
Adenine nucleotides and adenosine metabolism in pig kidney proximal tubule membranes
7
JOURNAL OF CELLULAR PHYSIOLOGY 157:77-83 11993) Adenine Nucleotides and Adenosine Metabolism in Pig Kidney Proximal Tubule Membranes jULlA BLANCO, ENRIC 1. CANELA, J O A N SAYOS, JOSEFA MALLOL, CARME LLUIS, AND RAFAEL FRANCO* Departanlent de Bioqui’rnica i Fisiologia, Facultat de Quirnica, Universitat dc Barcelona, Marti i Franqui.s, I, 08028 Barcelona, Catalonia, Spain Exogenous adenosine triphosphate (ATP) added to brush-border membrane vesi- cles was rapidly degraded mainly to inosine according to the high ecto-nucleotid- ase activities in these vesicles. In the absence of phosphate, inosine was slowly transformed into hypoxanthine, and xanthine oxidase and dehydrogenase activi- ties were not detected. The presence of ecto-adenosine deaminase and ecto- adenosine monophosphate (AMP) nucleotidase was shown. The ecto-adenosine deaminase was inhibited by deoxycoformycin and was also detected in rat renal brush-border membrane vesicles. Using orthovanadate, levamisole, and a, p- methylene adenosine diphosphate as possible inhibitors, alkaline phosphatase was shown to be the main agent responsible for ecto-AMP nucleotidase activity. In pig renal basolateral membrane vesicles and in whole cell extracts from pig renal cortex, ecto-AMP nucleotidase was the limiting factor in ATP degradation. Comparing the ATP catabolism in the whole cell cortical extract with the catabo- lism in the same sample precleared of membranes, it was shown that ecto- nucleotidase activity is mainly bound to the membranous components. It is also shown that the whole cell extract of pig renal cortex has hypoxanthine phos- phoribosyl transferase activity, and it seems probable that the rapid and specific formation of luminal inosine and its transport into the cell in competition with adenosine may start the purine salvage pathway through the synthesis of IMP from hypoxanthine. o 1993 WiIey-Liss, ltlc Cell adenine nucleotides and their metabolites are involved in the regulation of numerous intracellular processes in kidney (Spielman and Arend, 1991). Ade- nosine affects different renal physiological functions such as blood flow (Gouyon and Guignard, 19891, glom- erular filtration rate (Arend et al., 1987), urine flow, and sodium and potassium excretion rate (Collis et al., 19861, glomerular constriction related to the entry of calcium into glomerular cells (Oliveira et al., 19891, and the renin-angiotensin system (Deray et al., 1990). Adenosine is also important in the maintenance of kid- ney viability before transplantation (Ametani et al., 1990). Most of these adenosine effects are mediated by specific receptors (see Spielman and Arend, 1991 for references). Part of the intrarenal adenosine production comes from cellular adenosine release. In kidney (Siege1et al., 1983) as in other tissues (Meghji et al., 1988; Vanden- berghe et al., 1989) production of adenosine, and also inosine and hypoxanthine increases when catabolism of intracellular adenosine triphosphate (ATP) is acceler- ated, for instance in hypoxia and during enhanced en- ergy consumption. On the other hand, the extracellular production of adenosine from nucleotides is also impor- tant (hlic et al., 1990). The origin of extracellular nucleotides is unclear, although some come from blood through glomerular filtration, and a significant (I> 1993 WILEY-LISS, INC. amount may come from tubular cells by exocytosis, since proximal tubular exocytotic vesicles may dis- charge nucleotides into the luminal fluid. culic et al. (1990) demonstrated the presence of ectoenzymes in rat renal cortical brush-border membrane vesicles that produce adenosine as a final product using ATP, ade- nosine diphosphate (ADP), or adenosine monophos- phate (AMP) as substrate. The complete hydrolysis of ATP yielding adenosine is catalyzed by at least two separate ectoenzymes, one capable of hydrolyzing ATP and ADP, but not AMP, and another that hydrolyzes AMP. Extracellular adenosine acts on adenosine receptors and is then metabolized or internalized into the cell by means of nucleoside uptake systems (Le Hir and Du- bach, 1984; Angielski et al., 1983; Trimble and Coul- son, 1984; Franco et al., 1990). The adenosine uptake is important not only to maintain suitable extracellular nucleoside concentrations but also in the recovery of intracellular nucleotide levels after their depletion (Weinberg et al., 1988). Recovery from hypoxia re- Received February 26,1993; accepted May 11, 1993. *To whom reprint requestsicorrespondence should be addressed.
-
Upload
julia-blanco -
Category
Documents
-
view
213 -
download
0
Transcript of Adenine nucleotides and adenosine metabolism in pig kidney proximal tubule membranes

JOURNAL OF CELLULAR PHYSIOLOGY 157:77-83 11993)
Adenine Nucleotides and Adenosine Metabolism in Pig Kidney Proximal
Tubule Membranes jULlA BLANCO, ENRIC 1. CANELA, JOAN SAYOS, JOSEFA MALLOL, CARME LLUIS, AND
RAFAEL FRANCO* Departanlent de Bioqui’rnica i Fisiologia, Facultat de Quirnica, Universitat dc Barcelona,
Marti i Franqui.s, I , 08028 Barcelona, Catalonia, Spain
Exogenous adenosine triphosphate (ATP) added to brush-border membrane vesi- cles was rapidly degraded mainly to inosine according to the high ecto-nucleotid- ase activities in these vesicles. In the absence of phosphate, inosine was slowly transformed into hypoxanthine, and xanthine oxidase and dehydrogenase activi- ties were not detected. The presence of ecto-adenosine deaminase and ecto- adenosine monophosphate (AMP) nucleotidase was shown. The ecto-adenosine deaminase was inhibited by deoxycoformycin and was also detected in rat renal brush-border membrane vesicles. Using orthovanadate, levamisole, and a, p- methylene adenosine diphosphate as possible inhibitors, alkaline phosphatase was shown to be the main agent responsible for ecto-AMP nucleotidase activity. In pig renal basolateral membrane vesicles and in whole cell extracts from pig renal cortex, ecto-AMP nucleotidase was the limiting factor in ATP degradation. Comparing the ATP catabolism in the whole cell cortical extract with the catabo- lism in the same sample precleared of membranes, it was shown that ecto- nucleotidase activity is mainly bound to the membranous components. It is also shown that the whole cell extract of pig renal cortex has hypoxanthine phos- phoribosyl transferase activity, and it seems probable that the rapid and specific formation of luminal inosine and its transport into the cell in competition with adenosine may start the purine salvage pathway through the synthesis of IMP from hypoxanthine. o 1993 WiIey-Liss, ltlc
Cell adenine nucleotides and their metabolites are involved in the regulation of numerous intracellular processes in kidney (Spielman and Arend, 1991). Ade- nosine affects different renal physiological functions such as blood flow (Gouyon and Guignard, 19891, glom- erular filtration rate (Arend et al., 1987), urine flow, and sodium and potassium excretion rate (Collis et al., 19861, glomerular constriction related to the entry of calcium into glomerular cells (Oliveira e t al., 19891, and the renin-angiotensin system (Deray et al., 1990). Adenosine is also important in the maintenance of kid- ney viability before transplantation (Ametani et al., 1990). Most of these adenosine effects are mediated by specific receptors (see Spielman and Arend, 1991 for references).
Part of the intrarenal adenosine production comes from cellular adenosine release. In kidney (Siege1 et al., 1983) as in other tissues (Meghji et al., 1988; Vanden- berghe et al., 1989) production of adenosine, and also inosine and hypoxanthine increases when catabolism of intracellular adenosine triphosphate (ATP) is acceler- ated, for instance in hypoxia and during enhanced en- ergy consumption. On the other hand, the extracellular production of adenosine from nucleotides is also impor- tant ( h l i c et al., 1990). The origin of extracellular nucleotides is unclear, although some come from blood through glomerular filtration, and a significant (I> 1993 WILEY-LISS, INC.
amount may come from tubular cells by exocytosis, since proximal tubular exocytotic vesicles may dis- charge nucleotides into the luminal fluid. culic et al. (1990) demonstrated the presence of ectoenzymes in rat renal cortical brush-border membrane vesicles that produce adenosine as a final product using ATP, ade- nosine diphosphate (ADP), or adenosine monophos- phate (AMP) as substrate. The complete hydrolysis of ATP yielding adenosine is catalyzed by at least two separate ectoenzymes, one capable of hydrolyzing ATP and ADP, but not AMP, and another that hydrolyzes AMP.
Extracellular adenosine acts on adenosine receptors and is then metabolized or internalized into the cell by means of nucleoside uptake systems (Le Hir and Du- bach, 1984; Angielski et al., 1983; Trimble and Coul- son, 1984; Franco et al., 1990). The adenosine uptake is important not only to maintain suitable extracellular nucleoside concentrations but also in the recovery of intracellular nucleotide levels after their depletion (Weinberg et al., 1988). Recovery from hypoxia re-
Received February 26,1993; accepted May 11, 1993. *To whom reprint requestsicorrespondence should be addressed.

BLANCO E'I' AL. 78
quires the restoration of intracellular ATP concentra- tion. Two different pathways are able to restore nucle- otides. One starts from adenosine that is phosphorylated to AMP by adenosine kinase and the other starts from hypoxanthine or adenine that is transformed to IMP or AMP by phosphoribosyl transferase. In both cases availability of intracellular and extracellular sub- strates is required. The extracellular reaction sequence ATP +ADP+ adenosine participates in regulating the time-course of cellular response during crisis or signaling events (Gordon et al., 1989). Mandel et al. (1988) suggested that the increase in kidney cellular ATP requires extracellular adenosine or less effectively inosine and that adenosine obtained from the break- down of AMP may be preferably transported into the renal cells to synthesize AMP. Cadnapaphornchai et al. (1991) demonstrated that extracellular adenosine is es- sential for adenine nucleotide recovery in postischemic cultured renal tubule cells. The NBTI-sensitive nucleo- side uptake has an important role in renal postischemic ATP synthesis (van Waarde et al., 1992).
The aim of this paper was to investigate the possible existence of a rapid and specific nucleotide catabolism in the lumen of pig kidney proximal tubule membranes. Thus, we have determined the enzyme activities in- volved in ATP and adenosine catabolism in isolated pig renal brush-border and basolateral membrane vesicles of proximal tubules and compared them with those of the whole cell extract of pig renal cortex.
MATERIALS AND METHODS Materials
Adenosine, inosine, tetrabutyl ammonium phosphate (TBA) a, p-methyleneadenosine-5'-diphosphate, so- dium orthovanadate, and deoxycoformycin were pur- chased from Sigma. NAD', phosphoribosyl pyrophos- phate (PRPP), ATP, and calf alkaline phosphatase were from Boehringer Mannheim. ADP, AMP, IMP, hypo- xanthine, adenine, uric acid, and 1,1,2 trichloroethane (Freon) were from Merck. Heptane and high perfor- mance liquid chromatography (HPLC) grade acetoni- trile were from Carlo Erba. The remaining products were the best grade available. Deionized water further purified with a Millipore Milli-Q system was used throughout.
Preparation of homogenates Fresh pig kidneys were obtained from the local
slaughterhouse immediately after death of the ani- mals. Cortex, medullae, and papillae were removed af- ter kidney perfusion and dissection. Perfusion buffer, pH 7.4, contained (mM) 10 Tris, 140 NaCl, 4 KC1, 1 CaCl,. Tissues were homogenized in five volumes of 5 mM Tris-HC1 buffer, pH 7.4, containing 10 mM manni- to1 with a Polytron disrupter (Kinematica, PTA 20 TS rotor setting 5). Crude homogenates were immediately used for determination of purine levels or enzyme activ- ity analysis. Aliquots of these homogenates were cen- trifuged at 4"C, 80,000g, 75 min. The supernatants were collected and used as homogenates.
Preparation of luminal and basolateral membranes
Luminal brush-border membranes were isolated, from pig or male Wistar rat kidneys, by a magnesium
precipitation method as described by Lin et al. (1981). Basolateral membranes were isolated from pig kidneys by the method described by Kinsella et al. (1979). In both cases membranes were used immediately for pu- rine level determination or stored at -70°C until en- zyme activity assays. Purity of both preparations was assessed by the enrichment of alkaline phosphatase, EC 3.1.3.1 (brush-border fraction), or Na+-K* ATPase, EC 3.6.1.3 (basolateral fraction) as compared to the starting homogenate. Values of 11-15-fold in alkaline phosphatase activity in brush-border fraction and 12- fold in Na+-K' ATPase in basolateral fraction were obtained.
As reported previously (Blanco et al., 19921, luminal membranes were >85% right-side out vesicles and es- sentially composed of sealed vesicles; thus the activities studied in this paper correspond to ectoenzymes located in the lumen of the proximal tubule. In order to rule out any interference of intravesicular metabolism in some experiments vesicles were filtered through Schleicher and SchuellO.45 pm pore-size filters before HPLC anal- ysis. No significant difference was found when brush- border membrane vesicle suspensions were processed without the filtration step, indicating that intravesicu- lar metabolism is negligible in the presence of extracel- lular metabolism.
Enzyme activity assays Crude homogenates (final concentration 15 mg
protein/ml), the corresponding centrifuged homoge- nates, or membrane fractions (brush-border membrane vesicle final concentration 4 mg proteiniml, and baso- lateral membrane vesicles 3 mg proteinlml) were incu- bated at 25°C in 20 mM HEPES, 100 mM mannitol, 1 mM MgC1, buffer (pH 7.4, unless otherwise indicated) containing different purine compounds: ATP, IMP, ade- nosine, hypoxanthine, uric acid, NAD', or adenine (usually at a final concentration of 1 mM) in the ab- sence or presence of indicated inhibitors. Reaction ki- netics were followed by HPLC analysis. Samples of 100 pl were taken at the indicated times and deproteinized by addition of 100 p1 of 1.2 M perchloric acid solution. After 10 min incubation at O"C, samples were centri- fuged and the supernatants were delipidized by addi- tion of one volume of a mixture of Heptane-Freon (1:4) and neutralized with 0.6 M KOH solution. After cen- trifugation, organic phase and KClO, precipitate were discarded and the aqueous phase was removed and fro- zen until analysis. In all cases, results were corrected at each time for endogenous nucleoside and nucleotide concentration, as measured in a parallel control sample without any addition.
HPLC analysis HPLC analysis was performed using Pharmacia-
LKB HPLC equipment. Purine compounds were quan- tified on 20 p1 aliquots of samples at 254 nm or 310 nm (uric acid), with a 5 pm Spherisorb ODs-2 (PhaseSep) 250 x 4.6 mm reverse-phase column. Elution was per- formed in a binary gradient, buffer A: 30 mM KH,PO,, 7.5 mM TBA, pH 5.45; and buffer B: 30 mM KH,PO,, 7.5 mM TBA, pH 7.0, and acetonitrile 5010 v/v. The elution was a modification of the method described by Hammer et al. (1988) and consisted of three steps. Step
ADENINE NUCLEOTIDE METABOLISM IN KIDNEY 79 ;I Y m v) *
E
10.00 20.00 30.00
Time (min)
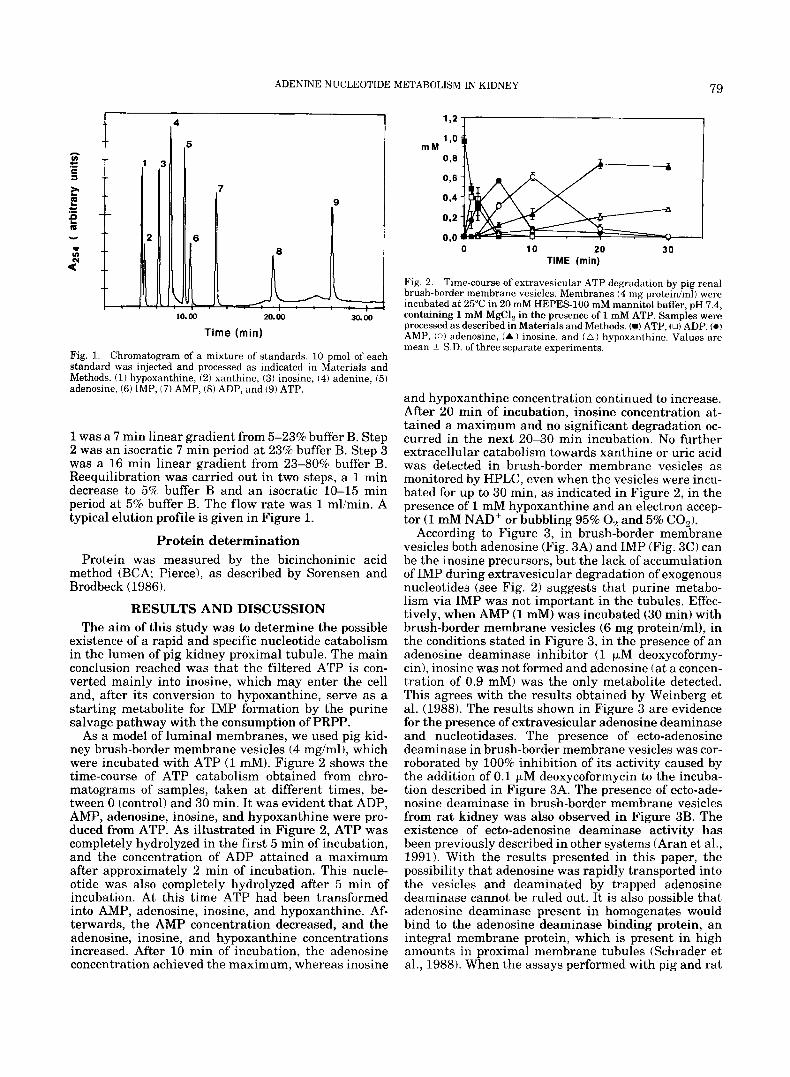
Fig. 1. Chromatogram of a mixture of standards. 10 pmol of each standard was injected and processed as indicated in Materials and Methods. (1) hypoxanthine, (2) xanthine, (3) inosine, (4) adenine, (5) adenosine, (6) IMP, (7) AMP, ( 8 ) ADP, and (9) ATP.
1 was a 7 rnin linear gradient from 5-23% buffer B. Step 2 was an isocratic 7 rnin period at 23% buffer B. Step 3 was a 16 rnin linear gradient from 23-80% buffer B. Reequilibration was carried out in two steps, a 1 min decrease to 5% buffer B and an isocratic 10-15 min period at 5% buffer B. The flow rate was 1 mlimin. A typical elution profile is given in Figure 1.
Protein determination Protein was measured by the bicinchoninic acid
method (BCA; Pierce), as described by Sorensen and Brodbeck (1986).
RESULTS AND DISCUSSION The aim of this study was to determine the possible
existence of a rapid and specific nucleotide catabolism in the lumen of pig kidney proximal tubule. The main conclusion reached was that the filtered ATP is con- verted mainly into inosine, which may enter the cell and, after its conversion to hypoxanthine, serve as a starting metabolite for IMP formation by the purine salvage pathway with the consumption of PRPP.
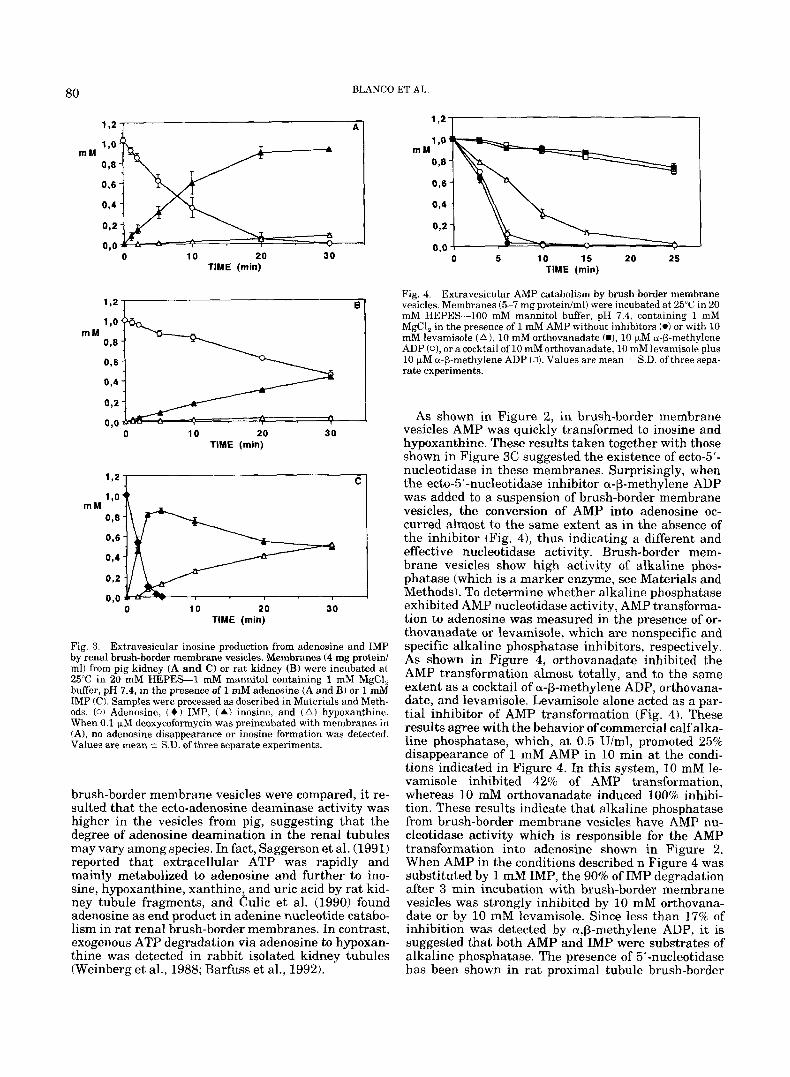
As a model of luminal membranes, we used pig kid- ney brush-border membrane vesicles (4 mgiml), which were incubated with ATP (1 mM). Figure 2 shows the time-course of ATP catabolism obtained from chro- matograms of samples, taken a t different times, be- tween 0 (control) and 30 min. It was evident that ADP, AMP, adenosine, inosine, and hypoxanthine were pro- duced from ATP. As illustrated in Figure 2, ATP was completely hydrolyzed in the first 5 min of incubation, and the concentration of ADP attained a maximum after approximately 2 min of incubation. This nucle- otide was also completely hydrolyzed after 5 min of incubation. At this time ATP had been transformed into AMP, adenosine, inosine, and hypoxanthine. Af- terwards, the AMP concentration decreased, and the adenosine, inosine, and hypoxanthine concentrations increased. After 10 min of incubation, the adenosine concentration achieved the maximum, whereas inosine
0 10 20 30 TIME (min)
Fig. 2. Time-course of extravesicular ATP degradation by pig renal brush-border membrane vesicles. Membranes (4 mg proteiniml) were incubated a t 25°C in 20 mM HEPES-100 mM mannitol buffer, pH 7.4, containing 1 mM MgCl, in the presence of 1 mM ATP. Samples were processed as described in Materials and Methods. (W ATP, (0 ) ADP, ( 0 )
AMP, (0) adenosine, (A 1 inosine, and ( A ) hypoxanthine. Values are mean i S.D. of three separate experiments.
and hypoxanthine concentration continued to increase. After 20 min of incubation, inosine concentration at- tained a maximum and no significant degradation oc- curred in the next 20-30 min incubation. No further extracellular catabolism towards xanthine or uric acid was detected in brush-border membrane vesicles as monitored by HPLC, even when the vesicles were incu- bated for up to 30 min, as indicated in Figure 2, in the presence of 1 mM hypoxanthine and an electron accep- tor (1 mM NAD' or bubbling 95% 0, and 5% C 0 2 ) .
According to Figure 3, in brush-border membrane vesicles both adenosine (Fig. 3A) and IMP (Fig. 3C) can be the inosine precursors, but the lack of accumulation of IMP during extravesicular degradation of exogenous nucleotides (see Fig. 2) suggests that purine metabo- lism via IMP was not important in the tubules. Effec- tively, when AMP (1 mM) was incubated (30 min) with brush-border membrane vesicles (6 mg protein/ml), in the conditions stated in Figure 3, in the presence of an adenosine deaminase inhibitor ( 1 pM deoxycoformy- cin), inosine was not formed and adenosine (at a concen- tration of 0.9 mM) was the only metabolite detected. This agrees with the results obtained by Weinberg et al. (1988). The results shown in Figure 3 are evidence for the presence of extravesicular adenosine deaminase and nucleotidases. The presence of ecto-adenosine deaminase in brush-border membrane vesicles was cor- roborated by 100% inhibition of its activity caused by the addition of 0.1 FM deoxycoformycin to the incuba- tion described in Figure 3A. The presence of ecto-ade- nosine deaminase in brush-border membrane vesicles from rat kidney was also observed in Figure 3B. The existence of ecto-adenosine deaminase activity has been previously described in other systems (Aran et al., 1991). With the results presented in this paper, the possibility that adenosine was rapidly transported into the vesicles and deaminated by trapped adenosine deaminase cannot be ruled out. It is also possible that adenosine deaminase present in homogenates would bind to the adenosine deaminase binding protein, an integral membrane protein, which is present in high amounts in proximal membrane tubules (Schrader et al., 1988). When the assays performed with pig and rat
80 BLANCO ET AL.
mM
mM
-.- 0 10 20 30
TIME (rnin)
0 10 20 30 TIME (mh)
0 10 20 30 TIME (min)
Fig. 3. Extravesicular inosine production from adenosine and IMP by renal brush-border membrane vesicles. Membranes (4 mg protein/ ml) from pig kidney (A and C) or rat kidney (B) were incubated a t 25°C in 20 mM HEPES-1 mM mannitol containing 1 mM MgCI, buffer, pH 7.4, in the presence of 1 mM adenosine (A and B) or 1 mM IMP (GI. Samples were processed as described in Materials and Meth- ods. (0) Adenosine, ( + ) IMP, (A) inosine, and (a) hypoxanthine. When 0.1 pM deoxycoformycin was preincubated with membranes in (A), no adenosine disappearance or inosine formation was detected. Values are mean i_ S.D. of three separate experiments.
brush-border membrane vesicles were compared, i t re- sulted that the ecto-adenosine deaminase activity was higher in the vesicles from pig, suggesting that the degree of adenosine deamination in the renal tubules may vary among species. In fact, Saggerson et al. (1991) reported that extracellular ATP was rapidly and mainly metabolized to adenosine and further to ino- sine, hypoxanthine, xanthine and uric acid by rat kid- ney tubule fragments, and h l i c e t al. (1990) found adenosine as end product in adenine nucleotide catabo- lism in rat renal brush-border membranes. In contrast, exogenous ATP degradation via adenosine to hypoxan- thine was detected in rabbit isolated kidney tubules (Weinberg et al., 1988; Barfuss et al., 1992).
0 5 10 15 20 25 TIME (min)
Fig. 4. Extravesicular AMP catabolism by brush-border membrane vesicles. Membranes (5-7 mg protein/mll were incubated at 25°C in 20 mM HEPES--100 mM mannitol buffer, pH 7.4, containing 1 mM MgCI, in the presence of 1 mM AMP without inhibitors (0 ) or with 10 mM levamisole (A), 10 mM orthovanadate (m), 10 pM a-P-methylene ADP (01, or a cocktail of10 mM orthovanadate, 10 mM levamisole plus 10 JLM a-P-methylene ADP (n). Values are mean -t S.D. of three sepa- rate experiments.
As shown in Figure 2, in brush-border membrane vesicles AMP was quickly transformed to inosine and hypoxanthine. These results taken together with those shown in Figure 3C suggested the existence of ecto-5'- nucleotidase in these membranes. Surprisingly, when the ecto-5'-nucleotidase inhibitor a-P-methylene ADP was added to a suspension of brush-border membrane vesicles, the conversion of AMP into adenosine oc- curred almost to the same extent as in the absence of the inhibitor (Fig. 4), thus indicating a different and effective nucleotidase activity. Brush-border mem- brane vesicles show high activity of alkaline phos- phatase (which is a marker enzyme, see Materials and Methods). To determine whether alkaline phosphatase exhibited AMP nucleotidase activity, AMP transforma- tion to adenosine was measured in the presence of or- thovanadate or levamisole, which are nonspecific and specific alkaline phosphatase inhibitors, respectively. As shown in Figure 4, orthovanadate inhibited the AMP transformation almost totally, and to the same extent as a cocktail of a-6-methylene ADP, orthovana- date, and levamisole. Levamisole alone acted as a par- tial inhibitor of AMP transformation (Fig. 4). These results agree with the behavior of commercial calf alka- line phosphatase, which, a t 0.5 Ulml, promoted 25% disappearance of 1 mM AMP in 10 min at the condi- tions indicated in Figure 4. In this system, 10 mM le- vamisole inhibited 42% of AMP transformation, whereas 10 mM orthovanadate induced 100% inhibi- tion. These results indicate that alkaline phosphatase from brush-border membrane vesicles have AMP nu- cleotidase activity which is responsible for the AMP transformation into adenosine shown in Figure 2. When AMP in the conditions described n Figure 4 was substituted by 1 mM IMP, the 90% of IMP degradation after 3 min incubation with brush-border membrane vesicles was strongly inhibited by 10 mM orthovana- date or by 10 mM levamisole. Since less than 17% of inhibition was detected by a$-methylene ADP, it is suggested that both AMP and IMP were substrates of alkaline phosphatase. The presence of 5'-nucleotidase has been shown in rat proximal tubule brush-border

81 ADENINE NUCLEOTIDE METABOLISM IN KIDNEY
membrane vesicles (culic e t al., 1990; Le Hir et al., 1985), but the data reported here with pig brush-border membrane vesicles together with the results of Wein- berg et al. (1988) with rabbit renal brush-border mem- brane vesicles indicate that ecto-5'-nucleotidase from these sources, if it exists, has a relative minor role in exogenous AMP catabolism under normoxic conditions. On the other hand, the nucleotidase activity of alkaline phosphatase is controversial. Some years ago, Une- mori-Aikawa (1971) described an alkaline phosphatase in the clam Meretrix meretrix lusoria with affinity for nucleotides, including AMP. Felix and Fleisch (1974) found pyrophosphatase and ATPase activity in purified calf bone alkaline phosphatase. More recently, Hilden and Madias (1990) described an alkaline phosphatase in canine kidney brush-border membranes that can act as ecto-ATPase. Further, Weinberg et al. (1988) pointed out the existence of nonspecific phosphatases as responsible for AMP degradation in isolated rabbit kidney tubules. These results agree with those de- scribed in this paper.
For comparative purposes, the ATP catabolism in pig kidney basolateral membrane vesicles was studied and the results are shown in Figure 5A. Comparing the kinetics in these two types of preparations, there was, in contraluminal vesicles, a slower decay of AMP (at 30 min the AMP concentration was still significant). Ade- nosine (1 mM) transformation into inosine (Fig. 5B) and the IMP (1 mM) transformation to inosine (Fig. 5C) were lower than those observed with brush-border membrane vesicles. These results agree with the AMP accumulation observed in basolateral membrane vesi- cles when exogenous ATP catabolism was determined (Fig. 5A). In these membranes, the alkaline phos- phatase activity was very low (less than 7% of alkaline phosphatase activity found in brush-border membrane vesicles).
Similar experiments were performed with crude re- nal cortex extract (15 mg/ml) (Fig. 6A). In this case, AMP concentration after 30 min of incubation was high, whereas di- and trinucleotides were undetectable after 10 min of incubation. The limiting step was also the AMP-nucleotidase activity, as indicated by the low concentration of adenosine and the slow degradation of AMP. When the same experiment was performed using a centrifuged aliquot of crude renal cortex extract, the results obtained were those described in Figure 6B. The apparent lack of AMP degradation should be noted, as well as the time required to transform the ATP into ADP. These results provide evidence that the nucleoti- dase activities in renal cortical extracts are mainly as- sociated to membrane fractions and indicate the ab- sence of cytosolic 5'-nucleotidase activity with affinity for AMP in pig renal cortex in these conditions. This could be due to the absence of the enzyme or to its inhibition by the excess of added ATP and ADP. To test this, 1 mM AMP or 1 mM IMP was added instead of 1 mM ATP in the conditions described in Figure 6B. Af- ter 20 min, less than 10% AMP disappearance was de- tected and less than 30% IMP was transformed, indicat- ing low cytosoluble 5'-nucleotidase activity. As concluded by Pie and Le Hir (1991) the low activity detected may represent ecto-5'-nucleotidase or alkaline phosphatase partially solubilized from membranes.
30 l o TIME (mi$' 0
m M
:::; 0.0 - I ~
0 10 20 30 TIME (min)
0 10 20 30 Time (min)
Fig. 5. Extravesicular metabolism in basolateral membranes. Mem- branes (4 mg proteiniml) were incubated a t 25°C in 20 mM H E P E S 100 mM mannitol buffer, pH 7.4, containing 1 mM MgCI, in the presence of (A) 1 mM ATP, (B) 1 mM adenosine, and (C) 1 mM IMP. Samples were processed a s described in Materials and Methods. (m) ATP, (0) ADP, (0 ) AMP, (0) adenosine, (+ ) IMP, (A 1 inosine, and ( A ) hypoxanthine. Values are mean 2 S.D. of three separate experiments.
As in brush-border or basolateral membrane vesicles, xanthine and uric acid formed from ATP or adenosine were undetectable in crude extracts of renal cortex. Since controversial results are reported in the litera- ture about kidney xanthine oxidase, the presence of xanthine oxidase or xanthine dehydrogenase activity in different kidney extracts was determined. Hypoxan- thine (1 mM) was incubated in 50 mM Tris-HC1 buffer, pH 7.4, with crude renal cortex (15 mg/ml), medulla (14 mg/ml), and papilla (4.5 mg/ml) extract (see Materials and Methods) in the absence or in the presence of oxy- gen (95% 0, plus 5% COz) or NADi (1 mM). No evi- dence of xanthine oxidase or xanthine dehydrogenase activity was obtained due to the lack of xanthine or uric acid formation measured by HPLC. This result agrees with those reported for rabbit renal tubules (Weinberg

82 BLANCO ET AL.
0 10 20 30 TIME (min)
0 20 40 60 80 100 120 TIME (rnin)
Fig. 6. Time-course of ATP degradation by renal cortex homoge- nates. A Crude cortex homogenates (15 mg proteiniml) or B: superna- tant of a centrifuged aliquot were incubated a t 25°C in the homogeni- zation buffer containing 1 mM MgCI, in the presence of 1 mM ATP. Samples were processed as described in Materials and Methods. (a) ATP, (0) ADP, (*) AMP, (0) adenosine, (A) inosine, ( A ) hypoxanthine, and ( + ) sum of nucleosides. Values are mean k S.D. of three separate experiments.
et al., 1988) and oxygen-deprived rabbit renal tubules (Buhl, 1982). In contrast, other reports suggest the presence of these activities in either isolated rat corti- cal tubules (Saggerson et al., 1991), ischemic rat cortex (Arnold et a]., 1986), perfused rat kidneys (Trimble and Coulson, 1984), or guinea pig kidney slices under normoxic conditions (Blanco et al., 1990).
Taken together, our results indicate, that in brush- border membrane vesicles, there is a rapid and specific formation of inosine from ATP. The physiological role of this transformation is unknown, but one hypothesis is that the filtered nucleotides may be transformed into inosine, enter the cell by the Na+-dependent and Na'- independent nucleoside transporters described for this system (Le Hir and Dubach, 1984; Trimble and Coul- son, 1984; Franco et al., 1990), and be reutilized via the purine salvage pathway to IMP with the sole consump- tion of PRPP and phosphate. To test this hypothesis we designed an experiment in which PRPP (2 mM) and adenine (1 mM) were incubated with brush-border membrane vesicles (4 mgiml) or crude renal cortex ex- tract (15 mg/ml) in a 50 mM Tris-HC1 buffer (pH 7.1) containing MgC1, (20 mM). In agreement with the hy- pothesis that extracellular purine salvage should not
occur, neither IMP nor AMP was detected in brush- border membrane vesicles. However, by the significant rise of AMP (0.12 mM) and IMP (0.12 mM) concentra- tions found when homogenates were incubated (30 min) with PRPP (2 mM) and adenine (0.2 mM) in the pres- ence of 0.36 mM of endogenous hypoxanthine, it was demonstrated that the purine salvage pathway func- tions in kidney. It should be noted that the homogenate already presented high concentrations of hypoxanthine to be reutilized by hypoxanthine-guanine phosphoribo- syl transferase. Although significant, the purine sal- vage pathway for hypoxanthine and adenine is low if compared with the high level of nucleotide catabolism. As stated by Weinberg et al. (19881, adenine was the main precursor of ATP, via the salvage pathway, in intact rabbit proximal tubules, probably denoting qual- itative differences between species. Our results are con- sistent with a greater necessity for rapid disappearance of nucleotides on the luminal side of tubules and their further transformation, mainly into inosine and, to a lesser extent, into adenosine and hypoxanthine. The adenosine so formed can stimulate A, adenosine recep- tors in brush-border membranes (Blanco et al., 1992) prior to its incorporation to the cell. The inosine and hypoxanthine may also be transported into the cells and transformed, via hypoxanthine, to IMP with the consumption of PRPP.
ACKNOWLEDGMENTS This work was supported by grant no. 91-0272 from
FIS-SS from Ministerio de Sanidad y Consumo from which Joan Sayos is recipient of a fellowship. Julia Blanco is recipient of a fellowship from the Program Formacion y Perfeccionamiento del Personal Investiga- dor from the Ministerio de Eduacion y Ciencia of the Spanish government.
LITERATURE CITED Ametani, M.S., Southard, J.H., and Belzer, F.O. (1990, Importance of
glutathione and adenosine in cold storage of the kidney. Transplant. Proc., 22r469A71.
Angielski, S., Le Hir, M., and Dubach, U.C. (1983) Transport of ade- nosine by renal brush-border membranes. Pflugers Arch., 397r75- 77.
Aran, J.M., Colomer, D., Matutes, E., Vives-Corrons, J.L., and Franco, R. (1991) Presence of adenosine deaminase on the surface of mono- clonal blood cells: Immunochemical localization using light and electron microscopy. J. Histochem. Cytochem., 39t1001-1008.
Arend, L.J., Bakris, G.L., Burnett Jr, J.C., and Megerian, C. (1987) Role of intrarenal adenosine in the renal hemodynamic response to contrast media. J. Lab. Clin. Med., I10:406411.
Arnold, P.E., van Putten, V.J., Lumlertgul, D., Burke, T.J., and Schrier, R.W. (1986) Adenine nucleotide metabolism and mitochon- drial Ca" ' transport following renal ischemia. Am. J . Physiol., 250:F357-F363.
Barfuss, D.W., McCann, W.P., and Katholi, R.E. (1992) Axial hetero- geneity of adenosine transport and metabolism in the rabbit proxi- mal tubule. Kidney Int., 41:1143-1149.
Blanco, J., Mallol, J., Lluis, C., Canela, E.I., and Franco, R. (1990) Adenosine metabolism in kidney slices under normoxic conditions. J. Cell. Physiol., 143r344-351.
Blanco, J., Canela, E.I., Mallol, J., Lluis, C., and Franco, R. (1992) Characterization of adenosine receptors in brush-border mem- branes from pig kidney. Br. J. Pharmacol., 107t671-678.
Buhl, M.R. (1982) Purine metabolism in ischemic kidney tissue. Dan. Med. Bull., 29:l-26.
Cadnapaphornchai, P., Kellner, D., Golembieski, A., and McDonald, F.D. (1991) Role of adenosine and theophylline on the recovery of adenine nucleotides in postischemic cultured renal tubular cells. J . Pharmacol. Exp. Ther., 257:774-780.

83 ADENINE NUCLEOTIDE METABOLISM IN KIDNEY
Collis, M.G., Baxter, G.S., and Keddie, J.R. (1986) The adenosine receptor antagonist 8-phenyltheophylline, causes diuresis and sali- uresis in the rat. J. Pharm. Pharmacol., 38:850-852.
Culic, O., Sabolic, I., and ganic-GrubiSic, T. 11990) The stepwise hy- drolysis of adenine nucleotides by ectoenzymes of rat renal brush- border membranes. Biochim. Biophys. Acta, 1030.143-151.
Deray, G., Sabra, R., Jackson, H.E.K., and Branch, R.A. (1990) Inter- action between angiotensin I1 and adenosine in mediating the vaso- constrictor response to intrarenal hypertonic saline infusion in the dog. J. Pharmacol. Exp. Ther., 252t631-635.
Felix, R., and Fleisch, H. (1974) The pyrophosphatase and (Ca"'- Mg' ' 1-ATPase activity of purified calf bone alkaline phosphatase. Biochim. Biophys. Acta, 350:84-94.
Franco, R., Centelles, J .J . , and Kinne, R.K.H. (1990) Further charac- terization of adenosine transport in renal brush-border membranes. Biochim. Biophys. Acta, 1024~241-248.
Gordon, E.L., Pearson, J.D., Dickinson, E.S., Moreau, D., and Slakey, L.L. (1989) The hydrolysis of extracellular adenine nucleotides by arterial smooth muscle cells. J. Biol. Chem., 264t1898618992.
Gouyon, J.-B., and Guignard, J.-P. 11989) Adenosine in the immature kidney. Dev. Pharmacol. Ther., 13t113-119.
Hammer, D.F., Unverferth, D.V., Kelley, R.E., Harvan, P.A., and Altschuld, R.A. (1988) Extraction and measurement of myocardial nucleotides, nucleosides and purine bases by high performance liq- uid chromatography. Anal. Biochem., 269t300-305.
Hilden, S.A., and Madias, N.E. (1990) Stimulation of canine kidney BBMV ATPase activity by acid pH in the presence of Zn2 ' . Membr. Biochem., 9t68-81.
Kinsella, J.L., Holohan, P.D., Pessaii, N.I., andRoss, C.R. (1979) Isola- tion of luminal and antiluminal membranes from dog kidney cortex. Biocbim. Biophys. Acta, 552:46&477.
Le Hir, M., and Dubach, U.C. (1984) Sodium gradient-energized con- centrative transport of adenosine in renal brush-border vesicles. Pfliigers Arch., 4OZr58-63.
Le Hir, M., Angielski, S., and Dubach, U.C. (1985) Properties of an ecto-5'-nucleotidase of the renal brush-border. Renal Physiol., 8t321-327.
Lin, J.T., Da Cruz, M.E.M., Riedel, S., and Kinne, R. (1981) Partial purification of dog kidney sodium-D-glucose cotransport system by affinity chromatography on a phlorttzin polymer. Biochim. Biophys. Acta, 64Ot43-54.
Mandel, L.J., Takano, T.S., Soltoff, S.P., and Murdaugh, S. (1988) Mechanism whereby exogenous adenine nucleotides improve rabbit renal proximal function during and after anoxia. J. Clin. Invest., 4t1255-1264.
Meghji, P., Middleton, K.M., and Newby, A.C. (1988) Absolute rates of adenosine formation during ischaemia in rat and pigeon hearts. Biochem. J. , 249r695-703.
Oliveira, A., Lamas, S., Rodriguez-Puyol, D., and L6pez-Novoa, J.M. (1989) Adenosine induces mesangial cell contraction by A,-type re- ceptors. Kidney Int., 35.1300-1305.
Pie, G., and Le Hir, M. (1991) The soluble "low K," 5'-nucleotidase of rat kidney represents solubilized ecto-5'-nucleotidase. Biochem. J., 273t409-413.
Saggerson, E.D., Carpenter, C.A., and Veiga, J.A.S. (1991) Stimula- tion of renal gluconeogenesis by exogenous adenine nucleotides. Biochim. Biophys. Acta, 755:119-126.
Schrader, W.P., Miczek, A.D., West,C.A., andSamsonoff, W.A. (1988) Evidence for receptor-mediate uptake of adenosine deaminase in rabbit kidney. J. Histochem. Cytochem., 36:1481-1487.
Siegel, N.J., Avison, H.J., Reilly, H.F., Alger, J.R., and Shulman, R.G. 11983) Enhanced recovery of renal ATP-MgCI, determined by '"P- NMR. Am. J. Physiol., 245:F530-F534.
Sorensen, K., and Brodbeck, U. (1986) A sensitive protein assay method using micro-titer platters. Experientia, 42r161-162.
Spielman, W.S., and Arend, L.J. (1991) Adenosine receptors and sig- naling in the kidney. Hypertension, 17.117-130,
Trimble, M.E., and Coulson, E. (1984) Adenosine transport in perfused rat kidney and renal cortical membrane vesicles. Am. J. Physiol., 246.F79PF803.
Unemori-Aikawa, Y. (1971) An alkaline phosphatase in the clam Mer- etrix rneretrix Lusoria (Gmelin), with affinities for nucleotides. Comp. Biochem. Physiol., 40Br347-358.
Vandenberghe, G., Vincent, M.F., and Bontemps, F. (1989) Pathways and control of adenine nucleotide catabolism in anoxic rat hepato- cytes. Biomed. Biophys. Acta, 48tS5-Sl0.
van Waarde, A,, Avison, M.J., Thulin, G., Gaudio, K.M., Schulman, R.G., and Siegel, N.J. (1992) Role of nucleoside uptake in renal postischemic ATP synthesis. Am. J. Physiol., 262tF109ZF1099.
Weinberg, J.M., Davis, J.A., Lawton, A., and Abarzua, M. (1988) Mod- ulation of cell nucleotide levels of isolated kidney tubules. Am. J. Physiol., 254:F311-F322.


















