
Matrix degradability controls multicellularity of 3D cell ...
date post
20-Dec-2015Category
view
213download
0
Abstract: The volvocine algae (Volvox and its close relatives) represent a unique opportunity for the study of origins of multicellularity. Several major evolutionary transitions have occurred within this relatively young group, which spans a large range of sizes and several levels of complexity. To explore the historical and adaptive bases of these major evolutionary transitions, we inferred phylogenetic relationships in the colonial volvocine algae using DNA sequence data. Using maximum parsimony reconstructions of ancestral character states, we traced the evolution of key innovations such as coloniality, large size, cellular differentiation, and novel developmental programs.
Origins of multicellularity
• Multicellularity has evolved several times independently.
• e.g. plants, animals, fungi, red and brown algae
• Complex multicellular organisms have functional specialization of cells.
• Specialization has a cost: some cells are specialized for functions other than reproduction, lowering fecundity.• This cost, along with multiple independent origins, suggests that multicellularity provides a substantial benefit.
References:1. A. G. Desnitski, Eur. J. Protistol. 31, 241 (1995). 2. D. Posada, K. Crandall, Bioinformatics 14, 817 (1998).3. F. Ronquist, J. P. Huelsenbeck, Bioinformatics 19, 1572 (2003). 4. W. P. Maddison, D. R. Maddison. http://mesquiteproject.org (2004).
Acknowledgements:We thank C. Solari for providing photographs and helpful advice, M. Buchheim for generously providing unpublished DNA sequences, and A. Coleman and H. Nozaki for helpful advice.
Conclusions:• Single origin of coloniality, no evidence of reversal to unicellular condition.• Most recent common ancestor of the Goniaceae and Volvocaceae was likely 16- or 32-celled. • At least four independent origins of large size (>32 cells), with at least two reductions in cell number.• At least three separate origins of germ-soma differentiation (partial or complete).• One possible reversal from complete to partial differentiation.• At least two losses of the ancestral developmental program.
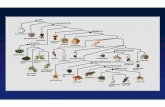
Figure 1. Subset of volvocine species: (A) Chlamydomonas reinhardtii – one undifferentiated cell, (B) Gonium pectorale – a clump of 8 undifferentiated cells, (C) Eudorina elegans – a spherical colony of 32 undifferentiated cells, (D) Pleodorina californica – a spherical colony of 32 cells of which a few are terminally differentiated, (E) Volvox carteri – a spherical colony of up to 4000 terminally differentiated somatic cells and a few specialized reproductive cells, and (F) Volvox aureus – a spherical colony of up to 2000 undifferentiated cells and a few specialized reproductive cells. Photos by C. Solari.
Volvocine green algae (Volvox and its close relatives) are an ideal system to study origins of multicellularity.
• More recent origin (35-60 MYA) than more complex multicellular groups
• Diverse group with unicellular, multicellular, and several intermediate stages
• Easily cultured, rapid generations
• Well-studied ecology, cytology, genetics, development
• Possible multiple origins of characters related to multicellularity
Major evolutionary transitions in the volvocine algae:
• From unicellular to colonial
• From simple clumps of cells to structured hollow spheres
• From undifferentiated cells to partial specialization
• From partial specialization to complete separation of reproductive and somatic functions
• Among sizes that span five orders of magnitude
• In Volvox, among four distinct developmental programs(1)
Methods:
• DNA sequences from five chloroplast genes (atpB, psaA, psaB, psbC, rbcL) combined into a supermatrix
• Best-fit model of nucleotide substitution inferred in Modeltest(2)
• Phylogeny inferred using Bayesian methods in MrBayes(3)
• Posterior probabilities calculated from 3.0 x 106 post-burnin generations
• Ancestral character states inferred using maximum parsimony in Mesquite(4)
Figure 3. Three levels of germ-soma differentiation mapped onto the Bayesian phylogeny. At least three separate origins of partial or complete functional specialization of cells are evident. Pleodorina californica and P. japonica may represent a reversal from G/S to GS/S.
Figure 2. Numbers of cells mapped onto the Bayesian phylogeny. At least four separate origins of large size (>32 cells) are evident. Gonium octonarium and Eudorina cylindrica represent probable reductions in cell number.
Figure 4. Four developmental programs(1) mapped onto the Bayesian phylogeny: (1), gonidia (reproductive cells) grow large and divide rapidly without further growth to produce equal-sized daughter cells; (2), an early asymmetric division creates large cells that develop into gonidia; (3), similar to (1) but divisions are slow; (4), gonidia are small and grow between cell divisions, which are symmetrical. At least two independent losses of the ancestral developmental program (program 1) are evident.
5%
Chlamydomonas moewusiiChlamydomonas reinhardtii
Chlamydomonas debaryanaVitreochlamys gloeocystiformisVitreochlamys ordinata
Vitreochlamys pinguisVitreochlamys aulata
Basichlamys sacculiferaTetrabaena socialis
Astrephomene perforataAstrephomene gubernaculifera
Gonium pectoraleGonium octonarium
Gonium quadratumGonium multicoccum
Gonium viridistellatumPlatydorina caudata
Volvox globatorVolvox barberi
Volvox rousselettiiVolvulina steinii
Volvulina compactaVolvulina pringsheimii
Pandorina morum APandorina colemaniae
Pandorina morum BPandorina morum C
Volvulina boldiiYamagishiella unicocca
Eudorina elegans AEudorina minodii
Eudorina elegans BEudorina unicocca
Volvox gigasEudorina elegans C
Pleodorina indicaPleodorina illinoisensis AEudorina elegans D
Pleodorina illinoisensis BEudorina cylindrica
Pleodorina californica
Volvox aureusVolvox africanus
Volvox dissipatrixVolvox tertius
Volvox obversusVolvox carteri f. nagariensis
Volvox carteri f. kawasakiensisVolvox carteri f.weismannia
Pleodorina japonica
99
80
99
98
8160
8599
2
4
1
Unknown
3
5%
Chlamydomonas moewusiiChlamydomonas reinhardtii
Chlamydomonas debaryanaVitreochlamys gloeocystiformisVitreochlamys ordinata
Vitreochlamys pinguisVitreochlamys aulata
Basichlamys sacculiferaTetrabaena socialis
Astrephomene perforataAstrephomene gubernaculifera
Gonium pectoraleGonium octonarium
Gonium quadratumGonium multicoccum
Gonium viridistellatumPlatydorina caudata
Volvox globatorVolvox barberi
Volvox rousselettiiVolvulina steinii
Volvulina compactaVolvulina pringsheimii
Pandorina morum APandorina colemaniae
Pandorina morum BPandorina morum C
Volvulina boldiiYamagishiella unicocca
Eudorina elegans AEudorina minodii
Eudorina elegans BEudorina unicocca
Volvox gigasEudorina elegans C
Pleodorina indicaPleodorina illinoisensis AEudorina elegans D
Pleodorina illinoisensis BEudorina cylindrica
Pleodorina californica
Volvox aureusVolvox africanus
Volvox dissipatrixVolvox tertius
Volvox obversusVolvox carteri f. nagariensis
Volvox carteri f. kawasakiensisVolvox carteri f.weismannia
GS/S: Undifferentiated cells (GS) and somatic cells (S)
G/S: Separate germ cells and somatic cells
GS: Undifferentiated cells (Germ-Soma)
Unknown
Pleodorina japonica
99
80
99
98
8160
8599
A Comparative Study of the Evolution ofMulticellularity in the VolvocalesMatthew D. Herron and Richard E. Michod
Department of Ecology and Evolutionary Biology, University of Arizona
5%
Chlamydomonas moewusii (1)Chlamydomonas reinhardtii (1)
Chlamydomonas debaryana (1)Vitreochlamys gloeocystiformis (1)Vitreochlamys ordinata (1)
Vitreochlamys pinguis (1)Vitreochlamys aulata (1)
Basichlamys sacculifera (4)Tetrabaena socialis (4)
Astrephomene perforata (128)Astrephomene gubernaculifera (128)
Gonium pectorale (16)Gonium octonarium (8)
Gonium quadratum (16)Gonium multicoccum (32)
Gonium viridistellatum (16)Platydorina caudata (32)
Volvox globator (16,000)Volvox barberi (32,000)
Volvox rousselettii (32,000)Volvulina steinii (16)
Volvulina compacta (16)Volvulina pringsheimii (16)
Pandorina morum A (16)Pandorina colemaniae (16)
Pandorina morum B (16)Pandorina morum C (16)
Volvulina boldii (16)Yamagishiella unicocca (32)
Eudorina elegans A (32)Eudorina minodii (32)
Eudorina elegans B (32)Eudorina unicocca (32)
Volvox gigas (2000)Eudorina elegans C (32)
Pleodorina indica (64)Pleodorina illinoisensis A (32)Eudorina elegans D (32)
Pleodorina illinoisensis B (32)Eudorina cylindrica (16)
Pleodorina californica (128)
Volvox aureus (2000)Volvox africanus (8000)
Volvox dissipatrix (16,000)Volvox tertius (1000)
Volvox obversus (2000)Volvox carteri f. nagariensis (4000)
Volvox carteri f. kawasakiensis (2000)Volvox carteri f.weismannia (2000)
1
4
>32
32
16
8
Unknown
Pleodorina japonica (128)
99
80
99
98
8160
8599
Tetrabaenaceae
Volvocaceae
Goniaceae
Chlamydomonadaceae




![Metachronal waves in the flagellar beating of Volvox and ... · colonial alga Volvox carteri is an ideal model organism for the study of flagella-driven flows [35]. Volvox comprises](https://static.fdocuments.in/doc/165x107/5fb2e1b931572466d6768af3/metachronal-waves-in-the-flagellar-beating-of-volvox-and-colonial-alga-volvox.jpg)












