Advisor: Dr. Stephen D. Liberles Erika Kristen Williams
57
Neural Mechanisms of Mechanosensation Within the Body The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Williams, Erika. 2018. Neural Mechanisms of Mechanosensation Within the Body. Doctoral dissertation, Harvard Medical School. Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:36923341 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA
Transcript of Advisor: Dr. Stephen D. Liberles Erika Kristen Williams

Neural Mechanisms ofMechanosensation Within the Body
The Harvard community has made thisarticle openly available. Please share howthis access benefits you. Your story matters
Citation Williams, Erika. 2018. Neural Mechanisms of MechanosensationWithin the Body. Doctoral dissertation, Harvard Medical School.
Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:36923341
Terms of Use This article was downloaded from Harvard University’s DASHrepository, and is made available under the terms and conditionsapplicable to Other Posted Material, as set forth at http://nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of-use#LAA


iii
Advisor:Dr.StephenD.Liberles ErikaKristenWilliams
Neuralmechanismsofmechanosensationwithinthebody
Abstract
Theabilitytodetectmechanicalforcesplaysacriticalroleinorganismbehaviorand
physiology.Oneofthefundamentalmeansbywhichweinteractwithourenvironmentis
throughtouch,whichincludestheabilitytosensemechanicaleventssuchaspressure,
impact,vibration,andchangesinjointposition.Similarly,oneofthefundamentalcues
usedbyinternalorgansystemstoregulatebehaviorandphysiologicalresponsesis
mechanicalforcewithinthebody.Sensorysystemsintheintestinaltractdetectstretchas
theseorgansfillwithandmovefood,playingapowerfulroleinthemodulationofeating
behavior.Inaddition,sensorysystemsalsomonitortheexpansionandrelaxationofthe
lungsduringbreathingtoregulaterespiration.Similarly,accuratemonitoringofpressure
withinthevascularsystemplaysakeyroleinregulationofcardiovascularfunction.Eating,
breathing,andbloodcirculationconstitutebasicneeds,yetourunderstandingofthe
sensoryneurobiologyincontrolofthesefunctionsislimited.Todate,themolecular
mechanosensorsrequiredremainunknown.However,thediscoveryofthemammalian
mechanosensorPiezo2raisestheinterestingpossibilitythatthismoleculeisnotonly
involvedindetectionofexternalmechanicalcuesinourskin,butmayalsosub-serve
detectionofmechanicalcueswithintheinternalorgansofthebody.

iv
TableofContentsIntroduction...........................................................................................................................................7Mechanosensationininternalorgans.....................................................................................................7ThemammalianmechanosensorPiezo2..............................................................................................25Recordingmechanosensitiveafferentsinthevagus........................................................................30
Experiment1:InVivoimagingofPiezo-2lineageneurons................................................35Introduction...................................................................................................................................................35Methods............................................................................................................................................................35Results..............................................................................................................................................................35Conclusions.....................................................................................................................................................39
Experiment2:InvivoimagingofneuronsthatexpressPiezo2intheadult.................40Introduction...................................................................................................................................................40Methods............................................................................................................................................................40Results..............................................................................................................................................................41Conclusions.....................................................................................................................................................43
Experiment3:DetectionofstretchstimuliinPiezo2knock-outganglia.......................44Introduction...................................................................................................................................................44Methods............................................................................................................................................................44Results..............................................................................................................................................................45Conclusions.....................................................................................................................................................48
Summary..............................................................................................................................................50Bibliography........................................................................................................................................50

v
Glossary
AAV–adeno-associatedvirus
DRG–dorsalrootganglion
GLP1R–GLP1receptor
GPR65–G-proteincoupledreceptor65
IGLE–intraganglioniclaminarending
IMA–intramusculararray
RAR–rapidly-adaptingstretchreceptor
SAR–slowly-adaptingstretchreceptor
siRNA–smallinterferingribonucleicacid

vi
Acknowledgements
Thankyoutomyfamily,theLiberlesLabpastmembersandpresent,theProgramin
Neuroscience,MDPhDProgram,andHSTProgramfortheconstantsupportand
inspiration.Thankyoutosomanyfortechnicalhelpandexpertise,andmostnotablythe
HarvardAnimalFacilities;withoutyounoneofthisworkwouldbepossible.Financial
supportforthisworkwasthroughF30CA177170andNIHMSTP-T32GM007754.

7
Introduction
Mechanosensationininternalorgans Internalorgansareinconstantmotion,andsimilarlyconstantlysubjecttoavariety
ofmechanicalforcesofcriticalphysiologicalsignificance.Eatingresultsinaccumulationof
foodinthestomachandmechanicaldistensionofthestomachwall.Neuraldetectionof
stomachstretchplaysacriticalroleinfeedingbehavior.Inaddition,digestioninvolvesthe
mechanicalpropulsionoffoodthroughsphinctersandoverapproximately25feetofadult
humanintestine,aprocesssimilarlymonitoredandregulatedbythenervoussystem.
Breathingischaracterizedbythemechanicalexpansionandrelaxationofthechestcavity
togeneratechangesinpressure,andsubsequentflowofairintoandoutofthelungs.
Sensoryneuronsmonitorrespiratorymovementtoregulatebreathfrequencyanddepth,
importantvariablesforadequateoxygenation,carbondioxideexcretion,andprotectionof
lungtissue.Lastly,thebeatingheartpumpsbloodandgeneratespulsatileflowand
pressurewithinthevasculature.Neuralmonitoringofbloodpressureinitiatesreflex
regulationofheartrateandsubsequentcardiovascularvariablestopreserveadequate
tissueperfusion.
Inthissection,wewillreviewmechanosensationindigestion,respiration,and
circulation,withafocusontransmissionofinformationfromperipheralorganstothe
brainstem.Withineachorgansystem,wewilldiscussphysiologicallyrelevantautonomic
reflexes,neuralresponseproperties,andsensoryneuronanatomy.Oneoftheprimary
neurallinksfrominternalorganstothebrainisthevagusnerve.Therefore,thevagus
nervewillfigureprominentlyinexperimentsanddiscussions,soabriefoverviewofits
generalanatomyandorganizationisprovided.

8
TheVagusNerve Thevagusnerveisoneofthemajorbody-to-brainneuralconnections,andmediates
afferentsensoryinformationtransmissionintothebrainstem,aswellasefferentmotor
commandtransmissionouttotargetorgans.Eightypercentofvagalfibersaresensory1,
andeachsensoryneuronextendsoneprojectionintotheperipherytocollectinformation
fromatargetorgan,andasecondprojectionintothebrainstemtotransmittheinformation
tocentralneuralcircuits.Thesensoryneuroncellbodiesresideinganglia,thenodoseand
jugular,locatedadjacenttothebaseoftheskull.Allsensoryneuronsusethe
neurotransmitterglutamatetocommunicatetocentralcircuits.Theremaining20%of
vagalfibersaremotorfibers1.Thecellbodiesofvagalmotorneuronsarelocatedwithin
nucleiinthebrainstem,andsendprojectionsoutfromthebraintoperipheraltarget
organs.Theneurotransmitterusedbythemotorneuronsisacetylcholine.Therefore,the
vagusnervecanbeconsideredananatomicalhighwaysharedbyasensorysystemcarrying
informationcentrally,andamotorsystemcarryingcommandsperipherally.Thesetwo
systemscommunicateclosely;thesensoryterminalsinthebrainstemareimmediately
adjacenttothenucleicontainingthemotorneuroncellbodies,andtheseneuronsare
knowntomakebothdirectandmulti-neuronnetworkconnectionstoeachothertoprovide
reflex-likecontrolofinternalorganstates2-4.Forthepurposesofareviewonperipheral
mechanosensation,wewillfocusontheresponseproperties,anatomy,andphysiological
rolesofvagalsensoryneurons.
Gastrointestinalstretch
Physiology
Mechanicaldistensionofthestomachplaysacriticalroleinregulationoffood
intake.Notonlydoesthisintuitivelycoincidewithnearlyuniversalhumanexperience
followingmeals,butalsocarefulexperimentationover60yearsagohaselaboratedonthe
physiologicalpowerandspecificsensorycuesrelevantforregulationofmealconsumption.
Theearliestexperimentstodemonstratetheimportanceofgastrointestinal
mechanosensationonfeedingbehaviorwereperformedindogswithsurgicallyaltered

9
uppergastrointestinaltracts5,6.Gastricfistulasallowedforartificialfillingofthestomach,
whileesophagostomyallowedforartificialdiversionoforallyingestedfoodoutofthe
esophagusandawayfromthestomach,dissociatingoralingestionandchangesinstomach
content.Fillingthestomachviaafistulatoavolumeequivalenttoapproximately40%ofa
normalmealsizejustpriortomealtimedecreasedoralconsumptionbyanequivalent(30-
50%)amount.Thedegreeofintakeinhibitionwasindependentofthecaloricvalueofthe
stomachfiller;intakeinhibitionwasthesameifthestomachwasfilledwithfood,or
caloricallyinertKarayagum,suggestingtherelevantsensorycuewasthemechanical
distensionandnotthechemicalfoodcontent5.Intriguingly,iffoodwasinfusedintothe
stomachatatimeotherthanduringeating,ithadnoimpactonshamfeeding,suggesting
thetemporalrelationshipbetweeningestionandstomachdistensioniscriticalforan
effect6.
Conversely,whenthefoodwasdivertedoutoftheesophagusandpreventedfrom
reachingthestomach,animalsfedforlonger.Furthermore,indiversioncasesinwhichthe
stomachwasdistendedeitherbyreplacementoffoodorbyinflationofarubberlatex
balloon,thiseffectwasreversedandanimalsingestedless5.Fromtheseexperiments,one
canconclude:1)Stomachdistensionisnotnecessarytoterminatefeedingbecauseanimals
willstopfeedingeveniffooddoesnotreachthestomach.2)Stomachdistensiondoes,
however,playanimportantroleinregulatingtheamountoffoodingestedbecausewithout
stomachdistension,animalseatmore,andwithitanimalseatless.3)Themechanical
distensionofthestomach,notthecaloriccontent,istherelevantgastriccueforacute
regulationofmealsize,andthiscuemustoccuratthesametimeasoralingestiontoimpact
acutefeedingbehavior.
Subsequentstudiesinrodentshaveconfirmedthesefindings,andalsodelineated
theeffectsofstomach-derivedversusintestine-derivedpost-ingestioncuesoneating
behavior.Inoneelegantseriesofexperiments7,8,ratsweresurgicallyimplantedwitha
pyloricsphinctercuffandagastriccatheter.Inflationofthepyloriccuffpreventedtransitof
stomachcontentstothesmallintestine,whileinfusionthroughthegastriccatheterallowed
forexperimenter-controlleddeliveryofgastriccontents.Theseexperimentsdemonstrated:
1)asindogs,infusionsinthestomachinhibitedfoodintakeindependentofthecaloric
contentoftheinfusedsolution,and2)incontrast,caloriccontentdidimpactthedegreeof

10
feedingsuppressionwhenthepyloriccuffwasleftopenandthecontentscouldpassinto
thesmallintestine7.Boththesefindingsareconsistentwithmechanicallymediated
inhibitionoffoodintakebythestomach,andchemicallymediatedinhibitionoffoodintake
bytheintestine.
Theinhibitionoffoodintakebygastricmechanosensationandintestinal
chemosensationisthoughttobedependentonintactvagalfibers.Animalsconsumelarger
mealsizesaftersurgeriesinwhichvagalinnervationofthestomachisdisrupted,and
infusionofvolumeintothestomachnolongersuppressesfoodintakeinvagotomized
animals9,10.Similarly,thesuppressionofmealsizebychemicalcuesintheintestineis
dependentonanintactvagusnerve11,12,thoughitshouldbenotednotallstudieshave
foundthiseffect13,14.Despitetheshortcomingsofnon-selectivevagotomyexperiments,
theseresultssuggestakeyroleforvagalafferentsinregulationoffeeding,and
demonstrateacleardistinctionbetweenvagalgastricmechanosensorsandintestinal
chemoreceptors.
Theexperimentsinanimalsaredirectlyrelevanttohumans.Surgicalmethodsto
induceweightlossinpeoplebearresemblancetoexperimentalmanipulationsofstomach
distension.Acentralgoalofbariatricsurgeryistoreducestomachcapacitysuchthatthe
sameamountoffoodwillcausegreaterdistension.Reductionofgastriccapacitycanbe
achievedviaseveraldifferentsurgicalalternatives,includingimplantationofavolume-
occupyingballoon(gastricballoon),applicationofaconstrictiveband(banding),stapling
ofthestomachtoreduceitseffectivevolume(verticalbandedgastroplasty),andstapling
andresectionmostofthestomachpouchtoleavearesidualtube-likegastricsleeve(sleeve
gastrectomy)15.Implantationofaballoontypicallyfilledtoavolumeof750-900milliliters
inthestomachofhumanpatientshasbeenshowntoimproveweightlosswhencompared
tonon-operativeinterventions16,17.Similarly,bandingandgastroplastyresultin~15%
weightlossmaintainedover>10years,whereascontrolsubjectsonlylost~2%;these
resultswereaccompaniedbyadecreaseinlong-termmortalitylikelysecondarytothe
reductionincardiovascularriskfactorsanddiabetesassociatedwithobesity18-20.Themost
effective,‘gold-standard’weightlosssurgeryisthegastricbypass,inwhichonlyavery
smallresidualstomachisconnectedtothedistalsmallintestinetobothreducethestomach
volumeandtheabsorptivecapabilitiesoftheintestine20,thoughsleevegastrectomyalone

11
isnearlyaseffectiveinachievingweightloss15.Inshort,surgicalreductionofthestomach
volumeisaneffectiveweightlosstoolinthefaceofthegrowingobesityepidemic,aneffect
likelymediatedinlargepartthroughmanipulationofvagalgastrointestinal
mechanosensation.
Responseproperties
Theimpactofmechanicalstomachdistensiononfeedingpromptedinvestigationof
theoperativesensorymechanisms.Onlyafewyearsfollowingtheinitialcaninephysiology
experimentsdescribedabove,Paintal,Iggo,andcolleaguesdescribedthefirst
electrophysiologicalrecordingsofvagalafferentsinresponsetodistensionofthestomach21,22.Ininitialexperiments,thecervicalvagustrunkwasseriallysub-dividedintofiber
strands,andresponsesineachstrandwererecordedwhilealatexballoonwasinflatedin
thestomach.Usingthismethod,gastricstretch-responsiveunitscouldbeisolatedfrom
unitsresponsivetoheartbeatsortherespiratorycycle,confirmingtheexistenceofa
uniquestomachstretchsensoryneuronclass.Furthermore,theendingsofthesefibers
werethoughttobelikelylocatedinthemuscularstomachwallbecausetheiractivitycould
beevokedbydigitalcompressionofthestomach,butnotbystrokingthemucosalor
peritonealgastricsurfaces21,22.
Gastricmechanoreceptorshaveconsistentresponseproperties.Allfibersresponded
todistensionquickly,exhibitedfiringratesof25-60impulses/secondduringthe
distension,andreturnedtobaselinelevelsofactivityimmediatelyfollowingreliefofthe
distension.Allbutoneofthefiberstestedwereeithernon-adaptingorslowlyadapting,
withlittlechangeinthefiringrateevenduringminuteormulti-minutelongdistensions.
Step-wiseincreasesinthevolumeofdistensiongeneratedalinearincreaseinthefiring
rate,thoughthethresholdofdistensionrequiredforfiberactivationvariedwidely21.
Furthermore,inductionofmuscularcontractionsofportionsofthestomachwallinwhich
thereceptiveendsofthesefiberswerefoundwouldelicitactivityinthegastricreceptors
thatwasevenmorerobustthanthatcausedbypassivedistension22.Therefore,the
relevantcueforfiberactivationisnotintra-gastricpressureperse,butthetensionwithin
themuscularwallofthestomach.Gastricstretchreceptorsarethereforeconsidered‘in-

12
series’tensionreceptors,andmonitornotonlypassivetensionofthestomachwallbutalso
activetensiongeneratedbymuscularcontractioninphysiologicallyrelevantranges.A
corollarytotheobservationthatgastricmechanoreceptorsarein-seriesisthattheslow
adaptationobservedintensionreceptorfiringratesmayatleastinpartreflectmuscular
relaxationofthestomachwall,andmaynotbeanintrinsicpropertyofthe
mechanoreceptor.Thesamepropertieswerefoundfordistension-sensitivevagalafferents
thatterminatedintheesophagusorthesmallintestine23.Similarpropertieshavebeen
subsequentlydescribedingastricmechanoreceptorsinmultiplestudies24-27.
Subsequentstudiessoughttoidentifythemolecularmechanismbywhich
mechanicalforcesaretransducedintoaneuralsignalwithinvagalgastrointestinal
mechanoreceptors.However,despitedecadesofrecordings,itisunclearwhethervagal
afferentsthemselvesaredirectlymechanosensitive,orwhethertheyaretransmitting
informationfrommechanosensitiveentericneuronsornon-neuronalgastrointestinalcells.
Threeargumentshavebeencitedtosupporttheideathatvagalafferentsaredirectly
mechanosensitive28.First,thelatencyofresponsefollowingspecificandrapidmechanical
deformationofthemuscularwalloftheesophagusis<6milliseconds,adelayhasbeen
citedastoorapidforamechanismotherthandirectmechanosensation.However,thedelay
betweenfiringofanupstreamneuronanditschemically-coupleddownstreamtargethas
beencalculatedat2millisecondsinthemammalianbrain,suggesting6millisecondsis
morethansufficientforsynaptictransmission,andcastingdoubtonthisclaim29.Second,
extracellularcalciumisconsideredanecessarycomponentforvesiculartransmitter
release,andvagalafferentmechanosensitivityispreservedbothwhenextracellular
calciumisremoved,andinthepresenceofthecalcium-channelblockerCd2+.However,
calcium-dependenttransmissionisnottheonlymeansofcell-cellcommunication;gap
junctionsforexamplewouldallowforrapidandextracellularcalciumindependentsignal
transduction30.Finally,pharmacologicinhibitionofcandidateneurotransmitterreceptors
(e.g.glutamatereceptors,purinergicreceptors)hasnoimpactonvagalmechanosensation,
evidencetakentoexcludearolefortheseputativesecond-messengers,thoughofcourse
thisrepresentsadrasticallyincompletesurveyofpotentialcell-cellsignalingmechanisms.
Insummary,thesedataarenotconclusive,leavingmuchambiguityaboutthetruesiteor

13
sitesofmechanosensationinthegastricdistensionsensorypathway.Themolecular
mechanosensor,unsurprisinglyinthiscontext,remainsunknown.
Anatomy
Despiteambiguityaboutthemechanismofmechanotransduction,themorphology
ofdistension-responsivevagalafferentsinthegastrointestinaltractisknown.Isolated
esophagealandstomachpreparationshaveallowedforlocalizationofmechanically
sensitive‘hotspots’,andconcurrentlabelingofvagalfibersinnervatingthosesites31,32.
Hot-spotsintheesophagusandthestomachareenrichedandlocalizedaround
intraganglioniclaminarendings(IGLEs),avagalterminaltypecloselyassociatedwith
myentericneuronganglialocatedbetweenthecircularandlongitudinalmusclelayers.
Consistentwithhot-spotstudies,geneticallylabeleddistension-sensitiveafferents,marked
byexpressionoftheG-protein-coupledreceptorGLP1R,formdenseIGLEterminals
throughoutthegastrointestinaltractandterminatecentrallywithinthemedialnucleusof
thesolitarytract33.
AsmallnumberofstudieshaveexaminedthestructureandlocationsofIGLEsin
hopesofprovidingcluesastothesitesofmechano-transduction.IGLEswereinitially
describedusingsilverstainingmethodsintheesophagus34aslamellatedendings
surroundingandenteringthecapsuleofcollectionsofentericneuroncellbodies.Electron
microscopyofvagalendingsinthesemyentericgangliashowedcloseapproximationof
mitochondria-richIGLEterminalswiththeganglionneuropilandcellbodies,butdidnot
revealaclassicsynapticultrastructureassociatedwiththesecontacts35.Theclose
associationbetweenIGLEsandentericgangliacouldsupportboththeideathatvagal
neuronsreceivesignalsfromprimarilymechanosensitiveentericcells,orthatenteric
gangliacouldprovidecriticalsupporttoprimarilymechanosensitivevagalneurons.
IGLEscanbefoundintheesophagus,stomachandproximalsmallintestine,
consistentwiththevagalresponsepropertiesidentifiedinallthesesites.Thegreatest
densityofIGLEsislocatedinthestomach,withdecreasingdensityalongtheproximal
intestinallength36-39.Someestimatessuggesttherearewellover1000individualIGLE
terminalsinthestomachwall,andthatthoughthedensityofinnervationinthesmall

14
intestineislower,thegreatersizeofthatorganwouldmeanalargerabsolutenumberof
intestinalIGLEsthaninthestomach38.ThehighIGLEdensityinthestomachisconsistent
withthephysiologicalimportanceofstomachstretchonfeedingbehavior.Inaddition,the
highnumberofIGLEsinothersitessuggeststhisterminaltypealsotransducesimportant
mechanicalinformationfromgastrointestinalsitesoutsidethestomach.
IGLEs,notably,arenottheonlyvagalterminaltypeinthemuscularwallofthe
gastrointestinaltract.Thesecondterminaltype,namedintra-musculararrays(IMAs),are
notnearlyaswidelydistributedasIGLEsbutratherseemtoterminatearoundthegastro-
esophagealjunction,gastro-duodenalsphincter,andthejunctionbetweentheproximal
andmiddlesegmentsofthecolon36.IMAscoursebetweenthelongitudinalsmoothmuscle
layers,forminglongbranchingparalleltractsthatformsynapse-likecontactswith
intrinsicallyoscillatory,gastriccontractionpace-makingcellstermedinterstitialcellsof
Cajal40.GiventheanatomicalassociationwithICCsandsphincters,IMA-formingneurons
arealsopositedtobemechanosensors,butdefinitiveevidencefortheirresponse
propertiesandphysiologicalroleremainselusive.
Summary
Mechanicaldistensionofthestomachregulatesmealsizeduringfeeding.Anintact
vagusnerveisrequiredforthedetectionofstomachdistensionandreductionofmealsize.
Aselectsubsetofvagalafferentsrespondstostomachstretchwithslowlyadapting
increasesinfiringrateinresponsetobothpassivedistensionandmuscularcontraction.
DistensionsensitivevagalafferentsformIGLE-typeterminalendingsintheesophagus,
stomachandintestinemusclewalls.However,keyquestionsremainabouttheidentityof
therelevantmechanosensorymolecularapparatus,theprimarysiteofmechanosensation,
andthecontributionsofnon-IGLEstomachmuscle-innervatingvagalsensoryafferents.

15
Lungstretch
Physiology
Thephysiologicalcontrolofrespirationhasbeenlinkedtothevagusnervesincethe
inceptionoftherespiratoryphysiologyfield.Earlyrespiratoryphysiologistsnotedthat
electricalstimulationofthetransectedvagusnervetrunkcouldinduceeitherexpiratoryor
inspiratoryeffort41.However,resultsweremixedandthephysiologicalrelevanceofthese
observationswasnotclarifieduntiltheworkofHeringandBreuerin1868,whenthe
artificialmanipulationofnerveswasabandonedinfavorofdescriptionofthephysiological
responsestoinflationanddeflationofthelungs42.
Breuernotedthatwhenthelungwasinflated,furtherinspiratorymovementswere
inhibited,andexpirationwaspromoted.Inspiratoryinhibitionoccurreddespite
developmentofprofoundhypoxiaandindependentlyofthechemicalcompositionofthe
gasusedtoinflatethelungs,suggestingtherelevantcuewaslesslikelychemical,andmore
likelymechanical.Thedegreeofinspiratoryinhibitionwasdirectlycorrelatedtothedegree
oflungexpansion.Thereflexalsoseemedtobephysiologicallyrelevantbecauseitcouldbe
observedbothifthelungwereartificiallyinflatedbeyondnormalphysiologicalvolumes,
andalsoifthetracheaweresimplyoccludedatthepeakofnaturalinspiration.Therelevant
stimuluswasthoughttobetheexpansionofthelungitselffortworeasons:1)Expansionof
collapsedlungsalsoblockedinspiratoryeffort,meaningthereflexwasintactevenwhen
intra-thoracicpressureswereabnormallylowandchestwallmovementscouldnot
contribute.2)Introductionofelevatedthoracicpressurethroughatubeinthesideofthe
chestdidnotimpactinspiratoryeffortaslongasthestateofexpansionofthelungswas
heldconstant,meaningthecriticalcuewasthemechanicalmovementofthelungandnot
theexternalpressuretowhichthelungwassubject.Thisreflexisclassicallyreferredtoas
theHering-Breuerinspiratoryreflex,andcanbeabolishedbycuttingbothvagusnerve
trunksintheneck42.
SubsequentphysiologicalstudieshaverevealedintriguingfeaturesoftheHering-
Breuerreflexthatcomplicatecurrentunderstandingofitsphysiologicalrole.First,a
similarreflexwasalsoinitiallydescribedinwhichholdingthelungsinexhalationwould
preventfurtherexhalation42;afindingthatsparkedmanydecadesofdebateaboutwhether

16
thesameneuralpathwayscouldunderlieboththeinspiratoryandexpiratoryreflexes43,44.
Second,aseriesofexperimentswereundertakentoinvestigatetherelevanceofthereflex
inman,whichrevealedfourintriguingfindings:1)inspiratoryinhibitiontolunginflation
wasobservedininfantsupuntil3-4daysoflife,duringwhichtimethereflexstrength
decreasedandultimatelywaslost45.2)Thereflexcouldonlybeobservedinadultsunder
anesthesiaor,inonesubject,unconsciousfromaheadinjury46,butnotinsleepingadults47.
3)Surgicalinstillationofanerve-blockingagenttothevagustrunkcouldblocktheHering-
Breuerreflexinanesthetizedpatients46.4)Incontrast,inawakesubjects,nerveblock
injectionsaroundtheninthandtenthcranialnervesatthebaseoftheskullresultedinno
changeinbreathing,thoughtheinjectionsintheseexperimentscausedprofound
hypertension(presumablysecondarytobaroreceptorblockade),andinsensitivityto
hypoxia(presumablysecondarytochemoreceptorblockade)48,49.Thesedatatogether
suggestthatthestrengthoftheHering-Breuerreflexchangeswithdevelopment,andis
influencedbysubjects’stateofconsciousness.Thequestionsraisedabouttheroleofthe
reflexinawake,normalhumansremainunanswered.
LunginflationinducesotherreflexeffectsbeyondtheHering-Breuerinhibitionof
inspiration.Twootherprominentreflexeffectsoccurwithlunginflation.First,
physiologicalinspirationcausesanincreaseinheartrate50.However,carefulexperiments
inwhichlungdistensionwasisolatedfromchangesinbloodflowinthecardiopulmonary
systemandfromchemicalchangesinthebloodsecondarytorespirationsuggestedthe
relevantcueinthisreflexisaugmentedcardiacvenousreturn,tobediscussedin
subsequentsections51.Secondly,inspirationalsotriggersareflexdecreaseofsystemic
vascularresistance52.Intheseexperiments,expansionofthelungsbyinjecting
physiologicallyrelevantvolumesofairresultedinadose-dependentsystemicvasodilation,
whilecollapseofthelungsresultedinsystemicvasoconstriction.LikeintheHering-Breuer
reflex,thereductioninsystemicvascularresistancewasindependentofthecomposition
andtemperatureofthegasusedtoinflatethelungs.Theeffectwasalsomaintainedwhen
thepressuresinthecarotidarteriesandtheaortawereheldconstant.Cuttingpulmonary
nervesandchemicallyinhibitingthesympatheticnervoussystematthestellateganglionin
theneckcouldbothabolishthereflex,suggestingitismediatedbycommunicationbetween
thevagusandsympatheticfibers.However,thesamecareinisolatingpulmonaryversus

17
cardiovascularcontributionstothisreflexwasnottakenasinotherexperiments,so
specificconclusionscannotbedrawn53.
Responseproperties
Asinthegastrointestinalstretchfield,thedescriptionoftheHering-Breuerreflex
drovethesearchfortheoperativeneuronalelements,andtheirdiscoveryonlyawaitedthe
developmentofelectrophysiologyequipmentandmethodssensitiveenoughtorecordfrom
singlenervefibers.Adrianwasthefirsttoidentifyindividualpulmonarystretch-sensitive
vagalafferentsin193354.ThepulmonarymechanoreceptorsAdrianidentifiedgenerate
largecurrentsinfast-conductingA-fibers,explainingtheiridentificationover10years
priortotheirmoreelusiveC-fibermediatedgastricmechanoreceptorcounterparts.Work
followingthisinitialdiscoveryidentifiedtwoclassesofmechanosensitivepulmonary
afferents,distinguishedbytheiradaptationrateandthresholdofactivation55.
Slowly-adaptingstretchreceptors(SARs),definedashavingadeclineoffiringrate
oflessthan55%withinthefirsttwosecondsofastretchstimulus,accountfor
approximately50%ofallpulmonaryinflation-sensitivevagalafferents55.SARsarenot
onlyactivatedbyexperimentalinflation,butalsofirecyclicallywiththerespiratorycycle,
canbeactivatedbyforcedlungdeflation,andarealsoactivatedbycontractionofsmooth
musclesurroundingtheairways56.MeasurementoftrachealpressurethresholdsofSAR
activationsuggeststhatatleasthalfofallSARsareactiveduringtherestingrespiratory
cycle57.Intriguingly,inhaledcarbondioxideinhibitsSARstretchsensitivity58,aneffect
mediatedwithinthelungbecauseCO2inhibitsSARsensitivitywithoutchangesinarterial
carbondioxidelevels59,60.However,otherthantheinhibitoryeffectofCO2,SARsare
notablyinsensitivetootherchemicalstimuli61,andtheoxygencontentoftheinhaledor
infusedgasdoesnotimpacttheresponsepropertiesofSARs54.SARsthereforehavea
mechano-specificreceptivefieldtorivaltheirsimilarlyslowlyadaptinggastricstretch
sensitivecousins.ThesepropertiesmakeSARsidealcandidatestomediatetheHering-
Breuerinspiratoryinhibitionreflex.
Incontrast,rapidly-adaptingstretchreceptors(RARs)arealsoactivatedbylung

18
inflation,butaredefinedashavingfiringratesthatdecrementbyover80%withinthefirst
twosecondsofastretchstimulus55.RARsaccountfor40%ofstretch-sensitivepulmonary
afferents,thoughthisvarieswithanimalmodelinvestigated62.RARsexhibitamarkedly
higherthresholdforactivationcomparedtoSARs;nearlynoRARsareactiveduring
breathingatrest,thoughthemajoritywouldbeactivatedatpressuresexhibitedduring
respiratoryeventssuchascoughing57.Inadditiontothesefeatures,RARsarealsosensitive
toanumberofirritatingchemicalmediatorsincludingpollutantssuchasozoneandwood
smoke,adiversityofchemicals,pulmonarycongestion,andintroductionofhypo-osmotic
solutionswithintheairways,earningthemthename‘irritantreceptors’63-67.Thismixed
high-thresholdmechanicalandchemicalreceptivefieldsuggestsfunctionaloverlapwith
lungafferentsthatarepurelychemo-sensitivewithoutanymechanicallysensitiveresponse
propertycomponent61.Forthesereasons,RARsaresuspectedtounderliereflexresponses
ofhyperpneaandbronchoconstrictiontooffensiveinhaledchemicalagents.Inaddition,
RARscanbesensitized,suggestingaroleinreactiveairwaydisease63.
WhileSARsandRARsareconsideredbroadcategoriesofmechanosensitivelung
afferents,detailedrecordingsrevealmanyfiberswithpropertiesintermediatebetween
SARsandRARsthataccountfortheremaining10%ofstretch-sensitiveafferentsperthe
SARandRARdefinitionsprovidedabove55,57,68.Therefore,itmaybemoreaccurateto
characterizefibersasbelongingtoaspectrumofadaptationandchemicalsensitivityrather
thantotwofullydiscreteneurongroups.However,withoutameanstoselectivelyand
specificallymanipulateRARsvs.SARs,theinnervationpatternsandphysiologicalrolesof
eachstretch-sensitivefibertyperemainsunclear.
Recentexperimentshavesuggestedthemolecularidentityofalung
mechanosensor69,andthesedatawillbepresentedindetailwithinthediscussionofthe
discoveryandelaborationofthemammalianmechanosensorPiezo2.
Anatomy
Vagalafferentstravelalongthemajorairwaysandprovideterminalsatalllevelsof
therespiratorytree,fromthetracheatothealveoli.TheanatomicallocationsofSARand

19
RARterminalshavebeeninferredfromfunctionalstudiesofterminalresponseproperties,
andtheirdetailedterminalanatomyinferredbasedonthestructuresofvagalafferents
foundatthesevariousanatomicalsites.
SARterminalsarethoughttobelocatedinthesmoothmusclesurroundingairways63.Threelinesofevidencesupportthisconclusion.First,SARsrespondtodirectmechanical
probingoftheairwaysmoothmuscle,aswellastoairwaysmoothmusclecontraction56,70.
Second,SARscontinuetofunctionafterthemucosaisstrippedfromisolatedexperimental
preparations,butceasetofunctionafterdisruptionofthesmoothmuscle56.Finally,SARs
arenotinhibitedintra-airwayanesthetics,supportingtheconclusionthattheirterminals
areisolatedfromtheairwaylumen56.Myelinatedvagalafferentterminalsinpulmonary
smoothmuscle,inferredfromthepropertieslistedabovetobeSARs,havebeen
characterizedwithbothlightanelectronmicroscopy.Asinothermusclemechanoreceptor
endings,theseterminalslosetheirmyelinsheathclosetotheirterminalsite,andformclose
contactswiththebasallaminaandconnectivetissuewithinthemuscularlayer71,72.The
distributionofSARsfromthetracheatothealveoliseemstoexhibitsomespecies
specificity,withanywherefrom40-90%beinglocatedintheintra-pulmonaryairways
versusinlargeextra-pulmonarysites73.
RARs,incontrast,arethoughttoterminateinthesubmucosaandairwayepithelium,
aconclusiondrawnfromthreepiecesofevidence.First,lightmucosalstrokinginsidethe
airway,aswellasinhalationofchemicalirritants,isabletoelicitresponsesinRARs,while
applicationofintra-airwaylocalanestheticssuchaslignocainecansuppresstheiractivity57,63,74,75.Second,thegreatestareaofRARresponsivenessisaroundthecarinaandlarge
airwaybranchpoints,whichisalsothesiteofgreatestintra-epithelialendingdensity57.
Finally,anintriguingandindirectobservationisthatanimalsthatlackthecoughreflex(e.g.
mice,ferrets),alsohaveveryfewintra-epithelialvagalterminalsandRARs62.The
sensitivityofRARstointrapulmonarycongestion,however,hasalsoraisedthepossibility
theyarefoundclosetolungvascularaswell76.RARsarelocatedinbothlargeandsmall
airways,andtendtohaveanatomicallyrestrictedreceptivefields.Forexample,inflationof
allairwaysandlungtissuedistaltoanRARreceptivesiteidentifiedbygentleintra-airway

20
mucosalstrokingfailstoelicitactivityoftheRAR57.
Intriguingly,geneticallytargetedlabelingofvagalafferentsthatexpressthe
purinergicreceptorP2RY1selectivelylabelsA-fibersthatinnervateneuroepithelialbodies
intheepithelialliningofthelung,aterminaltypethathadbeendescribedpreviouslybut
whosefunctionhasremainedunknown77,78.ActivationofP2RY1neuronsresultsinapnea
mirroringinhibitionofinflationviatheHering-Breuerreflex,andinsituhybridization
showedpartialoverlapbetweenP2ry1andthemechanosensorPiezo2,raisingthe
possibilitythisneuronpopulationisinvolvedintheHering-BreuerreflexandthatPiezo2
couldbetherelevantmechanosensorymolecule.
Summary
Mechanicaldistensionofthelungregulatesbreathingdepthandpatterns.Anintact
vagusnerveisrequiredforthedetectionoflungdistensionandchangesinrespiratory
physiology.Aselectsubsetofvagalafferentsrespondstolungstretch.Onesubsetisformed
ofslowlyadapting,low-threshold,puremechanosensorsthoughttoterminateinthe
smoothmusclesurroundingairwaysandmediatetheHeringBreuerreflex.Othersare
rapidlyadapting,high-threshold,mixedmechano-andchemo-receptorsthoughtto
terminateclosetotheairwayepitheliumandmediatecoughandbronchoconstrictionin
responsetoirritants.Howevermanyconclusionswithinthefielddependupon
assumptionsaboutstructure-functionrelationshipsthatstillrequirerigorousexamination,
andambiguityaboundsconcerningthemolecularmechanisms,anatomy,andphysiological
relevanceoflungmechanoreceptors.
Cardiovascularsystem
Physiology
Thevagalreflexregulationofthecardiovascularsystemholdsafundamentalplace
inthehistoryofneuroscience.OttoLoewiandHenryDalesharedtheNobelPrizein

21
PhysiologyandMedicinein1936forthediscoveryofthefirstneurotransmitter,
acetylcholine,releasedbythemotorfibersofthevagusnervetocauseadecreaseinheart
rate79.Justtwoyearslater,CorneilleHeymanwasawardedthesameprizeforthediscovery
thatsensorynervesinthelargevesselsleavingtheheartmonitorpressureandblood
chemistrytoregulatethecardiopulmonarysystem,inareflexarcthroughthevagalmotor
fibersinvestigatedinDaleandLoewi’swork.Thesecriticalexperimentsdelineatedthe
neuralpathwayofthebaroreceptorreflex,inwhichincreasesinbloodpressurecausea
reflexdecreaseinheartrateonabeat-to-beatbasis,allowingformaintenanceofaconstant
bloodpressure80,81.Thebaroreceptorreflexisimportantbothforpreventionof
hypertension,andofhypotension.Surgicalremovalofvagalafferentsintheaorticarchand
carotidbulbresultsinchronichypertension82.Inaddition,withoutreflexaccommodation
ofbloodpressure,movingfromalyingtoastandingpositionwouldresultinsucha
dramaticafallincerebralperfusionthatitwouldleadtolossofconsciousness.Vaso-vagal
syncopeandorthostasisare,infact,verycommonclinicalcomplaints.Furthermore,
decrementsinthebaroreceptorreflexwithageandtheconcomitantinabilitytoregulate
bloodpressurewithchangesinpositionisassociatedwithincreasedfalls,fractures,and
mortalityinpeople83.
Thebaroreceptorreflexisnottheonlycardiopulmonaryreflexmediatedbyvagal
sensoryfibers.Twootherprominentreflexesarecontrolledbyvagalafferents.Thefirst,
theBainbridgereflex,servestoregulateintravascularvolumestatus.Increasedatrial
volumeresultsintworeflexeffects:1)anincreaseinheartratetomovetheextravenous
bloodthroughtheheart,and2)anincreaseinurineoutputtoreduceintravascular
volume51,84.Initially,inspirationwasthoughtodrivethereflexincreaseinheartrate,as
notedpreviouslyinthediscussionofpulmonaryreflexphysiology.However,becausethe
heart,lungs,andgreatvesselsareallhousedinthethorax,changesinintrathoracic
pressureimpactsthedistributionofbloodinalltheseorgans.Reductionsinintrathoracic
pressureduringinspirationaugmentsthevenousreturnofthebloodtotheheart,and
carefulisolationofthisvariablefrominspirationitselfrevealedthecardiacchamber
dilationtobetherelevantcue51,84.
Thesecondreflex,termedtheBezold-Jarischreflex,isthoughttobeacardio-
protectivereflexinwhichbothmechanicalstretchandapplicationofirritantchemicalcues

22
tothemuscularheartwallcausesareflexbradycardia,hypotension,coronaryartery
dilatation,andapnea85,86.Reductionofheartratereducesmetabolicdemandofcardiac
tissue,systemichypotensionreducestheresistanceagainstwhichcardiacmyocytesneed
topumpblood,andcoronarydilationaugmentsperfusionofcardiactissue.Therefore,
thoughitisuncleartheextenttowhichmechanicalversuschemicalcuesdrivethisreflex,
theresultisreducedworkandimprovedoxygentationofcardiacmyocytes.Howthe
Bezold-Jarischreflexinteractswiththeothercardiopulmonaryreflexestomaintainnormal
bloodpressureandperfusionremainsanareaofuncertainty87.
Responseproperties
Assearchesfortheoperativeneuronalelementsinlungstretchwereunderway,
Adrian’scontemporariesweresimilarlyidentifyingtheneuronalelementswithchangesin
activitysynchronizedwiththebeatingheart.BronkandStellain1932werethefirstto
isolatesingleunitsinthecarotidsinusnervethatfiredcyclicallywithsystoleduringheart
beats,andchangedtheirfiringrateslinearlywithincreasesinbloodpressurefrom
40mmHgto120mmHg,therangeofnormalphysiologicalbloodpressuresintherabbit.
Theydeterminedthatincreasesinwhole-nervefiringratesinresponsetoincreasingblood
pressurewasbothafunctionofrecruitmentofmorefibers,andincreasesinfiringratein
individualfibers.Furthermore,thearterialbaroreceptorstheyidentified,aswithseveral
othervagalstretch-sensitiveelements,areslowly-adapting88.Subsequentstudieshave
confirmedvagalsensoryfibersareexquisitelytunedtopressurechangeswithinthegreat
vesselsexitingthehearttodeliverbloodtothebodyandbrain89-93.Intriguingly,despite
theirslowadaptationrate,vagalbaroreceptorsdoshifttheirbaselineandthresholds
followingprolongedexposuretohyperorhypo-tension,suggestingwhythissensory
apparatuswouldnotbesufficienttocounteractchronichypertensivedisease93.Thestress
historyofthevesselwallimpactsbaroreceptorresponsiveness92.
Mechanosensitiveafferentsinthecardiovascularsystemarenoonlylocatedinthe
majorvessels,butalsodetectmechanicalforcesintheheartwalls.Cardiacreceptorscanbe
furtherclassifiedbasedontheperiodofthecardiaccycleduringwhichtheiractivitypeaks.
Someafferentsaremaximallyactiveduringatrialcontraction,andexhibitincreasesin

23
firingratewithincreasesinheartrate,suggestingsensitivitytoactivechangesinatrialwall
stretch.Otherreceptorsrespondduringatrialfilling,andexhibitreducedactivitywith
increasesinheartrate.Thesereceptorsadaptslowlytochangesinpressure,suggesting
theyactprimarilyasmeasuresofoverallwallstrain.Bothoftheseatria-sensitivefibersare
exclusivelyfast-conductingA-fibers94,95.
Ventricularfibers,incontrast,aremuchlesscommonthanatrialfibers(ofcardiac
mechanosensors,75%arefoundintheatriaand25%intheventricles)andcanbe
classifiedfurtherintotwofunctionallydistinctgroups:onesetrespondsprimarilyto
ventricularcontraction,andtheothertogentleendocardialstroking96,97.Mostventricular
fibersareC-fibers,andalsorespondtointracardiacadministrationofabroadarrayof
chemicalcompounds.Thefunctionalsignificanceofthischemicalsensitivitymayrelateto
reflexesinvolvedinresponsetocardiacstressorischemia98-102.
However,thespecificmolecularmechanismsusedbycardiacmechanosensorsremain
undefined.Dissociatedaorticbaroreceptorneuronsexhibitmechanicallyinducedcurrents
thatcanbeblockedbygadolinium,suggestingthemolecularmechanosensormaybea
stretch-activatedionchannel103.Inaddition,bothamilorideanditsanalogue,benzamil,
inhibitedpressure-evokednerveactivityinbaroreceptorvagalafferents(Drummond,
2001),suggestingthatthechannelmaybearrelationtoENaCfamilymechanoreceptors.
However,alternativepossibilitiesabound,includingevidenceforexamplethatknockoutof
Trpc5attenuatesthebaroreceptorreflex104.Nocandidatetodatehasyetbeenshownto
fullyabolishthereflex.
Anatomy
Vagalafferentsinnervateallfourchambersoftheheart,theaorticarch,andmajor
brancharteriesintheneck.
Endingswithinthecardiacchambersconsistofthreeprimarytypes,1)complexun-
encapsulatedend-netsfoundmostdenselynearjunctionsofthegreatveinsenteringthe
atria,and2)anastomosingnetsoffineneuritesdistributedthroughoutatrialand
ventricularendocardium,someofwhichcloselycontactcardiacmyocytesand3)
terminalssurrounding“smallintenselyfluorescent”cellsofunknownfunctionin

24
cardiacganglia94,105-108.However,notably,withinthecardiacchambersitseemsthatthe
sameneuroncangiverisetomultipleoftheseendingtypes,complicatinginterpretationsof
structure-functionrelationships107.
Theneuronalendingsinthegreatvesselshavesimilarlybeencharacterizedinto
anatomicalsub-types.Approximately85%ofvessel-terminatingfibersdirectlyinnervate
thevesselwall,forming“flower-spray”and“end-net”typeterminals,whiletheremaining
15%terminateadjacenttoother“smallintensely-fluorescent”cellsproximaland
superficialtothewallitself105.Themostdetailedarchitecturalstudyofvagalafferentshas
beenperformedintheaorticarch,thesiteofmostelectrophysiologicallyactive
baroreceptors108.Aorticnervefibersinthisregionconsistbothofmyelinatedand
unmyelinatedfibersthatpassthroughthevesselwalladventitia,coilandterminatewithin
thevesselsmoothmusclemedialayer.Withinthislayer,myelinatedfiberslosetheirmyelin
sheath,andtheaxolemmaisdirectlyincontactwithextracellularconnectivetissue.The
tightapproximationofneuralelementswithelasticelementsofthewallspeakstoa
structure-functionrelationshipwellsuitedfortransductionofmechanicalforcesand
movementofthevesselwall.
Summary Sensorymonitoringofthemechanicalmovementoftheheartandgreatvessels
playsanimportantroleintheregulationofheartrateandbloodpressure.Aselectsubset
ofvagalafferentsrespondstochangesinpressureandstrainduringthecardiaccycle.Vagal
baroreceptors,arethoughttoinnervatethewallsoflargevessels,anddrivereflex
decreasesinheartrateinthesettingofhypertension,andreflexincreasesinthesettingof
hypotension.However,muchambiguityremainsconcerningtherelativerolesofheart
versusvessel-terminatingfibers,fibersofdifferentconductionvelocities,theinterplay
betweenchemicalandmechanicalstimuli,andofcoursethemolecularmechanisms
underlyingdetectionofpressureandstraininthissystem.

25
Conclusions Thevagusnervecontainsseveralafferentclassesthatmonitormechanicalforces
withinthegastrointestinal,respiratory,andcardiovascularsystems.Careful
electrophysiologicalcharacterizationandanatomicaltracinghasbeguntoelucidatesome
oftheterminaltypesandputativephysiologicalrolesoftheseimportantsensorycircuits.
Geneticidentificationofthespecificneuronsinvolvedhasjustbeguntounfold.Vagal
sensoryneuronsexpressingGLP1Rincludethegastrointestinalmechanosensors33.Vagal
sensoryneuronsexpressingP2RY1areA-fibersandinhibitrespiration,raisingthe
possibilitythesearetheslowly-adaptinglungmechanoreceptorsthatunderlietheHering-
Breuerreflex77.However,todate,theidentityofthereceptorresponsiblefor
mechanotransductionitselfininternalsensorysystemsremainsunknown.Discoveryofthe
molecularmechanismsatplaycouldresolvedebatesaboutthesiteofprimary
mechanosensation,moreclearlydefinetheneuronpopulationsinvolved,andalsoprovide
apharmacologicaltargetforpowerfulmanipulationoffundamentalautonomicreflexes.
ThemammalianmechanosensorPiezo2
Discovery Extensiveinvestigationintothemolecularidentityofmechanosensorshasyielded
discoveryofseveralnon-vertebratemechanosensitivechannelfamilies,includingthe
DEG/ENaC,TRP,TMCsandselectK+channels109.ThediscoveryofthePiezofamilyhas
offeredintriguingmolecularlyspecifiedmechanoreceptorsforqueryinknown
mechanosensitiveneuralsystems.
ThemembersofthePiezofamily,Piezo1andPiezo2,wereidentifiedinalarge-scale
screeningeffort110.Culturesofintrinsicallymechanosensitivemammaliancelllineswere
treatedwithsiRNAdesignedtoknockdownexpressionofproteinsofunknownfunction
predictedtocrossthecellmembraneatleasttwice,aconditionconsideredlikelyinarapid
mechanosensitivechannel.The71stcandidatesiRNAtestedtargetedFam38a,nownamed
Piezo1,andresultedinaseveral-foldreductionoftheintrinsicmechano-sensitivecurrent.
StructuralsimilarityledtoidentificationofthecloselyrelatedPiezo2.Overexpressionof

26
theseproteinsinheterologoussystemsconferredmechanosensitivitytootherwise
mechanicallyinsensitivecells.Furthermore,immunohistochemistryrevealedPiezo1tobe
localizedtothecellmembrane,thephysicalsiteofmechanotransduction.Piezo2wasfound
byinsituhybridizationtobeexpressedinasubsetofdorsalrootganglionneurons,and
knockdownofPiezo2inDRGculturesresultedinareductionofcellswithrapidly
inactivatingmechanosensitivecurrents,providingstrongevidencethatPiezo2playsarole
inmechanotransductioninknownmammalianmechanosensitiveneurons.Notably,slowly
inactivatingmechanosensitiveDRGneuronsremainedunperturbedbyknockdownof
Piezo2,suggestingadditionalmammalianmechanoreceptorsremaintobediscovered.
SubsequentworkprovidedfurtherevidencethatPiezo1andPiezo2wereinfactthe
molecularmechanosensitiveelement111.ExpressionofPiezoproteinsaloneinlipid
bilayerswassufficienttoconfermechanosensitivity.Furthermore,recordingsfrom
membranescontainingPIEZOproteinsshowsingle-channelcurrentsactivatedby
mechanicalforcethatcouldbeinhibitedwithrutheniumred,anon-selectivecationchannel
blocker.Photo-bleachingexperimentssuggestedthatPIEZOformstetramerswithinthecell
membrane.ThesedatatogethersuggestthatPiezo1andPiezo2assembleintopore-
forming,cationchannelscapableofmechanotransductionindependentofanyother
molecularelements.
Invivoproperties
Invertebratemechanosensation
Followingtheirdiscoveryandmolecularcharacterization,subsequentworksought
toelucidatetheinvivoexpressionpatternsandphysiologicalrelevanceofPiezo1and
Piezo2.ThefirststudytodemonstratetheinvivorelevanceofPiezomechanosensorswas
notinmammals,butratherinthefruitflyDrosophilamelanogaster112.ThePiezofamilyhas
onememberthatisexpressedinDrosophila,Dmpiezo.Expressionofafluorescentreporter
underthecontrolofDmpiezoresultedinfluorescenceinavarietyofsensoryneurons,
suggestingthepossibilityofaroleinmechanosensation.Dmpiezoknockoutflies,however,
didnotexhibitdeficitsincoordination,norinthebristlemechanoreceptorpotential,

27
suggestingDmpiezoisnotakeymechanoreceptorinadultflies.Incontrasthowever,
Dmpiezoknockoutlarvaeexhibitedreducedsensitivitytonoxiousmechanicalstimuli.
Isolationandrecodingofppkneurons,knowntobespecificallyimplicatedinlarvalnoxious
mechanicalsensitivity,demonstratedthatDmpiezoknockoutcausescompletelossof
mechanosensitivecurrentsinthisneuronclass.ThereforeDmpiezoplaysaroleinnoxious
mechanosensationinlarvae,makingitoneofthefirstionchannelstohavedemonstrated
bothinvitromechanosensitivityandaroleinbehaviorinvivo.
Vertebratedorsalrootgangliaandskinsensation
Withthedevelopmentofmousegenetictools,severalstudieshaveshownthat
Piezo2playsakeyroleinmultiplemechanosensorymodalitiesinmammals113,114.The
Piezo2proteincanbefoundonsensoryneuronterminalswithintheskin.Piezo2canalso
befoundwithinGolgitendonorgansandmusclespindlefibers,suggestinganadditional
roleinsensationofjointandlimbposition.Concordantwiththeseanatomicalresults,mice
lackingPiezo2showdeficitsindetectionofvibration,lighttouch,hairdeformation,low-
forcemechanicalstimuli,andproprioception,withoutdeficitsinresponsetopainor
temperature.Inaddition,Piezo2expressionisnotlimitedtoneurons;intriguingly,Piezo2
isalsoexpressedinMerkelcells,aspecializedcelltypeassociatedwithmechanosensitive
neuronterminals.Merkelcell-specificknockoutofPiezo2abolishedthe
mechanosensitivityofMerkelcells,andresultedinareductioninmechanosensitive
currentsandbehavioralresponsestolow-forcemechanicalstimuli115.Therefore,itislikely
thatPiezo2actsbothinneuronsandinMerkelcellstomediatelow-force
mechanosensation.
VertebratelunginflationandtheHeringBreuerreflex
Inadditiontoaroleinexternalsensationoftheskinandextremities,Piezo2has
alsobeenimplicatedinmechanosensationintherespiratorysystem69.Constitutive,global
deletionofPiezo2islethal;knockoutpupsarecyanotic,withlowarterialoxygen
saturation,abnormalrespiratoryrhythms,andsmallairspacesonhistologicalexamination

28
ofthelung,resultingindeathwithin24hoursofbirth.Totrytounderstandtherelevant
siteofPiezo2expressionforthislethalphenotype,differentmousedriverlineswereused
toselectivelyknock-outPiezo2intheendotheliumofbloodvessels,neuralcrestderivative
tissuessuchastheneuroepithelialbodiesinthelungandsensoryneuronswithinjugular
ganglia,thetrigeminalgangliaandofcoursethedorsalrootganglia,andincranialplacode-
derivedtissuessuchasthenodoseganglion.Piezo2knockoutspecificallyinthe
endotheliumorinthenodosegangliondidnotimpactpupsurvival.However,Piezo2
knockoutinthejugularganglion,dorsalrootganglion,andtrigeminalganglion
recapitulatedthephenotypeobservedwithglobalPiezo2deletion.Thesedatatogether
suggestthatPiezo2expressioninneuralcrest-derivedtissuesisnecessaryforsurvival,
andthatPiezo2lossfromtheneuralcrestresultsinabnormalpuprespiratorydynamics.
TheseobservationsbegthequestionoftherolePiezo2playsintherespiratory
systemintheadultanimal.Optogeneticactivationofneuronswithinthenodose-jugular
complexthateithercurrentlyexpressorhaveexpressedPiezo2duringdevelopment
resultsinapneaintheadult,implicatingthisneuronpopulationinadultrespiratory
control.ConditionaldeletionofPiezo2intheadultmousenodose,jugular,anddorsalroot
gangliaresultsinareductionofvagusnerveactivityinresponsetoinflationofthelung,a
findingconsistentwiththeroleofPiezo2inmechanosensationeitherwithintheneural
crestorplacode-derivedtissuesofthisstructure.
WhiledeletionofPiezo2fromtheplacode-derivednodosesensoryneuronsisnot
lethal,theseanimalsdoexhibitabnormalrespiratorycyclesasadults.Furthermore,vagal
responsestolungstretcharecompletelyablatedinPiezo2deletioninthenodoseganglion.
Thisresultraisesinterestingquestionsbecauseitsuggeststhatintheadultmouse,allvagal
lungmechanoreceptorsareinthenodoseganglion,andnoneintheneuralcrest-derived
jugularganglion.However,inpupslossofPiezo2intheneuralcrestbutnotthenodoseis
lethal.DoesthismeanthattheroleofPiezo2intheneuralcrestrequiredforsurvivalisnot
relatedtoadultdetectionoflunginflation?Howdothesensoryrolesofjugularversus
nodoseversusextra-vagalneuronsevolveoverthecourseofdevelopment?Ifdetectionof
lunginflationdependsonnodosePiezo2expression,andlossofdetectionoflunginflation
isnon-lethal,whatisthephysiologicalsignificanceofadultlunginflationmonitoring?
WhereelseisPiezo2deletedinbythedriverusedinnodosegangliondeletionexperiments,

29
anddoesthisresultinabnormalitiesthatcouldexplainthelossoflungstretchsensitivityin
theadult?ThedissociationoftheroleofPiezo2inneuralcrest–derivedversuscranial
placode-derivedtissuessuggestsitsub-servesmorethanonefunctionintherespiratory
system.
RoleofPiezo2inhumanmechanosensation
Piezo2playsanimportantroleinhumanmechanosensationanddevelopment.Gain-
of-functionmutationsinPiezo2havebeenshowntounderliearareautosomaldominant
formofdistalarthrogryposischaracterizedbymultipledistalcontractures,
ophthalmoplegia,ptosis,andrestrictivelungdisease116.Twofamilieswereidentifiedwith
similarfeatures;inoneaPiezo2pointmutationresultedinfasterrecoveryfrom
inactivation,andintheotheraPiezo2missensemutationcausedbothfasterrecoveryfrom
inactivationandslowerinactivationkinetics,bothmutationsthatwouldtranslateto
increasedsensitivitytomechanicalforces.Patientssurvivetilladulthoodandarefertile,
buthavecharacteristicfacieswithdeep-seteyes,anddifficultywithmusclemovement
includingoftheeye,back,neck,andbothlargeandsmalljointsoftheextremities.
Incontrast,twopatientsofdifferentancestryhavebeenidentifiedwithcomplete
Piezo2loss-of-functionmutations117.ThePiezo2variantsfoundinthesepatientseitherhad
Piezo2alleleswithprematurestopcodons,oroneallelewithaprematurestopcodonand
anotherwithamissensemutation.Thesevariantswereunabletogenerate
mechanosensitivecurrentswhenexpressedinheterologouscellsystems.
Thetwopatientswiththesemutationsbothpresentedwithasimilarconstellation
ofcharacteristics.Developmentalfeaturesincludedcongenitalhipdysplasia,finger
contractures,footdeformities,severeprogressivescoliosis,hypotonia,delayedwalkingand
headcontrol,andahistoryofshallowbreathingduringinfancy.Asadults,both
demonstratedaprofoundsensoryataxia,resultingindifficultyperformingreachingtasks
andaninabilitytowalkorstandwitheyesclosed,consistentwithdeficitsin
proprioception.Theywerealsounabletofeelavibratingtuningfork,hadseverelyreduced
lighttouchsensitivity,andexhibitedchanceperformanceoftwo-pointdiscriminationon

30
glabrousskin.Temperatureandpainsensitivitywasnormal.Theyalsohadintactlight
touchsensationonareasofhairyskin,suggestingaPiezo2-independentlighttouchsensory
modalityassociatedwithhair.Insummary,thesepatientsexhibitsimilarsensorydeficits
asmicethatlackPiezo2asadults.However,importantly,humanloss-of-functionofPiezo2
isnotnecessarilylethal.Furthercharacterizationofthemedicalhistoryduringinfancyand
characterizationoftherespiratorysystemofthesepatientswouldbeofgreatinterest.
Conclusions Piezo2isamammalianmechanosensorwithdemonstratedimportanceindetection
ofexternalmechanicalcues.Themolecularidentitiesofthemechanosensorsinvolvedin
detectionofinternalcuesremainunknown,thoughthephenotypesofPiezo2mutantmice
andhumansbothsuggestthatPiezo2couldbeinvolvedincriticalinternalphysiological
reflexes.Therefore,wesoughttodetermineiftheneuronsthathaveexpressedorcurrently
expressPiezo2inthenodoseganglionmediatedetectionofinternalstretch,andwhether
Piezo2knockoutcouldalterneuronresponseproperties.
Recordingmechanosensitiveafferentsinthevagus
Introduction SeveralexperimentalgoalsmustbemettoevaluatetheroleofPiezo2inmammalian
internalmechanosensation.Amethodtorecordtheactivityofvagalafferentsofknown
molecularidentityisrequired.Ideally,suchanexperimentaldesignwouldallowfor
simultaneousrecordingsfrommultipleneuronswithinthesameanimal.Similarly,stimulus
deliveryparadigmsmustenablerobustandconsistentactivationofmechanosensitive
vagalafferents.Thesegoalswereachievedusinginvivocalciumimagingwiththewell-
knowngeneticallyencodedcalciumindicatorGCaMP3inthenodoseganglion.These
methodsandresultsareelaboratedelsewhere33.However,forthepurposesof
understandingtheexperimentsherein,abriefsummaryisprovidedbelow.

31
Methods
Transgenicmiceusedforimagingexperimentsweregeneratedtoprovideganglion-
wideexpressionofthegeneticallyencodedcalciumindicatorGCaMP3,eitherthroughuse
ofaVglut2-ires-CrealleletodriveCre-dependentGcamp3expressioninallvagalsensory
neurons,orusingamouselineinwhichGcamp3isconstitutivelyexpressedinallcellsin
themousefrombirth.Thenodose/jugular/petrosalganglioncomplexwassurgically
exposed.Theconnectionstothebrainstemweretransected,andconnectionstothe
peripherypreserved.ImagingwasperformedwithaLeicaconfocalmicroscope,andthe
confocalpinholewasreducedtoavoidoverlappingcellsinthez-plane.Laserpowerdid
notexceed90uWtopreventbleachingandtissuedamage.Imageanalysiswasperformed
firstusingFiji;videoswerealignedsuchthatthesameneuronswereinthesamelocation
throughoutallimagingexperiments,neuronROIsweremanuallyselected,theaverage
intensitywithineachROIcalculatedforeachframe,andthesevalueswereexported.
MatlabwasusedtoconvertrawintensitiesintoaΔF/F,whereΔF/F=(intensity–average
baselineintensity)/averagebaselineintensity.
Recordingofneuronactivityofknownmolecularidentitywasachievedbycrossing
mouselinesthatexpressCrerecombinaseinasubsetofnodoseganglioncells,aCre-
dependenttdTomatoreporter,andtheconstitutiveGCaMP3allele.Becauseallcellsexpress
GCaMP3,andtargetedneuronswerevisualizedbytdTomatoexpression,thisenablesa
directcomparisonofresponsesinCre-positiveandCre-negativeneurons.Intriplecross
animals,thewavelengthrangecollectedforGCaMP3imagingwasrestrictedtoprevent
bleed-throughfromthetdTomatochannel.ThisresultedinbaselineGCaMP3fluorescence
intensitymeasurementsthatweresimilarbetweentdTomato-positiveandtdTomato-
negativeneurons(seedissertationthesis).InmyPhDdissertationwork,thisallowedfor
evaluationoftheresponsepropertiesofnodoseneuronsubsetsdefinedbyexpressionof
themarkersGpr65,Glp1r,P2ry1,Npy2r,andMc4r.
Gastricdistensionwasachievedbyeitheroftwomethods:1)inflationofasurgically
implantedlatexballoonaffixedtoasmallrodentfeedingneedleandsyringe,or2)inflation
ofthestomachwithnitrogengas(flowrate3-6mL/min,7-15secofinflation).Micewere
fastedovernightforgastrointestinalexperiments.Respiratorystimuliweredeliveredvia

32
trachealcannulaconnectedtogastanksthatwoulddeliverafixedgasflowrate.
Grossbloodpressuremanipulationswereattemptedbyrapidlyinjecting1-3mLof
lactatedringersintothemousevascularsupplyviafemoralveincatheter(4mice).
However,thisstimulusfailedtoelicitanyresponsesinthemicetested,andwas
abandoned.
Neuronswerecodedasresponsivetogastrointestinalmechanicalstimuliifeitherof
twocriteriaweremet:1)maximalGCaMP3fluorescencewas>sevenstandarddeviations
abovethebaselinemeanduringthestimulus,or2)ifmeanGCaMP3fluorescencewas>
threestandarddeviationsabovebaselinemeanduringtheentirestimulus.Neuronswere
categorizedasresponsivetointroductionofgasesinthelungiftheyexhibitedincreasesin
fluorescenceintensityduringthestimulusatleastthreestandarddeviationsorgreater
abovea30-secondbaselinemeanintensity.
Results Thedetailsoftheresultsprovidedarediscussedindetailinthedissertation
defense,andinpublication33.Briefly,however,invivocalciumimagingallowsforthe
identificationandcharacterizationofsensoryneuronpopulationsinthevagusnerveinthe
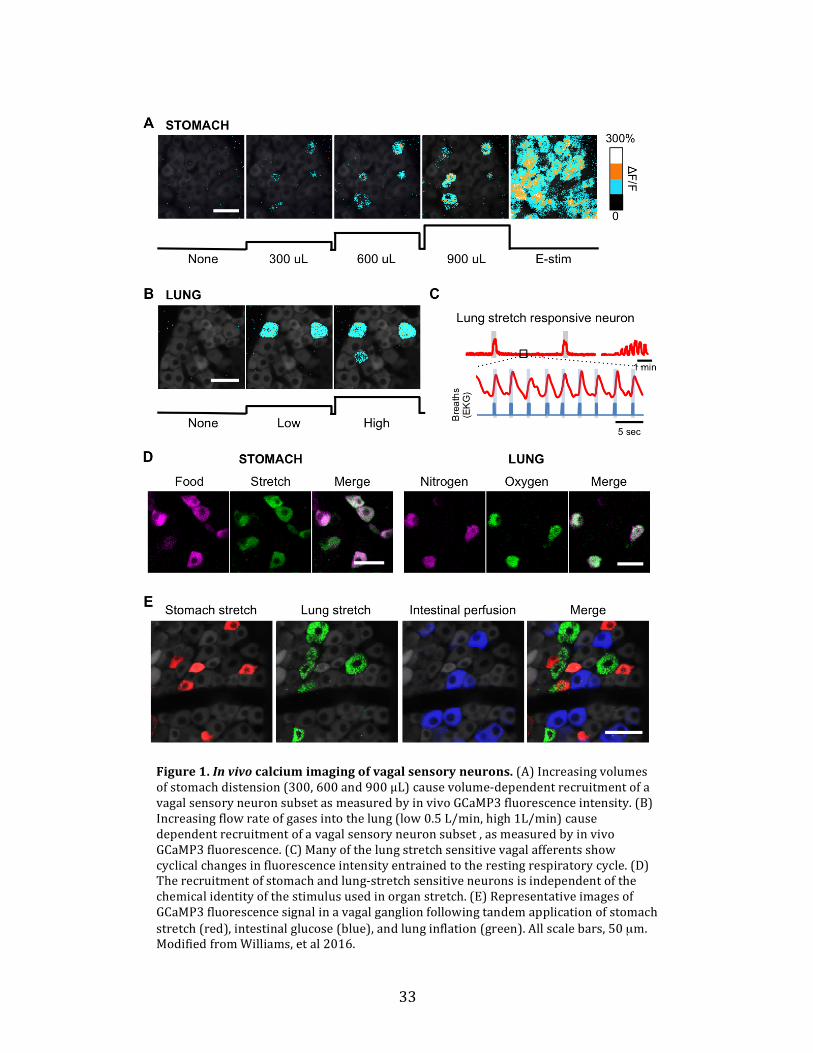
livingmouse.Stomachstretchcausesavolume-dependentrecruitmentofapproximately
17%ofallelectricallyresponsivevagalsensoryneurons(Figure1A).Lunginflation
similarlyrecruitedinaflow-dependentmannerapproximately4%ofvagalsensory
neurons,manyofwhichalsoshowedincreasesinactivitywiththerestingmouse
respiratorycycle(Figure1B,C).Thesemechanicallyresponsiveneuronsubsetswere
activatedindependentofthechemicalcompositionofthestimulusused(Figure1D).Inthe
caseofstomachstretch,thesameneuronswereactivatedbydistensionwithliquiddietor
nitrogengasinflation.Inthecaseoflungstretch,thesameneuronswereactivatedby
inflationwithnitrogen,oxygen,orroomair.Thesemechanicallyresponsivesubsetswere
uniquefromeachother,andfromtheneuronpopulationresponsivetochemicalstimuli
appliedinthesmallintestine(Figure1E).
33
Figure1.Invivocalciumimagingofvagalsensoryneurons.(A)Increasingvolumesofstomachdistension(300,600and900μL)causevolume-dependentrecruitmentofavagalsensoryneuronsubsetasmeasuredbyinvivoGCaMP3fluorescenceintensity.(B)Increasingflowrateofgasesintothelung(low0.5L/min,high1L/min)causedependentrecruitmentofavagalsensoryneuronsubset,asmeasuredbyinvivoGCaMP3fluorescence.(C)Manyofthelungstretchsensitivevagalafferentsshowcyclicalchangesinfluorescenceintensityentrainedtotherestingrespiratorycycle.(D)Therecruitmentofstomachandlung-stretchsensitiveneuronsisindependentofthechemicalidentityofthestimulususedinorganstretch.(E)RepresentativeimagesofGCaMP3fluorescencesignalinavagalganglionfollowingtandemapplicationofstomachstretch(red),intestinalglucose(blue),andlunginflation(green).Allscalebars,50µm.ModifiedfromWilliams,etal2016.

34
Finally,usingthetripletransgenicstrategytolabelspecificneuronsubsetswiththe
fluorescentproteintdTomatoallowedfortheidentificationoftheresponsepropertiesof
theseneuronpopulations.NeuronsmarkedbyexpressionofCreunderthecontrolofGlp1r,
orGLP1Rneurons,accountedformost(85%)ofthestomachstretchresponsivecells
(Figure2A).Incontrast,neuronsmarkedbyexpressionofCreunderthecontrolofGpr65
accountedformost(66%)oftheintestinallychemosensitiveneuronsubset(Figure2B).
Conclusions Thedevelopmentofinvivocalciumimaginginthenodoseganglionallowsfor
identificationofgastricandlungmechanosensitiveneuronpopulations,andconcurrent
identificationofmolecularlydefinedneuronsubtypes.Therefore,invivoimagingprovides
anexcellentexperimentalplatformtoinvestigatetheresponsepropertiesofPiezo2-
markedvagalsensoryafferents.
Figure2.Invivocalciumimagingofmolecularlydefinedvagalsensoryneuronsubsets.(A)RepresentativeimagesofGLP1RneuronsandincreasesinfluorescenceofthecalciumindicatorGCaMP3inresponsetofoodintheintestine,andtostomachstretch.Thestomachstretch,butnottheintestinalfoodresponsiveneuronsarecompletelycontainedwithintheGLP1Rneuronsubset.(B)Conversely,alloftheintestinefoodresponsiveneuronsarecontainedwithintheGPR65neuronsubset.Scalebar,50µm.ModifiedfromWilliamsetal,2016.

35
Experiment1:InVivoimagingofPiezo2
lineageneurons
Introduction IfPiezo2isthemechanosensorforinternalmechanicalstimuli,mechanosensitive
vagalafferentsshouldexpressPiezo2.Totestthishypothesis,weusedinvivocalcium
imagingtoexaminetheresponsepropertiesofneuronsthatexpressedaCre-dependent
tdTomatoreporterinPiezo2-Cremice.
Furthermore,theobservationthatselectiveactivationofP2RY1neuronscauses
apnearaisesthepossibilitythatP2RY1neuronsmayalsoplayaroleindetectionoflung
stretch.Insituhybridizationhasshownpartialoverlapbetweenneuronsthatexpress
P2ry1andPiezo277,furthersuggestingthatperhapstherelevantlung-stretchsensitive
populationconsistsoftheneuronsthatexpressbothP2ry1andPiezo2.Therefore,wealso
soughttoexaminethemechanicalsensitivityoftheP2RY1neuronsubsets.
Methods Wegeneratedthetripleknock-inmouselinePiezo2-ires-Cre;lox-tdTomato;Rosa26-
GCaMP3(Piezo2-GCaMP3*),andP2ry1-ires-Cre;lox-tdTomato;Rosa26-GCaMP3(P2ry1-
GCaMP3*),whichallowforaninvivonodoseganglionimagingapproachinvolving
constitutiveexpressionofthegeneticallyencodedcalciumindicatorGCaMP3,andthe
Piezo2-Cre-dependentexpressionoftdTomato.Imaging,stimulusadministration,and
analysiswereperformedasdescribedpreviously.
Results Piezo2-GCaMP3*micedemonstratedexpressionoftdTomatobroadlyinsatellite
glialcellssurroundingnodoseganglionneurons,andalsowithinasubsetofsensory
neurons.Of283electricallyresponsiveneurons(n=3mice),98(34.6%)expressed

36
tdTomato.P2ry1-GCaMP3*micealsodemonstratedexpressionoftdTomatobroadlyin
satelliteglialcellssurroundingnodoseganglionneurons,aninasubsetofneuronsaswell.
Of211electricallyresponsiveneurons(n=3mice),31(14.7%)expressedtdTomato.
InPIEZO2experiments,seventeenneuronswereidentifiedasresponsivetolung
stretchstimuli,aproportionofthetotalganglion(7.2%)consistentwithpriorestimatesof
thesizeofthelung-stretchresponsivepopulation.Ofthese17lungstretchresponsive
neurons,16werepositivefortdTomato(94%).Conversely,16of84(19%)tdTomato-
positiveneuronswereresponsivetolungstretch.Fromthesedataweconcludethatnearly
alllung-stretchresponsiveneuronshaveexpressedPiezo2,andthatlung-stretch
responsiveneuronsaccountforapproximatelyone-fifthofneuronsthathaveexpressed
Piezo2(Figure3A).
P2RY1
1
31
52
(211)
P2RY1 Merge Lung stretch
PIEZO2 1
100 98
(283)
PIEZO2 Merge Lung stretch
A B
Figure3.PIEZO2neurons,butnotP2RY1neuronscontainlungmechanoreceptors.(A)Representativeimagesandtime-resolvedresponsesof100sensoryneurons,(ΔF/F,colorscale)inresponsetolungstretch(greenbar,15seconds).Nearlyalllung-stretchresponsiveneuronsarecontainedwiththePIEZO2neuronpopulation.(B)Representativeimagesandtime-resolvedresponsesof52sensoryneurons,(ΔF/F,colorscale)inresponsetolungstretch(greenbar,15seconds).Noneofthelung-stretchresponsiveneuronsarecontainedwiththeP2RY1neuronpopulation.Scalebars,50µm.

37
Incontrast,inP2RY1experiments,twenty-threeneuronswereidentifiedas
responsivetolungstretch(11%),aproportionofthetotalganglionsomewhathigherthan
priorestimatesofthesizeofthelungstretchpopulation.Ofthese23lungstretch
responsiveneurons,2werepositivefortdTomato(8.6%),thoughtheresponseamplitudes
ofthesetwoneuronsweremarkedlyreducedcomparedtotdTomatonegativeresponsive
neurons.Fromthesedata,weconcludethattheP2RY1neuronpopulationlargelydoesnot
includelung-stretchsensitivecells(Figure3B).
InPIEZO2experiments,thirty-oneneuronswereidentifiedasresponsiveto
stomachstretch,aproportionoftheganglion(13.1%)consistentwithpriorestimatesof
thesizeofthestomach-stretchresponsivepopulation.Ofthese31stomachstretch
responsiveneurons,usingstandardcut-offstodefinerespondersversusnon-responders,
18(52.9%)werepositivefortdTomato(Figure4A).Anindependent-samplest-testwas
conductedtocomparethemeanresponseamplitude,expressedaspercentchangefrom
baseline,duringstomachstretchintdTomatopositiveversustdTomatonegativestomach-
stretchresponsiveneurons.Therewasasignificantdifferenceintheresponseamplitudes
fortdTomatopositive(M=66,SEM=16)andtdTomatonegative(M=11,SEM=5)
neurons;t(29)=2.83,p=0.008.Therefore,thoughbystandarddefinitionsforresponsive
versusnon-responsiveneurons,onlyhalfofstomach-stretchresponsivecellsaretdTomato
positive,thetdTomatopositiveneuronsweremuchmorestronglyresponsivethanthe
tdTomatonegativeneurons.Conversely,18of84(21%)tdTomatopositiveneuronswere
stomachstretchresponsive.Fromthesedataweconcludethatmoststronglyresponsive
stomachstretchsensitiveneuronshaveexpressedPiezo2,andthatstomach-stretch
responsiveneuronsaccountforanotherone-fifthofneuronsthathaveexpressedPiezo2.

38
Finally,thoughoneofthestomachstretchpreparationsfailedinP2RY1
experiments,datawasanalyzedfromtwoofthethreeanimalstested,yieldingatotalof
181electricallyresponsiveneurons.Ofthese181neurons,32(17.7%)wereresponsiveto
stomachstretch,aproportionsimilartopriorestimatesofthesizeofthestomachstretch
sensitivepopulation.Ofthe32stomachstretchresponsiveneurons,4(13%)werepositive
fortdTomato(Figure4B).Fromthesedataweconcludethatthemajorityofstomach
stretchsensitiveneuronshaveneverexpressedP2ry1.
B
PIEZO2
236
84
1
A PIEZO2 Merge Stomach stretch P2RY1 Merge Stomach stretch
P2RY1
181
1
31
Figure4.PIEZO2neurons,butnotP2RY1neuronscontainstomachmechanoreceptors.(A)Representativeimagesandtime-resolvedresponsesof236sensoryneurons,(ΔF/F,colorscale)inresponsetostomachstretch.Nearlyalllung-stretchresponsiveneuronsarecontainedwiththePIEZO2neuronpopulation.(B)Representativeimagesandtime-resolvedresponsesof181sensoryneurons,(ΔF/F,colorscale)inresponsetostomachstretch.Noneofthestomach-stretchresponsiveneuronsarecontainedwiththeP2RY1neuronpopulation.Allscalebars,50µm.Greenbarsundertherasterplotsindicatetimeofapplicationofstretchstimuli(Piezo220and30seconds,P2ry130seconds).

39
Conclusions NeuronsthathaveexpressedPiezo2atsomepointintheirlifetimeencompass
stomachandlungmechano-sensitivepopulations.Incontrast,neuronsthathaveexpressed
P2ry1intheirlifetimearenotresponsivetostomachorlungmechanicalstimuli.These
findingshavetwokeyimplications.
First,thesedatasupportthehypothesisthatPiezo2couldbethemolecular
mechanosensor.Asacorollary,suchaconclusionwouldalsosuggestthattheneurons
themselvescouldbetheprimarymechanosensorbecausetheyaremarkedbyexpressionof
aknownmolecularmechanosensor.Furtherexperimentsarerequired,however,to
demonstratetherelevanceofthePiezo2moleculeitself.Whilesomedataalreadysupport
theroleofPiezo2inlungmechanosensation,nothingisknownaboutPiezo2ingastric
mechanosensation.
Second,thesedatasupporttheconclusionthePiezo2-positive,P2ry1-negative
neuronscontaintherelevantlung-stretchresponsivepopulation.Thisobservationis
importantfortworeasons.First,itisacleardemonstrationthatP2RY1neuronsarenot
responsiblefortheHeringBreuerreflex,beggingthequestionwhatA-fibersthat
selectivelyinnervateneuroepithelialbodiesandwhocaninducecompletemouseapneaare
doing.P2ry1isauniquemolecularhandleonanunanticipatedvagalneuronsubset.
Second,becauselung-stretchresponsiveneuronsonlycoverasubsetofPIEZO2neurons,
additionalgeneticmarkersarerequiredtomorepreciselydefinethelungstretchsubset.
P2ry1canthereforeserveinfutureexperiments,suchasinanalysisofsingle-celldata,to
restrictinvestigationintoPiezo2-positive,P2ry1-negativeneurontypes.Suchexperiments
mightrevealcriticalmarkersormanipulablefeaturesoflung-stretchsensitiveneuron
subsetstobeexploitedinthefuture.

40
Experiment2:Invivoimagingofneurons
thatexpressPiezo2intheadult
IntroductionNeuronsthathaveexpressedPiezo2atsomepointintheirdevelopmentcontain
nearlyallthelungandstomachstretchresponders.However,acriticalshortcomingof
lineage-tracingexperimentsisthattheydonotdistinguishbetweenhistoric,
developmentalexpressionandactivefunctionalexpressionintheadult.ifPiezo2werethe
mechanosensoritself,onewouldalsoadditionallypredictthatmechanosensitiveneurons
expressPiezo2notonlyatsometimeduringdevelopment,butalsothattheyexpress
Piezo2inadulthood.Totestthishypothesisrequiresanexperimentalparadigminwhich
neuronsthatexpressPiezo2inadulthoodareselectivelylabeled,andinwhichtheir
responsepropertiescanberecordedtostretchstimuli.
MethodsWegeneratedthedoubleknock-inmouselinePiezo2-ires-Cre;Rosa26-GCaMP3,and
usedadeno-associatedvirusestodeliverflex-tdTomatovectorstoallnodosesensory
neurons.ThisresultedinexpressionoftdTomatoonlyinneuronsexpressingPiezo2-Creat
thetimeofinjectioninadulthood.Responsepropertiesoflabeledneuronscouldthenbe
recordedusinginvivocalciumimagingasinpriorexperiments.
Givenconcernsabouttheimpactofviralinjectionsonneuronhealthand
responsiveness,thesameexperimentswereperformedusingGlp1r-ires-Cre;Rosa26-
GCaMP3animalsaspotentialpositivecontrols.GLP1Rneuronshadpreviouslybeenshown
toincludethestomach-stretchresponsiveneuronpopulation33.
Adeno-associatedvirusinjectionswereperformedasdescribedpreviously33,77.
Briefly,miceareanesthetized,theleftnodoseganglionsurgicallyexposed,andAAVmixed
withFastGreendyeisinjectedintothebodyoftheganglion.Injectionsareconsidered
grosslysuccessfulwhentheganglionbodyfillswithbluedye.Theincisionintheneckis

41
closedandtheanimalsallowedtorecover.Imagingexperimentswereperformed6-7days
followinginjectionsinhopesofpreservingneuronviabilitywhileachievingreasonable
amountsofinfectionandconstructexpression.
Results InPiezo2animals,631electricallyresponsiveneuronswereanalyzed(n=7
animals).Ofthese631neurons,45weretdTomatopositive,suggestingacombined
infectionandexpressionefficiencyof7.1%.Whenthesameanalysiswasperformedbased
onlyuponmorphologicalidentificationofcellbodiesratherthanonresponsivenessto
electricalstimulation,811neuronswereidentifiedofwhich72weretdTomatopositive
(8.8%efficiency).ThesimilarityoftdTomatoexpressionfrequencyinallneuronsversusall
electricallyresponsiveneuronssuggeststhattdTomatoexpressionwasnotimpairinggross
neuronresponsiveness.
InGlp1ranimals,441electricallyresponsiveneuronswereanalyzed(n=4animals).
Ofthese441neurons,40weretdTomatopositive,suggestingacombinedinfectionand
expressionefficiencyof9.0%.Thisefficiencyisconcordantwiththeobservationthatinsitu
hybridizationforPiezo2andGlp1rinadultmousenodosegangliaresultsinlabeling
neuronalsubsetsofsimilarsize.
WefirstexaminedthedegreeofoverlapbetweentdTomato-expressingneurons
andstomachstretchresponsiveness.InGlp1rexperiments,71neuronsintotalresponded
tostomachstretch.Ofthe40tdTomato-positiveneuronsinGlp1ranimals,18(45%)were
responsivetostomachstretch(Figure5A).AChi-squaredtestforindependenceshowed
thattherewasasignificantrelationshipbetweentdTomatoexpressionandstretch
responsiveness,X2(2,N=441)=27.203,p<0.01(Figure5B).Thisresultisinaccordwith
theearlierobservationthatGLP1Rneuronscontainnearlyallthestomachstretch
responsiveneurons,andthatadultlabelingofGLP1RneuronslabelsnearlyallIGLE-type
endingsinthestomach33.Theseresultsserveasanimportantproofofconceptandpositive
control.
InPiezo2animalexperiments,54ofthe631neuronsanalyzedwereresponsiveto
stomachstretch(8.6%).Ofthesestomachstretchresponders,5(9.2%)weretdTomato

42
positive(Figure5A).Thesedatasuggesttwoobservations.First,asubsetofadultPiezo2-
expressingneuronscanrespondtostomachstretch.However,wewouldfurtherpredict
thatifPiezo2istherelevantstomachmechanosensor,thereshouldbegreater-than-chance
co-occurrenceoftdTomatoexpressionandstomachstretchresponsiveness.Weperformed
aChi-squaredanalysis,andfoundthattherewasnorelationshipbetweenstomachstretch
andadultPiezo2expression,X2(2,N=631)=0.40,p=0.53(Figure5B).Thesedatadonot
supportthehypothesisthatPiezo2istherelevantadultgastrointestinalmechanosensor.
WenextexaminedthedegreeofoverlapbetweentdTomato-expressingneurons
andlungstretchresponsiveness.OnlytwoGlp1ranimalshadlung-stretchstimuli
administered,resultinginanalysisof169electricallyresponsiveneurons.Seventeen
neuronswereresponsivetolungstretch(10%),aresponseratesimilartoprior
experimentalparadigms.NoneoftheseresponsiveneuronsweretdTomatopositive,
consistentwithpriorworkshowingnooverlapbetweenGLP1Rneuronsubsetslabeled
withgeneticlineage-tracingmethods,andlung-stretchresponsivesubsets.
InPiezo2experiments,ofthe631electricallyresponsiveneurons,29were
responsivetolungstretch(4.6%),similartopreviouslyreportedratesoflung-stretch
Figure5.ExpressionofPiezo2inadulthooddoesnotpreferentiallylabelstomachstretchsensitiveneurons.(A)Representativeimagesofneuronresponsestostomachstretch(green)inGlp1r-iresCreAAV-lox-tdTomatoandPiezo2-iresCreAAV-lox-tdTomatoneurons(pink).Scalebar,50µm.(B)PlotoftheobservednumberoftdTomatoandstomachstretchdouble-positiveneurons(PIEZO2,black;GLP1R,red)versustheexpectednumberofneuronscalculatedbasedoninfectionandresponserates(gray).**p<0.01forX2testforindependence.

43
responsiveness.Ofthese29lungstretchresponsiveneurons,3(10.3%)weretdTomato
positive.AChi-squaredanalysisshowedtherewasnorelationshipbetweenlungstretch
responsivenessandtdTomatoexpression,X2(2,N=631)=0.47,p=0.49.Thisresultis
surprisinginlightofthefindingsthatknockoutofPiezo2impactstheHering-Breuerreflex
andwhole-nerverecordingsofthevagusinresponsetolungstretch.
Conclusions TherelationshipbetweenGlp1r-drivenAAV-tdTomatoexpressionandstomach
stretchresponsivenessconfirmsthatstomach-stretchresponsiveneuronsexpressGlp1rin
adulthood.Moreimportantly,thisobservationsuggeststhatviralinfectionandinvivo
imagingcanbecompatibletechniquesinthisparadigm.However,thefindingthatthere
wasnorelationshipbetweenadultPiezo2-driventdTomatoexpressionandstomach
stretchresponsivenesscastsdoubtonthehypothesisthatPiezo2istherelevantmolecular
mechanosensorinstomachstretch.Rather,thisresultmightsuggestthatPiezo2playsan
importantdevelopmentalroleintheformationofIGLEs,butthatitsexpressionisnot
preservedintoadulthoodinthisneuronpopulation.
However,theobservationthatadultPeizo2-driventdTomatoexpressionandlung
stretchresponsivenessarenotrelatedissurprising.KnockoutofPiezo2frombirthinthe
neuronsubsetimagedintheseexperimentsabolisheswhole-nerveresponsestolung
stretch.Itispossiblethataninsufficientnumberofneuronswasqueriedinthisparadigm
torevealarelationship;lung-stretchresponsivecellsaremuchmorerarethatstomach-
stretchresponders.Despitemultipleimagingpreparations,thisresultdependsheavilyon
datafromlessthan30neurons.Furthermore,therobust-nessofneuronpopulationsto
viralinfectionmaydifferdependingonneurontype,makingthisexperimentsuitableto
examinestomach–stretch,butnotlung-stretchsensitivesubsets.
Alternatively,iflung-stretchresponsiveneuronstrulydonotexpressPiezo2in
adulthood,itwouldsuggestthatPiezo2intheneuronsthemselvesisnotcriticalforlung
mechanosensation.PerhapstheexpressionofPiezo2innon-neuronalelements,suchits
expressioninthenodosesatellitecellsorintargetorgans,issufficienttoconfer
mechanosensitivity.DriversusedtoknockoutPiezo2inpriorexperimentsarenotfully

44
nodose-neuronspecific.Similarly,anadditionalalternativeexplanationisthatPiezo2plays
animportantdevelopmentalroleinlungstretchsensitiveafferentsand/ortheir
surroundingstructuressuchthatitslosswouldconfermechano-insensitivitywithout
Piezo2beingtheactualmechanoreceptor.
Experiment3:Detectionofstretchstimuliin
Piezo2knock-outganglia
Introduction ThehypothesisthatPiezo2actsasthemechanosensorinvagalsensoryafferents
furthermoresuggeststwoadditionalpredictions.First,lossofexpressionofPiezo2should
resultinthelossofmechanicallyevokedresponses.Second,iftheneuronsarethedirect
mechanosensors,theselectivelossofPiezo2intheneuronsthemselvesbutnotinother
tissuesshouldbesufficienttoabolishresponses.Totestthesepredictionsrequiresan
experimentalparadigminwhichPiezo2expressionisdisruptedselectivelyinthenodose
ganglionneurons,andinwhichresponsepropertiescanstillberecorded.
Methods Usingaseriesofmousecrosses,wegeneratedPiezo2fl/fl-GCaMP3*,Piezo2fl/wt-
GCaMP3*,andPiezo2wt/wt-GCaMP3*animals.Anadeno-associatedviruscontainingaCAG-
Cre-mCherryconstructwaspurchasedfromSignaGenlaboratories(AAV1-CAG-Cre-
mCherry,SL101117).InfectionwiththisvectorisintendedtoinducetheexpressionofaCre
recombinaseandmCherryfusionalleletogeneraterednuclearfluorescenceinevery
neuronalsoexpressingCre.ExpressionofCrerecombinaseincellscontainingafloxed
Piezo2allelewouldresultingeneticknockoutofPiezo2.Imagingexperimentswere

45
performed14-21daysafterAAVinjectionsinhopesofprovidingsufficienttimefor
knockdownofPiezo2expression.SuccessfulexpressionofCre,andexcisionofthePiezo2
allelewereconfirmedusingPCRofgangliacollectedaftercompletionofimaging
experiments.
Results TwotothreeweeksfollowinginjectionofAAV1-CAG-Cre-mCherry,nodoseganglia
demonstratedwidespreadandrobustexpressionoftdTomatointheinfectedganglion,but
notintheuninfectedcontralateralganglion.Theredfluorescencewaslocalizedtothecell
nucleus,concordantwiththefluorescentproteinbeingtaggedtonucleus-localizedCre
recombinase.Inthefiveinjectedanimals,278electricallyresponsiveneuronswere
analyzed,ofwhich186(67%)containedamCherry-positivenucleus,suggestinghigh
infectionefficiencyintheseexperiments(Figure6A).PCRofganglionDNAextractedafter
imagingexperimentsshowedpositivereactionsforCrerecombinasefrominfected,butnot
fromuninfectedganglia.Furthermore,PCRreactionsspecificforthesequenceofthe
knockoutPiezo2allelewerepositiveonlyininfectedgangliafrommicecarryingatleast
onefloxedPiezo2allele(Figure6B).TheseresultssuggestnotonlythatCrewas
successfullyexpressed,butalsothatitwasfunctionalandexcisedthefloxedregionof
Piezo2.

46
Figure6.Validationofneuron-specific,adultdeletionofPiezo2.(A)Fluorescentmicroscopyimageofthenodoseganglion14-21daysfollowinginfectionwithAAV1-CAG-Cre-mCherry.showswidespreadexpressionofnuclearmCherry.Scalebar,200µm.ConfocalimagesofneuronswithintheganglionshownuclearmCherrylocalizationScalebarinsets50µm.(B)PolymerasechainreactiondemonstratestheexpressionofCrerecombinaseininfectedbutnotuninfectedganglia,andexcisionofexons43-45ofPiezo2onlyininjectedgangliaofanimalswithafloxedallele.Scalebar,50µm.

47
Offiveanimalssuccessfullytested,twowerePiezo2fl/wt,meaningthatPiezo2
expressionfromonealleleshouldalwaysbepreserveddespiteexpressionofCre.Inthese
animals,107electricallyresponsiveneuronswereanalyzed.Ofthese107neurons,7
(6.5%)wereresponsivetolungstretch,5ofwhichweremCherrypositiveand2ofwhich
weremCherrynegative.Furthermore,19(17.8%)werestomachstretchresponsive,of
which14weremCherrypositiveand5weremCherrynegative.Therefore,Creexpression
didnotabolishmechanosensitiveresponsesinnodosesensoryneurons(Figure7A).Chi-
squaredanalysisshowedtherewasnorelationshipbetweenlungstretchresponsiveness
andtdTomatoexpression,X2(2,N=107)=0.90,p=0.34,norbetweenstomachstretch
responsivenessandmCherryexpressioninanimals,X2(2,N=107)=1.88,p=0.17.These
datasuggestthatinjectionandconstructexpressionitselfdoesnotimpactmechanical
sensitivityofvagalsensoryneurons.
TheremainingthreeanimalstestedwerePiezo2fl/flanimals,meaningthatneurons
expressingmCherryintheseanimalsalsohaveexcisionofbothcopiesofthefloxedPiezo2
geneticsequence.Intheseanimals,171electricallyresponsiveneuronswereanalyzed.Of
these171neurons,25(14%)werestomach-stretchresponsive,ofwhich14weremCherry
positiveand11weremCherrynegative.Furthermore,4(2.3%)werelungstretch
responsive,ofwhich2weremCherrypositiveand2weremCherrynegative.Therefore,
evenwhenbothPiezo2alleleswerefloxed,neuronsexpressingCrerecombinase
maintainedtheirabilitytorespondtomechanicalstimuli(Figure7A).AChi-squared
analysisshowedtherewasnorelationshipbetweenstomachstretchresponsivenessand
mCherryexpressioninPiezo2-floxedanimals,X2(2,N=171)=0.0002,p=0.98,nor
betweenlungstretchresponsivenessandmCherryexpression,X2(2,N=171)=0.07,p=
0.80.Similarly,therewasnoattenuationinstomachstretchresponseamplitudein
mCherry-positiveneuronsinPiezo2fl/flanimals(Figure7B).Rather,mCherry-positive
neuronshadaslightlygreaterresponseamplitude(M=2.1,SD=1.3)thanmCherry-
negativeneurons(M=1.0,SD=1.02),t(22)=0.481,p=0.03.Toofewlungstretch
responsiveneuronswerepresentinthesepreparationsforrigorousstatisticalanalysisof
responseamplitudes,particularlygiventhesmallnumberofanimalstested.

48
Conclusions ThelackofrelationshipbetweenPiezo2excisionandeitherstomachorlungstretch
castsfurtherdoubtontheroleofPiezo2onlungandstomachmechanosensationinvagal
sensoryafferents.However,onetechnicalpointlimitsenthusiasmforthestrengthof
conclusionstobedrawninthisexperiment.WhilegenomicexcisionofthePiezo2exons
couldbeconfirmedgrosslybywhole-ganglionPCR,thisvalidationprovidesonlyaproxy
indicationthatproteinlevelsofPiezo2arealteredusingthisexperimentalmanipulation.
Forexample,thePiezo2knockout-specificPCRreactioncouldbepositiveincasesof
excisionofthefloxedPiezo2regioninonlyanincompletesubsetofneurons.Alternatively,
Figure7.Neuron-specific,adultdeletionofPiezo2doesnotabolishmechanicalresponses.(A)RepresentativeimagesofpreservedstomachandlungstretchresponsesinmCherry-positiveandnegativeneuronsinPiezo2fl/flandPiezo2fl/wtanimals.Scalebar,50µm.(B)Responseamplitudesforallstomachstretch(left)andlungstretch(right)responsiveneuronsinPiezo2fl/flanimals.TracesfromneuronsthatexpressmCherryareinred,whiletracesfromneuronsthatdonotexpressmCherryareinblack.

49
evenifknockoutwerecompleteatthegenomiclevel,thedurabilityofexpressedPiezo2
proteinisunknown,andmayextendbeyondthe2-3weeksbetweeninfectionandimaging
utilizedinthisexperimentalparadigm.However,intriguingly,theonlyotherexperimentto
knockdownPiezo2inadultanimalsgeneratedreductionsinskinsensitivitywithinone
weekofinductionofgenomicexcisionofthefloxedregionofPiezo269,suggestingthe2-3
weekwindowusedintheseexperimentsshouldbesufficient.Unfortunately,toolsto
evaluateproteinexpression,andspecificallyproteinexpressionatdistalsensoryterminals
withintargetorgans,arelacking.Interpretationofresultsmustthereforenecessarilybe
constrainedwiththisshortcominginmind.Theabsenceofarelationshipisanegative
resultthatcouldreflecttruebiology,oratechnicalhurdle.
WithincompletevalidationofPiezo2proteinexpressionknockdowninmindasa
prominenttechnicalshort-comingfordatainterpretation,thesedataarenotconcordant
withthehypothesisthatvagalsensoryneuronPiezo2isthesoleandnecessary
mechanosensorineitherstomachorlungstretch.Thisconclusionagreeswiththelackof
relationshipbetweentheadultexpressionofPiezo2andstomachstretchinprior
experiments,supportingthenotionthatPiezo2doesnotplayaroleintheadultgastric
mechanosensitiveneuronpopulation.
ThelackofrelationshipbetweenneuronalPiezo2knockoutandlungstretch
responsiveness,however,issurprising.ThesinglepriorexperimenttosuggestPiezo2
knockoutinadultsaltersrespiratorydynamicsandattenuatestheHeringBreuerreflex
usedatamoxifen-dependentAdvillin-CredrivertodeletePiezo2fromvagalanddorsalroot
ganglionneurons69.Theknockoutmicehad30%largertidalvolumes,andreachedpeak
expiratoryflowfasterthanwild-typecontrols.Theseanimalsalsohadlowerwholevagal
nerveactivityduringlunginflation.Onedifferencebetweenthisexperimentandvirally
mediatedknockdownisthatAdvillin-CrealsodrivesPiezo2knockoutoutsidethevagus
(e.g.inthedorsalrootganglia),whichcouldimpactrespiratorydynamics.However,this
wouldnotdirectlyexplainlowervagalwhole-nerverecordinglungstretchresponse
amplitudes.Inadditiontotamoxifen-inducedAdvillin-Cre-drivenPiezo2knockout,the
driverPhox2bwasusedtodeletePiezo2frombirthinnodoseneuronsandglia.Knockout
frombirthresultedinabolitionoftheHeringBreuerreflexandvagusnerveresponsesto
lunginflation.However,experimentsthatknockoutagenefrombirthcannotdistinguish

50
betweendevelopmentalandadultimpactofgeneloss.Therefore,thepossibilityremains
that,asinMerkelcellsintheskin,non-vagalPiezo2expressionplaysanimportantrolein
Piezomechanotransduction,andthatknockoutspecificallyinadultvagalsensoryneurons
isinsufficienttoabolishlungstretchmechanosensitivity.
SummaryDetectionofmechanicalforceswithinthebodyplaysacriticalroleinregulationof
organismphysiology.WeinvestigatedwhetherPiezo2,arecentlyidentifiedmammalian
mechanosensor,couldplayaroleindetectionofinternalmechanosensorycues.Wefound
thatlineagetracingofneuronsthathaveexpressedPiezo2labeledthelargemajorityof
lungandstomachmechanosensitiveneuronpopulationsinthenodoseganglionofthe
vagusnerve.However,neuronsthatexpressedPiezo2inadulthooddidnotpreferentially
labelmechanosensitivepopulations.Inaddition,adultandnodoseneuronspecificPiezo2
knockoutfailedtoabolishmechanosensitiveresponsesinnodosesensoryneurons,though
conclusionsfromthisexperimentarelimitedbytechnicalconcerns.Thesedatain
aggregatesuggestPiezo2maynotbethesolemechanosensorwithininternalsensory
systems,andfurthermoreraisethepossibilitythatextra-neuralPiezo2couldplayan
importantroleinthesesystemsasitdoesintheskin,orthatPiezo2playsanimportant
developmentalrole.Furtherexperimentationsuchas1)usingalternativemethodsto
knockoutPiezo2fromvagalsensoryneuronsandassociatedtissues,2)evaluationofadult
PIEZO2neuronanatomyintheperiphery,and3)investigationintotheroleofPiezo2inthe
cardiovascularsystem,wouldshedintriguinglightontotheroleofPIEZO2neurons,and
theroleofPiezo2expressionininternalmechanosensation.
Bibliography1. Foley,J.O.&DuBois,F.S.Quantitativestudiesofthevagusnerveinthecat.I.The
ratioofsensorytomotorfibers.JournalofComparativeNeurology67,49–67(1937).

51
2. Sawchenko,P.E.Centralconnectionsofthesensoryandmotornucleiofthevagusnerve.J.Auton.Nerv.Syst.9,13–26(1983).
3. Saper,C.B.Thecentralautonomicnervoussystem:consciousvisceralperceptionandautonomicpatterngeneration.Annu.Rev.Neurosci.25,433–469(2002).
4. Rinaman,L.,Card,J.P.,Schwaber,J.S.&Miselis,R.R.Ultrastructuraldemonstrationofagastricmonosynapticvagalcircuitinthenucleusofthesolitarytractinrat.JournalofNeuroscience9,1985–1996(1989).
5. Janowitz,H.&Grossman,M.I.Somefactorsaffectingthefoodintakeofnormaldogsanddogswithesophagostomyandgastricfistula.Am.J.Physiol.Regul.Integr.Comp.Physiol.159,143–148(1949).
6. Share,I.,Martyniuk,E.&Grossman,M.I.Effectofprolongedintragastricfeedingonoralfoodintakeindogs.JournalofPhyiology169,229–235(1952).
7. Phillips,R.J.&Powley,T.L.Gastricvolumeratherthannutrientcontentinhibitsfoodintake.Am.J.Physiol.271,R766–9(1996).
8. Powley,T.L.&Phillips,R.J.Gastricsatiationisvolumetric,intestinalsatiationisnutritive.Physiology&Behavior82,69–74(2004).
9. Phillips,R.J.&Powley,T.L.Gastricvolumedetectionafterselectivevagotomiesinrats.Am.J.Physiol.274,R1626–38(1998).
10. Gonzalez,M.F.&Deutsch,J.A.Vagotomyabolishescuesofsatietyproducedbygastricdistension.Science212,1283–1284(1981).
11. Walls,E.K.,Phillips,R.J.,Wang,F.B.,Holst,M.C.&Powley,T.L.Suppressionofmealsizebyintestinalnutrientsiseliminatedbyceliacvagaldeafferentation.Am.J.Physiol.269,R1410–9(1995).
12. Yox,D.P.,Stokesberry,H.&Ritter,R.C.Vagotomyattenuatessuppressionofshamfeedinginducedbyintestinalnutrients.Am.J.Physiol.260,R503–8(1991).
13. Ogawa,N.etal.Thevagalafferentpathwaydoesnotplayamajorroleintheinductionofsatietybyintestinalfattyacidinrats.NeuroscienceLetters433,38–42(2008).
14. Sclafani,A.&Lucas,F.Abdominalvagotomydoesnotblockcarbohydrate-conditionedflavorpreferencesinrats.Physiology&Behavior60,447–453(1996).
15. ColquittJL,P.K.L.E.F.G.Surgeryforweightlossinadults(Review).1–243(2013).16. Ponce,J.etal.TheREDUCEpivotaltrial_aprospective,randomizedcontrolled
pivotaltrialofadualintragastricballoonforthetreatmentofobesity.1–8(2015).doi:10.1016/j.soard.2014.12.006
17. Moura,D.etal.Effectivenessofintragastricballoonforobesity_Asystematicreviewandmeta-analysisbasedonrandomizedcontroltrials.SurgeryforObesityandRelatedDiseases12,420–429(2016).
18. Dixon,J.B.etal.Adjustablegastricbandingandconventionaltherapyfortype2diabetes:arandomizedcontrolledtrial.JAMA299,316–323(2008).
19. Favretti,F.etal.Laparoscopicadjustablegastricbandingin1,791consecutiveobesepatients:12-yearresults.ObesSurg17,168–175(2007).
20. Sjöström,L.etal.EffectsofbariatricsurgeryonmortalityinSwedishobesesubjects.N.Engl.J.Med.357,741–752(2007).
21. Paintal,A.S.Astudyofgastricstretchreceptors;theirroleintheperipheralmechanismofsatiationofhungerandthirst.JournalofPhyiology126,255–270(1954).

52
22. Iggo,A.Tensionreceptorsinthestomachandtheurinarybladder.J.Physiol.(Lond.)128,593–607(1955).
23. Iggo,A.GASTROINTESTINALTENSIONRECEPTORSWITHUNMYELINATEDAFFERENTFIBRESINTHEVAGUSOFTHECAT.QuarterlyJournalofExperimentalPhysiologyandCognateMedicalSciences42,130–143(1957).
24. Slattery,J.A.,Page,A.J.,Dorian,C.L.,Brierley,S.M.&Blackshaw,L.A.Potentiationofmousevagalafferentmechanosensitivitybyionotropicandmetabotropicglutamatereceptors.J.Physiol.(Lond.)577,295–306(2006).
25. AshleyBlackshaw,L.,Grundy,D.&Scratcherd,T.Involvementofgastrointestinalmechano-andintestinalchemoreceptorsinvagalreflexes"anelectrophysiologicalstudy.J.Auton.Nerv.Syst.18,1–10(2002).
26. Blackshaw,L.A.,Page,A.J.&Partosoedarso,E.R.Acuteeffectsofcapsaicinongastrointestinalvagalafferents.Neuroscience96,407–416(2000).
27. Jin,Y.-H.,Takemura,M.,Furuyama,A.&Yonehar,N.ReceptorsInhibitMechanosensitivityofPrimaryAfferentEndings.Pharmacology1–6(1999).doi:10.5772/34100
28. Zagorodnyuk,V.P.,Chen,B.N.,Costa,M.&Brookes,S.J.H.Mechanotransductionbyintraganglioniclaminarendingsofvagaltensionreceptorsintheguinea-pigoesophagus.J.Physiol.(Lond.)553,575–587(2004).
29. Regehr,B.L.S.A.W.G.TIMINGOFSYNAPTICTRANSMISSION.Annu.Rev.Physiol.61,521–542(1999).
30. Bennett,M.&Zukin,S.ElectricalCouplingandNeuronalSynchronizationintheMammalianBrain.Neuron41,495–511(2004).
31. Zagorodnyuk,V.P.&Brookes,S.J.Transductionsitesofvagalmechanoreceptorsintheguineapigesophagus.JournalofNeuroscience20,6249–6255(2000).
32. Zagorodnyuk,V.P.,Chen,B.N.&Brookes,S.J.Intraganglioniclaminarendingsaremechano-transductionsitesofvagaltensionreceptorsintheguinea-pigstomach.J.Physiol.(Lond.)534,255–268(2001).
33. Williams,E.K.etal.SensoryNeuronsthatDetectStretchandNutrientsintheDigestiveSystem.Cell166,209–221(2016).
34. NONIDEZ,J.F.Afferentnerveendingsinthegangliaoftheintermuscularplexusofthedog'soesophagus.J.Comp.Neurol.85,177–189(1946).
35. Neuhuber,W.L.Sensoryvagalinnervationoftheratesophagusandcardia:alightandelectronmicroscopicanterogradetracingstudy.J.Auton.Nerv.Syst.20,243–255(1987).
36. Wang,F.B.&Powley,T.L.Topographicinventoriesofvagalafferentsingastrointestinalmuscle.J.Comp.Neurol.421,302–324(2000).
37. Berthoud,H.R.&Powley,T.L.Vagalafferentinnervationoftheratfundicstomach:morphologicalcharacterizationofthegastrictensionreceptor.J.Comp.Neurol.319,261–276(1992).
38. Berthoud,H.R.,Patterson,L.M.,Neumann,F.&Neuhuber,W.L.Distributionandstructureofvagalafferentintraganglioniclaminarendings(IGLEs)intheratgastrointestinaltract.AnatomyandEmbryology195,183–191(1997).
39. fox,E.A.,Phillips,R.J.,Martinson,F.A.,Baronowsky,E.A.&Powley,T.L.Vagalafferentinnervationofsmoothmuscleinthestomachandduodenumofthemouse:morphologyandtopography.J.Comp.Neurol.428,558–576(2000).

53
40. Powley,T.L.etal.UltrastructuralevidenceforcommunicationbetweenintramuscularvagalmechanoreceptorsandinterstitialcellsofCajalintheratfundus.Neurogastroenterology&Motility20,69–79(2008).
41. Widdicombe,J.Reflexesfromthelungsandairways:historicalperspective.J.Appl.Physiol.101,628–634(2006).
42. Breuer,J.Theself-steeringofrespirationthroughtheNervusVagus,translatedbyE.Ullman.BreathingHering-BreuerCentenarySymposiumCibaFoundationSymposium,357–394(1970).
43. Head,H.OntheReglationofRespiration:PARTI.Experimental.J.Physiol.(Lond.)10,1–152.53(1889).
44. Head,H.OntheRegulationofRespiration:PartII.Theoretical.J.Physiol.(Lond.)10,279–290(1889).
45. Cross,K.W.,KLAUS,M.,TOOLEY,W.H.&WEISSER,K.Theresponseofthenew-bornbabytoinflationofthelungs.J.Physiol.(Lond.)151,551–565(1960).
46. Guz,A.,Noble,M.I.M.,Trenchard,D.,Cochrane,H.L.&Makey,A.R.Studiesonthevagusnerveinman:theirroleinrespiraoryandcirculatorycontrol.ClinicalScience293–304(1964).
47. Dejours,P.,RAYNAUD,J.,MONZEIN,P.&BECHTEL,Y.[StudyoftheHering-Breuerinspirationinhibitorreflexinmanduringnaturalsleep].J.Physiol.(Paris)54,320–321(1962).
48. Guz,A.,Noble,M.I.M.,Widdicombe,J.G.,Trenchard,D.&Mushin,W.W.TheeffectofbilateralblockofvagusandglossopharyngealnervesontheventilatoryresponsetoCO2ofconsciousman.RespirationPhysiology1,206–210(1966).
49. Guz,A.etal.Theroleofvagalandglossopharyngealafferentnervesinrespiratorysensation,controlofbreathingandarterialpressureregulationinconsciousman.ClinicalScience30,161–170(1966).
50. Anrep,G.V.,Pascual,W.&Rossler,R.Respiratoryvariationsoftheheartrate-I—Thereflexmechanismoftherespiratoryarrhythmia.ProceedingsoftheRoyalSocietyB119,191–213(1936).
51. Bainbridge,F.A.Therelationbetweenrespirationandthepulse-rate.J.Physiol.(Lond.)54,192–202(1920).
52. DeBurghDaly,M.,Hazzledine,J.L.&Ungar,A.Thereflexeffectsofalterationsinlungvolumeonsystemicvascularresistanceinthedog.J.Physiol.(Lond.)188,331–351(1967).
53. Looga,R.Reflexcardiovascularresponsestolunginflation:areview.RespirationPhysiology109,95–106(1997).
54. Adrian,E.D.Afferentimpulsesinthevagusandtheireffectonrespiration.J.Physiol.(Lond.)79,332–358(1933).
55. KNOWLTON,G.C.&LARRABEE,M.G.Aunitaryanalysisofpulmonaryvolumereceptors.Am.J.Physiol.147,100–114(1946).
56. Bartlett,D.,Jeffery,P.,Sant'Ambrogio,G.&Wise,J.C.Locationofstretchreceptorsinthetracheaandbronchiofthedog.J.Physiol.(Lond.)258,409–420(1976).
57. Widdicombe,J.G.Receptorsinthetracheaandbronchiofthecat.J.Physiol.(Lond.)123,71–104(1954).
58. Bartlett,D.&Sant'Ambrogio,G.Effectsoflocalandsystemichypercapniaonthedischargeofstretchreceptorsintheairwaysofthedog.RespirationPhysiology26,

54
91–99(1976).59. Sheldon,M.I.&Green,J.F.EvidenceforpulmonaryCO2chemosensitivity:effects
onventilation.JApplPhysiolRespirEnvironExercPhysiol52,1192–1197(1982).60. Matsumoto,S.,Takahashi,T.,Tanimoto,T.,Saiki,C.&Takeda,M.Effectsof
potassiumchannelblockersonCO2-inducedslowlyadaptingpulmonarystretchreceptorinhibition.TheJournalofPharmacologyandExperimentalTherapeutics290,974–979(1999).
61. Ho,C.Y.,Gu,Q.,Lin,Y.S.&Lee,L.Y.Sensitivityofvagalafferentendingstochemicalirritantsintheratlung.RespirationPhysiology127,113–124(2001).
62. Karlsson,J.A.,Sant'Ambrogio,G.&Widdicombe,J.Afferentneuralpathwaysincoughandreflexbronchoconstriction.J.Appl.Physiol.65,1007–1023(1988).
63. Widdicombe,J.Airwayreceptors.RespirationPhysiology125,3–15(2001).64. Pisarri,T.E.,Jonzon,A.,COLERIDGE,H.M.&COLERIDGE,J.C.Vagalafferentand
reflexresponsestochangesinsurfaceosmolarityinlowerairwaysofdogs.J.Appl.Physiol.73,2305–2313(1992).
65. Lai,C.J.&Kou,Y.R.Stimulationofpulmonaryrapidlyadaptingreceptorsbyinhaledwoodsmokeinrats.J.Physiol.(Lond.)508(Pt2),597–607(1998).
66. Mills,J.E.,Sellick,H.&Widdicombe,J.G.Activityoflungirritantreceptorsinpulmonarymicroembolism,anaphylaxisanddrug-inducedbronchoconstrictions.J.Physiol.(Lond.)203,337–357(1969).
67. Sellick,H.&Widdicombe,J.G.Stimulationoflungirritantreceptorsbycigarettesmoke,carbondust,andhistamineaerosol.J.Appl.Physiol.31,15–19(1971).
68. Yu,J.Spectrumofmyelinatedpulmonaryafferents.AJP:Regulatory,IntegrativeandComparativePhysiology279,R2142–8(2000).
69. Nonomura,K.etal.Piezo2sensesairwaystretchandmediateslunginflation-inducedapnoea.Nature541,176–181(2017).
70. Matsumoto,S.Effectsofvagalstimulationonslowlyadaptingpulmonarystretchreceptorsandlungmechanicsinanesthetizedrabbits.Lung174,333–344(1996).
71. Krauhs,J.M.Morphologyofpresumptiveslowlyadaptingreceptorsindogtrachea.Anat.Rec.210,73–85(1984).
72. Yamamoto,Y.,Atoji,Y.&Suzuki,Y.Calretininimmunoreactivenerveendingsinthetracheaandbronchioftherat.J.Vet.Med.Sci.61,267–269(1999).
73. Schelegle,E.S.&Green,J.F.Anoverviewoftheanatomyandphysiologyofslowlyadaptingpulmonarystretchreceptors.RespirationPhysiology125,17–31(2001).
74. Sampson,S.R.&Vidruk,E.H.Propertiesof‘irritant’receptorsincaninelung.RespirationPhysiology25,9–22(1975).
75. Armstrong,D.J.&LUCK,J.C.AcomparativestudyofirritantandtypeJreceptorsinthecat.RespirationPhysiology21,47–60(1974).
76. Kappagoda,C.T.,Skepper,J.N.,McNaughton,L.,Siew,E.E.&Navaratnam,V.Morphologyofpresumptiverapidlyadaptingreceptorsintheratbronchus.J.Anat.168,265–276(1990).
77. Chang,R.B.,Strochlic,D.E.,Williams,E.K.,Umans,B.D.&Liberles,S.D.VagalSensoryNeuronSubtypesthatDifferentiallyControlBreathing.Cell161,622–633(2015).
78. Adriaensen,D.etal.Pulmonaryintraepithelialvagalnodoseafferentnerveterminalsareconfinedtoneuroepithelialbodies:ananterogradetracingand

55
confocalmicroscopystudyinadultrats.CellTissueRes293,395–405(1998).79. Tansey,E.M.HenryDaleandthediscoveryofacetylcholine.ComptesRendus
Biologies329,419–425(2006).80. Heyman,J.F.&Heyman,C.Recherchesphysiologiquesetpharmacodynamiquessur
lateteisoleeduchien.TravailDeLInstitutdePharmacodynamieetdeTherapiedeLUniversitedeGand32,1–33(1926).
81. Heymans,C.ReflexogenicAreasoftheCardiovascularSystem.PerspectivesinBiologyandMedicine3,409–417(1960).
82. Nowak,S.J.CHRONICHYPERTENSIONPRODUCEDBYCAROTIDSINUSANDAORTIC-DEPRESSORNERVESECTION.Ann.Surg.111,102–111(1940).
83. Juraschek,S.P.etal.AssociationofHistoryofDizzinessandLong-termAdverseOutcomesWithEarlyvsLaterOrthostaticHypotensionAssessmentTimesinMiddle-agedAdults.JAMAInternMed177,1316–9(2017).
84. Bainbridge,F.A.Theinfluenceofvenousfillingupontherateoftheheart.J.Physiol.(Lond.)50,65–84(1915).
85. Paintal,A.S.ASTUDYOFVENTRICULARPRESSURERECEPTORSANDTHEIRROLEINTHEBEZOLDREFLEX.QuarterlyJournalofExperimentalPhysiologyandCognateMedicalSciences40,348–363(1955).
86. DAWES,G.S.&Widdicombe,J.G.TheafferentpathwayoftheBezoldreflex:theleftvagalbranchesindogs.BrJPharmacolChemother8,395–398(1953).
87. Campagna,J.A.&Carter,C.ClinicalrelevanceoftheBezold-Jarischreflex.Anesthesiology98,1250–1260(2003).
88. Bronk,D.W.&Stella,G.AfferentImpulsesintheCarotidSinusNerve:Therelationofthedischargefromsingleendorganstoarterialbloodpressure.TheJournalofCellularandComparativePhysiology1,113–130(1932).
89. COLERIDGE,H.M.&COLERIDGE,J.C.Cardiovascularafferentsinvolvedinregulationofperipheralvessels.Annu.Rev.Physiol.42,413–427(1980).
90. COLERIDGE,H.M.,COLERIDGE,J.C.&Schultz,H.D.CharacteristicsofCfibrebaroreceptorsinthecarotidsinusofdogs.J.Physiol.(Lond.)394,291–313(1987).
91. COLERIDGE,H.M.etal.ImpulsesinSlowlyConductingVagalFibersfromAfferentEndingsintheVeins,Atria,andArteriesofDogsandCats.CirculationResearch33,87–97(1973).
92. COLERIDGE,H.M.,COLERIDGE,J.C.,Poore,E.R.,Roberts,A.M.&Schultz,H.D.Aorticwallpropertiesandbaroreceptorbehaviouratnormalarterialpressureandinacutehypertensiveresettingindogs.J.Physiol.(Lond.)350,309–326(1984).
93. COLERIDGE,H.M.,COLERIDGE,J.C.,Kaufman,M.P.&DANGEL,A.Operationalsensitivityandacuteresettingofaorticbaroreceptorsindogs.CirculationResearch48,676–684(1981).
94. COLERIDGE,J.C.,HEMINGWAY,A.,HOLMES,R.L.&LINDEN,R.J.Thelocationofatrialreceptorsinthedog:aphysiologicalandhistologicalstudy.J.Physiol.(Lond.)136,174–197(1957).
95. Paintal,A.S.Astudyofrightandleftatrialreceptors.J.Physiol.(Lond.)120,596–610(1953).
96. COLERIDGE,H.M.,COLERIDGE,J.C.&KIDD,C.CARDIACRECEPTORSINTHEDOG,WITHPARTICULARREFERENCETOTWOTYPESOFAFFERENTENDINGINTHEVENTRICULARWALL.J.Physiol.(Lond.)174,323–339(1964).

56
97. Thorén,P.N.Characteristicsofleftventricularreceptorswithnonmedullatedvagalafferentsincats.CirculationResearch40,415–421(1977).
98. Armour,J.A.Myocardialischaemiaandthecardiacnervoussystem.Cardiovasc.Res.41,41–54(1999).
99. Kaufman,M.P.,Baker,D.G.,COLERIDGE,H.M.&COLERIDGE,J.C.StimulationbybradykininofafferentvagalC-fiberswithchemosensitiveendingsintheheartandaortaofthedog.CirculationResearch46,476–484(1980).
100. Benson,C.J.,Eckert,S.P.&McCleskey,E.W.Acid-evokedcurrentsincardiacsensoryneurons:Apossiblemediatorofmyocardialischemicsensation.CirculationResearch84,921–928(1999).
101. Thames,M.D.&Minisi,A.J.Reflexresponsestomyocardialischemiaandreperfusion.Roleofprostaglandins.Circulation80,1878–1885(1989).
102. Ustinova,E.E.&Schultz,H.D.Activationofcardiacvagalafferentsinischemiaandreperfusion.Prostaglandinsversusoxygen-derivedfreeradicals.CirculationResearch74,904–911(1994).
103. Sullivan,M.J.etal.NonVoltage-GatedCa2+InfluxThroughMechanosensitiveIonChannelsinAorticBaroreceptorNeurons.CirculationResearch80,861–867(1997).
104. Lau,O.-C.etal.TRPC5channelsparticipateinpressure-sensinginaorticbaroreceptors.NatureCommunications7,1–12(2016).
105. Cheng,Z.,Powley,T.L.,Schwaber,J.S.&Doyle,F.J.Alaserconfocalmicroscopicstudyofvagalafferentinnervationofrataorticarch:chemoreceptorsaswellasbaroreceptors.J.Auton.Nerv.Syst.67,1–14(1997).
106. MILLER,M.R.&KASAHARA,M.STUDIESONTHENERVEENDINGSINTHEHEART.Am.J.Anat.115,217–233(1964).
107. Cheng,Z.,Powley,T.L.,Schwaber,J.S.&Doyle,F.J.Vagalafferentinnervationoftheatriaoftheratheartreconstructedwithconfocalmicroscopy.J.Comp.Neurol.381,1–17(1997).
108. Krauhs,J.M.Structureofrataorticbaroreceptorsandtheirrelationshiptoconnectivetissue.J.Neurocytol.8,401–414(1979).
109. Chalfie,M.Neurosensorymechanotransduction.NatRevMolCellBiol10,44–52(2009).
110. Coste,B.etal.Piezo1andPiezo2AreEssentialComponentsofDistinctMechanicallyActivatedCationChannels.Science330,55–60(2010).
111. Coste,B.etal.Piezoproteinsarepore-formingsubunitsofmechanicallyactivatedchannels.Nature483,176–181(2013).
112. Kim,S.E.,Coste,B.,Chadha,A.,Cook,B.&Patapoutian,A.TheroleofDrosophilaPiezoinmechanicalnociception.Nature483,209–2012(2013).
113. Ranade,S.S.etal.Piezo2isthemajortransducerofmechanicalforcesfortouchsensationinmice.Nature516,121–125(2015).
114. Woo,S.-H.etal.Piezo2istheprincipalmechanotransductionchannelforproprioception.NatNeurosci18,1756–1762(2015).
115. Woo,S.-H.etal.Piezo2isrequiredforMerkel-cellmechanotransduction.Nature509,622–626(2015).
116. Coste,B.etal.Gain-of-functionmutationsinthemechanicallyactivatedionchannelPIEZO2causeasubtypeofDistalArthrogryposis.ProceedingsoftheNational

57
AcademyofSciences110,4667–4672(2013).117. Chesler,A.T.etal.TheRoleofPIEZO2inHumanMechanosensation.N.Engl.J.Med.
375,1355–1364(2016).