformants and digested by restriction enzymes and hybridized with a probe against 308
the pds (Fig. 3A). The endogenous pds gene was detected in all transformants and the 309
WT H. pluvialis strain, the size of 5.78 kb is corresponding to the size of the XhoI/XbaI-310
fragment that was isolated and subcloned from the genomic DNA library of H. pluvialis. 311
All tested transformants show additional copies of the engineered pds -L504R gene with 312
larger sizes than the endogenous pds WT gene. To verify the integration of the transgenes 313
into the nuclear genome of H. pluvialis, additional Southern blot analysis using a 314
hybridization probe against the ampicillin resistance cassette of the transformation vector 315
was performed (Fig. 3B). The data support the results obtained in Fig. 3A, where the pds 316
cDNA was used as a hybridization probe. Almost all transformants with the exception of 317
one of the eleven transformants tested yielded one or more bands as visible evidence for 318
transgene integration into the genome. No ampcillin resistance cassette band was detected 319
in transformant P3 and the WT H. pluvialis, although this transformant was about 20-fold 320
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

15
more resistant to norflurazon as the WT in an assay with increasing norflurazon 321
concentration on OHA plates (data not shown). Transformant P3 showed the additional 322
bands between 9.41 and 23.1 kb in the Southern blot using the pds cDNA hybridization 323
probe (Fig. 3A). This finding indicates a loss of the ampicillin resistance cassette and 324
perhaps parts of the transformation vector during the integration event of the transgene 325
into the nuclear genome of H. pluvialis. 326
The transformation stability was analyzed using the plasmid pBluescript-pds-327
L504R, harboring the engineered pds-L504R gene within the multiple cloning site. After 328
a growth period over 20 weeks (approx. 80 generations, growth rate 0.58 d-1) under 329
nonselective conditions, still 70% of the analyzed transformants retained their 330
norflurazon resistance and revealed a stable transgene integration within the genome 331
(data not shown). 332
The transformants show increased pds expression levels compared to the WT 333
To monitor whether the additional copies of transgene in the genome of the eleven 334
analyzed transformants change the expression level of the pds mRNA, Northern blot 335
analysis was carried out. For further comparison of the pds expression levels between the 336
different transformants and the WT, total RNA was extracted from WT cells before and 337
after 18 h and 36 h high light treatment. Recently, it was demonstrated that the expression 338
of genes encoding enzymes involved in the carotenoid biosynthesis are up regulated in 339
response to increased light intensities (37, 38). Exposure of low light grown WT cultures 340
to higher light intensities resulted in the accumulation of pds transcripts (Fig. 4A). The 341
pds transcript level in the transformants grown at low light conditions were analyzed 342
(Fig. 4A). Seven out of eleven transformants revealed an unchanged basal expression of 343
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

16
the pds gene under the tested condition. The transformants P3, P6, P10, P11 showed 344
increased levels of pds transcripts compared to the WT (Fig. 4A). To gain further 345
information whether the up-regulation of the pds transcripts is due to stress effects or to 346
additional copies of the pds gene in the nuclear genome, the expression of the bhy gene 347
was tested under the same conditions (Fig 4B). In earlier experiments, it was shown that 348
the bhy gene expression was only detected under stress conditions (37, 38). In all 349
transformants, expression of the bhy gene was below detection. However, light induction 350
of the bhy gene was observed in the WT cells included as a stress control. Since the 351
transcript level not necessarily matches the protein level, Western blot analysis using 352
Pds-antibodies were performed. The results revealed that the amount of Pds correlates 353
with the level of its transcripts. 354
355
The transformants show a mixture of pds WT and pds-L504R transcripts 356
To gain further information on the expression of the transgenic pds -L504R as well as the 357
functionality of the promoter and terminator region of the 5.7 kb pds gene, cDNA 358
synthesis and subsequent sequencing of the pds transcript pool was performed. The 359
cDNAs of the wild type H. pluvialis and of the two transformants P3 and P11 were 360
investigated by sequencing (data not shown). No additional mutations were found within 361
the pds cDNA except a mixture of ctg and cgc at codon position 504 for the 362
transformants P3 and P11. The obtained pds WT cDNA was identical with that present in 363
the EMBL GenBank. These findings indicate that the 2 kb promoter region of the 364
engineered 5.7 kb pds gene fragment isolated from a H. pluvialis genomic DNA library is 365
sufficient to drive transgene expression. 366
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

17
Carotenoids, chlorophylls and non-photochemical quenching of selected 367
transformants. Analysis of norflurazon resistant Synechococcus sp. mutants revealed 368
that phytoene desaturation is a rate-limiting step in carotenoid biosynthesis in 369
cyanobactria (8). Furthermore, previous results indicated that the heterologous expression 370
of the phytoene desaturase gene (crtI) from Erwinia uredovora in tobacco changed the 371
composition of leaf carotenoids (27). To obtain more information about these processes 372
in H. pluvialis, transformants were grown under low light intensities or 1h strong light 373
and carotenoid concentration and composition was determined in green cells (Table 1). 374
Transformants P3 and P11 both showed a higher carotenoid content as compared with the 375
WT. Additionally, the chlorophyll concentration in transformant P3 was lower, resulting 376
in a carotenoid/chlorophyll ratio which is significantly higher than in the WT. Subsequent 377
HPLC analysis of the carotenoid composition in transformants P3, P11 and WT 378
illuminated for 1h with strong light, revealed for both transformant a significant increased 379
of lutein, zeaxanthin and β-carotene levels. 380
In addition, it was investigated whether the phytoene desaturation is a rate-381
limiting step for astaxanthin synthesis. Therefore, the transformants were grown for 3d 382
under low light conditions, then exposed to continuous high light illumination and 383
astaxanthin accumulation determined. In a time course, transformant P3 showed a visible 384
red phenotype after 8h of high light exposure whereas the other transformants and the 385
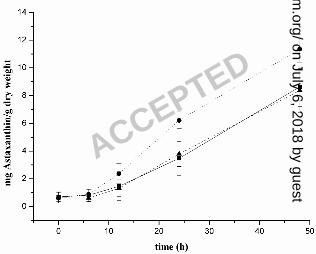
WT still remained green (Fig. 5). Quantification of astaxanthin from the two 386
transformants P3 and P11 and the WT cells after 48 h of high light illumination 387
confirmed that high light leads to enhanced astaxanthin levels in transformant P3 (11.4 ± 388
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

18
0.9 mg / g [dry weight]), whereas the levels in the WT (8.6 ± 1.4 mg / g [dry weight]) and 389
in transformant P11 (8.4 ± 1.6 mg / g dry weight) were 26% lower. 390
Chlorophyll fluorescence was determined with dark-adapted H. pluvialis WT and 391
transformants. The time course of non-photochemical quenching (NPQ) is shown in Fig. 392
6. WT cells reached a maximum value of 0.38 after 70 seconds. Transformants P3 and 393
P11 reached their maxima after the same time. However, the NPQ value was almost 2-394
fold higher for both. 395
396
DISCUSSION 397
398
The development of a transformation system depends on the availability of suitable 399
strong promoters, termination signals and the choice of promising reporter or resistance 400
genes. At present, there are no appropriate genetic tools available from H. pluvialis. The 401
pds/crtP gene from plants, algae and cyanobacteria is susceptible to the herbicide 402
norflurazon and four mutations in this enzyme from the cyanobacteria Synechoccocus sp. 403
PCC 7942 are known to acquire resistance (8). Since alignments of the amino acid 404
sequence of CrtP/Pds from Synechococcus sp. and H. pluvialis revealed a high sequence 405
similarity within the areas where the mutations were found, we followed a direct genetic 406
approach exchanging Leu504 to Arg in the phytoene desaturase from H. pluvialis (Fig. 407
1A). By in vitro enzyme analysis, it was shown that the engineered PDS-L504R was still 408
active, and exhibited a 43-fold higher resistance towards norflurazon compared to the 409
WT enzyme (Fig. 2). This decrease of norflurazone susceptibilty was sufficient for 410
selection of H. pluvialis transformed with the mutated pds gene. 411
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

19
After biolistic transformation of H. pluvialis and following selection on 412
norflurazon containing medium, the integration of the finally used transformation vector 413
pPlat-pds-L504R by Southern blot analysis was examined (Fig. 3). Using a pds cDNA 414
probe all transformants revealed additional copies of the pds gene within the nuclear 415
genome except the WT. For the 11 transformants that were analyzed is not fully 416
understood whether the transforming DNA was linked to the pds locus by homologous 417
recombination or the integration was due to heterologous recombination events within in 418
the genome. It was estimated for C. reinhardtii that homologous recombination events 419
take place at a ratio of 1:100 for a mutated protophorphyrinogen oxidase gene (ppx1) 420
when the biolistic method is used (31). A homologous recombination event by a single 421
crossover should result in a 6.4 kb and an 8.11 kb band in the Southern blot using the pds 422
cDNA probe. A homologous recombination event by a double crossover should lead to a 423
loss of the vector backbone and therefore the ampicillin resistance cassette. It is difficult 424
to estimate the correct size of the different bands that hybridize with the pds cDNA and 425
the ampicillin gene probe. Transformant 2 shows signals that could match this 426
assumption, but in general strong signals were found at the vector size of 8.7 kb. It is 427
more likely that the transforming DNA undergoes complex rearrangement and 428
concatemer formation before integration, resulting in complex tandem array of pPlat-pds-429
L504R DNA that is stable after integration into the genome. This has previously been 430
reported for integration of transgenes into the genome of the green alga Volvox carteri 431
that is taking place as tandem repeats due to homologous recombination events of the 432
plasmids forming circular concatemeric structures prior the integration into the genome 433
(1, 14). Interestingly, the transformants 5, 6, 10, 11 show very strong signals at the 8.7 kb 434
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

20
position, matching the vector size of 8.74 kb. To exclude that the intact plasmid was 435
maintained, the genomic DNA of these four transformants and the WT was restriction 436
digested with BstBI. This restriction enzyme does not cut the vector sequence. Using the 437
pds hybridization probe, the endogenous pds gene was detected in all transformants and 438
the WT H. pluvialis strain at a size of 9.4 kb (data not shown). The four transformants 439
show additional copies of the engineered pds -L504R gene with sizes about 23 kb and 440
larger (data not shown). 441
The changed expression level of the pds gene under standard growth conditions is 442
not linked to stress induction as shown by the bhy gene expression (Fig. 4). It is more 443
likely that the altered expression of the pds gene is due to multiple copies of the gene in 444
the genome of the transformants. Interestingly, the strength of expression is somehow 445
linked to the copy number of the transgene but there seem to be additional positional 446
effects. The integration locus of the introduced DNA in the genome seems to be an 447
important factor in determining the transgenic pds gene expression. Western blot analysis 448
revealed that pds gene transcript and protein level matched their degree of expression. 449
This result indicates that under standard growth conditions there is no posttranscriptional 450
regulation of the pds gene expression. 451
Furthermore, the analysis of total pds gene transcripts was performed for the WT 452
strain and the transformants P3 and P11. The two transformants showed a mixture of WT 453
and transgene transcripts indicating that the promoter is sufficient to drive transgenic pds-454
L504R gene expression and the endogenous pds gene is still intact. 455
The transformants were not only analyzed for transcription and protein expression 456
but also for possible effects of increased phytoene desaturase levels on carotenoid 457
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

21
metabolism. Carotenoid analysis of transformants P3, P11 has shown that total carotenoid 458
concentration was higher in P11 on a dry weight basis whereas this value was lower for 459
P3 (Table 1). Since the chlorophyll content was also lower, a pleiotropic effect on the 460
chloroplast can be assumed. Related to chlorophyll, total carotenoids are also increased 461
over the wild type. Both transformants synthesize higher levels of zeaxanthin and lutein, 462
as compared to the WT (Table 1). This is in accordance with findings that the 463
heterologous expression of a bacterial phytoene desaturase in tobacco changed the 464
composition of leaf carotenoids (27) and the carotenoid formation in tomato towards β-465
carotene at the expense of lycopene (32). Under light stress, the transformant P3 showed 466
an enhanced accumulation of astaxanthin that reached 26% higher levels after 48 h of 467
high light exposure as compared with the WT and transformant P11 (Fig. 5). All our 468
results indicate that the phytoene desaturating step is rate limiting for carotenoid 469
synthesis in the chloroplast of H. pluvialis under high light conditions. The degree of 470
non-photochemical quenching is related to the zeaxanthin content and the mount of 471
zeaxanthin relative to all xanthophyll cycle carotenoids calculated by the Z/(Z+A+V) 472
ratio (2). These zeaxanthin related parameters were both increased in transformants P3 473
and P11 (Tab. 1). In accordance with the higher zeaxanthin concentration, NPQ values 474
were almost 2-fold higher in the transformants (Fig 6). Consequently, NPQ measurement 475
may be used as a convenient screening procedure for transformants obtained with the 476
vector pPlat-pds-L504R. A video imaging of chlorophyll fluorescence has been proved 477
successful to detect Chlamydomonas xanthophyll cycle mutants (30). The results reported 478
here clearly demonstrate that the mutated pds gene can be used as dominant selective 479
marker for stable nuclear transformation of H. pluvialis. Thus, our transformation vector 480
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

22
pPlat-pds-L504R (Fig. 1C) resembles a basic tool for the genetic modification of this 481
alga. In addition, we report for the first time a specific genetic engineering of the 482
carotenoid biosynthesis pathway leading to increased carotenoid biosynthesis and an 483
accelerated accumulation of astaxanthin in H. pluvialis. The presented work resembles a 484
proof of concept for the genetic engineering of the carotenoid biosynthetic pathway in H. 485
pluvialis towards increasing astaxanthin production. 486
487
ACKNOWLEDGMENTS 488
This work was only possible due to the generous support from Prof. Dr. I. Adamska, 489
University of Konstanz, Germany and the Peter and Traudl Engelhorn Stiftung. We are 490
grateful to R. Miller-Sulger for excellent technical assistance and to Dr. H. Linden for 491
assistance in the preparation of the manuscript. Due thanks are expressed to Dr. C. 492
Hagen, University of Jena, Germany for the gift of the Pds-antibody and to Prof. Dr. P. 493
Kroth, University of Konstanz, Germany for the use of the Biolistic PDS-1000/He 494
system. 495
496
497
498
499
500
501
502
503
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

23
REFERENCES 504
505
1. Babinger, P., I. Kobl, W. Mages, and R. Schmitt. 2001. A link between DNA 506
methylation and epigenetic silencing in transgenic Volvox carteri. Nucleic. Acids. 507
Res. 29:1261-1271. 508
2. Baroli, I., and K.K. Niyogi. 2000. Molecular genetics of xanthophyll-dependent 509
photoprotection in green algae and plants. Phil.Trans. R. Soc. Lond. B 355:1385-510
1394. 511
3. Boussiba, S. 2000. Carotenogenesis in the green alga Haematococcus pluvialis: 512
cellular physiology and stress response. Physiol. Plant. 108:111-117. 513
4. Boussiba, S., L. Fan, and A. Vonshak. 1992. Enhancement and determination of 514
astaxanthin accumulation in green alga Haematococcus pluvialis. Methods 515
Enzymol. 213:386-391. 516
5. Breitenbach, J., C.F. Zhu, and G. Sandmann. 2001. Bleaching herbicide 517
norflurazon inhibits phytoene desaturase by competition with the cofactors. J. of 518
Agr. Food Chem. 49:5270-5272. 519
6. Böger, P., and G. Sandmann. 1990. Modern herbicides affecting typical plant 520
processes. In: Chemistry of Plant Protection, Vol 6: Bowers W.S., Ebing W., 521
Martin D., and R. Wegler (ed.) Springer Publ., Berlin, Heidelberg. 174-216. 522
7. Cerutti, H., A.M. Johnson, N.W. Gillham, and J.E. Boynton. 1997. A 523
eubacterial gene conferring spectinomycin resistance on Chlamydomonas 524
reinhardtii: integration into the nuclear genome and gene expression. Genetics 525
145:97-110. 526
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

24
8. Chamovitz, D., G. Sandmann, and J. Hirschberg. 1993. Molecular and 527
biochemical characterization of herbicide-resistant mutants of cyanobacteria 528
reveals that phytoene desaturation is a rate-limiting step in carotenoid 529
biosynthesis. J. Biol. Chem. 268:17348-17353. 530
9. Dixon, M. 1953. The determination of enzyme inhibitor constants. Biochem. J. 531
55: 170-171. 532
10. Eppler, K.S., L.C. Sander, R.G. Ziegler, S.A. Wise, and N.E. Craft. 1992. 533
Evaluation of reversed-phase liquid-chromatographic columns for recovery and 534
selectivity of selected carotenoids. J. Chromatogr. 20:89-101. 535
11. Fabregas, J., A. Dominguez, M. Regueiro, A. Maseda, and A. Otero. 2000. 536
Optimization of culture medium for the continuous cultivation of the microalga 537
Haematococcus pluvialis. Appl. Microbiol. Biotechnol. 53:530-535. 538
12. Grünewald, K., M. Eckert, J. Hirschberg, and C. Hagen. 2000. Phytoene 539
desaturase is localized exclusively in the chloroplast and up-regulated at the 540
mRNA level during accumulation of secondary carotenoids in Haematococcus 541
pluvialis (Volvocales, chlorophyceae). Plant Physiol. 122:1261-1268. 542
13. Guerin, M., M.E. Huntley, and M. Olaizola. 2003. Haematococcus astaxanthin: 543
applications for human health and nutrition. Trends Biotechnol. 21:210-216. 544
14. Hallmann, A., A. Rappel, and M. Sumper. 1997. Gene replacement by 545
homologous recombination in the multicellular green alga Volvox carteri. Proc. 546
Natl. Acad. Sci. USA 94:7469-7474. 547
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

25
15. Jakobiak, T., W. Mages, B. Scharf, P. Babinger, K. Stark, and R. Schmitt. 548
2004. The bacterial paromomycin resistance gene, aphH, as a dominant selectable 549
marker in Volvox carteri. Protist 155:381-393. 550
16. Johnson, E.A., and W.A. Schroeder. 1995. Microbial carotenoids. Adv. 551
Biochem. Eng. Biotechnol. 53:119-178. 552
17. Kajiwara., S., T. Kakizono, T. Saito, K. Kondo, T. Ohtani, N. Nishio, S. 553
Nagai, and N. Misawa. 1995. Isolation and functional identification of a novel 554
cDNA for astaxanthin biosynthesis from Haematococcus pluvialis, and 555
astaxanthin synthesis in Escherichia coli. Plant Mol. Biol. 29:343-352 556
18. Kobayashi, M., T. Kakizono, and S. Nagai. 1993. Enhanced carotenoid 557
biosynthesis by oxidative stress in acetate-induced cyst cells of a green unicellular 558
alga, Haematococcus pluvialis. Appl. Environ. Microbiol. 59:867-873. 559
19. Kovar, J.L., J. Zhang, R.P. Funke, and D. P. Weeks. 2002. Molecular analysis 560
of the acetolactate synthase gene of Chlamydomonas reinhardtii and development 561
of a genetically engineered gene as a dominant selectable marker for genetic 562
transformation. Plant Journal 29:109-117. 563
20. Lichtenthaler, H.K., and A. R. Wellburn. 1983. Determination of chlorophylls 564
a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1111:591-592. 565
21. Linden, H. 1999. Carotenoid hydroxylase from Haematococcus pluvialis: cDNA 566
sequence, regulation and functional complementation. Biochim. Biophys. Acta 567
1446:203-212. 568
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

26
22. Lorenz, R.T., and G. R. Cysewski. 2000. Commercial potential for 569
Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol. 570
18:160-167. 571
23. Lotan, T., and J. Hirschberg. 1995. Cloning and expression in Escherichia coli 572
of the gene encoding β-C-4-oxygenase, that converts β-carotene to the 573
ketocarotenoid canthaxanthin in Haematococcus pluvialis FEBS Letters 364:125-574
128. 575
24. Lumbreras, V., D. R. Stevens, and S. Purton. 1998. Efficient foreign gene 576
expression in Chlamydomonas reinhardtii mediated by an endogenous intron. 577
Plant J. 14:441-447. 578
25. Martinez-Ferez, I., A. Vioque, and G. Sandmann. 1994. Mutagenesis of an 579
amino acid responsible in phytoene desaturase from Synechocystis for binding of 580
the bleaching herbicide norflurazon. Pestic. Biochem. Physiol. 48:185-190. 581
26. Meng, C. X., C. Y.Teng, P. Jiang, S. Qin, and C. K. Tseng. 2005. Cloning and 582
characterization of beta-carotene ketolase gene promoter in Haematococcus 583
pluvialis. Acta Biochim. Biophys. Sin. Shanghai 37:270-275. 584
27. Misawa, N., K. Masamoto, T. Hori, T. Ohtani, P. Böger, and G. Sandmann. 585
1994. Expression of Erwinia phytoene desaturase gene not only confers multiple 586
resistance to herbicides interfering with carotenoid biosynthesis but also alters 587
xanthophyll metabolism in transgenic plants. Plant J. 6:481-489. 588
28. Misawa, N., Y. Satomi, K. Kondo, A. Yokoyama, S. Kajiwara, T. Saito, T. 589
Ohtani, and W. Miki. (1995) Structure and functional analysis of a marine 590
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

27
bacterial carotenoid biosynthesis gene cluster and astaxanthin biosynthetic 591
pathway proposed at the gene level. J. Bacteriol. 177:6575-6584. 592
29. Nelson, J. A., P. B. Savereide, and P. A. Lefebvre. 1994. The CRY1 gene in 593
Chlamydomonas reinhardtii: structure and use as a dominant selectable marker 594
for nuclear transformation. Mol. Cell Biol. 14:4011-4019. 595
30. Niyogi, K.K., O. Bjorkman, and A. R. Grossman. 1997. Chlamydomonas 596
xanthophyll cycle mutants identified by video imaging of chlorophyll 597
fluorescence quenching. Plant Cell 9:1369-1 380. 598
31. Randolph-Anderson, B. L., R. Sato, A. M. Johnson, E. H. Harris, C. R. 599
Hauser, K. Oeda, F. Ishige, S. Nishio, N. W. Gillham, and J. E. Boynton. 600
1998. Isolation and characterization of a mutant protoporphyrinogen oxidase gene 601
from Chlamydomonas reinhardtii conferring resistance to porphyric herbicides. 602
Plant Mol. Biol. 38:839-859. 603
32. Römer, S., P. D. Fraser, J. W. Kiano, C. A. Shipton, N. Misawa, W. Schuch, 604
and P. M. Bramley . 2000. Elevation of the provitamin A content of transgenic 605
tomato plants. Nature Biotechnol. 18:666-669. 606
33. Sandmann, G., and P. Böger. 1989. Inhibiton of carotenoid biosynthesis by 607
herbicides. In: Target sites of herbicide action. Böger P., and G. Sandmann (ed.) 608
CRC Press, Boca Raton, FL. 25-44 609
34. Schiedlmeier, B., R. Schmitt, W. Muller, M. M. Kirk, H. Gruber, W. Mages, 610
and D. L. Kirk. 1994. Nuclear transformation of Volvox carteri. Proc. Natl. 611
Acad. Sci. USA 91:5080-5084. 612
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

28
35. Simkin, A. J., J. Breitenbach, M. Kuntz, and G. Sandmann. 2000. In 613
vitro and in situ inhibition of carotenoid biosynthesis in Capsicum 614
annuum by bleaching herbicides. J. Agric. Food Chem. 48:4676-4680. 615
36. Sokolowsky, V., R. Kaldenhoff, M. Ricci, and V. E. A. Russo. 1990. Fast and 616
reliable mini-prep RNA extraction from Neurospora crassa. Fungal Genet. 617
Newslett. 37:41-43. 618
37. Steinbrenner, J., and H. Linden. 2001. Regulation of two carotenoid 619
biosynthesis genes coding for phytoene synthase and carotenoid hydroxylase 620
during stress-induced astaxanthin formation in the green alga Haematococcus 621
pluvialis. Plant Physiol. 125:810-817. 622
38. Steinbrenner, J., and H. Linden. 2003. Light induction of carotenoid 623
biosynthesis genes in the green alga Haematococcus pluvialis: regulation by 624
photosynthetic redox control. Plant Mol. Biol. 52:343-356. 625
39. Steiger, S., L. Schaefer, and G. Sandmann. 1999. High-light upregulation 626
of carotenoids and their antioxidative properties in the cyanobacterium 627
Synechocystis PCC 6803. J. Photochem. Photobiol. B 521:14-18. 628
40. Sun, Z., F. X. Cunningham, and E. Gantt. 1998. Differential expression of two 629
isopentenyl pyrophosphate isomerases and enhanced carotenoid accumulation in a 630
unicellular chlorophyte. Proc. Natl. Acad. Sci. USA 95:11482-11488. 631
41. Teng, C., S. Qin, J. Liu, D. Yu, C. Liang, and C. Tseng . 2002. Transient 632
expression of lacZ in bombarded unicellular green alga Haematococcus pluvialis. 633
J. Appl. Phycol. 14:497-500. 634
635
636
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

29
637
638
639
640
641
642
643
644
645
646
LEGENDS TO FIGURES 647
648
Fig. 1: Comparison of phytoene desaturase CrtP from Synechococcus sp. PCC 7942 649
including the four known amino acid exchanges and their resistance factors (RF) against 650
the bleaching herbicide norflurazon with the Pds from H. pluvialis (A). The ChloroP 651
program identified a chloroplast transit peptide (TP) at the N-treminus. The modified 652
codon in position 504 of the Pds from H. pluvialis, leading to an amino acid exchange 653
from leucine to arginine, corresponds to codon 436 in the CrtP from Synechococcus sp. 654
Intron and exon structure of the pds gene, the promoter sequence is indicated by the grey 655
filled box, the exon sequences by black filled boxes and the intron sequences by thin lines 656
(B). Map of H. pluvialis transformation vector pPlat-pds-L504R with indicated restriction 657
sites (C). The engineered pds gene (pds-L504R) is inserted into the NaeI site of the vector 658
pBluescript SK(-). The construct also contains the ampicillin resistance gene and the 659
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

30
Escherichia coli origin of replication (ColE1) and a multiple cloning site (MCS). The 660
sequence of this vector has been deposited at the Genebank (accession number 661
DQ404589). 662
663
Fig. 2: Dixon plot of the reciprocal in vitro phytoene to ζ-carotene conversion versus 664
increasing concentrations of the inhibitor norflurazon of a recombinant WT Pds (■) and 665
Pds Mut (▲) of H. pluvialis. The ki values for norflurazon inhibition were calculated 666
from the intercepts with the x-axis to be 0.14 µM and 5.98 µM for the heterologous 667
expressed Pds WT and Pds Mut, respectively. RF, Resistance factor. 668
669
Fig. 3: Southern blot analysis of H. pluvialis transformants P1-P11 and H. pluvialis wild 670
type (WT). DNA from transformants and WT was digested with XbaI and XhoI and 671
electrophoresed on a 0.8% agarose gel. For detection, a pds-specific cDNA probe (A) or 672
a specific probe against the ampicillin resistance cassette (B) was used. The arrow marks 673
the 5.78 kb XbaI/XhoI fragment containing endogenous pds gene. Numbers on the left 674
correspond to labeled λHindIII fragments used as size markers (M). 675
676
Fig. 4: Transcript analysis of phytoene desaturase gene (pds) and carotenoid hydroxylase 677
gene (bhy) in wild type H. pluvialis cells after 0, 18 and 36 h of high light and in 678
transformants P1-P11 under low light conditions (A). For comparison, total RNA (rRNA) 679
was stained with ethidium bromide. Western analysis of phytoene desaturase protein 680
(Pds) levels (B). 681
682
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

31
683
Fig. 5: Astaxanthin formation after exposition of H. pluvialis WT (■) and the 684
transformants P3 (●) and P11 (▲) to continuous high light. 685
686
Fig. 6: Kinetics of non-photochemical quenching (NPQ) of dark-adapted H. pluvialis 687
wild type (WT) (■) and the transformants P3 (●) and P11 (▲). 688
689
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from

32
Table 1 690
Pigment content of green Haematococcus pluvialis cells treated for 1h with strong light 691
692
WT P3 P11 693
---------------------------------------------------------------------------------------------- 694
Total carotenoids (mg/g dw)a 2.35+0.24 2.65+0.30 3.00+0.28 695
Chlorophylls (mg/g dw)a 31.72+2.41 26.28+3.47 31.32+2.40 696
697
Car/Chl. (%) 7.4 10 9.6 698
699
Carotenoid composition (µg/g dw) 700
Neoxanthin 37+6 36+5 18+4 701
Violaxanthin 79+1 76+2 65+7 702
Antheraxanthin 91+4 90+6 55+3 703
Lutein 1464+97 1596+36 1920+99 704
Zeaxanthin 142+18 202+20 214+11 705
ß-Carotene 532+46 648+30 728+84 706
707
V+A+Zb 312 368 334 708
Z/(V+A+Z) 0.46 0.55 0.64 709
---------------------------------------------------------------------------------------------- 710
asimilar total carotenoid and chlorophyll concentrations prior to high light illumination; 711
bV+A+Z is the sum of the xanthophyll cycle carotenoids violaxanthin, antheraxanthin 712
and zeaxanthin. 713
714
715
ACCEPTED on July 6, 2018 by guest
http://aem.asm
.org/D
ownloaded from


















