
About the Partners - CGIARciat-library.ciat.cgiar.org/Articulos_Ciat/9586940748.pdf · About the...
366
Transcript of About the Partners - CGIARciat-library.ciat.cgiar.org/Articulos_Ciat/9586940748.pdf · About the...


About the Partners
CIATThe International Center for Tropical Agriculture (CIAT) is a not-for-profit organization that conductssocially and environmentally progressive research aimed at reducing hunger and poverty andpreserving natural resources in developing countries. CIAT is one of the 15 centers funded mainlyby the 58 countries, private foundations, and international organizations that make up theConsultative Group on International Agricultural Research (CGIAR).
IITAIITA’s mission is to enhance the food security, income, and well-being of resource-poor people insub-Saharan Africa by conducting research and related activities to increase agricultural production,improve food systems, and sustainably manage natural resources, in partnership with nationaland international stakeholders.
AVRDCAVRDC is the principal international center for vegetable research and development in the world.Its mission is to reduce poverty and malnutrition in developing countries through improvedproduction and consumption of vegetables.
CIPThe International Potato Center (CIP) seeks to reduce poverty and achieve food security on asustained basis in developing countries through scientific research and related activities onpotato, sweetpotato, and other root and tuber crops and on the improved management of naturalresources in the Andes and other mountain areas.
ICIPEICIPE’s mission is to help alleviate poverty, ensure food security and improve the overall healthstatus of peoples of the tropics by developing and extending management tools and strategies forharmful and useful arthropods, while preserving the natural resource base through research andcapacity building.
About the Donors
DANIDAReducing poverty in developing countries is central to Danish development cooperation priorities.A number of crosscutting themes are built into DANIDA’s development assistance: women’sparticipation in development, the environment, promotion of democracy and observation of humanrights. These crosscutting themes are integrated into DANIDA’s development activities more generally.
DFIDThe United Kingdom’s Department for International Development (DFID) supports research in thesustainable management of natural resources that aims at improving the livelihoods of poor peoplein developing countries. DFID has funded commodity-based research on whiteflies and whitefly-transmitted viruses, primarily in Africa, for nearly a decade through its centrally funded CropProtection Programme (CPP). It is now aiming to build on this support by helping to create a globalframework that will allow different whitefly research projects and activities to coordinate theirefforts more effectively and produce research outputs which have positive and sustainable impactson poor peoples’ livelihoods.
NZAIDNZAID is New Zealand’s agency responsible for international assistance to developing countries.New Zealand’s aid helps to eliminate poverty and create a safe and just world, particularly in thePacific region.
USAIDUSAID is an independent federal government agency that receives overall foreign policy guidancefrom the Secretary of State. USAID supports long-term and equitable economic growth and advancesU.S. foreign policy objectives by supporting: economic growth, agriculture and trade; globalhealth; and, democracy, conflict prevention and humanitarian assistance.
USDAProvides leadership on food, agriculture, natural resources, and related issues based on soundpublic policy, the best available science, and efficient management.

ISBN 958-694-074-8
Base for Global Action
Edited by:Pamela K. Anderson and
Francisco J. Morales
With the collaboration ofAnnie L. Jones and Richard H. Markham
Whiteflyand Whitefly-borneViruses in the Tropics:Building a Knowledge

Centro Internacional de Agricultura TropicalInternational Center for Tropical AgricultureApartado Aéreo 6713Cali, ColombiaFax: +57 (2) 4450073E-mail: [email protected]
CIAT Publication No. 341ISBN 958-694-074-8Press run: 500Printed in ColombiaAugust 2005
Whitefly and whitefly-borne viruses in the tropics: Building a knowledge base for global action /edited by Pamela K. Anderson and Francisco J. Morales; with the collaboration of Annie L. Jonesand Richard H. Markham. Cali, CO : Centro Internacional de Agricultura Tropical (CIAT), 2005.351 p. -- (CIAT publication no. 341)ISBN 958-694-074-8
AGROVOC descriptors in English:1. Manihot esculenta. 2. Ipomoea batatas. 3. Sweet potatoes. 4. Vegetables. 5. Legumes.
6. Pest insects. 7. Aleyrodidae. 8. Bemisia tabaci. 9. Trialeurodes vaporariorum.10. Aleurotrachelus socialis. 11. Trialeurodes variabilis. 12. Bemisia tuberculata. 13. Trialeurodesabutiloneus. 14. Aleurocanthus woglumi. 15. Cassava mosaic virus. 16. Geminiviruses. 17. Vectors.18. Food security. 19. Poverty. 20. Pest control. 21. Disease control. 22. Biological control.23. Integrated control. 24. Parasitoids. 25. Natural enemies. 26. Resistance to chemicals.27. Socioeconomic environment. 28. PCR. 29. RAPD. 30. Mixed cropping. 31. Highlands.32. Capacity building. 33. Africa. 34. Central America. 35. Mexico. 36. Caribbean. 37. LatinAmerica. Local descriptors in English:
1. Cassava. 2. Whitefly. 3. Molecular techniques.
AGROVOC descriptors in Spanish:1. Manihot esculenta. 2. Ipomoea batatas. 3. Batatas. 4. Hortalizas. 5. Leguminosas.
6. Insectos dañinos. 7. Aleyrodidae. 8. Bemisia tabaci. 9. Trialeurodes vaporariorum.10. Aleurotrachelus socialis. 11. Trialeurodes variabilis. 12. Bemisia tuberculata. 13. Trialeurodesabutiloneus. 14. Aleurocanthus woglumi. 15. Virus del mosaico de la cassava. 16. Geminivirus.17. Vectores. 18. Seguridad alimentaria. 19. Pobreza. 20. Control de plagas. 21. Control deenfermedades. 22. Control biológico. 23. Control integrado. 24. Parasitoides. 25. Enemigosnaturales. 26. Resistencia a productos químicos. 27. Entorno socioeconómico. 28. PCR.29. RAPD. 30. Cultivo mixto. 31. Zona de montaña. 32. Creación de capacidad. 33. África.34. América Central. 35. México. 36. Caribe. 37. América Latina.
Local descriptors in Spanish:1. Yuca. 2. Mosca blanca. 3. Técnicas moleculares.
I. Anderson, Pamela K. II. Morales, Francisco José. III. Centro Internacional de AgriculturaTropical. IV. Ser.
AGRIS subject categories: H10 Pests of plants / Plagas de plantasH20 Plant diseases / Enfermedades de las plantas
LC classification: SB 945 .W4 W4
Copyright CIAT 2005. All rights reserved
CIAT encourages wide dissemination of its printed and electronic publications for maximum public benefit.Thus, in most cases colleagues working in research and development should feel free to use CIAT materialsfor noncommercial purposes. However, the Center prohibits modification of these materials, and weexpect to receive due credit. Though CIAT prepares its publications with considerable care, the Centerdoes not guarantee their accuracy and completeness.

Page
Foreword vii
IntroductionPamela K. Anderson 1
SECTION ONE
Whiteflies as Vectors of Plant Viruses in Cassava andSweetpotato in Africa
Chapter
1.1 IntroductionJames Legg 15
1.2 GhanaAnthony Cudjoe, Joseph Gyamenah and Braima James 24
1.3 BeninBrice Gbaguidi, Braima James and Symphorien Saizonou 30
1.4 NigeriaNnamdi C. Echendu, Joseph B. Ojo, Braima James andBrice Gbaguidi 35
1.5 CameroonNelson Ntonifor, Braima James, Brice Gbaguidi andAmbe Tumanteh 40
1.6 UgandaPeter Sseruwagi, James Legg and William Otim-Nape 46
1.7 KenyaJoseph Kamau, Peter Sseruwagi and Valente Aritua 54
1.8 TanzaniaJoseph Ndunguru, Simon Jeremiah, Regina Kapinga andPeter Sseruwagi 61
Contents
iii

4
Whiteflies and Whitefly-borne Viruses in the Tropics
PageChapter
1.9 MalawiMathews P. K. J. Theu and Peter Sseruwagi 68
1.10 MadagascarSahondramalala Ranomenjanahary, Jocelyn Ramelison andPeter Sseruwagi 72
1.11 The Diversity of Cassava Mosaic Begomoviruses in AfricaPeter Markham, Robert Briddon, Clotilde Roussot,John Farquhar, Geoffrey Okao-Okuja and James Legg 77
1.12 Serological Analysis of Sweetpotatoes Affected by SweetpotatoVirus Disease in East Africa
Valente Aritua, Richard Gibson and Josef Vetten 83
1.13 Factors Associated with Damage to Sweetpotato Crops bySweetpotato Virus Disease
Richard Gibson, Valente Aritua and Simon Jeremiah 89
1.14 Conclusions and RecommendationsJames Legg and Braima James 98
SECTION TWO
Whiteflies as Pests and Vectors of Viruses in Vegetable andLegume Mixed Cropping Systems in Eastern and
Southern Africa
2.1 IntroductionMohamed Ali Bob, Rebecca Raini and Bernhard Löhr 115
2.2 SudanGassim Dafalla and Musa Ahmed 118
2.3 KenyaMohamed Ali Bob, Benjamin Odhiambo, Gilbert Kibata andJason Ong’aro 129
2.4 TanzaniaIgnas Swai and Simon Slumpa 141
2.5 MalawiHarriet Thindwa and Patric Khonje 150
2.6 Tomato Yellow Leaf Curl Virus and Tomato Leaf Curl-likeViruses in Eastern and Southern Africa
Remi Nono-Womdim, Ignas Swai, Madan Mohan Lal Chadha andSylvia Green 157
2.7 Conclusions and RecommendationsLisbeth Riis, Thomas Njuguna, Rebecca Raini, Berhard Löhrand Mohamed Ali Bob 163
iv

5
Page
SECTION THREE
Whiteflies as Vectors of Viruses in Legume and VegetableMixed Cropping Systems in the Tropical Lowlands of
Central America, Mexico and the Caribbean
Chapter
3.1 IntroductionFrancisco Morales 173
3.2 MexicoFrancisco Morales, Rafael Rivera-Bustamante,Rafael Salinas Pérez, Irineo Torres-Pacheco, Raúl Díaz Plaza,Wilson Avilés Baeza and Genovevo Ramírez Jaramillo 177
3.3 GuatemalaFrancisco Morales, Abelardo Viana, Margarita Palmieri,Mónica Orozco and René Ruano 188
3.4 El SalvadorFrancisco Morales, Carlos Pérez, Priscila Henríquez,Reina de Serrano, Leopoldo Serrano and Claudio Nunes 197
3.5 HondurasFrancisco Morales, María Mercedes Doyle, Rogerio Trabanino,Carolina Nolasco, Elsa Barrientos, Luís Jara and Norman Escoto 204
3.6 NicaraguaFrancisco Morales, Alberto Sediles, Aurelio Llano andFalguni Guharay 210
3.7 Costa RicaFrancisco Morales, Luko Hilje, Juan Vallejos, Guillermo Sibaja,Carlos Araya and Rodolfo Araya 217
3.8 PanamaFrancisco Morales, Orencio Fernández and José Guerra 222
3.9 HaitiFrancisco Morales, Jackson Donis and Emmanuel Prophete 226
3.10 CubaFrancisco Morales, Gloria González, Carlos Murguido,Ana Echemendía, Yolanda Martínez, Yenín Hernández,Benito Faure and María Chailloux 230
3.11 Dominican RepublicFrancisco Morales, Emigdio Gómez, Augusto Villar andJulio Nin 237
3.12 Reproductive Crop Hosts of Bemisia tabaci (Gennadius)in Latin America and the Caribbean
Pamela K. Anderson, Avas Hamon, Pilar Hernández andJon Martin 243
Contents
v

6
Whiteflies and Whitefly-borne Viruses in the Tropics
Page
Chapter
3.13 Using Molecular Techniques to Analyse Whitefly Species andBiotypes in Latin America
Lee Calvert, Natalia Villarreal and Donald Frohlich 251
3.14 Conclusions and RecommendationsFrancisco Morales 263
SECTION FOUR
Whiteflies as Pests of Annual Crops in the TropicalHighlands of Latin America
4.1 IntroductionCésar Cardona 269
4.2 Colombia and EcuadorCésar Cardona, Aristóbulo López-Avila and Oswaldo Valarezo 274
4.3 Insecticide Resistance in Colombia and EcuadorCésar Cardona, Francisco Rendón, Isaura Rodríguez andAristóbulo López-Avila 285
4.4 Conclusions and RecommendationsCésar Cardona, Aristóbulo López-Avila and Oswaldo Valarezo 295
SECTION FIVE
Special Topics on Pest and Disease Management
5.1 Sustainable Integrated Management of Whiteflies throughHost Plant Resistance
Anthony Bellotti, Joseph Tohme, Michael Dunbier andGail Timmerman 303
5.2 Biological Control of Whiteflies by Indigenous Natural Enemiesfor Major Food Crops in the Neotropics
Anthony Bellotti, Jorge Peña, Bernardo Arias,José María Guerrero, Harold Trujillo, Claudia Holguín andAdriana Ortega 313
5.3 Progress of Cassava Mosaic Disease in Ugandan CassavaVarieties and in Varietal Mixtures
William Sserubombwe, Michael Thresh, James Legg andWilliam Otim-Nape 324
5.4 Management of the Cassava Mosaic Disease Pandemic inEast Africa
James Legg, James Whyte, Regina Kapinga and James Teri 332
ConclusionsFrancisco Morales 339
Acronyms and Abbreviations 345
vi

CIAT Publication No. 341
Integrated Pest and Disease Management Projectand
Communications Unit
Editorial assistance: Gladys Rodríguez
Production: Graphic Arts Unit, CIATOscar Idárraga (layout)Julio C. Martínez (cover design)
Printing: Arte Libro Impresores, Cali, Colombia

Page
Foreword vii
IntroductionPamela K. Anderson 1
SECTION ONE
Whiteflies as Vectors of Plant Viruses in Cassava andSweetpotato in Africa
Chapter
1.1 IntroductionJames Legg 15
1.2 GhanaAnthony Cudjoe, Joseph Gyamenah and Braima James 24
1.3 BeninBrice Gbaguidi, Braima James and Symphorien Saizonou 30
1.4 NigeriaNnamdi C. Echendu, Joseph B. Ojo, Braima James andBrice Gbaguidi 35
1.5 CameroonNelson Ntonifor, Braima James, Brice Gbaguidi andAmbe Tumanteh 40
1.6 UgandaPeter Sseruwagi, James Legg and William Otim-Nape 46
1.7 KenyaJoseph Kamau, Peter Sseruwagi and Valente Aritua 54
1.8 TanzaniaJoseph Ndunguru, Simon Jeremiah, Regina Kapinga andPeter Sseruwagi 61
Contents
iii

4
Whiteflies and Whitefly-borne Viruses in the Tropics
PageChapter
1.9 MalawiMathews P. K. J. Theu and Peter Sseruwagi 68
1.10 MadagascarSahondramalala Ranomenjanahary, Jocelyn Ramelison andPeter Sseruwagi 72
1.11 The Diversity of Cassava Mosaic Begomoviruses in AfricaPeter Markham, Robert Briddon, Clotilde Roussot,John Farquhar, Geoffrey Okao-Okuja and James Legg 77
1.12 Serological Analysis of Sweetpotatoes Affected by SweetpotatoVirus Disease in East Africa
Valente Aritua, Richard Gibson and Josef Vetten 83
1.13 Factors Associated with Damage to Sweetpotato Crops bySweetpotato Virus Disease
Richard Gibson, Valente Aritua and Simon Jeremiah 89
1.14 Conclusions and RecommendationsJames Legg and Braima James 98
SECTION TWO
Whiteflies as Pests and Vectors of Viruses in Vegetable andLegume Mixed Cropping Systems in Eastern and
Southern Africa
2.1 IntroductionMohamed Ali Bob, Rebecca Raini and Bernhard Löhr 115
2.2 SudanGassim Dafalla and Musa Ahmed 118
2.3 KenyaMohamed Ali Bob, Benjamin Odhiambo, Gilbert Kibata andJason Ong’aro 129
2.4 TanzaniaIgnas Swai and Simon Slumpa 141
2.5 MalawiHarriet Thindwa and Patric Khonje 150
2.6 Tomato Yellow Leaf Curl Virus and Tomato Leaf Curl-likeViruses in Eastern and Southern Africa
Remi Nono-Womdim, Ignas Swai, Madan Mohan Lal Chadha andSylvia Green 157
2.7 Conclusions and RecommendationsLisbeth Riis, Thomas Njuguna, Rebecca Raini, Berhard Löhrand Mohamed Ali Bob 163
iv

5
Page
SECTION THREE
Whiteflies as Vectors of Viruses in Legume and VegetableMixed Cropping Systems in the Tropical Lowlands of
Central America, Mexico and the Caribbean
Chapter
3.1 IntroductionFrancisco Morales 173
3.2 MexicoFrancisco Morales, Rafael Rivera-Bustamante,Rafael Salinas Pérez, Irineo Torres-Pacheco, Raúl Díaz Plaza,Wilson Avilés Baeza and Genovevo Ramírez Jaramillo 177
3.3 GuatemalaFrancisco Morales, Abelardo Viana, Margarita Palmieri,Mónica Orozco and René Ruano 188
3.4 El SalvadorFrancisco Morales, Carlos Pérez, Priscila Henríquez,Reina de Serrano, Leopoldo Serrano and Claudio Nunes 197
3.5 HondurasFrancisco Morales, María Mercedes Doyle, Rogerio Trabanino,Carolina Nolasco, Elsa Barrientos, Luís Jara and Norman Escoto 204
3.6 NicaraguaFrancisco Morales, Alberto Sediles, Aurelio Llano andFalguni Guharay 210
3.7 Costa RicaFrancisco Morales, Luko Hilje, Juan Vallejos, Guillermo Sibaja,Carlos Araya and Rodolfo Araya 217
3.8 PanamaFrancisco Morales, Orencio Fernández and José Guerra 222
3.9 HaitiFrancisco Morales, Jackson Donis and Emmanuel Prophete 226
3.10 CubaFrancisco Morales, Gloria González, Carlos Murguido,Ana Echemendía, Yolanda Martínez, Yenín Hernández,Benito Faure and María Chailloux 230
3.11 Dominican RepublicFrancisco Morales, Emigdio Gómez, Augusto Villar andJulio Nin 237
3.12 Reproductive Crop Hosts of Bemisia tabaci (Gennadius)in Latin America and the Caribbean
Pamela K. Anderson, Avas Hamon, Pilar Hernández andJon Martin 243
Contents
v

6
Whiteflies and Whitefly-borne Viruses in the Tropics
Page
Chapter
3.13 Using Molecular Techniques to Analyse Whitefly Species andBiotypes in Latin America
Lee Calvert, Natalia Villarreal and Donald Frohlich 251
3.14 Conclusions and RecommendationsFrancisco Morales 263
SECTION FOUR
Whiteflies as Pests of Annual Crops in the TropicalHighlands of Latin America
4.1 IntroductionCésar Cardona 269
4.2 Colombia and EcuadorCésar Cardona, Aristóbulo López-Avila and Oswaldo Valarezo 274
4.3 Insecticide Resistance in Colombia and EcuadorCésar Cardona, Francisco Rendón, Isaura Rodríguez andAristóbulo López-Avila 285
4.4 Conclusions and RecommendationsCésar Cardona, Aristóbulo López-Avila and Oswaldo Valarezo 295
SECTION FIVE
Special Topics on Pest and Disease Management
5.1 Sustainable Integrated Management of Whiteflies throughHost Plant Resistance
Anthony Bellotti, Joseph Tohme, Michael Dunbier andGail Timmerman 303
5.2 Biological Control of Whiteflies by Indigenous Natural Enemiesfor Major Food Crops in the Neotropics
Anthony Bellotti, Jorge Peña, Bernardo Arias,José María Guerrero, Harold Trujillo, Claudia Holguín andAdriana Ortega 313
5.3 Progress of Cassava Mosaic Disease in Ugandan CassavaVarieties and in Varietal Mixtures
William Sserubombwe, Michael Thresh, James Legg andWilliam Otim-Nape 324
5.4 Management of the Cassava Mosaic Disease Pandemic inEast Africa
James Legg, James Whyte, Regina Kapinga and James Teri 332
ConclusionsFrancisco Morales 339
Acronyms and Abbreviations 345
vi

ForewordPamela Anderson*
* Former coordinator for Phase 1 of theTropical Whitefly IPM Project; currentlyResearch Director at the CentroInternacional de la Papa (CIP), Lima, Peru.
This book is the product of thecollective knowledge accumulated over3 decades by hundreds of agriculturalprofessionals and scientists involved inthe fight against two of the worst pestsof all times: whiteflies and whitefly-transmitted viruses. Millions of farmersaffected by these pests have alsocontributed a great deal of valuableinformation regarding the nature of theproblem and the extent of the damagecaused by these pests in importantfood and industrial crops. Examples ofsuch are: cassava (Manihot esculentaCrantz), common bean (Phaseolusvulgaris L.), sweetpotato (Ipomoeabatatas [L.] Lam.), lima bean(Phaseolus lunatus L. var. lunatus),mung bean (Vigna radiata [L.]R. Wilczek), tomato (Lycopersiconesculentum Mill.), peppers (Capsicumspp. L.), squash (Cucurbita moschataDuchesne), melon (Cucumis melo L.),other cucurbits, cotton (Gossypiumhirsutum L.), tobacco (Nicotianatabacum L.), soybean (Glycine max [L.]Merr.), lettuce (Lactuca sativa L. var.capitata L.) and eggplant (Solanummelongena L.).
The devastating capacity of thesepests, and their global distribution in
the main tropical and sub-tropicalregions of the world, has been a majorchallenge to agricultural scientists inboth developing and industrializednations. Obviously, their impact hasbeen far greater in the rural areas ofdeveloping countries, where resource-poor farmers have lost both traditionalfood crops (e.g., common bean andcassava) and cash crops (e.g., tomatoand peppers) to these pests. Most of thenational agricultural research institutesand ministries of agriculture in thedeveloping countries affected by thesepests have implemented differentwhitefly control measures, includinglegal measures, which constitute aunique event in the history of mostdeveloping countries where agriculturein general is not regulated by the state.
The research activities described inthis book provide a general view of theproblem for agricultural scientists andlaypeople alike, and contributes a greatdeal of pertinent information on thenature and management of the problemin the different crops affected bywhiteflies and whitefly-transmittedviruses. In doing so, the TropicalWhitefly Integrated Pest ManagementProject (TWF-IPM) has paid considerableattention to agricultural sustainability,ecosystem health and human healthissues, related to the appalling misuseof highly toxic pesticides associated withwhitefly control practices in developingcountries.
vii

8
Whiteflies and Whitefly-borne Viruses in the Tropics
The information contained in thefollowing chapters will form the basis ofa worldwide effort aimed atdisseminating the knowledgeaccumulated on the nature of thewhitefly/virus problems and the mostefficient integrated pest and diseasemanagement measures available todevelop sustainable mixed cropping
systems. Moreover, this knowledgeshould ultimately contribute to improvethe livelihoods of resource-poor farmersaffected by these pests throughout thetropics, by safeguarding their foodstaples and high-value crops, reducingproduction costs and pesticidecontamination, and increasing theirfamily income.
viii

1
Introduction
IntroductionPamela K. Anderson*
The Problem of Whitefliesand Whitefly-Transmitted
Viruses in the Tropics
Although Mound and Halsey (1978)catalogued 1156 species of whiteflies(Homoptera: Aleyrodidae), only alimited number of whitefly species areconsidered pests of economicimportance. Two whitefly species arekey pests throughout the tropics:Bemisia tabaci (Gennadius) andTrialeurodes vaporariorum (Westwood).Five additional whitefly species areconsidered as important pests inspecific regions: Aleurotrachelus socialis(Bondar), Trialeurodes variabilis(Quaintance), Bemisia tuberculata(Bondar), Trialeurodes abutiloneus(Haldman) and Aleurocanthus woglumi(Ashby).
Whiteflies are phloem (sap) feeders.They cause direct damage in somehosts by extracting large quantities ofsap. The honeydew that they excrete,as a result of the copious sap intake,serves as substrate for sooty mouldfungi, which can also damage hosts byblocking photosynthesis. Sooty mouldcan discolour harvestable fruits andfibre, affecting the quality of produce,and can cause plant death in crops
such as potato (Solanum tuberosum L.),tomato (Lycopersicon esculentum Mill.)and common bean (Phaseolus vulgarisL.). In addition, B. tabaci is a vector ofplant begomoviruses (Geminiviridae:Begomovirus). These whitefly-transmitted viruses (WTVs) are amongthe most destructive plant viruses;early virus infection often results intotal crop loss.
Although the problems caused byB. tabaci, both as pest and vector, havebeen recognized for more than100 years, serious damage had beenlimited to a handful of crops inparticular geographic areas. Thisscenario has changed over the past2 decades. The known WTVs haveextended their geographic range andother WTVs are emerging in new cropsand geographic zones, globally.Whitefly infestations now have becomesevere in both traditional and non-traditional food and industrial cropsthroughout the tropics.
In Africa, cassava mosaic disease(CMD) was first described in 1894 fromTanzania (Warburg, 1894). In EastAfrica, the disease was not reported tocause serious losses until the 1920s. InWest Africa, it was first recorded in thecoastal areas of Nigeria, Sierra Leoneand Ghana in 1929 and had spreadnorthward by 1945. By 1987, CMD hadbeen reported from all countries inAfrica producing cassava (Manihot
* Former coordinator for Phase 1 of theTropical Whitefly IPM Project; currentlyResearch Director at the CentroInternacional de la Papa (CIP), Lima, Peru.

2
Whiteflies and Whitefly-borne Viruses in the Tropics
esculenta Crantz) (Fauquet andFargette, 1990). The disease had beencaused by African cassava mosaic virus(ACMV) in South Africa and Africa westof the Rift Valley, and by East Africancassava mosaic virus (EACMV) inMadagascar and Africa east of the RiftValley (Hong et al., 1993; Swanson andHarrison, 1994). ACMV and EACMV areboth B. tabaci-transmittedbegomoviruses.
In 1988, an extremely severe formof CMD was reported from north-eastern Uganda. Infected cassavaplants showed severe disease symptomsand produced little or no yield. Theepidemic caused total crop failure innorth-eastern Uganda (Otim-Nape etal., 1997). Each year since 1988, theepidemic moved progressivelysouthwards along a broad front at arate of approximately 20 km per year(Otim-Nape et al., 2000). By 1995, theepidemic had reached Kenya (Otim-Nape et al., 2000), with lossesestimated at 140,000 tons of cassavaper year, conservatively worthUS$14 million (Legg, 1999). The severeoutbreak in Uganda and neighbouringcountries has been associated with anovel recombinant (EACMV+ACMV)begomovirus, currently recognized asEACMV-Ug (Pita et al., 2001).
Soon after, Latin America wasstruggling with Bemisia-transmittedbean and tomato viruses. Bean goldenmosaic virus (BGMV) was first describedin Brazil in the early 1960s as a minordisease (5%-10% incidence) of commonbean in the state of São Paulo (Costa,1965). Since then, BGMV has become awidespread problem in Brazil and nowis known to cause epidemics in north-western Argentina and eastern Bolivia.Simultaneously, another begomovirus,Bean golden yellow mosaic virus(BGYMV), which causes similarsymptoms, was ravaging common beanproduction in over 12 countries of
Central America and the Caribbean(Morales, 1994). Yield loss from eitherBGMV or BGYMV is often 100%because of the high incidence of flowerabortion and the malformation of podsin infected plants (Morales andNiessen, 1988). BGMV and BGYMV areconsidered to be the limiting bioticconstraint to bean production in LatinAmerica (Gálvez and Morales, 1989).
Then, in the late 1980s, tomato-producing areas in Latin America andacross the tropics began to suffer fromhigh incidences of whitefly-bornebegomoviruses with devastatingeconomic consequences. While noformal assessment studies of crop losshad been undertaken for the tomatodiseases caused by begomoviruses, theempirical data were impressive. One ofthe more complete data sets came fromthe Dominican Republic (Polston andAnderson, 1997). In 1988, multiplebegomoviruses began to affect tomatoproduction in the Dominican Republic.Crop damage from 1988 to 1995ranged from 5% to 95%. Economiclosses in 1988 were estimated atUS$10 million dollars, with losses from1989 to 1995 totalling an estimatedUS$50 million dollars.
These scenarios were beingrepeated across the tropics, withsignificant consequences to foodsecurity and poverty alleviation effortsas well as human and ecosystemhealth.
Food security
In Africa, cassava is one of the mostwidely grown staple crops and secondonly to maize (Zea mays L.) in terms ofcalorie intake in sub-Saharan Africa.About 200 million people, or 40% ofthe sub-Saharan Africa population, relyon cassava for their well-being. In somecountries, people derive approximately1000 calories a day, or 50% of dailyfood intake, from cassava (IITA, 1988).

3
Introduction
In addition to the yield lossescaused by CMD, an estimated150,000 ha of cassava-growing landwas abandoned in Uganda during theheight of the epidemic; that is,equivalent to over 2.2 million metrictons (US$440 million). This causedfood shortages and famine in a numberof districts, particularly in the easternand northern regions where the cropwas the principle staple food (Otim-Nape, 1997).
In Latin America, common bean isone of the main staple foods,particularly among the rural and urbanpoor. In Central America, beans are themost important source of protein,usually being consumed three times aday. Central America, despite its smallarea (498,368 km2), devotes twice asmuch of its geographical area to thecultivation of beans (735,00 ha), whencompared to major bean producerssuch as Brazil (>5,000,000 ha). Beansalso are produced in some Caribbeanislands, such as Cuba (26,000 t), theDominican Republic (55,000 t) andHaiti (56,000 t) where they play animportant nutritional role in the diet ofthe lower socio-economic classes.Smallholdings cultivated by farmerswith limited resources arecharacteristic of bean production inCentral America and the Caribbean. InEl Salvador, for instance, 85% of thebean producers cultivate less than14 ha, and 50% of these producershave holdings of less than 3.5 ha.
Despite the large area planted tobeans in Central America, averageproductivity is low (495 kg/ha)compared to the average yield expected(over 1500 kg/ha) in most bean-producing regions of the USA and othertemperate countries in the world. Themain factor identified as responsible forthe low bean productivity in LatinAmerica has been the incidence ofbiotic constraints, particularly
diseases. Morales (1992) estimated thatby the early 1990s, approximately2,500,000 ha were under attack byBGMV/BGYMV and that at least1 million additional hectares could notbe planted to beans each year becauseof the possibility of total yield losses,mainly during the dry seasons, whenwhitefly populations peak. ThroughoutCentral America and the CaribbeanBasin, figures for crop damageindicated that the BGYMV infection wasdevastating. And there was continuingconsensus that BGYMV was the majorbiotic factor limiting bean production inLatin America (Morales, 1994).
Poverty alleviation
Many resource-poor producers in thetropics modified their traditionalcropping systems in the 1980s, toincorporate non-traditional cash andexport crops. The shift fromsubsistence to commercial agricultureis an opportunity to alleviate poverty inrural areas. Smallholder producersacross the tropics have begun tocultivate cash crops such as tomato(Lycopersicon esculentum), pepper(Capsicum spp. L.), melon (Cucumismelo L.), watermelon (Citrullus lanatus[Thunb.] Matsum. & Nakai), squash(Cucurbita moschata Duchesne), grape(Vitis vinifera L.), okra (Abelmoschusesculentus [L.] Moench), cucumber(Cucumis sativus L. var. sativus),cabbage (Brassica oleracea L.), eggplant(Solanum melongena L.), broccoli(Brassica oleracea L. var. italica Plenck)and ornamentals. However, most ofthese horticultural crops areparticularly susceptible to pestspreviously unknown to most small-scale farmers.
In the absence of adequatetechnical assistance for small-scalefarmers in the tropics, the new pestproblems were primarily dealt with by amyriad of pesticides appliedindiscriminately. To further complicate

4
Whiteflies and Whitefly-borne Viruses in the Tropics
this situation, a new biotype (B) ofB. tabaci was introduced in theAmericas in the early 1990s. Thisbiotype proved to be far morepolyphagous and fecund that theoriginal A biotype and has been animportant factor in most of the majoroutbreaks of whiteflies and whitefly-borne viruses. During the past decade,the outbreak of whitefly pests and theepidemics caused by whitefly-transmitted begomoviruses in cashcrops has been often devastating.Pesticide abuse is the norm in allwhitefly-stricken regions of the tropics,which increases production costs anddisqualifies contaminated (pesticideresidues) produce for export.
At present, many of the small-scalefarmers that attempted to diversifytheir cropping systems have failed todo so because of whitefly-associatedproblems. Prime agricultural landremains unexploited in manydeveloping countries during the dryseasons, when whitefly populationspeak, despite the availability of water(irrigation districts) in these areas. Thewelfare of developing countries andtheir low-income citizens is tightlylinked to the existence of cash-earningcommodities, principally agriculturalproducts.
The main force driving whiteflyoutbreaks and crop losses due towhitefly attack and WTVs is the lack ofqualified technical assistance to small-and medium-scale farmers in thetropics. The Tropical WhiteflyIntegrated Pest Management (TWF-IPM)Project is currently entering a finalphase when most of the informationpresented in this book and the lessonslearned during the validation of IPMpractices in Phase II will be translatedinto practical recommendations forresource-poor farmers throughout thetropics.
Human and ecosystem health
The reliance of farmers onagrochemicals to protect their cashcrops against whitefly pests andwhitefly-borne viruses has resulted inthe systematic destruction of naturalenemies that were once effective inproviding natural control, whiteflypopulations with high levels ofinsecticide resistance and the creationof new secondary pest problems. Therejection of contaminated produce ininternational markets has led to thesale of highly contaminated foodproducts in developing countries, withobvious detrimental healthconsequences for rural and urbanconsumers alike. Whiteflies provide aclassic example of the pesticidetreadmill. Insecticide abuse hasbecome a serious threat to theenvironment as well as a health hazardto producers and consumers.
About 10% of the world’spopulation lives in the main highlandsand mountainous areas of thedeveloping world (the Andes of SouthAmerica, the Africa ranges and theHimalayas). The management ofresources in those ecosystems affectsan additional 40% of the world’spopulation, which inhabits adjacentareas such as Inter-Andean valleys inthe Andean Zone. Traditionally, peoplein the highlands have beenmarginalized from major developmentefforts, with significant repercussionson poverty, migration and social unrestas well as environmental deteriorationin situ and downstream (IFPRI, 1995).
One major issue regarding thewelfare of those living in highland areasof Latin America is environmentaldegradation caused by excessivepesticide use. It is well known thatpesticide consumption in thedeveloping world is increasing rapidly.It was thought initially that increases

5
Introduction
in pesticide use in Latin America weremainly due to the growth of plantationcrops, an important source of exportrevenue (Bellotti et al., 1990). However,as pointed out by Whitaker (1993), thedeveloping country share of the worldagrochemical usage, currently valuedat US$10.6 billion, was forecast to risefrom 19% in 1988 to an estimated 35%by the year 2000. Much of thisprojected growth was attributed to awider and more intensive use ofchemical protection by smallholderfarmers. Unfortunately, highly toxicinsecticides represent up to 45% ofpesticide use in the developing world.
In the early 1980s, the greenhousewhitefly, Trialeurodes vaporariorum(Westwood), became a very serious pestof several hillside-grown crops in theAndes. Major outbreaks occurred in1987, 1991 and 1994 in selected areasof Colombia, northern Ecuador and theConstanza Valley in DominicanRepublic. For example, insecticide useby small-scale bean farmers, whichwas negligible until 1975-77(Schoonhoven and Cardona, 1980), hasincreased steadily and becameexcessive in Colombia and Ecuador.Surveys conducted in two regions ofColombia and the northern part ofEcuador (CIAT, 1994) revealed that100% of 893 farmers surveyed sprayedtheir crops in an attempt to control thegreenhouse whitefly. Highestinsecticide use occurred in theSumapaz region of Colombia wherefarmers make an average of11.1 applications per season.
Most alarming was the fact thatsome growers sprayed their crops up to24 times in a crop cycle of90-100 days, that is, every 3 to 4 days.Insecticide abuse on beans in Colombia(5.6 kg active ingredient per ha perseason) could be compared toinsecticide consumption for cotton(Gossypium hirsutum L.) (6.2 kg active
ingredient per hectare per season), acrop that has been known traditionallyas the worst offender in terms ofinsecticide use. And, of the insecticidesutilized for whiteflies, 78% wereclassified in toxicological category I(highly toxic) and most often applied inmixture with other insecticides, broad-spectrum fungicides and foliarfertilizers. Farmers usually did not takeprecautions when handling pesticides;up to 24% of those surveyed admit thatthey have been intoxicated at leastonce in the prior 10 years (CIAT, 1994).
In the east African highlands, acontributing factor to environmentaldegradation was the indiscriminateand/or excessive use of broad-spectrum insecticides as part of theintensive crop protection system inindustrial crops, especially cotton, andto some extent on plantation cropssuch as coffee (Coffea spp. L.) and tea(Camellia sinensis [L.] Kuntze). Theproblems of pesticide resistance andpest resurgence although initiallycreated on these target crops hadextended to the other seasonal cropsgrown in rotation and/or combinationwith the plantation crops, as in thecase of whiteflies.
The increasing importance ofhorticultural production also broughtwith it the attendant strategy ofpesticide-based protection in an effortto harvest damage-free produce.Intensive use of pesticides in vegetablecrops had become quite common inlarge areas of Sudan, Uganda, Kenya,Tanzania and Zimbabwe. In a survey ofvegetable farmers in Kenya it wasfound that the majority perceived thatthey would lose up to 90% of theirharvest if they did not use pesticides.By the early 1990s, these vegetablefarmers were applying up to 19 spraysa season, with a significant proportionof farmers spraying 9-12 times perseason (KARI-GTZ, 1994). It was

6
Whiteflies and Whitefly-borne Viruses in the Tropics
critical to begin work in eastern Africain hopes of intervening with alternativewhitefly crop protection measures,before the whitefly pests and WTVsreach the severity witnessed in theNeotropics.
There was an urgent need todevelop IPM systems that would reducepesticide use and help re-establish theecological equilibrium by means ofnon-chemical approaches to whiteflymanagement. For whitefly pests thisimplies identifying the principal crophosts, establishing economic injurylevels (EILs) and developing newapproaches to maintain whitefliesbelow the EIL. For whitefly vectors,however, the traditional IPM approachwill not suffice. Vectors must bestudied and managed within anepidemiological framework, that is,study and analysis of the WTV systemwith IPM intervention strategiesresulting from the epidemiologicalanalysis.
Origin and Organizationof the CGIAR TWF-IPM
Project
The Systemwide Programme onIntegrated Pest Management(SP-IPM)
The Consultative Group forInternational Agricultural Research(CGIAR) consists of 15 InternationalAgricultural Research Centres (IARCs)located around the world. Historically,each Centre has worked in a relativelyindependent and autonomous fashion.During the 1990s, the CGIARSecretariat recognized that significantbenefits could be derived by poolinghuman capital, infrastructure andeconomic resources to jointly tackleresearch problems that numerouscentres were working onindependently. New inter-centre
initiatives included the creation of theSP-IPM. Whitefly and WTV research hadbeen ongoing at several of the IARCssince the 1970s. Due to the growingimportance of the whitefly and WTVproblem, the TWF-IPM Project was thefirst inter-centre project approvedwithin the new programme. TheInternational Center for TropicalAgriculture (CIAT, the Spanishacronym) was designated as theconvening centre to organize the Inter-Centre Whitefly IPM Task Force and toformulate the Inter-Centre proposal onSustainable Integrated Management ofWhiteflies as Pests and Vectors of PlantViruses in the Tropics (the CGIARTWF-IPM Project).
A Task Force Meeting was held atCIAT in Cali, Colombia, from13-15 February 1996. The objectives ofthe meeting were to discuss: (a) thegoal, purpose, outputs and activitiesthat should be proposed for the project;and (b) a structure for the globalWhitefly IPM project, as well as how tolink and coordinate the institutions thatwould be involved in the project.
The Task Force Meeting included24 participants representing IARCs,National Agricultural Research Systems(NARS) and Advanced ResearchInstitutions (ARIs). After considerablediscussion on the nature of the whiteflyproblem, the Task Force agreed that itwas possible to define three whiteflyproblems that should be prioritized:
(1) Whiteflies as vectors in mixedcropping systems in low to midaltitudes of the tropics;
(2) Whiteflies as pests in mixedcropping systems in tropicalhighlands; and
(3) Whiteflies as vectors and pests ofthe semi-perennial cassava.
The first problem focused onwhiteflies as vectors of plant viruses in

7
Introduction
annual crops, especially legumes andvegetables, in the tropical lowlands.The second problem focused onwhiteflies as direct pests in annualcrops in the highlands. And the thirdproblem focused on whitefly pests andvectors in a semi-perennial crop,cassava.
Based on the initial Whitefly IPMTask Force Meeting, and within the CGframework for an eco-regional problemapproach (Bouma et al., 1995), theproject was structured into six sub-projects:
(1) Bemisia tabaci as a virus vector incassava and sweet potato in sub-Saharan Africa – led by IITA;
(2) Bemisia tabaci as a virus vector inmixed cropping systems of theCaribbean, Mexico and CentralAmerica – led by CIAT;
(3) Bemisia tabaci as a virus vector inmixed cropping systems of Easternand Southern Africa – led by theInternational Centre of InsectPhysiology and Ecology (ICIPE);
(4) Bemisia tabaci as a virus vector inmixed cropping systems of S.E.Asia – led by the Asian VegetableResearch and Development Center(AVRDC);
(5) Trialeurodes vaporariorum as adirect pest in the tropicalhighlands of Latin America – led byCIAT; and
(6) Whiteflies as direct pests oncassava in South America – led byCIAT.
There was consensus by the TaskForce on the project goal, projectpurpose and project outputs for theTWF-IPM project. The project goal is toimprove living conditions of ruralfamilies through the effectivemanagement of whiteflies, resulting inincreased crop production and a saferenvironment. The project purpose is toreduce crop losses due to whitefly
feeding damage and WTVs. The TaskForce agreed to organize the projectaround six outputs:
(1) Formation of an internationalnetwork for research on whitefliesand WTVs in the tropics;
(2) Characterization of whiteflyproblems in order to prioritizecritical target areas;
(3) Improvement of the understandingof whitefly pest and diseasedynamics in critical target areas;
(4) Development and testing of IPMstrategies and tactics;
(5) Strengthening of NARS researchcapacity, policy formulation andIPM implementation; and
(6) Assessment of project impact.
Phase 1 of the CGIAR TWF-IPMProject
The project was conceptualized in threephases, to be carried out over 10 years.Phase 1 of the project would focusprimarily on Output 1 (networkformation) and Output 2 (problemcharacterization) as the basis forPhase 2 work, with limited activities inOutput 3 (pest and disease dynamics)and Output 4 (development and testingof IPM strategies and tactics). TheProject was officially launched in 1997and evolved into a pan-tropicalpartnership. Five IARCs were included:
CIAT Centro Internacional deAgricultura Tropical
IITA International Institute ofTropical Agriculture
AVRDC Asian Vegetable Research andDevelopment Center
ICIPE International Centre of InsectPhysiology and Ecology
CIP Centro Internacional de laPapa

8
Whiteflies and Whitefly-borne Viruses in the Tropics
Twelve ARIs were also included:
NRI Natural Resources Institute(UK)
JIC John Innes Centre (UK)
BBA Biologische Bundesanstalt fürLand-und Fortwirtschaft(Germany)
CATIE Centro Agronómico Tropical deInvestigación y Enseñanza(Costa Rica)
UA University of Arizona-Tucson(USA)
UFL University of Florida-Gainesville (USA)
UW University of Wisconsin-Madison (USA)
MSU Montana State University(USA)
FSAC Florida State ArthropodCollection
USDA- U.S. Department of(ARS) Agriculture-Agricultural
Research Service
CSIRO Commonwealth Scientific andIndustrial ResearchOrganisation (Australia)
C&F New Zealand Crop and FoodResearch
And 31 National AgriculturalResearch and Extension Systems(NARES) in Latin America, theCaribbean, Africa and Asia wereincluded from:
Mexico, Guatemala, Honduras,El Salvador, Nicaragua, Costa Rica,Panama, El Salvador, Cuba, Haiti,Dominican Republic, Colombia,Ecuador, Sudan, Benin, Ghana, Nigeria,Cameroon, Kenya, Uganda, Tanzania,Malawi, Madagascar, Nepal, Sri Lanka,
Bangladesh, Thailand, Malaysia,Vietnam, Indonesia and thePhilippines.
This collaborative partnership waspossible due to the support of sixdonor partners:
DANIDA Danish InternationalDevelopment Agency
ACIAR Australian Centre forInternational AgriculturalResearch
USAID United States Agency forInternational Development
MFAT Ministry of Foreign Affairsand Trade (New Zealand,now New Zealand Aid)
USDA- U.S. Department of(ARS) Agriculture-Agricultural
Research Service
DFID Department for InternationalDevelopment (UK)
The CGIAR TWF-IPM Project waslaunched in 1997 with the DANIDA-funded Phase 1 project on SustainableIntegrated Management of Whiteflies asPests and Vectors of Plant Viruses inthe tropics. This project coveredactivities in Output 1 (networkformation) and Output 2(characterization) activities in23 African and Latin American partnercountries in Sub-projects 1, 2, 3 and5. ACIAR then approved funding for theproject Sustainable IntegratedManagement of Whiteflies as Pests andVectors of Plant Viruses in S.E. Asia,which covered Output 2(characterization) activities in the eightAsian countries in Sub-project 4. TheUSAID-funded project on BiologicalControl of Whiteflies by IndigenousNatural Enemies for Major Food Cropsin the Neotropics and the MFAT-fundedproject on Sustainable IntegratedManagement of Whiteflies through Host

9
Introduction
Plant Resistance contributed to Output2 (characterization) and Output 4 (IPM)activities in Sub-project 6. USDA-ARSsigned a Scientific CooperativeAgreement to link the ARS nationalresearch on whitefly IPM research withthe international effort. This agreementon Integrated Pest ManagementStrategies for Whitefly-transmittedViruses supported Output 1 (networkformation) and preliminary Output 3(disease dynamics) research activitieson epidemiology of whitefly-transmittedbegomoviruses.
The CMD epidemic in easternAfrica was of particular concern to theTWF-IPM Project for humanitarianreasons. The USAID Office of ForeignDisaster Assistance (OFDA) grantedspecial funding for the EmergencyProgramme to Combat the CassavaMosaic Disease Pandemic in EastAfrica. The objective of this disasterassistance is to boost production ofcassava in Uganda, Kenya andTanzania and enhance both short- andlonger-term food security through theimplementation of an emergencyprogram to multiply and disseminateCMD-resistant cassava (Output 4activities). And DFID funded the projecton Promotion of and Technical Supportfor Methods of Controlling Whitefly-borne Viruses in Sweet Potato in EastAfrica; sweetpotato (Ipomoea batatas[L.] Lam.) served as the principal foodsecurity crop in the midst of the CMDepidemic.
Whiteflies and Whitefly-BorneViruses in the Tropics: Building aKnowledge Base for Global Actionreports the results from Phase 1 of theCGIAR TWF-IPM Project. The purposeof Output 2 (problem characterization)was to gather, review, generate andanalyse baseline data relevant to thediagnosis and characterization ofwhitefly and WTV problems in thetropics, in order to propose a sound
research agenda for improvedunderstanding of pest and diseasedynamics, IPM development and IPMimplementation. Several of the projectcoordinators have published reviewarticles (Polston and Anderson, 1997;Legg, 1999; Morales, 2000; Bellotti andArias, 2001; Morales, 2001; Moralesand Anderson, 2001; Morales andJones, 2004). Baseline data weregenerated through extensive surveywork in each of the participatingtropical countries. Surveys includedthe collection and identification ofbiological specimens (whitefly species,whitefly biotypes, geminiviruses, andB. tabaci natural enemies) andcollection and analysis of socio-economic data and IPM practices,based on producer interviews. Allpartners have presented preliminaryresults of the extensive survey workand IPM measures in international,regional and national meetings. Thisbook is the first presentation ofdetailed results generated by thePhase 1 projects contributing to theCGIAR TWF-IPM Project. For furtherinformation on specific methodologiesused to produce the results publishedin this book, please contact theinformation and communicationassistant of the project(www.tropicalwhiteflyipmproject.cgiar.org).
References
Bellotti, A. C.; Arias, B. 2001. Host plantresistance to whiteflies withemphasis on cassava as a casestudy. Crop Prot. 20:813-823.
Bellotti, A. C.; Cardona, C.; Lapointe, S.L. 1990. Trends in pesticide use inColombia and Brazil. J. Agric.Entomol. 7:191-201.
Bouma, J.; Kuyvenhoven, A.; Bouman, B.A. M.; Luyten, J. C. (eds.). 1995.Eco-regional approaches forsustainable land use and foodproduction. Kluwer AcademicPublishers, Dordrecht, NL.

10
Whiteflies and Whitefly-borne Viruses in the Tropics
CIAT (Centro Internacional de AgriculturaTropical). 1994. Annual report.CIAT-Bean Program, Cali, CO.p. 171-179.
Costa, A. S. 1965. Three whitefly-transmitted virus diseases of beansin São Paulo, Brazil. FAO PlantProt. Bull. 13:121-130.
Fauquet, C.; Fargette, D. 1990. Africancassava mosaic virus: Etiology,epidemiology and control. Plant Dis.74:404-411.
Gálvez, G.; Morales, F. J. 1989. Whitefly-transmitted viruses. In: Schwartz,H. F.; Pastor-Corrales, M. A. (eds.).Bean production problems in thetropics. Centro Internacional deAgricultura Tropical (CIAT), Cali,CO. p. 379-406.
Hong, Y. G.; Robinson, D. J.; Harrison, B.D. 1993. Nucleotide sequenceevidence for the occurrence of threedistinct whitefly-transmittedgeminiviruses in cassava. J. Gen.Virol. 74:2437-2443.
IFPRI (International Food Policy ResearchInstitute). 1995. A 2020 vision forfood, agriculture, and theenvironment. InternationalConference, 13-15 June 1995,Washington, DC. 145 p.
IITA (International Institute of TropicalAgriculture). 1988. Strategic plan1989-2000. Ibadan, NG.
KARI-GTZ (Kenya Agricultural ResearchInstitute - German Agency forTechnical Cooperation). 1994. Cropprotection measures of Kenyanvegetable farmers and their use ofpesticides, knowledge, attitude andpractice survey. GTZ-IntegratedPest Management HorticultureProject Report, Nairobi, KE. 45 p.
Legg, J. 1999. Emergence, spread andstrategies for controlling thepandemic of cassava mosaic virusdisease in east and central Africa.Crop Prot. 18:627-637.
Morales, F. J. 1992. Annual progressreport (1992) and five-year report(1988-1992). International Centerfor Tropical Agriculture (CIAT), Cali,CO.
Morales, F. J. (ed.). 1994. Bean goldenmosaic: Research advances. CentroInternacional de AgriculturaTropical (CIAT), Cali, CO. 193 p.
Morales, F. J. (ed.). 2000. Bean goldenmosaic virus and other diseases ofcommon bean caused by whitefly-transmitted geminiviruses in LatinAmerica. Centro Internacional deAgricultura Tropical (CIAT), Cali,CO. 169 p.
Morales, F. J. 2001. Conventionalbreeding for resistance to Bemisia-transmitted geminiviruses. CropProt. 20:825-834.
Morales, F. J.; Anderson, P. K. 2001. Theemergence and dissemination ofwhitefly-transmitted geminivirusesin Latin America. Arch. Virol.146(3):415-441.
Morales, F. J.; Jones, P. G. 2004. Theecology and epidemiology ofwhitefly-transmitted viruses inLatin America. Virus Res. 100:57-65.
Morales, F. J.; Niessen, A. I. 1988.Comparative responses of selectedPhaseolus vulgaris germplasminoculated artificially and naturallywith Bean golden mosaic virus. PlantDis. 72:1020-1023.
Mound, L. A.; Halsey, S. H. 1978.Whitefly of the world: A systematiccatalogue of the Aleyrodidae(Homoptera) with host plant andnatural enemy data. John Wiley,Chichester, GB. 340 p.
Otim-Nape, G. W.; Bua, A.; Thresh, J. M.;Baguma, Y.; Ogwal, S.; Semakula,G. N.; Acola, G.; Byabakama, B.;Martin, A. 1997. Cassava mosaicvirus disease in Uganda: Thecurrent pandemic and approachesto control. Natural ResourcesInstitute (NRI), Chatham, GB. 65 p.

11
Introduction
Otim-Nape, G. W.; Bua, A.; Thresh, J. M.;Baguma, Y.; Ogwal, S.; Ssemakula,G. N.; Acola, G.; Byabakama, B.;Colvin, J.; Cooter, R. J.; Martin A.2000. The current pandemic ofcassava mosaic virus disease inEast Africa and its control. CABInternational, Wallingford, GB.100 p.
Pita, J. S.; Fondong, V. N.; Sangaré, A.;Otim-Nape, G. W.; Ogwal, S.;Fauquet, C. M. 2001.Recombination,pseudorecombination andsynergism of geminiviruses aredeterminant keys to the epidemic ofsevere cassava mosaic disease inUganda. J. Gen. Virol. 82:655-665.
Polston, J. P.; Anderson, P. K. 1997. Theemergence of whitefly-transmittedgeminiviruses in tomato in theWestern Hemisphere. Plant Dis.81:1358-1369.
Schoonhoven, A. van; Cardona, C. 1980.Insects and other bean pests inLatin America. In: Bean productionproblems: Disease, insect, soil andclimatic constraints of Phaseolusvulgaris. Centro Internacional deAgricultura Tropical (CIAT), Cali,CO. p. 363-412.
Swanson, M. M.; Harrison, B. D. 1994.Properties, relationships anddistribution of cassava mosaicgeminiviruses. Trop. Sci. 34:15-25.
Warburg, O. 1894. Die Kulturpflazenusambaras. Mitt. Dtsch. Schutz.7:131.
Whitaker, M. J. 1993. The challenge ofpesticide education and training fortropical smallholders. Int. J. PestManage. 39(2):117-125.

blanca 12

SECTION ONE
Whiteflies as Vectors of Plant Virusesin Cassava and Sweetpotato in Africa
SECTION ONE
Whiteflies as Vectors of Plant Virusesin Cassava and Sweetpotato in Africa

BLANCA 14

15
Introduction
CHAPTER 1.1
IntroductionJames Legg*
More than 1200 species of whitefly(Homoptera: Aleyrodidae) have beendescribed worldwide, of which roughlyone quarter occurs in Africa (Moundand Halsey, 1978). Of these diversewhitefly species, only a few are ofsignificant economic importance. Theseinclude: Aleurocanthus spiniferus(Quaintance), Aleurocanthus woglumiAshby, Aleurodicus dispersus Russell,Aleurothrixus floccosus (Maskell),Aleyrodes proletella (Linnaeus), Bemisiatabaci (Gennadius), Dialeurodes citri(Ashmead), Parabemisia myricae(Kuwana), Siphoninus phillyreae(Haliday), Trialeurodes vaporariorum(Westwood) and Vasdavidius indicus(David and Subramaniam) Russell.Principal among these, however, isB. tabaci, which is a pest on a widerange of field crops throughout thecontinent.
B. tabaci was first described fromtobacco (Nicotiana tabacum L.) inGreece and is thought to haveoriginated from the Old World (Frohlichet al., 1996). It has been recognized asan important agricultural pest in Africasince at least the 1930s, with several of
the earliest reports relating to damagein cotton (Gossypium hirsutum L.)(Kirkpatrick, 1931) and cassava(Manihot esculenta Crantz) (Kufferathand Ghesquière, 1932; Storey andNichols, 1938). By the 1950s,B. tabaci was reported causing majorlosses in cotton as a result ofpopulation resurgence of this specieswhen it developed resistance topesticides applied against other pests(Joyce, 1955). And, by the 1980s,B. tabaci had been associated with amajor pandemic of cassava mosaicdisease (CMD) in East Africa (Otim-Nape et al., 1997; Legg, 1999).
Cassava and Sweetpotatoin Sub-Saharan Africa
According to international productionstatistics (FAO, 2004), Africaproduces almost half of the world’scrop of cassava and East Africa is oneof the world’s most important areasfor growing sweetpotato (Ipomoeabatatas [L.] Lam). Nigeria is theworld’s leading producer of cassava,while Uganda ranks third, worldwide,in total sweetpotato production.These two crops are of majorimportance to sub-Saharan Africaand play vital roles in sustaining foodsecurity, particularly in view of theirability to provide reasonable yieldseven under conditions of poor soilfertility or drought (Jennings, 1970).
* International Institute of TropicalAgriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda, and Natural Resources Institute,University of Greenwich, Chatham Maritime,Kent, UK.

16
Whiteflies and Whitefly-borne Viruses in the Tropics
Cassava was brought to Africa inthe sixteenth century by Portugueseseafarers and, perhaps because of thisrelatively recent introduction, has fewmajor pests and diseases. This is incontrast to the wide diversity of pestsand diseases that affect this crop in itsnative South America (Bellotti et al.,1994). The two most importantarthropod pests of cassava in Africa,the cassava mealybug (Phenacoccusmanihoti Matile-Ferrero) and thecassava green mite (Mononychellustanajoa [Bondar]) were both introducedfrom South America in the early 1970s(Yaninek and Herren, 1988;Neuenschwander, 1994). They spreadrapidly to all cassava-producing zonesof mainland Africa during the 1980sand 1990s but both have beencontrolled successfully through theimplementation of continent-widebiological control programs (Herren andNeuenschwander, 1991; Yaninek et al.,1993).
The principal diseases of cassavain Africa are CMD and cassavabacterial blight. The latter, like the twoarthropod pests described above, wasintroduced into Africa in the 1970s andspread rapidly to all cassava-growingareas (Boher and Verdier, 1994). Thedisease is caused by the bacteriumXanthomonas axonopodis pv. manihotis(Xam). Although, under certaincircumstances, damage may be verysevere and result in total crop loss,outbreaks with serious effects areusually localized. Cassava bacterialblight is therefore less economicallyimportant than CMD.
CMD was first described more thana century ago (Warburg, 1894) fromwhat is modern-day Tanzania. It wasassumed from the early years of thetwentieth century that the disease wascaused by a virus, since no pathogenswere visible on affected plants and thecondition was shown to be graft
transmissible (Zimmerman, 1906). Itwas not until the 1970s, however, thatthe viral etiology was confirmed (Bockand Woods, 1983). CMD is caused bybegomoviruses (Bock and Woods,1983; Hong et al., 1993), which aretransmitted by B. tabaci (Storey andNichols, 1938; Chant, 1958; Dubern,1979). Cassava mosaic begomovirusesoccurring in Africa are thought to beindigenous to that continent; it isassumed that they moved into cassavafrom wild host plants following theintroduction of cassava from SouthAmerica (Swanson and Harrison,1994).
Cassava was subsidiary inimportance to other staple crops untilthe early part of the twentieth centurybut promotion of the crop as a faminereserve by colonial authorities in the1920s and 1930s rapidly led to it beingmore widely grown and consumed(Hillocks, 2002). It appears that thisincrease in cultivation area andintensity, coupled possibly with theunrestricted movement of germplasmbetween countries, catalyzed thecontinent-wide spread of CMD. Newoccurrences as well as the increasingimportance of CMD were reported fromnumerous countries in East and WestAfrica, including Sierra Leone(Deighton, 1926), Uganda (Hall, 1928),Ghana (Dade, 1930), Nigeria (Golding,1936) and Madagascar (François,1937). Losses due to CMD in Africahave been estimated at between12 and 23 million tons annually,equivalent to between US$1200 andUS$2300 million (Thresh et al., 1997).Following the success of biologicalcontrol in reducing losses caused bycassava mealy bug and cassava greenmite, CMD generally is regarded as themost important biotic constraint oncassava production in Africa.
Sweetpotato in Africa is attackedby a greater diversity of pests and

17
Introduction
diseases than is cassava, although, aswith cassava, few are of majoreconomic importance. The mostimportant pests include thesweetpotato weevil (Cylas spp.), thestriped sweetpotato weevil (Alcidodesdentipes [Oliver]), the sweetpotatobutterfly (Acraea acerata Hewitson), leafblight (Alternaria spp.) and sweetpotatovirus disease (SPVD). The latter hasbeen ranked as the single mostimportant constraint to sweetpotatoproduction in the East African regionand is the most important diseasethroughout Africa (Geddes, 1990). Itresults from co-infection of sweetpotatoplants with the aphid-borneSweetpotato feathery mottle virus(SPFMV) and the B. tabaci-borneSweetpotato chlorotic stunt virus(SPCSV) (Schaefers and Terry, 1976;Gibson et al., 1998).
Status of CMD and SPVDResearch
Since its description more than acentury ago, CMD has receivedsignificant research attention. Fauquetand Fargette (1990) and Thresh et al.(1994; 1998) have reviewed a widerange of studies on the biological andmolecular characterization, etiology,epidemiology, yield loss and control ofthese viruses. Fishpool and Burban(1994) and Legg (1994) have reviewedstudies on B. tabaci as the vector ofcassava mosaic geminiviruses,including work on biotypecharacterization, population dynamics,virus transmission and movementwithin cassava fields. The emergence ofa severe form of CMD in the 1990s(Otim-Nape et al., 1997; Legg andOgwal, 1998; Legg, 1999) has had adevastating effect on cassavacultivation across a large area of Eastand Central Africa. Rapid expansion of
the pandemic has been associated witha new, more virulent, cassava mosaicbegomovirus (Deng et al., 1997; Zhouet al., 1997). Super-abundant B. tabacipopulations associated with thepandemic also have been shown toresult from a synergistic interactionwith severely CMD-diseased cassavaplants, in which the diseased plantsappear to promote B. tabaci populationincrease, possibly through the diseaseimproving the food quality of the plantfor B. tabaci (Colvin et al., 1999).However, evidence has been presentedfor a link between a distinct B. tabacigenotype cluster and the pandemic inUganda (Legg et al., 2002).
In contrast to CMD, knowledge ofSPVD and the role of B. tabaci in theecology of the disease is extremelylimited. Studies have been restrictedlargely to virus characterization,etiology and transmission, virus-virusinteractions, and yield lossassessments (Schaefers and Terry,1976; Hahn, 1979; Gibson et al., 1997;1998). Although the disease is knownto occur widely in sub-Saharan Africa,virtually nothing is known about itsprevalence in the major sweetpotatocultivation areas and at the inceptionof this study published incidence datawere available only for Uganda (Arituaet al., 1998).
Although significant progress hadbeen made previously in characterizingwhitefly-transmitted viruses andwhiteflies associated with CMD andSPVD, at the time of launching thepresent study, significant gaps inunderstanding remained (Table 1). Thiswas most apparent for B. tabaci, wherevery little research other than one-timeassessments of abundance and limitedpopulation dynamics studies had beendone outside Uganda and the IvoryCoast.
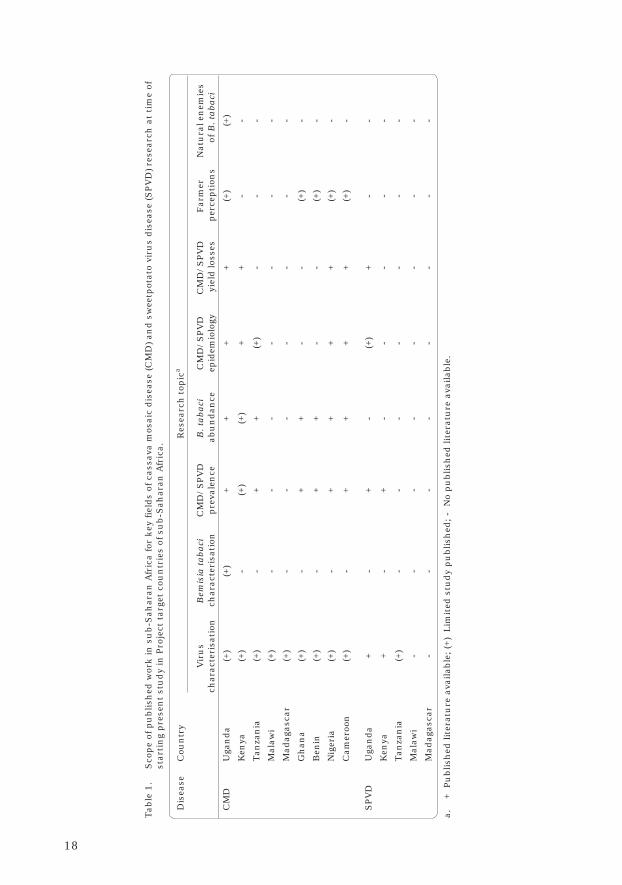
18
Whiteflies and Whitefly-borne Viruses in the Tropics
Table
1.
Sco
pe
of p
ublish
ed w
ork in
su
b-S
ah
ara
n A
fric
a for
key
fie
lds
of c
ass
ava
mos
aic
dis
ease
(C
MD
) an
d s
wee
tpot
ato
vir
us
dis
ease
(S
PV
D) re
searc
h a
t ti
me
ofst
art
ing
pre
sen
t st
udy
in P
roje
ct t
arg
et c
oun
trie
s of
su
b-S
ah
ara
n A
fric
a.
Dis
ease
Cou
ntr
yR
esea
rch
top
ica
Vir
us
Bem
isia
ta
ba
ciC
MD
/S
PV
DB
. ta
ba
ciC
MD
/S
PV
DC
MD
/S
PV
DF
arm
erN
atu
ral e
nem
ies
char
acte
risa
tion
ch
arac
teri
sati
onpre
vale
nce
abu
ndan
ceep
idem
iolo
gy y
ield
los
ses
per
cepti
ons
of
B. ta
ba
ci
CM
DU
gan
da
(+)
(+)
++
++
(+)
(+)
Ken
ya(+
)-
(+)
(+)
++
--
Tan
zan
ia(+
)-
++
(+)
--
-
Mal
awi
(+)
--
--
--
-
Mad
agas
car
(+)
--
--
--
-
Gh
ana
(+)
-+
+-
-(+
)-
Ben
in(+
)-
++
--
(+)
-
Nig
eria
(+)
-+
++
+(+
)-
Cam
eroo
n(+
)-
++
++
(+)
-
SPV
DU
gan
da
+-
+-
(+)
+-
-
Ken
ya+
-+
--
--
-
Tan
zan
ia(+
)-
--
--
--
Mal
awi
--
--
--
--
Mad
agas
car
--
--
--
--
a.
+ Pu
blish
ed lit
eratu
re a
vailable
; (+
) L
imit
ed s
tudy
pu
blish
ed; -
No
pu
blish
ed lit
eratu
re a
vailable
.

19
Introduction
Sub-project on Whitefliesas Vectors of Viruses of
Cassava and Sweetpotatoin Sub-Saharan Africa
Within the framework of the TropicalWhitefly Integrated Pest Management(TWF-IPM) Project, the purpose of thediagnostic phase of the sub-project wasto gather, generate and analysebaseline data relevant to the diagnosisand characterization of whiteflyproblems and whitefly-transmittedvirus problems in cassava andsweetpotato. The rationale behind thediagnostic phase was therefore toprovide essential baseline informationon CMD, SPVD and whiteflies oncassava and sweetpotato, to serve asthe basis for the development andimplementation of targeted andappropriate IPM strategies insubsequent phases of the project.
The sub-project involvedcollaboration among three internationalagricultural research centres, onenational agricultural researchorganization in each of nine Africancountries, and three other researchinstitutions outside Africa (Table 2).The inclusion of nine countries at thisstage of the project and the use of astandardized protocol provided aunique opportunity to obtain data thatcould be compared from country tocountry and region to region. The scopeof the study also facilitated the firstsystematic collection andcharacterization of whitefly-transmittedviruses, whitefly and whitefly naturalenemy specimens from cassava andsweetpotato in sub-Saharan Africa.
Collaboration and selection oftarget areas
Countries were identified forparticipation in the sub-projectprimarily on the basis of theimportance of cassava and/orsweetpotato cultivation to their
farmers, although security concernsprecluded the participation of some ofthe major producers of each crop. Thenine African countries that wereextensively surveyed—Ghana, Benin,Nigeria, Cameroon, Uganda, Tanzania,Kenya, Malawi and Madagascar—together represented 62% of cassavaand 56% of sweetpotato production inAfrica (FAO, 1999). Zambia wasinvolved in the project only to a limitedextent, implementing a sweetpotatovirus diagnostic survey in collaborationwith the Natural Resources Institute,UK. Sweetpotato work was carried outonly in eastern and southern Africa,because the crop is not grown widelyin West Africa. Table 2 summarizes theroles of each of the partners involvedin the diagnostic phase.
Implementation of the workplan, results and analysis
Diagnostic surveys were conductedaccording to protocols set out in thestandardized methodology agreedamong the project partners. In eachcountry, three or four target areas wereidentified for the survey. The criteriafor selection of these areas were thatthey should be major root crop-producing zones and preferably shouldhave contrasting agro-ecologicalcharacteristics. In each ecozone, thesites were selected at about 20-kmintervals along roads. Where nocassava farms were encountered at the20-km point, the next suitable sitethereafter was selected. Field data andbiological specimens were collectedfrom 3-6 month-old cassava plantingson surveyed farms and producers wereinterviewed using the standardizedproject questionnaires.
Between 10 and 42 locations wereused for sampling in each target area,and at each location a single set ofsamples and data was collected. Thedata set comprised: (1) a producerquestionnaire relating to perceptions

20
Whiteflies and Whitefly-borne Viruses in the Tropics
Table 2. Partners involved in the diagnostic phase of the sub-project on Whiteflies as Vectors ofViruses of Cassava and Sweetpotato in sub-Saharan Africa and their roles.
Institution Country Rolea
International agricultural research centres:
International Institute of Tropical Agriculture (IITA) Uganda/Benin Co-ordination
Centro Internacional de la Papa (CIP) Uganda SPVD epidemiology
International Centre of Insect Physiology and Kenya Whitefly and naturalEcology (ICIPE) enemy species
identification
National agricultural research organizations/institutes:
Plant Protection and Regulatory Services Directorate Ghana Diagnostic survey(PPRSD)
Institut National de Recherche Agronomique Benin Diagnostic surveydu Bénin (INRAB)
National Root Crops Research Institute (NRCRI) Nigeria Diagnostic survey
Institut de Recherche Agronomique (IRA) Cameroon Diagnostic survey
National Agricultural Research Organisation (NARO) Uganda Diagnostic survey
Tanzania Root and Tuber Crops Programme (TRTCP) Tanzania Diagnostic survey
Kenya Agricultural Research Institute (KARI) Kenya Diagnostic survey
Chitedze Research Station (CRS) Malawi Diagnostic survey
Centre National de Recherche Appliquée auDéveloppement Rural (FOFIFA) Madagascar Diagnostic survey
Mount Makulu Research Station Zambia Diagnostic survey(SPVD only)
Specialized research organizations:
Natural Resources Institute (NRI) United Kingdom Epidemiology of CMDand SPVD
Biologische Bundesanstalt für Land und Germany Sweetpotato virusFortwirtschaft (BBA) diagnostics
John Innes Centre (JIC) United Kingdom Moleculardiagnostics ofcassava viruses andwhiteflies
a. CMD, cassava mosaic disease; SPVD, sweetpotato virus disease.
and management of whiteflies andwhitefly-transmitted virus problems;(2) biological samples of whiteflies,virus-diseased leaf and stem tissues,and whitefly natural enemies; and(3) field assessments of diseaseincidence, virus symptoms and whiteflyabundance. Before carrying out thesurvey, lead scientists in each countrywere trained in survey methods.Surveys were conducted betweenNovember 1997 and September 1998.The results are reported in thesubsequent country chapters in this
section. Data from the producerquestionnaires for all countries wereentered into a single database fromwhich they could be summarized andqueried. Field data from all countrieswere summarized and integrated todevelop maps (Chapter 1.14, thisvolume). Studies conducted inpartnership with research institutionsoutside Africa were initiated in early1998 and completed by mid-1999;results are reported later in this section(Chapters 1.11, 1.12 and 1.13) or inSection 5 (Chapter 5.3).

21
Introduction
References
Aritua, V.; Adipala, E.; Carey, E. E.;Gibson, R. W. 1998. The incidenceof sweetpotato virus disease andvirus resistance of sweetpotatogrown in Uganda. Ann. Appl. Biol.132:399-411.
Bellotti, A. C.; Braun, A. R.; Arias, B.;Castillo, J. A.; Guerrero, J. M. 1994.Origin and management ofneotropical cassava arthropodpests. Afr. Crop Sci. J. 2:407-418.
Bock, K. R.; Woods, R. D. 1983. Etiologyof African cassava mosaic disease.Plant Dis. 67:944-955.
Boher, B.; Verdier, V. 1994. Cassavabacterial blight in Africa: The stateof knowledge and implications fordesigning control strategies. Afr.Crop Sci. J. 2:505-509.
Chant, S. R. 1958. Studies on thetransmission of cassava mosaicvirus by Bemisia spp. (Aleyrodidae).Ann. Appl. Biol. 46:210-215.
Colvin, J.; Otim-Nape, G. W.; Holt, J.;Omongo, C.; Seal, S.; Stevenson, P.;Gibson, R.; Cooter, R. J.; Thresh, J.M. 1999. Factors driving the currentepidemic of severe cassava mosaicdisease in East Africa, In: Procs.VIIth International Plant VirusEpidemiology Symposium, 11-16April 1999, Almeria, ES. p. 76-77.
Dade, H. A. 1930. Cassava mosaic. In:Yearbook. Department ofAgriculture, GH. p. 245-247.
Deighton, F. C. 1926. Annual report ofthe Lands and ForestryDepartment. Freetown, SL. p. 1-2.
Deng, D.; Otim-Nape, G. W.; Sangare, A.;Ogwal, S.; Beachy, R. N.; Fauquet,C. M. 1997. Presence of a new virusclosely associated with cassavamosaic outbreak in Uganda. Afr. J.Root Tuber Crops 2:23-28.
Dubern, J. 1979. Quelques propriétés dela mosaïque africaine du manioc. I:La transmission. J. Phytopathol.96:25-39.
Fauquet, C.; Fargette, D. 1990. Africancassava mosaic virus: Etiology,epidemiology and control. Plant Dis.74:404-411.
Fishpool, L. –D .C.; Burban, C. 1994.Bemisia tabaci: The whitefly vectorof African cassava mosaicgeminivirus. Trop. Sci. 34:55-72.
FAO (Food and Agriculture Organizationof the United Nations). 1999.Cassava production data 1998.Available in: http://www.fao.org
FAO (Food and Agriculture Organizationof the United Nations). 2004.Cassava production data 2003.Available in: http://www.fao.org
François, E. 1937. Un grave peril: Lamosaïque du manioc. Agron. Colon.26:33-38.
Frohlich, D. R.; Brown, J. K.; Bedford, I.;Markham, P. 1996. Mitochondrial16S ribosomal subunit as amolecular marker in Bemisia, andimplications for populationvariability. In: Gerling, D.; Mayer,R. T. (eds.). Bemisia 1995:Taxonomy, biology, damage, controland management. Intercept,Andover, GB. p. 143-145.
Geddes, A. M. 1990. The relativeimportance of crop pests in sub-Saharan Africa. Bulletin no. 36,Natural Resources Institute (NRI),Chatham, GB.
Gibson, R. W.; Mwanga, R. O. M.; Kasule,S.; Mpembe, I.; Carey, E. E. 1997.Apparent absence of viruses inmost symptomless field-grownsweetpotato in Uganda. Ann. Appl.Biol. 130:481-490.
Gibson, R. W.; Mpembe, I.; Alicai, T.;Carey, E. E.; Mwanga, R. O. M.;Seal, S. E.; Vetten, H. J. 1998.Symptoms, aetiology and serologicalanalysis of sweetpotato virusdisease in Uganda. Plant Pathol.47:95-102.
Golding, F. D. 1936. Cassava mosaic inSouthern Nigeria. Bull. Dept. Agric.11:1-10.

22
Whiteflies and Whitefly-borne Viruses in the Tropics
Hahn, S. K. 1979. Effects of sweetpotatovirus disease (SPVD) on growth andyield of sweetpotato. Exp. Agric.15:253-256.
Hall, F. W. 1928. Annual report.Department of Agriculture, Uganda.Government Printer, Entebbe, UG.35 p.
Herren, H. R.; Neuenschwander, P. 1991.Biological control of cassava pestsin Africa. Ann. Rev. Entomol.36:257-283.
Hillocks, R. J. 2002. Cassava in Africa. In:Hillocks, R. J.; Thresh, J. M.;Bellotti, A. C. (eds.). Cassava:Biology, production and utilization.CAB International, Wallingford, GB.p. 41-54.
Hong, Y. G.; Robinson, D. J.; Harrison,B. D. 1993. Nucleotide sequenceevidence for the occurrence of threedistinct whitefly-transmittedgeminiviruses in cassava. J. Gen.Virol. 74:2437-2443.
Jennings, D. L. 1970. Cassava in Africa.Field Crop Abstr. 23:271-278.
Joyce, R. J. V. 1955. Cotton spraying inthe Sudan Gezira. II. Entomologicalproblems arising from spraying.FAO Plant Prot. Bull. 3:97-103.
Kirkpatrick, T. W. 1931. Leaf-curl ofcotton in the Sudan. Bull. Entomol.Res. 21:127-137.
Kufferath, H.; Ghesquière, J. 1932. Lamosaïque du manioc. Compte-Rendu Soc. Biol. Belge 109:1146-1148.
Legg, J. P. 1994. Bemisia tabaci: Thewhitefly vector of cassava mosaicgeminiviruses in Africa: Anecological perspective. Afr. Crop Sci.J. 2:437-448.
Legg, J .P. 1999. Emergence, spread andstrategies for controlling thepandemic of cassava mosaic virusdisease in east and central Africa.Crop Prot. 18:627-637.
Legg, J. P.; Ogwal, S. 1998. Changes inthe incidence of African cassavamosaic geminivirus and theabundance of its whitefly vectoralong south-north transects inUganda. J. Appl. Entomol.122:169-178.
Legg, J. P.; French, R.; Rogan, D.; Okao-Okuja, G.; Brown, J. K. 2002. Adistinct, invasive Bemisia tabaci(Gennadius) (Hemiptera:Sternorrhyncha: Aleyrodidae)genotype cluster is associated withthe epidemic of severe cassavamosaic virus disease in Uganda.Mol. Ecol. 11(7):1219-1229.
Mound, L. A.; Halsey, S. H. 1978.Whitefly of the world. A systematiccatalogue of the Aleyrodidae(Homoptera) with host plant andnatural enemy data. John Wiley,Chichester, GB. 340 p.
Neuenschwander, P. 1994. Control of thecassava mealy bug in Africa:Lessons from a biological controlproject. Afr. Crop Sci. J. 2:369-384.
Otim-Nape, G. W.; Bua, A.; Thresh, J. M.;Baguma, Y.; Ogwal, S.; Semakula,G. N.; Acola, G.; Byabakama, B.;Martin, A. 1997. Cassava mosaicvirus disease in Uganda: Thecurrent pandemic and approachesto control. Natural ResourcesInstitute (NRI), Chatham, GB. 65 p.
Schaefers, G. A.; Terry, E. R. 1976. Insecttransmission of sweetpotatodiseases in Nigeria. Plant Pathol.66:642-645.
Storey, H. H.; Nichols, R. F. W. 1938.Studies on the mosaic of cassava.Ann. Appl. Biol. 25:790-806.
Swanson, M. M.; Harrison, B. D. 1994.Properties, relationships anddistribution of cassava mosaicgeminiviruses. Trop. Sci. 34:15-25.
Thresh, J. M.; Fargette, D.; Otim-Nape,G. W. 1994. The viruses and virusdiseases of cassava in Africa. Afr.Crop Sci. J. 2:459-478.

23
Introduction
Thresh, J. M.; Otim-Nape, G. W.; Legg,J. P.; Fargette, D. 1997. Africancassava mosaic disease: Themagnitude of the problem? Afr. J.Root Tuber Crops 2:13-19.
Thresh, J. M.; Otim-Nape, G. W.;Thankappan, M.; Muniyappa, V.1998. The mosaic diseases ofcassava in Africa and India causedby whitefly-borne geminiviruses.Rev. Plant Pathol. 77:935-945.
Warburg, O. 1894. Die KulturpflanzenUsambaras. Mitt. Dtsch. Schutz.7:131.
Yaninek, J. S.; Herren, H. R. 1988.Introduction and spread of thecassava green mite Mononychellustanajoa (Bondar) (Acari:Tetranychidae), an exotic pest inAfrica and the search forappropriate control methods: Areview. Bull. Entomol. Res. 78:1-13.
Yaninek, J. S.; Onzo, A.; Ojo, J. B. 1993.Continent-wide releases ofneotropical phytoseiids against theexotic cassava green mite in Africa.Exp. Appl. Acarol. 17:145-160.
Zhou, X.; Liu, Y.; Calvert, L.; Muñoz, C.;Otim-Nape, G. W.; Robinson, D. J.;Harrison, B. D. 1997. Evidence thatDNA-A of a geminivirus associatedwith severe cassava mosaic diseasein Uganda has arisen byinterspecific recombination. J. Gen.Virol. 78:2101-2111.
Zimmerman, A. 1906. DieKrauselkrankheit des Maniok.Pflanzer 2:145.

24
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 1.2
GhanaAnthony Cudjoe*, Joseph Gyamenah* and
Braima James**
Introduction
Cassava mosaic disease (CMD) is amajor food production constraint inAfrica (Thresh et al., 1994). It is causedby begomoviruses, which are vectoredby the whitefly Bemisia tabaci(Gennadius) and can be spreadthrough infected planting material.Yield losses caused by the disease arehigher if vegetative planting materialssprout with symptoms (cuttinginfection) than if initially disease-freematerial becomes infected by B. tabacifollowing sprouting (current seasonwhitefly-borne infection) (Fauquet andFargette, 1990). In Ghana, recent workon Bemisia whiteflies and CMD hasbeen carried out under the auspices ofa regional project, EcologicallySustainable Cassava Plant Protection(ESCaPP) (Yaninek et al., 1994).Unpublished results of ESCaPPdiagnostic surveys showed cassava(Manihot esculenta Crantz) to be themost widely planted arable crop in thecountry (45% site incidence), followedby maize (Zea mays L.) (15%) and yam(Dioscorea spp.) (5%).
Whitefly survey samples consistedlargely of B. tabaci. B. afer (Priesner
and Hosny) was unevenly distributedand less abundant (Sotomey et al.,1995). Across ecozones and seasons,CMD was the most common cassavapest constraint both in terms of fielddata and farmer perceptions. In thefield, total plant incidence of CMD was76% in the dry season and 68% in thewet and, in 56% of the villages, farmersranked CMD as the main disease. Onan ascending 1-5 scale of damageclasses, 22% of plants were in class2 (mild), 51% in class 3 (moderate),26% in class 4 (severe) and none inclass 5 (very severe). Previous attemptsto develop CMD control measures inGhana have included preliminaryevaluation of local cassava varieties inrelation to their susceptibility to CMDand laboratory studies to producevirus-free tissue culture materials oflocal germplasm (Bieler et al., 1996;Kissiedu et al., 1996). In subsequentcollaborative epidemiology trials,whitefly infection was consistentlylowest in the variety TMS 30001 whencompared to other improved or localvarieties (Legg et al., 1997; IITA, 1998;James et al., 1998). The role ofparasitoids, other natural enemies,genetic heterogeneity in Bemisiapopulations and the identity of cassavaviruses/viral strains are yet to beinvestigated in the country.
This chapter gives results of acountrywide survey in Ghana,conducted in November and December
* Plant Protection and Regulatory ServicesDirectorate (PPRSD), Ministry of Food andAgriculture, Ghana.
** International Institute of Tropical Agriculture(IITA), Biological Control Center for Africa,Cotonou, Benin.

25
Ghana
1997, on 80 farms. These included25 sites in each of the rainforest andtransition forest, 10 sites in the“coastal” savannah and 20 sites in thewet savannah (Figure 1).
symptoms in various local names.Some of these, for example, bankyekwata and kwata, described theappearance of CMD symptoms. Suchindigenous knowledge will facilitatefarmer participatory learning andresearch activities. Bemisia whiteflyabundance averaged 1.3 adults percassava shoot tip in the rainforest,1.0 in transition forest, 11.8 in coastalsavannah and 0.3 in wet savannah (seealso Figure 2 in Chapter 1.14, thisvolume). Whitefly mean abundance(logarithm transformed counts) wassignificantly higher in the coastalsavannah than in each of the otherecozones (t > 12.1; P < 0.05). Thenatural enemy fauna associated withthe whiteflies included thehymenopteran parasitoid Encarsiasophia (Girault and Dodd), which wasreared from Bemisia mummies at foursurvey sites.
Increased Socio-EconomicUnderstanding
Cassava production was characterizedby small landholdings, typically lessthan 1 ha in size, and a preponderanceof local landraces. The intensity ofcassava farming decreased along therainforest-savannah axis. The five mostprofitable crops listed by farmers werecassava (45%), yam (18%), maize(11%), cowpea (Vigna unguiculata [L.]Walp.) (6%) and tomato (Lycopersiconesculentum Mill.) (5%). These othercrops were planted in association with,or near to, cassava fields but cassavawas the most common crop (40% siteincidence) planted in nearby fields.Among the associated crops, bothcowpea and tomato are known hostplants of B. tabaci. However,experimental data from other countriesin Africa show that the B. tabacibiotype that occurs on cassava islargely restricted to that crop (Burbanet al., 1992; Legg et al., 1994).
Figure 1. Areas surveyed for whitefly incidenceand cassava mosaic disease inGhana.
Burkina Faso
Côted’Ivoire
Togo
Humid forest
Ghana
Derivedsavannah
Southern guinea savannah
Northern guinea savannah
Coastal derivedsavannah
Increased BiologicalUnderstanding
B. tabaci, B. afer and Trialeurodesvaporariorum (Westwood) wereidentified from adult whitefly samplescollected from cassava in Ghana.Whilst B. tabaci occurred at all surveysites, B. afer was recorded from only3.8% of the sites and comprised 1.9%of the 151 specimens determined.T. vaporariorum was collected from onlyone location. Generally, farmers wereunaware of Bemisia whiteflies and didnot recognize them by specific localnames; only 2% of them knew that theinsects caused serious problems tocassava. By contrast, 54% of farmersknew that CMD caused seriousproblems and described the disease

26
Whiteflies and Whitefly-borne Viruses in the Tropics
Priority cassava pests listed byfarmers were vertebrates (16%), weeds(13%), termites and weevils (10% each)and CMD (6%). Overall, CMD incidencewas mainly attributable to cuttinginfection (Figure 2). The incidence ofcutting infection averaged 60% in therainforest, 55% in the transition forest,63% in the coastal savannah and 51%in the wet savannah. The incidence ofwhitefly infection, transformed to allowfor the effect of multiple infection(Gregory, 1948), was significantlyhigher in the coastal savannah andrainforest than in the transition forestor wet savannah (t > 2.9; P < 0.05).Whitefly abundance showed nocorrelation with the incidence ofwhitefly infection. In terms of reducingCMD incidence, the transition forestand wet savannah would seem toprovide better sites for multiplication ofclean planting material.
to pest-induced losses. In view of thehigher proportion of plants in themoderate and severe damage categoriesin the transition forest, significant rootyield losses could be expected in thisecozone. However, yield loss estimatesprovided by farmers appeared to beunrealistically high. For example, about26% of farmers estimated losses at50% and 20% of farmers at 75%. Suchloss estimates would need to bevalidated, especially since 91% of thefarmers reported that they sell theircassava harvest.
Whitefly infection Cutting infection
Coa
stal
sava
nn
ah
Rai
nfo
rest
Tra
nsi
tion
fore
st
Wet
sava
nn
ah
Per
cen
tage
of pla
nts
wit
h C
MD
sym
pto
ms
100
80
60
40
20
0
Figure 2. Cassava mosaic disease (CMD)incidence and source of infection inthe ecozones of Ghana.
Per
cen
tage
of pla
nts
wit
hdiffe
ren
t C
MD
sev
erit
ies
Slightdamage
Moderatedamage
Seriousdamage
Rainforest
Wet savannah
100908070605040302010
0
Figure 3. Cassava mosaic disease (CMD)damage severity in the ecozones ofGhana.
Transition forest
Coastal savannah
Across ecozones, 24%-32% ofplants showed no CMD symptoms.About 10% of the plants showedserious (score 4) to severe (score 5)damage symptoms (Figure 3). Theplants were 3-6 months old, a growthstage during which storage rootformation and development is initiatedand root yield is particularly vulnerable
Farmers’ opinion was almostequally divided on whether or not CMDwas becoming more severe, and asizeable proportion (34%) consideredthat the disease was not a yearlyoccurrence. Extension and trainingsupport for whitefly/CMD problemsappeared low, since 89% of farmersreported receiving no technicalinformation or assistance with theseproblems. In view of this, it is notsurprising that only 1% of farmerspracticed any deliberate whitefly/CMDcontrol. Local varieties predominated(54% site incidence) and 18% of thefarmers ranked Busuminsia as theirsingle most preferred variety, followedby three other local varieties, Santom

27
Ghana
(6.2%), Bakentenma (5%) and Kokoo(5%). Adoption of improved varietieswas low: only 11% of farmers plantedTMS 30572 (originating from theInternational Institute of TropicalAgriculture [IITA]) and “Agric” varieties(a locally adapted material of IITAorigin) recommended by the nationalextension service on the basis of theirCMD resistance. Agronomic featureswere more important criteria forselecting planting material than weredisease-specific characteristics.
Where disease-specific selectioncriteria were used, only 10% of thefarmers reported that the method waseffective, while 9% noticed that the“clean” material became diseased afterplanting. Since the single mostimportant primary source of plantingmaterial was farmers’ own fields(Figure 4), adoption of appropriateprocedures for selecting plantingmaterial would significantly reducerecycling of cassava mosaic viruses.About 60% of farmers reported thatthey encountered shortage of plantingmaterial in some years. Shortages likethese, coupled with the observed highincidence of cutting infection, implythat rapid multiplication schemeswould be required to ensure timelyavailability of clean and healthyplanting materials at farm level. Nofarmer reported pesticide use as acontrol method against the whiteflies orthe disease but about 61% of farmers
rogue diseased plants, mostly (55% offarmers) within the first month ofplanting.
Conclusions
This study has increased field-levelunderstanding of whiteflies and CMDincidence, and provides a strong basisfor the development of ecologicallysound management practices forwhitefly/CMD problems in Ghana. Theincidence of CMD infection by B. tabaciwas low and the disease appears to bespread mainly through infectedcuttings. The preponderance of localcassava varieties over proven CMD-resistant improved varieties contributesto the incidence of the disease in thecountry. Two key issues that need to beaddressed are the quantification ofyield losses in the preferred localcultivars and, associated with this, anassessment of why CMD-resistantvarieties have not been adopted widely.
Assuming that yield losses warrantthe use of CMD control measures,farmer participatory learning activitieswill promote farm-level selection ofparent planting material that is CMDsymptom free for making cuttings.Related to this, farmer-led rapidmultiplication schemes would berequired to increase the availability andadoption of CMD-resistant varieties,provided that evaluations linked tosuch schemes indicate that some ofthese varieties are acceptable tofarmers. Research should also evaluatefarmers’ preferred local varieties aspossible sources of CMD resistance inbreeding programs since this approachhas provided important new impetus toresistance breeding programs forcassava elsewhere (Dixon et al., 1992).
Little is known about temporal andspatial changes in the abundance ofB. tabaci in Ghana, particularly in
Own farm
Neighbour
Extension
Market
Relative frequency (%) of plantingmaterial source
Figure 4. Farmers’ sources of cassavaplanting material in Ghana.
0 10 20 30 40 50 60 70

28
Whiteflies and Whitefly-borne Viruses in the Tropics
relation to its ability to transmitcassava mosaic viruses over thegrowing period of the crop. Fieldstudies therefore might be necessary toquantify such changes, assess whetherparasitism by E. sophia has anysignificant impact on the insect’sabundance and, in epidemiology trials,to ascertain the ability of B. tabaci tospread cassava mosaic viruses to arange of cassava varieties.
National research capacity needs tobe enhanced in order to allow projectpartners to undertake further researchon the whitefly/CMD issues raisedhere. In this regard, training of nationalprogram partners in appropriate areaswould strengthen the collaborativelinkages initiated in the pilot phase ofthe project and provide a strong basefor providing sustainable technicalsupport to cassava farmers. Key areasfor strengthened national researchcapacity include post-graduate degreetraining in B. tabaci bionomics andcassava mosaic virology, and field-based training of technicians andextension workers in participatoryprocesses and facilitation. Technicianand extension training will increasescientific literacy in farmingcommunities and foster action researchand learning with farmers. This willimprove farmers’ currently poor accessto information, will help themunderstand the causes and nature ofthe disease problem and will highlighteffective cultural control options suchas planting material selection androguing.
References
Bieler, P.; Ekanayake, I. J.; Dossou, R.;Sanni, A. O.; Ahihou, K.; Alhassan,A. Y.; Ahiabu, R.; Yaninek, J. S.1996. Production of high qualitycassava planting materials: Acollaborative effort. Trop. RootTuber Crops Bull. 9(1):1-3.
Burban, C.; Fishpool, L. D. C.; Fauquet,C.; Fargette, D.; Thouvenel, J. -C.1992. Host associated biotypeswithin West African populations ofthe whitefly Bemisia tabaci (Genn.)(Hom., Aleyrodidae). J. Appl.Entomol. 113:416-423.
Dixon, A. G. O.; Asiedu, R.; Hahn, S. K.1992. Cassava germplasmenhancement at IITA. In: Akoroda,M. O.; Arene, O. B. (eds.).Promotion of root crop-basedindustries: An incentive for researchand development. Procs. 4thTriennial Symposium of theInternational Society for TropicalRoot Crops-Africa Branch (ISTRC-AB), 5-8 December 1989, Kinshasa,CD. p. 83-87.
Fauquet, C.; Fargette, D. 1990. Africancassava mosaic virus: Etiology,epidemiology, and control. PlantDis. 74:404-411.
Gregory, P. H. 1948. The multipleinfection transformation. Ann. Appl.Biol. 35:412-417.
IITA (International Institute of TropicalAgriculture). 1998. EcologicallySustainable Cassava PlantProtection (ESCaPP) in SouthAmerica and Africa. Africa: Projectfindings and recommendations.Ibadan, NG.
James, B. D.; Legg, J. P.; Gbaguidi, J. B.;Annang, D.; Asiabaka, C. C.;Cudjoe, A. R.; Echendu, T. N. C.;Maroya, N. G.; Ntonifor, N.; Ogbe,F.; Salawu, R. A.; Tumanteh, J. A.1988. Understanding farm-levelepidemiology of cassava mosaicdisease. In: Abstracts, 7th TriennialSymposium of the InternationalSociety for Tropical Root Crops-Africa Branch (ISTRC-AB), 11-17October 1988, Cotonou, BJ. p. 122.
Kissiedu, A. F. K.; Asare Bediako, A.;Afuakwa, J. J.; Okai, E.; Missah, A.1996. National Root and TuberCrops Improvement Project(NRTCIP), Ghana-highlights ofactivities and achievements 1992-93. Trop. Root Tuber Crops Bull.,Special Edn. March 1996. p. 3-5.

29
Ghana
Legg, J. P.; James, B. D. 1998. Tacklingwhitefly and whitefly-borne virusconstraints to cassava andsweetpotato in Africa: Aims,approach, and progress of theCGIAR whitefly IPM project. In: 7thTriennial Symposium of theInternational Society for TropicalRoot Crops-Africa Branch (ISTRC-AB), 11-17 October 1988, Cotonou,BJ. p. 64.
Legg, J. P.; Gibson, R. W.; Otim-Nape, G.W. 1994. Genetic polymorphismamongst Ugandan populations ofBemisia tabaci (Gennadius)(Homoptera: Aleyrodidae), vector ofAfrican cassava mosaic geminivirus.Trop. Sci. 34:73-81.
Legg, J. P.; James, B. D.; Cudjoe, A.;Saizonou, S.; Gbaguidi, B.; Ogbe,F.; Ntonifor, N.; Ogwal, S.; Thresh,J.; Hughes, J. 1997. A regionalcollaborative approach to the studyof ACMD epidemiology in sub-Saharan Africa. Afr. Crop Sci. J.3:1021-1033.
Sotomey, M.; James, B. D.; Gbaguidi, B.1995. Les espèces de Bemisia sur lemanioc en Afrique occidentale etcentrale. In: 11th Meeting andScientific Conference of the AfricanAssociation of Insect Scientists,6-11 August 1995, Yamoussoukro,CI. p. 4.
Thresh, J. M.; Fishpool, L. D. C.; Otim-Nape, G. W.; Fargette, D. 1994.African cassava mosaic virusdisease: An under-estimated andunsolved problem. Trop. Sci.34:3-14.
Yaninek, J. S.; James, B. D.; Bieler, P.1994. Ecologically SustainableCassava Plant Protection (ESCaPP):A model for environmentally soundpest management in Africa. Afr.Crop Sci. J. 2:553-562.

30
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 1.3
BeninBrice Gbaguidi*, Braima James* and
Symphorien Saizonou**
Introduction
Preliminary research on cassava pestsand diseases in Benin was carried outas part of a regional project forEcologically Sustainable Cassava PlantProtection (ESCaPP) (Yaninek et al.,1994). Unpublished results of ESCaPPdiagnostic surveys show cassava(Manihot esculenta Crantz) to be thepredominant arable crop (87% siteincidence) in the country and the nextmost common crops to be maize (Zeamays L.) with 30% site incidence andcowpea (Vigna unguiculata [L.] Walp.)with 26%. Summaries of plantprotection aspects of the survey showcassava mosaic disease (CMD) causedby cassava mosaic begomoviruses(CMBs) and vectored by the whiteflyBemisia tabaci (Gennadius) to be themost common biotic constraint oncassava in the country, with total plantincidence of 49% in the dry season and57% in the wet. Survey samples ofred-eyed whitefly nymphs comprisedB. tabaci and B. afer (Priesner andHosny) although the latter was lessabundant, occurring in only 4% of thesamples (Sotomey et al., 1995). TheCMD damage score was low, theaverage falling within class 2
(indicating “slight damage” on a1-5 scale) across ecozones andseasons. During the surveys, in focusgroup interviews at village level,farmers at 68% of survey sites ratedCMD as the main disease problem. Insubsequent preliminary CMDepidemiology trials (Legg et al., 1997;IITA, 1998; James et al., 1998), currentseason whitefly infection wasconsistently lowest in the varietyTMS 30001 when compared with otherimproved or local varieties. In a relatedpopulation dynamics study, theparasitoid Encarsia sophia (Girault andDodd) was identified from Bemisiamummies at each of six trial sites inthe transition forest, and wet and drysavannahs (James and Gbaguidi,unpublished data). Geneticheterogeneity in B. tabaci populations,which is known to influence CMDepidemiology (Burban et al., 1992; Legget al., 1994), and the identities ofcassava viruses and virus strains areyet to be investigated in the country.
This chapter reports the results ofa countrywide diagnostic survey ofwhiteflies and CMD in Benin,conducted in November and December1997, on 60 farms distributed in thetransition forest (28 sites), drysavannah (19 sites) and wet savannah(13 sites) (Figure 1). Farmerrespondents in the survey were mostlymen (93%) and landowners with over5 years of cassava farming experience.
* International Institute of Tropical Agriculture(IITA), Biological Control Center for Africa,Cotonou, Benin.
** Service Protection des Végétaux et ContrôlePhytosanitaire, Porto Novo, Benin.

31
Benin
Based on the survey results, thechapter identifies areas requiring moreresearch attention to further ourunderstanding of CMD in the countryand to serve as a basis for developingand implementing an integratedapproach to managing this disease.
Increased BiologicalUnderstanding
The co-existence of B. tabaci and B.afer on cassava in Benin wasconfirmed by the determination ofthese species from adult specimens. Invarious local languages, farmersrecognized whiteflies simply as“insects”. On the other hand, amongthe various local names used todescribe CMD, the names edjekpo,
ekpo, goudou and kpanro referspecifically to the resemblance of CMDsymptoms to those of leprosy. Thisindigenous knowledge will greatlyfacilitate farmer participatory researchand learning activities—activities thatseem especially important given that ahigh proportion of farmers did notknow that whiteflies (60% farmers) andCMD (50% farmers) caused seriousproblems to cassava. Whilst B. tabacioccurred at all 60 survey sites, B. aferwas recorded in 50% of the sites andcomprised 15% of the 212 specimensdetermined. Whitefly abundance waslow and averaged 3.2 adults percassava shoot tip in the transitionforest, 0.5 in the wet savannah and0.9 in the dry savannah. Whitefly meanabundance (logarithm transformedcounts) was significantly higher in thetransition forest than in either wet ordry savannah (t > 5.04; P < 0.05). Theparasitoids E. sophia and Eretmocerussp. were identified in natural enemycollections. The presence of E. sophiawas confirmed at 15 sites andEretmocerus sp. at only one site.Diseased whiteflies were not observedin the field and hence noentomopathogens were collected.Biotypes of cassava viruses identifiedare reported elsewhere (Chapter 1.11,this volume).
Increased Socio-EconomicUnderstanding
Across ecozones, cassava was typicallyproduced on farms of less than0.25 ha. The improved variety “Agric”,originating from the InternationalInstitute of Tropical Agriculture (IITA),was grown by 13% of farmers. Thelocal variety Odounbo, grown by 7% offarmers, was the next most popularvariety, while the remainder compriseda wide diversity of local cultivars. Fifty-eight percent of farmers rankedcassava as their most profitable crop,
Figure 1. Areas surveyed for whiteflyincidence and cassava mosaicdisease in Benin.
Northern guineasavannah
Southern guineasavannah
Derivedsavannah
Forest
Nigeria
Togo
Benin

32
Whiteflies and Whitefly-borne Viruses in the Tropics
compared to 33% for maize, 10% eachfor cowpea and groundnut (Arachishypogaea L.) and 5% for cotton(Gossypium hirsutum L.). These cropswere planted also in association withcassava or near to cassava fields.Cowpea, groundnut and cotton areknown crop host plants of B. tabaci,although experimental data from othercountries in Africa show that theB. tabaci biotype occurring on cassavais more or less restricted to that crop(Burban et al., 1992; Legg et al., 1994).Cassava was also the most commoncrop planted in nearby fields (38% ofall instances) and the intensity ofcassava farming gradually decreasedfrom the transition forest (2.4 nearbyfields) to wet savannah (1.9 nearbyfields) and dry savannah (0.2 nearbyfields).
Average CMD incidence rangedfrom 25% to 49% across ecozones(Figure 2), with infected cuttingsproviding the more important source ofinfection. The incidence of cuttinginfection did not differ significantlybetween ecozones. The incidence ofwhitefly infection, transformed to allowfor the effect of multiple infection(Gregory, 1948), differed significantlyacross ecozones and was higher inboth the transition forest and wetsavannah than in the dry savannah
(t > 2.06; P < 0.05). Across ecozones,whitefly abundance showed nocorrelation with the incidence ofwhitefly infection. The pattern ofdamage was similar across ecozones,with most plants showing no CMDsymptoms and, of those visibly infectedwith the disease, only a very smallproportion showing moderate or severedamage (Figure 3). This is important inthe context of the age of cassava farmssampled. The plants were 3-6 monthsold at the time of the survey, an age atwhich storage roots form and begin todevelop and root yield is particularlyvulnerable to pest-induced losses.None of the farmers reported total lossof their cassava to CMD, probably areflection of the mild disease symptomsobserved. However, 25% of farmersattributed losses of at least 25% of theyield to the disease. Whilst this highloss estimate seems to conflict with thelow damage severity observed, 58% offarmers indicated that CMD isbecoming more severe.
60
40
20
0
Whitefly infection Cutting infection
Drysavannah
Transitionforest
Wetsavannah
Per
cen
tage
of pla
nts
wit
h C
MD
sym
pto
ms
Figure 2. Cassava mosaic disease (CMD)incidence and source of infection inthe ecozones of Benin.
Per
cen
tage
of pla
nts
wit
hdiffe
ren
t C
MD
sev
erit
ies
Slightdamage
Moderatedamage
Seriousdamage
Wet savannah
100908070605040302010
0
Transition forest
Dry savannah
Figure 3. Cassava mosaic disease (CMD)damage severity in the ecozones ofBenin.
Most farmers (78%) undertook nospecific control measures against CMDor whiteflies, and less than 10% hadreceived technical information orassistance on them. Among farmerswho attempted to control the whitefly

33
Benin
or disease problem, the main methodsused were weeding and application ofwood ash. Even though no farmerreported the use of resistant varietiesas a whitefly/disease control method,further questioning revealed that 8% offarmers listed “Agric” and 5% listedOdounbo as varieties they had plantedon the basis of recommendations thatthey were tolerant to CMD. Twentypercent of these farmers indicated thatthe better performance of thesevarieties was mainly because of bettersprouting and establishment of thecutting. Agronomic features were moreimportant than pest/disease resistanceas criteria for the selection of plantingmaterial. About 12% of the selectioncriteria related to disease-free stems,health of stems and disease resistance,whilst 23% related to size, maturity,better yield, colour and size of theparent stem.
Where disease-specific criteria wereused for selecting planting material,only 10% of the farmers reported thatthe method was effective and 7%noticed that the “clean” materialbecame diseased after planting.Because the single most frequentlycited sources of planting material werefarmers’ own fields (30%) andneighbours’ fields (22%), adoption ofappropriate planting material selectionwould be expected to reduce recyclingof CMBs considerably. Whilst nofarmer indicated roguing as a specificCMD control method, 7% of themconducted roguing to prevent andreduce disease and 2% because of poorplant growth; 7% of the farmersreported some impact from this controlmeasure. However, the current highincidence of cutting infection wouldmake roguing very labor intensive andunattractive for adoption, unlesscarried out in combination with theselection of CMD-free plantingmaterial. Pesticide use was rare.
Conclusions
In Africa, cassava root yield losses dueto CMD have been estimated at28%-40% annually (Thresh et al.,1994). In Benin, there appears to be acontradiction between field data andfarmers’ perception of the CMDproblem. The research data indicateonly a very small proportion of cassavaplants with moderate or severe diseasedamage but 25% of the farmersattribute high losses to the disease. Insuch cases, farmer participatory trialswould help farmers and researchersjointly to better understand therelationship between disease incidenceand yield, and to validate lossestimates. The studies would helpestablish whether losses to CMD reallyare significant in Benin and, if so,convince farmers of the need to adoptintegrated management practicesagainst the disease. Such an approachwill provide for action learning byfarmers to improve their access toinformation and understanding of thecauses and nature of the diseaseproblem. For example, cutting infectionwas found to be much more importantthan Bemisia whiteflies in the spread ofCMD, suggesting that a participatorylearning and extension effort focusingon planting material selection andsanitation and roguing could have apositive impact.
Generally, training of nationalprogram partners would strengthen thecollaborative links established in thepilot phase of the project and furtherenhance national capacity toundertake priority CMD and whiteflyresearch activities. For example, fieldstudies on the population dynamics ofB. tabaci would be needed tounderstand temporal and spatialchanges in the insect’s abundance inrelation to its ability to spread CMD.There is also the need to quantify theeffectiveness of parasitism by E. sophia,

34
Whiteflies and Whitefly-borne Viruses in the Tropics
particularly as a possible factorinfluencing the importance of thevector in spreading CMD. In view of thelow abundance of B. afer in thecountry, it would not be appropriate tocommit further resources toinvestigating its role in CMDepidemiology.
References
Burban, C.; Fishpool, L. D. C.; Fauquet,C.; Fargette, D.; Thouvenel, J. -C.1992. Host associated biotypeswithin West African populations ofthe whitefly Bemisia tabaci (Genn.)(Hom., Aleyrodidae). J. Appl.Entomol. 113:416-423.
Gregory, P. H. 1948. The multipleinfection transformation. Ann. Appl.Biol. 35:412-417.
IITA (International Institute of TropicalAgriculture). 1998. EcologicallySustainable Cassava PlantProtection (ESCaPP) in SouthAmerica and Africa. Africa: Projectfindings and recommendations.Ibadan, NG.
James, B. D.; Legg, J. P.; Gbaguidi, J. B.;Annang, D.; Asiabaka, C. C.;Cudjoe, A. R.; Echendu, T. N. C.;Maroya, N. G.; Ntonifor, N.; Ogbe,F.; Salawu, R. A.; Tumanteh, J. A.1998. Understanding farm-levelepidemiology of cassava mosaicdisease. In: Abstracts, 7th TriennialSymposium of the InternationalSociety for Tropical Root Crops-Africa Branch (ISTRC-AB), 11-17October 1988, Cotonou, BJ. p. 122.
Legg, J. P.; Gibson, R. W.; Otim-Nape, G.W. 1994. Genetic polymorphismamongst Ugandan populations ofBemisia tabaci (Gennadius)(Homoptera: Aleyrodidae), vector ofAfrican cassava mosaic geminivirus.Trop. Sci. 34:73-81.
Legg, J.; James, B. D.; Cudjoe, A.;Saizonou, S.; Gbaguidi, B.; Ogbe,F.; Ntonifor, N.; Ogwal, S.; Thresh,J.; Hughes, J. 1997. A regionalcollaborative approach to the studyof ACMD epidemiology in sub-Saharan Africa. Afr. Crop Sci. J.3:1021-1033.
Sotomey, M.; James, B. D.; Gbaguidi, B.1995. Les espèces de Bemisia sur lemanioc en Afrique occidentale etcentrale. In: 11th meeting andscientific conference of the AfricanAssociation of Insect Scientists,6-11 August 1995, Yamoussoukro,CI. p. 4.
Thresh, J. M.; Fishpool, L. D. C.; Otim-Nape, G. W.; Fargette, D. 1994.African cassava mosaic virusdisease: An under-estimated andunsolved problem. Trop. Sci.34:3-14.
Yaninek, J. S.; James, B. D.; Bieler, P.1994. Ecologically SustainableCassava Plant Protection (ESCaPP):A model for environmentally soundpest management in Africa. Afr.Crop Sci. J. 2:553-562.

35
Nigeria
CHAPTER 1.4
NigeriaNnamdi C. Echendu*, Joseph B. Ojo**,
Braima James*** and Brice Gbaguidi***
Introduction
Cassava (Manihot esculenta Crantz) isan important staple food crop in sub-Saharan Africa. In Nigeria,unpublished results of diagnosticsurveys by a project for EcologicallySustainable Cassava Plant Protection(ESCaPP) (Yaninek et al., 1994) showcassava to be the most important cropin the country, followed by yam(Dioscorea spp.) and maize (Zea maysL.). Cassava has the potential to yield60 t/ha of storage roots (Nweke andLynam, 1997). However, average yieldsin farmers’ fields are usually less than10 t/ha (FAO, 1989). The shortfall isassociated largely with pest anddisease problems, of which cassavamosaic disease (CMD) is one of themost important. In sub-Saharan Africaas a whole, losses to CMD have beenestimated at 15-27 million tons,representing 15%-24% of totalproduction (Thresh et al., 1997). Thedisease is caused by begomovirusestransmitted by the whitefly Bemisiatabaci (Gennadius) and is spread alsoby man through planting of infectedplanting materials (Hahn et al., 1981).The ESCaPP survey results showedCMD to be the single most common
pest constraint on cassava in Nigeriawith, on average, 85% of plants showingsymptoms in the dry season and 79% inthe wet (IITA, 1997). The surveysindicated the co-existence of B. tabaciand B. afer (Priesner & Hosny) oncassava in Nigeria, although B. afer wasless abundant and widespread thanB. tabaci (Sotomey et al., 1995).
In Nigeria, studies are continuing toexamine the begomoviruses infectingcassava and their variability (Ogbeet al., 1998). These have recorded thepresence of both African cassava mosaicvirus (ACMV) and East African cassavamosaic virus (EACMV), although earlierless comprehensive studies hadsuggested the occurrence of only ACMVin Nigeria (Swanson and Harrison,1994). Genetic heterogeneity in B. tabacipopulations, which is known toinfluence CMD epidemiology (Burban etal., 1992; Legg et al., 1994), has yet tobe investigated in the country. Inpreliminary CMD epidemiology trials,current season whitefly infection wasconsistently lowest in the varietyTMS 30001 when compared to otherimproved or local varieties (Legg et al.,1997; IITA, 1998; James et al., 1998).The development and deployment ofresistant varieties has been the mainapproach to CMD management pursuedin Nigeria (Hahn et al., 1980; 1981).Little attention has been given to thepotential for using natural enemies tomanage the whitefly vector, althoughJerath (1967) suggested that the
* National Root Crops Research Institute(NRCRI), Umudike, Nigeria.
** International Institute of TropicalAgriculture (IITA), Ibadan, Nigeria.
*** IITA, Biological Control Center for Africa,Cotonou, Benin.

36
Whiteflies and Whitefly-borne Viruses in the Tropics
parasitoid Encarsia strenua (Silvestri)and a predatory mite, Typhlodromus sp.,were possible biocontrol agents.
This report presents results of acountrywide survey of Bemisia whitefliesand CMD conducted in November andDecember 1997, on 80 farmsdistributed across the main cassava-producing agro-ecological zones ofNigeria (Figure 1). Sites were sampled inrainforest (25), transition forest (25), wetsavannah (20) and dry savannah (10)zones, and cassava plantings selectedwere 3 to 4 months old. This chaptersummarizes the results of the surveyand identifies areas requiringsubsequent attention in order to furtherour understanding of the disease and toserve as a basis for developing andimplementing the integratedmanagement of CMD in Nigeria.
were found in samples collected. Somefarmers recognized whiteflies and 17.5%of farmers knew that the insects causeda serious problem to cassava. In variouslocal languages, farmers recognizedBemisia whiteflies simply as “insects”.About 24% of farmers knew that CMDcaused serious problems and farmers indifferent localities described the diseasein various local languages as akpa,ekikang, ikwukwo, kpuojuju andnkpuk—mostly terms that refer to theappearance of the disease symptoms.Bemisia abundance was less than threeadults per cassava shoot tip (defined asthe terminal shoot with the first fiveopen leaves) in each of the ecozones.
Five species of hymenopterousparasitoids of Bemisia were recorded:Encarsia sophia (Girault and Dodd)comprising 93.7% of samples,Eretmocerus sp., 3.5%, Encarsia sp.(luteola group), 0.7%, Encarsia lutea(Masi), 1.2% and Encarsia mineoiViggiani, 0.8%. Potential whiteflypredators recorded were Cheilomenessulphurea (Olivier) and phytoseiid mites.Diseased whiteflies were not observed inthe field and entomopathogens thereforewere not collected. The diversity incassava viruses identified is reported inChapter 1.11 (this volume). Othercassava pests listed by farmers weregrasshoppers, reported by 15% offarmers, termites by 6.3%, andvertebrates by 2.5%.
Increased Socio-EconomicUnderstanding
Cassava farm sizes averaged0.38 hectares in the rain and transitionforests, 0.56 hectares in the wetsavannahs and 0.10 hectares in the drysavannahs. Both local and improvedvarieties were recorded in farmers’fields, although local varietiespredominated. The most widelyoccurring improved varieties were
Northern guineasavannah
Southern guineasavannah
Derived savannah
Humid forest
Niger
Benin
Cameroon
Nigeria
Figure 1. Areas surveyed for whitefly incidenceand cassava mosaic disease inNigeria.
Increased BiologicalUnderstanding
The insect vector of CMD, B. tabaci,occurred at all survey sites, while asecond whitefly species, Trialeurodesvaporariorum (Westwood), hithertounreported on cassava, was identifiedfrom nymphs collected from one locationin the country. No examples of B. afer

37
Nigeria
TMS 30572 (18.8%) and TMS 30555(2.5%). There was a marked differencein the incidence of improved varietiesbetween ecozones: they were cultivatedin 80% of sampled fields in thetransition forest but occurred in onlyabout 30% of fields in the rainforestand wet savannah and were notrecorded at all in the dry savannah.There was a direct negative correlationbetween the frequency of cultivation ofimproved varieties and CMD incidence.Farmers ranked cassava as their mostprofitable crop followed by maize, yam,sorghum (Sorghum bicolor [L.] Moench)and cowpea (Vigna unguiculata [L.]Walp.), in descending order. Of thecrops planted in association withcassava or in adjacent fields, cowpea isa known host plant of B. tabaci.Experimental data from other countriesin Africa, however, show that the B.tabaci biotype occurring on cassava ismore or less restricted to that crop(Burban et al., 1992; Legg et al., 1994).
Average CMD incidence rangedfrom 45% to 83% across ecozones, withcuttings providing by far the moreimportant source of infection (43% to83%) (Figure 2). The incidence ofwhitefly infection, transformed to allowfor the effect of multiple infection(Gregory, 1948), was low in each of theecozones. Among plants with disease,slight damage symptoms (score 2 on afive-point scale from no damage tosevere damage) predominated(Figure 3). Plant damage severity variedlittle by ecozone. Moderate damagesymptoms (score 3) were morepronounced in the rainforest (30% ofplants) and wet savannah (20% ofplants) than in the other ecozones,whilst serious (score 4) to severe(score 5) damage symptoms werenegligible (Figure 3). Plants were lessthan 4 months old at the time of thesurveys, a growth stage during whichstorage root formation and developmentis initiated and root yield is particularly
vulnerable to pest-induced losses. Veryfew farmers (1.2%) reported total yieldloss in the first planting season but18.8% of farmers attributed losses of atleast one quarter of their yield to thedisease. However, about 70% of thefarmers provided no yield lossestimates for the first or second crop.
Whitefly infection Cutting infection
Dry
sava
nn
ah
Rai
nfo
rest
Tra
nsi
tion
fore
st
Wet
sava
nn
ah
Per
cen
tage
of pla
nts
wit
h C
MD
sym
pto
ms
100
80
60
40
20
0
Figure 2. Cassava mosaic disease (CMD)incidence and source of infection inthe ecozones of Nigeria.
Per
cen
tage
of pla
nts
wit
hdiffe
ren
t C
MD
sev
erit
ies
Slightdamage
Moderatedamage
Seriousdamage
Rainforest
Wet savannah
100908070605040302010
0
Transition forest
Dry savannah
Figure 3. Cassava mosaic disease (CMD)damage severity in the ecozones ofNigeria.
Nearly all farmers reported thatCMD occurred every year, while slightlymore than half of them confirmed thatCMD was most severe during the earlygrowth stages of cassava. More thanthree-quarters of farmers (77.5%)reported that they had received notechnical information or assistance on

38
Whiteflies and Whitefly-borne Viruses in the Tropics
CMD or whiteflies. Among farmers whoattempted to manage the whitefly/disease problem, the main practicesused were selection of plantingmaterials, application of wood ash,roguing and proper weeding. Agronomicfeatures were more important thanpest/disease resistance as criteria forthe selection of planting material. Sincethe most frequently cited sources ofplanting material were neighbours’ andfarmers’ own fields (Figure 4), adoptionof appropriate strategies for selectingplanting material would be expected toreduce the incidence of CMDappreciably. About 53% of farmersrogued CMD affected plants, usuallywithin 3 months of planting. Mostfarmers (61%), however, rated roguingas partially effective, with only 4%considering it to be highly effective. Inthe choice of varieties, 25% of farmerscited “Agric” as the single mostcommonly planted variety and indicatedthat the choice was based onrecommendations that it was resistantto CMD. Other CMD-resistant varietiescommonly recorded were TMS 30572,TMS 30001 and TMS 30555. No farmerused pesticides against CMD and fewerthan 2% of farmers used pesticidesagainst whiteflies.
based on the relatively mild symptomsthat were observed in the survey. Thereis an obvious need, however, to obtainyield loss data for the commonly grownvarieties in order to make an accurateassessment of economic impact. In viewof the overwhelming importance ofcutting infection, as compared withvector-borne infection, in the spread ofCMD, research and extension efforts onassuring the health of planting materialshould be prioritized. The farmers’generally high level of awareness ofCMD and their willingness to roguediseased plants to prevent or slow itsspread could provide a good basis forestablishing participatory experimentsto help farmers multiply clean plantingmaterial of their preferred varieties.Action learning activities, involvingfarmers and extension agents, couldhelp improve their access to informationand their understanding of the causesand nature of the disease problem. Thiswill provide opportunities to promotecultural control options such asappropriate methods of selectingplanting material, sanitation androguing. Additionally, training ofnational program partners wouldstrengthen national capacity toinvestigate and promote the sustainablemanagement of whiteflies and CMD.
Acknowledgements
The assistance of Dr. M. A. Bob of theInternational Center of Insect Physiologyand Ecology in the identification ofwhitefly species and natural enemies isrecognized.
References
Burban, C.; Fishpool, L. D. C.; Fauquet,C.; Fargette, D.; Thouvenel, J. -C.1992. Host associated biotypeswithin West African populations ofthe whitefly Bemisia tabaci (Genn.)(Hom., Aleyrodidae). J. Appl.Entomol. 113:416-423.
Neighbour
Own farm
Market
Research
Extension
Relative frequency (%) of plantingmaterial source
Figure 4. Farmers’ sources of cassava plantingmaterial in Nigeria.
0 5 10 15 20 25 30 35 40
Conclusions
In Nigeria, whilst incidence of CMD ishigh, the economic impact of thedisease is believed to be only moderate,

39
Nigeria
FAO (Food and Agriculture Organizationof the United Nations). 1989.Production year book. Rome, IT.
Gregory, P. H. 1948. The multipleinfection transformation. Ann. Appl.Biol. 35:412-417.
Hahn, S. K.; Terry, E. R.; Leuschner, K.1980. Breeding cassava forresistance to cassava mosaicdisease. Euphytica 29:673-683.
Hahn, S. K.; Terry, E. R.; Leuschner, K.;Singh, T. P. 1981. Cassavaimprovement strategies forresistance to major economicdiseases and pests in Africa. In:Procs. Triennial Root CropsSymposium, 1980, InternationalDevelopment Research Centre(IDRC), Ottawa, CA. p. 279.
IITA (International Institute of TropicalAgriculture). 1997. EcologicallySustainable Cassava PlantProtection (ESCaPP) diagnosticsurvey results: Dry and wet seasonplant protection summaries.Ibadan, NG.
IITA (International Institute of TropicalAgriculture). 1998. EcologicallySustainable Cassava PlantProtection (ESCaPP) in SouthAmerica and Africa. Africa: Projectfindings and recommendations.Ibadan, NG.
James, B. D.; Legg, J. P.; Gbaguidi, J. B.;Annang, D.; Asiabaka, C. C.;Cudjoe, A. R.; Echendu, T. N. C.;Maroya, N. G.; Ntonifor, N.; Ogbe,F.; Salawu, R. A.; Tumanteh, J. A.1998. Understanding farm-levelepidemiology of cassava mosaicdisease. In: Abstracts, 7th TriennialSymposium of the InternationalSociety for Tropical Root Crops-Africa Branch (ISTRC-AB),11-17 October 1998, Cotonou, BJ.p. 122.
Jerath, M. L. 1967. A review of 50 yearsapplied entomology in Nigeria: Rootcrops. Entomol. Soc. Niger. p. 5-12.
Legg, J. P.; Gibson, R. W.; Otim-Nape, G.W. 1994. Genetic polymorphismamongst Ugandan populations ofBemisia tabaci (Gennadius)(Homoptera: Aleyrodidae), vector ofAfrican cassava mosaic geminivirus.Trop. Sci. 34:73-81.
Legg, J. P.; James, B. D.; Cudjoe, A.;Saizonou, S.; Gbaguidi, B.; Ogbe, F.;Ntonifor, N.; Ogwal, S.; Thresh, J.;Hughes, J. 1997. A regionalcollaborative approach to the study ofACMD epidemiology in sub-SaharanAfrica. Afr. Crop Sci. J. 3:1021-1033.
Nweke, F. I.; Lynam, J. K. 1997. Cassavain Africa. Afr. J. Root Tuber Crops2:10-13.
Ogbe, F. O.; Atiri, G. I.; Thottappilly, G.;Dixon, A. G. O.; Mignouna, H. D.;Quin, F. M. 1998. Evidence of doubleinfection and random occurrence ofcassava begomoviruses in sub-Saharan Africa. In: Procs. 7thTriennial Symposium of theInternational Society for TropicalRoot Crops-Africa Branch (ISTRC-AB), 11-17 October 1988, Cotonou,BJ. p. 524-529.
Sotomey, M.; James, B. D.; Gbaguidi, B.1995. Les espèces de Bemisia sur lemanioc en Afrique occidentale etcentrale. In: 11th Meeting andScientific Conference of the AfricanAssociation of Insect Scientists,6-11 August 1995, Yamoussoukro,CI. p. 4.
Swanson, M. M.; Harrison, B. D. 1994.Properties, relationships anddistribution of cassava mosaicgeminiviruses. Trop. Sci. 34:15-25.
Thresh, J. M.; Otim-Nape, G. W.; Legg, J.P.; Fargette, D. 1997. African cassavamosaic disease: The magnitude of theproblem? Afr. J. Root Tuber Crops2:13-19.
Yaninek, J. S.; James, B. D.; Bieler, P.1994. Ecologically SustainableCassava Plant Protection (ESCaPP): Amodel for environmentally soundpest management in Africa. Afr. CropSci. J. 2:553-562.

40
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 1.5
CameroonNelson Ntonifor*, Braima James**,
Brice Gbaguidi** and Ambe Tumanteh***
Introduction
Cassava mosaic begomovirusestransmitted by Bemisia tabaci andcausing cassava mosaic disease(CMD) are among the most importantvector-borne pathogens of crop plantsin sub-Saharan Africa (Geddes, 1990).Yield losses attributable to CMD areestimated at 28%-40% (Thresh et al.,1994). Fauquet and Fargette (1990)reported yield losses ranging from20% to 95% for particular varieties ofcassava (Manihot esculenta Crantz)under specific conditions. Theincidence and effects of CMD indifferent ecozones are influenced byenvironmental factors, the intensity ofcassava cultivation, the relativesusceptibility and sensitivity toinfection of cassava genotypes grown,and the virulence and abundance ofvirus and vector species (Fargette andThresh, 1994). Disease epidemiologymay be influenced also by theincidence of Bemisia tabaci(Gennadius), the principal vector ofCMD, and of B. afer (Priesner andHosny), a non-vector species that alsooccurs on cassava (Robertson, 1987;
Fishpool and Burban, 1994; Legg,1995), as well as by man through thetransport and replanting of infectedcassava cuttings. In Cameroon,Fondong et al. (1997), using infectionof initially CMD-free cuttings as anindicator, showed that the spread ofthe disease was more rapid in thelowland forest than in the mid-altitude forest and savannah areas. Itwas also observed that CMD spreadsfaster in monocultures of cassava orwhen cassava is intercropped withcowpea (Vigna unguiculata [L.] Walp.)than in cassava intercropped withmaize (Zea mays L.) (Fondong et al.,1997). In a regional CMDepidemiology trial covering Benin,Cameroon, Ghana and Nigeria,varietal differences in the rate of CMDinfection were observed amongimproved and local varieties (James etal., 1998).
Given that many factors influenceCMD epidemiology, a detailedunderstanding of the various cassavaagro-ecologies, the associated whiteflyspecies composition and thecultivation practices of the farmersmay be helpful in designing the mostappropriate CMD control strategy.Furthermore, since the disease isprevalent in almost all the majorcassava production areas of Africa,potential advantages are to be gainedin collecting such baseline data fromdifferent areas using a common
* Department of Biological Sciences,University of Buea, Cameroon.
** International Institute of TropicalAgriculture (IITA), Biological Control Centerfor Africa, Cotonou, Benin.
*** Institute of Agricultural Research forDevelopment, Ekona, Cameroon.

41
Cameroon
protocol, to enable valid comparisonsto be made between countries and thusfacilitate the search for effectivesolutions. This chapter reports thestatus of the CMD problem, the speciescomposition and relative abundance ofBemisia whitefly populations, and thecassava production practices in threemajor cassava agro-ecologies ofCameroon.
Surveys carried out in Novemberand December 1997, and later inMarch 1998, followed the methodspreviously agreed among projectpartners. Seventy fields were surveyedin three cassava growing regionschosen to represent the major agro-ecological regimes under which cassavais grown in Cameroon: South-WestProvince, representing rainforest(26 fields); Central/South Province,representing transition forest(28 fields); and North-West Province,representing wet savannah (16 fields)(Figure 1).
Increased BiologicalUnderstanding
Two whitefly species, B. tabaci andB. afer, were identified on cassava inCameroon. B. tabaci was the prevalentspecies, comprising at least 92% ofwhitefly adult collections in each of thezones surveyed. Whitefly abundanceaveraged 4.1 adults per cassava shoottip in the rainforest, 3.1 in the transitionforest and 2.4 in the wet savannah.Logarithm-transformed whitefly countswere significantly lower in the wetsavannah than in each of the otherecozones (t > 3.11; P < 0.05). Twoparasitoid species, Encarsia sophia(Girault and Dodd) and Eretmocerus sp.were hatched from Bemisia mummies;E. sophia predominated in thecollections.
Total CMD incidence averaged 44%in the rainforest, 72% in the transitionforest and 45% in the wet savannah.Incidence of stem cutting infectioncontributed far more to total CMDincidence than did whitefly infection(Figure 2) and was significantly higher inthe transition forest than in either of theother ecozones (t = > 3.09; P < 0.05). Apossible reason for the high incidence ofcutting infection observed in thetransition forest is that at least 90% offarmers in the region grow CMD-susceptible local cassava varieties whosetender leaves are consumed as leafyvegetables. The incidence of whiteflyinfection, transformed to allow for theeffect of multiple infection (Gregory,1948), was significantly lower in thetransition forest than in either of theother ecozones (t > 6.27; P < 0.05). In allecozones, most of the diseased plantshad a mean damage severity score of2 (“slight damage”, on an ascendingdamage severity scale of 1 to 5) and theproportion of plants with damageseverity score 4 was negligible; therewere no plants with damage score 5(Figure 3).
North-West
South-West
Nigeria
Eq, guinea
CentralAfrican
Republic
Cameroon
Gabon Congo
South-Central
Figure 1. Areas surveyed for whiteflyincidence and cassava mosaicdisease in Cameroon.

42
Whiteflies and Whitefly-borne Viruses in the Tropics
Increased Socio-EconomicUnderstanding
Cassava farmers were mostly women(87%). Sixty-six percent of farmersowned land whilst 20% rented land forfarming; the remaining 14% offered noresponse on the issue of land tenure.Farm size averaged 0.29 hectares(2.5 nearby fields) in South WestProvince, 0.6 hectares (2.4 nearbyfields) in Central/South Province and0.32 hectares (4.2 nearby fields) inNorth-West Province. Farmers grewcassava mostly on land that had beenworked for more than 5 years (60% of
sites). Fourteen percent of the farmswere on land that had been worked forbetween 2 and 5 years, and 26% onland that had been cultivated for lessthan 2 years. Only 21% of cassavafarmers practiced crop rotation.Nineteen percent of farmers rankedcassava as the most profitable crop and90% of farmers sold all or part of theircassava crop.
Farmers generally attached nosignificance to whiteflies and in theirlocal languages they simply referred tothem as “flies” or “white insects”. Incontrast, 51% of farmers recognizedCMD as a problem and in locallanguages referred to the diseasesymptoms variously as “crazy disease”,“curl”, “jelly cassava”, “leaf curl”,“leprosy”, kalle, khumbo, nome orpama. Use of these local names, whichdescribe the symptoms, will be helpfulin training farmers to improve theirunderstanding of the causes andnature of the disease.
No farmer reported total yieldlosses attributable to CMD; however,16% of farmers reported losses of 25%in the first season, 20% reported 50%losses and 10% reported 75% losses.These figures are surprising in view ofthe mild damage observed in thesurvey, which, based on experienceelsewhere, would not be expected tolead to significant yield loss. Mostfarmers indicated that climate affectedwhitefly/CMD problems and statedthat the problem was most severeduring the early growth stages ofcassava. This period corresponds toMarch-April during the first season andto August during the second growingseason in Cameroon. Even thoughmost farmers recognized CMD as aproblem that occurs every year, 61% ofthem had received no technicalinformation or assistance on theproblem.
Figure 3. Cassava mosaic disease (CMD)damage severity in the ecozones ofCameroon.
Per
cen
tage
of pla
nts
wit
hdiffe
ren
t C
MD
sev
erit
ies
Slightdamage
Moderatedamage
Seriousdamage
Transition forest
80706050403020100
Rainforest
Wet savannah
80706050403020100
Whitefly infection Cutting infection
Transitionforest
Wetsavannah
Per
cen
tage
of pla
nts
wit
h C
MD
sym
pto
ms
Figure 2. Cassava mosaic disease (CMD)incidence and source of infection inthe ecozones of Cameroon.
Rainforest

43
Cameroon
Farmers undertook variousmeasures to combat the whitefly/CMDproblem. The common practices wereroguing, choice of resistant varieties,selection of planting material and theapplication of wood ash or of fertilizers(Figure 4). Less than 2% of farmersused pesticides against whiteflies and/or CMD. Even though it might besupposed that the high incidence ofcutting infection observed in the surveywould make roguing an unattractivestrategy, it was nevertheless thepredominant practice.
CMD-resistant varieties. Farmer-ledrapid multiplication schemes toproduce CMD-resistant varieties maytherefore be relatively easy to promoteunder such circumstances by buildingupon previous technology transferexperiences and farmer trainingprogrammes in the country (Dahniyaet al., 1994; Akoroda, 1997; Bakia etal., 1999). In selecting plantingmaterial, the five most importantcriteria were yield (34.3% of farmers),followed by stem health (25.7% offarmers), absence of diseases (17.1%of farmers), stem size (5.7%) and stemmaturity (4.3%). Most farmersexpressed willingness to changeplanting dates for CMD control but30% of them indicated unwillingnessto alter planting date.
Conclusions
There was only limited evidence ofCMD being spread by its whiteflyvector in farmers’ fields in Cameroon.Infected planting material was themajor source of CMD and a majorextension effort will be required toencourage farmers to select disease-free and healthy planting materials. Ifeffective measures to improve crophealth are to be developed, theimportance of latent infection will haveto be investigated. The expressedwillingness of farmers to alter plantingdates and their conviction that CMDincidence varies with climaticconditions paves the way forparticipatory action research tooptimize times of planting in thevarious ecozones so as to reduce CMDincidence and damage. In suchstudies, it may be useful to monitorthe population dynamics of B. tabaci tosee whether its abundance varies atthe different planting times. Evaluationof parasitism by existing naturalenemies (E. sophia, for example) mayhelp show whether biological control
Roguing
Varieties
Selection
Fertilizers
Relative frequency (%) of practice
0 5 10 15 20 25 30 35
Wood ashPesticides
Figure 4. Cassava mosaic disease managementpractices used by farmers inCameroon.
In their choice of varieties, 86% ofthe farmers were aware of thedifferences in susceptibility to thedisease among cassava varieties.Among improved varietiesrecommended for their diseaseresistance, “Agric” (of InternationalInstitute of Tropical Agriculture, Nigeria[IITA] origin), chosen by 47% offarmers, was the most commonlyselected. The variety Pawpaw leaf wasthe single most frequently plantedCMD-resistant local variety (chosen by9% of farmers). Other CMD-resistantlocal varieties cited were Metta agric,Mfont and Ndongo.
Most farmers (96%) indicated thatin selecting planting materials theyintentionally chose parent plants withminimal or without CMD symptoms.This practice would inherently favour

44
Whiteflies and Whitefly-borne Viruses in the Tropics
contributes significantly to regulatingpopulations of the vector. Suchinformation will contribute tounderstanding the role of B. tabaci inthe epidemiology of CMD.
Even though the CMD damageobserved in the survey was slight,farmers provided high yield lossestimates. This suggests the need forfarmer participatory yield loss trials toquantify losses both of storage rootsand leaves, especially since the latterare widely consumed as leafyvegetables, but losses have been littlestudied. Farmer participatory varietalselection trials also need to beconducted on the local cassavavarieties (such as Pawpaw leaf ), whichfarmers report to be CMD-resistant,and comparisons made with cultivarswhose response to CMD is welldocumented. Such studies will providevaluable input into breedingprogrammes that aim to achievesustainable high yields in varietiespreferred by farmers and to increaseadoption of such varieties.
References
Akoroda, M. O. 1997. Cassava technologytransfer: Lessons from Cameroonand Nigeria. Afr. J. Root TuberCrops 2:257-260.
Bakia, B.; Ambe, J. T.; James, B. 1999.Technology transfer strategies: Thecase of sustainable cassava plantprotection in Cameroon. Afr. J. RootTuber Crops 3(2):23-27.
Dahniya, M. T.; Akoroda, M. O.; Alvarez,M. N.; Kaindaneh, P. M.; Ambe-Tumanteh, J.; Okeke, J. E.; Jalloh,A. 1994. Development anddissemination of appropriate rootcrops packages to farmers in Africa.In: Ofori, F.; Hahn, S. K. (eds.).Tropical root crops in a developingeconomy. Procs. Ninth Symposiumof the International Society forTropical Root Crops (ISTRC), 20-26October 1991, Accra, GH. p. 2-9.
Fargette, D.; Thresh, J. M. 1994. Theecology of African cassava mosaicgeminivirus. In: Blakeman, J.P.;Williamson, B. (eds.). Ecology ofplant pathogens. CAB International,Wallingford, GB. p. 269-282.
Fauquet, C.; Fargette, D. 1990. Africancassava mosaic virus: Etiology,epidemiology and control. Plant Dis.74:404-411.
Fishpool, L. D. C.; Burban, C. 1994.Bemisia tabaci: The whitefly vector ofAfrican cassava mosaic geminivirus.Trop. Sci. 34:55-72.
Fondong, V. N.; Adipala, A.; Thresh, J. M.1997. Spread of African cassavageminivirus at four agroecologicallocations of Cameroon. Afr. CropSci. J. 3:1035-1049.
Geddes, A. M. 1990. The relativeimportance of crop pests in sub-Saharan Africa. Bulletin no. 36,Natural Resources Institute (NRI),Chatham, GB.
Gregory, P. H. 1948. The multipleinfection transformation. Ann. Appl.Biol. 35:412-417.
James, B. D.; Legg, J. P.; Gbaguidi, J. B.;Annang, D.; Asiabaka, C. C.;Cudjoe, A. R.; Echendu, T. N. C.;Maroya, N. G.; Ntonifor, N.; Ogbe,F.; Salawu, R. A.; Tumanteh J. A.1988. Understanding farm-levelepidemiology of cassava mosaicdisease. In: Abstracts, 7th TriennialSymposium of the InternationalSociety for Tropical Root Crops-Africa Branch (ISTRC-AB), 11-17October 1988, Cotonou, BJ. p. 122.
Legg, J. P. 1995. The ecology of Bemisiatabaci (Gennadius) (Homoptera:Aleyrodidae), vector of Africancassava mosaic geminivirus inUganda. Ph.D. thesis, University ofReading, GB.

45
Cameroon
Robertson, I. A. D. 1987. The role ofBemisia tabaci (Gennadius) in theepidemiology of ACMV in EastAfrica: Biology, population dynamicsand interaction with cassavavarieties. In: African cassava mosaicdisease and its control, Procs.International Seminar, 4-8 May1987, Yamoussoukro, CI. CentreTechnique de Coopération Agricoleet Rurale, Wageningen, NL, andInstitut Français de RechercheScientifique pour le Développementen Coopération, Paris, FR. p. 57-63.
Thresh, J. M.; Fishpool, L. D. C.; Otim-Nape, G. W.; Fargette, D. 1994.African cassava mosaic virusdisease: An under-estimated andunsolved problem. Trop. Sci.34:3-14.

46
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 1.6
UgandaPeter Sseruwagi*, James Legg** and
William Otim-Nape***
Introduction
Cassava mosaic disease (CMD) is themost important viral disease of cassava(Manihot esculenta Crantz) (Otim-Nape,1993) in Uganda. It is caused bybegomoviruses transmitted by thewhitefly vector, Bemisia tabaci(Gennadius) and spread throughvirus-infected planting material(Harrison, 1987).
The first recording of CMD inUganda was in 1928 (Martin, 1928).Severe epidemics from 1933 to 1944were successfully controlled byplanting resistant cassava varieties andthrough phytosanitation measuresenforced by local government statute(Jameson, 1964). The disease then wasconsidered a relatively minor problemuntil the late 1980s, when severeepidemics were again reported innorthern Luwero District (Otim-Nape,1988). By 1990, CMD had severelyaffected other cassava-growing districtsof northern and eastern Uganda (Otim-Nape et al., 1997). In 1993 and 1994,the widespread crop losses associated
with the CMD epidemic, coupled withdrought, led to food shortages andlocalized famine (Otim-Nape et al.,1997). Between 1994 and 1998, theCMD epidemic continued to expand,affecting much of southern Uganda.Significant progress has been made incontrolling the disease, particularly innorth-eastern parts of the country,which were some of the first to beaffected (Thresh et al., 1994) but theepidemic continues to spread insouthern and western districts. Theprincipal control method has been thedeployment of resistant varieties.
Sweetpotato virus disease (SPVD) iscorrespondingly the most serious viraldisease of sweetpotato (Ipomoea batatas[L.] Lam.) in Uganda (Bashaasha et al.,1995), although its distribution andimpact on sweetpotato production havescarcely been studied until recently.Aritua et al. (1998) provide informationon the disease for only two of thecountry’s (at that time) 45 districts,Mpigi and Soroti (Figure 1). SPVDresults from co-infection of Sweetpotatochlorotic stunt virus, transmitted byB. tabaci and Sweetpotato featherymottle virus, transmitted by the aphidMyzus persicae (Sulzer) (Gibson et al.,1998).
In order to enhance understandingof whiteflies and whitefly-transmittedviruses on cassava and sweetpotato inUganda, surveys were conducted
* International Institute of TropicalAgriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda.
** IITA-ESARC, Kampala, Uganda, and NaturalResources Institute, University ofGreenwich, Chatham Maritime, Kent, UK.
*** Namulonge Agricultural and AnimalProduction Research Institute (NAARI),Kampala, Uganda.

47
Uganda
between November 1997 and January1998. It was anticipated that the datacollected would be useful in helpingresearchers in Uganda monitor thedevelopment of the CMD epidemic,target control interventions moreeffectively and, in the longer term,develop sound integrated pestmanagement (IPM) strategies forcassava and sweetpotato.
Increased BiologicalUnderstanding
Whitefly species and abundance
The survey identified two whiteflyspecies, B. tabaci and B. afer (Priesnerand Hosny). Of the 261 samplesidentified on cassava, 95.8% wereB. tabaci and 4.2% B. afer. OnlyB. tabaci was recorded from sweetpotato.
Whitefly populations on cassava(counted on the top five leaves for asingle shoot of each of 30 plants persampled field) were highest in the south-western districts of Masaka and Rakaiand lowest in Masindi (the north-western limit of the survey area) andTororo (on Uganda’s eastern border withKenya), with mean abundance rangingfrom 0.7 to 13.1 per top five leaves(Table 1) (see also Figure 2 in Chapter1.14, this volume). The pattern ofwhitefly populations on sweetpotato(counted by disturbing leaves andmaking 10 serial counts of 1 min each ofwhitefly adults observed) was similar tothat on cassava: abundance wasgreatest in the central districts ofMukono and Mpigi and lowest inMasindi and Apac. Mean adult whiteflyabundance ranged from 0.9 to 14.8 perminute count. In similar studies, Arituaet al. (1998) found greater whiteflyabundance on sweetpotato in thesouthern and central zones, which arecharacterized by more evenly distributedrainfall than the northern and easternzones.
The data indicate a higher whiteflypopulation on both crops in southernand south-western Uganda than in thenorthern and eastern districts of thecountry. Assessing the significance ofthese differences is difficult based onrecords collected on a single occasion,during what appears to be thecontinuing spread of an epidemic.Numerous factors such as physical
Figure 1. Cassava- and sweetpotato-growingareas surveyed for whitefly incidencein Uganda.
LakeVictoria
Uganda
LiraApac
Masindi
Luwero
Mubende
Masaka
Rakai
Mpigi
Mukono
IgangaTororo
Rallisa
Surveys were carried out in12 districts of Uganda (Figure 1) bymulti-disciplinary teams from theNational Agricultural ResearchOrganisation and the InternationalInstitute of Tropical Agriculture (IITA).Districts were chosen to representmajor cassava and sweetpotatoproducing areas. Farmers’ fields ofcassava 3-5 months after planting andof sweetpotato 3 months after plantingwere selected at regular intervals alongmain roads traversing each district.Data were collected according to thestandardized survey methods, exceptthat the number of fields variedbetween the districts because thesurvey areas were selected on thebasis of agro-ecological differencesrather than administrative boundaries(Table 1).

48
Whiteflies and Whitefly-borne Viruses in the Tropics
Table
1.
Inci
den
ce (%
) of
cass
ava
mos
aic
dis
ease
(C
MD
) an
d s
wee
tpot
ato
vir
us
dis
ease
(S
PV
D), d
isea
se s
ever
ity
an
d w
hit
efly
abu
ndan
ce o
n c
ass
ava
an
dsw
eetp
otato
in
sel
ecte
d d
istr
icts
of U
gan
da, su
rvey
ed d
uri
ng
Nov
ember
an
d D
ecem
ber
1997.
Dis
tric
tN
o.C
ass
ava
aS
wee
tpot
atoa
fiel
ds
Wh
itef
lyC
MD
Wh
itef
lyS
PV
Dco
un
tsco
un
tsW
hit
efly
Cu
ttin
gTot
alS
ever
ity
Inci
den
ceS
ever
ity
infe
ctio
nin
fect
ion
inci
den
ce(1
-5 s
cale
)(1
-5 s
cale
)
Mas
indi
7
0.7
11 (19)
34
45
3.0
0.9
3
2.0
Apac
6
1.4
22 (1
40)
71
93
3.0
1.9
9
2.3
Lir
a7
1.4
11 (89)
81
92
3.0
1.4
3
2.3
Igan
ga6
1.6
20 (80)
64
84
2.8
11.2
16
2.1
Tor
oro
7
0.9
16 (73)
69
85
3.6
1.5
15
2.8
Pal
lisa
7
3.0
21 (28)
15
36
3.0
3.4
6
2.6
Mu
kon
o7
3.7
13 (47)
64
77
3.0
14.8
10
2.4
Mpig
i7
4.6
17 (59)
61
78
2.8
14.7
29
2.7
Lu
wer
o6
3.0
12 (56)
71
83
2.6
7.8
7
2.2
Mu
ben
de
7
3.4
24 (29)
7
31
2.7
3.8
9
2.9
Masa
ka
613.1
50 (91)
17
67
3.0
3.2
14
2.7
Rakai
711.1
44 (60)
3
47
2.7
2.0
8
2.7
Ave
rage
3.9
22 (65)
46
68
2.9
5.5
11
2.5
a.
Fig
ure
s are
mea
ns
for
each
dis
tric
t. W
hit
efly
cou
nts
, w
hit
efly
abu
ndan
ce o
n c
ass
ava
(n
um
ber
of w
hit
eflies
per
top
fiv
e le
ave
s) a
nd o
n s
wee
tpot
ato
(per
min
ute
cou
nt)
; w
hit
efly
in
fect
ion
, figu
res
in p
are
nth
eses
tra
nsf
orm
ed t
o m
ult
iple
in
fect
ion
un
its
to a
llow
for
mu
ltip
le in
fect
ion
(G
rego
ry, P. H
. 1948. Th
e m
ult
iple
infe
ctio
n t
ran
sfor
mati
on. A
nn
. A
ppl. B
iol. 3
5:4
12-4
17); s
ever
ity
of d
isea
se is
mea
sure
d o
n a
n a
scen
din
g 1-5
sca
le, fr
om low
to
seve
re.

49
Uganda
environment, host plant healthcondition and natural enemies havebeen shown to influence whiteflypopulations on cassava (Legg, 1995).Nonetheless, this general geographicaltrend in abundance does parallel thecurrent trend in disease severity.
Disease incidence andsymptom severity
CMD was present in all cassava fieldssampled and overall incidence (theaverage proportion of affected plants)throughout the survey was 68% (seealso Figure 2 in Chapter 1.14, thisvolume). Disease incidence was highestin the northern districts of Apac andLira and lowest in Mubende. Symptomswere moderate (2.6 to 3.0 on anascending scale of 1 to 5) with theexception of Tororo District, where thedisease was severe (Table 1).
The pattern of CMD incidence inUganda indicates a north-to-southprogression. In areas of epidemicexpansion in the south (Masaka andRakai Districts) the main source ofinfection is whitefly transmission,whereas in post-epidemic areas to thenorth (Masindi, Lira and Apac Districts)whitefly transmission is less evidentand diseased cuttings are the mainsource of infection (Table 1). Therelatively low CMD incidence recordedin Pallisa District is because of theNational Cassava Programme’s recentintroduction of CMD-resistant varieties,Nase 1 (TMS 60142), Nase 2(TMS 30337) and Nase 3 (TMS 30572),originating from the IITA breedingprogramme. Similarly, diseaseincidence and infection were relativelylow for the districts neighbouringPallisa-Kumi, immediately to the north,and Soroti, to the south.
SPVD occurred in 60 of the80 sweetpotato fields sampled.Generally, SPVD incidence was greater
in southern areas along the shoreline ofLake Victoria, with the highest incidencein Mpigi District on the northern shoreof the lake, and the lowest in Masindiand Lira, further to the north. Overall,SPVD symptom severity was moderatein all the districts surveyed.
Natural enemy species anddistribution
Whitefly natural enemies collected in thesurvey included predators andparasitoids. Few predators werecollected. This was probably due to thelimited time available during the surveyfor collecting samples, the relatively lowpopulations of whiteflies on cassava andsweetpotato, and the difficulty ofobserving arthropods actually feeding onwhitefly immature or adult stages.
The predator Conwentzia africanaMeinander was recorded from16 locations in three of the four targetareas. Three species of whiteflyparasitoids were identified: Encarsiasophia (Girault and Dodd), Encarsia sp.(luteola group) and Eretmocerus sp.E. sophia was the most widespread andnumerous, occurring through most ofthe surveyed area (Table 2). Eretmocerussp. occurred principally in south-western Uganda. Encarsia sp. (luteolagroup) was only identified from easternUganda. The absence of records ofparasitoids from some of the districtssurveyed, particularly those in thenorth, is thought to be largely becauseof the relatively low abundance ofwhiteflies recorded in these locations.Seasonal and environmental factorsclearly play an important role in theabundance of whiteflies and theirparasitoids, and in order to collect morecomprehensive data, the surveys wouldhave to be repeated, perhaps at3-monthly intervals throughout theyear. Little is known about the role ofnatural enemies in the populationdynamics of B. tabaci on cassava andsweetpotato.

50
Whiteflies and Whitefly-borne Viruses in the Tropics
Increased Socio-EconomicUnderstanding
Farmers’ assessment ofwhitefly-related problems
Only a small proportion of cassava andsweetpotato farmers (22%) recognizedwhiteflies. In contrast, the diseasesCMD and SPVD were more widelyrecognized (CMD by 100% of cassavafarmers and SPVD by 48% ofsweetpotato farmers) and had specificlocal names in most of the locationswhere they were prevalent. Bothdiseases were commonly referred towith words describing the mottling ordeformation of the leaves induced bythe disease, and a number of nameswere common for both crops. Virtuallyall cassava farmers recognized CMD asa production constraint, whereas forSPVD only 35% of producersconsidered it a problem. Most cassavafarmers (74%) considered that CMDwas becoming more severe.Sweetpotato producers were similarlypessimistic about the outlook for
SPVD, with 65% of the producers whorecognized SPVD as a problemconsidering that it was becoming moresevere. Farmers expressed no clearopinion as to the time of year at whichwhitefly/disease problems areespecially severe. The picture of thespread of the CMD epidemic providedby farmers’ first record of severe CMDin their fields, indicating a north-to-south progression, is in closeagreement with scientific records of theexpansion of the severe CMD epidemic(Otim-Nape et al., 1997; Legg andOgwal, 1998).
Managing whiteflies andwhitefly-transmitted viruses
A common response amongst farmersto the CMD problem has been toabandon production of the cropaltogether. Whilst cassava farmersassessed losses to CMD at more than75% of production, sweetpotatofarmers estimated losses to SPVD to beless than 25%. Farmers attempting tocontrol CMD and SPVD mainly usedroguing, while some applied wood ashon the infected plants. Only 6.5% offarmers reported having receivedinformation on control practicesthrough extension and/or researchchannels. Selection of planting materialfree of disease symptoms was the mostwidely used disease control practice forboth cassava and sweetpotato, and thesecond most widely reported wasroguing of plants that developedsymptoms. However, most farmersreported the two methods to be onlypartially effective in controlling thediseases.
Thirteen out of 19 farmers growingresistant varieties of cassava and zeroout of 11 farmers growing resistantsweetpotato varieties cited resistanceas a control method. However, onlyabout 24% of cassava farmers growingresistant varieties found theirperformance to be superior to that of
Table 2. Occurrence of parasitoids rearedfrom mummies collected fromcassava at each of the sampling sitesin Uganda.
District Parasitoid occurrence per sampling sitea
Encarsia Eretmocerus Encarsia sp.sophia sp. (luteola group)
Masindi - - -
Apac - - -
Lira - - -
Iganga 3/6 1/6 1/6
Tororo 2/7 - -
Pallisa 2/7 - -
Mukono 6/7 1/7 -
Mpigi 4/7 3/7 -
Luwero - - -
Mubende 2/7 - -
Masaka 1/6 3/6 -
Rakai 3/7 2/7 -
a. Dashes indicate the absence of any rearedparasitoids.

51
Uganda
local varieties. Assessments of thebenefits of disease-resistantsweetpotato varieties were more mixed,with some farmers reporting nodifference in performance betweenresistant and local varieties. About20% of both cassava and sweetpotatofarmers attributed differences in thedisease response of the differentvarieties to some kind of resistance toinfection, inherent in the variety. Onthe other hand, 28% of cassavaproducers and 18% of sweetpotatoproducers had no idea why suchdifferences should occur. Very fewfarmers reported using chemicalpesticides to control either CMD orSPVD. Of these farmers, almost allused dimethoate, an organophosphateinsecticide, so their intention waspresumably to control the whiteflyvector. Most of these farmers usedpesticides on the recommendation ofneighbours or relatives, sprayed whensymptoms were seen, and did so onone to three occasions during theproduction cycle. However, manyreported that the approach wasineffective.
Whitefly-related cropproduction costs in affectedareas
Since farming families produce mostcassava and sweetpotato for homeconsumption rather than sale, it wasdifficult to assess the value of lostproduction attributable to CMD andSPVD and costs incurred in trying tomanage these diseases. As mentionedabove, the most widely practicedmanagement tactics were selection ofdisease-free planting material androguing, both of which involveinvestment of additional labour.Because labour is usually provided bymembers of farming households, ratherthan hired, this represents a time-related opportunity cost rather than adirect financial one. More focusedfollow-up studies are required to
provide an understanding of theeconomic costs and benefits ofmanaging these diseases.
Conclusions
The results of the survey indicate thatfarmers’ approaches to managing thewhitefly-borne virus diseases, CMD andSPVD, are generally ad hoc and basedon only a partial understanding of theproblem. Since researchers alreadyhave a considerable body of knowledgerelating to these problems that is notbeing applied at farm level, thissuggests that information flow betweenfarmers and researchers is weak.However, the absence of a clearlydefined and well-articulated IPMapproach to managing CMD and SPVDcould be a contributory factor.Researchers believe that such anapproach would combine the use ofboth local and improved resistantvarieties with the selection of cleanplanting material and subsequentroguing of diseased plants—planthealth methods that are already widelypracticed.
The development andenhancement of channels ofinformation flow from research to farmlevel is clearly critical to the successfuladoption of IPM of whiteflies andwhitefly-transmitted viruses. A numberof initiatives have recently beenimplemented in Uganda to strengthenboth the extension system and linkagesbetween research and extension and tofoster a participatory approach toworking with farmers to developappropriate and sustainable IPMstrategies. It is hoped that these willimprove the efficiency of the two-wayflow of information between researchersand farmers in the years ahead,facilitating the more widespreadadoption of IPM strategies as theybecome available.

52
Whiteflies and Whitefly-borne Viruses in the Tropics
Future work should focus mainlyon managing CMD and SPVD, throughmultiplication and distribution of theexisting disease-resistant varieties, andon developing, evaluating, multiplyingand distributing new disease-resistantmaterials. Since most farmers reportedthe planting of clean cuttings androguing of diseased plants to beineffective, there is a need to find outunder what conditions these measurescould be both effective and adoptableby farmers. Future work should alsoconsider the possibility of community-based phytosanitation, which offers theprospect of widespread reductions inCMD or SPVD inoculum pressure andthereby reduced rates of spread. Toolittle is currently known about theactivity of parasitoids or other naturalenemies of B. tabaci on either cassavaor sweetpotato to offer the prospect oftheir immediate use within an IPMstrategy. However, both introductionsof exotic parasitoids and augmentationthrough habitat management havebeen used successfully in the UnitedStates to improve management ofB. tabaci (Goolsby et al., 2000). It maybe appropriate therefore to exploresimilar approaches to using parasitoidssuch as E. sophia for the managementof B. tabaci as vector of CMD andSPVD, although this is always likely tobe a measure of secondary importanceto the major control approaches ofresistant varieties and phytosanitation.
Acknowledgements
The technical support of Dr. NicoleSmit of the Centro Internacional de laPapa (CIP), Dr. Benson Odongo,Mr. Valentine Aritua, Mr. Titus Alicai,Mr. William Sserubombwe andMr. Solomon Ogwal of the NationalAgricultural Research Organisation ofUganda and Mr. Geoffrey Okao-Okujaof IITA, is acknowledged. The authors
also acknowledge Dr. Mohammed AliBob of the International Center ofInsect Physiology and Ecology for hisidentification of whitefly species andnatural enemies.
References
Aritua, V.; Adipala, E.; Carey, E. E.;Gibson, R. W. 1998. The incidenceof sweetpotato virus disease andvirus resistance of sweetpotatogrown in Uganda. Ann. Appl. Biol.132:399-411.
Bashaasha, B.; Mwanga, R .O. M.; Ocittip’Obwoya, C. O.; Ewell, P. T. 1995.Sweetpotato in the farming and foodsystems of Uganda: A farm surveyreport. Centro Internacional de laPapa (CIP), sub-Saharan AfricaRegion, Nairobi, KE/NationalAgricultural Research Organisation(NARO), Kampala, UG. 63 p.
Gibson, R. W.; Mpembe, I.; Alicai, T.;Carey, E. E.; Mwanga, R. O. M.;Seal, S. E.; Vetten, H. J. 1998.Symptoms, aetiology and serologicalanalysis of sweetpotato virusdisease in Uganda. Plant Pathol.47:95-102.
Goolsby, J. A.; Ciomperlik, M. A.; Kirk, A.A.; Jones, W. A.; Legaspi, B. C.;Legaspi, J. C.; Ruiz, R. A.; Vacek, D.C.; Wendel, L. E. 2000. Predictiveand empirical evaluation forparasitoids of Bemisia tabaci(Biotype “B”), based onmorphological and molecularsystematics. In: Austin, A.; Dowton,M. (eds.). Hymenoptera: Evolution,biodiversity, and biological control.4th International HymenopteristsConference, 1999, Canberra.Commonwealth Scientific andIndustrial Research Organisation(CSIRO), Collingwood, Victoria, AU.p. 347-358.
Gregory, P. H. 1948. The multipleinfection transformation. Ann. Appl.Biol. 35:412-417.

53
Uganda
Harrison, B. D. 1987. Properties andgeographical variation ofgeminivirus isolates from mosaic-affected cassava. In: African cassavamosaic disease and its control.Procs. International Seminar,4-8 May 1987, Yamoussoukro, CI.Centre Technique de CoopérationAgricole et Rurale, Wageningen, NL,and Institut Français de RechercheScientifique pour le Développementen Coopération, Paris, FR. p. 270.
Jameson, J. D. 1964. Cassava mosaicdisease in Uganda. East Afr. Agric.For. J. 29:208-213.
Legg, J. P. 1995. The ecology of Bemisiatabaci (Gennadius) (Homoptera:Aleyrodidae), vector of Africancassava mosaic geminivirus inUganda. Ph.D. thesis, University ofReading, GB.
Legg, J. P.; Ogwal, S. 1998. Changes inthe incidence of African cassavamosaic geminivirus and theabundance of its whitefly vectoralong south-north transects inUganda. J. Appl. Entomol. 122:169-178.
Martin, E. F. 1928. Report of themycologist. In: Annual Report of theDepartment of Agriculture.Government Printer, Entebbe, UG.p. 31.
Otim-Nape, G. W. 1988. Cassavasituation in Luwero district. Amission report by the Uganda RootCrops Programme. NamulongeResearch Station, Kampala, UG.
Otim-Nape, G. W. 1993. Epidemiology ofthe African cassava mosaicgeminivirus disease (ACMD) inUganda. Ph.D. thesis, University ofReading, GB.
Otim-Nape, G. W.; Bua, A.; Thresh, J. M.;Baguma, Y.; Ogwal, S.; Semakula,G. N.; Acola, G.; Byabakama, B.;Martin, A. 1997. Cassava mosaicvirus disease in Uganda: Thecurrent pandemic and approachesto control. Natural ResourcesInstitute (NRI), Chatham, GB. 65 p.
Thresh, J. M.; Fishpool, L. D. C.; Otim-Nape, G. W.; Fargette, D. 1994.African cassava mosaic virusdisease: An under-estimated andunsolved problem. Trop. Sci.34:3-14.

54
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 1.7
KenyaJoseph Kamau*, Peter Sseruwagi** and
Valente Aritua***
Introduction
Cassava mosaic disease (CMD) iscaused by cassava mosaicbegomoviruses (CMBs), which aretransmitted by the whitefly Bemisiatabaci (Gennadius). Further spread mayoccur through farmers distributing andplanting virus-infected cuttings(Harrison, 1987). The disease causeshigher production losses than any othervirus disease of cassava in Kenya (Bock,1994b). Sweetpotato virus disease(SPVD) results from co-infection ofsweetpotato (Ipomoea batatas [L.] Lam.)by two distinct viruses: Sweetpotatochlorotic stunt virus, transmitted by B.tabaci, and Sweetpotato feathery mottlevirus, transmitted by the aphid, Myzuspersicae (Sulzer) (Gibson et al., 1998).Although SPVD is the most prevalentviral disease of sweetpotato in farmers’fields in Kenya (Carey et al., 1998), itseffects on growth and yield have yet tobe quantified.
The early history of CMD in Kenyais obscure but work on cassava underthe East African breeding programme inthe mid-1950s is described by Doughty
(1958) and resulted in the release ofCMD-resistant hybrids to researchstations in the country. However,farmers did not replace their localtraditional Kenyan cultivars of cassava(Manihot esculenta Crantz) with thisimproved germplasm, except for46106/27 “Kaleso”, which becamewidely grown in Coast Province (Bock,1994b).
Research on CMD and its whiteflyvector only became important in Kenyaduring the 1970s, when a major plantvirology project was initiated. The workof this project initially emphasized theisolation of the putative Cassava mosaicvirus (Bock, 1975), which was latershown to cause CMD (Bock and Woods,1983). Subsequent epidemiologicalwork by Bock (1983) noted that theincidence of CMD was generally high incoastal and western Kenya, where itexceeded 80% in some districts andapproached 100% on individual farmsbut the spread of disease into initiallyCMD-free plantings was generally slow.Additional studies associated with thisproject were the description of thepopulation dynamics of B. tabaci (Seif,1982; Robertson, 1987), assessments ofyield loss (Seif, 1981) and thedevelopment of control methods (Bock,1994a). During the 1990s, a series ofsurveys conducted to reassess theincidence of the disease in the countryindicated increased spread andincidence of CMD in farmers’ fields.
* Kenya Agricultural Research Institute(KARI), National Dryland Farm ResearchCenter, Katumani, Kenya.
** International Institute of TropicalAgriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda.
*** Makerere University, Kampala, Uganda.

55
Kenya
Recent epidemiological studies onCMD in the region have providedevidence of the spread of severe CMDfrom Uganda to western Kenya (Legg,1999). This is associated with thespread of a novel and particularlyvirulent CMB (Harrison et al., 1997;Legg, 1999). Major losses have beenexperienced already in WesternProvince, and districts of NyanzaProvince are now threatened. Effortshave been made to control the diseaseby introducing resistant material intowestern Kenya from the breedingprogram of the International Instituteof Tropical Agriculture-Eastern andSouthern Africa Regional Center (IIITA-ESARC) in Namulonge, Uganda.Continuing work by the cassavaprogram of the Kenya AgriculturalResearch Institute (KARI) emphasizesthe evaluation and multiplication of theinitial stock of resistant material, incollaboration with internationalresearch institutions and networkssuch as the East Africa Root CropsResearch Network (EARRNET) (Legg etal., 1999).
Research on SPVD began duringthe 1950s, when Sheffield (1957)reported the presence of a severesweetpotato mosaic disease onsweetpotato fields in East Africa. Morerecently, surveys have been carried outto assess the occurrence andimportance of sweetpotato viruses inKenya (Carey et al., 1998) and theCentro Internacional de la Papa (CIP)now has a major germplasmintroduction and development programbased in Kenya. Whilst cultivarresistance has been identified as theprincipal strategy for managing SPVD,little detailed research has been doneon yield loss or the epidemiology of thedisease.
A study was conducted in mid-1998 to identify whiteflies and whitefly-transmitted viruses prevalent in Kenya
and to assess producers’ knowledge ofwhiteflies and the diseases theytransmit on cassava and sweetpotato.Three “target areas” representing themajor cassava and sweetpotato growingareas in the country were selected:Coast Province, Western Province andNyanza Province (Figure 1). Withinthese areas, 3 to 5-month-old cassavafields and 3-month-old sweetpotatofields were randomly selected atregular intervals along main roads.
Coast
Kenya
Western
Nyanza
Tanzania
Uganda
EthiopiaSudan
Som
alia
Figure 1. Cassava- and sweetpotato-growingareas surveyed for whitefly incidencein Kenya.
Increased BiologicalUnderstanding
Whitefly species andabundance
Whitefly nymph samples were obtainedfrom most but not all surveyed sitesand, for each sample, from one to threenymphs were identified to species level.Two whitefly species, B. tabaci andBemisia afer (Priesner and Hosny), wereidentified on both cassava andsweetpotato. Seventy whitefly nymphsamples were identified from cassava;

56
Whiteflies and Whitefly-borne Viruses in the Tropics
of which 58 were B. tabaci and 12 wereB. afer. Only 35 whitefly samples wereidentified from sweetpotato(attributable to the lower populationson that crop), comprising 31 B. tabaciand four B. afer. The two speciesoccurred throughout the three targetareas.
Adult whitefly populations wererelatively low on both cassava andsweetpotato (Table 1). Whiteflies oncassava were more numerous in CoastProvince, while whiteflies onsweetpotato were more abundant inNyanza Province. Mean adult whiteflynumbers ranged from 0.2 to 2.9 pertop five leaves on cassava and 0.5 to4.4 per minute count on sweetpotato inthe surveyed regions (see also Figure 2,Chapter 1.14, this volume).
Disease incidence andsymptom severity
Mean CMD incidence in the surveyedregions was 51.3% and incidence washighest in Western Province and lowestin Nyanza (Table 1). Diseased cuttingsprovided the main source of infectionin all three target areas. CMD symptomseverity was mild in the Coast andNyanza Provinces and more severe inWestern Province. SPVD incidenceranged between 3% and 53% in
farmers’ fields. The disease was moreprevalent in Western and NyanzaProvinces than in Coast Province.
Whitefly parasitoids
The survey identified two species ofaphelenid parasitoids; 15 individualparasitoids were identified, comprising13 Encarsia sophia (Girault and Dodd)and two Eretmocerus sp. Little isknown, however, about the role ofnatural enemies in the populationdynamics of whiteflies in Kenya.
Increased Socio-EconomicUnderstanding
Farmers’ assessment ofwhitefly-related problems
About half of the producers surveyed(52% of cassava farmers and 46% ofsweetpotato farmers) were able torecognize whiteflies, although thenames they used were non-specific. Ofthe farmers who recognized whiteflieson either crop, 61% considered them aproduction problem.
Most cassava farmers (88%)recognized CMD as a disease of theircrop, whilst only 58% of sweetpotatofarmers recognized SPVD. The
Table 1. Incidence (%) of cassava mosaic disease (CMD) and sweetpotato virus disease (SPVD),disease severity and whitefly abundance on cassava and sweetpotato in three provinces ofKenya, surveyed during May and June 1998.
Province Cassavaa Sweetpotatoa
No. Whitefly CMD No. Whitefly SPVDfields counts fields counts incidence
Whitefly Cutting Total Severityinfection infection incidence
Coast 17 2.9 19 (35.9) 37 56 2.3 15 0.5 0.4
Western 15 0.2 14 (65.9) 71 85 2.9 18 2.5 6.9
Nyanza 18 0.6 2 (2.3) 11 13 2.2 17 4.4 6.7
Average - 1.2 11.7 (21.6) 39.7 51.3 2.5 - 2.5 4.7
a. Figures are means for each province. Whitefly counts, whitefly abundance on cassava (number ofwhiteflies per top five leaves) and on sweetpotato (per minute count); whitefly infection, figures inparentheses transformed to multiple infection units to allow for multiple infection (Gregory, P. H.1948. The multiple infection transformation. Ann. Appl. Biol. 35:412-417); severity of diseasemeasured on an ascending 1-5 scale, from low to severe.

57
Kenya
respective diseases were recognized asa production constraint by 68% ofcassava farmers and 44% ofsweetpotato farmers. Most cassavafarmers (82%) considered CMD tooccur every year, whilst only 52% ofsweetpotato farmers believed this to bethe case. Farmers had different viewsregarding the trend of symptomseverity for CMD and SPVD. Fifty-fourpercent of cassava farmers consideredthat CMD severity was increasing,whilst only 40% of sweetpotato farmersconsidered that SPVD was becomingmore important. In both cases, amajority of cassava (72%) andsweetpotato (62%) farmers believedthat climate influences disease severity.The two diseases were thought to bemore severe during periods of lowrainfall by 70% of cassava farmers and44% of sweetpotato farmers. This wasconsistent with farmers’ views that thetwo diseases were more severe duringperiods of moderate temperature.
Yield loss estimates due to CMDvaried between regions. In WesternProvince, 21.4% of farmers estimatedlosses at >75%. This is understandablein view of the impact that the CMDpandemic has had in this area (Legg,1999). Yield loss estimates were ratedat between 25% and 50% by 79% offarmers in Coast Province and at only25% by 47% of farmers in NyanzaProvince. At the time of the survey, theCMD pandemic had affected onlyWestern Province and losses recordedelsewhere were consequently less. Only14% of sweetpotato farmers consideredSPVD to be a significant source of croploss, estimating these losses at only25%.
Managing whiteflies andwhitefly-transmitted viruses
Cassava and sweetpotato producersreported a number of managementtactics, including selection of disease-free planting material, roguing of
diseased plants and use of insecticides.In extreme cases, heavily infestedcrops were simply abandoned. Themost widely used practices, however,were roguing and selection. Just overone third (38%) of cassava farmersused roguing, with 28% of farmersconsidering that this practice wouldreduce CMD incidence, 6% that itwould prevent the spread of CMD and2% that it would reduce the number ofpoorly growing plants. Amongsweetpotato farmers, 30% usedroguing, 18% believing that it wouldreduce SPVD incidence and 8%believing that it would prevent spreadof the disease. Both cassava andsweetpotato farmers rogued infectedplants before the crop was 4 monthsold. Selection of clean plantingmaterial, based on the absence ofdisease symptoms, was used as adisease management tactic by 32% ofcassava farmers and 12% ofsweetpotato farmers. However, eventhose farmers who used roguing andselection of clean cuttings consideredthese measures to be only partiallyeffective in controlling the diseases.
Only one cassava farmer and noneof the sweetpotato farmers mentionedchemical control as a control method.The cassava farmer used theorganophosphate insecticidedimethoate, applied it more than10 times and sprayed when whitefliesand damage were observed in the field.Few farmers reported abandoning theproduction of cassava (12%) orsweetpotato (2%) but many reportedoccasional shortage of clean plantingmaterial. None of the farmers grewimproved varieties, although 42% ofcassava farmers and 20% ofsweetpotato farmers noted differencesin response to CMD and SPVD betweenvarieties. However, most farmers whonoted differences between varieties hadno idea what caused the differences.Only 4% of cassava farmers and 2% of

58
Whiteflies and Whitefly-borne Viruses in the Tropics
sweetpotato farmers considered thedifferences to be the result ofdifferences in levels of resistance todisease. Very few cassava (10%) andsweetpotato (4%) farmers had receivedany technical assistance in themanagement of whiteflies and whitefly-transmitted viruses on their crops.However, more than 30% of farmers ofeach crop were willing to monitorwhitefly and disease problems if itwould help in the management ofeither CMD or SPVD.
Conclusions
Farmers noted that CMD and SPVDare becoming increasingly importantconstraints to cassava and sweetpotatoproduction in the country. Somefarmers were attempting to control thediseases, mainly using roguing andselection, initiatives that in generalthey considered to be developed fromtheir own knowledge. However, farmersrecognized the methods employed incontrolling the two diseases as beingonly partially effective. During thissurvey in 1998, no farmer reportedusing resistant varieties. The limitedunderstanding that farmersdemonstrated of the causes and effectsof CMD and SPVD, coupled with theirreliance on their “own” controlmethods, suggests weaknesses in thesystems of information flow, both fromresearch through extension to farmersand from farmers back to research.
During the survey conducted inthis study, the impact of the pandemicof severe CMD has been captured boththrough the description of theunusually high incidence and severityof the disease in Western Province andthrough the estimates of major yieldlosses provided by farmers interviewed.Considerable experience in tacklingthis problem has been developed
already in neighbouring Uganda (Otim-Nape et al., 1997) with the focus on thedeployment of host plant resistancesupported by some phytosanitation. Inthe immediate future, some of thisexperience clearly needs to betransferred to the situation in westernKenya where the CMD pandemiccontinues to expand. CMD-resistantgermplasm will need to be introducedfrom Uganda to Kenya and evaluated inparticipatory trials with farmers, whilstat the same time encouraging themaintenance of the quality of plantingmaterial through the use of selectionand roguing in multiplication plots.Given the evident shortcomings in theunderstanding of both CMD and SPVDin Kenya, participatory learning shouldbe a key feature of any future controlprograms. SPVD is clearly not as acutea problem in Kenya as is CMD.However, the report by farmers thatSPVD is becoming more important is aconcern and future work should aimtherefore at identifying resistantgermplasm and evaluating this andpossible phytosanitary approaches toSPVD control through participatoryresearch with farmers.
Acknowledgements
The technical support of Mr. Nzioki andMr. Simiyu of the KARI is highlyappreciated. The whitefly andparasitoid specimens were mountedand identified by Dr. Mohammed AliBob of the International Center ofInsect Physiology and Ecology.
References
Bock, K. R. 1975. Maize streak, cassavalatent and similar viruses. In:Abstracts, Third InternationalCongress for Virology, Madrid, ES.p. 93.

59
Kenya
Bock, K. R. 1983. Epidemiology ofcassava mosaic disease in Kenya.In: Plumb, R. T.; Thresh, J. M.(eds.). Plant virus epidemiology.Blackwell Scientific Publications,Oxford, GB. p. 337-347.
Bock, K. R. 1994a. Control of Africancassava mosaic geminivirus byusing virus-free planting material.Trop. Sci. 34:102-109.
Bock, K. R. 1994b. The spread of Africancassava mosaic geminivirus incoastal and western Kenya. Trop.Sci. 34:92-101.
Bock, K. R.; Woods, R. D. 1983. Etiologyof African cassava mosaic disease.Plant Dis. 67:944-955.
Carey, E. E.; Mwanga, R. O. M.; Fuentes,S.; Kasule, S.; Macharia, C.;Gichuki, S. T.; Gibson, R. W. 1998.Sweetpotato viruses in Uganda andKenya: Results of a survey. In:Procs. Sixth Triennial Symposium ofthe International Society of TropicalRoot Crops-Africa Branch (ISTRC-AB), 22-28 October 1995, Lilongwe,MW. p. 457-461.
Doughty, L. R. 1958. Cassava breeding forresistance to mosaic and brownstreak viruses. In: Annual report.East Afr. Agric. For. Res. p. 48-51.
Gibson, R. W.; Mpembe, I.; Alicai, T.;Carey, E. E.; Mwanga, R. O. M.;Seal, S. E.; Vetten, H. J. 1998.Symptoms, aetiology and serologicalanalysis of sweetpotato virusdisease in Uganda. Plant Pathol.47:95-102.
Gregory, P. H. 1948. The multipleinfection transformation. Ann. Appl.Biol. 35:412-417.
Harrison, B. D. 1987. Properties andgeographical variation ofgeminivirus isolates from mosaic-affected cassava. In: Africancassava mosaic disease and itscontrol. Procs. InternationalSeminar, 4-8 May 1987,Yamoussoukro, CI. CentreTechnique de Coopération Agricoleet Rurale, Wageningen, NL, andInstitut Français de RechercheScientifique pour le Dévéloppementen Coopération, Paris, FR. p. 270.
Harrison, B. D.; Zhou, X.; Otim-Nape, G.W.; Liu, Y.; Robinson, D. J. 1997.Role of a novel type of doubleinfection in the geminivirus-induced epidemic of severe cassavamosaic in Uganda. Ann. Appl. Biol.131:437-448.
Legg, J. P. 1999. Emergence, spread andstrategies for controlling thepandemic of cassava mosaic virusdisease in east and central Africa.Crop Prot. 18:627-637.
Legg, J. P.; Kapinga, R.; Teri, J.; Whyte, J.B. A. 1999. The pandemic ofcassava mosaic virus disease inEast Africa: Control strategies andregional partnerships. Roots6(1):10-19.
Otim-Nape, G. W.; Bua, A.; Thresh, J. M.;Baguma, Y.; Ogwal, S.; Semakula,G. N.; Acola, G.; Byabakama, B.;Martin, A. 1997. Cassava mosaicvirus disease in Uganda: Thecurrent pandemic and approachesto control. Natural ResourcesInstitute (NRI), Chatham, GB. 65 p.
Robertson, I. A. D. 1987. The role ofBemisia tabaci (Gennadius) in theepidemiology of ACMV in EastAfrica: Biology, populationdynamics and interaction withcassava varieties. In: Africancassava mosaic disease and itscontrol. Procs. InternationalSeminar, 4-8 May 1987,Yamoussoukro, CI. CentreTechnique de Coopération Agricoleet Rurale, Wageningen, NL, andInstitut Français de RechercheScientifique pour le Dévéloppementen Coopération, Paris, FR. p. 57-63.

60
Whiteflies and Whitefly-borne Viruses in the Tropics
Seif, A .A. 1981. Seasonal fluctuation ofadult population of the whitefly,Bemisia tabaci, on cassava. InsectSci. Appl. 1:363-364.
Seif, A. A. 1982. Effect of cassava mosaicvirus on yield of cassava. Plant Dis.66:661-662.
Sheffield, F. M. L. 1957. Virus diseases ofsweetpotato in East Africa. 1.Identification of viruses and theirinsect vectors. Phytopathology47:582-587.

61
Tanzania
CHAPTER 1.8
TanzaniaJoseph Ndunguru*, Simon Jeremiah*,
Regina Kapinga** and Peter Sseruwagi***
Introduction
Cassava (Manihot esculenta Crantz) andsweetpotato (Ipomoea batatas [L] Lam.)are important staple food crops,especially in the rural communities ofTanzania. The two crops have a longhistory of providing food security in thecountry, particularly during famine.However, their production is currentlythreatened by cassava mosaic disease(CMD) and sweetpotato virus disease(SPVD). CMD is caused by cassavamosaic begomoviruses (CMBs)transmitted by the whitefly Bemisiatabaci (Gennadius) and through virus-infected planting material (Harrison,1987). On the other hand, SPVDresults from co-infection by two distinctviruses, Sweetpotato chlorotic stunt virustransmitted by B. tabaci andSweetpotato feathery mottle virustransmitted by the aphid Myzuspersicae (Sulzer) (Gibson et al., 1998).
CMD was first reported in Tanzaniaunder the name “Krauselkrankheit”(Warburg, 1894), although it was not
recorded as causing serious lossesuntil the 1920s. Between 1920 and1960, comprehensive studies wereconducted in the country, emphasizingthe development of CMD-resistantvarieties through a breeding programconducted at Amani in the UsambaraMountains (Jennings, 1994). Resistantvarieties developed by the programmewere effective in controlling CMD andrestored the crop’s productivity. Dataobtained in 1989 and 1990 during thefirst phase of the Collaborative Study ofCassava in Africa (COSCA) indicatedthat Tanzania had the lowest CMD-incidence (37%) of the six countriessurveyed (Thresh et al., 1994). Anotherextensive survey, conducted between1993 and 1994 in mainland Tanzaniaand the islands of Zanzibar andPemba, rated the incidence of CMD at28%, with infected cuttings (24%)providing the major source of infection(Legg and Raya, 1998). During thelatter part of the 1990s, CMD researchhas been reinvigorated and hasincluded work to assess thedevelopment of the disease in thecountry.
Recent epidemiological studies onCMD elsewhere in East Africa haveprovided evidence of the north-to-southspread of an epidemic of severe CMD,firstly within Uganda (Otim-Nape et al.,1997; Legg and Ogwal, 1998; Chapter1.6, this volume) and subsequently intoparts of western Kenya and Tanzania
* Root and Tubers Programme of the LakeZone Agricultural Research andDevelopment Institute (LZARDI), Ukiriguru,Mwanza, Tanzania.
** Centro Internacional de la Papa (CIP),Kampala, Uganda.
*** International Institute of TropicalAgriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda.

62
Whiteflies and Whitefly-borne Viruses in the Tropics
Kagera
Mwanza
D.R. Congo
Rwanda
Kenya
Ugandabordering Uganda (Legg, 1999). Thisexpansion has been associated withthe spread of a novel CMB variant(Harrison et al., 1997). The pandemicposes an immediate threat to cassavaproduction in the areas of Tanzaniabordering Uganda and eventually to thecountry as a whole. Research towardscontrolling the disease is the focus ofcontinuing work by the Root andTubers Programme of the Lake ZoneAgricultural Research and DevelopmentInstitute (LZARDI) based at Ukiriguru,Mwanza. The institute is supported inthis effort by other internationalresearch institutions and networks,including the International Institute ofTropical Agriculture-Eastern andSouthern Africa Regional Center(IITA-ESARC), the East African RootCrops Research Network (EARRNET)and the Southern African Root CropsResearch Network (SARRNET). SPVD,and its effects on the production ofsweetpotato, has received less attentionin Tanzania. However, the diseaseposes a threat to the production of thecrop if left unchecked.
Diagnostic surveys were conductedin mid-1998. Three target areas werechosen to represent the major cassavaand sweetpotato growing areas in thecountry: the “lake zone” (i.e., borderingLake Victoria), which includes Mwanza,Shinyanga, Mara and Kagera regions;the “southern coast”, which includesLindi and Mtwara regions; and the“northern coast”, which includesTanga, Dar-es-Salaam and Pwaniregions (Figure 1). The study aimed atidentifying whiteflies and whitefly-transmitted viruses and characterizingproducer knowledge and responses tothese problems on cassava andsweetpotato in Tanzania.
Increased BiologicalUnderstanding
Whitefly species andabundance
Three whitefly species were identifiedon cassava: B. tabaci, B. afer (Priesnerand Hosny) and Trialeurodes ricini(Misra). Only B. tabaci and B. afer wereidentified on sweetpotato. Of303 whitefly samples collected oncassava, 175 were B. tabaci,127 B. afer and one T. ricini. The105 whitefly samples from sweetpotatocomprised 89 B. tabaci and 16 B. afer.The species B. tabaci and B. afer werefound in all three survey areas. B. aferwas more common on cassava than onsweetpotato. It comprised 26% of thewhiteflies in samples collected oncassava and 9% of those onsweetpotato from the lake zone, 53%on cassava and 11% on sweetpotatofrom the southern coast, and 45% oncassava and 27% on sweetpotato fromthe northern coast (see also Figure 2,Chapter 1.14, this volume).
Figure 1. Cassava- and sweetpotato-growingareas surveyed for whitefly incidencein Tanzania.
Mara
Tanzania
Mozambique
Shinyanga
Tanga
Dar-es-Salaam
Pwani
Lindi
Mtwara
LakeVictoria
Burundi

63
Tanzania
Adult whiteflies were moreabundant on cassava in the northerncoast (5.6 per top five leaves) than ineither the southern coast (1.1) or thelake zone (0.7). On sweetpotato,estimates of whitefly abundance werehighest in the lake zone (40.1 per1-min count) and lowest in thesouthern coast (2.8)
Disease incidence andsymptom severity
CMD incidence was generally low in thethree target areas. The highestincidence was recorded in the northerncoast (33.6%) and the lowest (9.4%) inthe southern coast (Table 1). The mainsource of infection in the lake zone(69.3% of the total infection in area)and northern coast (56.8% of the totalinfection in area) was cutting infection.However, whitefly-borne infection wasthe predominant source of infection inthe southern coast. CMD symptomswere mild in the lake zone andsouthern coast and slightly moresevere in the northern coast.
The incidence of SPVD was highestin the southern coast (12.4%), followedby the lake zone (11.3%), and lowest inthe northern coast (4.2%) survey area(Table 1). SPVD symptoms wererelatively mild in most of the surveyedregions, with the exception ofShinyanga region in the lake zone,where the disease was very severe (4.5).The low incidence of SPVD shouldenable the use of phytosanitarymeasures, including selection ofplanting material and roguing in themanagement of the disease.
Whitefly parasitoids
The survey identified two whiteflyparasitoids, Encarsia sophia (Giraultand Dodd) and Encarsia sp. (luteolagroup). E. sophia was by far the morewidely recorded, with 43 samplesidentified in the southern coast and
19 in the northern coast samples,while only one sample of Encarsia sp.(luteola group) was recorded, in thenorthern coast. Little is known aboutthe role of natural enemies in thepopulation dynamics of whiteflies inTanzania.
Increased Socio-EconomicUnderstanding
Farmers’ assessment ofwhitefly-related problems
A small proportion of both cassava(30%) and sweetpotato (11.7%) farmerswere able to recognize whiteflies on thetwo crops, and even the names given tothe insect were non-specific. Moreover,of the farmers who recognized thewhiteflies only 16.7% (on cassava) and3.3% (on sweetpotato) considered thema problem in the production of theirrespective crops.
Most cassava farmers (70%) couldrecognize CMD as a disease of cassava,58% recognized it as a constraint tocrop production and 68% as a problemoccurring every year in their crops. Incontrast, only 38% of sweetpotatofarmers recognized SPVD as a diseaseof sweetpotato, 32% recognized it as aconstraint to production and 41% as ayearly recurring problem. One third ofcassava farmers considered that CMDseverity was increasing, whilst aslightly smaller proportion ofsweetpotato farmers considered thatSPVD was becoming more important.Forty percent of cassava farmers and32% of sweetpotato farmers believedthat climate influenced the diseases;two-thirds believed that they were moreprevalent during periods of low rainfalland high temperatures. Cassavafarmers attributed higher losses toCMD than did sweetpotato farmers toSPVD. Using five categories (zero,quarter, half, three quarters and total

64
Whiteflies and Whitefly-borne Viruses in the Tropics
Table
1.
Inci
den
ce (%
) of
cass
ava
mos
aic
dis
ease
(C
MD
) an
d s
wee
t pot
ato
vir
us
dis
ease
(S
PV
D), d
isea
se s
ever
ity
an
d w
hit
efly
abu
ndan
ce o
n c
ass
ava
an
d s
wee
tpot
ato
in
sel
ecte
d r
egio
ns
of T
an
zan
ia, 1998.
Su
rvey
are
aR
egio
nC
ass
ava
aS
wee
t pot
ato
a
No.
Wh
itef
lyC
MD
No.
Wh
itef
lyS
PV
Dfiel
ds
cou
nts
fiel
ds
cou
nts
Wh
itef
lyC
utt
ing
Tot
alS
ever
ity
Inci
den
ceS
ever
ity
infe
ctio
nin
fect
ion
inci
den
ce
Lake
zon
eK
age
ra6
2.2
0 (0)
30.7
30.7
2.6
518.0
20.0
3.1
Mara
50.1
1.3
(1.4
)
7.3
8.6
2.6
655.9
5.3
2.8
Sh
inya
nga
50.4
0.7
(0.9
)20.7
21.4
2.6
425.3
2.0
4.5
Mw
anza
40.2
3.3
(4.1
)18.0
21.3
2.4
561.1
18.0
3.1
Mea
n0.8
1.2
(1.7
)19.8
21.0
2.6
40.1
11.5
3.3
Sou
ther
n c
oast
Lin
di
61.0
2.8
(2.8
)
0
2.8
2.8
9
2.9
8.3
2.1
Mtw
ara
14
1.2
8.1
(9.2
)
7.9
16.0
2.5
11
2.7
16.4
2.4
Mea
n1.2
6.5
(7.1
)
5.5
12.0
2.7
2.8
12.8
2.3
Nor
ther
n c
oast
Pw
ani
78.9
15.4
(18.0
)
6.7
22.1
3.0
13
1.8
9.2
2.7
DS
Mb
34.3
14.4
(16.3
)
4.4
18.8
3.0
5
5.0
3.3
2.0
Tan
ga10
3.6
13.7
(29.5
)46.3
60.0
3.1
2
5.7
02.0
Mea
n5.8
14.4
(21.0
)26.2
40.6
3.0
3.1
6.8
2.5
a.
Fig
ure
s are
mea
ns
for
each
reg
ion
. W
hit
efly
cou
nts
, w
hit
efly
abu
ndan
ce o
n c
ass
ava
(n
um
ber
of w
hit
eflies
per
top
fiv
e le
ave
s) a
nd o
n s
wee
t pot
ato
(per
min
ute
cou
nt)
; w
hit
efly
in
fect
ion
, figu
res
in p
are
nth
eses
tra
nsf
orm
ed t
o m
ult
iple
in
fect
ion
un
its
to a
llow
for
mu
ltip
le in
fect
ion
(G
rego
ry, P. H
. 1948. Th
e m
ult
iple
infe
ctio
n t
ran
sfor
mati
on. A
nn
. A
ppl. B
iol. 3
5:4
12-4
17); s
ever
ity
of d
isea
se m
easu
red o
n a
n a
scen
din
g 1-5
sca
le, fr
om low
to
seve
re.
b.
DS
M, D
ar
es S
ala
am
.

65
Tanzania
loss) to assess yield loss attributable toeither CMD or SPVD, 20% of thecassava farmers believed that CMDcauses at least 50% yield loss, while23% considered the disease to causeonly 25% yield loss. Among thesweetpotato farmers, over 50%considered that SPVD caused only25% yield loss.
Managing whiteflies andwhitefly-transmitted viruses
Both cassava and sweetpotato farmersattempted to control CMD and SPVDby roguing and selecting disease-freeplanting material. Roguing was usedfor disease management by 23% ofcassava farmers and 5% of sweetpotatofarmers. Farmers growing cassava gavethree reasons for roguing diseasedplants: reducing the spread of thedisease (18%), poor growth (3%) andpreventing disease (2%). However, only5% of sweetpotato farmers consideredthat roguing would reduce SPVD. Mostcassava farmers rogued infected plantsbefore the crop was 4 months old. Forsweetpotato, there was no specificperiod during which roguing was done.Seventy-three percent of cassavafarmers compared with 47% ofsweetpotato farmers selected cleanplanting material, with absence ofdisease symptoms as the selectioncriterion for the two diseases. In bothcases, however, farmers considered thetwo principal methods to be onlypartially effective in managing thediseases.
Only one sweetpotato farmer andnone of the cassava farmers mentionedchemical control as a managementtactic. The sweetpotato farmer used theorganophosphate pesticide dimethoate,applied 3 times, when whiteflies anddamage were observed in the field. Fewfarmers reported abandoning theproduction of cassava (10%) andsweetpotato (3%) but many reportedoccasional shortage of clean planting
material. The use of host plantresistance as a management option wasmentioned by only 2% of cassavafarmers but not mentioned at all bysweetpotato farmers. Only 30% ofcassava farmers and 22% ofsweetpotato farmers noted differencesin response to CMD and SPVD betweenvarieties. Those that did so mainlyattributed it to differences in levels ofresistance to disease. In general, veryfew cassava (8%) and sweetpotato (2%)farmers had received any technicalassistance in the management ofwhiteflies and associated virus diseaseson either crop. However, most farmerswere willing to change the plantingdates of their crop and monitor whiteflyand disease problems if it would help inthe management of either disease.
Conclusions
Both diseases covered in this study arebecoming more prevalent in Tanzania(Chapters 1.13 and 1.14, this volume)and therefore measures to reduce theirimpact are needed urgently.
Farmers are more aware of CMDthan of SPVD. However, theirknowledge of management options forthe two diseases is weak, implying poorcommunication among researchers,extension workers and farmers.Farmers’ knowledge of these diseasesshould be enhanced throughparticipatory training, whileresearchers, farmers and otherstakeholders should work together todevelop and validate cost-effectivedisease and vector managementstrategies and encourage their wideradoption by producers.
Varieties that are resistant to CMDand SPVD, acceptable to farmers andwell adapted to local conditions areurgently needed. The development,farm-level evaluation and wider

66
Whiteflies and Whitefly-borne Viruses in the Tropics
distribution of improved varieties ofcassava and sweetpotato should bestrengthened and expanded.
The whitefly parasitoid E. sophiaoccurred frequently in the areassurveyed, and studies elsewhere inEast Africa have shown that parasitoidscan cause B. tabaci nymph mortalityrates of up to 50%. The deployment ofresistant varieties and phytosanitationmeasures probably will be the mostreadily applicable CMD and SPVDmanagement measures in Tanzania.However, there may be scope in thefuture for improved management of thewhitefly vector through thedevelopment and application ofmeasures designed to conserve andaugment the activity of whitefly naturalenemies. The farmer field school (FFS)approach is being widely used inTanzania, and FFS in root crop growingareas may provide an ideal opportunityto strengthen farmers’ knowledge ofCMD and SPVD and develop a holistic,integrated approach to theirmanagement, incorporating all possiblecomponents including host plantresistance, cultural methods andbiological control.
Acknowledgements
The technical support of Dr. RichardGibson of the Natural ResourcesInstitute, UK, Mr. Mathew Raya,Mr. Geoffrey Mkamiro, Mr. RaphaelMsabaha of Naliendele AgriculturalResearch Institute and Ms. HellenKiozya and Ms. Esther Masumba ofKibaha Research Institute is highlyappreciated. The authors alsoacknowledge Dr. Mohammed Ali Bob ofthe International Center of InsectPhysiology and Ecology for hisidentification of whitefly species andnatural enemies.
References
Gibson, R. W., Mpembe, I.; Alicai, T.;Carey, E. E.; Mwanga, R. O. M.;Seal, S. E.; Vetten, H. J. 1998.Symptoms, aetiology andserological analysis of sweetpotatovirus disease in Uganda. PlantPathol. 47:95-102.
Gregory, P. H. 1948. The multipleinfection transformation. Ann. Appl.Biol. 35:412-417.
Harrison, B. D. 1987. Properties andgeographical variation ofgeminivirus isolates from mosaic-affected cassava. In: Africancassava mosaic disease and itscontrol, Procs. InternationalSeminar, 4-8 May 1987,Yamoussoukro, CI. CentreTechnique de Coopération Agricoleet Rurale, Wageningen, NL, andCentre Technique de CoopérationAgricole et Rurale, Wageningen, NLand Institut Français de RechercheScientifique pour le Dévéloppementen Coopération, Paris, FR. p. 270.
Harrison, B. D.; Zhou, X.; Otim-Nape, G.W.; Liu, Y.; Robinson, D. J. 1997.Role of a novel type of doubleinfection in the geminivirus-induced epidemic of severe cassavamosaic in Uganda. Ann. Appl. Biol.131:437-448.
Jennings, D. L. 1994. Breeding forresistance to African cassavamosaic geminivirus in East Africa.Trop. Sci. 34:110-122.
Legg, J. P. 1999. Emergence, spread andstrategies for controlling thepandemic of cassava mosaic virusdisease in east and central Africa.Crop Prot. 18:627-637.
Legg, J. P.; Ogwal, S. 1998. Changes inthe incidence of African cassavamosaic geminivirus and theabundance of its whitefly vectoralong south-north transects inUganda. J. Appl. Entomol.122:169-178.

67
Tanzania
Legg, J. P.; Raya, M. 1998. Survey ofcassava virus diseases inTanzania. Int. J. Pest Manage.44:17-23.
Otim-Nape, G. W.; Bua, A.; Thresh, J.M.; Baguma, Y.; Ogwal, S.;Semakula, G. N.; Acola, G.;Byabakama, B.; Martin, A. 1997.Cassava mosaic virus disease inUganda: The current pandemicand approaches to control.Natural Resources Institute (NRI),Chatham, GB. 65 p.
Thresh, J. M.; Fishpool, L. D. C.; Otim-Nape, W.; Fargette, D. 1994.African cassava mosaic virusdisease: An under-estimated andunsolved problem. Trop. Sci.34:3-14.
Warburg, O. 1894. Die KulturpflanzenUsambaras. Mitt. Dtsch. Schutz.7:131.

68
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 1.9
MalawiMathews P. K. J. Theu* and Peter Sseruwagi**
Introduction
Cassava mosaic disease (CMD) iscaused by cassava mosaicbegomoviruses, transmitted by thewhitefly Bemisia tabaci (Gennadius)and through virus-infected plantingmaterial (Harrison, 1987). Thedisease has gained significance as amajor constraint to production inmany areas of Malawi where cassava(Manihot esculenta Crantz) is grown,especially in the lowlands, whereconditions are warmer; infectionreportedly exceeds 90% in someplantings (Nyirenda et al., 1993).
A survey was conducted in threetarget areas selected to represent themajor cassava growing areas inMalawi: the central lakeshore(Salima and Nkhotakhota, at500-800 m altitude); the northernlakeshore (Rumphi and Nkhata Bay,500-800 m); and the central plateau(Dedza, Lilongwe and Dowa,1000-1500 m) (Figure 1). The keyobjectives of the study were toidentify whiteflies and whitefly-transmitted viruses and to
characterize producer knowledge ofthese problems on cassava in Malawi.Although assessments of sweetpotatovirus disease (SPVD) also were plannedinitially, surveys were done at a time(July-September 1998) when crops ofsweetpotato (Ipomoea batatas [L.] Lam.)mostly had been harvested, so diseaseand vector incidence could not beassessed adequately.
* Chitedze Research Station, Lilongwe, Malawi.** International Institute of Tropical
Agriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda.
Figure 1. Areas surveyed for whitefly incidenceand cassava mosaic disease inMalawi.
Rumphi
NkhataBay
Mozambique
Tanzania
Zambia
Nkhotakota
Malawi
Zimbabwe
Dowa
SalimaLilongwe
Dedza

69
Malawi
Increased BiologicalUnderstanding
Whitefly species andabundance
Two whitefly species, B. tabaci andB. afer (Priesner and Hosny), wereidentified on cassava. Altogether,212 whitefly samples were collected onthe crop. Of these, 65 were B. tabaciand 147 were B. afer. The relativeabundance of the two species variedamong survey areas. B. afer wasespecially abundant in the central mid-altitude plateau, where it comprised97% of the whitefly samples collected.The adult whitefly population did notvary significantly among the regionssurveyed (Table 1).
Disease incidence andsymptom severity
The incidence of CMD ranged from11.4% in the central plateau surveyarea to 62.2% for the central lakeshore(Table 1). The use of diseased cuttingswas the main source of infection in allthree survey areas. Symptoms of CMDwere moderately severe in each of thesurvey areas and averaged 2.8 overall.
Table 1. Cassava mosaic disease (CMD) incidence (%), disease symptoms and whitefly counts inselected areas of Malawi, 1998.
Target area Provinces/ No. Whitefly CMDa
districts fields counts Whitefly Cutting Total Severityinfection infection incidence
Central Salima andlakeshore Nkhotakota 20 1.5 27.5 (54.7) 34.7 62.2 2.8
Northern Rumphi andlakeshore Nkhata Bay 9 1.0 19.6 (34.4) 32.6 52.2 2.7
Central Dedza, Lilongweplateau and Dowa 12 1.3 3.3 (3.7) 8.1 11.4 3.0
Mean 1.3 16.8 (25.4) 25.1 41.9 2.8
a. Figures are means for each region. Whitefly counts, whitefly abundance on cassava (number ofwhiteflies per top five leaves); whitefly infection, figures in parentheses transformed to multipleinfection units to allow for multiple infection (Gregory, P. H. 1948. The multiple infectiontransformation. Ann. Appl. Biol. 35:412-417); severity of disease measured on an ascending1-5 scale, from low to severe.
Whitefly parasitoids
Only one species of whitefly parasitoid,Encarsia sophia (Girault and Dodd) wasidentified and this was from thenorthern lakeshore.
Increased Socio-EconomicUnderstanding
Farmers’ assessment ofwhitefly-related problems
Forty-four percent of the farmersinterviewed were able to recognizewhiteflies on the crop but only 32%considered them a problem. Mostfarmers (61%) were able to recognizeCMD as a disease of cassava and gaveit a range of local names. Less thanhalf the farmers (42%) recognized thedisease as a problem to cassavaproduction and most (64%) noted thatit occurs yearly. Only 22% believedthat CMD was becoming more severe,with 1997 noted as the year of mostsevere disease symptoms. About onequarter (27%) of the farmers believedthe disease to be affected by climate.There was no consensus on the effectof rainfall on CMD but 39% of farmers

70
Whiteflies and Whitefly-borne Viruses in the Tropics
considered that the disease was mostsevere during periods of hightemperature. Farmers’ estimates of thelosses attributable to CMD variedconsiderably, ranging from “none” to“total crop loss”.
Managing whiteflies andwhitefly-transmitted viruses
Only 7.3% of farmers attempted tocontrol CMD; 2.4% rogued, 3.7%selected for healthy planting materialat planting, while some 1.2%de-topped CMD-infected plants toreduce plant shoots with diseasesymptoms in the field. Farmers whoselected their planting material mainlyused the absence of disease symptomsas the selection criterion. Farmers whorogued gave three reasons for doingso—prevention of disease spread (7%),reduction of disease (7%) and removalof poorly growing plants (5%). Most ofthe roguing was done before the cropwas 4 months old. Only 5% of farmersconsidered selection and 7%considered roguing as highly effectivein controlling the disease. Somefarmers (10%) reported the occasionalshortage of clean planting material,with 5% reporting abandoningproduction of the crop in 1990 and1992. This may have been associatedwith the severe damage caused by thecassava mealybug, Phenacoccusmanihoti Matile-Ferrero, at this time.Improved varieties, primarily developedfor their superior yield and agronomiccharacteristics, were grown by only 5%of the farmers, who considered theirperformance to be better than that oflocal varieties. Very few farmers (5%)had received any technical assistancein the management of whiteflies.However, a large proportion (40%) offarmers were willing to monitor whiteflyand disease problems if it would helpin the management of the disease.
Conclusions
Prospects for managing CMDsuccessfully are good in most locationssampled, since the use of diseasedcuttings was the main source ofinfection. However, despite the majorityof farmers recognizing the disease as aproduction constraint to cassava, theirknowledge of the disease was weak.The incidence of CMD was relativelyhigh in the lakeshore survey areas and,given that moderate symptoms wereobserved, it is likely that significantyield losses are being experienced.Given that there appears to be verylimited CMD spread due to the activityof whiteflies, and that B. afer, a non-vector, is the predominant cassavawhitefly species in Malawi,phytosanitation may provide aneffective approach to CMDmanagement. However, before anappropriate regime for the use ofroguing and the selection of CMD-freeplanting material can be proposed,basic information will be required onyield losses and rates of infection in themajor cassava-growing areas for themost commonly grown varieties. Theseresults should be used to develop aprogramme of participatory evaluationwith farmers of both phytosanitationand new varieties. This should beclosely linked with an educationalprogramme targeted at increasingfarmers’ knowledge of CMD and thewhitefly vector and of the range ofpossible management approaches.
Acknowledgements
The authors gratefully recognize thetechnical support of Mr. McDonaldKavala, Mr. Elias Kamsikili andMr. Eric Mazuma. Assistance fromDr. Mohammed Ali Bob of theInternational Center of Insect

71
Malawi
Physiology and Ecology in theidentification of whitefly species andnatural enemies was highlyappreciated.
References
Gregory, P. H. 1948. The multipleinfection transformation. Ann. Appl.Biol. 35:412-417.
Harrison, B. D. 1987. Properties andgeographical variation ofgeminivirus isolates from mosaic-affected cassava. In: African cassavamosaic disease and its control.Procs. International Seminar,4-8 May 1987, Yamoussoukro, CI.Centre Technique de CoopérationAgricole et Rurale, Wageningen, NL,and Institut Français de RechercheScientifique pour le Dévéloppementen Coopération, Paris, FR. p. 270.
Nyirenda, G. K. C.; Munthali, D. C; Phiri,G. S. N.; Sauti, R. F. N.; Gerling, D.1993. Integrated pest managementof Bemisia spp. whiteflies in Malawi.Makoka Research Station,Thondwe, MW.

72
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 1.10
MadagascarSahondramalala Ranomenjanahary*,
Jocelyn Ramelison* and Peter Sseruwagi**
Introduction
Cassava mosaic disease (CMD), causedby cassava mosaic begomovirusestransmitted by the whitefly Bemisiatabaci (Gennadius) and through virus-infected planting material (Harrison,1987), is the most important viraldisease of cassava (Manihot esculentaCrantz) in Madagascar. The diseasewas first reported in Madagascar in1932, although it was of only minorimportance at this time (François,1937). Epidemics were first reported in1934, from an area to the west of LakeAlatroa on the central plateau and,between this time and the end of the1930s, the epidemic spread to cover allcassava-growing areas of the island(Cours, 1951). Symptoms in localvarieties were very severe and cassavacultivation was abandoned on a widescale. The impact was so great that thegovernment ordered urgent attention tobe given to the problem (Frappa, 1938)and a major programme for thedevelopment of resistant varieties wasinitiated (Cours, 1951; Cours-Darne,1968). The first of these varieties waswidely disseminated in the mid-1940s.Since this time, CMD appears to have
become a problem of relatively minorsignificance.
Sweetpotato virus disease (SPVD)results from co-infection of sweetpotato(Ipomoea batatas [L.] Lam.) by whitefly-transmitted Sweetpotato chlorotic stuntvirus and aphid-transmittedSweetpotato feathery mottle virus(Gibson et al., 1998). SPVD has notbeen reported as a major problem inMadagascar, although the country isone of the largest producers ofsweetpotato in Africa.
In order to provide a basicunderstanding of the incidence ofwhiteflies, CMD and SPVD inMadagascar, a study was conducted infour survey areas that represent themajor cassava and sweetpotato growingareas: Antananarivo (central mid-altitude plateau), Fianarantsoa (easternhumid coast), Toliara (south-westernsemi-arid coast) and Mahajanga(northern sub-humid coast) (Figure 1).This initial phase of the study aimed atdocumenting the status of whitefliesand whitefly-transmitted viruses and atdescribing producer knowledge of theseproblems on cassava and sweetpotato,with a view to prioritizing furtheractions and eventually developingintegrated management strategies totackle the most serious problemsidentified. The survey was carried outduring the months of June and August1998.
* Laboratoire de Pathologie Végétale,Ambatobe, Madagascar.
** International Institute of TropicalAgriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda.

73
Madagascar
Increased BiologicalUnderstanding
Whitefly species andabundance
Two whitefly species, B. tabaci andB. afer (Priesner and Hosny) wereidentified on both cassava andsweetpotato. Of 415 whitefly samplescollected on the two crops, 118 wereB. tabaci and 297 were B. afer. Adultwhitefly populations on cassava weregreatest in the Antananarivo surveyarea and lowest in Mahajanga (Table 1,see also Figure 3 in Chapter 1.14, thisvolume). Whiteflies were very scarce onsweetpotato.
Disease incidence andsymptom severity
The highest disease incidence (71%)was recorded in Fianarantsoa, while
the lowest (31%) was recorded inAntananarivo (Table 1). The planting ofinfected cuttings was much moreimportant than whitefly transmissionas a source of infection and comprised86% of total infection. CMD symptomseverity was relatively mild inAntananarivo but was severeelsewhere.
SPVD was of very low incidence orabsent, except in the Toliara surveyarea, where an incidence of 18% wasrecorded. SPVD symptoms were severein Antananarivo but very mild wherethe disease occurred elsewhere.
Increased Socio-EconomicUnderstanding
Farmers’ assessment ofwhitefly-related problems
Thirteen percent of cassava farmersbut only 3% of sweetpotato farmersrecognized whiteflies. Scarcely any ofthese farmers considered whiteflies tobe a problem on either crop. Almostone fifth (19%) of cassava farmers wereable to recognize CMD as a disease ofcassava and gave the disease a range oflocal names, whereas no sweetpotatofarmers recognized SPVD. Only 6% ofcassava farmers considered CMD as aproduction constraint to cassava. Theynoted that the disease occurs yearlyand that it was becoming more severe.Climate affects CMD and SPVDincidence according to 30% of cassavafarmers and 5% of sweetpotatofarmers. In particular, 19% of cassavafarmers noted that disease is moresevere during periods of little rain andhigh temperature but sweetpotatofarmers did not report this effect. Noneof the farmers could estimate the levelof yield loss attributable to eitherdisease. The lack of recognition ofCMD, first as a disease and second asa constraint to production, is
Mozambique
Tanzania Madagascar
Figure 1. Cassava- and sweetpotato-growingareas surveyed for whitefly incidencein Madagascar.
Mahajanga
Antananarivo
Fianarantsoa
Toliara

74
Whiteflies and Whitefly-borne Viruses in the Tropics
Table
1.
Inci
den
ce o
f ca
ssava
mos
aic
dis
ease
(C
MD
) an
d s
wee
tpot
ato
vir
us
dis
ease
(S
PV
D), d
isea
se s
ever
ity
an
d w
hit
efly
abu
ndan
ce o
n c
ass
ava
an
d o
nsw
eetp
otato
in
sel
ecte
d r
egio
ns
of M
adaga
scar,
1998.
Pro
vin
ceC
assa
vaa
Sw
eetp
otat
oa
No.
Wh
itef
lyC
MD
No.
Wh
itef
lyS
PV
Dfiel
ds
cou
nts
fiel
ds
cou
nts
Wh
itef
lyC
utt
ing
Tot
alS
ever
ity
Inci
den
ceS
ever
ity
infe
ctio
nin
fect
ion
inci
den
ce
An
tan
anar
ivo
35
7.1
5.6
(7.8
)25.5
31.0
2.6
11
0.0
1
2.6
3.3
Fia
nara
nts
oa22
4.3
7.1
(21.9
)63.9
71.0
3.5
15
0
2.4
2.0
Tol
iara
12
5.9
6.4
(11.2
)39.7
46.0
3.2
70.1
017.6
2.3
Mah
ajan
ga42
2.5
6.9
(11.0
)34.0
41.0
3.3
60
0
-
Mea
n5.0
6.5
(11.6
)40.8
47.3
3.1
-0.0
3
5.7
2.5
a.
Fig
ure
s are
mea
ns
for
each
pro
vin
ce. W
hit
efly
cou
nts
, w
hit
efly
abu
ndan
ce o
n c
ass
ava
(n
um
ber
of w
hit
eflies
per
top
fiv
e le
ave
s) a
nd o
n s
wee
tpot
ato
(per
min
ute
cou
nt)
; w
hit
efly
in
fect
ion
, figu
res
in p
are
nth
eses
tra
nsf
orm
ed t
o m
ult
iple
in
fect
ion
un
its
to a
llow
for
mu
ltip
le in
fect
ion
(G
rego
ry, P. H
. 1948. Th
e m
ult
iple
infe
ctio
n t
ran
sfor
mati
on. A
nn
. A
ppl. B
iol. 3
5:4
12-4
17); s
ever
ity
of d
isea
se m
easu
red o
n a
n a
scen
din
g 1-5
sca
le, fr
om low
to
seve
re.

75
Madagascar
surprising, particularly given thehistory of previous CMD epidemics inMadagascar and the severity of thedisease symptoms observed.
Managing whiteflies andwhitefly-transmitted viruses
Selection of clean planting material wasthe only disease management approachused—as reported by 40% of cassavafarmers and 13% of sweetpotatofarmers. Sixteen percent of cassavafarmers and 3% of sweetpotato farmersused the absence of disease symptomsas selection criterion but bothconsidered the method to be onlypartially effective. Only 27% of cassavafarmers and 3% of sweetpotato farmerswere able to recognize differencesbetween varieties of their respectivecrops and even fewer (only 2% ofcassava farmers and no sweetpotatofarmers) could describe or characterizethese differences. Cassava farmersgrowing improved varieties noted thattheir performance was only slightlybetter than the local varieties. Only 4%of cassava farmers and no sweetpotatofarmers received technical assistancein whitefly or disease management.None of the farmers considered thatenvironmental factors such as certainpatterns of weather gave them any ideaabout what whitefly populations ordisease levels would result and mostwere unwilling to change the plantingdates of their crops even if it mighthelp reduce the effects of whitefliesand/or disease. The farmers’reluctance is understandable, however,given the relatively short rainy seasonin much of Madagascar that leavesthem little scope to adjust plantingdates.
Conclusions
The incidence of CMD was low in manyfarmers’ fields, compared with figuresreported from countries on the African
mainland, at least in three of the foursurvey areas. Infection was mainlyattributable to the use of diseasedplanting material. However, the highseverity of the symptoms in plantingswhere the disease does occur suggeststhat farmers are probably sustainingsignificant yield losses. It would bevaluable to quantify yield losses in localcultivars being grown in each of thetarget areas. Such an exercise shouldbe carried out with the fullparticipation of farmers and could beused simultaneously to introduce newgermplasm and evaluate itsperformance and acceptability.
The sharp disparity in SPVDincidence between Toliara and theother three survey areas is difficult toexplain. Although the abundance ofwhitefly adults was greatest in thistarget area, whitefly populations weregenerally very low compared to thoserecorded in other countries. Inaddition, at the time the survey wasconducted, B. afer, a non-vector, wasthe predominant species on bothcassava and sweetpotato. More detailedepidemiology and population dynamicsstudies investigating changes inabundance of the two whitefly speciesand disease spread over time would berequired in order to identify thereason(s) behind this difference.
Both cassava and sweetpotatofarmers had inadequate knowledge ofthe principal diseases and the optionsavailable for managing them. Aparticipatory research and trainingeffort would enhance farmers’knowledge and would help researchersunderstand the constraints to farmers’adoption of resistant varieties andother potential management tactics. Itwould be valuable also to continue tomonitor the incidence of the twodiseases in the country and strengthenthe national breeding program, tocompare local sources of resistance

76
Whiteflies and Whitefly-borne Viruses in the Tropics
with those available elsewhere and sodevelop resistant varieties acceptable tofarmers.
Acknowledgements
Appreciation is extended to Ms. BakolyRavoniarimelina, Centre National deRecherche sur l’Environnement (CNRE)and Mr. Ernest Randrianarisoa,Departement d’Entomologie, Facultédes Sciences, Universitéd’Antananarivo, who participated indata collection, and Dr. Mohammed AliBob, of the International Center ofInsect Physiology and Ecology, for hisidentification of whitefly species.
References
Cours, G. 1951. Le manioc à Madagascar.Mémoires de l’Institut Scientifiquede Madagascar Série B Biol. Vég.3:203-400.
Cours-Darne, G. 1968. Improving cassavain Africa. The Abidjan Conference.Agr. Res. Prior. Econ. Dev. Afr.2:330-339.
François, E. 1937. Un grave peril: Lamosaïque du manioc. Agron. Colon.26:33-38.
Frappa, C. 1938. Les insectes nuisiblesau manioc sur pied et auxtubercules de manioc en magasin àMadagascar. Rev. Bot. Appl. 28:17-29.
Gibson, R. W.; Mpembe, I.; Alicai, T.;Carey, E. E.; Mwanga, R. O. M.;Seal, S. E.; Vetten, H. J. 1998.Symptoms, aetiology and serologicalanalysis of sweetpotato virusdisease in Uganda. Plant Pathol.47:95-102.
Gregory, P. H. 1948. The multipleinfection transformation. Ann. Appl.Biol. 35:412-417.
Harrison, B. D. 1987. Properties andgeographical variation ofgeminivirus isolates from mosaic-affected cassava. In: African cassavamosaic disease and its control.Procs. International Seminar,4-8 May 1987, Yamoussoukro, CI.Centre Technique de CoopérationAgricole et Rurale, Wageningen, NL,and Institut Français de RechercheScientifique pour le Développementen Coopération, Paris, FR. p. 270.

77
Diversity of African Cassava Mosaic Disease
CHAPTER 1.11
The Diversity of Cassava MosaicBegomoviruses in Africa
Peter Markham*, Robert Briddon*, Clotilde Roussot*,John Farquhar*, Geoffrey Okao-Okuja** and James Legg***
Introduction
Cassava mosaic disease (CMD) is themost widespread and economicallyimportant disease of cassava (Manihotesculenta Crantz) in tropical Africa. Adisease of cassava was first describedin East Africa in the nineteenth century(Warburg, 1894) and cassava mosaic,as it came to be called, wassubsequently reported as spreadingthroughout the cassava-growing areasof Central and West Africa (Calvert andThresh, 2002). In more recent times,CMD has been reported as occurring atvarying levels of incidence throughoutthe cassava belt of Africa and losseshave been estimated at between 12%and 25% of total production (Thresh etal., 1997).
In 1983, the first sequence of ageminivirus to be determined andpublished was that of African cassavamosaic virus (ACMV) (Stanley and Gay,1983). Later, this virus was included inthe genus Begomovirus of the familyGeminiviridae (Briddon and Markham,1995), consisting of viruses withbipartite genomes transmitted by the
whitefly Bemisia tabaci (Gennadius).For some time, ACMV was thought tobe the only geminivirus speciesassociated with CMD, despite thesuggestion by Harrison and Robinson(1988), based upon serologicalanalysis, that CMD was regionallydistinct. Some 10 years after thepublication of the sequence of ACMV,Hong et al. (1993) characterized afurther CMD-associated geminivirus.Although based upon only thesequence of the DNA A genomiccomponent (and therefore lacking anycorroborative information on infectivity,since this is determined by theB genome component) this virus wasgiven the name East African cassavamosaic virus (EACMV). A furthergeminivirus species has been foundsince in southern Africa and namedSouth African cassava mosaic virus(SACMV) (Berrie et al., 1998).
During the 1990s, an unusuallysevere and damaging form of CMDspread rapidly through Uganda and thewider East African region and came tobe referred to as the CMD “pandemic”(Otim-Nape et al., 1997; Legg, 1999). Anovel recombinant geminivirus wasshown to be consistently associatedwith the pandemic (Zhou et al., 1997),commonly in dual infections withACMV that gave rise to the severestsymptoms (Harrison et al., 1997). Theso-called Uganda variant has beenshown to be a recombinant between
* John Innes Centre (JIC), Norwich ResearchPark, Colney, Norwich, UK.
** International Institute of TropicalAgriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda.
*** IITA-ESARC, Kampala, Uganda, andNatural Resources Institute, University ofGreenwich, Chatham Maritime, Kent, UK.

78
Whiteflies and Whitefly-borne Viruses in the Tropics
ozambique
nzania
EACMV and ACMV, with ACMVproviding an approximately 400 bpfragment of the coat protein gene; thisvirus is therefore best described asEACMV-Uganda (EACMV-Ug).Subsequent studies have identified DNAB components for EACMV-Ug, haveconfirmed the occurrence in Uganda ofEACMV and have demonstratedinfectivity through the generation ofinfectious clones (Pita et al., 2001).
At the beginning of this project itwas apparent that the differentbegomovirus species and strainsassociated with CMD did not occurnecessarily in different regions of Africa.It was therefore a major objective of thework to survey the diversity of cassavamosaic geminiviruses in the maincassava growing areas of Africa anddetermine their geographicaldistribution.
Methods and Materials
Cassava is normally propagated bystem cuttings and national collaboratorstherefore collected stem samples fromCMD-infected plants during theirproject surveys, and shipped these tothe John Innes Centre (JIC), UK. Thematerial was planted in compost incontainers (7.6 cm diameter) andmaintained at about 25 °C (± 5 °C) inglasshouses, with supplementarylighting between October and April.
Samples of nucleic acids forpolymerase chain reaction (PCR)analysis were extracted from 0.1 gsamples of infected leaf tissue using theNucleon Phytopure plant DNAextraction kit (Amersham) essentially asdescribed by the manufacturer. Themain modification of the procedure wasthat liquid nitrogen was used in placeof dry ice. Samples were stored at–200 °C and used directly in the PCRreactions.
The PCR procedure and the primersutilized have been described previously(Briddon and Markham, 1994). Typically1 µL, or 1 µL of a 10-fold dilution, of theDNA extract was used in theamplification reactions. Reaction volumesof 100 µL were used and amplificationproducts were cleaned further by phenolextraction and ethanol precipitationbefore use in the restriction fragmentlength polymorphism (RFLP) analysis.
Typically 10 µL (of the original100 µL reaction volume) of the PCRproduct was used for restriction enzymedigestion in a 100 µL reaction volumefollowing the manufacturers’ (Gibco-BRL)instructions. After a 4-hour digestion,RFLP products were resolved on 1%agarose gels in TBE buffer stained withethidium bromide. For most cases thissufficed to determine the restrictionpattern. However, for a few samples theamplification was poor. This necessitatedblotting of the gel to Hybond membranes(Amersham) and probing withradioactively labeled probes. As a rule, allgels were blotted to Hybond forsubsequent analysis.
Restriction patterns with particularviruses were predicted from publishedsequences (ACMV: Stanley and Gay,1983; Morris et al., 1990. EACMV: Honget al., 1993. EACMV-Ug: Zhou et al.,1997).
For each of the virus speciesencountered in the restriction mappingprocedures, and any viruses withrestriction maps that did not conform toknown patterns, full length DNA Agenomic components were cloned forsequence analysis. Clones were producedby PCR amplification with specificabutting, non-overlapping primers(Briddon et al., 1993). The sequences ofthese primers were obtained by cloningthe PCR products produced withuniversal primers into pGem T-Easyvectors (Promega). Sequences were

79
Diversity of African Cassava Mosaic Disease
determined by dideoxynucleotide chaintermination sequencing using the PCR-based BIG DYE kit (Perkin ElmerCetus) and specific internal primers(Gibco-BRL). Reaction products wereresolved on an ABI 377 automatedsequencer. Sequence information wasstored, assembled and analysed usingVersion 7 of the program library of theGenetics Computer Group.
Results
Table 1 summarizes the analysis of thesamples. About 13% of samples re-grew initially without symptoms,although 14% of these (2% of the total)did eventually show symptoms aftermore than 3 months. Less than 1%recovered after initially showingsymptoms. Table 2 shows the viruses
Table 1. Summary of analysis of cassava cuttings collected during field surveys. Sampled plants wereall showing symptoms when collected.a
Country Total no. Dead Re-grown Re-grown Healthy > Infected > DNA PCRof stems healthy infected infected healthy extract DNA-A
Tanzania 80 9 0 71 0 0 39 26
Kenya 45 15 0 30 0 2 14 5
Uganda 126 28 11 87 0 0 65 24
Zimbabwe 1 0 0 1 0 0 1 1
Malawi 23 19 2 2 0 0
Madagascar 193 1 39 153 1 2 30 18
Benin 96 46 7 43 5 2 35 15
Nigeria 68 21 27 20 7 0 25 12
Ghana 4 0 0 4 0 0 4 0
Cameroon 32 24 4 4 0 0 4 0
Totalb 671 163 (24) 90 (13) 418 (62) 13 (14) 6 (1) 217 (51) 101 (47)
a. “Healthy” indicates plants showing no disease symptoms after about 3 months of re-growth.“Healthy > infected” indicates plants that initially showed no symptoms on re-growth but developeddisease later; “Infected > healthy” indicates the converse. Other data: not sampled 39 (9%); yet tobe identified 68 (16%).
b. ( ) Percentage of totals; for DNA extract ( ) percentage of successful viral DNA extractions madefrom cassava mosaic disease samples; for PCR DNA-A ( ) percentage of DNA A productssuccessfully amplified from viral DNA.
Table 2. Summary of virus species isolated following restriction mapping using Mlu1, Dra1 and EcoR1on polymerase chain reaction products following the use of universal primers to amplifyBegomovirus genome component A.
Country Virus speciesa Total
ACMV EACMV EACMV-Ug SACMV Dual
Tanzania 3 5 1 8
Kenya 1 3 1 4
Uganda 6 2 15 3 23
Zimbabwe 1 1
Madagascar 12 4 16
Benin 12 3 15
Nigeria 8 8
Totalsb 42 (56) 17 (23) 15 (20) 1 [5] 75
a. ACMV, African cassava mosaic virus; EACMV, East African cassava mosaic virus; EACMV-Ug,EACMV-Uganda; SACMV, South African cassava mosaic virus; Dual, combinations.
b. ( ) percentage of total; [ ] included in total as single infections.

80
Whiteflies and Whitefly-borne Viruses in the Tropics
identified from the samples analysed:ACMV comprised 56%, EACMVcomprised 43% (of which 53% wereEACMV and 47% EACMV-Ug) and 7%were dual infections (of which allexcept one were combinations of ACMVand EACMV-Ug). Samples from WestAfrica comprised less than one-third ofthe total identified (31%) but of theseonly 13% were EACMV, while this virusaccounted for 56% of East Africasamples.
Full-length clones of the DNA Acomponents of CMD-associatedbegomoviruses were obtained fromisolates originating from Uganda andfrom Zimbabwe. The sequence of theclone originating from Uganda showsthis to be highly similar to thesequence of EACMV-Ug reported byZhou et al. (1997). The DNA Acomponent of this virus is essentiallyEACMV with an approximately 450 bpinsertion of an ACMV sequence in thecentre of the coat protein gene. Therestriction pattern of the virusoriginating from Zimbabwe was entirelynovel, with no counterpart in thepublished literature. Sequence analysisof this clone showed it to have mostsimilarity to SACMV (Berrie et al.,1998). The sequence published byBerrie et al. (1998) covers only the coatprotein gene of SACMV to which thesequence of the Zimbabwe clone shows86% similarity. In the absence of thefull sequence of SACMV we mustprovisionally conclude that theZimbabwe virus is distinct from, butmost closely related to, SACMV. Effortsare now under way to obtain the DNAB genomic components of these twoviruses for infectivity studies. Attemptsalso are continuing to produce full-length clones of EACMV and ACMVoriginating from Madagascar. Althoughamplification of these viruses has beenachieved, it has not yet proven possibleto clone them.
Discussion
The results obtained during thisscreening project indicate that a PCR/RFLP-based approach is ableconsistently to identify the virusesassociated with CMD and identifyspecies or strains that differsignificantly from those previouslyreported. It is unclear whether aprotocol based solely upon PCRamplification with diagnostic primersfor each virus species/strain couldachieve the same.
It is surprising, based on experienceof related viruses, to find the geneticstability which is evident from the verylow variability detected both in thesequence analysis of clones and fromthe RFLP analysis. However, thisstability makes RFLP analysis aparticularly useful diagnostic tool forthese viruses. This method also enablesplants that are dual-infected to becorrectly diagnosed because the overlaidRFLP patterns are evident. Possibly themajor drawback to the technique is thathigh-quality samples of nucleic acidsare essential but these may be difficultto extract from cassava. This was mostevident in the case of the dried cassavaleaf samples sent to JIC from Africa,from which no successful PCRamplification was achieved. The reasonfor this failure remains unclear. It ispossible that sample degradation occursunder tropical (hot and humid)conditions; fungal growth wasparticularly evident on some of the leafsamples. This problem has beenencountered previously where evensamples dried under optimal conditionswere not suitable for DNA extractionand subsequent begomovirus PCRamplification. However, the cassava leafsamples collected from the glasshouseand dried immediately yieldedamplification products more than 1 yearlater, after storage at room temperaturebut in a dry atmosphere.

81
Diversity of African Cassava Mosaic Disease
It is clear from the recent reports ofnew strains/species of begomovirusesidentified in association with CMD thatthe diversity of these agents is greaterthan initially expected. The proceduresutilized for the analysis of the diversityof begomoviruses infecting cassavaacross Africa were devised to be allencompassing. The use of so-called“universal primers”, designed withdegeneracy so as to amplify all whitefly-transmitted begomoviruses, ensuresthat all possible begomoviruses in aparticular sample are amplified. Theuse of specific primers in an analysislike this would risk missing possiblenew viruses, or mutants of viruses,which have been described previously.The universal primers used (Briddonand Markham, 1994) are wellcharacterized and have been able thusfar to amplify all begomoviruses againstwhich they have been tested.
The use of RFLP analysis on PCR-amplified DNA allows several diagnosticrestriction enzymes to be used on asingle sample to identify a virus. Theinitial identification of a virus species/strain by a particular restrictionpattern was subsequently confirmed bysequence analysis of a full-lengthclone. Full characterization of a clonerequires infectivity studies on cassava(Briddon et al., 1998). Our work hasshown that the use of alternate hosts,such as Nicotiana benthamiana Domin(Solanaceae), can lead to selection ofmutants with atypical biologicalcharacters (Liu et al., 1997) and thus isnot appropriate for diagnostic studies.
Acknowledgements
The authors wish to thank the DanishInternational Development Agency forproviding funds for this project, and allthe participants of the SystemwideProgram on Integrated PestManagement whose enthusiasm and
determination guaranteed its initiation.We also wish to thank all members ofthe project team who sent materialduring their regional surveys; withouttheir help and endeavours, the projectwould not have been possible. We alsothank Dr. Richard Gibson and Mr. IanRobertson for sending extra samplesfor inclusion. The work was alsosupported by the John Innes Centre,which is grant-aided by theBiotechnology and Biological SciencesResearch Council. The viruses wereimported and held under the authorityof the Ministry of Agriculture Fisheriesand Food, under the Plant Health(Great Britain) Order 1993, LicenceNumbers PHL11/2557(4/1998) andPHL/2300(6/1998).
References
Berrie, L. C.; Palmer, K. E.; Rybicki, E. P.;Rey, M. E. C. 1998. Molecularcharacterization of a distinct SouthAfrican cassava infectinggeminivirus. Arch. Virol. 143:2253-2260.
Briddon, R. W.; Markham, P. G. 1994.Universal primers for the PCRamplification of dicot-infectinggeminiviruses. Mol. Biotechnol.1:202-205.
Briddon, R. W.; Markham, P. G. 1995.Geminiviridae. In: Murphy, F.A.;Fauquet, C. M.; Bishop, D. H. L.;Ghabrial, S. A.; Jarvis, A. W.;Martelli, G. P.; Mayo, M. A.;Summers, M. D. (eds.). Virustaxonomy: Sixth report of theInternational Committee onTaxonomy of Viruses. Springer,Vienna, AT. p. 158-165.
Briddon, R. W.; Prescott, A. G.; Lunness,P.; Chamberlin, L. C. L.; Markham,P. 1993. Rapid production of full-length, infectious geminivirusclones by abutting primer PCR(AbP-PCR). J. Virol. Methods 43:7-20.

82
Whiteflies and Whitefly-borne Viruses in the Tropics
Briddon, R. W.; Liu, S.; Pinner, M. S.;Markham, P. G. 1998. Infectivity ofAfrican cassava mosaic virus clonesto cassava by biolistic inoculation.Arch. Virol. 143:1487-1492.
Calvert, L. A.; Thresh, J. M. 2002. Theviruses and virus diseases ofcassava. In: Hillocks, R. J.; Thresh,J. M.; Bellotti, A. C. (eds.). Cassava:Biology, production and utilization.CAB International, Wallingford, GB.p. 237-260.
Harrison, B. D.; Robinson, D. J. 1988.Molecular variation in vector-borneplant viruses: Epidemiologicalsignificance. Philos. Trans. R. Soc.,Lond., B, Biol. Sci. 321:447-462.
Harrison, B. D.; Zhou, X.; Otim-Nape, G.W.; Liu, Y.; Robinson, D. J. 1997.Role of a novel type of doubleinfection in the geminivirus-inducedepidemic of severe cassava mosaicin Uganda. Ann. Appl. Biol.131:437-448.
Hong, Y. G.; Robinson, D. J.; Harrison, B.D. 1993. Nucleotide sequenceevidence for the occurrence of threedistinct whitefly-transmittedgeminiviruses in cassava. J. Gen.Virol. 74:2437-2443.
Legg, J. P. 1999. Emergence, spread andstrategies for controlling thepandemic of cassava mosaic virusdisease in east and central Africa.Crop Prot. 18:627-637.
Liu, S.; Bedford, I. D.; Briddon, R. W.;Markham, P. 1997. Efficientwhitefly transmission of Africancassava mosaic geminivirusrequires sequences from bothgenomic components. J. Gen. Virol.78:79-94.
Morris, B.; Coates, L.; Lowe, S.;Richardson, K.; Eddy, P. 1990.Nucleotide sequence of theinfectious cloned DNA componentsof African cassava mosaic virus(Nigerian strain). Nucleic Acids Res.18:197-198.
Otim-Nape, G. W.; Bua, A.; Thresh, J. M.;Baguma, Y.; Ogwal, S.; Semakula,G. N.; Acola, G.; Byabakama, B.;Martin, A. 1997. Cassava mosaicvirus disease in Uganda: Thecurrent pandemic and approachesto control. Natural ResourcesInstitute (NRI), Chatham, GB. 65 p.
Pita, J. S.; Fondong, V. N.; Sangare, A.;Otim-Nape, G. W.; Ogwal, S.;Fauquet, C. 2001. Recombination,pseudorecombination andsynergism of geminiviruses aredeterminant keys to the epidemic ofsevere cassava mosaic disease inUganda. J. Gen. Virol. 82:655-665.
Stanley, J.; Gay, M. G. 1983. Nucleotidesequence of cassava latent virusDNA. Nature 301:260-262.
Thresh, J. M.; Otim-Nape, G. W.; Legg, J.P.; Fargette, D. 1997. Africancassava mosaic disease: Themagnitude of the problem? Afr. J.Root Tuber Crops 2:13-19.
Warburg, O. 1894. Die KulturpflanzenUsambaras. Mitt. Dtsch. Schutz.7:131.
Zhou, X.; Liu, Y.; Calvert, L.; Muñoz, C.;Otim-Nape, G. W.; Robinson, D. J.;Harrison, B. D. 1997. Evidence thatDNA-A of a geminivirus associatedwith severe cassava mosaic diseasein Uganda has arisen byinterspecific recombination. J. Gen.Virol. 78:2101-2111.

83
Sweetpotato Virus Disease in East Africa
CHAPTER 1.12
Serological Analysis of SweetpotatoesAffected by Sweetpotato Virus Diseasein East Africa
Valente Aritua*, Richard Gibson** andJosef Vetten***
Introduction
Infection of sweetpotato (Ipomoeabatatas [L.] Lam.) by viruses is a majorcause of yield reduction worldwide. Atleast 13 viruses are reported to infectsweetpotato naturally (Moyer andLarsen, 1991). Most of them are insect-transmitted, mainly by whitefly oraphid species. In East Africa,symptoms of sweetpotato virus disease(SPVD) were first reported onsweetpotato in 1945 (Hansford, 1945).However, the presence of viruses wasnot demonstrated until 1957 whenSheffield (1957) associated two viruseswith sweetpotato plants having virus-like symptoms, virus A being aphid-transmitted and virus B being whitefly-transmitted. Virus A was lateridentified as Sweetpotato feathery mottlevirus (SPFMV) but the identity of thewhitefly-transmitted virus remainedunclear. Subsequent efforts todetermine the range of virusesoccurring in East African sweetpotatocrops confirmed SPFMV as the mostfrequently found virus but alsorevealed the presence of Sweetpotatomild mottle virus (SPMMV), Sweetpotatolatent virus (SPLV) and Sweetpotatochlorotic fleck virus (SPCFV) (Carey
et al., 1998). SPFMV has a worldwidedistribution, is readily spread byaphids such as Myzus persicae (Sulzer)in a non-persistent manner butprobably is not spread by seedtransmission (Cadena-Hinojosa et al.,1981a). Various strains of SPFMV havebeen differentiated, mostly bysymptoms produced on certainsweetpotato cultivars. In USA, twostrains are recognized—the russetcrack strain or internal cork strain,which causes internal necrosis of thestorage roots of some cultivars, and thecommon strain, which causes no suchsymptoms. In East Africa, SPFMV-infected sweetpotato plants typicallyexhibit no symptoms on either roots orfoliage. SPMMV is considered to bewhitefly-borne (Hollings and Stone,1976).
The whitefly-transmittedcomponent virus of SPVD has beennamed as Sweetpotato chlorotic stuntvirus (SPCSV) largely on the basis ofthe symptoms of West African isolatesexpressed in the indicator plantIpomoea setosa Ker Gawl. (Schaefersand Terry, 1976). SPCSV issynonymous (Gibson et al., 1998) withSweetpotato sunken vein virus (Cohenet al., 1992) and SPVD-associatedclosterovirus. An East African strain ofSPCSV (SPCSV-SEA) was identifiedusing monoclonal antibodies (MABs)prepared against an Israeli isolate ofSPCSV as well as against bacterially
* Makerere University, Kampala, Uganda.** Natural Resources Institute (NRI),
University of Greenwich, ChathamMaritime, Kent, UK.
*** Biologische Bundesanstalt für Land undForstwirtschaft, Braunschweig, Germany.

84
Whiteflies and Whitefly-borne Viruses in the Tropics
expressed coat protein of a Kenyanisolate (Hoyer et al., 1996; Gibson etal., 1998). Subsequent analyses usingenzyme-linked immunosorbent assays(ELISA) led to the description of twodistinct serotypes: SEA1 as originallydescribed from Kenya, and SEA2 (Alicaiet al., 1999). In East Africa, SPCSVcauses only moderate stunting andyellowing or purpling of the lower andmiddle leaves when it occurs alone(Gibson et al., 1998). Co-infection ofSPFMV and SPCSV is very commonand the severe symptoms of SPVDresult from a synergistic interactionbetween SPCSV and SPFMV. SPVD isthe most important disease ofsweetpotato in Africa (Geddes, 1990).Depending on the genotype, SPVDsymptoms comprise leaf strapping,mottling, vein clearing, puckering andstunting of sweetpotato plants(Schaefers and Terry, 1976).
The respective distributions ofSPMMV, SPCSV and its synergisticpartner, SPFMV, are still poorlyunderstood in the major sweetpotatogrowing regions of East Africa,although it has long been known thatthese viruses can cause large yieldlosses (Mukiibi, 1977). The growingimportance of sweetpotato for foodsecurity in Africa has increasedinternational exchange of sweetpotatogermplasm and thus increased theneed to identify and establish thedistribution of the common virusesoccurring on the crop. Suchinformation is an importantprerequisite for the development andappropriate release of cultivarsresistant to the prevailing viruses/diseases.
The following work investigates thedistributions of SPMMV, SPCSV and itsserotypes, and SPFMV in Uganda,Kenya and Tanzania.
Collection and Analysis ofSamples
Project partners carried out fieldsurveys according to a commonprotocol. Additional information on thetiming and coverage of the surveys aregiven in the respective country papersin the present volume (Chapters 1.6,1.7 and 1.8). The authors analysedsamples collected by project partnersas follows: those from Uganda weretested fresh at Namulonge ResearchInstitute; dried samples from Kenyaand Tanzania were analysed at theBiologische Bundesanstalt für Landund Forstwirtschaft (BBA),Braunschweig, Germany. The sampleswere analysed using polyclonal antiseraor monoclonal antibodies to SPCSV,SPFMV and SPMMV using tripleantibody sandwich (TAS), doubleantibody sandwich (DAS), ornitrocellulose membrane (NCM) ELISA.
More than 200 sweetpotato leafsamples were collected from plantsshowing possible symptoms of viralinfection. Field symptoms observedconsisted of leaf strapping, yellowing,purpling, mottling, vein clearing andstunting of sweetpotato plants. Sinceincidence varied among and within thethree countries, different numbers ofsamples were collected from eachcountry and target area.
Reactions of the DiseasedSamples
Table 1 shows the numbers of samplesby country and target area that testedpositive for SPFMV, SPSCV-SEA and/or SPMMV. Both SPFMV and SPCSVoccurred throughout the sampledareas. Overall, SPFMV was mostfrequently detected, in 151 out of the243 samples. It was most frequently
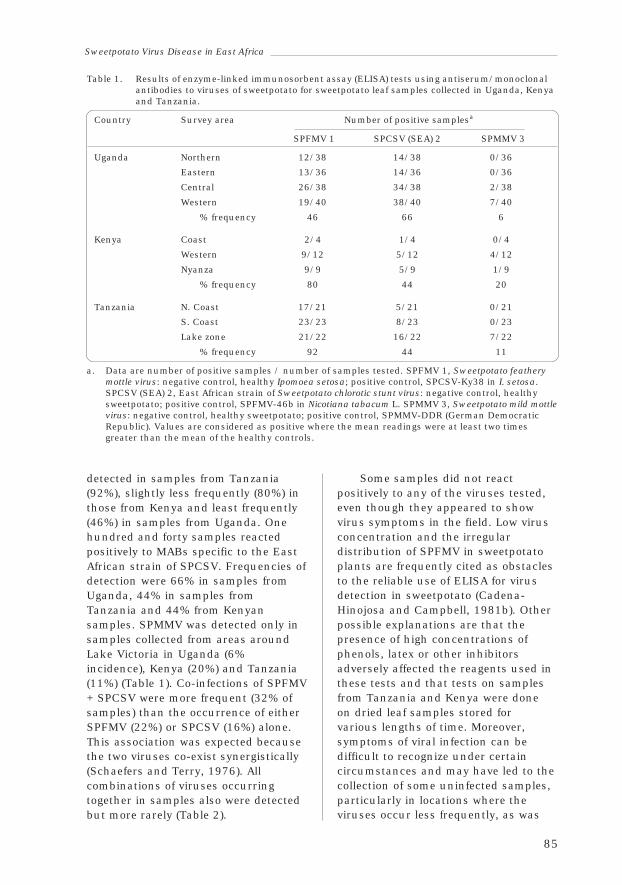
85
Sweetpotato Virus Disease in East Africa
detected in samples from Tanzania(92%), slightly less frequently (80%) inthose from Kenya and least frequently(46%) in samples from Uganda. Onehundred and forty samples reactedpositively to MABs specific to the EastAfrican strain of SPCSV. Frequencies ofdetection were 66% in samples fromUganda, 44% in samples fromTanzania and 44% from Kenyansamples. SPMMV was detected only insamples collected from areas aroundLake Victoria in Uganda (6%incidence), Kenya (20%) and Tanzania(11%) (Table 1). Co-infections of SPFMV+ SPCSV were more frequent (32% ofsamples) than the occurrence of eitherSPFMV (22%) or SPCSV (16%) alone.This association was expected becausethe two viruses co-exist synergistically(Schaefers and Terry, 1976). Allcombinations of viruses occurringtogether in samples also were detectedbut more rarely (Table 2).
Some samples did not reactpositively to any of the viruses tested,even though they appeared to showvirus symptoms in the field. Low virusconcentration and the irregulardistribution of SPFMV in sweetpotatoplants are frequently cited as obstaclesto the reliable use of ELISA for virusdetection in sweetpotato (Cadena-Hinojosa and Campbell, 1981b). Otherpossible explanations are that thepresence of high concentrations ofphenols, latex or other inhibitorsadversely affected the reagents used inthese tests and that tests on samplesfrom Tanzania and Kenya were doneon dried leaf samples stored forvarious lengths of time. Moreover,symptoms of viral infection can bedifficult to recognize under certaincircumstances and may have led to thecollection of some uninfected samples,particularly in locations where theviruses occur less frequently, as was
Table 1. Results of enzyme-linked immunosorbent assay (ELISA) tests using antiserum/monoclonalantibodies to viruses of sweetpotato for sweetpotato leaf samples collected in Uganda, Kenyaand Tanzania.
Country Survey area Number of positive samplesa
SPFMV 1 SPCSV (SEA) 2 SPMMV 3
Uganda Northern 12/38 14/38 0/36
Eastern 13/36 14/36 0/36
Central 26/38 34/38 2/38
Western 19/40 38/40 7/40
% frequency 46 66 6
Kenya Coast 2/4 1/4 0/4
Western 9/12 5/12 4/12
Nyanza 9/9 5/9 1/9
% frequency 80 44 20
Tanzania N. Coast 17/21 5/21 0/21
S. Coast 23/23 8/23 0/23
Lake zone 21/22 16/22 7/22
% frequency 92 44 11
a. Data are number of positive samples / number of samples tested. SPFMV 1, Sweetpotato featherymottle virus: negative control, healthy Ipomoea setosa; positive control, SPCSV-Ky38 in I. setosa.SPCSV (SEA) 2, East African strain of Sweetpotato chlorotic stunt virus: negative control, healthysweetpotato; positive control, SPFMV-46b in Nicotiana tabacum L. SPMMV 3, Sweetpotato mild mottlevirus: negative control, healthy sweetpotato; positive control, SPMMV-DDR (German DemocraticRepublic). Values are considered as positive where the mean readings were at least two timesgreater than the mean of the healthy controls.

86
Whiteflies and Whitefly-borne Viruses in the Tropics
the case in northern and easternregions of Uganda. Alternatively, sinceall samples tested were selected on thebasis of apparent virus symptoms, thelower rates of detection of virus insamples from more easterly areas mayindicate the presence there of virusesother than SPCSV, SPFMV or SPMMV(i.e., viruses for which no tests werecarried out).
In Uganda, the presence of twoserotypes of SPCSV was confirmedusing two MABs (Hoyer et al., 1996),
both serotypes reacting with one(MAB 2G8) but only SEA2 reacting withthe other (MAB 6D12) (Alicai et al.,1999). Serotype SEA2 was detected in66 of the 96 SPCSV-positive samples,while SEA1 was detected in 30 of theSPCSV positive samples. SerotypeSEA2 was found in all districts exceptfor Tororo, Masindi and Iganga,although it was found most frequentlyin the central and western surveyareas. Serotype SEA1 was distributedmore evenly across the country and, incontrast to the SEA2, occurred more
Table 2. Single and multiple occurrences of viruses of sweetpotato in leaf samples from three EastAfrican countries.
Country Nothing Sweetpotato virusesa
detectedSPFMV SPCSV SPMMV SPFMV + SPFMV + SPCSV + SPFMV +alone alone alone SPCSV SPMMV SPMMV SPCSV +
SPMMV
Uganda 36 15 40 1 54 0 1 5
Tanzania 0 30 0 0 29 1 0 6
Kenya 2 9 0 1 9 2 1 1
Total 38 54 40 2 92 3 2 12
% 16 22 16 1 38 1 1 5
a. SPFMV, Sweetpotato feathery mottle virus; SPCSV, Sweetpotato chlorotic stunt virus and SPMMV,Sweetpotato mild mottle virus.
Table 3. Distribution of two Sweetpotato chlorotic stunt virus (SPCSV) serotypes in sweetpotato leafsamples from 12 districts of Uganda.
Target area District No. positive samples SPCSV serotype
MAB 2G8 MAB 6D12 SEA2 SEA1
Northern Apac 4 1 3 1
Lira 4 1 3 1
masindi 5 5 0 5
Total 6 7
Eastern Tororo 1 1 0 1
Palisa 8 6 2 6
Iganga 5 5 0 5
Total 2 12
Central Luwero 11 0 11 0
Mpigi 9 2 7 2
Mukono 11 4 7 4
Total 25 6
Western Masaka 13 3 10 3
Rakai 12 0 12 0
Mubende 13 2 11 2
Total 33 5

87
Sweetpotato Virus Disease in East Africa
frequently in the northern and easternsurvey areas (Table 3). These resultsconfirm the heterogeneity of SPCSV, atleast at the serotype level. Furtherstudies would therefore be useful toinvestigate the diversity of this group ofviruses and clarify any implications fordisease management.
Conclusions
SPCSV is a common whitefly-bornevirus in sweetpotato throughout EastAfrica, often occurring together withSPFMV, with which it actssynergistically to produce the severedisease SPVD. In contrast, SPMMV islargely restricted to the Lake Victoriaregion of East Africa.
Differences in the distribution oftwo SPCSV serotypes have beenconfirmed. Both SEA1 and SEA2 arewidely distributed throughout Ugandabut SEA1 is more prevalent in northernand eastern Uganda and SEA2 is moreprevalent in central and westernUganda. These areas correspond withareas of lesser and greater whiteflyabundance and slower and faster ratesof spread (Aritua et al., 1998) butwhether this relationship is causal isunclear.
Acknowledgements
The technical assistance of Mr. TitusAlicai during sample analysis isgratefully acknowledged. The CropProtection Programme of the UKDepartment for InternationalDevelopment funded Dr. RichardGibson’s work.
References
Alicai, T.; Fenby, N. S.; Gibson, R. W.;Adipala, E.; Vetten, H. J.; Foster, G.D.; Seal, S. E. 1999. Occurrence oftwo serotypes of Sweetpotatochlorotic stunt virus in East Africaand their associated differences incoat protein and HSP70 homologuegene sequences. Plant Pathol.48:718-726.
Aritua, V.; Adipala, E.; Carey, E. E.;Gibson, R. W. 1998. The incidenceof sweetpotato virus disease andvirus resistance of sweetpotatogrown in Uganda. Ann. Appl. Biol.132:399-411.
Cadena-Hinojosa, M. A.; Campbell, R. N.1981a. Characterisation of fouraphid transmitted sweetpotatoviruses. Phytopathology 71:1086-1089.
Cadena-Hinojosa, M. A.; Campbell, R. N.1981b. Serological detection offeathery mottle virus strains insweetpotatoes and Ipomoeaincarnata. Plant Dis. 65:412-414.
Carey, E. E.; Mwanga, R. O. M.; Fuentes,S.; Kasule, S.; Macharia, C.;Gichuki, S. T.; Gibson, R. W. 1998.Sweetpotato viruses in Uganda andKenya: Results of a survey. In:Procs. Sixth Triennial Symposiumof the International Society ofTropical Root Crops-Africa Branch(ISTRC-AB), 22-28 October 1995,Lilongwe, MW. p. 457-461.
Cohen, J.; Franck, A.; Vetten, H. J.;Lesemann, D. E.; Loebenstein, G.1992. Purification and properties ofclosterovirus-like particlesassociated with a whitefly-transmitted disease of sweetpotato.Ann. Appl. Biol. 121:257-268.
Geddes, A. M. 1990. The relativeimportance of crop pests in sub-Saharan Africa. Bulletin no. 36,Natural Resources Institute (NRI),Chatham, GB.

88
Whiteflies and Whitefly-borne Viruses in the Tropics
Gibson, R. W.; Mpembe, I.; Alicai, T.;Carey, E. E.; Mwanga, R. O. M.;Seal, S. E.; Vetten, H. J. 1998.Symptoms, aetiology and serologicalanalysis of sweetpotato virusdisease in Uganda. Plant Pathol.47:95-102.
Hansford, C. G. 1945. Ugandan plantdiseases. Part IV. Diseases ofsweetpotato. East Afr. Agric. J.10:126-127.
Hollings, M.; Stone, O. M. 1976.Purification and properties ofsweetpotato mild mottle, a whitefly-borne virus from sweetpotato(Ipomoea batatas) in East Africa.Ann. Appl. Biol. 82:511-528.
Hoyer, U.; Maiss, E.; Jekmann, W.;Lessemann, D. E.; Vetten, H. J.1996. Identification of coat proteingene of sweetpotato sunken veinclosterovirus isolate from Kenyaand evidence for serologicalrelationship among geographicallydiverse closterovirus isolates fromsweetpotato. Phytopathology86:744-750.
Moyer, J. W.; Larsen, R. C. 1991.Management of insect vectors ofviruses infecting sweetpotato. In:Jansson, R. K.; Raman, K. V. (eds.).Sweetpotato pest management: Aglobal perspective. Westview Press,Denver, Colorado, USA. p. 341-358.
Mukiibi, J. 1977. Effects of mosaic viruson the yields of sweetpotato inUganda. Procs. Fourth InternationalSymposium on Tropical Root Crops.International DevelopmentResearch Centre (IDRC), Ottawa,CA. p. 169-170.
Schaefers, G. A.; Terry, E. R. 1976. Insecttransmission of sweetpotatodiseases in Nigeria. Plant Pathol.66:642-645.
Sheffield, F. M.L. 1957. Virus diseases ofsweetpotato in East Africa. 1.Identification of viruses and theirinsect vectors. Phytopathology47:582-587.

89
Factors Associated with Damage
CHAPTER 1.13
Factors Associated with Damage toSweetpotato Crops by SweetpotatoVirus Disease
Richard Gibson*, Valente Aritua** andSimon Jeremiah***
Introduction
Sweetpotato virus disease (SPVD) is themost destructive disease of sweetpotato(Ipomoea batatas [L]) in Africa (Geddes,1990). It is caused by a combinedinfection of the aphid-borneSweetpotato feathery mottle virus(SPFMV) and the whitefly-borneSweetpotato chlorotic stunt virus(SPCSV) (Schaefers and Terry, 1976;Gibson et al., 1998b). Infection withSPFMV alone causes no obvioussymptoms in African sweetpotatovarieties (Gibson et al., 1997), whileinfection with SPCSV alone causes onlymoderate stunting, yellowing orpurpling of middle and lower leavesand some yield loss (Gibson et al.,1998b). By contrast, SPVD causessevere plant stunting and small,distorted leaves with either chloroticmosaic or vein clearing (Schaefers andTerry, 1976). The yield of affectedplants generally is reduced by morethan 50% (Mukiibi, 1977; Hahn, 1979;Ngeve and Boukamp, 1991). It appearsthat SPCSV has a synergistic effect onSPFMV (Rossel and Thottappilly, 1987;Aritua et al., 1998a): field plants that
are infected with SPCSV quicklydevelop SPVD, either through anactivation of a latent SPFMV infectionor through increased susceptibility toaphid-borne spread of the virus. In thismanner, infection with SPCSV is thetrigger for SPVD and damage by thedisease has long been linked with highpopulations of its vector, the whiteflyBemisia tabaci (Gennadius) (Sheffield,1957).
SPCSV is transmitted semi-persistently by B. tabaci; feeds lasting afew hours are required for both efficientacquisition and inoculation (Cohen etal., 1992). B. tabaci that colonizesweetpotato in Africa cannot readilycolonize cassava (Manihot esculentaCrantz) and vice-versa (Burban et al.,1992; Legg, 1996). As a result, theremay be no direct link between whiteflynumbers on sweetpotato and whiteflynumbers on cassava, or betweentransmission of SPCSV and of Africancassava mosaic virus, even wheresweetpotato and cassava areintercropped. Thus, the epidemic ofcassava mosaic disease and B. tabaci incassava crops in East Africa seems tohave had little effect on the incidenceof SPVD. This is particularly fortunatebecause sweetpotato often provides themost readily available alternative cropto cassava.
Sweetpotato is an important cropfor subsistence farmers throughout
* Natural Resources Institute (NRI),University of Greenwich, ChathamMaritime, Kent, UK.
** Makerere University, Kampala, Uganda.*** Root and Tubers Programme of the Lake
Zone Agricultural Research andDevelopment Institute (LZARDI), Ukiriguru,Mwanza, Tanzania.

90
Whiteflies and Whitefly-borne Viruses in the Tropics
Uganda, generally being growncontinuously throughout the year.Planting occurs whenever soil moistureis adequate and so is concentratedduring, although not exclusively limitedto, the rains, which in most areas ofUganda occur from March to June andfrom September to November. Cropsare usually maintained for at least6 months, cropping frequently beingextended for up to 1 year by piecemealharvesting, using a stick to tease outindividual mature tubers (Bashaasha etal., 1995).
SPVD has been reported to beprevalent at moderate altitudes, 50 to600 m above sea level, in East Africa(Sheffield, 1953). However, in Uganda,it was rare in the eastern districts ofSoroti, Tororo and Busia but reacheddamaging levels of incidence in centralMpigi District, as well as in thesouthern districts of Masaka andRakai, even though all locations wereat a similar mid-altitude (Aritua et al.,1998b). The main aim of this projecttherefore was to understand theunderlying causes of the differentincidences, so as to provide a soundbase from which to developmanagement practices. Recent work(Gibson et al., 1997; Aritua et al.,1998b) identified three factors as beingassociated with low SPVD incidence:
(1) Whiteflies were rare on sweetpotatoin some localities (for example, theeastern districts of Uganda);
(2) The predominant sweetpotatovariety is resistant; and
(3) Farmers select planting materialfrom symptomless parents.
Our work therefore focused onlinks between SPVD incidence and (a)whitefly numbers, (b) varietalresistance and (c) farmers’ plant healthmanagement practices.
Methods and Results
Monthly surveys of whiteflies andSPVD were carried out at 10 farmers’fields around each of six towns inUganda (Soroti, Busia, Iganga,Namulonge, Kanoni and Masaka), fromJanuary 1998 to April 1999. All thesites are at about the same altitudeand lie roughly along a south-east/north-west axis. Sweetpotato fields,4 to 8 months old, were chosen whiletravelling along a rural road or patharound each town, stopping atintervals of about 1 km or at the firstfield observed thereafter, differentplantings being sampled each month.Whitefly numbers in each field wereassessed by counting the number ofwhitefly adults observed while turningover leaves for a 1-minute period. Tendifferent parts of each crop weresampled in this manner. A moredetailed account of the samplingmethod is provided in Aritua et al.(1998b). SPVD was assessed bycounting the proportion of affectedplants among 30 plants selected atrandom along a V-shaped trackthrough each field.
Whiteflies were relatively few atSoroti and Busia except in February1998 (Figure 1) and, in accord withthis, SPVD was also rare there(Figure 2). Similarly, there were slightlymore of both whiteflies and SPVD atIganga. Although farmers’ fields atNamulonge, Kanoni and Masaka allhad relatively large and similarnumbers of whiteflies, SPVD atNamulonge averaged only 2% and atMasaka, 6%, reaching 18% at Kanoni.Whitefly numbers were relatively highat all sites from January to April(inclusive). At those sites where SPVDwas common, its incidence remainedrelatively low until February, consistentwith there being a latent period ofabout 1 month (Gibson et al., 1998a)between infection by the whiteflies and

91
Factors Associated with Damage
appearance of SPVD. The incidence ofSPVD then remained relatively highuntil August, partly perhaps as a resultof continuing infection but probablyalso because of the survival of affectedplants.
From the point of view ofunderstanding the relationshipbetween vector dynamics and diseaseincidence over time, sampling farmers’fields had the disadvantages that theywere not necessarily planted with the
Mea
n w
hit
eflies
per
min
ute
Date of observation
Soroti Busia Iganga Namulonge KanoniMasaka
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar Apr
35
30
25
20
15
10
5
0
Figure 1. Mean whitefly numbers in six districts of Uganda (January 1998-April 1999). Each datapoint represents the mean for 10 fields.
Mea
n S
PV
D in
ciden
ce (%
)
25
20
15
10
5
0
Date of observation
Soroti Busia Iganga Namulonge KanoniMasaka
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar Apr
Figure 2. Sweetpotato virus disease (SPVD) incidence in farmers’ fields in six districts of Uganda(January 1998-April 1999).

92
Whiteflies and Whitefly-borne Viruses in the Tropics
same cultivars and that the plantingmaterial already might have beeninfected and, to a varying degree, withSPVD. In order to eliminate thesevariables, three on-farm trials wereplanted at each of the three mostcontrasting sites, Soroti, Namulongeand Kanoni, using planting materialfrom Namulonge Agricultural andAnimal Production Research Institute(NAARI) farm, with one series ofplantings in the second rains of 1997(Aritua et al., 1999) and another in thesecond rains of 1998. Whiteflynumbers and SPVD incidence weremonitored monthly, from January toJune, using the same procedures as inthe surveys of farmers’ fields. In bothseries, the results were very similar tothose obtained by monitoring farmers’fields:
(1) No SPVD and few whiteflies (resultsnot shown) were found at Soroti;
(2) Whiteflies were abundant at bothNamulonge and Kanoni (Table 1);
(3) But SPVD increased in incidencerapidly only at Kanoni (Figure 3);and
(4) There was no correlation betweenwhitefly numbers and final SPVDincidence (Table 1).
An index of the level of localinoculum for each site at Namulongeand Kanoni was also calculated from“area of each field of sweetpotato within100 m of each trial” × “incidence ofSPVD in it”¸ “distance to the trial plot”.Results are tabulated (Table 1) for thefirst series of trials in which there wasmore virus spread. The value for localinoculum correlated closely with SPVDincidence at both sites, suggesting thatthe greater spread of SPVD at Kanoniwas due to the greater incidence ofSPVD in nearby farmers’ fields.Examination of records of farmers’crops in the two localities also revealedthat a highly SPVD-resistant varietycalled New Kawogo predominated atNamulonge and the surroundingvillages and this may explain whySPVD was rare in the farmers’ fieldsthere. At Kanoni, more susceptiblevarieties predominated in farmers’fields (Aritua et al., 1999).
Links between SPVD incidence,crop data and farmers’ management
Table 1. Relationship between sweetpotato virus disease (SPVD) incidence and (1) whitefly counts,(2) local inoculum and (3) whitefly numbers × local inoculum.
Farm SPVD Whitefliesb Local Whiteflies ×incidencea inoculumc inoculum
Namulonge A 4.0 11.4 0 0
Namulonge B 1.5 13.2 100 1320.0
Namulonge C 0 14.0 120 1680.0
Kanoni D 18.5 10.5 1043 10951.5
Kanoni E 36.0 13.4 2298 30793.2
Kanoni F 23.5 9.4 1519 14278.6
Correlationd - -0.284 0.990 0.969
P - N.S. < 0.001 < 0.001
a. SPVD incidence (%): final assessment (June) in each on-farm trial.b. Whiteflies: mean monthly (January-June) whitefly counts from each on-farm trial.c. Local inoculum: index calculated from SPVD incidence × area of field ÷ distance from field for fields
within 100-m radius of trial plots.d. Correlation of SPVD incidence with data in corresponding column.
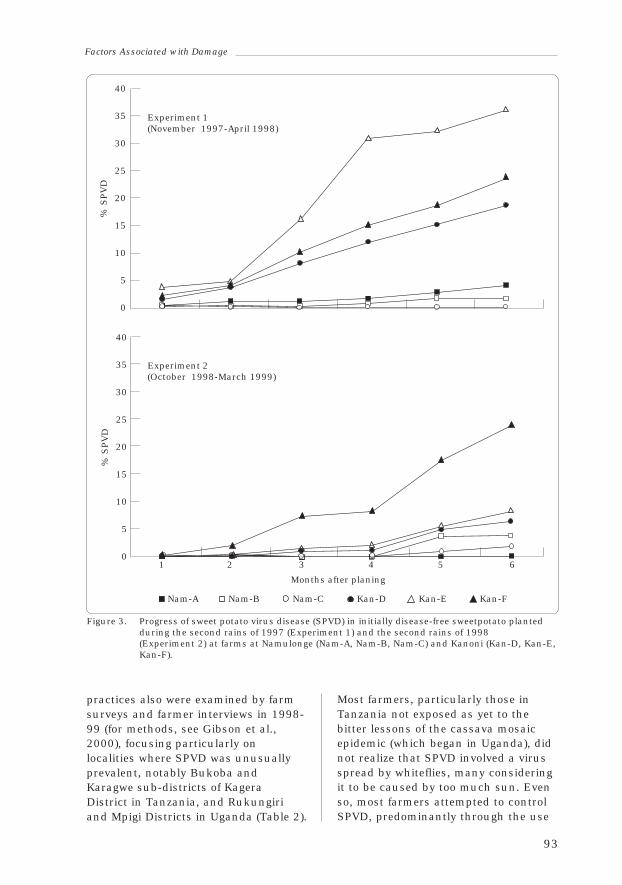
93
Factors Associated with Damage
practices also were examined by farmsurveys and farmer interviews in 1998-99 (for methods, see Gibson et al.,2000), focusing particularly onlocalities where SPVD was unusuallyprevalent, notably Bukoba andKaragwe sub-districts of KageraDistrict in Tanzania, and Rukungiriand Mpigi Districts in Uganda (Table 2).
Most farmers, particularly those inTanzania not exposed as yet to thebitter lessons of the cassava mosaicepidemic (which began in Uganda), didnot realize that SPVD involved a virusspread by whiteflies, many consideringit to be caused by too much sun. Evenso, most farmers attempted to controlSPVD, predominantly through the use
Nam-A Nam-B Nam-C Kan-D Kan-E Kan-F
% S
PV
D
40
35
30
25
20
15
10
5
0
% S
PV
D
40
35
30
25
20
15
10
5
0
Experiment 1(November 1997-April 1998)
Experiment 2(October 1998-March 1999)
Months after planing
1 2 3 4 5 6
Figure 3. Progress of sweet potato virus disease (SPVD) in initially disease-free sweetpotato plantedduring the second rains of 1997 (Experiment 1) and the second rains of 1998(Experiment 2) at farms at Namulonge (Nam-A, Nam-B, Nam-C) and Kanoni (Kan-D, Kan-E,Kan-F).

94
Whiteflies and Whitefly-borne Viruses in the Tropics
of plant health management tactics(Table 2). In particular, most farmerscarefully selected their plantingmaterial from unaffected parent plants.A few also removed (rogued) diseasedplants from young crops. However,farmers often planted new crops closeto old diseased crops, even thoughincidence of SPVD and the distancebetween crops were significantly(P < 0.05) and negatively correlated.Farmers also appeared to make noattempt to remove diseased plants fromold and abandoned crops even whenthey were close to newly planted fields.This is consistent with farmers lackingknowledge that SPVD is caused by aninsect-transmitted virus.
Although most farmers knew ofSPVD-resistant varieties, few rankedresistance as a valuable managementpractice (Table 2), apparently becausemost resistant varieties had a poorand/or late yield. Despite this, theyranked new varieties with superioryield characteristics and resistance astheir top requirement.
Conclusions
Our results confirm that SPVD variesconsiderably in incidence among sitesand that part of the variation inincidence is associated with differencesin the numbers of B. tabaci infestingthe crop. Thus, the explanation for therarity of SPVD in Soroti and Busiaseems to be that whiteflies are rarethroughout much of the year (Figure 1)but this still leaves unanswered thequestion of why fewer whiteflies shouldbe present in Soroti and Busia than inMpigi District. Our investigation wastoo limited to identify positively thereason(s) for this. However, a majordifference between Soroti and BusiaDistricts and Mpigi District is that thefirst rains in Soroti and Busia Districtsoccur later and the second rains earlierthan in Mpigi District. This leads to amore prolonged dry season between thesecond and the first rains in Soroti andBusia, during which time mostvegetation dries out, natural bushfiresare common and the resulting naturalvegetation predominantly consists oflow-growing bushes and grasslands.Rainfall being more evenly spread
Table 2. Importance to farmers of different measures that they already are using to managesweetpotato virus disease (SPVD) (adapted from Gibson et al., 2000).
Locality Percentage of farmersa giving the rank indicated No. of farmersto a control measure
Symptomless Roguing Resistant Controlling Interviewedplanting material cultivars SPVDb
1st 2nd 3rd 1st 2nd 3rd 1st 2nd 3rd
Tanzaniac
Bukoba 100 0 0 0 33 0 0 0 0 15 (75) 20
Karagwe 80 0 0 20 10 0 0 0 0 10 (36) 28
Uganda
Rukungiri 82 14 0 11 50 7 7 14 18 44 (88) 50
Mpigi 88 10 0 2 10 27 10 54 7 41 (82) 50
Soroti 100 0 0 0 17 17 0 17 0 6 (12) 50
Overall 87 9 0 7 28 13 6 25 9 116 (60) 198
a. Among those claiming to use specific measures to control SPVD.b. Number of farmers (in parentheses, percentage of farmers among those interviews) claiming to use
specific control measures for SPVD.c. Farmers in other districts in Tanzania were not questioned on this subject.

95
Factors Associated with Damage
throughout the year in Mpigi,vegetation continues to grow year-round and such fires rarely occur. As aresult, sweetpotato crops in this regionoften occur in locations sheltered bytall trees and other vegetation.
High incidences of SPVD wereassociated with:
(1) Large average numbers ofwhiteflies found on the sweetpotatocrop during the year;
(2) High peak whitefly populations,which occurred during the hot dryseason (January-March); and
(3) Relatively large amounts of SPVD-affected plants nearby.
The importance of local inoculumwas shown directly by our on-farm fieldtrials but was indicated also by surveydata showing that SPVD incidenceincreased as sweetpotato croppingintensity increased. A requirement forlocal inoculum is consistent withSPCSV being only semi-persistentlytransmitted and with B. tabaci being arelatively weak flier. Using pesticides toreduce the population of the whiteflyvector is highly unlikely to be a usefultactic in SPVD management since, evensetting aside environmental risks,human health concerns and thetendency of whiteflies to developresistance to insecticides, the crop hastoo low a value to justify theinvestment in insecticides. Choice ofappropriate varieties is unlikely to beeffective in reducing vector populationsbecause earlier work has not identifiedmajor differences in the numbers ofwhiteflies on different sweetpotatovarieties (Aritua et al., 1998a).However, local inoculum could bereduced by:
(1) Planting disease-free cuttings(already used by most farmers);
(2) Roguing plants that developsymptoms (used by a few farmersearly in the season);
(3) Removal of crop debris (both rootsand foliage as both re-grow readilyand are often diseased) from oldfields (not currently used);
(4) Separation of new crops from olddiseased ones (not currently used);and
(5) Increased use of resistant varieties(favoured only in localities whereacceptable resistant varieties wereavailable).
Our work also suggests that it maybe particularly important to apply planthealth management tactics during thehot dry season because this is whenthe whitefly vectors are mostabundant. However, it is important toemphasize that, although they mayseem obvious, none of these strategieshave been tested yet in field trials, toensure that they are effective andadoptable by farmers, and so cannot beofficially recommended. Furthermore,our farmer interviews indicated thatmost farmers do not have a soundunderstanding of the cause and sourceof SPVD. Some of these controlpractices might therefore appear tofarmers to be inappropriate.Consequently, both participatoryresearch and training programs will beessential if plant health managementstrategies are to be developed andwidely adopted.
Farmers in areas severely affectedby SPVD gave highest priority to theneed for superior, virus-resistantvarieties (Table 3). Our work showedthat resistant varieties are valuable notonly because they suffer less SPVDdamage but also because theirwidespread cultivation reducesincidence of SPVD in remaining stands

96
Whiteflies and Whitefly-borne Viruses in the Tropics
Table 3. Aspects of sweetpotato virus disease (SPVD) management on which farmers in severelyaffected localities would most like to receive assistance (adapted from Gibson et al., 2000).
Locality Percentage of farmersa giving a particular rank No. of farmersto the indicated management tactic
Superior, resistant Technical Chemical Requestingb Interviewed cultivars information control
1st 2nd 3rd 1st 2nd 3rd 1st 2nd 3rd
Bukoba 100 0 0 0 0 0 0 0 0 14 (70) 20
Rukungiri 60 21 0 23 6 2 17 15 0 48 (96) 50
Overall 69 16 0 18 5 2 13 11 0 62 (89) 70
a. Among those requesting specific measures to manage SPVD.b. Number of farmers (in parentheses, percentage among those interviewed) requesting assistance
with management of SPVD.
of susceptible varieties. Althoughfarmers also gave a low ranking to theuse of their currently availableresistant varieties (Table 2), this wasbecause most farmers perceived theirresistant landraces to have majorflaws. The higher yielding, resistantvarieties such as those selected atNAARI in Uganda and at theAgricultural Research Institute inUkiriguru, Tanzania, taking account offarmer preferences, should overcomethis obstacle.
Acknowledgements
We thank the many farmers whopatiently answered our questions andallowed their fields to be examined andused for research purposes. We thankthe Director of NAARI and the Staff ofthe Ugandan Potato Programme fortheir support and the use of NAARIfacilities. We thank Mr. I. Mpembe,Mr. R. P. Msabaha and Mr. J.Ndunguru for their collaborativeinputs.
References
Aritua, V.; Alicai, T.; Adipala, E.; Carey,E. E.; Gibson, R. W. 1998a. Aspectsof resistance to sweetpotato virusdisease in sweetpotato. Ann. Appl.Biol. 132:387-398.
Aritua, V.; Adipala, E.; Carey, E. E.;Gibson, R. W. 1998b. The incidenceof sweetpotato virus disease andvirus resistance of sweetpotatogrown in Uganda. Ann. Appl. Biol.132:399-411.
Aritua, V.; Legg, J. P.; Smit, N. E. J. M.;Gibson, R. W. 1999. Effect of localinoculum on the spread ofsweetpotato virus disease:Widespread cultivation of aresistant sweetpotato cultivar maybe protecting susceptible cultivars.Plant Pathol. 48:655-661.
Bashaasha, B.; Mwanga, R. O. M.; Ocittip’Obwoya, C.; Ewell, P. T. 1995.Sweetpotato in the farming and foodsystems of Uganda: A farm surveyreport. Centro Internacional de laPapa (CIP), sub-Saharan AfricaRegion, Nairobi, KE/NationalAgricultural Research Organisation(NARO), Kampala, UG. 63 p.
Burban, C.; Fishpool, L. D. C.; Fauquet,C.; Fargette, D.; Thouvenel, J. -C.1992. Host-associated biotypeswithin West African populations ofthe whitefly Bemisia tabaci(Genn.)(Hom., Aleyrodidae). J. Appl.Entomol. 113:416-423.
Cohen, J.; Franck, A.; Vetten, H. J.;Lesemann, D. E.; Loebenstein, G.1992. Purification and properties ofclosterovirus-like particlesassociated with a whitefly-transmitted disease of sweetpotato.Ann. Appl. Biol. 121:257-268.

97
Factors Associated with Damage
Geddes, A. M. 1990. The relativeimportance of crop pests in sub-Saharan Africa. Bulletin no. 36,Natural Resources Institute (NRI),Chatham, GB.
Gibson, R. W.; Mwanga, R. O. M.; Kasule,S.; Mpembe, I.; Carey, E. E. 1997.Apparent absence of viruses inmost symptomless field-grownsweetpotato in Uganda. Ann. Appl.Biol. 130:481-490.
Gibson, R. W.; Mpembe, I.; Alicai, T.;Carey, E. E.; Mwanga, R. O. M.;Seal, S. E.; Vetten, H. J. 1998a.Symptoms, aetiology andserological analysis of sweetpotatovirus disease in Uganda. PlantPathol. 47:95-102.
Gibson, R. W.; Jeremiah, S. C.; Msabaha,R. P. 1998b. Virus diseases ofsweetpotato and cassava in theLake Zone of Tanzania. Report no.2380, Natural Resources Institute(NRI), Chatham, GB.
Gibson, R. W.; Jeremiah, S. C.; Aritua, V.;Msabaha, R. P.; Mpembe, I.;Ndunguru, J. 2000. Sweetpotatovirus disease in sub-SaharanAfrica: Evidence that neglect ofseedlings in the traditional farmingsystem hinders the development ofsuperior resistant landraces.Phytopathology 148:441-447.
Hahn, S. K. 1979. Effects of sweetpotatovirus disease (SPVD) on growth andyield of sweetpotato. Exp. Agric.15:253-256.
Legg, J. P. 1996. Host-associated strainswithin Ugandan populations of thewhitefly Bemisia tabaci (Genn.)(Hom., Aleyrodidae). J. Appl.Entomol. 120:523-527.
Mukiibi, J. 1977. Effects of mosaic viruson the yields of sweetpotato inUganda. Procs. Fourth InternationalSymposium on Tropical Root Crops.International DevelopmentResearch Centre (IDRC), Ottawa,CA. p. 169-170.
Ngeve, J. M.; Boukamp, J. C. 1991.Effects of sweetpotato virus diseaseon the yield of sweetpotatogenotypes in Cameroon. Exp. Agric.27:221-225.
Rossel, H. W.; Thottappilly, G. 1987.Complex virus diseases ofsweetpotato. In: Exploration,maintenance and utilization ofsweetpotato genetic resources.Centro Internacional de la Papa(CIP), Lima, Peru. p. 291-302.
Schaefers, G. A.; Terry, E. R. 1976. Insecttransmission of sweetpotatodiseases in Nigeria. Plant Pathol.66:642-645.
Sheffield, F. M. L. 1953. Virus diseases ofsweetpotato in parts of Africa. Emp.J. Exp. Agric. 21:184-189.
Sheffield, F. M. L. 1957. Virus diseases ofsweetpotato in East Africa.1. Identification of viruses and theirinsect vectors. Phytopathology47:582-587.

98
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 1.14
Conclusions and RecommendationsJames Legg* and Braima James**
Introduction
The first phase of the Tropical WhiteflyIntegrated Pest Management (TWF-IPM)Project (see Introduction to thisvolume) provided a unique opportunityto assess whitefly and whitefly-bornevirus problems in a diverse range ofcropping systems across the tropicsand to obtain a broad range ofdiagnostic information as a basis forfurther concerted action. Sub-Project 4,Whiteflies as virus vectors in cassavaand sweetpotato in sub-Saharan Africa,had the ambitious target ofestablishing collaborative researchlinkages among more than 15 partnerinstitutions and implementingdiagnostic surveys in nine Africancountries. Activities began inSeptember 1997 and were completed inmid-1999. This chapter reviews theresults obtained, considers theexperiences gained in setting-up,implementing and completing thediagnostic phase of research, anddiscusses the implications forsubsequent work on enhancing themanagement of whitefly-borne diseases
of cassava (Manihot esculenta Crantz)and sweetpotato (Ipomoea batatas [L.]Lam.).
Increased BiologicalUnderstanding
Whitefly species andabundance
Field data collection and theexamination of whitefly nymphspecimens collected by project partnersduring surveys of cassava andsweetpotato in participating countriesprovided the most comprehensiveinformation ever obtained for whiteflieson these crops in Africa. More than2000 specimens were identified andfour species were recorded. The twospecies that have been reportedpreviously on cassava, Bemisia tabaci(Gennadius) and B. afer (Priesner andHosny), were the only ones occurringwidely on cassava and sweetpotato andwere recorded from most locations. Twospecies were identified that have notbeen reported previously from cassava.These were the greenhouse whiteflyTrialeurodes vaporariorum Westwood,identified from two locations in Ghanaand Nigeria, and T. ricini (Misra)identified from a single site in the LakeVictoria zone of Tanzania. Whilst theoccurrence of late instar nymphs ofthese two species does suggest
* International Institute of TropicalAgriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda, and Natural Resources Institute,University of Greenwich, Chatham Maritime,Kent, UK.
** IITA, Biological Control Center for Africa,Cotonou, Benin.

99
Conclusions and Recommendations
colonization of cassava, the very lowfrequency of occurrence means thatthey are of little significance.
B. tabaci and B. afer both occurredon cassava and sweetpotato, althoughB. afer was relatively less frequent onsweetpotato and in Uganda was notrecorded from this crop. There was animportant geographical zonation in therelative abundance of the two species(Figures 1-3). Whilst B. tabacipredominated in countries in theequatorial belt (Ghana, Benin, Nigeria,Cameroon, Uganda and Kenya), thetwo species occurred at similarfrequencies in Tanzania, to the south,and B. afer predominated in the mostsoutherly countries, Malawi andMadagascar. The reasons for thispattern of occurrence are not clear,particularly since no singleenvironmental factor appears tocorrelate consistently with the speciesratio. In Malawi, however, there was asignificant difference in the ratiobetween survey areas. In the twosurvey areas (1 and 2) along the shoreof Lake Malawi (475-700 m altitude),similar numbers of the two specieswere recorded; whilst in survey area 3,the central plateau (1000-1500 maltitude), 97% of specimens identifiedwere B. afer (Chapter 1.9, this volume).
The key difference between thelakeshore and plateau environments istemperature and it seems likely thatthe low temperatures that occur in themid-altitude environments of southernAfrica are unfavourable for B. tabaci.This possibility is supported bypublished information indicating thatthe development period of this speciesis significantly increased (Gerling et al.,1986) and fecundity significantlyreduced (Azab et al., 1971) attemperatures below 15 ºC. Moredetailed ecological studies andmolecular characterization of the twospecies will be required to clarify these
issues, particularly since the currentsurveys were conducted only at onetime in the year and therefore maynot be fully representative. It isimportant to highlight, however, thatB. afer has never been shown totransmit viruses. The distinctionbetween the two species thereforebecomes important if errors are not tobe made in describing relationshipsbetween adult whitefly populationsand the spread of viruses causingcassava mosaic disease (CMD) orsweetpotato virus disease (SPVD).
Geographical patterns of variationin abundance of cassava whiteflieswere partly confounded with thedifferent patterns of distribution ofthe two species (Figures 1-3).Although survey teams were trainedto differentiate the adult stages of thetwo species, it was not alwayspossible for them to do so duringcounting. Some patterns indistribution were apparent, however.In West Africa, where B. tabaci wasthe dominant species, abundance wasgenerally greater in the more humidrainforest and transition forestecozones. Conversely, populationswere smaller in the drier northernsavannah zones. This is a patternthat has been recognized before, bothin the Ecologically SustainableCassava Plant Protection (ESCaPP)surveys of 1993-94 (IITA, 1998) andthrough earlier studies of CMDepidemiology in Ivory Coast (Fauquetet al., 1987; 1988). In East Africa, twocentres of whitefly abundance wererecognized: the northern shoreline ofLake Victoria, in Uganda, and thenorthern Tanzania/southern Kenyacoast. As for West Africa, these arealso the areas in the region wherehumidity is greatest. It is notable,however, that abundance of B. tabaciin the “Lake Crescent” agro-ecologicalzone in Uganda, comprising thesouthern districts of Iganga, Mukono,

100
Whiteflies and Whitefly-borne Viruses in the Tropics
Whit
efly
abundan
ce0-1 1-3
3-9
9-2
7
27-8
1
CM
D i
nci
den
ce
20%
-30%
30%
-40%
40%
-50%
50%
-60%
60%
-70%
70%
-80%
Not
su
rvey
ed
200
0200
400
600
Kil
om
eter
s
Ghan
a
Ben
in
Nig
eria
Cam
eroon
B. a
fer
B. ta
ba
ci
Fig
ure
1.
Wh
itef
ly a
bu
ndan
ce, ca
ssava
mos
aic
dis
ease
(C
MD
) in
ciden
ce a
nd B
emis
ia t
aba
ci/B
. afe
r ra
tio
for
Su
b-P
roje
ct 4
cou
ntr
ies
in W
est
Afr
ica.

101
Conclusions and Recommendations
Whitefly abundance0-1
1-3
3-9
9-27
27-81
CMD incidence
20%-30%
30%-40%
40%-50%
50%-60%
60%-70%
Not surveyed
Uganda
Kenya
Tanzania
Malawi
B. afer
B. tabaci
100 0 100 200
Kilometers
Figure 2. Whitefly abundance, cassava mosaic disease (CMD) incidence and Bemisia tabaci/B. aferratio for Sub-Project 4 countries in East Africa.

102
Whiteflies and Whitefly-borne Viruses in the Tropics
Whitefly abundance0-1
1-3
3-9
9-27
27-81
CMD incidence
20%-30%
30%-40%
40%-50%
50%-60%
60%-70%
Not surveyed
Madagascar
B. afer
B. tabaci
100 0 100 200
Kilometers
300 400
Figure 3. Whitefly abundance, cassava mosaic disease (CMD) incidence and Bemisia tabaci/B. aferratio in Madagascar.

103
Conclusions and Recommendations
Mpigi, Masaka and Rakai, is recorded ashaving increased significantly during thesecond half of the 1990s (Legg, 1995;Otim-Nape et al., 1997; Legg and Ogwal,1998). This is now thought to haveresulted from a synergistic interactionwith an unusually severe form of CMD,which spread through Uganda duringthis period (Colvin et al., 1999) andcontinues to spread in the East andCentral African region (Legg, 1999). Theinteraction has occurred in all agro-ecologies in Uganda but it remains to beseen if the same will occur in the drieragro-ecologies to the south, in Tanzania.In Madagascar, there was no clearpattern of variation in abundance(Figure 3), although abundance in thesouthern central areas was lower thanelsewhere.
Variation in abundance ofsweetpotato whiteflies was much greaterthan for cassava whiteflies (Table 1).Very high populations were recordedaround the shores of Lake Victoria,whilst whiteflies were not recorded at allfrom many locations in Madagascar. Itis likely that seasonal effects can partlyexplain this degree of variation, sincesweetpotato is grown for a much shorterperiod (typically 4-6 months) thancassava (typically 10-24 months). Thus,although cassava crops may be availablethroughout the year, in drier areas,sweetpotato cultivation outside the maingrowing season is confined to very smallareas used for the maintenance ofplanting material. Associated with this,there is evidence showing that B. tabacion sweetpotato colonize a wide range ofcrop plants and weed species, incontrast to B. tabaci on cassava, whichappear to be largely restricted tocassava (Burban et al., 1992; Legg et al.,1994).
Whitefly-transmitted viruses incassava
Stem samples of CMD-diseased cassavawere sent from all nine principal
participating countries to the JohnInnes Centre (JIC) for diagnostics andcharacterization. Chapter 1.11 (thisvolume) reports the results of thiswork. Until 1996, it was consideredthat two cassava mosaicbegomoviruses were occurring inAfrica, with largely disjunctdistributions (Swanson and Harrison,1994). African cassava mosaic virus(ACMV) was reported to occur in muchof West, Central and southern Africa,and East African cassava mosaic virus(EACMV) was said to be restricted tocoastal East Africa, Madagascar,Malawi and Zimbabwe. More recentdiagnostic surveys have shown thatEACMV is not restricted to East Africa(Offei et al., 1999; Ogbe et al., 1999),that the distributions of EACMV andACMV overlap (Ogbe et al., 1996; 1997)and that mixed virus infections occur(Harrison et al., 1997). Results fromthe TWF-IPM Project (Chapter 1.11,this volume) have extended ourunderstanding further by establishingthe first record of EACMV from Beninin West Africa; identifying a novelbegomovirus from cassava inZimbabwe, which appears to be closelyrelated to South African cassava mosaicvirus (Rey and Thompson, 1998); andhighlighting the occurrence of virusmixtures in East Africa. Dual infectionsare a relatively uncommon occurrence(< 10%) and it is significant that allthese were recorded from the part ofEast Africa affected by the pandemic ofunusually severe CMD (Otim-Nape etal., 1997; Legg 1999; Chapter 1.1, thisvolume).
Although virus diagnoses havebeen carried out on only a limitednumber of samples, a relationship isapparent between the viruses affectingcassava plants and the disease caused.In West Africa, diversity seems to belimited and symptoms of disease aregenerally mild to moderate (Table 1). InEast and southern Africa, by contrast,

104
Whiteflies and Whitefly-borne Viruses in the Tropics
Table 1. Summary of data on the incidence of cassava mosaic disease (CMD), sweetpotato virusdisease (SPVD) and abundance of Bemisia tabaci, from Sub-Project 4 diagnostic surveys.
Country Survey area Fieldsa CMD Wfinfc CMD Cassava SPVD SP(%)b (miu) sev.d WFe (%)f WFg
Uganda North 20 (20) 77 48.0 3.0 1.2 5.0 1.40
East 20 (20) 68 46.0 3.1 1.8 12.0 5.40
South/Central 20 (20) 79 53.0 2.8 3.8 15.0 12.40
West 20 (20) 48 55.0 2.8 9.2 10.0 3.00
Kenya Coastal Province 17 (15) 56 36.0 2.3 2.9 0.5 0.50
Western Province 15 (18) 85 64.0 2.9 0.3 6.0 2.30
Nyanza Province 18 (17) 11 2.3 2.2 0.5 7.0 4.90
Madagascar Central plateau 35 (11) 31 7.8 2.6 7.1 3.0 0.01
Humid coast 22 (15) 71 11.0 3.5 4.3 2.0 0
Dry coast 12 (7) 46 11.0 3.2 5.9 18.0 0.10
Semi-humid coast 42 (6) 41 11.0 3.3 2.6 0 0
Tanzania Lake zone plateau 20 (20) 21 1.7 2.6 0.7 11.0 40.10
Semi-humid coast 20 (20) 12 7.1 2.7 1.2 13.0 2.80
Humid coast 20 (20) 39 21.0 3.0 5.8 6.0 3.10
Malawi Central lakeshore 20 62 55.0 2.8 1.5 - -
Northern lakeshore 9 52 34.0 2.7 1.0 - -
Central plateau 12 11 3.7 3.0 1.3 - -
Ghana Coastal savannah 10 76 44.0 2.2 11.8 - -
Rainforest 25 69 23.0 2.2 1.5 - -
Transition forest 25 74 54.0 2.5 1.0 - -
Guinea savannah 20 68 44.0 2.4 0.3 - -
Benin Transition forest 28 26 3.4 2.1 4.2 - -
Wet savannah 19 43 0.5 2.1 0.7 - -
Dry savannah 13 49 3.4 2.1 3.4 - -
Nigeria Rainforest 25 83 3.9 2.4 3.1 - -
Transition forest 25 32 2.7 2.1 2.6 - -
Wet savannah 20 56 0 2.4 1.0 - -
Dry savannah 10 45 2.4 2.2 0.3 - -
Cameroon South-west 26 54 21.0 2.2 4.1 - -
North-west 16 55 21.0 2.4 2.4 - -
Centre-south 28 73 5.6 2.3 3.1 - -
a. Fields: Figures in parentheses indicate number of sweetpotato fields sampled.b. CMD %: Total incidence of CMD, calculated as the average percentage of plants showing symptoms
of virus disease from 30 plant samples recorded in each dataset (i.e., one field).c. Wfinf (miu): Incidence of plants infected with CMD whitefly in the current season, transformed to
allow for multiple infection units (Gregory, P. H. 1948. The multiple infection transformation. Ann.Appl. Biol. 35: 412-417).
d. CMD sev.: Severity of CMD symptoms, based on an arbitrary 1-5 scale, where 1 indicates low and5 indicates high severity of symptoms.
e. Cassava WF: Mean whitefly abundance for cassava recorded as the average number of adultwhiteflies counted on the five uppermost leaves of 30 plant samples recorded in each field.
f. SPVD %: Total incidence of SPVD, calculated as the average percentage of plants showingsymptoms of virus disease from 30 plant samples recorded in each field.
g. SP WF: Mean whitefly abundance for sweetpotato recorded as the average number of adultwhiteflies counted in ten 1-minute counts made at random in the sampled field.

105
Conclusions and Recommendations
complexity appears greater, dualinfections are more prevalent andsevere disease is more widespread,although in most instances (4/5) thedual infections were in areas affectedby the East African CMD pandemic(Legg, 1999). An association betweenvirus infection and epidemiology issimilarly apparent (Table 1). In EastAfrica, current-season whitefly-borneinfection is greatest in Uganda, wherethe “Uganda variant” of EACMV(EACMV-Ug) was reported (Chapter1.11, this volume) and where EACMV-Ug/ACMV mixtures were common, asreported earlier by Harrison et al.(1997); whilst in Nigeria, where onlyACMV was reported, current-seasonwhitefly-borne infection was veryinfrequent.
The incidence of CMD variedconsiderably within sub-regions. InWest Africa, disease incidence rangedfrom 36% (Benin) to 72% (Ghana);whilst in East and southern Africa, itranged from 21% (Tanzania) to 68%(Uganda). In all countries, theproportion of diseased plants that hadarisen from planting diseased cuttings(cutting infection) was greater than thatderived from current season whitefly-borne infection. Many factors influenceCMD incidence, including the virus(es)infecting the plant, cultivar response tothe virus(es), vector abundance andtransmission efficiency, the physicalenvironment, and cropping practices(including vector or diseasemanagement measures). Most of thesefactors remain poorly defined for manyof the participating countries, althoughthe surveys have provided quantitativedata relating to some aspects. In thisrespect it would be useful to comparethe epidemiology of CMD betweenregions, using common cultivars forreference, as proposed by Legg et al.(1997).
Whitefly-transmitted viruses insweetpotato
Sweetpotato leaf samples withsymptoms of virus disease fromUganda, Kenya, Tanzania and Zambiawere analysed. Chapter 1.12 discussesin detail the results of virus diagnosesfrom Uganda, Kenya and Tanzania, andKaitisha and Gibson (1999) discussthose from Zambia. The East Africanstrain (SEA) of Sweetpotato chloroticstunt virus (SPCSV) occurredthroughout the region and, in Uganda,Aritua et al. (Chapter 1.12, thisvolume) have discussed distributionsfor the two distinct serotypes (SEA1 andSEA2) described by Alicai et al. (1999).The other whitefly-borne virusidentified from project surveys wasSweetpotato mild mottle virus (SPMMV),although this was restricted to areasaround the shores of Lake Victoria(Chapter 1.12, this volume). SPVD,which results from co-infection ofsweetpotato plants with SPCSV and theaphid-borne Sweetpotato feathery mottlevirus (SPFMV), occurred throughoutareas sampled. The disease was mostfrequent in south-western Uganda, theLake Victoria zone of Tanzania, south-eastern coastal Tanzania and the semi-arid south-western part of Madagascar.Diagnostic tests related SPVD to theco-occurrence of SPCSV and SPFMVfor each of these regions (Chapter 1.12,this volume). No samples fromMadagascar were tested.
Whitefly natural enemies
The agreed survey protocol for theTWF-IPM Project provided guidelinesfor the collection of parasitoids,predators and entomopathogens duringthe course of diagnostic surveys.Limitations in the methods proposed,and the short period of time availablefor natural enemy collection at eachsampling site, meant that the collectionof natural enemies was lesscomprehensive than had been

106
Whiteflies and Whitefly-borne Viruses in the Tropics
anticipated. More intensive andtargeted surveys would be required forthe adequate characterization ofpredators and entomopathogens inparticular.
Survey teams in most countriescollected parasitized whitefly mummiesand reared adult parasitoids fromthem. The parasitoids weresubsequently mounted and identifiedby Dr. Mohammed Ali Bob of theInternational Center of InsectPhysiology and Ecology (ICIPE).Dr. A. Polaszek of the British Museum(Natural History) verified representativespecimens. Five species of parasitoidwere recorded: Encarsia sophia (Giraultand Dodd), Encarsia lutea (Masi),Encarsia mineoi Viggiani, Encarsia sp.(luteola group) and Eretmocerus sp.(Table 2). Only two species, E. sophiaand Eretmocerus sp., were widelydistributed, parasitizing whiteflies onboth cassava and sweetpotato.E. sophia was an order of magnitudemore common that Eretmocerus sp.,although it is considered that easyrecognition of the former, resultingfrom the black coloration of themummy from which it emerges, mayhave partially biased the sampling.Whilst over 30 species of parasitoidhave been described as parasitizingB. tabaci worldwide (Gerling, 1986),only limited information is availableabout Bemisia parasitoids in Africa,and virtually no information relating tocassava or sweetpotato (Fishpool and
Burban, 1994). Whilst this componentof the TWF-IPM Project’s diagnosticphase had deficiencies, it isnevertheless considered that theinformation obtained has provided animportant baseline from which todevelop future studies. Key issues thatwill need to be more fully addressedinclude the variation in parasitoiddiversity between regions, of whichthere was evidence in this study, andthe role (if any) of the differentparasitoid species in the populationdynamics of Bemisia spp. and theepidemiology of CMD.
Increased Socio-EconomicUnderstanding
Farmers’ assessment ofwhitefly-related problems
There was a clear relationship betweenfarmers’ assessments of CMD andSPVD and the prevalence and severityof the diseases as described from thefield data collection. In Uganda, allfarmers both recognized CMD andconsidered it a problem for theircassava production. In the countries ofWest Africa, on the other hand,although typically about 70% offarmers were able to recognize CMD,less than 50% considered it a problem.A smaller proportion of sweetpotatofarmers recognized SPVD but, where itwas recognized, most farmers
Table 2. Aphelinid parasitoids identified from diagnostic surveys of Sub-Project 4.
Country Encarsia Eretmocerus Encarsia sp. Encarsia Encarsiasophia sp. (luteola group) lutea mineoi
Uganda 62 46 3 - -
Kenya 13 2 - - -
Tanzania 62 - 1 - -
Ghana 4 - - - -
Benin 16 2 - - -
Nigeria 557 21 4 7 5
Cameroon 324 38 - - -

107
Conclusions and Recommendations
considered it a problem. Much smallerproportions of farmers were able torecognize whiteflies and manyexpressed surprise when informed oftheir role as disease vectors. Therewere significant differences betweencountries in this respect, however, withthe extremes being Madagascar, whereno farmers reported recognizingwhiteflies, and Benin, where 70%recognized them.
Farmers had names for whitefliesand whitefly-transmitted diseases in allcountries, but in many cases thesewere non-specific, translating simply as“insect” or “disease”. More specificnames were commonest for CMD,which was often described usinganthropomorphic disease equivalentssuch as “leprosy”. Names for SPVDwere often exactly the same as thoseused for CMD. Where such names arein common usage, potential exists fortheir use, perhaps with minormodification, in training and extensionprograms in the future. Estimates ofloss to the respective diseases weremost frequently in the range of 25%-75% for cassava and 25% or less forsweetpotato, although most surveyteams considered that these were likelyto be overestimates, particularly inWest African countries, where diseasesymptoms (in this case only CMD) weretypically mild. Participatory yield lossstudies would be helpful in many of thesurveyed countries to evaluate lossesscientifically and reconcile suchestimates with farmer perceptions. Theshortcoming of this and otherresponses emphasized the need to linkproducer interviews with the collectionof more objective field data.
Managing whiteflies andwhitefly-transmitted viruses
There were widely divergent responseson the use of vector and diseasemanagement measures. In Ghana andMadagascar, less than 5% of farmers
reported doing anything at all tocontrol CMD. This result wasparticularly anomalous for Ghana,since many farmers reported relativelyhigh losses. Management practiceswere most widely reported in Ugandaand Cameroon, where most farmersindicated the use of at least onepractice. The most cited methods ofdisease management were thephytosanitary techniques of roguing,and selection of disease-free stems.This has to be set against thepredominance of the use of diseasedcuttings as planting material in almostall areas surveyed. Selection of disease-free planting material similarly wascited widely as a management tacticbut the health status of stems wasnormally less important as a criterionfor planting material selection thanwere agronomic characters.
Although it appears that farmersover-estimated their use of roguing andselection of healthy cuttings, theyunder-reported use of disease-resistantvarieties. Use of cassava varietiesrecommended for resistance to CMDwas most widespread in Cameroon,where 48% of farmers reported growingvariety “Agric” in the knowledge that itwas resistant to CMD. In Uganda, morethan 15% of sweetpotato farmers werefound to be growing SPVD-resistantvarieties but none of these realized thatthis was a method of managing SPVD.
Very few farmers reported usingpesticides to control whiteflies orwhitefly-borne viruses. Occasionally,sweetpotato farmers reported pesticideuse but this was more likely to be forthe purpose of controlling other insectpests such as the sweetpotato butterflyAcraea acerata Hewitson. Chapter 1.13(this volume) gives a more detailedassessment of the approaches ofsweetpotato farmers to SPVD control.Few farmers reported having receivedany technical assistance with respect

108
Whiteflies and Whitefly-borne Viruses in the Tropics
to the control of either CMD or SPVDand, as a result, most considered thatthe management methods being usedwere their own. The only exception tothis was Cameroon, where 39% offarmers reported that they had receivedtechnical support relating to whitefliesand whitefly-borne diseases.
Recommendations
The overall picture that emerges fromthis assessment of farmers’ awarenessof whiteflies and whitefly-bornediseases in cassava and sweetpotatoand their approach to diseasemanagement is one of weakunderstanding of the nature of theproblem and a correspondinglyhaphazard implementation ofmanagement tactics. Much of theresponsibility for this lies with theresearch (both national andinternational) and extension systems.Researchers have failed to provide awell-defined and well-tested strategy formanaging CMD and SPVD. Moreover,weakness in the extension systems ofmost of the participating countries hasmeant that even where managementrecommendations have been defined,these are not reaching the majority offarmers (as is particularly the case forthe control of whitefly-borne diseases ofcassava and sweetpotato in Uganda). Inorder to address this situation, keyobjectives for future work on whitefliesand whitefly-borne diseases, for bothcassava and sweetpotato, should be toencourage researchers, extensionagents and farmers to work together to:
(1) Define exactly where diseasemanagement is needed and wherenot;
(2) Develop effective, clearly definedand readily implemented vectorand disease management methods(which based on current knowledgeare most likely to include choosing
disease-resistant varieties, selectingdisease-free planting material and/or roguing of diseased plants); and
(3) Facilitate the adoption of thesemethods by using participatorytraining and all media channelsavailable to improve the access offarmers to current knowledge.
Sub-Project 4 of the TWF-IPMProject has been successful inachieving part of the first objective.Following the completion of thediagnostic phase, there is a clearerunderstanding of the distribution ofwhitefly-borne viruses, what theirimpact is in terms of disease incidenceand epidemiology and how farmers areresponding to these diseases. Theproject also has been uniquelysuccessful in obtaining comparableinformation of this type from nine of themajor cassava-producing countries ofAfrica and four of the majorsweetpotato-producing ones; thesecountries together account for morethan 50% of African production of eachcrop. In achieving this, the project hasestablished a network of researchersworking on whiteflies and whitefly-borne disease of root crops in sub-Saharan Africa. In order to completethe first objective and to begin toaddress the second, the followingissues, a number of which have beenhighlighted in the country chapters,need to be addressed.
(1) How severe are yield losses in theCMD-diseased local landraces thatstill dominate cassava productionin Africa? Knowledge of this isparticularly critical, for example, inGhana, where farmers report highlosses yet appear to make littleattempt to control them. A realisticestimate of current yields and yieldlosses would help direct researchand extension efforts moreeffectively.

109
Conclusions and Recommendations
(2) What are the conditions underwhich tactics for improving planthealth, especially selection ofdisease-free cuttings and roguingof diseased plants, are most likelyto be effective?
(3) Where is host plant resistancerequired as a foundation forsustainable management ofcassava and/or sweetpotato (ascompared with a strategy focusingon improving the husbandry ofland races) and are particularvirus/vector/environmentalcharacteristics common to theseareas?
(4) Where conditions demand the useof host plant resistance, as hasbeen the case in Uganda during thepandemic of severe CMD, what arethe constraints to the adoption ofnewly developed CMD-resistantvarieties?
(5) Are there alternative, novelapproaches to crop healthmanagement that can be used tosupplement existing methods?Although both biological controland host plant resistance areregarded conventionally as beingmore appropriate for use againstdirect pests, rather than virusvectors, is there scope for theirdeployment?
(6) Where are successes currentlybeing realized in CMD and/orSPVD management and how canexperience from these situations beincorporated into research andextension efforts elsewhere?
The second phase of the TWF-IPMProject proposes to address keyresearch and implementation issuessuch as those indicated above. In theformulation of this vitally importantphase of the project, attention needs tobe paid to collaborative mechanisms in
addition to technical issues; the firstphase, described in this volume, hasprovided some important lessons inthis regard.
(1) The coordination structuredeveloped during the first phasehas provided an effective means bywhich a global-scale project can beexecuted. Effective interaction,facilitated by electroniccommunication, is vital for theefficient working of this structure.
(2) At the outset of the next phase ofthe project, all partners at sub-project level should participate inan introductory meeting, duringwhich opportunities are providedfor critical review of the proposedwork plans and budgets, and fortraining in additional skillsrequired for projectimplementation. This was not doneduring the first phase and, as aresult, it was difficult toconsistently train partners innational research systems and toprovide all partners with anappreciation of the continental andglobal contexts of the project.
(3) The sub-project should budget forannual review meetings for allprincipal partners. If resources arelimited, the number of partnersshould be restricted to allow fullparticipation of each.
(4) If a large number of nationalresearch system partners are to beincluded, as was the case in thefirst phase of Sub-Project 4, thetask of co-ordination and provisionof research support, executed bythe co-ordinating internationalcentre, should be divided betweenmajor regions covered. In the firstphase of Sub-Project 4, theInternational Institute of TropicalAgriculture (IITA) provided co-ordination and support through

110
Whiteflies and Whitefly-borne Viruses in the Tropics
offices in both West and EastAfrica. This division ofresponsibility was considered to bea key factor in the successfulimplementation of the sub-projectalthough such a division demandsan effective exchange of ideas andinformation between the regions.
Whiteflies continue to pose atremendous challenge to farmers,researchers and development workersin Africa, as well as in the wider globalcontext. However, significant successesare being realized in meeting thischallenge both in terms ofcharacterizing the problems and indeveloping strategies to address themin a sustainable manner. In the Africancontext, at stake are the livelihoods ofmany millions of producers, whodepend on cassava and sweetpotato assources of cash income, andconsumers, both rural and urban, whodepend on these crops to provideaffordable staple foods. It is to behoped, therefore, that initiatives tostrengthen management of whitefly-transmitted virus diseases of these twomajor food crops, including the secondphase of the TWF-IPM Project, will beable to attract a level of supportcommensurate with the magnitude ofthe problem.
Acknowledgements
A grant from the Danish InternationalDevelopment Assistance funded thediagnostic phase of the TWF-IPMProject, Sub-Project 4. Each of theproject partners providedcomplementary support. The authorswould like to extend particular thanksto Dr. Mohammed Ali Bob of ICIPE,who identified all the whitefly andparasitoid samples for Sub-Project 4.
References
Alicai, T.; Fenby, N. S.; Gibson, R. W.;Adipala, E.; Vetten, H. J.; Foster, G.D.; Seal, S. E. 1999. Occurrence oftwo serotypes of Sweetpotato chloroticstunt virus in East Africa and theirassociated differences in coatprotein and HSP70 homologuegene sequences. Plant Pathol.48:718-726.
Azab, A. K.; Megahed, M. M.; El-Mirsawi,D. H. 1971. On the biology ofBemisia tabaci (Genn.). Bull. Soc.Entomol. Egypte 55:305-315.
Burban, C.; Fishpool, L. D. C.; Fauquet,C.; Fargette, D.; Thouvenel, J. -C.1992. Host associated biotypeswithin West African populations ofthe whitefly Bemisia tabaci (Genn.)(Hom., Aleyrodidae). J. Appl.Entomol. 113:416-423.
Colvin, J.; Otim-Nape, G. W.; Holt, J.;Omongo, C.; Seal, S.; Stevenson, P.;Gibson, R.; Cooter, R. J.; Thresh, J.M. 1999. Factors driving the currentepidemic of severe cassava mosaicdisease in East Africa. In: Procs.VIIth International Plant VirusEpidemiology Symposium, 11-16April 1999, Almeria, ES. p. 76-77.
Fauquet, C.; Fargette, D.; Thouvenel,J. -C. 1987. Epidemiology of Africancassava mosaic on a regional scalein the Ivory Coast. In: Procs.International Seminar on Africancassava mosaic disease and itscontrol, 4-8 May, Yamoussoukro, CI.Centre Technique de CoopérationAgricole et Rurale, Wageningen, NL,and Institut Français de RechercheScientifique pour le Développementen Coopération, Paris, FR.p. 131-135.
Fauquet, C.; Fargette, D.; Thouvenel,J. -C. 1988. Some aspects of theepidemiology of African cassavamosaic virus in Ivory Coast. Trop.Pest Manag. 34:92-96.
Fishpool, L. D. C.; Burban, C. 1994.Bemisia tabaci: The whitefly vector ofAfrican cassava mosaic geminivirus.Trop. Sci. 34:55-72.

111
Conclusions and Recommendations
Gerling, D. 1986. Natural enemies ofBemisia tabaci, biologicalcharacteristics and potential asbiological control agents: A review.Agric. Ecosyst. Environ. 17:999-110.
Gerling, D.; Horowitz, A. R.; Baumgärtner,J. 1986. Autecology of Bemisia tabaci.Agric. Ecosyst. Environ. 17:5-20.
Gregory, P. H. 1948. The multiple infectiontransformation. Ann. Appl. Biol.35:412-417.
Harrison, B. D.; Zhou, X.; Otim-Nape, G.W.; Liu, Y.; Robinson, D. J. 1997.Role of a novel type of doubleinfection in the geminivirus-inducedepidemic of severe cassava mosaic inUganda. Ann. Appl. Biol. 131:437-448.
IITA (International Institute of TropicalAgriculture). 1998. EcologicallySustainable Cassava Plant Protection(ESCaPP) in South America andAfrica. Africa: Project findings andrecommendations. Ibadan, NG.
Kaitisha, G. C.; Gibson, R. W. 1999.Identification of sweetpotato featherymottle and chlorotic stunt viruses insweetpotato in Zambia. In: Akoroda,M. O.; Teri, J. M. (eds.). Procs.Scientific Workshop of the SouthernAfrican Root Crops Research Network(SARRNET), 17-19 August 1998,Lusaka, ZM. p. 293-299.
Legg, J. P. 1995. The ecology of Bemisiatabaci (Gennadius) (Homoptera:Aleyrodidae), vector of Africancassava mosaic geminivirus inUganda. Ph.D. thesis, University ofReading, GB.
Legg, J. P. 1999. Emergence, spread andstrategies for controlling thepandemic of cassava mosaic virusdisease in east and central Africa.Crop Prot. 18:627-637.
Legg, J. P.; Ogwal, S. 1998. Changes in theincidence of African cassava mosaicgeminivirus and the abundance of itswhitefly vector along south-northtransects in Uganda. J. Appl.Entomol. 122:169-178.
Legg, J. P.; Gibson, R. W.; Otim-Nape, G.W. 1994. Genetic polymorphismamongst Ugandan populations ofBemisia tabaci (Gennadius)(Homoptera: Aleyrodidae), vector ofAfrican cassava mosaic geminivirus.Trop. Sci. 34:73-81.
Legg, J. P.; James, B.; Cudjoe, A.;Saizonou, S.; Gbaguidi, B.; Ogbe,F.; Ntonifor, N.; Ogwal, S.; Thresh,J.; Hughes, J. 1997. A regionalcollaborative approach to the studyof ACMD epidemiology in sub-Saharan Africa. Afr. Crop Sci. J.3:1021-1033.
Ogbe, F. O.; Songa, W.; Kamau, J. W.1996. Survey of the incidence ofAfrican cassava mosaic and EastAfrican cassava mosaic viruses inKenya and Uganda using amonoclonal antibody baseddiagnostic test. Roots 3:10-13.
Ogbe, F. O.; Legg, J.; Raya, M. D.;Muimba-Kankolongo, A.; Theu, M.P.; Kaitisha, G.; Phiri, N. A.; Chalwe,A. 1997. Diagnostic survey ofcassava mosaic viruses in Tanzania,Malawi and Zambia. Roots 4:12-15.
Ogbe, F. O.; Atiri, G. I.; Robinson, D.;Winter, S.; Dixon, A.; Quin, F. M.;Thottappilly, G. 1999. First report ofEast African cassava mosaicbegomovirus in Nigeria. Plant Dis.83:398.
Otim-Nape, G. W.; Bua, A.; Thresh, J. M.;Baguma, Y.; Ogwal, S.; Semakula,G. N.; Acola, G.; Byabakama, B.;Martin, A. 1997. Cassava mosaicvirus disease in Uganda: Thecurrent pandemic and approachesto control. Natural ResourcesInstitute (NRI), Chatham, GB. 65 p.
Rey, M. E. C.; Thompson, G. 1998.Cassava mosaic virus disease inSouth Africa. Roots 5:3-5.
Swanson, M. M.; Harrison, B. D. 1994.Properties, relationships anddistribution of cassava mosaicgeminiviruses. Trop. Sci. 34:15-25.

BLANCA 112

SECTION TWO
Whiteflies as Pests and Vectors ofViruses in Vegetable and
Legume Mixed Cropping Systems inEastern and Southern Africa
SECTION TWO
Whiteflies as Pests and Vectors ofViruses in Vegetable and
Legume Mixed Cropping Systems inEastern and Southern Africa

BLANCA 114

115
Introduction
CHAPTER 2.1
IntroductionMohamed Ali Bob, Rebbeca Raini and
Bernhard Löhr*
In some parts of Africa such as in theSudan and Tanzania the countrysideresembles a graveyard, not through theravages of civil war or natural disasterbut because of one pest, the whiteflyBemisia tabaci (Gennadius), which issucking the life out of Africa’s crops.Bemisia, which was originally only aproblem in industrial crops such astobacco (Nicotiana tabacum L.) andcotton (Gossypium hirsutum L.) is nowmuch more widespread and despite allefforts to control this pest, it hasstayed one step ahead of man’s actionsagainst it. Bemisia has done this notonly by developing resistance to manyof the chemicals commonly used butalso by diversifying its tastes and itshabitat to a much wider range of staplecrops such as cassava (Manihotesculenta Crantz) and common bean(Phaseolus vulgaris L.) to high-valuehorticultural crops such as tomato(Lycopersicon esculentum Mill.) andmelon (Cucumis melo L.). On-farmsurveys in the region have shown thatthe number of producers who haveabandoned their crops because oftomato leaf curl diseases is on a sharpincrease. In the Sudan especially,whiteflies have forced entire areas outof production, depriving producers offood and cash income.
Selection of Target Cropsand Target Countries
Tomato is one of the most widelycultivated vegetable crops in the region,predominantly grown by small-scaleproducers for fresh consumption (GTZ,1995). National scientists have rankedB. tabaci as the most important pest/vector in tomato in Tanzania andKenya, and the second most importantin Malawi after red spider mite (Varelaand Pekke, 1995). In the Sudan,B. tabaci is considered the crop pest ofhighest economic importance(Eveleens, 1983; Ahmed et al., 1987).
Knowledge Available onWhitefly-Related Problems
Despite these opinions, no quantitativedocumentation was available on theeconomic importance of whiteflies aspests and/or vectors in vegetablecropping systems in the countries ofthis sub-project prior to itscommencement. There was no clearmap of the areas affected in thesecountries and related information oninsecticide use and the cost of controlin the affected areas. Knowledge onwhitefly-transmitted viruses invegetables, the host range and naturalenemies was also lacking in mostpartner countries before this projectbegan. Information on the status ofwhitefly resistance to insecticides was
* International Center of Insect Physiology andEcology (ICIPE), Nairobi, Kenya.

116
Whiteflies and Whitefly-borne Viruses in the Tropics
available only from the Sudan (seeChapter 2.2).
Although the history of whiteflyresearch in the Sudan goes back to the1930s (Kirkpatrick, 1931), asubstantial collection of grey literatureon whitefly and whitefly-transmittedviruses from local stations/projectreports in the Sudan revealed that theresearch has focused almost entirelyon whitefly-related problems in cottonproduction. Whitefly-related problemsin vegetable production appear morerecent, for example, Tomato yellow leafcurl virus (TYLCV) was first identified in1965 (Yassin and Nour, 1965).Nevertheless, research on whitefly-related problems in vegetable crops hasbeen limited.
In Tanzania, TYLCV was firstidentified in 1990 (Czosnek et al.,1990). Later, Chiang et al. (1997)identified a different virus, namedToLCV-Tz, which has complicated thescenario in Tanzania. No furtherdocumentation on whitefly-relatedproblems in vegetable crops is availablefrom this country. In Kenya andMalawi, documentation on whitefly-related problems is only available oncitrus (Citrus spp. L.) and cassava.
Planning andCo-ordination
In order to form the basis for thediagnostic activities for the first phase,a planning and methodology workshopwas held, with the participation ofscientists from the national teams, atthe International Center of InsectPhysiology and Ecology (ICIPE) in April1997. A detailed work plan for the firstphase of the project was developed,methodologies were discussed andviews and experiences were sharedamong the participants.
A progress meeting held at theICIPE followed this up in June 1998.Progress of the two African sub-projects was presented, experiencesshared and priority research areas forPhase 2 were considered. Theparticipating co-ordinators andnational teams made 10 presentationson whitefly research.
Finally, a research agenda forPhase 2 was outlined at a jointplanning workshop held at the ICIPE inSeptember 1999. Research prioritieswere thoroughly discussed and prioritysetting was carried out with fullparticipation of all national scientists ofSub-Project 4, co-ordinators andcollaborators from advanced researchinstitutions.
Continent-Wide Appeal
Additionally, the formation of anAfrican Whitefly and GeminivirusNetwork was spearheaded by the sub-project at the 12th Meeting of theAfrican Association of Insect Scientistsheld in June 1997 in Stellenbosch,South Africa. Thirty-two researchersfrom 12 countries in Africa participatedand expressed interest in participationin a regional information exchangenetwork led by ICIPE.
References
Ahmed, A. H. M.; Elhang, E A., Bashir, H.H. 1987. Insecticide resistance inthe cotton whitefly (Bemisia tabaciGenn.) in the Sudan Gezira. Trop.Pest Manag. 33:67.
Chiang, B. T.; Nakhla, M. K.; Maxwell, P.;Schoenfelder, M.; Green, S. K.1997. A new geminivirus associatedwith a leaf curl disease of tomato inTanzania. Plant Dis. 81:111.

117
Introduction
Czosnek, H.; Navot, N.; Laterrot, H. 1990.Geographical distribution of Tomatoyellow leaf curl virus. A first surveyusing a specific DNA probe.Phytopathol. Mediterr. 29:1-6.
Eveleens, K. G. 1983. Chemical insectcontrol in the Sudan Gezira:Analysis of a crisis crop. Crop Prot.2:273-287.
GTZ (Deutsche Gesellschaft fürTechnische Zusammenarbeit).1995. Major pests of tomatoes ineastern and southern Africa:Compilation of past research workand identification of IPMopportunities. GTZ-Integrated PestManagement (IPM) HorticultureProject Report, Nairobi, KE. 32 p.
Kirkpatrick, T. W. 1931. Further studieson leaf-curl of cotton in the Sudan.Bull. Entomol. Res. 22:323-363.
Varela A. M.; Pekke, A. 1995. Proceedingsof the Tomato Planning Workshopfor Eastern and Southern AfricaRegion, 16-20 October 1995,Harare, ZW. Deutsche Gesellschaftfür Technische Zusammenarbeit(GTZ)- Integrated Pest Management(IPM) Horticulture Project, Nairobi,KE. 32 p.
Yassin, A. M.; Nour, M. A. 1965. Tomatoleaf curl disease, its effects on yieldand varietal susceptibility. SudanAgric. J. 1:3-7.

118
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 2.2
SudanGassim Dafalla* and Musa Ahmed**
Introduction
Geographical context
Diagnostic surveys were conductedduring 1997-99 following themethodology agreed among projectpartners, and covering Gezira, Butana,Dindir, Hasaheisa, Kamlin, Managil,Umulgra, White Nile, El Jabalain, UmRawaba, Er Rahad, Dongola, Merewe,Gedaref and Nahr el Rahad in central,eastern, northern and western Sudan(Figure 1). Most of the surveyed sitesare low lying (< 500 m above sea level).The climate is hot and dry for most ofthe year, with temperatures in summerreaching more than 40 °C and relativehumidity less than 10%, and irrigationis commonly used. The driest of thesurvey areas are Dongola and Merewein the Northern State. Gedaref regionand some areas in Kordufan Provinceare at a somewhat higher altitude(> 500 m above sea level) and receiveenough rainfall for rain-fed vegetableproduction.
The emergence of Bemisia tabacias a pest and virus vector
The whitefly species Bemisia tabaci(Gennadius) is considered to be the
economically most important crop pestin the Sudan (Eveleens, 1983; Ahmedet al., 1987). In addition to the directdamage this whitefly causes throughfeeding, it disseminates a number ofviruses that affect fibre and vegetablecrops grown in the country.
Research on B. tabaci and whitefly-transmitted viruses (WTVs) in Sudanstarted in the late1920s, whenrecurrent outbreaks of Cotton leaf curlvirus (CLCuV), transmitted by B. tabaci,threatened to end cultivation of cotton(Gossypium hirsutum L.) in the Gezira(Kirkpatrick, 1931). Since then, aconsiderable research effort has beenfocused on B. tabaci and WTVs in
* Plant Pathology Centre, Faculty ofAgricultural Science, University of Gezira,Wad-Medani, Sudan.
** Agricultural Research Corporation,Wad-Medani, Sudan.
Figure 1. Areas surveyed for whitefly incidencein the Sudan.
Towns
Surveyedareas
Wadi Halpha
Argo
Berbetti
Al Khurtum
Pibor Post
Sudan

119
Sudan
cotton, leading to the introduction ofCLCuV-resistant varieties in the Gezirain the late 1960s. However, research onwhitefly-related problems in vegetablecrops has been limited. Vegetables areseverely affected by WTVs and theincidence of WTVs in a given seasonconstitutes the major factordetermining the success or failure oftomato (Lycopersicon esculentum Mill.),cucurbits, okra (Abelmoschusesculentus [L.] Moench), pepper(Capsicum annuum L.) and commonbean (Phaseolus vulgaris L.) in thatseason. It is estimated that, onaverage, 75% of the fruit yield oftomato, watermelon (Citrullus lanatus[Thunb.] Matsum. & Nakai), muskmelon (Cucumis melo L.) and okra islost each year to WTVs.
Yassin and Nour (1965) firstreported Tomato leaf curl virus,transmitted by B. tabaci. Laterinvestigations showed that the virusfrom Sudan is similar to Tomato yellowleaf curl virus (TYLCV) from the EastMediterranean region. In recent years,severe yellowing symptoms wereobserved in watermelon and melongrown in many parts of the country,similar to those described for theB. tabaci-transmitted Watermelonchlorotic stunt virus (WmCSV) in Yemen.CLCuV not only affects cotton but alsohas been reported by many workers tocause heavy damage in okra in mostparts of the country. Both cotton andokra belong to the mallow family.According to some workers (cited byDafalla and Sidig, 1997) the whitefly-transmitted African cassava mosaicdisease is still the most destructive incassava in Western Equatoria Provincein southern Sudan.
At producer level, research effortshave not been reflected sufficiently inthe successful management of B. tabaciand WTVs, especially in vegetables. Inthe absence of such management
strategies, the problem has becomemore complicated and has defiedvegetable producers’ efforts toovercome it and secure a return fortheir efforts. The need to design viableintegrated control strategies, based ona strong scientific foundation andknowledge of the field situation, hasemerged as a major challenge forresearch.
Little information was available onthe whitefly-related problems ofvegetable production in Sudan prior tothe commencement of the TropicalWhitefly Integrated Pest Management(TWF-IPM) Project. Many findings fromthe survey conducted in the first,diagnostic phase of the project are newand add substantially to our knowledgebase on biological and socio-economicaspects of the whitefly and WTVproblems of vegetable cropping systemsin Sudan.
Increased BiologicalUnderstanding
Characterization ofbegomoviruses and whiteflybiotypes
Samples of whitefly adults and nymphswere collected mostly from tomato, witha few from lablab (Lablab purpureus [L]Sweet) and Nalta jute (Corchorusolitorius L.), on 80 farms. One hundredand fifty specimens were processed andmounted on 76 slides and sent to theInternational Center of InsectPhysiology and Ecology (ICIPE) foridentification. All the mounted nymphspecimens were identified as B. tabaci.
In an experimental field at theUniversity of Gezira, severe silver leafsymptoms were noticed on squash(Cucurbita pepo L.), heavily infestedwith whiteflies. These symptoms havebeen associated elsewhere with the

120
Whiteflies and Whitefly-borne Viruses in the Tropics
B. tabaci B-biotype (Shapiro, 1995) butmolecular characterization of thesewhitefly specimens is still pending. Ingeneral, some variability has beennoticed in the patterns of behaviourand efficiency in virus transmission,which also may be circumstantialevidence of the existence of differentBemisia biotypes.
Host plants on which whiteflieswere found to reproduce, identified inthe course of surveys in Sudan,belonged to the following families:Amaranthaceae, Asclepiadaceae,Asteraceae, Brassicaceae,Commelinaceae, Convolvulaceae,Cucurbitaceae, Euphorbiaceae,Lamiaceae, Leguminosae, Malvaceae,Solanaceae, Tiliaceae and Verbenaceae(Table 1).
Many of the WTVs in vegetables inSudan have been well characterized butmore information on the inter-seasonalvariation of virus incidence and on thedistribution and economic importanceof various WTVs in the different regionsis needed. The field surveys revealedthat TYLCV, WmCSV, Okra leaf curlvirus and Pepper leaf curl virus are themost widespread and economicallyimportant WTVs in Sudan. Tomato,watermelon, melon, okra, pepper andcommon bean were among the mostaffected crops.
Other viruses detected includeTomato vein thickening virus, whichcauses another important disease oftomato in Sudan and Bean mild mosaicvirus (BMMV). Cucumber vein yellowingvirus (CVYV) and a virus inducingpotyvirus-like yellowing have beenobserved in cucurbits but the latter isyet to be identified. In legumes, ageminivirus and a Closterovirus thataffect cowpea (Vigna unguiculata [L.]Walp.) and common bean have beenobserved and are yet to be identified.Symptoms of another putative WTV,
whose identity is yet to be established,were also noticed on pepper.
Some weeds, Acalypha indica L.,Datura stramonium L. and Solanumcoagulans Forskål, were identified asTYLCV reservoirs. Many other non-cultivated host plants of B. tabaci mayact as WTV reservoirs. Continuouscultivation of tomato and othersolanaceous vegetables, coupled withthe abundance of alternative hosts forvector and viruses, is very likely tocontribute to the build up of diseaseproblems.
Disease incidence and symptomseverity
The Gezira Irrigation Scheme is the areawith the most severe and persistentWTV disease problems in the country.Incidences of 80-100% were frequentlyrecorded in Gezira, Hasaheisa, Managil,Umelgura, Dindir and Butana. In half ofthe tomato fields in the Gezira Province,100% TYLCV incidence was recorded.B. tabaci was found to be moreabundant in the southern part of Gezirathat receives less rainfall than thenorthern region. High whiteflypopulations and TYLCV incidence wereobserved in Central Sudan, significantlylower populations and TYLCVincidences (1%-9%) were observed inNahr el Rahad (Eastern State), Dongolaand Merewe (Northern State), and inUmm Rawaba and Er Rahad (WesternState). No TYLCV symptoms wereobserved in Gedaref Province (Babiker,Khor-Garab, El Ramla, Basunda, Doka)and the Abu Habil Basin. Vegetablegardens along the Blue Nile River wereonce the sole suppliers of tomatooutside the period suitable for rain-fedcultivation. This is no longer the casebecause of severe TYLCV epidemics inrecent years.
The overall impression provided bythe surveys is that B. tabaci does not

121
Sudan
Table 1. Reproductive host plants of Bemisia tabaci (Gennadius) in the Sudan.
Family Species Common name
Crops
Brassicaceae Raphanus sativus L. Radish
Cucurbitaceae Citrullus lanatus (Thunb.) Matsum & Nakai Watermelon
Cucumis melo L. Melon
Cucumis sativus L. Cucumber
Cucurbita pepo L. Squash
Euphorbiaceae Manihot esculenta Crantz Cassava
Leguminosae Cajanus cajan (L.) Millsp. Pigeon pea
Cassia alexandrina Mill. Senna
Lablab purpureus (L.) Sweet Lablab
Lens culinaris Medik. subsp. culinaris Lentil
Phaseolus vulgaris L. Common bean
Vicia faba L. Broad bean
Lamiaceae Ocimum basilicum L. Basil
Malvaceae Abelmoschus esculentus (L.) Moench Okra
Gossypium hirsutum L. Cotton
Hibiscus cannabinus L. Kenaf
Solanaceae Capsicum annuum L. Pepper
Lycopersicon esculentum Mill. Tomato
Tiliaceae Corchorus olitorius L. Nalta jute
Non-cultivated hosts
Amaranthaceae Achyranthes aspera L.
Amaranthus tricolor L.
Asclepiadaceae Leptadenia sp.
Asteraceae Ageratum conyzoides L.
Bidens pilosa L.
Sonchus sp.
Commelinaceae Commelina benghalensis L.
Convolvulaceae Ipomoea cordofana Choisy
Euphorbiaceae Acalypha indica L.
Euphorbia aegyptiaca Boiss.
Euphorbia heterophylla L.
Leguminosae Rhynchosia hirta (Andrews) Meikle & Verdc.
Malvaceae Abutilon pannosum (G. Foster) Schltdl.
Solanaceae Datura stramonium L.
Solanum incancum L.
Solanum nigrum L.
Verbenaceae Lantana camara L.
cause major problems outside of areaswhere cotton is cultivated. Forexample, in the Abu Habil Basin, whereno cotton is grown, tomato is typicallyproduced without whitefly infestation.Similarly, OLCV is present in manyparts of the country where cotton is
grown, whereas WmCSV is present inalmost all melon production areas.
Natural enemy species
Seventeen collections of whiteflyparasitoids and predators were made

122
Whiteflies and Whitefly-borne Viruses in the Tropics
and preliminary identification made.The most common parasitoids ofB. tabaci were found to be Encarsialutea (Masi) and Eretmocerus mundusMercet. The following natural enemiesthat were found in the vegetable-basedsystems in the country also could beinvolved in regulating B. tabacipopulations: the coccinellids Coccinellaundecimpunctata L., Hippodamiavariegata (Goeze), Cheilomenessulphurea (Olivier) and Scymnus sp.;the lacewings Chrysoperla pudica(Navás) and Chrysoperla spp.; piratebugs, Orius sp.; other predatory truebugs Campylomma sp. and spiders.
Increased Socio-economicUnderstanding
Farmers’ assessment ofwhitefly-related problems
More than 95% of vegetable productionin the surveyed areas is carried out onsmall-scale holdings of 2 to 5 ha.Among tomato producers in Sudan,40% farmed their own land, 38% usedrented land, while 22% were squatters.Virtually all farmers interviewed (99%)were men. Almost all the producersinterviewed (94%) regarded tomato asthe most profitable of the horticulturalcrops they could grow and most (74%)have cultivated tomato for more than5 years. The varieties of tomato mostwidely grown in Sudan wereMoneymaker, Strain B, Peto 86,Pearson, Early pack and Ace. Mostproducers (65%) bought their tomatoseed from the local market, while 28%used their own seed and 7% importedseed from other countries. Othercommonly grown vegetable cropsinclude cucurbits (especially melon andcucumber), okra, common bean,pepper, eggplant (Solanum melongenaL.), sweetpotato (Ipomoea batatas [L.]Lam.), radish (Raphanus sativus L.),onion (Allium cepa L.), carrot (Daucus
carota L. subsp. sativus [Hoffm.]Arcang. var. sativus Hoffm.) and leafyvegetables. Most producers (82%)practised crop rotation.
Whiteflies and/or WTVs were foundto be the greatest cause of concern tomost tomato producers in the survey.Other destructive pests reported wereHelicoverpa armigera (Hübner),Liriomyza sativae Blanchard,Scrobipalpa sp. and Keiferialycopersicella (Walshingham).
Most tomato producers in Sudan(75% overall, including all those inCentral Sudan) were able to recognizewhiteflies and even more (89%)recognized TYLCV. However, 65% ofthem did not know that TYLCV andB. tabaci were inter-related. Themajority (84%) believed that whitefliesand/or TYLCV were serious productionconstraints in their farms. Whereas65% of the producers reported thatTYLCV was a problem, only 35%reported that both the whitefly and thedisease were problematic, and nonereported a problem with whitefly alone.In contrast, most producers in Gedaref,Northern and North KordufanProvinces, where TYLCV incidence wasvery low (0%-9%), could not recognizewhiteflies or TYLCV nor did they havelocal names for them.
All producers who recognizedwhiteflies and TYLCV had names forboth and in most cases the producersgave very specific names for theproblems they cause. Local names forwhiteflies include asala, biadah,dubbana, dubbana beida, zubaba andzubaba beida. These literally mean“flies” or “white flies”, except asala,which refers to the honeydew that theinsects secrete. Names given to theTYLC include hurug (burning), saratan(cancer) and karmata, karmasha orkurmut (all of which allude to leafdeformation).

123
Sudan
Historically, Sudan has beenregarded as one of the countries worsthit by whitefly problems, especially inthe cotton production system. Thisscenario has changed recently and thewhitefly problem in cotton seems tohave become less acute. Highlyeffective insecticide treatments, oftencontaining insect growth regulators,have reduced the breeding success ofwhiteflies on cotton. Nonetheless, thewhitefly has maintained its status as ahighly injurious pest of tomato,cucurbits, pepper, okra and commonbean, and the numbers of producerswho have abandoned tomatocultivation because of TYLCV are onthe increase.
Farmers’ perceived yield loss towhitefly and/or TYLCV in tomato wason average 62%. According to thesurvey, 26% of producers, mainly inGezira, reported total yield loss, 38%reported losing three-quarters of theiryield, 10% reported losing half, 10%reported losing one-quarter of theiryield and 16%, mainly from Gedarefand North Kordufan, reported no loss.Of the producers interviewed, 34%(most of them in Gezira State) reportedabandoning their tomato crops in atleast 1 year because of whitefly/TYLVCproblems and most reported that thisoccurred in 1997. The perceived lossescan be compared usefully with therecorded incidence of TYLCVsymptoms. Among all surveyed tomatoproducers, 92% had some virussymptoms in their tomato crop and50% had more than 25% incidence. Itshould be emphasized that 31% of thetomato producers had an 80% TYLCincidence in their tomato crops andthese outbreaks were mainly confinedto the greater Gezira. TYLC wassignificantly less severe (1%-9%) inwestern, northern and eastern states,and absent in Gedaref and the AbuHabil Basin. Most tomato producers(65%) believe that they have whitefly/
TYLCV problems every year. Themajority of producers (86%) believethat there is a direct relationshipbetween the climate and the incidenceof whiteflies and/or whitefly-transmitted disease, with 63% ofproducers associating high incidencewith hot and dry seasons, while only9% believe that they have the problemall year round.
Estimation of diseaseincidence and yield losses
The damage associated with B. tabaciand WTVs is always more serious insummer crops, especially during thehot season from March to August(during which there are three plantingperiods: “early summer” in March, “latesummer” in May and the “mainplanting season” in July) and totalfailure of the tomato crop due to TYLCVis common. This is despite the lowerprevalence of the vector in the summerseason. Whitefly populations build upduring the winter (October-January)and peak in December-January. Up to100% infection by TYLCV can occurduring a mild winter but less yielddepression is associated with thesewinter infections.
In 1998, tomato grown during theMay-July season suffered severe attackby TYLCV and this crop failedcompletely. Watermelon likewisesuffered severe losses from WmCSV.An exceptionally heavy rainy seasonfrom mid-August to mid-October,however, seemed to alter the whiteflysituation later in the year, withpopulation levels dropping sharply andtomato crops grown during October-November being comparatively lessaffected by TYLCV. A field experimentconducted at the University of Gezirarevealed that early infection by TYLCVreduced the total number of flowers perplant, percent fruit setting and thequality of fruits. The infection resultedin 67% reduction in the number of

124
Whiteflies and Whitefly-borne Viruses in the Tropics
mature fruits and 84% reduction in theweight of marketable fruits per plant.Severe infection resulted in 86%reduction in the number of greenmature fruits, 87% reduction in thenumber of ripe fruits and 93%reduction in the weight of marketablefruits per plant. Flower bud initiationsubstantially decreased as the infectionbecame more severe or when infectionoccurred early in the plant’s growthcycle but the proportion of flowerabortion and fruit setting was notsignificantly affected. Affected plantshad reduced foliage cover and fruitswere exposed to direct sunlight, whichcaused white blotches (sunscald) andreduced their market value. The resultsshow that TYLCV infection can affectall yield components, from flowerproduction through to the quantity andquality of fruits.
WmCSV is less importanteconomically than TYLCV but isconsidered to be the most importantsingle cause of high yield loss ofcucurbits in eastern and central Sudanand has affected the country’s melonexports seriously in recent years. Cropsaffected include watermelon, muskmelon and, to a lesser extent, snakemelon (Cucumis melo L. subsp. melovar. flexuosus [L.] Naudin) and squash.Melon producers incur a 64% loss inrevenue because of WmCSV. The virusis prevalent in Gezira, Upper AtbaraRiver and southern Sinnar, where freshmarket varieties such as CharlestonGrey, Congo Red and Sugar Baby aregrown. Watermelon in large areas ofSouthern Blue Nile, Shuwak andDinder inland delta were totallydevastated by WmCSV in1998-99, andup to 80% yield losses occurred inGalia melon (a variety of musk melon)grown for export in Khartoum andGezira States. However, this virus isonly sporadically important inKordufan and western Sudan wherelocal strains of watermelon are grown
for seed production and water storage.WmCSV incidence of 82% was observedon Galia melon grown for commercialpurposes at the Gezira University farmin 1997. Complete absence of “netting”on the fruit surface was evident in someseverely affected plants, while thehealthy ones produced fruits entirelycovered with netting (an importantquality that correlates positively withsweetness of the fruit). The severelyaffected plants gave 1.3 fruits per plant,while healthy plants gave 2.1 fruits perplant. The average incidence of WmCSVin Galia melon from eight sites in theSelait Irrigation Scheme (KordufanProvince) was 70% and data from thesesites indicated a positive correlationbetween the level of virus incidence andyield loss.
OLCV restricts okra production inSudan. Symptoms of the virus appearedto be more severe during the late winterand early summer growing seasons,with Gezira being the worst hit area.The virus had less effect on localvarieties of okra. BMMV may constitutea potential hazard, especially in thenewly established export-orientedvegetable production area in KhartoumState. This virus is less important and isconsidered a late season disease innorthern Sudan.
Pesticide use
The cost of controlling whiteflies andWTVs accounts for more than 30% ofthe total production costs of tomato inSudan. A small proportion ofproducers (4%) spent as much asUS$300-400 per hectare on pesticidetreatment. Other producers estimatedtheir pest control costs at US$200-299per hectare (14% of producers),US$100-199 per hectare (21% ofproducers), US$50-99 per hectare (27%of producers) and US$0-49 per hectare(34% of producers). The costs ofchemicals and their application varygreatly among sites.

125
Sudan
Only 10% of producers reportedreceiving technical advice orinformation regarding the managementof pest and disease problems. Fortypercent of producers received advicefrom other vegetable producers orneighbours, 30% from commercialsales agents, while 20% had notreceived any advice but appliedinsecticides according to their ownjudgement. The absence of an effectiveextension service has left the farmersto make their own decisions onwhitefly/WTV management. Theconstraints on farmers’ knowledge ofwhitefly management can besummarized as:
(1) Low level of education and limitedknowledge of modern techniques ofvegetable production;
(2) Poor ability to choose the right cropand a suitable variety of that crop;
(3) Poor knowledge on pesticide usageand related hazards; and
(4) Inability to monitor market trendsas a basis for deciding what togrow and when to grow it.
Because of the lack of extensionservices in vegetable farming, chemicalcontrol of the whitefly remains theprincipal pest management strategy.The great majority of tomato producersin the country (80%) used insecticides,mostly pyrethroids andorganophosphates, to combat thewhitefly/TYLCV problem on theirfarms. The most popular pesticidesapplied by the producers werefenpropathrin, fenvalerate andcypermethrin (pyrethroids), omethoate,malathion, dimethoate andchlorpyriphos (organophosphates),methomyl and carbaryl (carbamates),and endosulfan (organochlorine).
Most producers believed that thepesticides were indispensable in
tomato farming. A significantproportion of producers (30%) mademore than 10 insecticide applicationsper season, 26% made 9 or10 applications, and a few producersapplied pesticides as many as 24 to30 times per season. Ten percent ofproducers practiced crop rotation tocombat the problem. A few farmersused cultural control methods such asburying of infected plants and inter-cropping with repellent crops such ascoriander (Coriandrum sativum L.) andfenugreek (Trigonella foenum-graecumL.). Others claimed that directly sowntomato, that is, without transplanting,was less affected by TYLCV. Tenpercent of producers, mainly inGedaref, Western State and NorthernState did not practice any sort ofmanagement of the problem.
Lack of knowledge of the properuse of pesticides was evident in allareas surveyed. Farmers were found tobe ignorant of the judicious use ofpesticides and alternative whiteflycontrol methods. Some tomato growersoverused chemicals, while others didnot spray until symptoms of damagewere observed, and subsequentintensive chemical application failed tosave the crop. Producers were ignorantof the interval required between thelast spraying and harvest, and someeven used the taste or smell of thepesticides to judge their effectiveness.Lack of protective clothing andequipment exposed the farmers toserious health hazards. Inappropriatechemicals such as fungicides(propiconzale) and others without anylabels were being used in an attempt tocontrol the whitefly and WTV problem.Recommended dosages were notfollowed and virtually all vegetableproducers mixed one tomato sauce tin,containing about 50 mL, of anyinsecticide, irrespective of itsformulation or active ingredient, with4 gallons (18 L) of water. A few

126
Whiteflies and Whitefly-borne Viruses in the Tropics
producers used twigs to apply pesticide,instead of a knapsack sprayer, resultingin poor coverage and greater loss ofpesticides. Most producers (67%)applied insecticides as a preventivemeasure, 17% applied insecticideswhen they observed whitefly/TYLCVdamage, while 8% applied insecticidesaccording to calendar.
Vegetable growers in Gezira andRahad illegally obtain part of theirsupply of chemicals from the stocks ofcotton schemes. These include highlytoxic “cocktails” of insecticide,exclusively recommended for use oncotton. Their experience of severewhitefly outbreaks in previous seasonsleads most of them to use excessivequantities of these chemicals. As aresult, environmental contamination,occupational health hazards andproduction costs are on the increase. Inaddition, the overuse of chemicals tocontrol whitefly seemed to have resultedin the build-up of previously secondarypests such as H. armigera, L. sativaeand Scrobipalpa sp. Nevertheless,several producers suffered completeloss of even the treated crop.
Several researchers (e.g., Ahmed etal., 1987; Dittrich et al., 1990) havedocumented the resistance status ofB. tabaci in cotton to variousinsecticides in the Sudan. Theyreported that B. tabaci showed highlevels of resistance to manyorganophosphates and syntheticpyrethroids and moderate resistance tocarbamates and organochlorines. Thespecific levels of resistance reportedvary depending on the source of thesamples, the history of insecticideapplication in that area, the time of theyear when bioassays were done and thesensitivity of the whitefly populationused. However, the reports wereprimarily based on dose responses ofwhiteflies in the cotton monoculturesystem. Pesticide resistance of
whiteflies in the mixed vegetablecropping systems in the Sudan has notbeen documented.
Sudan is the only country in EastAfrica that has adopted IPM as itsofficial crop protection policy. Thegovernment’s position is well defined ina recent publication entitled “SudanCountry Strategy Note, 1997-2001:Partnership towards SustainableHuman Development”. The documentemphasizes that the government of theSudan is pursuing an integratedprogram for environmentallysustainable development. However,implementation of the policy is minimal,particularly, in the case of vegetablefarming, because of farmers’ ignoranceof alternatives to chemical control,which in turn is mainly attributed to thelack of effective extension services inthis sector. In the absence ofalternatives, chemical control of thewhitefly therefore remains the principalstrategy for managing WTVs.
Strengthened ResearchCapacity
Sudan’s participation in the diagnosticphase of the TWF-IPM Project hasprovided a substantial body ofknowledge covering various aspects,biological and socio-economic, of thewhitefly/WTV problems in vegetable-based cropping systems in the country.Sudan’s participation in the planningworkshop in April 1997 and theco-ordination meeting in June 1998provided national researchers with achance to learn from, and share ideasand experiences with, scientists fromother countries. A Ph.D. project on“Distribution of whitefly species (andbiotypes) and their natural enemies invegetable-based cropping systems inSudan” was undertaken in collaborationwith the University of Gezira under theauspices of the project.

127
Sudan
Conclusions
The work conducted during thediagnostic phase of the TWF-IPMProject has provided, for the first time,a biological and socio-economiccharacterization of the problems linkedto B. tabaci in the vegetable croppingsystems of Sudan. The studies haveprovided a better understanding of thebiology of this important vector, thedistribution of its natural enemies, theepidemiology of the viruses it transmitsand the socio-economic implications ofvirus disease problems for producers.
Farmers viewed the B. tabaci/TYLCV problem as the most importantconstraint on tomato production. Theareas identified as hot spots for TYLCVinclude Gezira, Managil, Umelgura andHasaheisa Provinces, all of which arecharacterized by irrigated productionand are within cotton production areas.The main problems associated withB. tabaci in vegetable productionsystems in Sudan are:
(1) Tomato: transmission of TYLCVand TVTV, and direct feeding byB. tabaci during early crop stages;
(2) Watermelon and musk melon:transmission of WmCSV, CVYV,and a virus inducing potyvirus-likeyellowing symptoms;
(3) Okra: transmission of OLCV, sootymould induced by secretion ofhoneydew by B. tabaci, and directfeeding damage;
(4) Peppers: transmission of TYLCV;and
(5) Vegetable legumes: symptoms of abegomovirus and closterovirusobserved on cowpea and commonbean but transmission by B. tabaciis not yet confirmed.
The surveys documented alarmingmisuse of pesticides on vegetables. Theinformation gained can help tosensitize policy makers in Sudan to thealarming magnitude of the whitefly andWTV problems and to the dangersinherent in the associated misuse ofpesticides. The knowledge gained hasprovided a sound foundation fordeveloping a future research agenda.The following priorities arerecommended:
(1) Research should be conducted onthe following IPM components:
(a) Evaluation of availableTYLCV-resistant or -toleranttomato varieties for immediateincorporation into IPM plans;
(b) Screening of exotic tomatovarieties for resistance toB. tabaci and/or TYLCV;
(c) Breeding for resistance againstB. tabaci and WTVs, whileutilizing resistant strains ofcrops;
(d) Evaluation of alternative andnovel pest and diseasemanagement methods,including the use of botanicals,biological control and culturalmethods; and
(e) Investigation of the efficiency ofnatural enemies in vectorcontrol.
(2) Available IPM options should betested and refined, with the closeparticipation of producers, underon-farm conditions; participatoryresearch should be complementedwith intensive training of producersand other stakeholders.
(3) Resistance of whiteflies toinsecticides commonly used invegetable farming should be

128
Whiteflies and Whitefly-borne Viruses in the Tropics
monitored, using a standardizedmethodology in Sudan andelsewhere to enable validcomparisons to be made.
(4) Appropriate techniques to evaluateinsecticide residues in fruits andvegetables should be developed andapplied.
(5) The whiteflies observed inducingsilverleaf in Gezira for the first timein 1998 should be characterizedusing molecular techniques andbiological assays.
The recommended research willprovide a platform for the developmentand adoption of sustainable productionpractices for vegetables in Sudan,leading to improved well-being forproducers and consumers.
References
Ahmed, A. H. M.; Elhang, E. A.; Bashir,H. H. 1987. Insecticide resistance inthe cotton whitefly (Bemisia tabaciGenn.) in the Sudan Gezira. Trop.Pest Manag. 33:67.
Dafalla, G. A.; Sidig, S. A. 1997.Management of the whitefly-viruscomplex in Sudan. Food andAgriculture Organization (FAO)Plant Production and ProtectionPaper 143. Rome, IT. 214 p.
Dittrich, V.; Ruesch, O.; Ernest, G. H.1990. Resistance mechanism in thesweetpotato whitefly (Homoptera:Aleyrodidae) populations fromSudan, Turkey, Guatemala andNicaragua. J. Econ. Entomol.83:1665-1670.
Eveleens, K. G. 1983. Chemical insectcontrol in the Sudan Gezira:Analysis of a crisis crop. Crop Prot.2:273-287.
Kirkpatrick, T. W. 1931. Further studieson leaf-curl of cotton in the Sudan.Bull. Entomol. Res. 22:323-363.
Shapiro, J. P. 1995. Insect-plantinteractions and expression ofdisorders induced by the silverleafwhitefly, Bemisia argentifolii. In:Gerling, D.; Mayer, R. T. (eds.).Bemisia: Taxonomy, biology,damage, control and management.Intercept, Andover, GB. p. 167-171.
Yassin, A. M.; Nour, M. A. 1965. Tomatoleaf curl disease, its effects on yieldand varietal susceptibility. SudanAgric. J. 1:3-7.

129
Kenya
CHAPTER 2.3
KenyaMohamed Ali Bob*, Benjamin Odhiambo**,
Gilbert Kibata** and Jason Ong’aro**
Introduction
Geographical context
Field surveys covering 94 farms wereconducted in 1997-98 following themethodology agreed among projectpartners. The farms represented:Kirinyaga, Thika, Muranga, Kiambu,and Nairobi Districts in CentralProvince; Machakos, Athi River, Meru,Kitui, and Kibwezi Districts in EasternProvince; Kwale, Kilifi, and Taita TavetaDistricts in Coast Province; Kajiadoand Naivasha Districts in Rift ValleyProvince; Kisii, Migori, Homa Bay, andSiaya Districts in Nyanza Province; andVihiga and Busia Districts in WesternProvince (Figure 1). The districts wereselected based on the prevalence of thecultivation of tomato (Lycopersiconesculentum Mill.). Table 1 gives themean annual temperatures andprecipitation for each of the surveyedregions.
In the surveyed regions, tomato isplanted throughout the year with theexceptions noted below. EasternProvince is a region of semi-arid lowermid-altitude land becoming transitionalsemi-humid towards Central Province;here, tomato is not planted in July.
* International Center of Insect Physiology andEcology (ICIPE), Nairobi, Kenya.
** Kenya Agricultural Research Institute (KARI),Nairobi, Kenya.
Towns
Surveyedareas
Eastern
Kenya
NortheasternWestern
Central
Nairobi
Nakuru
Kitale
Kisumu
Figure 1. Tomato-growing areas surveyed forwhitefly incidence in Kenya.
Table 1. Mean annual temperatures andprecipitation for the surveyed regionsof Kenya.
Region/ Mean min.-max. MeanProvince temperature precipitation
(°C) (mm)
Eastern 15-26 728
Rift Valley 14-27 591
Central 13-25 1166
Coast 21-30 872
Western and 16-29 1289Nyanza
Whitefly surveys took place betweenNovember and March of 1997-98 and1998-99. Rift Valley Province consists

130
Whiteflies and Whitefly-borne Viruses in the Tropics
of transitional lower to higher mid-altitude land. In the surveyed areas ofKajiado and Naivasha Districts in theRift Valley, tomato is not planted inJanuary and July. Whitefly surveyswere carried out in September 1998and March 1999. Central Provinceconsists of sub-humid highlands;whitefly surveys were carried out herebetween November and March in1997-98 and in 1998-99. In the CoastProvince, a region of sub-humidcoastal lowlands, tomato is notplanted from November to January.Whitefly surveys were carried out inJuly 1998. In Western and NyanzaProvinces (sub-humid to humid uppermid-altitude land and lower highlands)tomato is not planted in January.Whitefly surveys in this region werecarried out in February and November1998.
El Niño rains devastated tomatoproduction in Kenya from September1997 to June 1998. Whiteflies werescarce during this period.
The emergence of Bemisiatabaci as a pest and virusvector
Tomato is one of the most widelycultivated vegetable crops in EastAfrica, predominantly grown by small-scale producers for fresh consumption(GTZ, 1995), while in Kenya inparticular, vegetable-growing both forlocal consumption and for exportprovides an increasingly importantsource of income to farmers. In 1995,Kenyan national scientistsparticipating in a regional workshopranked the whitefly Bemisia tabaci(Gennadius) as the most importantpest/vector in tomato (Varela andPekke, 1995). This whitefly hasbecome an important productionconstraint to crops of the familiesSolanaceae, Leguminosae, Malvaceae,
Cucurbitaceae and Euphorbiaceae,particularly in the semi-arid EasternProvince of Kenya. Extension workersand scientists have reportedinformally the occurrence of severalwhitefly-transmitted viruses: Tomatoyellow leaf curl virus (TYLCV), Cowpeamild mottle virus (CPMMV),Sweetpotato mild mottle virus (SPMMV)and Watermelon chlorotic stunt virus(WmCSV).
Despite the perceived importanceof these horticultural crops and theirvirus disease problems in Kenya,previously no systematic attempt hasbeen made to document theirdistribution and incidence. Virtuallyall work related to whitefliesconducted in Kenya has focused oncassava mosaic disease (CMD), itsprevalence in the country, itstemporal and spatial distribution andassociated yield loss (Seif, 1982; Bock,1987). Some research also had beenconducted on the citrus woollywhitefly Aleurothrixus floccosus(Maskell), which is reported to havebecome a common pest of citrus(Citrus spp. L.) as well as coffee(Coffea arabica L.) and guava (Psidiumguajava L.) in Kenya (Löhr, 1997). Inaddition, the whitefly parasitoids,Eretmocerus mundus Mercet. andEncarsia sublutea (Silvestri), havebeen recorded in Kenya (Dr. AbdurabiSeif, personal communication, 1998).Kenya’s participation in the TropicalWhitefly Integrated Pest Management(TWF-IPM) Project provided anopportunity to gain an overview of thegeographical range and economicimportance of whiteflies, whitefly-transmitted viruses and whiteflynatural enemies in the country—information that would serve as asound basis for planning furtherresearch and pest managementefforts.

131
Kenya
Increased BiologicalUnderstanding
Characterization ofbegomoviruses and whiteflybiotypes
During the surveys in Kenya,325 collections of whitefly adults andnymphs were made. From these,549 specimens of whitefly nymphswere mounted on 310 slides andidentified to species level. Most of thewhiteflies collected were B. tabaci(57%), while Trialeurodes vaporariorum(Westwood) represented 34%. B. afer(Priesner and Hosney) wereoccasionally found (7%) in fields ofcassava (Manihot esculenta Crantz)adjacent to tomato fields. A number ofother whitefly species (2%) also wereencountered, including Trialeurodesricini (Misra), A. floccosus, Siphoninusphillyreae (Haliday) and Bemisia spp.Table 2 shows the composition ofwhitefly species based on the identifiedspecimens from each Province.
Disease incidence andsymptom severity
Whiteflies in the surveyed farms werefound breeding on host plants thatbelong to the following families:Acanthaceae, Amaranthaceae,Asteraceae, Commelinaceae,Convolvulaceae, Cucurbitaceae,Euphorbiaceae, Lamiaceae,Leguminosae, Malvaceae, Rutaceae,
Solanaceae and Verbenaceae (Table 3).Surprisingly, whiteflies were foundbreeding only occasionally on tomato inKenya. In Muranga, Kiambu and MweaDistricts in Central Province, it wasdifficult to find a single whitefly nymphin a whole tomato field. In these areas,whiteflies, mainly T. vaporariorum, werefound breeding on crops such ascommon bean (Phaseolus vulgaris L.)and cassava and on non-cultivatedplants, most significantly Euphorbiaheterophylla L., in all areas surveyed.However, in Kihara (Nairobi) andKigumo (Muranga) in Central Provinceand in Nguruman in Rift ValleyProvince, whiteflies, mainlyT. vaporariorum, were found breedingreadily on tomato.
Host plant feeding preference andstatus as reproductive hosts(oviposition and offspring survival) ofB. tabaci was studied in field cages atthe International Center of InsectPhysiology and Ecology (ICIPE),Nairobi, in 1997 using adult insectscollected from a tomato field nearby.Among the four host plants studied,common bean was the most preferredhost plant, followed by lablab (Lablabpurpureus [L.] Sweet), cowpea (Vignaunguiculata [L.] Walp.) and okra(Abelmoschus esculentus [L.] Moench).There were significant differencesbetween plants with regard to theparameters tested. These resultsconcur with observations in the fieldduring the survey.
Table 2. Species composition (%) of whitefly samples from surveyed regions of Kenya.
Province Species compositiona
B.t. T.v. B.a. T.r. A.f. S.p.
Eastern 61 29 6 0 0 4
Rift Valley 20 80 0 0 0 0
Central 35 60 4 1 0 0
Coast 79 0 21 0 0 0
Western 90 0 7 3 0 0
Nyanza 79 6 5 0 10 0
a. B.t., Bemisia tabaci; T.v., Trialeurodes vaporariorum; B.a., Bemisia afer; T.r., Trialeurodes ricini;A.f., Aleurothrixus floccosus; S.p., Siphoninus phillyreae.

132
Whiteflies and Whitefly-borne Viruses in the Tropics
Table
3.
Rep
rodu
ctiv
e h
ost
pla
nts
of w
hit
efly
spec
ies
enco
un
tere
d d
uri
ng
surv
eys
in K
enya
.
Wh
itef
ly s
pec
ies
Cro
ps
Non
-cu
ltiv
ated
hos
ts
Fam
ily
Spec
ies
Fam
ily
Spec
ies
Bem
isia
ta
ba
ci (G
enn
adiu
s)C
onvo
lvu
lace
aeIp
omoe
a b
ata
tas
(L.) L
am.
Aca
nth
acea
eA
syst
asi
a s
chim
per
i T.
An
der
s.C
ucu
rbit
acea
eC
itru
llus
lan
atu
s (T
hu
nb.) M
atsu
m &
Nak
aiA
mar
anth
acea
eA
chyra
nth
es a
sper
a L
.C
ucu
rbit
a p
epo
L.
Ach
yra
nth
es s
icu
la L
.M
omor
dic
a c
ha
ran
tia
L.
Ast
erac
eae
Age
ratu
m c
onyzo
ides
L.
Eu
ph
orbia
ceae
Ma
nih
ot e
scu
len
ta C
ran
tzA
spili
a m
ossa
mbic
ensi
s (O
liv.
) W
ild
Leg
um
inos
aeC
rota
lari
a s
p.
Bid
ens
pilo
sa L
.La
bla
b p
urp
ure
us
(L.) S
wee
tE
mili
a d
isci
folia
(O
liv.
) C
. Jef
frey
Vig
na
un
guic
ula
ta (L.) W
alp
.G
alin
soga
pa
rvif
lora
Cav.
Cro
tala
ria s
p.
Gu
ten
ber
gia
cor
dif
olia
Ben
th. ex
Oliv.
Tit
hon
ia d
iver
sifo
lia (H
emsl
.) A
. G
ray
Ph
ase
olu
s vu
lga
ris
L.
Com
mel
inac
eae
Com
mel
ina
ben
gha
len
sis
L.
Lam
iace
ae
Oci
mu
m b
asi
licu
m L
.C
onvo
lvu
lace
aIp
omoe
a tri
colo
r C
av.
Mal
vace
aeA
bel
mos
chu
s es
cule
ntu
s (L
.) M
oen
chJa
cqu
emon
tia
tam
nif
olia
(L). G
rise
b.
Gos
sypiu
m h
irsu
tum
L.
Eu
ph
orbia
ceae
Eu
ph
orbia
het
erop
hylla
L.
Sol
anac
eae
Ca
psi
cum
an
nu
um
L.
Leg
um
inos
aeR
hyn
chos
ia h
irta
(A
ndre
ws)
Mei
kle
& V
erdc.
Lyco
per
sico
n e
scu
len
tum
Mill.
Lam
iace
ae
Leo
not
is n
epet
ifol
ia (L.) R
. B
r.S
ola
nu
m m
elon
gen
a L
.O
cim
um
kili
ma
nd
sha
ricu
m G
uer
ke
Mal
vace
aeS
ida
acu
ta B
urm
f.
Sol
anac
eae
Da
tura
str
am
oniu
m L
.N
ica
nd
ra p
hysa
lod
es (L.) G
aer
tn.
Ver
ben
acea
eLa
nta
na
ca
ma
ra L
.A
ster
acea
eTit
hon
ia d
iver
sifo
lia (H
emsl
.) A
. G
ray
Tri
ale
uro
des
va
por
ari
oru
m (W
estw
ood)
Cu
curb
itac
eae
Cu
curb
ita s
pp.
Eu
ph
orbia
ceae
Eu
ph
orbia
het
erop
hylla
L.
Mom
ord
ica
ch
ara
nti
a L
.Leg
um
inos
aePh
ase
olu
s vu
lga
ris
L.
Mal
vace
aeA
bel
mos
chu
s es
cule
ntu
s (L
.) M
oen
chS
olan
acea
eLyco
per
sico
n e
scu
len
tum
Mill.
Sol
an
um
mel
onge
la L
.
Bem
isia
afe
r (P
ries
ner
an
d H
osn
ey)
Con
volv
ula
ceae
Ipom
oea
ba
tata
s L. Lam
.Leg
um
inos
aePh
ase
olu
s vu
lga
ris
L.
Mal
vace
aeG
ossy
piu
m h
irsu
tum
L.
Ast
erac
eae
Tit
hon
ia d
iver
sifo
lia (H
emsl
.) A
. G
ray
Tri
ale
uro
des
ric
ini (
Mis
ra)
Sol
anac
eae
Lyco
per
sico
n e
scu
len
tum
Mill.
Lam
iace
ae
Leo
not
is n
epet
ifol
ia (L.) R
. B
r.
Ale
uro
thri
xus
floc
cosu
s (M
ask
ell)
Ru
tace
ae
Cit
rus
spp.
Sip
hon
inu
s ph
illyre
ae
(Haliday)
Un
iden
tified
tre
e

133
Kenya
Samples of 10 plants per field, onaverage, collected from all surveyedareas, were squashed on nylonmembranes and sent to the John InnesCentre, UK, for hybridization.Hybridization signals varied fromstrong to very weak, probably becauseof differences in the degree of nucleicacid homology. This could suggestdifferent strains of begomoviruses ordifferent begomoviruses altogether. Thesignal strength also may have beeninfluenced by the sampling method,because the virus titre may have beenlow in the particular tissues sampled orat that stage of infection.
At Kibwezi, begomovirus symptomswere observed on sweet pepper(Capsicum annuum L.) and a lowincidence of WmCSV (about 5%) wasrecorded on watermelon (Citrulluslanatus [Thunb.] Matsum. & Nakai). Avariety of mosaic symptoms that needfurther investigation also were noticedin pepper and watermelon. There weresymptoms that suggest that tomato inthis region should be tested forpotyviruses such as Potato virus Y(PVY). Aphid infestation levels werevery high at Kibwezi and some of thesymptoms observed could be a result ofaphid-borne viruses. High incidenceand severe symptoms of CMD wereobserved in Mombasa and KilifiDistricts (Coast Province) and in KituiDistrict (Eastern Province). Thesymptoms appeared more severe thanthose observed in western Kenya.Whether these symptoms are causedby infections of East African cassavamosaic virus (EACMV) or are associatedwith the occurrence of the Ugandanvariant needs to be ascertained.
Leaf curl symptoms were foundduring the survey on non-cultivatedhost plants. These were Achyranthesaspera L., E. heterophylla and Nicandraphysalodes (L.) Gaertn. These plantsare therefore likely to act as reservoirs
of TYLCV. Severe begomovirussymptoms also were observed on aweed identified as Gutenbergiacordifolia Benth. ex Oliv. E.heterophylla was found to be a majorreproductive host of both B. tabaci andT. vaporariorum at any site wherewhitefly infestation occurs and thisplant species is present. Therefore, therole of E. heterophylla in survival of thedisease and the vector outside thecrop-growing season needs to beinvestigated.
Natural enemy species
Forty whitefly parasitoid samples wereobtained from the surveys andpreliminary identification was carriedout. Encarsia sophia (Girault and Dodd)was found to be the predominantspecies, while Eretmocerus sp. wasencountered in the Central Province.Encarsia formosa Gahan was onlycollected from Mwea in the CentralProvince. At Nguruman, Rift Valley, thelevel of parasitization appeared higherthan at other survey sites in Kenya.
Some of the 24 predators collectedwere found feeding on the whiteflypupae. Most were coccinnellids, while afew were spiders and mites but nonewere identified to species level.Macrolophus caliginosus Wagner, bugsknown to prey on T. vaporariorum inother parts of the world, were collectedat several locations in Nguruman (RiftValley Province), Kibwezi (Easternprovince) and Mwea (Central Province).
Geographical range of whiteflyand whitefly-transmitted virusinfestation
Areas identified as “hot spots” forTYLCV infection include Kibwezi, Kitui,Machakos, Athi River (EasternProvince) and Naivasha (Rift ValleyProvince). Low incidence of TYLCVsymptoms was observed in all othersurveyed provinces with the exception

134
Whiteflies and Whitefly-borne Viruses in the Tropics
of Western and Nyanza Provinces,where no TYLCV symptoms wereobserved.
B. tabaci was found to be the mostcommon whitefly species in the country(Table 3). T. vaporariorum was commonin the highlands and was encounteredfrequently in the Central, Rift Valleyand Eastern Provinces. T. vaporariorumwas not recorded from the Coastal andWestern Provinces. The abundancepattern and economic importance ofB. tabaci/TYLCV differed significantlybetween provinces. It was found toattain particularly high epidemicincidence (>25% of the tomato withTYLCV symptoms) in Eastern Province(Kibwezi 30%-100%, Kitui 30%-100%,Machakos and Athi River 15%-30%)and in Rift Valley Province (Naivasha15%-40%), where whiteflies and TYLCVwere of great concern to the tomatoproducers. This suggests that theimpact of the B. tabaci/TYLCV complexis generally more severe in the dryareas.
In Coast Province, where theweather is hot and humid throughoutthe year, B. tabaci constituted 79% ofthe collected whitefly specimens.However, only a moderate incidence ofTYLCV symptoms (5%-25%) wasobserved. In Western Province,B. tabaci constituted 90% of thecollected whitefly specimens and inNyanza Province, 79% (weather is alsohot and humid throughout the year)but no TYLCV symptoms were observedon tomato, other vegetable crops, orweeds, and collected samples werenegative when hybridized using a probeto the original Israeli isolate of TYLCV.In Western and Nyanza Provinces, mostproducers plant tomato as a secondarycrop after the main staple cereal crops,maize (Zea mays L.) and sorghum(Sorghum bicolor [L.] Moench). TheB. tabaci population at Kibwezi(Eastern Province) is usually very high
from December to early March (hot anddry season) but the whiteflies virtuallydisappear from April to August (longrainy season followed by cold season),despite the presence of the same cropsin the irrigated fields throughout theyear.
High T. vaporariorum populationswere observed in Nguruman (Rift ValleyProvince) where the underside of leavesof tomato, common bean and eggplant(Solanum melongena L.) could be foundcovered with adults and nymphs. Inthis area, the infestation level onlydrops during a brief rainy season,around April-May. A similar level ofinfestation by the same whiteflyspecies was observed in the relativelycooler area of Kihara (CentralProvince). Direct feeding damage byT. vaporariorum was observed in bothCentral Province (Kihara and Mwea)and Rift Valley Province (Nguruman),where it was the predominant whiteflyspecies.
Since completion of the survey,very high whitefly infestations havebeen reported in the cut-flower andornamental plants on export-orientedflower and vegetable farms aroundLake Naivasha (Rift Valley Province).The highest TYLCV incidences wereobserved at altitudes ranging fromabout 500 to 1000 m altitude (Table 4).B. tabaci was the dominant whiteflyspecies up to an altitude of about1700 m, above which T. vaporariorumbecame dominant.
Increased Socio-economicUnderstanding
Farmers’ assessment ofwhitefly-related problems
Smallholders dominate vegetableproduction in Kenya and account for70% of the total production (Mulandi,1998). Producers in the survey areas

135
Kenya
were surveyed using agreed protocols.Most of the interviewed producers(75%) used their own land, while 13%used rented lands and 12% had otherarrangements. The great majority ofproducers (80%) were men.
Tomato is one of the mostimportant vegetable crops in thecountry. Virtually all interviewedproducers (98%) regarded tomato astheir most profitable vegetable crop and58% of the producers interviewed havecultivated tomato for more than5 years. Main varieties of tomato grownin the country were Moneymaker andCal-J, while other varieties includeRoma, Marglobe, Petomech, EarlyBeauty, Mecheast, Bonny Best, Heinz,Floradade, Marmande, Ponderosa andHotset. However, no tomato varietygrown in Kenya was known to beresistant or tolerant to TYLC. Mostproducers (75%) bought their tomatoseed from the local market, while 20%used their own seed and 5% reportedbuying directly from the seedcompanies. Other vegetable cropsinclude kale (Brassica oleracea L.),common bean, sweet pepper, cabbage(Brassica oleracea var. capitata L.),eggplant, sweetpotato (Ipomoea batatas[L.] Lam.), okra, bitter gourd(Momordica charantia L.), melon(Cucumis melo L.) and onion (Allium
cepa L.). Most producers (94%)practised crop rotation.
Fifty-five percent of the interviewedproducers listed whiteflies and/orassociated virus diseases among theirprincipal pests and diseases andranked these problems, on average, astheir third-most-important problem intomato. For tomato producers inKenya, late blight Phytophthorainfestans (Mont.) de Bary is the maincause of concern as a yield-limitingfactor. Other pest and diseaseproblems were early blight (Alternariasolani [Ell. and Mart.] Jones andGrout), red spider mites (Tetranychusurticae Koch.), bollworms (Helicoverpaarmigera [Hubner]), leaf miners(Lyriomyza spp.), bacterial wilt (Erwiniatracheiphila [Smith]), aphids, cutworms(Spodoptera litura [Fabricius]),nematodes (Rhabditida:Steinernematidae & Heterorhabditae),end rot (Godronia cassandrae f.vaccinii), thrips (Thrips palmi Karny)and rats (Rattus sp.).
Most producers (88%) were able torecognize whiteflies, while only 64% ofproducers were able to recognize thesymptoms of TYLCV and only 1% knewthat whiteflies and TYLCV were inter-related. Although only about half of theproducers (55%) listed whiteflies and
Table 4. Maximum incidence (% of sampled plants showing disease symptoms) of Tomato yellow leafcurl virus (TYLCV) encountered and species composition (%) of whitefly samples collected atdifferent altitudes in Kenya.
Altitudea (m) TYLCV Whitefly species compositionb
incidence B.t. T.v. T.r. B.a. S.p. A.f. B. spp.
20-270 8 72 6 0 14 8 0 0
520-770 100 81 6 0 13 0 0 0
770-1020 80 33 65 2 0 0 0 0
1020-1270 30 66 25 1 8 0 0 0
1270-1520 15 81 16 0 1 0 0 2
1520-1770 27 77 9 2 5 0 7 0
1770-2020 39 43 53 0 4 0 0 0
a. No survey was conducted in the 270-520 altitude range.b. B.t., Bemisia tabaci; T.v., Trialeurodes vaporariorum; T.r., Trialeurodes ricini; B.a., Bemisia afer;
S.p., Siphoninus phillyreae; A.f., Aleurothrixus floccosus; B. spp., Bemisia spp.

136
Whiteflies and Whitefly-borne Viruses in the Tropics
associated viruses among theirprincipal pests and diseases, 79% ofinterviewed producers believed thatwhiteflies cause problems in the cropson their farms. Among theseproducers, 50% attributed theproblems to both whiteflies and TYLCV,47% attributed the problem towhiteflies only, while only 3%mentioned TYLCV alone.
Areas where producers were notable to recognize whiteflies were mainlyin Western Province (Funyula, Mokonjaand Buhuma), Nyanza Province (SouthUgenya and Nyabondo) and CoastProvince (Shimba Hills and Kikambala),where whiteflies were scarce. Mostproducers in these localities also couldnot recognize TYLCV symptoms andwere not aware of its association withthe vector. Some thought that TYLCVwas either a soil-borne or seed-borneproblem, while some confused it withblight.
Most producers who recognizedwhiteflies and TYLCV symptoms hadnames for both but in most cases theproducers gave non-specific names forwhiteflies (e.g., “insects”) and forTYLCV (e.g., “disease”). Most of thenames given to the whiteflies literallymean “flies” or “small insects” and localnames include ebichuni (insect),keguneta, kimbulutwa (butterfly), kudni(insect), mbuu (mosquito), ngaturia,okogataa (mites), oulolo (white moulds),rwagi rweru (white mosquito), twihurutatweru (small white butterflies), ume(aphids), umuu (moulds), vipuvutevidogo and obororo (plant fleas). Namesgiven to the disease problem mean “leafcurling”, “folding” or “stunted plantgrowth” and local names include chanamatawi (combing of leaves), gathuri(stunted old man), gikware, gukunjamathangu (folding of leaves), kanyaria(the devastating disease), kugogonyara(curly) and mbaa (blight).
Estimation of diseaseincidence and yield losses
Among the surveyed farms, 55% hadTYLCV symptoms in their tomato crop.However, very low incidences (1%infection) were found in most parts ofCentral, Eastern and CoastalProvinces. Incidence of TYLCVsymptoms of more than 30% wasrecorded in 11 out of 94 farms. Thesewere in Kibwezi (3), Kitui (3),Machakos/Athi River (1) and Naivasha(4). High incidence (80%-100%) wasonly recorded in two farms in EasternProvince, in Kibwezi and Kitui areas.The incidence in Kibwezi resulted in atotal loss of the tomato crop. Severeleaf curling plus subsequent yielddepression of nearly 100% in sweetpepper was also observed in oneexperimental plot in Kibwezi. Despitethe presence of B. tabaci in westernKenya (Western and Nyanza Provinces),no symptoms of TYLCV infection wererecorded in this part of the country.
On average, producers’ estimate ofthe yield loss in tomato production was30%. According to the survey, 1%reported a total yield loss, 5% reportedthree-quarter yield loss, 26% reportedhalf yield loss and 50% of theproducers reported one-quarter yieldloss due to the whitefly/TYLCVcomplex. The problem was moreserious in drier areas where irrigationwas practised and this could bebecause of the presence of TYLCVreservoirs and the virus vector.Eighteen percent of the producersinterviewed, mainly from WesternProvince, thought that their tomatocrops did not suffer any yield lossbecause of the whitefly/TYLCVcomplex. However, 8% of producersinterviewed reported abandoning theirtomato crops in at least 1 year becauseof the problem. These were fromEastern, Rift Valley, Central and CoastProvinces. Half of them reported thatthe loss occurred in 1997.

137
Kenya
Seventy-nine percent of tomatoproducers believe that they encounterthe whitefly/TYLCV complex everyyear. Almost one-third of the producers(30%) believed that the incidence andseverity of whitefly-transmitted viruses,especially in tomato, was exceptionallyhigh in early 1997 before the El Niñophenomenon. Most (89%) also believethat there is a direct relationshipbetween the seasonal weather changesand the whitefly/disease incidence.Seventy-five percent of producersreported that the period fromDecember to March (hot and dryseason) was the time when theyexperience the most serious whitefly/TYLCV problem, while few believed thatthey have the problem all year round.Producers gave estimation of the costsinvolved (see below).
Costs estimated by producersin control of whitefly/TYLCV
complex per hectare of tomato
US$ % producers0-49 1350-99 17
100-199 30200-299 17300-400 23
Pesticide use
Almost half of the producers (43%)received advice and recommendationson the use of insecticides fromtechnical advisors, 37% from otherproducers, neighbours or familymembers, 10% from pesticidessalesmen, while 10% claimed to usechemicals on their own initiative. Mostproducers (85%) made their owndecision on what, when and whichinsecticides to apply, while only 10%relied on the technical advice theyreceived. Half of the producers (48%)applied insecticides as a preventivemeasure, 25% applied insecticideswhen they observed whitefly/TYLCVdamage, while 18% applied insecticides
routinely according to the calendar.The high level of insecticide usagecoupled with prophylactic chemicalspraying could easily lead to thewhiteflies developing resistance, whichwould hamper their control.
Because of the lack of virus-resistant tomato cultivars, producersresort to the use of pesticides tocombat the whitefly/TYLCV complex.The use of pesticides in mostproduction areas in Kenya hasincreased, even though it is hazardousand sometimes ineffective anduneconomical. Eighty-six percent oftomato producers used insecticides,mostly pyrethroids andorganophosphates, to combat thewhitefly problem on their farms.However, 9% of the producers did notpractice any sort of control against theproblem, while 4% practised croprotation as a means of combating it.Some producers used a Bacillusthuringiensis formulation (Dipel) forcontrol of whiteflies. One producer inMwea (Central Province) reported usingash, in addition to commercialchemicals, to manage the whitefly/TYLCV complex on his farm. Anotherproducer in Kihara (Central Province)used a home-prepared blend ofbotanicals and a detergent to managethe whitefly problem but apparentlywith little success. Chemical controlwas in many cases directed againstblight (i.e., using fungicides). The mostcommonly used insecticides for controlof the whitefly/TYLCV complex in thecountry were pyrethroids (lambda-cyhalothrin and cypermethrin) andorganophosphates (dimethoate anddiazinon). Other commonly usedinsecticides include alphacypermethrin (pyrethroid), amitraz(amidine), malathion, fenitrothion, andomethoate (organophosphates) andendosulfan (organochlorines).Acaricides such as dicofol, which has

138
Whiteflies and Whitefly-borne Viruses in the Tropics
little effect on insects (EXTOXNET,1998), were also commonly appliedagainst whiteflies. The use of fungicidessuch as metalaxyl, copper sulphate andMancozeb (ethylenebisdithiocarbamate), in an attempt tocombat TYLCV was very common.Almost half the producers (48%) madebetween one and four insecticideapplications per crop season to controlthe whitefly/TYLCV complex, while only8% applied nine to ten applications.Most producers in Nyanza and WesternProvinces did not apply insecticides orpractice any sort of control measureagainst the whitefly/TYLCV complex.
There was lack of knowledge onproper use of pesticides. Some tomatogrowers overused chemicals, whileothers did not spray until symptoms ofdamage were observed, and subsequentintensive chemical application failed tosave the crop. In many cases, producerscontinued spraying on a crop alreadyseverely affected by the virus, using ahigher dosage in the hope of curing theplants. Producers were ignorant aboutthe safety period between the lastspraying of the crop and date of sale orconsumption. Some producers said theydid not care about the pre-harvestinterval as long as the produce lookedgood for sale. Some of the localagrochemical suppliers bought largevolumes of pesticides and measured outsmall volumes according to the financialcapability of the buyers. The applicationof inappropriate pesticides and doserates was common and led to controlfailure. Chemicals without any labelswere usually bought from local dealers.
For the first time in Kenya,information was generated on whiteflyresistance to insecticides in selectedareas of the country. Assessment of theresistance of B. tabaci and T.vaporariorum to methomyl (acarbamate), cypermethrin andbifenthrin (pyrethroids) using both
glass-vial and leaf-dip bioassays wascarried out at Kibwezi, Kitui (EasternProvince), Nguruman (Rift ValleyProvince) and Mwea and Kihara (CentralProvince). Generally, T. vaporariorumwas found to be more resistant to thethree insecticides than was B. tabaci. Ofthe three insecticides used in the tests,cypermethrin was the most commonlyused in the survey farms and bifenthrinthe least. The resistance levels deducedfrom the analysis showed that both B.tabaci and T. vaporariorum were moreresistant to cypermethrin than to eitherbifenthrin or methomyl. The resultsobtained suggest that repeatedexposure of whiteflies to specificinsecticides could have selected themfor resistance to the chemicals.However, this was not a universalconclusion because there were somedeviations from the general trend. Forinstance, a 6.8-fold resistance tobifenthrin was detected at Kihara wherethe insecticide had reportedly neverbeen used. Whiteflies from Kajiado werethe most resistant to cypermethrin,giving a resistance factor value (RF) of87 in the leaf-dip method, while thosefrom Nairobi were the most resistant tomethomyl.
Strengthened ResearchCapacity
The present work has substantiallyimproved our understanding of thewhitefly problem in the major vegetableproduction areas of Kenya. Quantitativeinformation on whiteflies and naturalenemy species diversity and theoccurrence of the whitefly-transmitteddiseases, mainly TYLCV, as well as thesocio-economic understanding of theproducers’ perception and practicesrelating to management of the whitefly/TYLCV complex, has provided Kenyanresearchers with a solid background forsetting a research agenda for furtherwork, some of which may be carried out

139
Kenya
under Phase 2 of the TWF-IPM Project(see Introduction to this volume). Thefirst, diagnostic, phase of the projectalso contributed to capacity buildingthrough a planning workshop and amid-term meeting for all participatingscientists, training of one M.Sc. thesisstudent in assessment of insecticideresistance and training of one B.Sc.student in host plant preferencestudies.
Conclusions
The following areas in Kenya have beenidentified as hot spots for futureresearch on whiteflies and whitefly-transmitted viruses: Kibwezi, Kitui,Machakos/Athi River and Naivasha.The survey revealed that the prevalentwhitefly species in the country areB. tabaci, T. vaporariorum, B. afer,T. ricini, A. floccosus and S. phillyreae.The fact that 55% of the surveyedfarms had TYLCV symptoms indicatesa potential for whitefly/TYLCVproblems in Kenya. The majorconstraint in producers’ knowledge onthe whitefly/TYLCV complex was acomplete lack of understanding (99% offarmers) of the connection between thedisease and the vector.
To minimize the devastating effectsof whitefly-transmitted viruses intomato, sustainable vector- anddisease-management options need tobe developed, tested and deployed,including the use of resistant and/ortolerant cultivars, as well as culturaland biological control practices. Thedevelopment of such options needs tobe based on, and supported by,appropriate research. In addition, weneed to understand the source ofwhitefly infestations and the reasonsbehind the striking between-fieldvariability in whitefly population levelsthat is often observed among fields inclose proximity to each other. For
example, very heavy whitefly infestationwas observed in two adjacent farms atKihara but not in any of the othernearby farms. The contrast could havebeen due to the insects’ feedingpreference, because the two farms hadeggplant and squash (both preferredhosts) that were not on the other farmsbut this possibility would need to beinvestigated further. We need tounderstand why there is low or noincidence of TLCV in western Kenya,despite the prevalence of the vector.The potential of local isolates andcommercial formulations ofentomopathogenic fungi, as well asparasitoids and predators, for controlof whiteflies could be evaluated. Thereis also a need to look at other whitefly-transmitted viral diseases and at therole of weeds as reservoirs ofbegomoviruses.
There is a need for training ofentomologists and virologists in areasof detection, identification andcharacterization of whitefly-transmittedviruses, whiteflies and their naturalenemies, as well as training inepidemiological researchmethodologies. The continuedinvolvement of scientists from ICIPEand the Asian Vegetable Research andDevelopment Center (AVRDC), whohave provided technical support duringthe first phase of this project, would behelpful. Collaborative work involvingthe network of regional stations of theKenya Agricultural Research Institute(KARI) should be extended.
References
Bock, K. R. 1987. Some aspects of theepidemiology of African cassavamosaic virus in coastal districts ofKenya. In: Procs. InternationalSeminar on African cassava mosaicdisease and its control, 3-4 May1987, Yomoussokro, CI.p. 125-130.

140
Whiteflies and Whitefly-borne Viruses in the Tropics
EXTOXNET (Extension ToxicologyNetwork). 1998. Pesticideinformation profile, 3 Nov 1998.Available in: http://pmep.cce.cornell.edu/profiles/extoxnet/carbaryl-dicrotophos/dicofol-ext.html. (Accessed 2 May 2000)
GTZ (Deutsche Gesellschaft fürTechnische Zusammenarbeit).1995. Major pests of tomatoes ineastern and southern Africa:Compilation of past research workand identification of IPMopportunities. GTZ-Integrated PestManagement (IPM) HorticultureProject Report, Nairobi, KE. 32 p.
Löhr, B. 1997. The citrus wooly whitefly,a new pest in eastern and southernAfrica. Agrofor. Today Oct-Dec:21-22.
Mulandi, M. A. S. 1998. The status of thehorticultural industry, challengesand strategies, now and beyond theyear 2000. Horticultural CropsDevelopment Authority, Nairobi,KE. 34 p.
Seif A. A. 1982. Effect of the cassavamosaic virus on yield of cassava.Plant Dis. 66:661-662.
Varela, A. M.; Pekke, A. 1995.Proceedings of Tomato PlanningWorkshop for Eastern and SouthernAfrica Region, 16-20 October 1995,Harare, ZW. Deutsche Gesellschaftfür Technische Zusammenarbeit(GTZ)- Integrated Pest Management(IPM) Horticulture Project, Nairobi,KE. 32 p.

141
Tanzania
CHAPTER 2.4
TanzaniaIgnas Swai* and Simon Slumpa**
Introduction
Geographical context
The following regions were covered inthe surveys conducted during the1997-99 period: Arusha, Babati,Arumeru, Dodoma, Kongwa, Mpwapwa,Njombe, Iringa, Mwanga, Rombo,Moshi, Hai, Same, Mkuu, Mbeya,Vwawa, Tukuyu, Morogoro, Tanga,Lushoto and Zanzibar (Figure 1).Surveys followed the methodologyagreed among the project partners.
The surveyed areas can be groupedinto distinct eco-climatic zones(Table 1). The northern highlands coverthe Arusha and Kilimanjaro regionsand Usambara Highlands in the Tangaregion. The zone experiences two rainyseasons: short rains (vuli) fromNovember to December and long rains(masika) from March to May. The maindry season is from June to September.The coastal lowlands include suchareas as Muheza, Tanga, Ruvu, Dar-es-Salaam, Kibaha and Zanzibar Island.The rainfall pattern is the same as forthe northern highlands but thelowlands are hotter. The CentralPlateau includes Dodoma and western
* Horticultural Research and TrainingInstitute (HORTI-Tengeru), Arusha,Tanzania.
** Selian Agricultural Research Institute (SARI),Arusha, Tanzania.
Towns
Surveyedareas
Tanzania
Kilimanjaro
Tanga
Arusha
Dodoma
LakeVictoria
Moshi
Zanzibar
Iringsa
Njombe
Mbeya
Figure 1. Regions covered in the 1997-99survey, Tanzania.
parts of Morogoro regions, where therainfall pattern is variable andunpredictable. The Morogoro region istransitional and receives higher rainfallthan the Dodoma region. Dodomaexperiences only one rainy season,around November to January, andproduction of vegetables in this zone isseasonal. Irrigation is common in bothMorogoro and Dodoma.
The emergence of Bemisiatabaci as a pest and virusvector
The whitefly species Bemisia tabaci(Gennadius), the vector of Tomato leafcurl virus (ToLCV) and Tomato yellow
Kagera
Mwanza
Mara
Shinyanga
Kigoma
TaboraRukwa Singida
Pwani
Morogoro
RuvumaMtwara
Lindi

142
Whiteflies and Whitefly-borne Viruses in the Tropics
leaf curl virus (TYLCV), is ranked as themost important insect pest of tomato(Lycopersicon esculentum Mill.) inTanzania (Varela and Pekke, 1995). B.tabaci also transmits other viraldiseases in common bean (Phaseolusvulgaris [L.]), cowpea (Vigna unguiculata[L.] Walp.), sweetpotato (Ipomoeabatatas [L.] Lam.) and cassava (Manihotesculenta Crantz), while the woollywhitefly Aleurothrixus floccosus(Maskell) is widespread in citrus (Citrusspp. L.) growing areas. No informationwas available on the geographical rangeof whitefly species and their socio-economic significance in Tanzania priorto the country’s participation in theTropical Whitefly Integrated PestManagement (TWF-IPM) Project.
ToLCV was first reported inTanzania in 1990 (Czosnek et al., 1990).Additional survey studies conducted bythe Asian Vegetable Research andDevelopment Centre (AVRDC)-AfricaRegional Program in Tanzania from1994-97 showed that several tomatosamples with typical leaf curl symptomsdid not hybridize with the Egyptian orIsraeli ToLCV-DNA probes. Polymerasechain reaction (PCR) analysis performedwith the above tomato samplesindicated that in addition to TYLCV, avirus different from all previously
characterized tomato geminiviruses ofthe Old World occur in Tanzania andwas tentatively named Tomato leaf curlvirus-Tanzania (TLCV-Tan; Chiang et al.,1997), now ToLCV-Tz.
Increased BiologicalUnderstanding
Characterization ofbegomoviruses and whiteflybiotypes
During the 1997-99 period, 179 samplesof whitefly adults and nymphs werecollected from vegetable croppingsystems in Tanzania, of which 407nymph specimens were histologicallyprocessed and mounted on 213 slides.Identification of whiteflies conducted bythe International Center of InsectPhysiology and Ecology (ICIPE) revealedthat B. tabaci was the most commonwhitefly species, representing 78% of themounted specimens. B. afer (Priesnerand Hosney) represented 8% andTrialeurodes vaporariorum (Westwood),7%. Other whitefly species made up 7%of the specimens and includedTrialeurodes ricini (Misra), B. hirta Bink-Moenen, Orchamoplatus citri (Takahashi),Tetraleurodes andropogon (Dozier) andAleurothrixus floccosus (Maskell).
Table 1. Mean annual temperature, precipitation and altitude range of the surveyed regions inTanzania.
Region (Province)a Mean min.-max. Mean Altitudetemperature precipitation range
(°C) (mm) (m)
Arusha 11-21 913 485-1660
Dodoma 16-29 556 329-378
Iringa 14-25 633 396-561
Kilimanjaro (Same) 14-26 1130 244-1600
Kilimanjaro (Moshi) 16-26 549 120-850
Mbeya 10-27 1132 424-536
Morogoro 19-30 854 450-900
Tanga (Lushoto) 19-30 1006 366-549
Zanzibar 21-30 1436 20-60
a. Same is on the windward side and Moshi, the leeward side, of Mt. Kilimanjaro.

143
Tanzania
The reproductive host plants ofwhiteflies identified from the surveys inTanzania belong to the followingfamilies: Amaranthaceae, Asteraceae,Commelinaceae, Convolvulaceae,Cucurbitaceae, Euphorbiaceae,Lamiaceae, Leguminosae, Myrtaceae,Rutaceae, Solanaceae and Verbenaceae.Table 2 gives the scientific names of thehost plant species on which eachwhitefly species was found to bereproducing. Whiteflies were abundanton common bean but no whitefly-transmitted disease symptoms wereobserved.
ToLCV symptoms were observed inAchyranthes aspera L., Euphorbiaheterophylla L. and Nicandraphysalodes (L.) Gaertn. Nono-Womdimet al. (1996) identified TYLCV, thecausative virus in these hosts. Thesenon-cultivated host plants arewidespread in the country and mayserve as major reservoirs of TYLCV.
Disease incidence and symptomseverity
Areas identified as “hot spots” forToLCV incidence in Tanzania includeMorogoro, Dodoma, Kilimanjaro(especially Same) and Arusha (Figure 1).Fifteen percent of the surveyed farmshad 100% ToLCV incidence in theirtomato crop. It should be highlightedthat all surveyed farms in Morogoro(Dakawa, Kariakoo, Kipela, Kibundi,Kiruka and Bigwa) had 100% TLCincidence. Up to 100% of the crop alsowere reported to be affected inKilimanjaro (Marwa, Bangalala andSame) and Dodoma (Mbalala, Chikulaand Mpwapwa). Nine percent of thesurveyed farms had ToLCV incidencesranging from 70% to 99%. These werein Arusha (Baraa), Kilimanjaro(Mijongomeni, Kivulini and Mkolowoni)and Dodoma (Msolota). Ten percent ofthe surveyed farms had ToLCVincidences ranging from 50% to 69%,while 65% of the farms had incidences
ranging from 1%-19%. The lowestincidences of ToLCV symptoms,1%-2%, were observed in the southernpart of Tanzania (Iringa, Mbeya, Tukuyuand Njombe) and in Zanzibar. Therewere no distinct ToLCV-free zones foundwithin the surveyed areas of Tanzania.ToLCV incidence was highest during thesummer months (hot season), fromDecember to March. Yield loss due toToLCV depends on the susceptibility ofthe tomato cultivar, the crop stage atthe time of infection and environmentalconditions (Nono-Womdim et al., 1999).
Twenty-three tomato accessions,including progenies of crosses betweenLycopersicon chilense Dunal LA 1969and cultivated tomato, were field-screened for resistance to ToLCV byAVRDC (Nono-Womdin et al., 1999).Two commercial F1 hybrids, Fiona andTyking, had the highest level ofresistance. Plants of these varieties weresymptomless and they did not hybridizewith Israel TYLCV-DNA probe. Thevarieties PSR-403511 and PSR-407111were also resistant to ToLCV buthybridization tests showed that a fewsymptomless plants contained ToLCVDNA. The following progenies of crossesbetween L. chilense LA 1969 andL. esculentum had 30%-60% resistantplants: Chilytlc 94-1, 94-2, 94-4, 94-5,94-6 and MultichilTLC. Tests on twocommonly grown commercial tomatovarieties, Moneymaker and Roma,showed 100% susceptibility.
Natural enemy species
Samples of whitefly parasitoids andpredators were collected during thesurveys. Preliminary identification of thesampled parasitoids showed that mostwere Encarsia sophia (Girault andDodd), while Eretmocerus sp. wascollected on only two occasions. Anumber of predators, coccinellids andpredatory bugs were also collected in thevicinity of whitefly populations in thesurvey fields.

144
Whiteflies and Whitefly-borne Viruses in the Tropics
Table
2.
Rep
rodu
ctiv
e h
ost
pla
nts
of w
hit
efly
spec
ies
enco
un
tere
d d
uri
ng
surv
eys
in T
an
zan
ia.
Wh
itef
ly s
pec
ies
Cro
ps
Non
-cu
ltiv
ated
hos
ts
Fam
ily
Spec
ies
Fam
ily
Spec
ies
Bem
isia
ta
ba
ci (G
enn
adiu
s)C
onvo
lvu
lace
aeIp
omoe
a b
ata
tas
(L.) L
am.
Am
aran
thac
eae
Ach
yra
nth
es a
sper
a L
.E
uph
orbia
ceae
Ma
nih
ot e
scu
len
ta C
ran
tzA
ma
ran
thu
s sp
.Leg
um
inos
aeD
esm
odiu
m s
p.
Ast
erac
eae
Age
ratu
m c
onyzo
ides
L.
Vig
na
un
guic
ula
ta (L.) W
alp
.B
iden
s pilo
sa L
.Ph
ase
olu
s vu
lga
ris
L.
Ga
linso
ga p
arv
iflo
ra C
av.
Myr
tace
aePsi
diu
m g
ua
java
L.
Com
mel
inac
eae
Com
mel
ina
sp.
Sol
anac
eae
Lyco
per
sico
n e
scu
len
tum
Mill.
Eu
ph
orbia
ceae
Eu
ph
orbia
het
erop
hylla
L.
Sol
an
um
tu
ber
osu
m L
.Leg
um
inos
aeTep
hro
sia s
p.
Lam
iace
ae
Leo
not
is n
epet
ifol
ia (L.) R
. B
r.V
erben
acea
eLa
nta
na
ca
ma
ra L
.
Bem
isia
afe
r (P
ries
ner
an
d H
osn
ey)
Eu
ph
orbia
ceae
Ma
nih
ot e
scu
len
ta C
ran
tzLeg
um
inos
aePh
ase
olu
s vu
lga
ris
L.
Tri
ale
uro
des
va
por
ari
oru
m (W
estw
ood)
Leg
um
inos
aeD
esm
odiu
m s
p.
Eu
ph
orbia
ceae
Eu
ph
orbia
het
erop
hylla
L.
Vig
na
un
guic
ula
ta (L.) W
alp
Ph
ase
olu
s vu
lga
ris
L.
Tri
ale
uro
des
ric
ini (
Mis
ra)
Cu
curb
itac
eae
Cit
rullu
s la
na
tus
(Th
un
b.) M
atsu
m &
Nak
aiA
mar
anth
acea
eA
chyra
nth
es a
sper
a L
.E
uph
orbia
ceae
Ma
nih
ot e
scu
len
ta C
ran
tzA
ster
acea
eG
alin
soga
pa
rvif
lora
Cav.
Leg
um
inos
aeD
esm
odiu
m s
p.
Ph
ase
olu
s vu
lga
ris
L.
Bem
isia
hir
ta B
ink-M
oen
enE
uph
orbia
ceae
Eu
ph
orbia
het
erop
hylla
L
Orc
ha
mop
latu
s ci
tri (
Takah
ash
i)Pla
nt u
nid
enti
fied
Tet
rale
uro
des
an
dro
pog
on (D
ozie
r)Pla
nt u
nid
enti
fied
Ale
uro
thri
xus
floc
cosu
s (M
ask
ell)
Ru
tace
ae
Cit
rus
spp.

145
Tanzania
Geographical range of whiteflyand whitefly-transmitted virusinfestation
B. tabaci was the most commonwhitefly species throughout thecountry (Table 3). B. afer was commonin cassava in the areas of Iringa,Arusha, Mbeya and Kilimanjaro(Moshi). T. vaporariorum wasencountered in Arusha, Dodoma,Iringa, Kilimanjaro (Same), Mbeya andTanga (Lushoto) but not in Morogoro,Kilimanjaro (Moshi) or Zanzibar.T. ricini was common in Arusha,Iringa and Mbeya. A. floccosus was
collected only from citrus. The fewspecimens identified as B. hirta requireadditional confirmation.
Up to 100% ToLCV incidence wasfound at altitudes ranging from 260 to1060 m, while ToLCV incidence of upto 70% was encountered as high as1460 m altitude (Table 4). B. tabaciwas the prevalent whitefly species upto an altitude of 1260 m, while thetwo Trialeurodes species mentionedabove were prevalent at higheraltitudes (Table 4).
Table 3. Species composition (%) of whitefly samples collected in regions of Tanzania.
Provincea Species compositionb
B.t. B.a. T.v. T.r. B.h. O.c. T.a. A.f.
Arusha 48 18 8 14 2 0 0 10
Dodoma 90 2.5 2.5 5 0 0 0 0
Iringa 37.5 25 25 12.5 0 0 0 0
Kilimanjaro (Same) 96 2 2 0 0 0 0 0
Kilimanjaro (Moshi) 89 11 0 0 0 0 0 0
Mbeya 40 17 33 10 0 0 0 0
Morogoro 99 0 0 0 0 1 0 0
Tanga (Lushoto) 91 2 5 2 0 0 0 0
Zanzibar 83 0 0 0 0 0 17 0
a. Same is on the windward side and Moshi, the leeward side, of Mt. Kilimanjaro.b. B.t., Bemisia tabaci; B.a., B. afer; T.v., Trialeurodes vaporariorum; T.r., T. ricini; B.h., B. hirta;
O.c., Orchamoplatus citri; T.a., Tetraleurodes andropogon; A.f., Aleurothrixus floccosus.
Table 4. Maximum incidence (% of sampled plants showing disease symptoms) of tomato leaf curl(TLC) encountered and species composition (%) of whitefly samples collected at differentaltitudes in Tanzania.
Altitude (m) Maximum Whitefly species compositiona
TLCincidence B.t. T.v. T.r. B.a. A.f. B.h. T.a. O.c.
0-60 4 83 0 0 0 0 0 17 0
60-260 100 91 9 0 0 0 0 0 0
260-460 100 88 3 4 5 0 0 0 0
460-660 100 66 18 7 9 0 0 0 0
660-860 100 84 0 0 14 0 2 0 0
860-1060 100 44 0 0 0 56 0 0 0
1060-1260 50 75 13 12 0 0 0 0 0
1260-1460 70 0 0 100 0 0 0 0 0
1460-1660 15 14 43 0 43 0 0 0 0
a. B.t., Bemisia tabaci; T.v., Trialeurodes vaporariorum; T.r., T. ricini; B.a., B. afer; A.f., Aleurothrixusfloccosus; B.h., B. hirta; T.a., Tetraleurodes andropogon; O.c., Orchamoplatus citri.

146
Whiteflies and Whitefly-borne Viruses in the Tropics
Increased Socio-economicUnderstanding
Farmers’ assessment ofwhitefly-related problems
Tomato is one of the most importantvegetables cultivated in Tanzania. Most(79%) of the tomato producersinterviewed used their own land forproduction, while 17% used rentedland. A vast majority of the producers(92%) were men. Almost all theproducers (96%) regarded tomato astheir most profitable vegetable cropsand 62% of producers have beeninvolved in vegetable farming for morethan 5 years. Although tomatoproduction has increased in the last5 years to meet increased demand forprocessing and fresh markets, theaverage yields remain low at about10-14 tons per hectare.
The main tomato varietiescultivated in Tanzania are Marglobe,Moneymaker and Roma VF, all ofwhich are highly susceptible to severalviral diseases. Most producers (60%)obtain planting material from the localmarket, 36% use their own seeds and4% get planting material from othertomato producers. Other commonlygrown vegetable crops include cowpea,pea (Pisum sativum L.), common bean,cabbage (Brassica oleracea L. var.capitata L.), onion (Allium cepa L.), Irishpotato (Solanum tuberosum L.),eggplant (Solanum melongena L.), sweetpepper (Capsicum annuum L.), okra(Abelmoschus esculentus [L.] Moench),sweetpotato, Amaranthus spp.,watermelon (Citrullus lanatus [Thunb.]Matsum. & Nakai), melon (Cucumismelo L.), cucumber (Cucumis sativus L.var. sativus) and other cucurbits.Whiteflies also attack most of thesecrops. The great majority (94%) of theproducers interviewed practice croprotation.
Producers ranked their pest anddisease problems in tomato andassociated vegetable crops in Tanzania(in order of importance) as follows: lateblight (Phytophthora infestans [Mont.]de Bary), TLC, fruit borers (Helicoverpaarmigera [Hübner]), whiteflies(Homoptera: Aleyrodidae), aphids(Aphididae), Fusarium wilt, early blight(Alternaria solani [Ell. and Mart.] Jonesand Grout), red spider mite(Tetranychus urticae Koch.), diamondback moth (Plutella xylostella [L.]),angular leaf spot (Pseudomonassyringae pv. Lachrymans), bruchids(Coleoptera: Bruchidae), bacterial leafspot (Xanthomonas campestris pv.visicatoria), leaf miner (Lyriomyza spp.),beanfly (Ophiomyia spp.), cutworm(Spodoptera litura [Fabricius]), powderymildew (Leveillula taurica), black rot(X. campestris pv. campestris), fruit fly(Drosophila melanogaster), bacterialcanker (Clavibacter michiganensis),bean rust (Uromyces phaseoli), Tomatomosaic virus, Septoria leaf spot (Septorialycopersici), nematodes (Meloidogynespp.), Cucumber mosaic virus, thrips(Thysanoptera: Thripidae), cabbagesawfly (Nematus spp.) and soft rot(Erwinia carotovora).
Most producers (90%) were able torecognize the whitefly and 84% alsorecognized ToLCV. Only 11% of theinterviewed producers knew thatwhiteflies and ToLCV were interrelated.None of the interviewed producers fromMorogoro (where the disease incidencewas 100% on all surveyed farms) knewthe cause of ToLCV; they did not havenames for the whiteflies other than“insects”, nor did they have precisenames for ToLCV. Most producers(75%) believed that the whitefliescaused problems on their farms.Almost half of them (43%) believed thatboth whiteflies and ToLCV causedproblems, whereas 30% attributed theproblems to ToLCV only and 27% towhiteflies only.

147
Tanzania
Local names given to whitefliesincluded chawa (lice), inzi weupe (whiteflies), kibanda, kifizi, kipe weupe,kipepeo (small butterflies), kurukury,mbuu, msubi weupe, mvumuu, ndaka,sughru (small flying insects), sunhuu,suru, tukorokotwa, wadudu (insects)and wadudu weupe (white insects). Thelocal names given to ToLC includedrasta (dreadlocks), ugonjwa wa kukunja(curling disease), ukoma (leprosy),dume (sterile), ghojo, kibangi, kudulala,kutu (rust), bondia, majani, masai,mdamango, mwanga bondia, ngofu,ngumi (boxer), and kibangi and kobe(tortoise).
Estimation of diseaseincidence and yield losses
Among the surveyed farms, 99% hadTLC symptoms in their tomato cropand 34% had ToLCV incidences above25%. Common symptoms include leafcurling, yellowing, chlorosis of leafmargins, leaf distortion, reduction inleaf size, shortening of the internodes,stunting, excessive branching andflower abscission (Nono-Womdim et al.,1999). The average perceived yield lossin tomatoes due to the whitefly/ToLCVcomplex was 45%. Some producers(9%) reported a total yield loss, 13%reported three-quarters yield loss, 32%reported half yield loss, 42% reportedone-quarter yield loss and only 5% ofthe producers reported no yield loss.
Most of the producers interviewed(74%) believed that they had whiteflyand/or virus problems every year and26% reported that the most severeproblems occurred in 1997. Mostproducers (94%) also believed thatthere is a relationship between climateand whitefly/ToLCV occurrence. Themajority (90%) believe that the problemis worst during the hot season,between December and March, whilethe problem is less pronouncedbetween August and December. The
producers mostly attributed theincrease in severity to weatherchanges, particularly long dry spells.Other factors mentioned by someproducers were ineffective insecticides,resistance to insecticides and old cropremains (acting as a source ofinfestation and virus reservoir).
Producers gave estimation of thecosts involved in crop protectionmeasures (see below). The costs weremainly incurred in the purchasing ofchemical insecticides and labor fortheir application.
Costs estimated by producersin control of whitefly/TYLCV
complex per hectare of tomato
US$ % producers0-49 4050-99 18
100-199 26200-299 9300-400 7
Pesticide use
One-third of producers (35%) receivedrecommendations on managementpractices from technical advisors, 35%from other tomato producers, 3% fromsales agents, 10% from others and17% relied on their own judgement.Among those producers who appliedinsecticides, the decision on whatinsecticides to apply and when wasmade by the producer in 93% of thecases, while 6% relied on technicaladvice.
Use of pesticides was the mostcommon option for managing thewhitefly/ToLCV problem and waspracticed by 76% of those interviewed.Cultural control was practiced by 11%of the producers, while 13% did notpractice any control. The pesticidesmost commonly applied by vegetableproducers in Tanzania for control ofwhitefly/disease problems on their

148
Whiteflies and Whitefly-borne Viruses in the Tropics
farms are profenofos, chlorpyrifos,fenitrothion, dimethoate, pirimiphos-methyl, chlorpyrifos and diazinon(organophosphates); deltamethrin,lambda-cyhalothrin and fenvalerate(pyrethoids); thiodan and endosulfan(organochlorines); and methomyl(carbamate).
Producers who made 10 insecticideapplications per season to manage thewhitefly/ToLCV complex accounted for21% of those interviewed. Those whomade from five to eight applicationscomprised 31% of interviewees and asimilar percentage made from one tofour applications. However, 18%reported that they did not applyinsecticides. Most producers (62%)applied insecticides as a preventivemeasure, 26% made the applicationwhen they observed damage and 6%made the application according to thecalendar.
Strengthened ResearchCapacity
This survey has substantially improvedour understanding of the magnitude ofthe whitefly-associated problems in themajor vegetable production areas ofTanzania. The work conducted underthis first, diagnostic, phase of the TWF-IPM Project also has strengthenedcollaboration between the nationalagricultural research organization andinternational research institutionssuch as ICIPE and AVRDC.
Conclusions
TYLCV is the most important whitefly-transmitted viral disease in tomatocropping systems in Tanzania. Thedisease causes significant yield loss intomato production in various parts ofthe country. The lowlands of Morogoro
and Dodoma were identified as theprincipal “hot spots” for ToLCV, whereincidences of symptoms of 100% werefrequently encountered. Moderatelyhigh incidences were also observed inthe Arumeru Highlands near Arushaand Kilimanjaro.
Commercial cultivars of tomatogrown in Tanzania are susceptible toTYLCV. Some breeding lines from theAfrican Regional Program (ARP) ofAVRDC at Arusha have shownsignificant tolerance to TYLCV. AVRDC-ARP in collaboration with theHorticultural Research and TrainingInstitute (HORTI-Tengeru) have begunefforts to screen a broad array oftomato germplasm and incorporate theresistance genes into cultivated tomatovarieties.
Further work in Tanzania, underthe second phase of the TWF-IPMProject, should focus on evaluation ofgermplasm for use in breeding forresistance to TYLCV and ToLCV, anddevelopment of IPM options for themanagement of the whitefly vector.
References
Chiang, B. T.; Nakhla, M. K.; Maxwell, P.;Schoenfelder, M.; Green, S. K.1997. A new geminivirus associatedwith a leaf curl disease of tomato inTanzania. Plant Dis. 81:111.
Czosnek, H.; Navot, N.; Laterrot, H. 1990.Geographical distribution of Tomatoyellow leaf curl virus. A first surveyusing a specific DNA probe.Phytopathol. Mediterr. 29:1-6.
Nono-Womdim, R.; Swai, I. S.; Green, S.K.; Gebre-Selassie, K.; Laterrot, H.;Marchoux, G.; Opena, R.T. 1996.Tomato viruses in Tanzania:Identification, distribution anddisease incidence. J. S. Afr. Hort.Sci. 6:41-44.

149
Tanzania
Nono-Womdim, R.; Swai, I. S.; Green,S. K.; Chadha, M. L. 1999.Tomato yellow leaf curl virus andTomato leaf curl-like virus inEastern and Southern Africa.Paper presented at final IPMworkshop held at theInternational Center of InsectPhysiology and Ecology (ICIPE),Nairobi, KE. 2 p.
Varela, A. M.; Pekke, A. 1995.Proceedings of Tomato PlanningWorkshop for Eastern and SouthernAfrica Region, 16-20 October 1995,Harare, ZW. Deutsche Gesellschaftfür Technische Zusammenarbeit(GTZ)-Integrated Pest Management(IPM) Horticulture Project, Nairobi,KE. 32 p.

150
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 2.5
MalawiHarriet Thindwa and Patric Khonje*
Introduction
Geographical context
Malawi lies between latitude 9° 22' and17° 7' South, and between longitudes32° 40' and 35° 55' East. The climate ischaracterized by three main seasons:cool and dry from May to August, warmand dry from September to November,and warm and wet from December toApril. The 5-month rainy season(November to March) begins earlier insouthern and central regions of thecountry. Annual rainfall ranges from600 mm in the lower Shire Valley andthe Karonga Lake Shore Plains to over3000 mm in high-elevation areas.Temperatures range between 20 °C and35 °C but may approach and surpass40 °C in the Rift Valley areas duringOctober and November (Mkanda et al.,1995).
Surveys were carried out in themain areas of the country growingtomato (Lycopersicon esculentum Mill.)during the period 1997 to 1999. Areassurveyed were Mzuzu, Mzimba, NkhataBay, Salima, Lilongwe, Dedza, Ntcheuand Thyolo (Figure 1).
The emergence of Bemisia tabacias a pest and virus vector
In Malawi, the whitefly species Bemisiatabaci (Gennadius) has been reported asthe vector of cassava mosaic disease(Swanson and Harrison, 1994). Varying
* Bvumbwe Agricultural Research Station,Limbe, Malawi.
Figure 1. Tomato-growing areas surveyed forwhitefly incidence in Malawi.
Towns
Surveyedareas
Lake
Malawi
Kusungu
Lilongwe
Kumuz Intl
Karonga
Chileka Intl
Mzuzu
Dlantyre
Zomba

151
Malawi
levels of infestation by the citrus woollywhitefly Aleurothrixus floccosus(Maskell) also have been recorded inregions of the country growing citrus(Citrus spp. L.) (Löhr, 1996).
Although whiteflies were suspectedof transmitting viral diseases invegetable crops, especially tomato, noreliable information on this subject wasavailable from Malawi prior to thesurvey conducted under the TropicalWhitefly Integrated Pest Management(TWF-IPM) Project. The occurrence ofwhitefly species, their geographicaldistribution and their natural enemiesin the vegetable cropping system alsowere unknown.
Increased BiologicalUnderstanding
Characterization ofbegomoviruses and whiteflybiotypes
Selection of survey sites and othermethodologies followed the standardsagreed with other project partners.Questionnaires were used to gatherinformation from vegetable producers.The incidence of plants affected bytomato yellow leaf curl was estimatedon their farms, whiteflies were sampledfor species identification and parasitoidsand suspected natural enemies werecollected.
During the surveys, 130 collectionsof adults and nymphs of whiteflies weremade. Of these, 159 specimens wereprocessed and mounted on 67 slides.Preliminary identification carried out atthe International Center of InsectPhysiology and Ecology (ICIPE) revealedthat Bemisia afer (Priesner and Hosney)is the most prevalent species in thecountry, accounting for 67% of samplesidentified. Of the remaining specimens,B. tabaci comprised 30%, and
Trialeurodes vaporariorum (Westwood),T. ricini (Misra) and Aleyrodes proletella(Linnaeus) 1% each. The speciescomposition of samples from thevarious areas surveyed is reported inTable 1.
Table 1. Species composition (%) of whiteflysamples collected in tomato-producing regions of Malawi.
Province Species compositiona
B.a. B.t. T.v. T.r. A.p.
Dedza 56 34 5 0 5
Lilongwe 86 14 0 0 0
Mzimba 86 14 0 0 0
Mzuzu 100 0 0 0 0
Nkhata Bay 53 47 0 0 0
Ntcheu 55 15 10 20 0
Salima 33 67 0 0 0
Thyolo 54 42 0 0 4
a. B.a., Bemisia afer; B.t., B. tabaci; T.v.,Trialeurodes vaporariorum; T.r., T. ricini;A.p., Aleyrodes proletella.
The reproductive host plants fromwhich whitefly nymphs were collectedbelong to the families Solanaceae,Euphorbiaceae, Leguminosae andVerbenaceae (Table 2). B. tabaci wasfound breeding on tomato in only a fewareas.
Symptoms of tomato yellow leafcurl were observed in most of thesurveyed sites. Samples of about 10plants per field were collected from allthe affected farms, squashed on nylonmembranes and sent to the John InnesCentre (JIC) for hybridization. Theresults confirmed the presence ofTomato yellow leaf curl virus (TYLCV).
Disease incidence andsymptom severity
Areas identified as “hot spots” (highincidence of TYLCV) include Dedza,Mzimba, Thyolo and Ntcheu (Figure 1).The highest incidences of TYLCV wereobserved at altitudes ranging from1085 to 1485 m (Table 3).

152
Whiteflies and Whitefly-borne Viruses in the Tropics
Table
2.
Rep
rodu
ctiv
e h
ost
pla
nts
of w
hit
efly
spec
ies
reco
rded
in
Mala
wi.
Wh
itef
ly s
pec
ies
Cro
ps
Non
-cu
ltiv
ated
hos
ts
Fam
ily
Spec
ies
Fam
ily
Spec
ies
Bem
isia
afe
r (P
ries
ner
an
d H
osn
ey)
Eu
ph
orbia
ceae
Ma
nih
ot e
scu
len
ta C
ran
tzLeg
um
inos
aePh
ase
olu
s vu
lga
ris
L.
Bem
isia
ta
ba
ci (G
enn
adiu
s)E
uph
orbia
ceae
Ma
nih
ot e
scu
len
ta C
ran
tzE
uph
orbia
ceae
Eu
ph
orbia
het
erop
hylla
L.
Leg
um
inos
aeM
ucu
na
dee
rin
gia
na (B
ort.
) M
err.
Ver
ben
acea
eLa
nta
na
ca
ma
ra L
.Ph
ase
olu
s vu
lga
ris
L.
Sol
anac
eae
Lyco
per
sico
n e
scu
len
tum
Mill.
Tri
ale
uro
des
va
por
ari
oru
m (W
estw
ood)
Leg
um
inos
aePh
ase
olu
s vu
lga
ris
L.
Tri
ale
uro
des
ric
ini (
Mis
ra)
Leg
um
inos
aePh
ase
olu
s vu
lga
ris
L.
Ale
yro
des
pro
lete
lla (L
inn
aeu
s)Leg
um
inos
aeM
ucu
na
dee
rin
gia
na (B
ort.
) M
err.

153
Malawi
The survey revealed that B. afer isthe predominant whitefly species inmost parts of Malawi (Table 1). Thisspecies was found on cassava (Manihotesculenta Crantz), common bean(Phaseolus vulgaris L.) and unidentifiedweeds but not on other vegetable crops.B. tabaci was also common in mostparts of the country (Table 1).pT. vaporariorum was collected fromDedza and Ntcheu, while T. ricini wascollected from Ntcheu only andA. proletella from Dedza and Thyolo.High numbers of nymphs werecollected from Nkhata Bay, Dedza andNtcheu, the latter two being the onlyregions from which more than threewhitefly species were identified. Onfarms in Salima, very few whiteflynymphs could be found foridentification and the region hadrelatively low TYLCV incidences(0%-11%). B. tabaci was thepredominant whitefly species up to analtitude of about 900 m, while B. aferbecame more abundant at higheraltitudes (Table 3).
Natural enemy species
Nine collections of whitefly parasitoidsand 10 of predators were made duringthe survey. Among these were Encarsiasophia (Girault and Dodd), coccinellids(Coleoptera: Coccinellidae) andpredatory bugs (Heteroptera:Anthocoridae).
Increased Socio-economicUnderstanding
Farmers’ assessment ofwhitefly-related problems
Most of the producers interviewed(86%) grow tomato on their own land,while 12% rent the land. The majorityof producers (88%) were men. Tomatowas regarded as the most profitablevegetable crop by 98% of theproducers. Only 27% of them havebeen growing tomato for more than5 years. Tomato varieties commonlygrown in Malawi include Moneymaker,Red Khakhi, Vercles and Heinz. Thetomato seed was bought from the localmarket by 35% of the producers, 33%used their own seed (from the previousseason), 21% obtained seed from theAgriculture Trading Company (ATC)and 11% used imported seeds. Tomatois grown both under rain-fed andirrigated conditions.
Other common vegetable crops inthe country include common bean,cabbage (Brassica oleracea var. capitataL.), Irish potato (Solanum tuberosumL.), mustard (Sinapis alba L. subsp.alba), onion (Allium cepa L.), pumpkin(Cucurbita spp.), rape (Brassica napusL. var. napus), sweetpotato (Ipomoeabatatas [L.] Lam.), turnip (Brassicarapa L. subsp. rapa), okra
Table 3. Maximum incidence (%) of Tomato yellow leaf curl virus (TYLCV) and species composition (%)of whitefly samples encountered at increasing altitudes in Malawi.
Altitude TYLCV Whitefly species compositionb
(m) incidenceaB.a. B.t. T.v. T.r. A.p.
485-685 14 31 56 0 0 13
685-885 14 38 63 0 0 0
885-1085 7 64 27 0 0 9
1085-1285 52 79 21 0 0 0
1285-1485 49 59 24 5 12 0
1485-1685 5 66 23 6 0 5
a. A TYLCV incidence of 94% was observed in one farm in the highest altitude range (1485-1684 m),while other farms in this altitude range had incidences of 0%-5%.
b. B.a., Bemisia afer; B.t., B. tabaci; T.v., Trialeurodes vaporariorum; T.r., T. ricini; A.p., Aleyrodesproletella.

154
Whiteflies and Whitefly-borne Viruses in the Tropics
(Abelmoschus esculentus [L.] Moench),pepper (Capsicum annuum L.), eggplant(Solanum melongena L.), and carrot(Daucus carota L. subsp. sativus[Hoffm.] Arcang. var. sativus Hoffm.).Cassava is also a major staple crop.Most producers (94%) practice croprotation.
The main pests and diseasesrecorded on tomato, ranked accordingto importance, were late blight(Phytophthora infestans [Mont.] deBary), red spider mites (Tetranychusspp.) TYLCV, whiteflies, bacterial wilt(Ralstonia solanacearum) and cutworms(Spodoptera litura [Fabricius]).
Eighty-three percent of producerscould recognize whiteflies and 82%could recognize TYLCV. None of theproducers seemed to know theinterrelationship between TYLCV andwhiteflies and confused TYLCVsymptoms with red spider mitedamage, aphid damage, heavy raindamage, fungal diseases, nitrogendeficiency and soil-borne diseases.Most of those interviewed (83%)thought that whiteflies and TYLCVcaused problems on their farms.Almost half of the producers (46%)complained about the whitefly only,17% of the disease only and 37%complained about both the disease andthe whitefly.
Local names given to whitefliesinclude msambe, aphids, flies, pests,vegetable lice, white aphids, cobweb,chinoni, white lice, white ants, mites,small flies, lice and white butterflies.Local names given to TYLCV includedblight, leprosy, chisaka (bushy top),kaligwiti (bushy top), crinkled leaves,katungana, ciguduli, bunchy top,cicsaka, kafumbata and malformation.
Estimation of diseaseincidence and yield losses
Among the surveyed farms, 75% hadTYLCV symptoms in their tomato crop.However, only 6% of the farms hadincidences of TYLCV symptoms above25%, while 16% of the farms hadincidences ranging from 10% to 25%,and 53% of the producers hadincidences ranging from 0.5% to 9%.The highest incidence (94%) wasreported on a farm in Dedza.
On average, the producers’estimate of yield loss in tomato due tothe whitefly/TYLCV complex was 49%.According to the survey, 14% reportedtotal yield loss, 14% reported three-quarter yield loss, 35% reported halfyield loss, 29% reported one-quarteryield loss and 9% did not report anylosses due to the whitefly/TYLCVcomplex. Those who reported incurringtotal loss were from the Dedza andNtcheu Provinces, and a singleproducer from Mzimba. Producers whoreported abandoning tomato growing,at least once, due to the problem madeup 21% of those interviewed; 38% ofthem abandoned their farms in 1997.
Most producers (68%) believed thatthey had whitefly/TYLCV problemsevery year. The most severe attack wasin 1997 according to 43% of therespondents. These producers werefrom Ntcheu (Kasamba), Dedza(Kalilombe, Chimlambe, Jere), NkhataBay (Kashonga, Chipayika), Salima(Matumba), Mzimba (Chizuminja) andMzuzu (Kaboko). Nearly all producers(96%) believe there is a directrelationship between the climate andwhitefly/disease incidence. There wasno obvious consensus among theproducers as to what weatherconditions trigger whitefly and TYLCVoutbreaks. The whitefly/TYLCVproblems were reported to be worstfrom January to June and again fromSeptember to October. Some producers

155
Malawi
(40%) believe that the whitefly/TYLCVincidence is higher when it is dry andhot, while others (30%) said theyexperience most problems when it iswet and cold. The opinions of theremaining producers were scatteredbetween the remaining combinations ofdry, moderate, wet and hot, moderate,and cold weather.
Producers gave estimation of thecosts involved in crop protectionmeasures (see below).
Costs estimated by producers incontrol of whitefly/TYLCV complex
per season of tomato production
US$ % producers0-49 2650-99 16
100-199 19200-299 14300-400 9
>400 16
Many producers (42%) had notreceived recommendations onmanagement practices for whiteflies.Those who had received adviceobtained it from various sources: 34%received recommendations fromtechnical advisors, 15% from othervegetable producers, 6% from familymembers or neighbours and 3% fromsales agents. Most producers (90%)managed the whitefly/virus problem bymeans of synthetic pesticides, 4% bycultural means and 6% practiced nocontrol at all. Other control methodssuch as legislative measures, naturalinsecticides (e.g., botanicals and bio-pesticides), biological control and theuse of resistant varieties, were notreported.
Pesticide use
The decision on what insecticide toapply and when was made by theproducer himself in 58% of the cases,by technical advisors (29%), sales
people (3%) or a family member (2%),while 7% did not use insecticides ontheir farms. Half the producers (52%)applied insecticides as a preventivemeasure, while 33% applied themwhen damage was observed and 5%applied insecticides according to thecalendar. Few producers (5%) reportedmaking more than 10 insecticideapplications per season, while 4%made 9-10 applications, 37% of theproducers made 5-8 applications, 40%made 1-4, and 14% applied noinsecticides to manage the whitefly/disease problem.
The insecticides most widely usedin the management of the whitefly/TYLCV problem in Malawi include:fenitrothion, malathion and dimethoate(organophosphates); cypermethrin andlambda-cyhalothrin (pyrethroids);carbaryl and sevin (carbamates).Farmers were ignorant of the fact thatchemicals with the same activeingredient often are sold under varioustrade names and they therefore riskusing the same active ingredientthroughout, rather than alternatingwith others. The use of fungicides wascommon because producers confusedTYLCV symptoms with blight. Miticidessuch as dicofol were used also in anattempt to control whiteflies butEXTOXNET (1998) reports that thischemical has little effect on insects.
Strengthened ResearchCapacity
Scientists from ICIPE trained andaccompanied the national scientistsduring the beginning of the survey. Theproject has equipped nationalscientists with a better understandingof the severity of whitefly-relatedproblems in the country’s vegetableproduction. New information on thestatus of whiteflies and the whitefly-transmitted viruses in tomato now is

156
Whiteflies and Whitefly-borne Viruses in the Tropics
available. Understanding ourproducers’ perceptions and practices inrelation to whitefly problems has beena significant achievement of thisproject.
Conclusions
Whitefly/TYLCV “hot spots” in Malawiinclude Dedza, Mzimba, Thyolo andNtcheu. In both Dedza and Ntcheu,several farmers reported havingabandoned their tomato crops at leastonce due to whiteflies/TYLCV. For thisreason, these two areas should betargeted for further research in thesecond phase of the project. B. afer,followed by B. tabaci, is the prevalentwhitefly species in most parts of thecountry and the most widespreadparasitoid was E. sophia.
Although many producersrecognized whiteflies and TYLCV, theyoften confused TYLCV symptoms withthose of other pest and disease ornutrient deficiencies. Producers alsowere confused about the climaticconditions that trigger outbreaks ofwhiteflies and TYLCV.
Incidences of TYLCV in Malawiwere not as high as in neighbouringTanzania. Only 6% of the surveyedfarms had TYLCV incidence above25%. Although TYLCV incidences weregenerally low, the disease was
encountered in all surveyed areas,indicating that the whitefly/TYLCVcomplex is a potential threat toMalawi’s vegetable production.
References
EXTOXNET (Extension ToxicologyNetwork). 1998. Pesticideinformation profile, 3 Nov 1998.Available in: http://pmep.cce.cornell.edu/profiles/extoxnet/carbaryl-dicrotophos/dicofol-ext.html (Accessed 2 May2000)
Löhr, B. 1996. The citrus woolly whiteflyin eastern and southern Africa:Distribution, indigenous naturalenemies and control strategies. In:Procs. International Society ofCitriculture. Vol. 1, p. 619-624.
Mkanda, F. X.; Kainja, S.; Kalindekafe,M.; Chavula, G. M. 1995. Malawi:Greenhouse gas inventory andassessment of climate changevulnerability. Interim report onclimate change country studies,March 1995. US Global ChangeResearch Information Office(GCRIO). Available in: http://www.gcrio.org/CSP/IR/IRmalawi.html (Accessed 11 May2000)
Swanson, M. M.; Harrison, B. D. 1994.Properties, relationships anddistribution of cassava mosaicgeminiviruses. Trop. Sci. 34:15-25.

157
Tomato Yellow Leaf Curl Virus in E. and S. Africa
CHAPTER 2.6
Tomato Yellow Leaf Curl Virus andTomato Leaf Curl-like Viruses inEastern and Southern Africa
Remi Nono-Womdim*, Ignas Swai*,Madan Mohan Lal Chadha** and Sylvia Green*
Introduction
Tomato (Lycopersicon esculentum Mill.)is one of the most popular vegetables ineastern and southern Africa. It is ahigh-value crop, providing a goodsource of income to small-scalefarmers. Tomato is consumed both infresh and processed forms. Theprocessing industry often provides goodrural employment opportunities andproducts for export.
The productivity of tomato inAfrica, particularly in eastern andsouthern Africa, is unfortunatelyamong the world’s lowest. For example,the mean yield per hectare of tomato insouthern African countries ranges from1.5 to 14 tons per hectare as comparedto the world average of 25 tons perhectare based on the 1989 Food andAgriculture Organization (FAO)production estimates. Several fungal,bacterial and viral diseases havedrastically hampered the cultivation oftomato in eastern and southern Africa(AVRDC 1996; 1997; 1998). Amongviral pathogens, Tomato yellow leaf curlvirus (TYLCV) and tomato leaf curl-like
viruses account for most losses infarmers’ fields (unpublished results).However, adequate information on theepidemiology of TYLCV and associatedviruses in eastern and southern Africaremain scanty.
This chapter refers to theemergence of TYLCV and associatedviruses in tomato in eastern andsouthern Africa and describes thedevelopment of our understanding oftheir identification and management.
Impact and AssessmentLosses
Over the past decade, TYLCV and leafcurl-like viruses have causedsignificant losses for tomato farmers ineastern and southern Africa. Yieldlosses depend on the growing season,tomato cultivars and growth stage atwhich plants become infected. Recenton-farm surveys indicated that theaverage perceived yield losses in tomatowere up to 40% in Malawi, 50% inKenya, 75% in Tanzania and 100% inSudan (CIAT, 1998). In Sudan, yieldlosses due to TYLCV have beenreported to vary in different regions(Yassin and Nour, 1965; Makkouk etal., 1979). Moreover, Ioannau andIordanou (1985) indicated that lossesdue to TYLCV vary between 50% and82% depending on the time ofinfection.
* Asian Vegetable Research and DevelopmentCenter (AVRDC), Africa Regional Program.
** Southern Africa Development Community(SADC)-AVRDC-Collaborative Network forVegetable Research and Development inSouthern Africa (CONVERDS), Arusha,Tanzania.

158
Whiteflies and Whitefly-borne Viruses in the Tropics
In East Africa, TYLCV was firstreported in the Sudan (Yassin andNour, 1965). The disease has sincespread to Tanzania (Czosnek et al.,1990; Nono-Womdim et al., 1996),Malawi and Zambia (AVRDC, 1996).Recent survey studies have indicatedthat this virus occurs in Namibia andSwaziland (AVRDC, 1998), and inKenya and Uganda (Table 1). A similardisease caused by a distinct virus,tentatively named “Tomato leaf curlvirus-Tanzania” (TLCV-Tan; nowaccepted as ToLCV-Tz) (Chiang et al.,1997) has been reported in the centralregion of Tanzania (Nono-Womdim etal., 1996).
Diagnosis
In east and southern African countries,TYLCV is primarily identified by thepresence of symptoms that consist ofyellowing, leaf curling and stunting.However, visual diagnosis is not usefulfor the implementation of effectivecontrol strategies since the symptomsare very clear on old plants that arealready fruiting.
Selected monoclonal antibodiesdeveloped against African cassavamosaic virus (ACMV) or Indian cassavamosaic virus (ICMV) have been used todetect different isolates of TYLCV(Macintosh et al., 1992; Givord et al.,1994). These antibodies also were usedto identify TYLCV isolates from Tanzania(unpublished results). It has been shownthat many whitefly-transmittedbegomoviruses are serologically related(Roberts et al., 1984). Therefore,serological tests for identification ofTYLCV are unspecific and do notdifferentiate strains or different leaf curlviruses.
Nucleic acid hybridization assay iscommonly used for the detection ofTYLCV. Many cDNA probes preparedagainst different strains of TYLCV arenow available (Czosnek et al., 1990;Kheyr-Pour et al., 1991). Some of theseprobes currently are being used toidentify leaf curl isolates from easternand southern Africa.
Etiology
TYLCV belongs to the Geminiviridaefamily and has small geminate particlesmeasuring about 20 × 30 nm (Harrisonet al., 1977; Czosnek et al., 1988). Thegenome of TYLCV consists of a circularsingle-stranded DNA encapsulated by asingle capsid protein (Navot et al., 1991).Many isolates from eastern andsouthern Africa are closely related toTYLCV from Israel (TYLCV-Is).
Table 1. Occurrence of Tomato yellow leaf curlvirus (TYLCV) in eastern andsouthern Africa.
Country Year of surveya
1996 1997 1998
Kenya 16/43 23/81 -
Tanzania - 73/271 162/344
Uganda - 5/35 10/58
a. Number of infected samples/number oftested samples on which squash blothybridization tests were performed using aspecific DNA probe for TYLCV from Israel(TYLCV-Is).
Tomato plants infected with TYLCVexhibit severe and varied symptoms.Common symptoms consist of leafcurling, yellowing, chlorosis of leafmargins, leaf distortion, reduction inleaf size, shortening of the internodesor stunting. In some cases, infectedplants show excessive branching andflower abscission. Symptoms of TYLCVvary with strain, tomato cultivars,plant age at the time of infection andenvironmental conditions. In easternand southern Africa, epidemics ofTYLCV occur during the summermonths.

159
Tomato Yellow Leaf Curl Virus in E. and S. Africa
TYLCV has been reported to have awide host range, including plantspecies belonging to Acanthaceae,Asteraceae, Canicaceae, Euphorbiacea,Leguminosae, Malvaceae, Oxalidaceae,Pedaliaceae, Plantaginaceae andSolanaceae (Singh and Reddy, 1993). InTanzania, TYLCV was detected in threenewly identified hosts, Achyranthesaspera, Euphorbia heterophylla andNicandra physalloides (Nono-Womdimet al., 1996). These weeds arewidespread and serve as a majorreservoir of TYLCV.
TYLCV is transmitted by thewhitefly Bemisia tabaci (Gennadius), notby seeds. In four African countries,Kenya, Malawi, Tanzania and Sudan,several populations of B. tabaci havebeen collected from TYLCV-infectedtomato samples and characterized(CIAT, 1998).
TYLCV diseases exhibit varioussymptoms indicating the existence ofdifferent strains. Preliminaryidentification studies conducted in1994 showed that several tomatosamples from Tanzania, with typicalleaf curl and yellowing symptoms, didnot hybridize with the Egyptian and/orIsrael DNA probes (Nono-Womdim etal., 1996). Two of these Tanzanian leafcurl samples were positive forbegomovirus by polymerase chainreaction (PCR) using a primer whichspecifically amplifies part of thereplicase (AC1) open reading frame(ORF), the intergenic region and thecoat protein (AV1) ORF of whitefly-transmitted begomoviruses.Furthermore, the nucleotide sequenceof the PCR fragment was comparedwith the sequences of several distinctbegomoviruses, including TYLCV-Is,TYLCV-Th, TYLCV-Sar, TYLCV-Ind, etc.that infect tomato, as well as with thesequence of ACMV, ICMV and Mungbean yellow mosaic virus (MYMV). It wasfound that the Tanzanian isolate shows
less than 80% nucleotide identitieswith these other begomoviruses(Chiang et al., 1997). Since it has beenproposed that two isolates of the samebegomovirus species have nucleotidesequence greater than 90%, theTanzanian begomovirus, TYLCV-Tz, isdifferent from all previouslycharacterized begomoviruses from theOld World. Recent survey studies haveshown that TYLCV-Tz is nowwidespread in Tanzania (Table 2). A fewleaf curl samples from Malawi haveshown a weak positive reaction insquash blot hybridization with the DNAprobe of TYLCV-Tz. These resultsindicate that this virus, or anotherclosely related begomovirus, might bepresent in this country.
Recently, another study conductedon the identification of leaf curl virusesof tomato indicated the presence of anewly identified virus in Uganda thathas 73% homology with TYLCV-Is and78% homology with ToLCV-Tz. The leafcurl virus from Uganda is tentativelynamed ToLCV-Ug (Charles Sskyewa,personal communication, 1999).
These findings indicate thepresence of at least three different leafcurl viruses of tomato in eastern and
Table 2. Preliminary results on thedistribution of tomato leaf curldisease in Tanzania using twospecific DNA probes against tomatoyellow leaf curl virus-Egypt (TYLCV-Eg) and Tomato leaf curl virus-Tanzania (ToLCV-Tz).
Region Disease incidencea
TYLCV-Is ToLCV-Tz
Arusha 13/40 39/40
Dodoma 79/113 96/113
Kilimanjaro 12/30 29/30
Morogoro 34/70 60/70
Zanzibar 5/29 6/29
a. Number of positive samples in DNAhybridization tests/number of samples withleaf curl symptoms.

160
Whiteflies and Whitefly-borne Viruses in the Tropics
southern Africa. The causes of this newemergence of leaf curl viruses in Africahave not been investigated yet.
Management
Several management strategies tocontrol TYLCV and associate viruseshave been reported (Singh and Reddy,1993; Polston and Anderson, 1997).These strategies include culturalpractices, control of the insect vectorand the use of resistant varieties. Ineastern and southern Africancountries, the implementation ofcultural practices such as use of virus-free seedlings, reduction of virusinoculum (roguing), elimination ofsources of infection and intercroppingto control TYLCV has not beenperformed at farmers’ field level. ManyAfrican farmers still cannot recognizeTYLCV in order to apply these culturalpractices for management. However,the use of insecticides, primarily tocontrol whiteflies and not TYLCV, isvery common in eastern and southernAfrica (CIAT, 1998).
Experiments have been conductedsince 1995 at AVRDC Africa RegionalProgram to identify tomato varietieswith resistance to TYLCV (Table 3). Thevarieties Fiona and Tyking are resistantto TYLCV in Tanzania. Plants of thesespecies are symptomless and TYLCV isnot detected by DNA hybridization.Generally, resistance to TYLCV is quitespecific. For instance, tomato cultivarswith resistance or tolerance to TYLCVfrom the Old World are susceptible totomato leaf curl viruses from theWestern Hemisphere (Polston andAnderson, 1997). The situation seemsto be different in Africa. In recent fieldscreening tests, Fiona, Tyking andTY52, which are resistant to TYLCV,were found to resist ToLCV-Tz as well.Such lines will provide an efficientcontrol strategy for the management of
TYLCV and ToLCV-Tz since bothviruses occur in mixed infections inTanzania.
Conclusion
This chapter summarizes recentoutbreaks of TYLCV and leaf curl-likeviruses in eastern and southern Africa.The virus was first reported in Sudanin the 1960s, where it has been athreat for several decades. Then, in1990, it was found in Tanzania. Todate, TYLCV is widespread in manycountries of the subregion. It is notwell known whether TYLCV hasoccurred in these countries from sometime ago, as in Sudan, or whether ithas spread southwards fromMediterranean countries.
Until 1997, TYLCV was the onlybegomovirus reported to cause diseaseon tomato in eastern and southernAfrica. Two additional viruses, namely
Table 3. Field screening of Lycopersiconaccessions for resistance to Tomatoyellow leaf curl virus.
Lycopersicon Final diseaseaccessions incidence (%)
Chepertylc C-1 964-2 95FI-3 100J-7 100L-7 100Sin-3 100Siv-5 100Siv-6 100
Columbian 36 100EC-104395 100Jackal 55LA 3214 100LA 3216 95Lignon C8-6 100PSR-403511 5PSR-407111 9Roza 100Ty-20 548476 75Fiona 0Tyking 0MoneyMaker 100Roma 100

161
Tomato Yellow Leaf Curl Virus in E. and S. Africa
ToLCV-Tz and ToLCV-Ug, which aredifferent from TYLCV, have beenreported recently (Chiang et al., 1997;Charles Sskyewa, personalcommunication, 1999). Recent surveysindicate that many tomato leaf curlsamples do not hybridize with theavailable cDNA probes in nucleic acidhybridization tests. This may indicatethe existence of additional Africanindigenous leaf curl viruses.
In Tanzania, tomato varieties withresistant to TYLCV under fieldconditions have been identified. Theseresistant tomatoes are hybrid varietiesthat many farmers in eastern andsouthern Africa cannot afford.
At AVRDC, a breeding programmehas been initiated aimed atincorporating the TYLCV resistancederived from Lycopersicon chilense(Zakay et al., 1991) into cultivatedtomato. However, until open-pollinatevarieties with resistance to TYLCVbecome available, there is a need todevelop alternative control strategiesfor African farmers that will includecontrol of the insect vector and culturalpractices.
References
AVRDC (Asian Vegetable Research andDevelopment Center). 1996.Progress report 1995. Shanhua,Tainan, TW.
AVRDC (Asian Vegetable Research andDevelopment Center). 1997.Progress report 1996. Shanhua,Tainan, TW.
AVRDC (Asian Vegetable Research andDevelopment Center). 1998.Progress report 1997. Shanhua,Tainan, TW.
Chiang, B. T.; Nakhla, M. K.; Maxwell, P.;Schoenfelder, M.; Green, S. K.1997. A new geminivirus associatedwith a leaf curl disease of tomato inTanzania. Plant Dis. 81:111.
CIAT (Centro Internacional de AgriculturaTropical). 1998. Sustainableintegrated management of whitefliesas pests and vectors of plantviruses in the tropics. Progressreport. Cali, CO. 131 p.
Czosnek, H.; Ber, R.; Antignus, Y.; Cohen,S.; Navot, N.; Zamir, D. 1988.Isolation of Tomato yellow leaf curlvirus, a geminivirus. Phytopathology78:508-512.
Czosnek, H.; Navot, N.; Laterrot, H. 1990.Geographical distribution of Tomatoyellow leaf curl virus. A first surveyusing a specific DNA probe.Phytopathol. Mediterr. 29:1-6.
Givord, L.; Fargette, D.; Kounounguissa,B. R.; Thouvenel, J. C.; Walter, B.;Van Regenmortel, M. H. V. 1994.Detection of geminiviruses fromtropical countries by a doublemonoclonal antibody ELISA usingantibodies to African cassavamosaic virus. Agron. 14:327-333.
Harrison, B. D.; Barker, H.; Bock, K. R.;Guthrie, E. J.; Meredith, G.;Atkinson, M. 1977. Plant viruseswith circular single-stranded DNA.Nature 270:760-762.
Ioannau, N.; Iordanou, N. 1985.Epidemiology of Tomato yellow leafcurl virus in relation to thepopulation density of its whiteflyvector, Bemisia tabaci (Genn.). Tech.Bull. 71, Agricultural ResearchInstitute, Nicosia, CY. 7 p.
Khey-Pour, A.; Bendahmane, M.; Matzeit,V.; Accotto, G. P.; Crespi, S.;Groenenborn, B. 1991. Tomatoyellow leaf curl virus from Sardiniais a whitefly-transmittedmonopartite geminivirus. NucleicAcids Res. 19:6763-6769.

162
Whiteflies and Whitefly-borne Viruses in the Tropics
Macintosh, S.; Robinson, D. J.; Harrison,B. D. 1992. Detection of threewhitefly-transmitted geminivirusesoccurring in Europe by tests withheterologous monoclonalantibodies. Ann. Appl. Biol. 121:297-303.
Makkouk, K. M.; Shehab, S.; Majdalani,S. E. 1979. Tomato yellow leaf curl:Incidence, yield losses andtransmission in Lebanon.Phytopath. Z. 96:263-267.
Navot, N.; Pichersky, E.; Zeidan, M.;Zamir, D.; Czosnek, H. 1991.Tomato yellow leaf curl virus: Awhitefly-transmitted geminiviruswith a single genomic component.Virology 185:151-161.
Nono-Womdim, R.; Swai, I. S.; Green, S.K.; Gebre-Selassie, K.; Laterrot, H.;Marchoux, G.; Opena, R. T. 1996.Tomato viruses in Tanzania:Identification, distribution anddisease incidence. J. S. Afr. Hort.Sci. 6:41-44.
Polston, J. E.; Anderson, P. 1997. Theemergence of whitefly-transmittedgeminiviruses in tomato in thewestern hemisphere. Plant Dis.81:1358-1369.
Roberts, M. I.; Robinson, D. J.; Harrison,B. D. 1984. Serologicalrelationships and genomehomologies among geminiviruses. J.Gen. Virol. 65:1723-1730.
Singh, S. J.; Reddy, M. K. 1993. Leaf curlvirus disease of tomato and itsmanagement. Vatika from the Seedand Plant People 2:5-21.
Yassin, A. M.; Nour, M. A. 1965. Tomatoleaf curl disease, its effect on yieldand varietal susceptibility. SudanAgric. J. 1:3-7.
Zakay, Y.; Navot, N.; Zeidan, M.; Kedar,N.; Rabinowitch, H. D.; Czosnek, H.;Zamir, D. 1991. ScreeningLycopersicon accessions forresistance to the Tomato yellow leafcurl virus: Presence of viral DNA andsymptom development. Plant Dis.75:279-281.

163
Conclusions and Recommendations
CHAPTER 2.7
Conclusions and RecommendationsLisbeth Riis, Thomas Njuguna, Rebecca Raini,
Berhard Löhr and Mohamed Ali Bob*
Results of CountryStudies
Bemisia tabaci (Gennadius) research inthe eastern African region has beenfocused so far on cotton (Gossypiumhirsutum L.) in Sudan and on cassava(Manihot esculenta Crantz) in Kenya,Tanzania and Malawi. In recent years,vegetables have become the crops mostseverely affected by B. tabaci-transmitted viruses. Little attentionhas been directed to vegetablestherefore little information wasavailable prior to the commencement ofthe System-wide Tropical WhiteflyIntegrated Pest Management (TWF-IPM)Project in this region. For the first time,information about critical aspects ofB. tabaci characterization on thevegetable-based cropping system hasbeen reported from the eastern Africaregion.
Characterization ofbegomovirus and whiteflybiotypes
B. tabaci is the most dominatingwhitefly species in the region as awhole. All specimens of whiteflycollected in Sudan were B. tabaci; inTanzania, B. tabaci made up 78% ofthe collected specimens and in Kenya,65%. The species Trialeurodesvaporariorum (Westwood), Bemisia afer
(Priesner & Hosney), Siphoninusphillyreae (Haliday), Aleurothrixusfloccosus (Maskell), Trialeurodes ricini(Misra), Orchamoplatus citri(Takahashi), Tetraleurodes andropogon(Dozier) and Aleyrodes proletella(Linnaeus) were occasionallyencountered in Malawi, Kenya andTanzania, of which the last four specieswere registered for the first time inTanzania. Bemisia hirta spec. nov. wasencountered once but is subject toconfirmation. A. floccosus wasencountered on citrus. In the EastAfrican highlands, B. tabaci was thedominating whitefly species from sealevel up to 1300-1700 m above sealevel, above which T. vaporariorum andT. ricini dominated. In southern Africa(Malawi), B. tabaci only dominated upto 900 m above sea level, above whichB. afer was the dominating species.
Disease incidence andsymptom severity
The Phase 1 survey revealed silver leafsymptoms on squash (Cucurbita pepoL.) in Sudan. Elsewhere, thesesymptoms are associated with B. tabacibiotype-B as the causal pest. This isthe first observation of silver leafsymptoms in the region and mayindicate that this feared biotype-B hasreached Sudan. Characterization of theB. tabaci specimens collected at thesite where the silver leaf symptomswere found is underway and thebiotype-B is yet to be confirmed.
* International Center of Insect Physiology andEcology (ICIPE), Nairobi, Kenya.

164
Whiteflies and Whitefly-borne Viruses in the Tropics
B. tabaci was found widespread ontomato (Lycopersicon esculentum Mill.),pepper (Capsicum spp. L.) and eggplant(Solanum melongena L.), in theSolanaceae; watermelon (Citrulluslanatus [Thunb.] Matsum. & Nakai),melon (Cucumis melo L), squash,cucumber (Cucumis sativus L. var.sativus) and bitter gourd (Momordicacharantia L.), in the Cucurbitaceae;common bean (Phaseolus vulgaris L.),lablab (Lablab purpureus [L] Sweet),Leguminosae family; cassava(Euphorbiaceae); okra (Abelmoschusesculentus [L.] Moench), cotton,sweetpotato (Ipomoea batatas [L.] Lam.),Desmodium and basil (Ocimum basilicumL.), in the Labiateae. In addition tothese families, B. tabaci also was foundfrequently on wild plant species withinthe families of Acanthaceae,Amaranthaceae, Asteraceae,Commelinaceae and Verberaceae.
Whiteflies in association withTomato yellow leaf curl virus (TYLCV)had been studied in the Sudan (Dafallaand Sidig, 1997) and Tomato leaf curlvirus (ToLCV) symptoms in Tanzaniahad been found due to a unique strainnamed ToLCV-Tz (Chiang et al., 1997).Since tomato was known as one of themost profitable crops, and whiteflieshad been ranked as the most/secondmost important pest/vector in tomato(Varela and Pekke, 1995), tomato wasselected as the target crop in anextensive survey on whiteflies andwhitefly-transmitted viruses (WTVs) invegetable production in the region.
Tomato yellow leaf curl (TYLC)symptoms have been identified ascaused by TYLCV in Sudan, Kenya andMalawi. TYLC/tomato leaf curl (ToLC)symptoms were found in more than93% of the surveyed fields of tomato inSudan and Tanzania, in 72% in Malawiand 55% in Kenya. More than 25%TYLC incidence was found in 50% ofthe fields in Sudan, 34% in Tanzania,
11% in Kenya and 6% in Malawi. Inthe East African highlands, high ToLCincidence (>70%) was found from500-1000 m above sea level in Kenyabut from 50-1500 m above sea level inTanzania. TYLC symptoms areparticularly severe during the hotseasons (December to March inTanzania and Kenya; March toSeptember in Sudan).
Reservoirs for TYLCV in the wildvegetation were found in plant speciesbelonging to the families of Solanaceaeand Euphorbiaceae, in particular, andin Amaranthaceae.
In Sudan, the experienced teamsurveyed WTVs in all commonly grownvegetable crops. TYLCV in tomato is byfar the most economically importantB. tabaci-transmitted virus in Sudan.Next ranks Watermelon chlorotic stuntvirus (WmCSV) and Okra leaf curl virus(OLCV). TYLCV and WmCSV areparticularly severe during the hotseason growing periods (March-September) and OLCV during the latewinter and early summer (January-March). Other severe WFTV wereTomato vein thickening virus (TVTV) intomato, Cucumber vein yellowing virus(CVYV) and a yellowing-inducing poty-like virus not fully identified inwatermelon and musk melon (Cucumismelo L.). Less important WTVs, yetrepresenting a potential hazard, areTYLCV in pepper, WmCSV in snakemelon (Cucumis melo L. subsp. melovar. flexuosus [L.] Naudin) and squash,a begomovirus and a closterovirus notyet fully identified in Vigna sp. andcommon bean (Phaseolus vulgaris L.var. vulgaris), Bean mild mosaic virus(BMMV) and Pepper leaf curl virus(PepLCV).
The African Regional Program (ARP)of the Asian Vegetable Research andDevelopment Center (AVRDC) hasidentified three tomato accessions

165
Conclusions and Recommendations
(Fiona, Tyking and TY52) resistant toTYLCV and ToLCV-Tz. Under heavydisease pressure, these accessions aresymptomless and the viruses are notdetected by DNA hybridization.Furthermore, two tomato accessions(PSR-403511 and PSR-407111) wereidentified as tolerant to TYLCV; only5%-9% disease incidence was detectedunder heavy disease pressure.
Natural enemy species
Encarsia sophia (Girault and Dodd) wasthe predominant whitefly parasitoid inKenya, Tanzania and Malawi, but wasnot encountered during the survey inthe Sudan. Encarsia lutea (Masi) andEretmocerus mundus Mercet. werepredominant in the Sudan but not inthe other countries, althoughEretmocerus spp. were occasionallyencountered in Kenya and Tanzania.Encarsia formasa (Gahan) wasencountered in the highlands of Kenya.Predators of coccinelids, lacewings,bugs, spiders and mites were foundadjacent to whitefly populations andtheir identification at species level isyet to be confirmed.
Estimation of diseaseincidence and yield losses
In Sudan, the “hot spots” for B. tabaciand transmitted viruses are the aridand irrigated cotton-growing areas inGezira and Khartoum States, at400-500 m, where the problem issevere throughout the year but peakingin March to September. Where cottonis not grown, the incidence is less. InTanzania, the hot spots for B. tabaciand TYLC are transitional areas inMorogor, Dodoma, Kilimanjaro andArusha, ranging from 100-1450 m. InKenya, the hot spots are semi-arid totransitional areas in Eastern Province(Kibwezi, Kitui and Machacos) and RiftValley (Naivasha), 700-1200 m. InMalawi, the hot spots are in southernareas of the Lake Malawi region
(Dedza, Mzimba, Thyolo and Ntcheu),600-1500 m. In Tanzania, Kenya andMalawi the problem is most severeduring the hot dry season, December toMarch. Irrigation in the semi-arid andarid regions allows for continuouscultivation of crops favoured byB. tabaci and transmitted viruses,which guarantees continuousavailability of reproductive hosts duringthe year.
In hot spot areas throughout theregion, yield losses due to B. tabaci andtransmitted viruses ranged between50% and100%. As an average of allsurveyed tomato fields, the yield lossesdue to B. tabaci and TYLC were 49% inMalawi, 45% in Tanzania and 30% inKenya. In Sudan, the survey revealedthat an average of 62% fruit yields oftomato, watermelon, musk melon andokra are lost annually due to B. tabaci-transmitted viruses. WmCSV wasimplicated in up to 80% yield losses ofexport Galia melon in Khartoum andGezira States during the 1998-99season, and completely wiped out theproduction in southern Blue Nile,Shuwak and Dinder inland delta. Inthe region in general, more than 65%of producers reckoned that they hadwhitefly/disease problems in theirtomato crop every year and 1997 wasthe year with the most severe attack(prior to the onset of the El Niño rains).On average, producers in the regionspend US$145 per hectare combatingwhiteflies.
Farmers’ assessment ofwhitefly-related problems
In Sudan, farmers ranked B. tabaci andits transmitted viruses as the mosteconomically and most damaging cropprotection problem in tomato in thecountry. In Tanzania, farmers rankedB. tabaci and its transmitted viruses asa second problem in tomato afterbacterial blight. In Kenya and Malawi,

166
Whiteflies and Whitefly-borne Viruses in the Tropics
B. tabaci and its transmitted virusesranked less important. Nevertheless,79% of the interviewed farmers inKenya found that the problems were onthe increase compared with 74% inTanzania, 68% in Malawi and 65% inSudan.
It was generally believed that thewhitefly/disease complex is mostdamaging during dry and hot seasons(December through March in Tanzaniaand Kenya, March through August inthe Sudan). In Malawi, however,controversial opinions on theseasonality of outbreaks were foundamong the interviewed producers.Interestingly, in the Sudan, TYLCVincidence was most severe during thehot summer months despite lessprevalence of the vector, whosepopulation peaked during the coolerwinter months (December throughJanuary).
Pesticide use
Eighty three percent of the farmersinterviewed in the region appliedpesticides in their tomato crops. Thepercentage of farmers making morethan nine pesticide application pertomato cropping season was 56% inSudan, 21% in Tanzania, 9% in Malawiand 8% in Kenya. The use of syntheticinsecticides was the most prevailingcontrol method and in most cases theproducer alone made the decision onwhat insecticide to use and when(average 84%). Half of the producerssprayed insecticides as a preventionmeasure and 25% of them would startspraying after damage had beenobserved. Five to six sprayings perseason was practiced on average but inthe Sudan, 56% of the interviewedfarmers made nine or more applicationsper season. Pyrethroids andorganophosphates were the mostcommonly used insecticides in theregion.
Only 35% of the interviewedproducers in Sudan, 11% in Tanzania,1% in Kenya and none in Malawi knewabout the inter-relationship betweenwhiteflies and ToLCV symptoms. InMorogoro in Tanzania, where 100%ToLCV incidence was found on all thesurveyed farms, none of the producersknew about its inter-relationship withwhiteflies. In Malawi, almost allproducers confused TYLC symptomswith other damage such as damagefrom red spider mite, aphids, heavyrain, fungal diseases, nitrogendeficiency and soil-borne diseases.
Sudan is the only country in theregion that has adopted IPM as itsofficial policy of crop protection aimed atcombating the whitefly problem.Government support for IPM is welldefined in the recent publication, SudanCountry Strategy Note, 1997-2001:Partnership towards SustainableHuman Development. The documentemphasizes that the Government of theSudan is in pursuit of an integratedprogram for environmentallysustainable development. However,there is at present a laxity inimplementation and in cultural andlegislative measures. There are norestrictions on the number of pesticideapplications. Sudan has no extensionservice at all, whereas technicaladvisors from extension services havevisited about one-third of theinterviewed producers in the othercountries.
Strengthened ResearchCapacity
The TWF-IPM Project has helped inincreasing the capacity in whiteflyresearch by training one M.Sc. studentfrom Kenya and one Ph.D. student fromSudan. With assistance/input from theInstitute of International Education(IIE), London, and the John Innes

167
Conclusions and Recommendations
Centre (JIC), project scientists weretrained in conventional taxonomy andDNA techniques. A short-termattachment also was completed on theeffect of host plants on the ovipositionand survival of the sweetpotato whitefly.After long fruitless efforts, finallycultures of Bemisia and Trialeurodeswere successfully established at theInternational Center of InsectPhysiology and Ecology (ICIPE). TheCentre, AVRDC and the nationalpartners are planning work ontransmission studies and hostsuitability studies in Phase 2. The hotspot areas of whitefly problems inpartner countries have been identified.Besides confirming the local occurrenceof several known natural enemies ofwhiteflies, additional genera/species ofpredators/parasitoids have beenidentified. Perceived on-farm losses dueto tomato geminiviruses and theaverage cost to control whitefly/TYLCper hectare of tomato in one seasonwere documented.
ICIPE has extended assistance inthe identification of whitefly species andnatural enemy collections from surveysacross nine countries in Africa to thesubproject on whiteflies on cassava andsweetpotato, led by the InternationalInstitute of Tropical Agriculture (IITA).Likewise, ICIPE has extendedassistance in the identification ofwhiteflies and natural enemies tocountries that were not participatingofficially in this project, for example,Uganda and Senegal. The Center hasestablished a reference collection forAfrican whiteflies in the past 2 yearsfrom 10 countries within the frameworkof the two African whitefly subprojects.This is in connection with a collection ofabout 2000 samples of whiteflies andmore than 4000 nymph specimensmounted on permanent slides, inaddition to about 400 whiteflyparasitoids and predators collected. It ishoped that this collection will further
help understand whiteflies in futurestudies and training.
This project demonstrates howeffective collaboration can result inproper identification of researchagendas and their correctimplementation. This has been possibleonly because of the joint efforts of thenational programmes and thecollaborators in a well-coordinatedapproach. The joint efforts of the ICIPEscientists and the national teams havefurther strengthened ICIPE’spartnership with national programmesin Africa. This mode of collaborationand the complementation betweenICIPE, AVRDC, JIC and nationalagricultural research systems scientistshas helped link up the expertise ofthese centres and the local knowledgeand experiences of the nationalprogrammes. The adoption of astandardized methodology and beingable to modify it to suit specificrequirements has simplified work andmade it possible to compare results andshare experiences.
Recommendations
As a result of a joint planning workshopin September 1999, the partners inSub-project 3 (Whiteflies as vectors ofviruses in vegetable and mixed croppingsystems in eastern Africa) made thefollowing recommendations for thesecond project phase.
It was proposed that Phase 2should include Uganda in addition toSudan, Tanzania and Kenya. The focusshould remain on B. tabaci as vector ofviral diseases. The target croppingsystem in Kenya and Uganda willremain tomato. Target cropping systemswill include both tomato andwatermelon in Tanzania and tomato,cucurbits and okra in the Sudan. Thereis a need to continue the

168
Whiteflies and Whitefly-borne Viruses in the Tropics
characterization of whiteflies and WTVsin target cropping systems. Thischaracterization comprehends bothcharacterization at molecular level,including bio-typing of whiteflies, andbiological characterization of bothwhiteflies and WTVs with emphasis onhost range, host preferences andinteraction between the whiteflies andthe viruses.
The aim is to continuestrengthening the global whiteflynetwork set up by the TWF-IPM Project.Through this network, scientists fromthe region will enjoy the informationflow of knowledge and participation inexperimental research in order to boostthe whitefly research in their respectivecountries with new knowledge anddisseminate new control methods to thenational extension services andvegetable producers. Initiatives taken tocreate an African Whitefly Networkshould continue.
More evidence must be recorded onyield depression due to whiteflies andtheir transmitted viruses. Geographicalinformation on agricultural regions,major crops, target crops, whitefly/virus hot spots, topographical andclimatic data, crop seasonality andspatial patterns of cropping systemsmust be collected in order to applygeographic information systems to theinformation created.
There is an urgent need for theimmediate development of IPMcomponents viable under the socio-economic conditions in the region.Selection, multiplication anddissemination of tomato varietiesresistant/tolerant to whitefly and/orToLCVs were given first priority in thedevelopment of the IPM component.AVRDC should gather and multiply a“package” of resistant germplasm fromall over the world and distribute it tothe national partners for screening,
selection, multiplication anddissemination.
Development of physical barriersfor vector control was given secondpriority in developing the IPMcomponent. Different physicalscreening methods to prevent earlyinvasion of whiteflies should begathered from all over the world andtested as physical control measuresunder the socio-economic and climaticconditions in each partner country.
Testing rational use of conventionaland non-conventional insecticides wasgiven third priority in developing theIPM component. Producers urgentlyneed alternative control methods inorder to meet the new requirements ofpesticide residue-free products for theexport market. Regional monitoring ofinsecticide resistance of whiteflies willprovide useful information to convincepolicy makers of the need fordeveloping alternative control methods.Non-conventional and indigenousmethods from all over the world suchas oil, non-phytotoxic soaps, botanicalsand ash should be gathered in a“package” and tested in partnercountries by means of a standardizedmethodology that will allowcomparisons across ecosystems.Evaluation of the effect of bothconventional and non-conventionalcontrol methods on natural enemiesand biological control agents such asparasitoids and entomopathogens willreveal information on their potentialenvironmental impact and theirsuitability in organic farming.
Understanding of the mechanismbehind effective cultural practices inorder to define appropriate croppingsequences also was put on the agenda.Such studies may require several yearsand experimental cropping seasonsand may not provide immediateapplicable information for producers as

169
Conclusions and Recommendations
the target stakeholders. The priority ofsuch studies was therefore set to followthe above IPM components. Suchstudies would evaluate the effect ofdiversity-based cropping in time andspace, planting dates, planting density,ground cover, roguing, in-field weedmanagement, crop residue managementand micro-environment modification.
Studies on biological control ofwhitefly vectors were given less priority.Since low vector populations can createa severe virus outbreak, it is believedthat biological control may have lowimpact in the control of virus outbreaksdriven by the movement of whiteflyvectors. It was decided to startgathering cases of successful legalcontrol methods and relateddocumentation. When sufficientevidence has been provided through theproject work, the aim is to startreinforcing official regulation andinspection for area-wide management,including planting date, harvestingdate, crop free periods, destruction ofcrop residues, irrigation cut-off, etc.
The research capacity in thepartner countries must be strengthenedthrough training of national scientistsin detection, identification andcharacterization of whiteflies, WTVs,natural enemies of whiteflies andepidemiology. Furthermore, yearlyinformation exchange and co-ordinationmeetings will be needed.
When feasible, producers must beactively involved in the development ofthe IPM component, including thebrainstorming, planning, realizationand evaluation of experiments. Traininghandouts on rational control ofwhiteflies and WTVs must be producedfor the extension service and producers,and farmers’ organizations in hot spotareas must be sensitized, in particular.
Decision support tools must beavailable for policy makers and newsmedia must be used currently as theproject develops.
Acknowledgements
The ICIPE-coordinated subproject hasderived valuable input in geminiviruscharacterization from JIC in the UK andfrom AVRDC in Tanzania. Dr. JamesLegg (IITA, Uganda) offered valuablesupport in linking with the IITA-ledwhitefly subproject on cassava/sweetpotato systems. The project alsohas received valuable assistance fromDr. Andrew Polaszek (CAB InternationalBioscience, UK Centre) and Dr. EddieUeckermann of the Plant ProtectionResearch Institute (PPRI), South Africain the identification of some whiteflynatural enemies. This book is a jointproduct of all partners and the ICIPEco-ordinating unit scientists.
References
Chiang, B. T.; Nakhla, M. K.; Maxwell, P.;Schoenfelder, M.; Green, S. K. 1997.A new geminivirus associated with aleaf curl disease of tomato inTanzania. Plant Dis. 81: 111.
Dafalla, G. A.; Sidig, S. A. 1997.Management of the whitefly-viruscomplex in Sudan. Food andAgriculture Organization (FAO) PlantProduction and Protection Paper143. Rome, IT. 214 p.
Varela A. M.; Pekke, A. 1995. Proceedingsof the Tomato Planning Workshopfor Eastern and Southern AfricaRegion, 16-20 October 1995, Harare,ZW. Deutsche Gesellschaft fürTechnische Zusammenarbeit (GTZ)-Integrated Pest Management (IPM)Horticulture Project, Nairobi, KE.32 p.

BLANCA 170

SECTION THREE
Whiteflies as Vectors of Viruses inLegume and Vegetable Mixed
Cropping Systems in the TropicalLowlands of Central America, Mexico
and the Caribbean
SECTION THREE
Whiteflies as Vectors of Viruses inLegume and Vegetable Mixed
Cropping Systems in the TropicalLowlands of Central America, Mexico
and the Caribbean

BLANCA 172

173
Introduction
CHAPTER 3.1
IntroductionFrancisco Morales*
Establishing anInternational Network
Organizing an international network onwhitefly-transmitted viruses in a regionas extensive as Central America,Mexico and the Caribbean is, forvarious reasons, no easy task. First,whitefly-borne viruses have affectedmore than a dozen countries andcommercial crops in this region.Second, most of these countries lackthe necessary research infrastructureor resources to study plant viruses.Third, most of the research conductedin the region has been focused on thewhitefly Bemisia tabaci (Gennadius)(Homoptera: Aleyrodidae) as a pest andonly rarely as a vector. And, finally,communication among researchers inthe region remains difficult. However,this was the first task facing partnersin Sub-project 2 of the TropicalWhitefly Integrated Pest Management(TWF-IPM) Project. Sub-project 2,Sustainable integrated management ofwhiteflies as pests and vectors of plantviruses in the tropics, is led by theCentro Internacional de AgriculturaTropical (CIAT) on behalf of theSystemwide Programme on IntegratedPest Management.
Fortunately, a Regional Action Planfor Whitefly Management had been
created in 1992 under the initiative ofvarious international institutions suchas the Centro Agronómico Tropical deInvestigación y Enseñanza (CATIE) inCosta Rica, the Pan AmericanAgricultural School at Zamorano inHonduras and the InstitutoInteramericano de Cooperación para laAgricultura (IICA). The network haspromoted annual meetings of regionalscientists interested in controlling thisimportant pest, mainly under thecoordination of CATIE. Anotherimportant network that operates in theregion is the Programa CooperativoRegional de Frijol para Centro América,México y El Caribe (PROFRIJOL),financed by the Swiss Government topromote the production of commonbean, Phaseolus vulgaris L.(Leguminosae), in the region. A mainproduction problem of common bean inthe lowlands and the mid-altitudevalleys of Central America, Mexico andthe Caribbean has been Bean goldenyellow mosaic virus (Geminiviridae:Begomovirus), a disease caused by awhitefly-transmitted virus. BothPROFRIJOL and CIAT have givenconsiderable attention to this problemsince 1978 through a collaborativeresearch agreement. The existence ofthese networks greatly facilitated thetask of organizing an internationalnetwork to focus specifically on virusestransmitted by the whitefly B. tabaci inlegume and horticultural crops in theregion.
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.

174
Whiteflies and Whitefly-borne Viruses in the Tropics
To further strengthen this network,Sub-project 2 selected threesubregional coordinators and threeadvanced research institutions withprevious research experience in each ofthe three subregions. The firstsubregion selected was CentralAmerica and the advanced researchinstitution chosen to provide technicalsupport was the Plant PathologyDepartment of the University ofWisconsin, Madison, USA. Thesubregional coordinator was Dr. LukoHilje, an entomologist at CATIE, andthe responsible scientist for theUniversity of Wisconsin wasDr. Douglas Maxwell. Subregion 2 wasMexico, and its support researchinstitution was the University ofArizona, in Phoenix, USA. Thesubregional coordinator was Dr. RafaelRivera Bustamante of the Centro deInvestigaciones y Estudios Avanzados(CINVESTAV) in Irapuato Mexico, andthe responsible scientist for theUniversity of Arizona was Dr. Judith K.Brown. Subregion 3 was theCaribbean, and its support institutionwas the University of Florida,Bradenton, USA. The subregionalcoordinator was Dr. Colmar Serra, ofthe Instituto Superior de Agricultura(ISA), Dominican Republic, and theresponsible scientist at the Universityof Florida, Dr. Jane Polston. Later onduring the development of the project,Drs. Hilje and Serra stepped down fromtheir respective coordination roles andthe coordinator of Sub-project 2,Dr. Francisco Morales, assumed theseresponsibilities.
Assessing Socio-economicand Environmental
Impact
The large number of countries includedin Sub-project 2 precluded theimplementation of socio-economicsurveys in each country. Nevertheless,
four countries in Central America—Guatemala, El Salvador, Honduras andCosta Rica—were selected to conductthese studies, based on the followingconsiderations. First, nationalinstitutions were available that werewilling to conduct the surveys. Second,a predominant small-scale farmingsystem could be identified in the threecountries and study areas selected.Third, a truly diversified agriculture,including legumes and horticulturalcrops, could be found in the studyareas selected. These conditions werenot satisfied in most of the areassurveyed in Mexico or the Caribbeanregion, with the exception of Cuba,which was excluded on the basis ofother logistical constraints.
Another consideration was that, inMexico and the Caribbean region,statutory measures to counter whiteflyand virus epidemics have beensuccessfully implemented. The nationalprogram scientists who have had theresponsibility of implementing theregulatory measures will discuss thecase of the Dominican Republic(Chapter 3.11, this volume).
Emergence of Whitefly-Transmitted Viruses
A number of historical, economic andbiological factors have been identifiedas being associated with the emergenceof whitefly-borne viruses in legumesand horticultural crops in the regioncovered by Sub-project 2. These will bediscussed in the individual countrypapers and in the concluding chapterof this section (Chapter 3.14, thisvolume). However, in introducing thissection, two crosscutting trends can behighlighted. First, the presence ofwhitefly-transmitted viruses isfrequently associated with thecultivation of large-scale commercial ornon-traditional horticultural crops,

175
Introduction
especially for export. Second, thedisintegration of national agriculturalresearch programs throughout thestudy region has had a negative impacton the management of whitefly-relatedproduction problems. Basically, thelack of technical assistance to controlthe whitefly vector has left producers inthe hands of pesticide vendors. As aconsequence, pesticide abuse hasresulted, with serious biological andenvironmental consequences.
The economic situation in LatinAmerica has been deteriorating sincethe late 1970s, when the oil crisisgenerated a gigantic debt for mostcountries in the region. Possessing onlyan incipient industrial capacity, mostcountries had to resort to the export ofnon-traditional crops such as soybean(Glycine max [L.] Merr.) and vegetables.This strategy produced complexchanges in the region’s agriculturalenvironment that national agriculturalresearch programs in Central America,Mexico or the Caribbean have notaddressed. The cropping systemsinvolved have been difficult to analysein this project because the productionlosses caused by whitefly-transmittedviruses have forced growers to abandonand replace crops frequently.Nevertheless, the Sub-project partnershave been able to document the rapidturnover of crops in whitefly-affectedareas and to analyse some of theconsequences.
Looking Ahead
A wealth of information, characterizingproblems associated with whitefly-transmitted viruses affecting legumesand horticultural crops in CentralAmerica, Mexico and the Caribbeanwas accumulated during thispreliminary phase of the TWF-IPMProject. Considerable progress wasmade in the use of a geographic
information system to analyse the datacollected, and in the creation of aninteractive system to allow potentialusers to access these data. The amountof data collected across this diverseregion considerably exceededexpectations and further analysis willbe required to extract all the valuableinformation it contains. However, whenthis task is complete, the knowledgegained will provide a sound foundationfor developing and implementingsustainable management strategies forwhiteflies and whitefly-borne viruses inthe region. Figure 1 shows the areasaffected by whitefly-transmittedviruses.
In the following chapters, thereader will find a brief description ofthe different historical, geographic,biological and socio-economic factorsthat contributed to the emergence ofthe whitefly B. tabaci as a major pestand vector of viruses (referred to hereas geminiviruses or, more specifically,as begomoviruses). Concludingchapters draw together resultsregarding the biotypes andreproductive hosts of B. tabaci in thisregion and, based on a synthesis of allresults, make recommendations forfurther research and implementationdirected towards achieving moreeffective and sustainable managementof whiteflies and whitefly-borne virusdiseases in the region.
Acknowledgements
The following chapters were written bythe senior author based on resultsobtained at the Plant VirologyLaboratory (CIAT, Cali, Colombia) fromsamples collected by the senior authoror collaborators in participatingcountries, as well as on the availableliterature and personal observationsand experience in the region. Hence,co-authors are not responsible for

176
Whiteflies and Whitefly-borne Viruses in the Tropics
omissions or errors found in thesechapters. Data provided bycollaborating national scientists aredutifully acknowledged in thecorresponding chapters. Severalnational scientists and studentscollaborating in the project conductedsocio-economic studies. Most of thesurvey data presented have beenpublished as theses by their mainauthors, also included as co-authors inthe corresponding chapters. Specialacknowledgements are due to theprofessional personnel of the CIATPlant Virology Laboratory, whoprocessed most of the samples ofwhiteflies and viruses analysed in this
extensive survey: Claritza Gómez, RaúlSedano, Ana Karine Martínez, AnaCecilia Velasco, Mauricio Castaño, JoséArroyave, Iván Lozano, Natalia Villarealand Maritza Cuervo. Special thanks aredue to Ms. María del Pilar Hernández,CIAT’s whitefly taxonomist, forexamining all the whitefly specimenscollected during this investigation. Lastbut not least, we recognize the work ofDr. Lee Calvert, who supervised themolecular characterization of B. tabacibiotypes and Trialeurodes vaporariorum(Westwood) (Homoptera: Aleyrodidae)specimens collected during thisinvestigation.
Figure 1. Regions in the Neotropics affected by whitefly-transmitted viruses (shaded areas).
Mexico
GuatemalaEl Salvador
Costa Rica
Panama
Nicaragua
Honduras
Cuba
Haiti

177
Mexico
CHAPTER 3.2
MexicoFrancisco Morales*, Rafael Rivera-Bustamante**,
Rafael Salinas Pérez***, Irineo Torres-Pachecoψ,Raúl Díaz Plazaψ, Wilson Avilés Baezaψ
and Genovevo Ramírez Jaramilloψ
Introduction
Geographical context
Mexico is the largest country inMiddle America, with an area of1,958,201 km2. The country can bedivided into different regions, thelargest being the Central Plateau,which extends from the United Statesborder in the north to the Isthmus ofTehuantepec in the south. At itsnorthern end, the plateau is about1320 m above sea level, rising to morethan 2650 m south of Mexico City, thecountry’s capital. This Central Plateauis divided into two parts—the Mesa delNorte extends from the United Statesborder to near San Luis Potosí, and theMesa Central from San Luis Potosí tojust south of Mexico City. The MesaCentral is a less arid, higher and flatterplateau than the Mesa del Norte.Although the Central Plateau containsmany agricultural basins and valleys,the altitude limits the survival andactivity of the whitefly Bemisia tabaci(Gennadius). Instead, other whitefliessuch as Trialeurodes vaporariorum
(Westwood) predominate. The latterspecies can become a major pest onsome crops, including common bean(Phaseolus vulgaris L.), tomato(Lycopersicon esculentum Mill.), potato(Solanum tuberosum L.) and otherhorticultural crops grown above 900 maltitude; but it is not a vector ofbegomoviruses. Figure 1 shows themain agricultural regions affected bywhitefly-transmitted begomoviruses.
The Central Plateau is flanked bythe Sierra Madre, two mountain rangesrunning west and east of the plateau.To the south is the Balsas Depression,a hot, dry and broken area withnumerous small basins. Further southis the Mesa del Sur, with small isolatedvalleys, 1300 m to 1650 m above sealevel. The Valley of Oaxaca is thelargest and most densely populated ofthese valleys. The Southern Highlandsconsist of a series of mountain rangesand plateaux, including the SierraMadre del Sur. The Isthmus ofTehuantepec is a narrow stretch oflowland (up to about 330 m altitude)with a hilly central area. The presenceof B. tabaci south of the CentralPlateau has not been well documentedand it seems that T. vaporariorum maypredominate as a pest of agriculturalsignificance. The Chiapas Highlandsare an extension of the mountainranges of Central America. The Sierrade Soconusco runs along the PacificCoast. The Grijalva river valley leads
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** Centro de Investigaciones y EstudiosAvanzados, Irapuato, Mexico.
*** Instituto Nacional de InvestigacionesForestales y Agropecuarias (INIFAP),Sinaloa, Mexico.
ψ INIFAP, based at Mérida, Yucatán, Mexico.

178
Whiteflies and Whitefly-borne Viruses in the Tropics
northwest into the Tabasco Plain, anextension of the Yucatán Peninsula,bordering the Gulf of Mexico. Thispeninsula consists of flat, limestoneterrain, seldom exceeding 160 m abovesea level. The northernmost coastalarea of the Yucatán Peninsula is animportant horticultural area and hasbeen affected by B. tabaci and differentbegomoviruses transmitted by thisvector.
The main horticultural areas ofMexico affected by B. tabaci are thecoastal lowlands that lie east and westof the Central Plateau. The GulfCoastal Plain extends some 1400 kmfrom the Texas border to the YucatánPeninsula. The triangular northernportion is more than 160 km wide nearthe US border, narrowing southwardsuntil it encounters the Sierra MadreOriental close to the sea, north ofTampico. This is a swampy area withseveral lagoons and is called the ZonaLagunera. South of this constriction,the Gulf Coastal Plain runs, narrowand irregular, all the way to theYucatán Peninsula. The narrowerPacific coastal lowlands begin near theMexicali Valley in the north and end,also some 1400 km to the south, near
Tepic, Nayarit. Parts of these aridregions have been irrigated since the1930s and support intensive croppingsystems of traditional and non-traditional (export) crops. Finally, BajaCalifornia is an isolated strip of aridland, about 1280 km long and 160 kmwide. Most of the eastern side of thispeninsula is mountainous, reachingelevations of almost 3000 m andsloping downwards towards the west.
The emergence of Bemisiatabaci as a pest and virusvector
The first report of B. tabaci as a vectorof plant viruses in Mexico, in the early1950s, is linked to the production ofcotton (Gossypium hirsutum L.) in theValley of Mexicali, Baja California(Cárdenas et al., 1996). The diseaseobserved was probably “cotton leafcrumple”, caused by a begomovirustransmitted by B. tabaci, previouslyobserved in the United States near theColorado River Valley (Brown, 1994).Around that time, B. tabaci was alsoassociated with a disease of tomato(Lycopersicon esculentum Mill.), called‘chino del tomate’, in the Valley of
Sonora
San Luis Potosí
Tamaulipas
Veracruz
YucatánNayarit
Sinaloa
BajaCalifornia
South BajaCalifornia
Chiapas
Mexico
Figure 1. The main agricultural regions affected by whitefly-transmitted begomoviruses, Mexico.
QuintanaRoo

179
Mexico
Culiacán, Sinaloa, in north-westernMexico (Gallegos, 1978). In 1962,B. tabaci was observed attacking cottonin the Soconusco region of the southernstate of Chiapas (Cardenas et al., 1996).
In 1978, B. tabaci causedconsiderable damage to horticulturalcrops in the Huasteca region, includingparts of the states of Hidalgo,Tamaulipas, San Luis Potosí andVeracruz, where the Gulf Coastal Plaintapers to form a narrow strip thatreaches all the way to the northernmostpoint of the Yucatán Peninsula. Adisease believed to be Bean goldenyellow mosaic virus (BGYMV) wasobserved for the first time in the north-western state of Sinaloa, in 1974(López, 1974). When this diseaseoccurred later (1990) in the state ofSonora, it was shown to be caused notby BGYMV but by a distinct virus, Beancalico mosaic virus (BCaMV) (Brown etal., 1990). BCaMV induces morestriking golden mosaic symptoms,which eventually resemble a bleachingeffect. The virus is not closely related toBGYMV but rather to Squash leaf curlvirus (SLCV), a virus previouslydescribed attacking cucurbits inCalifornia (Flock and Mayhew, 1981).BGYMV emerged on the Gulf Coast ofVeracruz in 1977. In 1979, it affectedcommon bean plantings in LasHuastecas and in 1980 affected thesame crop in the state of Chiapas(López-Salinas, 1994). Finally, in 1980,B. tabaci reached the YucatánPeninsula, particularly the state ofYucatán, where it caused severe yieldlosses to horticultural crops as a pestand virus vector (Díaz-Plaza et al.,1996).
Most of the subsequentdissemination of whitefly-transmittedbegomoviruses in Mexico has beenassociated with the boom inhorticultural and other export cropsthat took place in the 1980s. The
cultivation of melon (Cucumis melo L.),soybean (Glycine max [L.] Merr.), tomatoand peppers (Capsicum spp. L.), hascreated conditions favourable for thereproduction of B. tabaci away from itstraditional hosts such as cotton. Thesenon-traditional export crops arecurrently planted in B. tabaci-affectedregions such as the north-west, LasHuastecas and Yucatán. Among themost important viruses affectinghorticultural crops in Mexico may bementioned: Tomato chino (leaf crumple)virus, described in Sinaloa in 1970(Gallegos, 1978); Serrano golden mosaicvirus, in north-eastern Mexico (Sánchezet al., 1996) and Texas pepper virus(Stenger et al., 1990), both of which areregarded as Pepper golden mosaic virus(PepGMV); (Brown et al., 1993); PepperHuasteco yellow vein virus (PHYVV) inpepper and tomato in Las Huastecasand Sinaloa (Hou et al., 1996); and“El Tigre” virus of pepper in Tamaulipas(Brown, 1989). In Yucatán, chilli pepper(Capsicum spp.), tomato, squash(Cucurbita spp. L.), melon, andwatermelon (Citrullus lanatus [Thunb.]Matsum. & Nakai) have been attackedsince the 1980s by B. tabaci and thebegomoviruses this whitefly speciestransmits (Díaz-Plaza et al., 1996).
More recently, Tomato mottle virus(ToMoV), a begomovirus originallydescribed from Florida, United States,was detected in the Yucatán Peninsula,also affecting tomato (Garrido andGilbertson, 1998). In addition, PHYVVwas found in Habanero varieties of chillipepper and in pepper, and PepGMV wasfound in Habanero. PHYVV had alreadybeen detected infecting Jalapeñovarieties of chilli pepper in theneighbouring state of Quintana Roo(Díaz-Plaza et al., 1996). Meanwhile, anOld World begomovirus, Tomato yellowleaf curl virus (TYLCV) has made itsappearance in the Yucatán Peninsula(Ascencio-Ibáñez et al., 1999). This virushas caused yield losses in the

180
Whiteflies and Whitefly-borne Viruses in the Tropics
Caribbean estimated at millions ofUS dollars and thus poses an evidentthreat to agriculture in Mexico.
Advances in BiologicalResearch
The participation of Mexicanresearchers in Sub-project 2 of theTropical Whitefly Integrated PestManagement (TWF-IPM) Projectprovided an opportunity to gain anupdated and more systematic view ofthe status of whitefly-borne viruses inthe country. The Centro deInvestigaciones y Estudios Avanzados(CINVESTAV), located at Irapuato,Guanajuato, was selected as the leadinstitution in Mexico. This Centre
conducts research on begomovirusesaffecting horticultural crops in Mexico.Dr. Rafael Rivera-Bustamante conductedsurveys in collaboration with Dr. IrineoTorres-Pacheco, co-ordinator of thehorticulture program of the InstitutoNacional de Investigaciones Forestales yAgropecuarias (INIFAP) at that time.
In the course of this project, sampleswere taken from crop plants showingbegomovirus-like symptoms at a rangeof sites in major horticultural productionareas of Mexico (Table 1) and assayedusing monoclonal antibodies (Cancinoet al., 1995). In addition, a wide-rangingsurvey was conducted of viruses affectingtomato and chilli in Mexico (Table 2).The results obtained demonstrate thatbegomoviruses can attack the main
Table 1. Analyses of plants affected by begomovirus-like diseases in Mexico.
Location Plant (year) Begomovirusa Other viruses
MAB-BS MAB-GA
Los Mochis, Sinaloa Common bean (93) + - -
Los Mochis, Sinaloa Common bean (93) + - -
J. Rosas, Guanajuato Tomato + - -
Dzemul, Yucatán Squash + - -
Dzemul, Yucatán Chilli - - -
Dzemul, Yucatán Chilli - - -
Dzemul, Yucatán Chilli + - -
Los Mochis, Sinaloa Common bean + - -
Los Mochis, Sinaloa Common bean + - -
Los Mochis, Sinaloa Common bean + - -
Los Mochis, Sinaloa Squash + na -
Los Mochis, Sinaloa Squash + na -
Los Mochis, Sinaloa Squash + na -
Culiacán, Sinaloa Common bean + na -
Culiacán, Sinaloa Common bean + na -
Culiacán, Sinaloa Common bean + - -
Culiacán, Sinaloa Common bean + - -
Culiacán, Sinaloa Soybean + na -
Culiacán, Sinaloa Soybean + na -
Culiacán, Sinaloa Squash + na -
Culiacán, Sinaloa Squash - na -
Etchojoa, Son. Common bean + - -
a. MAB-BS, a broad spectrum monoclonal antibody used to detect bi-partite begomoviruses;MAB-GA, a monoclonal antibody used to detect the original Middle American isolates of Beangolden yellow mosaic virus-Guatemala; and na, not analysed.
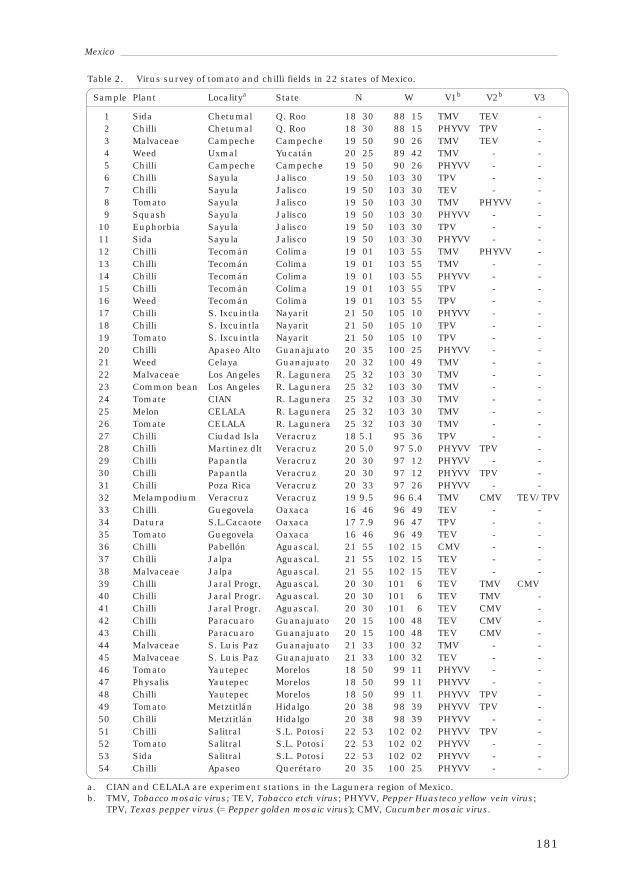
181
Mexico
Table 2. Virus survey of tomato and chilli fields in 22 states of Mexico.
Sample Plant Localitya State N W V1b V2b V3
1 Sida Chetumal Q. Roo 18 30 88 15 TMV TEV -2 Chilli Chetumal Q. Roo 18 30 88 15 PHYVV TPV -3 Malvaceae Campeche Campeche 19 50 90 26 TMV TEV -4 Weed Uxmal Yucatán 20 25 89 42 TMV - -5 Chilli Campeche Campeche 19 50 90 26 PHYVV - -6 Chilli Sayula Jalisco 19 50 103 30 TPV - -7 Chilli Sayula Jalisco 19 50 103 30 TEV - -8 Tomato Sayula Jalisco 19 50 103 30 TMV PHYVV -9 Squash Sayula Jalisco 19 50 103 30 PHYVV - -
10 Euphorbia Sayula Jalisco 19 50 103 30 TPV - -11 Sida Sayula Jalisco 19 50 103 30 PHYVV - -12 Chilli Tecomán Colima 19 01 103 55 TMV PHYVV -13 Chilli Tecomán Colima 19 01 103 55 TMV - -14 Chilli Tecomán Colima 19 01 103 55 PHYVV - -15 Chilli Tecomán Colima 19 01 103 55 TPV - -16 Weed Tecomán Colima 19 01 103 55 TPV - -17 Chilli S. Ixcuintla Nayarit 21 50 105 10 PHYVV - -18 Chilli S. Ixcuintla Nayarit 21 50 105 10 TPV - -19 Tomato S. Ixcuintla Nayarit 21 50 105 10 TPV - -20 Chilli Apaseo Alto Guanajuato 20 35 100 25 PHYVV - -21 Weed Celaya Guanajuato 20 32 100 49 TMV - -22 Malvaceae Los Angeles R. Lagunera 25 32 103 30 TMV - -23 Common bean Los Angeles R. Lagunera 25 32 103 30 TMV - -24 Tomate CIAN R. Lagunera 25 32 103 30 TMV - -25 Melon CELALA R. Lagunera 25 32 103 30 TMV - -26 Tomate CELALA R. Lagunera 25 32 103 30 TMV - -27 Chilli Ciudad Isla Veracruz 18 5.1 95 36 TPV - -28 Chilli Martinez dlt Veracruz 20 5.0 97 5.0 PHYVV TPV -29 Chilli Papantla Veracruz 20 30 97 12 PHYVV - -30 Chilli Papantla Veracruz 20 30 97 12 PHYVV TPV -31 Chilli Poza Rica Veracruz 20 33 97 26 PHYVV - -32 Melampodium Veracruz Veracruz 19 9.5 96 6.4 TMV CMV TEV/TPV33 Chilli Guegovela Oaxaca 16 46 96 49 TEV - -34 Datura S.L.Cacaote Oaxaca 17 7.9 96 47 TPV - -35 Tomato Guegovela Oaxaca 16 46 96 49 TEV - -36 Chilli Pabellón Aguascal. 21 55 102 15 CMV - -37 Chilli Jalpa Aguascal. 21 55 102 15 TEV - -38 Malvaceae Jalpa Aguascal. 21 55 102 15 TEV - -39 Chilli Jaral Progr. Aguascal. 20 30 101 6 TEV TMV CMV40 Chilli Jaral Progr. Aguascal. 20 30 101 6 TEV TMV -41 Chilli Jaral Progr. Aguascal. 20 30 101 6 TEV CMV -42 Chilli Paracuaro Guanajuato 20 15 100 48 TEV CMV -43 Chilli Paracuaro Guanajuato 20 15 100 48 TEV CMV -44 Malvaceae S. Luis Paz Guanajuato 21 33 100 32 TMV - -45 Malvaceae S. Luis Paz Guanajuato 21 33 100 32 TEV - -46 Tomato Yautepec Morelos 18 50 99 11 PHYVV - -47 Physalis Yautepec Morelos 18 50 99 11 PHYVV - -48 Chilli Yautepec Morelos 18 50 99 11 PHYVV TPV -49 Tomato Metztitlán Hidalgo 20 38 98 39 PHYVV TPV -50 Chilli Metztitlán Hidalgo 20 38 98 39 PHYVV - -51 Chilli Salitral S.L. Potosí 22 53 102 02 PHYVV TPV -52 Tomato Salitral S.L. Potosí 22 53 102 02 PHYVV - -53 Sida Salitral S.L. Potosí 22 53 102 02 PHYVV - -54 Chilli Apaseo Querétaro 20 35 100 25 PHYVV - -
a. CIAN and CELALA are experiment stations in the Lagunera region of Mexico.b. TMV, Tobacco mosaic virus; TEV, Tobacco etch virus; PHYVV, Pepper Huasteco yellow vein virus;
TPV, Texas pepper virus (= Pepper golden mosaic virus); CMV, Cucumber mosaic virus.

182
Whiteflies and Whitefly-borne Viruses in the Tropics
horticultural crops grown in thecountry: common bean, tomato, chilliand squash. Additionally, a begomoviruswas detected in soybean plants inSinaloa State. Soybean is a host forB. tabaci in South America but its rolein Mesoamerica as a reproductive hostfor this whitefly seems to have beensecondary to that of other crops suchas cotton and some horticultural crops.Nevertheless, B. tabaci reportedlydestroyed over 5000 ha of soybean inthe Valle del Fuerte, Sinaloa, in 1994(INIFAP, 1995).
As noted above, golden mosaicsymptoms in common bean in northernMexico are caused by BCaMV, which isrelated to SLCV, a cucurbit virusoriginally recorded from the UnitedStates. In this project, we surveyedthree key locations in the north-westernregion of Mexico—Etchojoa in Sonora,and Los Mochis and Culiacán in Sinaloa
state—and took samples of commonbean plants affected by symptoms of thedisease. A partial molecularcharacterization of the viral isolatescollected (Table 3) shows that BCaMV isstill present in common bean plantingsin north-western Mexico, specifically inLos Mochis. This isolate is still closelyrelated to an isolate of BCaMV collectedin the same locality in 1986. The samplefrom Culiacán represents a begomovirusclosely related to BCaMV but alreadyevolving into a new species (as indicatedby a sequence homology of less than90%). Perhaps of even greater interest isthat the common bean begomovirusfrom Sonora is still identifiable as anisolate of the original SLCV, constitutingthe first direct evidence of the evolutionof this virus.
Selected whitefly samples were alsocollected on different crops in north-western Mexico. Table 4 presents the
Table 3. Comparative sequence homologies (%) of common bean begomoviruses from north-westernMexico (Sonora and Sinaloa) and other whitefly-transmitted begomoviruses.
Virus isolate ORFa Common bean begomovirusesb
BGYMV-GA BCaMV SLCV BCaMV-MO BDMV
Etchojoa, Sonora AC1 57.0 77.2 96.0 76.8 61.0AV1 72.2 75.4 97.8 74.9 74.2
Los Mochis, Sinaloa AC1 69.8 89.7 70.5 88.3 68.8AV1 74.4 92.2 76.1 92.2 73.9
Culiacán, Sinaloa AC1 74.3 83.0 69.1 81.8 74.6AV1 73.0 86.1 79.2 85.7 74.2
a ORF, open reading frame; AC1, replicase; and AV1, capsid protein.b BGYMV-GA, Bean golden yellow mosaic virus-Guatemala; BCaMV, Bean calico mosaic virus;
SLCV, Squash leaf curl virus; MO, Mochis; BDMV, Bean dwarf mosaic virus.
Table 4. Identification of Bemisia tabaci biotypes from Sinaloa, northern Mexico.
Locality Crop Biotype A Biotype B
Los Mochis Common bean X -Los Mochis Common bean X -Los Mochis Common bean - XLos Mochis Common bean - XLos Mochis Common bean - X
Culiacán Eggplant X -Culiacán Eggplant X -Culiacán Eggplant X -Culiacán Eggplant X -Culiacán Eggplant X -

183
Mexico
results of the molecular biotyping ofthe whiteflies collected. As can beobserved, there is a mixed populationof biotypes A and B of B. tabaci.
Finally, the project supported workconducted by INIFAP researcher,Genovevo Ramírez-Jaramillo, based inMérida, Yucatán, on “the bio-ecology ofthe whitefly B. tabaci in the YucatánPeninsula.” This work—which alsoincluded the Government of the Stateof Yucatán, the Secretary ofAgriculture, Livestock and RuralDevelopment, the Regional ResearchCentre for the South-east and theFundación Produce de Yucatán—involved developing a geographicalinformation system (GIS)-based modelto predict the probability of whiteflyand begomovirus attack.
According to the classification ofToledo (1989), Mexico can be dividedinto five major ecological zones.Interestingly, the northernmost portionof the Yucatán Peninsula belongs tothe Dry Tropics (Trópico Seco), the sameecological zone as the northern PacificPlains, where the B. tabaci thrives. Inthis dry zone, the annual precipitationranges between 600 and 1500 mm andthe mean annual temperature is over20 °C. Although Yucatán is not one ofthe country’s main horticulturalregions, in 1992 this peninsulacultivated over 11,000 ha of Habaneroand Jalapeño chilli pepper, watermelonand tomato (listed in declining order ofproduction area). However, theagriculture of the peninsula includes adiversity of other crops, includingspecies that can act as reproductivehosts of B. tabaci such as soybean andcotton of which over 500 ha of each aregrown.
The GIS-based model for whiteflyand virus outbreaks drew on threecomponents of a database developed byINIFAP: (1) a digital elevation model,
(2) a climate database and (3) a soilsdatabase. For the biologicalparameters, a mean monthlytemperature of 23-27 °C and annualrainfall of less than 50 mm werechosen as defining areas with a highprobability of B. tabaci infestation.Whitefly population dynamics inYucatán were influenced to a largeextent by the amount of rainfall butonly minimally by the temperature,which remains within a range that isfavourable for whitefly development(Figure 1). Thus, the peak populationsof B. tabaci occurred in the months ofMay and June, following 4-5 months(December-April) of low precipitation(0-18 mm/month). Based on all ofthese parameters, risk assessmentmaps for the Yucatán Peninsula wereproduced. The assessments will allowrecommendations to be issuedregarding the most appropriateplanting dates for those crops mostlikely to be affected by B. tabaci, bothas a pest and vector of plant viruses.
Socio-economic Analysis
The results of the biological surveysconducted during the course of theproject show that whitefly-transmittedviruses and B. tabaci affect severalcrops of social and economicimportance throughout Mexico, withthe exception of the Central Highlands,where other whitefly speciespredominate.
In the Mexicali Valley, BajaCalifornia and the San Luis RíoColorado region of Sonora, B. tabaciattacked fields of cotton (damagingsome 14,000 ha), summer melon(causing the loss of 1500 ha) andsesame (Sesamum indicum L.)(damaging 7500 ha) in the 1991-92season, causing nearly US$10,000,000in production losses (López, 1996).Some 3000 ha of Serrano chilli and

184
Whiteflies and Whitefly-borne Viruses in the Tropics
tomato crops were lost between 1992and 1995 in the central region of SanLuis Potosí State, because of damageby B. tabaci and begomoviruses. Othercrops colonized by B. tabaci in thisregion were common bean, cotton,tomatillo (Physalis ixocarpa B.), squash,melon, watermelon, cucumber(Cucumis sativus L. var. sativus),sweetpotato (Ipomoea batatas [L.] Lam.)and sunflower (Helianthus annuus L.)(Hernández et al., 1996).
B. tabaci caused severe yield lossesto various horticultural crops,especially melon, in the ComarcaLagunera, Durango, in 1993 (Moraleset al., 1996). B. tabaci attacked melon,chilli and cotton in the state ofCoahuila, northern Mexico. Theinvestigation that was conducted at thetime showed that melon was anexcellent host, cotton was a good hostand chilli did not adequately supportthe reproduction of B. tabaci (Nava andRiley, 1996). Sonora has been one ofthe states in northern Mexico mostaffected by infestations of B. tabaci onvarious crops. An epidemiological studyin the Valley of the Yaqui indicated thatone of the main reproductive hosts ofthe whitefly in the valley was soybean(Pacheco, 1996). The incidence ofwhitefly-transmitted begomoviruses incommon bean plantings in the valley ofEl Fuerte, Sinaloa, caused yield lossesestimated at 20%-30% in the entireregion, between February and June1994 (Salinas et al., 1996). B. tabacihas been increasing its populations oncucumber in the valley of Culiacán,Sinaloa, since 1993, causing significantyield losses in over 9000 ha of this crop(Avilés and Valenzuela, 1996).
In the state of Nayarit, the fall-winter cropping cycle has been reducedby 50% because of the direct andindirect damage caused by B. tabaci asa pest and vector of viruses in tomato,
common bean, chilli and eggplant(Solanum melongena L.) (Ortíz et al.,1996). Viruses transmitted by B. tabacihave severely affected tomato plantingsin the state of Guerrero. In order toharvest tomato, farmers have resortedto treating their fields twice a weekwith insecticides (Barajas, 1996). Andin the state of Yucatán, B. tabaci andwhitefly-borne begomoviruses haveaffected horticultural crops such as hotpeppers, tomato, squash, melon andwatermelon since the 1980s (Díaz-Plazaet al., 1996).
The social and economic impact ofthese pest and disease outbreaks mustbe enormous. In the mid-1990s,horticultural and fruit crops occupiedover 1 million hectares in Mexico.About 7 million tons of produce, mainlytomato, melon and watermelon wereexported (principally to the UnitedStates, Canada, Japan and Brazil). Therest, some 35 million tons of squash,melon, pepper, tomato, potato,eggplant and chillis, was absorbed bythe national market for localconsumption, underlining theimportance of these crops to thedomestic economy. The production ofhorticultural crops demandsconsiderable human labour. Forinstance, tomato requires 271 work-days per cropping cycle as comparedwith only 24 for maize (Zea mays L.).Production costs for maize wereUS$430 per hectare compared withUS$3800 per hectare for tomato in1990. The marketing of horticulturalproducts is difficult because of theirperishable nature and price instability.In 1988, over 50% of the farmersproducing horticultural products inMexico had less than 5 ha of land(FIRA, 1997). Overall, the incidence ofwhitefly-transmitted viruses affects alarge portion of Mexico’s ruralpopulation, many of whom can beconsidered small-scale farmers.

185
Mexico
Strengthened ResearchCapacity
In addition to its research activities,CINVESTAV offers training to visitingscientists from developing countries.During the course of this project, twoCuban scientists conducted researchon Taino tomato mottle virus (TToMoV),a whitefly-transmitted begomovirusaffecting tomato in Cuba (Ramos et al.,1997). The project also contributedlimited funds for research on viruses ofeconomic importance in horticulturalcrops to the Horticulture Program ofINIFAP, and paved the way forcontinuing collaboration betweenINIFAP and the current TropicalWhitefly Project. (The state of Yucatánis the main target area for the currentPhase II of the project.)
Current Status ofWhitefly/Begomovirus
Problems
The whitefly/begomovirus situation inMexico is rather complex because ofthe size and diversity of the country—and because of the introduction oremergence of different whitefly-transmitted viruses in both the northand the south. In the north, theintensive cropping systems for non-traditional export crops share not onlya common border but also commonwhitefly/begomovirus problems withthe equally intensive agriculturalsystems of the south-western UnitedStates. In the south, commercial andsmall-scale subsistence agriculture co-exist as an extension of the croppingsystems found in Central America.Thus, it is not surprising to find someNorth American begomoviruses innorthern Mexico and some CentralAmerican begomoviruses in southernMexico.
Whereas the begomovirus situationon common bean has been partiallycontrolled through the use of resistantcultivars, the situation of otherhorticultural crops affected by whitefly-transmitted begomoviruses such astomato, pepper, chilli and melon is notyet resolved in most regions.Nevertheless, Mexico has madeconsiderable progress in implementingintegrated pest management (IPM)measures such as biological control,physical barriers (micro-tunnels) andother relevant cultural practices, and indeveloping the necessary knowledgebase, including, for instance, viruscharacterization. Mexico is also at theforefront in using GIS technology formore effective management ofagricultural resources.
More research is needed to studythe epidemiology of B. tabaci biotypesand other whitefly species inagricultural regions with altitudes above1000 m, including those in the states ofOaxaca, Guerrero, Morelos, Mexico, SanLuis Potosí, Guanajuato and Durango.Suitable IPM measures should berapidly implemented and evaluated toreduce dependence on chemicalinsecticides for whitefly control.
Acknowledgement
Project research and capacity-buildingactivities in Mexico were supported byfunds made available through theTWF-IPM Project by the Danish Agencyfor Development Assistance (DANIDA).
References
Ascencio-Ibáñez, J. T.; Díaz-Plaza, R.;Méndez-Lozano, J.; Monsalce-Fonnegra, J. I.; Arguello-Astorga, G.;Rivera-Bustamante, R. 1999. Firstreport of Tomato yellow leaf curl inYucatán, Mexico. Plant Dis. 83:1178.

186
Whiteflies and Whitefly-borne Viruses in the Tropics
Avilés, M.; Valenzuela, G. 1996.Respuesta de diferentes materialesde pepino al ataque de mosquitablanca (B. tabaci) en el Valle deCuliacán, Sin. In: Resumen, Tallerde Moscas Blancas, Acapulco, MX.p. 196.
Barajas, R. 1996. Intervalos de aplicaciónde insecticida contra la mosquitablanca Bemisia tabaci comparadocon acolchado plástico en jitomate.In: Resumen, Taller de MoscasBlancas, Acapulco, MX. p. 211.
Brown, J. K. 1989. A whitefly-transmittedgeminivirus from peppers with tigredisease. Plant Dis. 73:610.
Brown, J. K. 1994. Current status ofBemisia tabaci as a plant pest andvirus vector in agro-ecosystemsworldwide. FAO Plant Protect. Bull.42:3-32.
Brown, J. K.; Chapman, M. A.; Nelson, M.R. 1990. Bean calico mosaic, a newdisease of common bean caused bya whitefly-transmitted geminivirus.Plant Dis. 74:81.
Brown, J. K.; Idris, A. M.; Fletcher, D. C.1993. Sinaloa tomato leaf curl virus,a newly described geminivirus oftomato and pepper in west coastalMexico. Plant Dis. 77:1262.
Cancino, M.; Abouzid, A. M.; Morales, F.J.; Purcifull, D. E.; Polston, J. E.;Hiebert, E. 1995. Generation andcharacterization of threemonoclonal antibodies useful indetecting and distinguishing Beangolden mosaic virus isolates.Phytopathology 85:484-490.
Cárdenas, J. A.; Pérez, F.; Nieves, F.1996. Campaña contra la mosquitablanca en México. In: Resumen,Taller de Moscas Blancas,Acapulco, MX. p. 167-169.
Díaz-Plaza, R.; Avilés-Baeza, W.; Torres-Pacheco, I.; Rivera-Bustamante, R.1996. Geminivirus en la regiónhortícola de Yucatán, México. In:Resumen, Taller de MoscasBlancas, Acapulco, MX. p. 193.
FIRA (Fideicomisos Instituidos enRelación con la Agricultura). 1997.Rentabilidad de 15 cultivos anualesfinanciados en el ciclo agrícola PV/1996. Boletín Informativo N. 293.Banco de Mexico, MX. 34 p.
Flock, R. A.; Mayhew, D. E. 1981. Squashleaf curl, a new disease of cucurbitsin California. Plant Dis. 65:75-76.
Gallegos, H. M. L. 1978. Enchinamientodel tomate (chino disease oftomato). In: Enfermedades decultivos en el estado de Sinaloa.Secretaría de Agricultura yRecursos Hidráulicos (SARH), MX.p. 119.
Garrido, E. R.; Gilbertson, R. L. 1998.First report of tomato mottlegeminivirus infecting tomatoes inYucatán, Mexico. Plant Dis. 82:592.
Hernández, V.; Obregón, J.; Contreras, J.L. 1996. Identificación dehospederos de mosquita blancaBemisia tabaci en la zona mediapotosina. In: Resumen, Taller deMoscas Blancas, Acapulco, MX.p. 177.
Hou, Y. M.; Garrido, E. R.; Stingley, S.;Guzmán, P.; Gilbertson, R. L. 1996.Characterization of the infectiousclones of the Sinaloa strain ofpepper huasteco geminivirus.Phytopathology 86:S43.
INIFAP (Instituto Nacional deInvestigaciones Forestales Agrícolasy Pecuarias). 1995. Mosquitablanca en el noroeste de México.Memoria Científica no. 3. MX. 73 p.
López, G. H. 1974. Aumente susrendimientos en frijol en el Valle deCuliacán. Circular no. 12, Centrode Investigaciones Agrícolas delSureste (CIAS)-Instituto Nacional deInvestigacíon Agraria (INIA)-Secretaría de Agricultura yRecursos Hidráulicos (SARH), MX.

187
Mexico
López, R. 1996. Experiencia sobre elcontrol integral de la mosca blancaBemisia tabaci en el distrito dedesarrollo rural 002, Rio Colorado,Sonora. In: Resumen, Taller deMoscas Blancas, Acapulco, MX.p. 162.
López-Salinas, E. 1994. Situación delmosaico dorado del frijol en México-Sur. In: Morales, F. J. (ed.). Mosaicodorado del frijol: Avances deinvestigación. ProgramaCooperativo Regional de Frijol paraCentro América, México y el Caribe(PROFRIJOL)-Swiss Agency forDevelopment and Cooperation(SDC), Centro Internacional deAgricultura Tropical (CIAT), Cali,CO. p. 28-33.
Morales, Y.; Ramírez, M.; Pedroza, A.1996. Dinámica poblacional demosquita blanca Bemisia sp. en dospredios de melón con diferentesprácticas de control. In: Resumen,Taller de Moscas Blancas,Acapulco, MX. p. 178.
Nava, C. U.; Riley, D. G. 1996. Desarrollo,sobrevivencia y fecundidad deBemisia argentifolii (Homoptera:Aleyrodidae) a diferentestemperaturas y plantas hospederas.In: Resumen, Taller de MoscasBlancas, Acapulco, MX. p. 179.
Ortíz, M.; Arroyo, J. M.; Ramos, A.;Gonzáles, J. 1996. Especies demosquitas blancas, sus enemigosnaturales y hospederos en el estadode Nayarit. In: Resumen, Taller deMoscas Blancas, Acapulco, MX.p. 204.
Pacheco, J. J. 1996. Crecimientopoblacional de la mosquita blancade la hoja plateada (Bemisiaargentifolii) en el valle del Yaqui,Sonora. In: Resumen, Taller deMoscas Blancas, Acapulco, MX.p. 181.
Ramos, P. L.; Guerra, O.; Peral, R.;Oramas, P.; Guevara, R. G.; Rivera-Bustamante, R. 1997. Taino tomatomottle virus, a new bi-partitegeminivirus from Cuba. Plant Dis.81:1095.
Salinas, R.; Macías, C. J.; López, A. B.1996. Caracterización de virus enfrijol a través de diferenciales bajocondiciones de campo. In:Resumen, Taller de MoscasBlancas, Acapulco, MX. p. 192.
Sánchez, V.; Frías G; Aguirre, L.; Bravo,L.; Cotéz, L.; Quesada, J.; Ruíz, R;Hernández, M.; Rangel, U.; Garzón,J. 1996. Agro-ecology of a whitefly-transmitted virus in an arid andisolated Serrano pepper growingregion in Northeastern Mexico.Phytopathology 86:S117.
Stenger, D. C.; Duffus, J. E.; Villalon, B.1990. Biological and genomicproperties of a geminivirus isolatedfrom pepper. Phytopathology80:704-709.
Toledo, V. M. 1989. La producción ruralen México: Alternativas ecológicas.Ed. Fundación Universo 21, MX.76 p.

188
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 3.3
GuatemalaFrancisco Morales*, Abelardo Viana**,
Margarita Palmieri***, Mónica Orozco***and René Ruanoψ
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** Proyecto Regional de Frijol para CentroAmérica, México y el Caribe (PROFRIJOL),Guatemala City, Guatemala.
*** Universidad del Valle, Guatemala City,Guatemala.
ψ Instituto de Ciencia y Tecnología Agrícolas(ICTA), Guatemala City, Guatemala.
Introduction
Geographical context
Of the seven countries that comprisethe isthmus of Central America,Guatemala is physically and culturallythe most diverse. The highlands of thesouthern half of Guatemala descendsouthward to the Pacific Coast to formthe fertile “Piedmont” or “Coffee Belt”(500-1500 m above sea level) and thehot, grass- and forest-covered lowlandsknown as “The Coast” (0-500 m). Thenorthern regions of Guatemala aretropical lowlands covered by rain forestand scattered savannahs, which areonly sparsely populated (West andAugelli, 1977). However, it is thesouth-eastern region of the country(“El Oriente”), formed by low- and mid-altitude (200-1000 m) valleys, whichhas suffered the worst attacks from thewhitefly Bemisia tabaci (Gennadius)and different begomovirusestransmitted by this whitefly species.This region has drastically changed itstraditional agricultural environment tojoin the boom in export crops, thus
creating complex and fragile croppingsystems. Figure 1 shows the mainagricultural regions affected bywhitefly-transmitted begomoviruses.
SantaRosa
El Progreso
Escuintla
Zacapa
Jutiapa
Baja Verapaz
Jalapa
Guatemala
Figure 1. The main agricultural regionsaffected by whitefly-transmittedbegomoviruses, Guatemala.
The emergence of Bemisiatabaci as a pest and virusvector
The civil war in the United States in the1860s created an increased demand forcotton (Gossypium hirsutum L.). Soon,cotton produced in the Pacific lowlandsof the country comprised 20% ofGuatemala’s exports. This species wasprobably one of the first commerciallycultivated hosts of the whiteflyB. tabaci. A second boom of cottonproduction took place in the 1950s and

189
Guatemala
Sonora
1960s, and at this time the first virusestransmitted by whiteflies were observedin Guatemala and neighbouringcountries (Gill, 1994).
Tobacco (Nicotiana tabacum L.) isanother popular export crop thatcontinues to be grown in Guatemala.The cultivation of tobacco in south-eastern Guatemala has been blamedalso for increasing the population ofthe virus vector B. tabaci in the 1970s,thus provoking the first outbreaks ofBean golden yellow mosaic virus(BGYMV) in common bean (Phaseolusvulgaris L.) in that region of Guatemala(Morales, 1994).
In 1974, the HorticulturalResearch Division of the Instituto deCiencia y Tecnología Agrícolas (ICTA)initiated extension work in the ZacapaDepartment with a view to producingexport crops such as melon (Cucumismelo L.). At that time, the mainproduction problems identified wereoutdated agronomic practices,inadequate cultivars and the poorquality of the produce, which couldonly meet the relatively low standardsof the local markets. A similar industryto produce tomato (Lycopersiconesculentum Mill.) was also initiated inthe late 1970s in south-easternGuatemala (Gaytán, 1979). These cropsprovided better reproductive hosts forB. tabaci than either common bean orwild plant species. The populations ofthe vector soon soared and then newbegomoviruses emerged to attack thenew crops. The arrival of the B biotypeof B. tabaci, a more aggressive andpolyphagous variant of the originalspecies, further aggravated yield losses,with the new whitefly acting as both apest and a vector of damaging plantviruses (Dardón, 1992).
The first collaborative project thatset out to tackle the new and growingthreat of whitefly-borne viruses was the
Programa Cooperativo Regional deFrijol para Centro América, México y elCaribe (PROFRIJOL), initiated by theCentro Internacional de AgriculturaTropical (CIAT) to address the beangolden yellow mosaic problem. Thisproject, initiated in 1978 with supportfrom the United Nations DevelopmentProgramme (UNDP) and subsequentlyfunded (from 1980) by the SwissDevelopment Cooperation (SDC), iscredited with the development ofseveral BGYMV-resistant (DOR) linesthat have subsequently been widelyadopted as commercial varieties inMexico, Central America, theCaribbean and South America.
In Guatemala, the project beganwork under the auspices of ICTA in thesouth-eastern Monjas Valley, about960 m above sea level. Until the 1970s,this valley had been planted totraditional crops, mainly common beanand maize (Zea mays L.). In the 1980s,horticultural crops such as tomato andbroccoli (Brassica oleracea L. var. italicaPlenck) made their appearance in thevalley. Other non-traditional crops—pepper (Capsicum spp. L.), cucumber(Cucumis sativus L. var. sativus) andmore recently watermelon (Citrulluslanatus [Thunb.] Matsum. & Nakai) andgrape (Vitis vinifera L.)—have alsofound a niche in this valley over thelatter part of the 1990s. Tobacco is alarge-scale commercial crop that wasfirst planted in the valley during theearly 1970s; its production collapsed inthe late 1980s, only to make acomeback in recent years.
BGYMV, appearing in the mid-1970s, was the first whitefly-transmitted virus to cause severe yieldlosses in the Monjas Valley(Rodríguez, 1994). Tobacco wasprobably the most importantreproductive host of B. tabaci. The localcommon bean cultivars, Pecho Amarilloand Rabia de Gato, soon succumbed to

190
Whiteflies and Whitefly-borne Viruses in the Tropics
Table 1. Antigenic properties of common beanbegomoviruses from Guatemala.
Isolate Locality MAB-BSa MAB-GAb
1 Monjas + +
2 Tecpán + -
3 Cuyuta + -
4 Monjas + -
5 Monjas + -
6 San Jerónimo + -
7 Quiaté + -
a. MAB-BS, a broad spectrum monoclonalantibody used to detect bi-partitebegomoviruses.
b. MAB-GA, a monoclonal antibody used todetect the original Middle American isolatesof Bean golden yellow mosaic virus-Guatemala.
the disease. Fortunately, thePROFRIJOL project began to showresults and in due course released thefirst disease-resistant breeding lines:ICTA-Quetzal, ICTA-Jutiapán andICTA-Tamazulapa (Rodríguez, 1994).The causal agent, initially believed tobe Bean golden mosaic virus (BGMV),was characterized at the molecularlevel, thanks to collaborative researchfinanced by the United States Agencyfor International Development andcarried out at the University ofWisconsin and at CIAT. TheGuatemalan virus in the early 1990swas shown to be a distinct species,different from the Brazilian isolates ofBGMV (Faria et al., 1994) but with aconsiderable degree of homology andcausing a very similar disease; it wastherefore given the name Bean goldenyellow mosaic virus. Maize is not a hostof the whitefly B. tabaci and is notattacked by these particularbegomoviruses, although maize inAfrica is infected by differentgeminiviruses (Geminiviridae:Mastrevirus) transmitted byleafhoppers.
Advances in BiologicalResearch
Further research, undertaken incollaboration with the University ofFlorida, was intended to producemonoclonal antibodies (MABs) againstdifferent isolates of BGYMV, using theGuatemalan (MAB-GA) isolate ofBGYMV. At the same time, the projectsought to produce a broad-spectrummonoclonal antibody (MAB-BS),capable of detecting most whitefly-transmitted begomoviruses (Cancinoet al., 1995). The results shown inTable 1 indicate that these objectiveswere realized, in as much as MAB-BShas been able to detect all bi-partitebegomoviruses encountered to date inGuatemala, but MAB-GA reacts only
with the original Middle-AmericanBGYMV isolates. The result that MAB-GA, raised against the original isolatefrom Monjas, does not react with isolatessubsequently collected from this areasuggests that some change in theantigenic properties of the virus hasoccurred or that a new isolate has beenintroduced and become established inthe area. Begomoviruses are known toadapt to different host plants and vectorspecies. It has been suggested that thechanges observed in the antigenicreaction of the Monjas isolates may beassociated with the displacement of theA biotype of B. tabaci by the B biotype,which is believed to have occurred in the1990s.
Characterization of whitefliescollected from sites across Guatemalaconfirms that the B biotype has displacedthe A biotype almost completely (Table 2).This would be consistent with thehypothesis that the antigenic changes inthe virus detected with the specific MABmay be related to the presence of a newvector. To explore this possibility further,the coat protein of a recent isolate ofBGYMV from the locality of Monjas waspartially sequenced and compared withthe original isolate. Table 3 shows thecomparative results obtained.
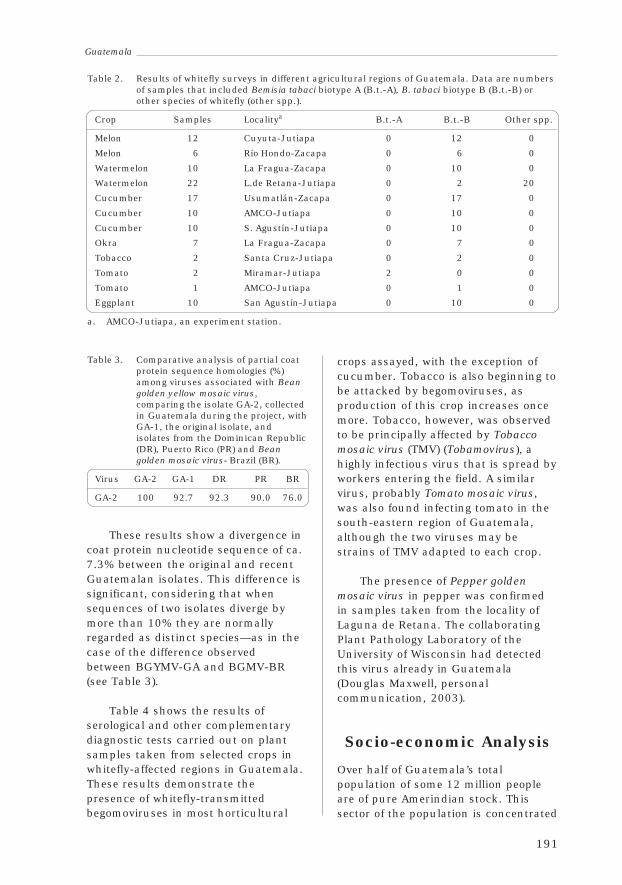
191
Guatemala
Table 2. Results of whitefly surveys in different agricultural regions of Guatemala. Data are numbersof samples that included Bemisia tabaci biotype A (B.t.-A), B. tabaci biotype B (B.t.-B) orother species of whitefly (other spp.).
Crop Samples Localitya B.t.-A B.t.-B Other spp.
Melon 12 Cuyuta-Jutiapa 0 12 0
Melon 6 Río Hondo-Zacapa 0 6 0
Watermelon 10 La Fragua-Zacapa 0 10 0
Watermelon 22 L.de Retana-Jutiapa 0 2 20
Cucumber 17 Usumatlán-Zacapa 0 17 0
Cucumber 10 AMCO-Jutiapa 0 10 0
Cucumber 10 S. Agustín-Jutiapa 0 10 0
Okra 7 La Fragua-Zacapa 0 7 0
Tobacco 2 Santa Cruz-Jutiapa 0 2 0
Tomato 2 Miramar-Jutiapa 2 0 0
Tomato 1 AMCO-Jutiapa 0 1 0
Eggplant 10 San Agustín-Jutiapa 0 10 0
a. AMCO-Jutiapa, an experiment station.
Table 3. Comparative analysis of partial coatprotein sequence homologies (%)among viruses associated with Beangolden yellow mosaic virus,comparing the isolate GA-2, collectedin Guatemala during the project, withGA-1, the original isolate, andisolates from the Dominican Republic(DR), Puerto Rico (PR) and Beangolden mosaic virus- Brazil (BR).
Virus GA-2 GA-1 DR PR BR
GA-2 100 92.7 92.3 90.0 76.0
These results show a divergence incoat protein nucleotide sequence of ca.7.3% between the original and recentGuatemalan isolates. This difference issignificant, considering that whensequences of two isolates diverge bymore than 10% they are normallyregarded as distinct species—as in thecase of the difference observedbetween BGYMV-GA and BGMV-BR(see Table 3).
Table 4 shows the results ofserological and other complementarydiagnostic tests carried out on plantsamples taken from selected crops inwhitefly-affected regions in Guatemala.These results demonstrate thepresence of whitefly-transmittedbegomoviruses in most horticultural
crops assayed, with the exception ofcucumber. Tobacco is also beginning tobe attacked by begomoviruses, asproduction of this crop increases oncemore. Tobacco, however, was observedto be principally affected by Tobaccomosaic virus (TMV) (Tobamovirus), ahighly infectious virus that is spread byworkers entering the field. A similarvirus, probably Tomato mosaic virus,was also found infecting tomato in thesouth-eastern region of Guatemala,although the two viruses may bestrains of TMV adapted to each crop.
The presence of Pepper goldenmosaic virus in pepper was confirmedin samples taken from the locality ofLaguna de Retana. The collaboratingPlant Pathology Laboratory of theUniversity of Wisconsin had detectedthis virus already in Guatemala(Douglas Maxwell, personalcommunication, 2003).
Socio-economic Analysis
Over half of Guatemala’s totalpopulation of some 12 million peopleare of pure Amerindian stock. Thissector of the population is concentrated

192
Whiteflies and Whitefly-borne Viruses in the Tropics
Table 4. Results of assays carried out on selected plant samples from Guatemala.
Sample Locality Plant Reactiona EMb
MAB-BS PTY1
1 Zacapa Melon + - nt
2 Zacapa Melon - + nt
3 Monjas Tobacco + nt nt
4 Monjas Tobacco + nt TMV
5 El Ovejero Tobacco - nt TMV
6 L. de Retana Tomato + nt nt
7 L. de Retana Tomato - - nt
8 L. de Retana Tomato + nt nt
9 Zacapa Tomato - - nt
10 Las Conchas Tomato - nt TMV
11 Las Conchas Tomato - nt TMV
12 Las Flores Tomato + nt nt
13 L. de Retana Tomato + nt nt
14 L. de Retana Tomato + nt nt
15 Plan de la Cruz Tomato + nt nt
16 Plan de la Cruz Tomato + nt nt
17 Monjas Tomato + nt nt
18 Monjas Tomato + nt nt
19 Monjas Tomato + nt nt
20 L. de Retana Pepper + nt nt
21 L. de Retana Pepper - nt -
22 L. de Retana Pepper + nt nt
23 L. de Retana Cucumber - nt -
24 L. de Retana Cucumber - nt -
a. Positive or negative reactions to a broad spectrum monoclonal antibody used to detect bi-partitebegomoviruses (MAB-BS); reactions to a potyvirus-specific monoclonal antibody (PTY1); and nt, notests for these samples.
b. Detection of Tobacco mosaic virus (TMV) by electron microscopy (EM); and nt, no tests for thesesamples.
in the highlands, north and west ofGuatemala City, as well as in theregion of Petén. South-easternGuatemala is the region with the lowestproportion of native Americans. Thepresence of mestizos or ladinos in theOriente of Guatemala explains the moredynamic and complex croppingsystems in these drier lowlands,including the prevalence of non-traditional export crops.
A special study was conducted inthe municipality of Monjas, JalapaDepartment, where Amerindians stillmake up over 70% of the totalpopulation. The Monjas Valley is
located at an altitude of 960 m and hasan average annual precipitation of900 mm, most of which falls betweenMay and October. The meantemperature is 23.7 °C and the meanrelative humidity is 80%. Ecologically,the Monjas Valley is classified both asdry subtropical forest and temperatesubtropical humid forest. Most of theagricultural land is private property,although it may have been acquired by“right” and not through a commercialtransaction.
Table 5 shows the existingcropping systems in the Monjas Valley.Maize and common bean are evidently

193
Guatemala
Table 5. Cropping systems in the MonjasValley, Jalapa, Guatemala.
Crop Years of Irrigation Rain-fedplanting
Common bean >50 - +
Maize >50 - +
Tobacco 25 + +
Tomato 15 + -
Broccoli 10 + -
Onion 10 - +
Pepper 7 + +
Cucumber 5 + -
Sweet corn 3 + -
Grape 3 + -
the most traditional crops in the valley.Tobacco has been planted for over20 years as a commercial crop butalmost disappeared in the late 1970sbecause of market problems, comingback into favour only in the late 1990s.Onion (Allium cepa L.) is another cropthat has been planted in the valley forover 10 years but it has never becomeprevalent. Broccoli has been a verypopular crop in the valley in the pastdecade and pepper has beencommercially cultivated during the past7 years. Cucumber has been plantedcommercially for the past 5 years andwatermelon for the past 2 years,although it has been planted for localconsumption for decades in the valley.Finally, sweet corn (Zea mays L. subsp.mays) and grape have been introducedin the past 3 years.
Table 5 also shows the associationbetween commercial crops and the useof irrigation in the Monjas Valley.Subsistence crops such as maize andcommon bean are planted during therainy season. The use of irrigationdemonstrates the intention of localgrowers to minimize risk in the case ofhigh-value crops.
Table 6 shows the perception thatgrowers have of the whitefly B. tabaci,both as a pest and virus vector in the
crops listed above, and the number ofpesticides used in each crop. Theresults clearly illustrate that pesticideabuse occurs in most of the cropsaffected by whiteflies. In the case oftomato, only five of the 11 pesticidesused are specific to the control ofwhitefly; the remaining six pesticidesare applied as “repellents” in the beliefthat the “smell” of these productsrepels the whitefly. The low use ofinsecticides on broccoli is interestingand responds to demand from foreignbuyers (many of whom only allow bio-pesticides to be used).
In general terms, whiteflies are themain pest on common bean, tomato,broccoli and tomato. Other pests suchas white grubs and lepidopterouslarvae also cause significant damage totobacco, tomato and sweet corn. Slugsand chrysomelids are perceived asimportant pests of common bean in thevalley. Diamond-back moth, Plutellaxylostella (Linnaeus), is also a pest ofbroccoli; and the corn ear worm,Helicoverpa zea (Boddie) causes severedamage to maize and sweet corn. Theexistence of insect pests other thanwhiteflies in the various crops grown inthe Monjas Valley hampers efforts toreduce the use of chemical pesticides
Table 6. Growers’ perception of whiteflyproblems and growers’ use ofpesticides in selected croppingsystems of the Monjas Valley, Jalapa.
Crop Whitefly Pesticides usedproblem (no.)
Common bean Yes 8
Maize No 1
Tobacco Yes 9
Tomato Yes 11
Broccoli Yes 1
Onion No -
Pepper Yes 5
Cucumber Yes 10
Sweet corn No 1
Grape No -

194
Whiteflies and Whitefly-borne Viruses in the Tropics
against whiteflies. In fact, most growersof non-traditional export crops use“cocktails”, or “bombs”, as pesticidemixtures are locally called.
Whereas growers recognizeB. tabaci as a pest, they do not fullyunderstand its role as a vector ofviruses. However, they readily associatethe bright yellowing shown by infectedcommon bean plants with a disease(bean golden yellow mosaic). In thecase of tomato, growers call symptomsexpressed by begomovirus-infectedplants acolochamiento (curling). In thecase of tobacco, growers recognize thesevere malformation and stuntingsymptoms shown by virus-infectedplants. However, they cannotdistinguish between whitefly-transmitted begomoviruses and TMV.
According to most growers,whiteflies are a regular pest, year afteryear, particularly during the summertime because of the highertemperatures and relative humidity.Growers also perceive the complexcropping systems in the valley as anegative factor because they claim thatthese crops represent more foodsources for the whitefly. Finally,growers blame ratoon crops for thesurvival of whiteflies between summerand winter seasons. Growers believe inthe existence of whitefly-resistant ormore generally pest- and disease-resistant crops, particularly tomato,tobacco and common bean. However,the “resistance” of tomato and tobaccoseems to be associated with theimplementation of more successfulchemical control practices in the valley.In reality, the case of common bean isa good example of genetically controlledvirus resistance obtained through awell-designed breeding programme.
A general question on the ability ofgrowers to predict seasons of high orlow whitefly incidence was generally
misinterpreted. Most growersresponded that they were always readyto control whiteflies in seedbeds, aftertransplanting and during the vegetativeand early reproductive phases of thecrop. In fact, some growers indirectlyanswered the question, associatingmild winters and an abundance ofalternative crops with higher whiteflypopulations and crop damage.
Strengthened ResearchCapacity
The Tropical Whitefly Integrated PestManagement (TWF-IPM) Projectfinanced two case studies conducted byICTA agronomists, in the departmentsof Jalapa, Jutiapa and Baja Verapaz.These departments used to betraditional agricultural areas, devotedto the planting of food staples such asmaize and common bean. The boom inexport crops, however, rapidlydisplaced traditional crops anddrastically changed the socio-economicstructure of the rural communitiesfound in these departments. Thestudies conducted in these regionsprovided information on the positiveaspects (potential higher income) andnegative consequences (new bioticproblems and pesticide abuse) of non-traditional export crops. The first studywas conducted in 15 ruralcommunities, located in threemunicipalities of the departments ofJalapa and Jutiapa. The second casestudy was conducted in five ruralcommunities in the municipalities ofSan Jerónimo, Rabinal and San MiguelChicaj. This study helped to developnon-chemical strategies for whiteflycontrol and generated information ofvalue to ICTA in its attempts to adoptsuitable research policies to assistsmall-scale farmers.
Although training in advancedlaboratory techniques was not

195
Guatemala
contemplated in this preliminary phaseof the project, it became apparent thata number of agricultural scientists inCentral America, Mexico and theCaribbean have adequate laboratoryfacilities to conduct such work. Thesescientists are becoming increasinglyreluctant to act simply as “guides” or“providers” of biological samples toforeign scientists from advancedresearch laboratories that often visitdeveloping countries in search ofinteresting scientific materials.Although the aim of this project was toconduct research in a trulycollaborative manner, in which nationalscientists have preferential access tothe results of all research undertaken,the possibility of training nationalscientists in advanced moleculartechniques became a need. The tworesearch areas in which advancedtechniques are being implemented inthis project are in the molecularidentification of whitefly biotypes andof the begomoviruses they transmit.
In Guatemala, the identification ofbegomoviruses is being done at theUniversity of San Carlos with thesupport of the Plant PathologyLaboratory of the University ofWisconsin. The molecularcharacterization of B. tabaci biotypes isbeing carried out at the University ofEl Valle, Guatemala City, which hasadequate facilities to conduct researchin the area of molecular biology but didnot have the technology to identifywhitefly biotypes. Consequently, aresearch assistant from the relevantlaboratory received 1 week of intensivetraining at CIAT on the molecularcharacterization of B. tabaci biotypesand was able to carry out this worksuccessfully upon return toGuatemala.
Thus, the project helped nationalscientists to conduct surveys thatimproved both the biological and socio-
economic knowledge base on whichresearch and implementation ofagricultural policy are founded. Theresearch sector of Guatemala was alsostrengthened through the training ofscientists from advanced researchlaboratories in the country intechniques used for the identification ofwhitefly biotypes and thebegomoviruses that affect food andexport crops of social and economicimportance.
Current Status ofWhitefly/Begomovirus
Problems
The whitefly/begomovirus problems inGuatemala are concentrated in thesouth-eastern agricultural regions ofthe country (Figure 1). This regioncontains most of the country’s mid-altitude valleys (at 500 to 1000 maltitude) with an average annualrainfall below 1500 mm, andtemperatures between 20 and 25 ºC;these valleys are also characterized byhighly diverse cropping systems,including irrigation. All of these factorsfavour the reproduction of B. tabaci.Moreover, the concentration of non-traditional export crops in this region isassociated with the intensive use ofpesticides, regularly used to protectthese high-value crops. Pesticide abuseis another predisposing factor for theestablishment of B. tabaci as a pest.Any crop planted in the Pacificlowlands will be severely attacked bywhitefly-transmitted begomovirusesparticularly because of the highertemperatures, which increase diseaseseverity.
The current begomovirus situationis becoming more complex as newwhitefly-transmitted viruses areintroduced from neighbouringcountries, or evolve from theindigenous virus population, or arise

196
Whiteflies and Whitefly-borne Viruses in the Tropics
through genetic recombination betweenthe existing begomoviruses that affectthe diverse crops such as commonbean, tomato, pepper, squash(Cucurbita spp. L.), melon and tobaccothat are grown in this area. Only in thecase of common bean has there been asustained effort on the part of thenational program and CIAT (with Swissfunding) to develop and release virus-resistant cultivars. For the remainingcrops, the emphasis has been onchemical control, although large agro-export companies have adopted someIPM packages. South-easternGuatemala provides an ideal site toconduct a case study with small-scalefarmers on the implementation ofpractical IPM measures to combat B.tabaci and the begomoviruses that thisspecies transmits to food andindustrial crops in this region.
Acknowledgements
The authors are grateful to thefollowing technical staff of ICTA whoassisted in the research: Carlos PaizGarcía, Edgardo Carrillo, Edin Palma,Iván Esquivel, Viany Gillespie, JoséLuis Ordóñez and T. Mairor R. Osorio.The work was supported by a grantfrom the Danish Agency forDevelopment Assistance (DANIDA), viathe TWF-IPM Project.
References
Cancino, M.; Abouzid, A. M.; Morales, F.J.; Purcifull, D. E.; Polston, J. E.;Hiebert, E. 1995. Generation andcharacterization of threemonoclonal antibodies useful indetecting and distinguishing Beangolden mosaic virus isolates.Phytopathology 85:484-490.
Dardón, D. E. 1992. Las moscas blancasen Guatemala. In: Hilje, L.; Arboleda,O. (eds.). Las moscas blancas enAmérica Central y el Caribe.Memorias del Taller Centroamericanoy del Caribe sobre moscas blancas,Turrialba, CR. p. 38-41.
Faria, J. C.; Gilbertson, R. L.; Hanson, S.F.; Morales, F. J.; Ahlquist, P.;Loniello, A. O.; Maxwell, D. P. 1994.Bean golden mosaic geminivirustype II isolates from the DominicanRepublic and Guatemala: Nucleotidesequences, infectiouspseudorecombinants, andphylogenetic relationships.Phytopathology 84:321-329.
Gaytán, M. A. 1979. Proyecto deexportación de melones en el Vallede Zacapa. XXV Reunión annualPrograma CooperativoCentroamericano para elMejoramiento de CultivosAlimenticios (PCCMCA), Tegucigalpa,HN.
Gill, A. 1994. Problemática del complejomosca blanca-virus en algodón enCentroamérica. In: Mata, M.; Dardón,D. E.; Salguero, V. E. (eds.). Biologíay manejo del complejo mosca blanca-virosis. Memorias III TallerCentroamericano y del Caribe sobremosca blanca, Antigua, GM.p. 23-38.
Morales, F. J. 1994. Bean golden mosaic:Research advances. CentroInternacional de Agricultura Tropical(CIAT), Cali, CO. 193 p.
Rodríguez, R. 1994. Situación actual delmosaico dorado del frijol enGuatemala. In: Morales, F. J. (ed.).Bean golden mosaic. 1994 researchadvances. Programa CooperativoRegional de Frijol para CentroAmérica, México y el Caribe(PROFRIJOL)-Swiss DevelopmentCooperation (SDC). CentroInternacional de Agricultura Tropical(CIAT), Cali, CO. p. 34-39.
West, R. C.; Augelli, J. P. 1977. MiddleAmerica: Its lands and peoples.Second edition. Prentice-Hall, NJ,USA. 494 p.

197
El Salvador
CHAPTER 3.4
El SalvadorFrancisco Morales*, Carlos Pérez**,
Priscila Henríquez**, Reina de Serrano**,Leopoldo Serrano*** and Claudio Nunesψ
Introduction
Geographical context
El Salvador lies entirely within avolcanic region but most of its territoryis formed by mid to low altitude valleys(up to 1000 m). About 40% of ElSalvador consists of agricultural land—the highest proportion in LatinAmerica, where it is the smallestcountry but the most denselypopulated. Given the shortage of land,the higher slopes are dedicated tocrops such as coffee (Coffea arabica L.).Farmers make up over 50% of thepopulation, primarily producingsubsistence crops such as maize (Zeamays L.) and common bean (Phaseolusvulgaris L.).
Cotton (Gossypium hirsutum L.),undoubtedly the second mostimportant commercial crop in ElSalvador in past decades (for export toJapan), is mostly found in the Pacificlowlands. Sugarcane (Saccharum
officinarum L.) became an importantcash crop at the turn of the centuryand a third of the sugar produced isexported to the United States. Rice(Oryza sativa L.) is another importantcrop as well as some cucurbits such aswatermelon (Citrullus lanatus [Thunb.]Matsum. & Nakai) (Pastor, 1988). Morerecently, the production of melon(Cucumis melo L.), pepper (Capsicumspp. L.) and tomato (Lycopersiconesculentum Mill.) has greatly increasedbecause of their relatively high marketvalue; even so, some of these productscontinue to be imported because thelocal market absorbs most of thecountry’s produce. Common bean iswidely grown in El Salvador, occupyingabout 75,000 ha (1995). However, totalcommon bean production does notsatisfy internal demand and, in 1994-95, El Salvador imported 14,600 tonsof common bean (Parada and Pérez,1996).
Because of the relatively small sizeof the country, there are fewecologically distinct areas other thanthose defined by altitude, where thewhitefly Bemisia tabaci (Gennadius)cannot thrive. Hence, the whitefly/begomovirus problem is distributedfairly evenly throughout the country.Figure 1 shows the main agriculturalregions affected by whitefly-transmittedbegomoviruses.
* Centro Internacional de AgriculturaTropical (CIAT), Cali, Colombia.
** Centro Nacional de Tecnificación Agrícola(CENTA), El Salvador.
*** Universidad del Salvador.ψ Universidad Técnica Latinoamericana,
El Salvador.

198
Whiteflies and Whitefly-borne Viruses in the Tropics
Sonora
The emergence of Bemisiatabaci as a pest and virusvector
Granillo and co-workers (1974) firstreported B. tabaci as a vector of plantviruses in El Salvador. The first virusand whitefly outbreaks occurred in thePacific coastal area, affecting cotton,kenaf (Hibiscus cannabinus L.) andcommon bean. As in the case ofGuatemala, we find here a closeassociation between cotton and thewhitefly B. tabaci. Among the bioticproblems that limit common beanproduction in the country, the mainpest and disease problems are B. tabaciand Bean golden yellow mosaic virus(BGYMV). Zaumeyer and Smith (1964)first reported this virus in El Salvador.BGYMV was more commonly observedin the Pacific lowlands, reaching up to100% incidence in most affectedcommon bean plantings in thedepartments of La Libertad, La Paz, SanVicente and Usulutan. BGYMV hasreached most of the adjacent mid-altitude valleys in the departments ofLa Libertad and Ahuchapan, and it canbe found now wherever common bean isgrown in the country. The introductionof new crops, particularly horticulturalcrops, further complicated the dynamicsof whitefly populations in El Salvador.The Zapotitán Valley (La Libertad) is agood example of a traditionalagricultural area where crops such as
maize and common bean were largelydisplaced by high-value horticulturalcrops (e.g., tomato, pepper andcucurbits). However, these vegetablecrops have been eliminated graduallyfrom the valley because of the damagecaused by whitefly-transmitted viruses.
Advances in BiologicalResearch
The first generation of black-seeded,BGYMV-resistant common beanvarieties produced in Guatemala in thelate 1970s made no impact on thecontrol of BGYMV in El Salvador,because Salvadorans prefer bright red-coloured beans. It was not until 1992,when the first red-seeded bean varietieswere developed and released, that beancould be cultivated again in these mid-altitude valleys. The BGYMV isolatesfrom El Salvador are very invasive insusceptible common bean cultivarssuch as Rojo de Seda, the preferredcommon bean cultivar in the country.Nevertheless, the BGYMV isolates fromEl Salvador were shown to be closelyrelated to the Middle American BGYMVisolates from Guatemala and PuertoRico (Table 1).
Cuscatlan
Sonsonate SanVicente
Santa Ana
LaLibertad
El Salvador
Ahuachapan
Figure 1. The main agricultural regionsaffected by whitefly-transmittedbegomoviruses, El Salvador.
Table 1. Serological reactions of selectedbegomoviruses from El Salvador.
BGYMVa Year MAB-BSb MAB-GAc
isolate collected
Guatemala 1992 + +
El Salvador 1992 + +
Puerto Rico 1992 + +
El Salvador 1998 + -
El Salvador 1999 + -
El Salvador 1999 + -
a. BGYMV, Bean golden yellow mosaic virus.b. MAB-BS, a broad spectrum monoclonal
antibody used to detect bi-partitebegomoviruses.
c. MAB-GA, a monoclonal antibody used todetect the original Middle American isolatesof BGYMV-Guatemala.

199
El Salvador
During the course of this project,individual virus samples were collectedfrom common bean plants showingtypical BGYMV symptoms. Table 1shows the results of the serologicaltests, with the broad spectrum andspecific (Guatemalan) monoclonalantibodies developed for BGYMV, forBGYMV isolates collected in 1992, 1998and 1999. The results show that, as of1998, there have been changes in thecoat protein composition of BGYMVisolates in El Salvador (no reaction withthe specific MAB-GA antiserum).
In order to determine the currentcomposition of whitefly species/biotypesin El Salvador, a collection of immaturewhiteflies found on economicallyimportant crops affected bybegomoviruses and/or B. tabaci wasundertaken in selected agriculturaldepartments of the country. The resultsobtained (Table 2) show an interestingsituation of mixed populations ofbiotype A and B of B. tabaci, biotype Bbeing predominantly associated withcucurbits.
The next step was to investigatethe proportion of whitefly-transmittedviruses in the different crops grown inwhitefly-affected agricultural regions.Table 3 shows the results from thisinvestigation. Basically, it is evidentthat whiteflies are not the only group ofvectors transmitting viruses tohorticultural crops in El Salvador.Many of the samples tested alsocontained potyviruses transmitted byaphids. It is interesting to note that thecucurbit locally called pipián (Cucurbitaargyrosperma C. Huber subsp.argyosperma) had not been recordedpreviously in Central America as a hostof whitefly-transmitted viruses. For thefour cucurbit species assayed in theabove survey, aphid-transmittedpotyviruses appear as the main viralproblem, with the exception of pipián.For common bean, BGYMV waspresent but the relatively low numberof samples collected probably reflectsthe displacement of this food legumefrom areas traditionally growingcommon bean. The results obtained fortomato and pepper suggest a complex
Table 2. Results of whitefly biotype surveys in different agricultural regions of El Salvador.
Cropa Sample Department B.t.-Ab B.t.-Bb Other
Tomato 1 Chalatenango 9 - 1
Chilli 2 Chalatenango 4 5 1
Watermelon 3 La Libertad 7 - 3
Loroco 4 La Libertad - 3 7
Chilli 5 La Libertad 7 - 3
Ayote 6 Santa Ana 6 - 4
Watermelon 7 La Paz 8 1 1
Melon 8 La Paz 2 7 1
Pipián 9 La Paz 4 - 6
Tomato 10 Chalatenango 5 - 5
Tomato 11 Chalatenango 18 - 2
Watermelon 12 Santa Ana 9 - 1
Tomato 13 Ahuachapán 8 - 2
Cucumber 14 Ahuachapán 4 3 3
Ayote 15 Ahuachapán 4 4 2
Watermelon 16 Santa Ana 2 6 2
a. Loroco (Fernaldia pandurata [A. DC.] Woodson), ayote (Cucurbita moschata Duchesne), pipián(Cucurbita argyrosperma C. Huber subsp. argyrosperma).
b. Bemisia tabaci A and B biotypes.

200
Whiteflies and Whitefly-borne Viruses in the Tropics
virus problem in these crops. In pasttests, one pepper sample and onetomato sample collected earlier in theZapotitán Valley had tested positive forthe presence of begomoviruses.
A BGYMV isolate from San Andrés,El Salvador, was selected for partialsequencing to determine the degree ofmolecular divergence with respect toBGYMV isolates characterized in theearly 1990s. Table 4 shows thecomparative results obtained with twopartial regions of the BGYMV genome:the viral replicase (AC1) and the coatprotein (AV1). As can be observed, theSalvadoran BGYMV-ES isolate has notchanged drastically over the pastdecade and it can still be considered anisolate of the Middle American BGYMVspecies (sequence homologies above90%). The differences observed at the
coat protein level (AV1) probably reflectminor changes in certain aminoacidsas a result of the introduction of a newwhitefly biotype. Differences at thereplicase level (AC1) may represent anevolutionary trend. The significantdifferences between the MiddleAmerican BGYMV species and theBrazilian BGMV species reflect theirdifferent origins or divergentevolutionary paths.
A visit to one of the mosttraditional agricultural areas of ElSalvador, the Zapotitán Valley, showedthe profound changes that haveoccurred in the cropping systems ofthis country. This irrigation district,thanks to its proximity to the capital,San Salvador, was the breadbasket ofEl Salvador, producing most of thebasic food crops demanded by the local
Table 3. Serological analyses of plants showing virus-like symptoms associated with the presence ofwhiteflies in El Salvador.
Samples Planta Begomovirus Potyvirus Other None
45 Pipián 25 11 0 9
9 Watermelon 0 9 1 -
11 Cucumber 0 7 0 4
6 Ayote 1 2 0 3
6 Common bean 2 0 0 4
38 Tomato 0 2 2 36
19 Pepper 0 4 0 0
1 Okra 0 0 0 0
1 Loroco 0 1 CMVb 0
1 Chilipuca (lima bean) 1 0 0 0
a. Pipián (Cucurbita argyrosperma C. Huber subsp. argyrosperma), ayote (Cucurbita moschataDuchesne), loroco (Fernaldia pandurata [A. DC.] Woodson), chilipuca (Phaseolus lunatus L.var. lunatus).
b. CMV, cucumovirus.
Table 4. Partial sequence homology (%) between a Bean golden yellow mosaic virus (BGYMV) isolatefrom El Salvador (ES) and related BGYMV/Bean golden mosaic virus (BGMV) isolatescharacterized in the early 1990s.
ORFa Isolatesb
BGYMV-DR BGYMV-PR BGYMV-GA BGMV-BR BGYMV-ES
AC1 90.5 88.8 91.7 69.4 100
AV1 94.3 91.5 93.8 78.2 100
a. ORF, open reading frame; AC1, viral replicase gene; and AV1, coat protein gene.b. DR, Dominican Republic; PR, Puerto Rico; GA, Guatemala; and BR, Brazil.
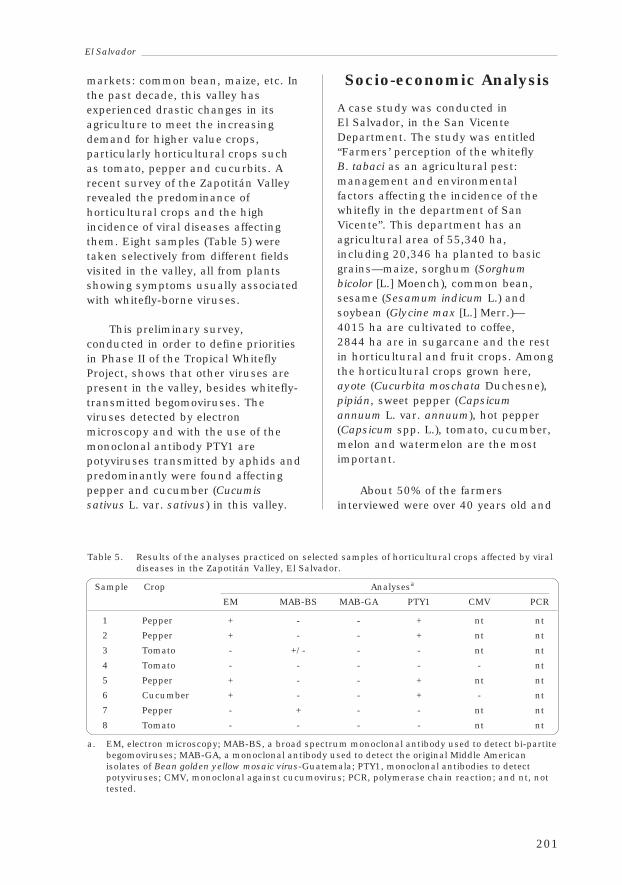
201
El Salvador
markets: common bean, maize, etc. Inthe past decade, this valley hasexperienced drastic changes in itsagriculture to meet the increasingdemand for higher value crops,particularly horticultural crops suchas tomato, pepper and cucurbits. Arecent survey of the Zapotitán Valleyrevealed the predominance ofhorticultural crops and the highincidence of viral diseases affectingthem. Eight samples (Table 5) weretaken selectively from different fieldsvisited in the valley, all from plantsshowing symptoms usually associatedwith whitefly-borne viruses.
This preliminary survey,conducted in order to define prioritiesin Phase II of the Tropical WhiteflyProject, shows that other viruses arepresent in the valley, besides whitefly-transmitted begomoviruses. Theviruses detected by electronmicroscopy and with the use of themonoclonal antibody PTY1 arepotyviruses transmitted by aphids andpredominantly were found affectingpepper and cucumber (Cucumissativus L. var. sativus) in this valley.
Socio-economic Analysis
A case study was conducted inEl Salvador, in the San VicenteDepartment. The study was entitled“Farmers’ perception of the whiteflyB. tabaci as an agricultural pest:management and environmentalfactors affecting the incidence of thewhitefly in the department of SanVicente”. This department has anagricultural area of 55,340 ha,including 20,346 ha planted to basicgrains—maize, sorghum (Sorghumbicolor [L.] Moench), common bean,sesame (Sesamum indicum L.) andsoybean (Glycine max [L.] Merr.)—4015 ha are cultivated to coffee,2844 ha are in sugarcane and the restin horticultural and fruit crops. Amongthe horticultural crops grown here,ayote (Cucurbita moschata Duchesne),pipián, sweet pepper (Capsicumannuum L. var. annuum), hot pepper(Capsicum spp. L.), tomato, cucumber,melon and watermelon are the mostimportant.
About 50% of the farmersinterviewed were over 40 years old and
Table 5. Results of the analyses practiced on selected samples of horticultural crops affected by viraldiseases in the Zapotitán Valley, El Salvador.
Sample Crop Analysesa
EM MAB-BS MAB-GA PTY1 CMV PCR
1 Pepper + - - + nt nt
2 Pepper + - - + nt nt
3 Tomato - +/- - - nt nt
4 Tomato - - - - - nt
5 Pepper + - - + nt nt
6 Cucumber + - - + - nt
7 Pepper - + - - nt nt
8 Tomato - - - - nt nt
a. EM, electron microscopy; MAB-BS, a broad spectrum monoclonal antibody used to detect bi-partitebegomoviruses; MAB-GA, a monoclonal antibody used to detect the original Middle Americanisolates of Bean golden yellow mosaic virus-Guatemala; PTY1, monoclonal antibodies to detectpotyviruses; CMV, monoclonal against cucumovirus; PCR, polymerase chain reaction; and nt, nottested.

202
Whiteflies and Whitefly-borne Viruses in the Tropics
40% were between 25-40 years of age.Only 25% of the farmers interviewedhad secondary (high school) educationand 41% had received technicalassistance in the selection ofpesticides to control the whiteflyproblem. Pesticide salesmen are highlyinfluential in the selection and use ofinsecticides for all crops. Most farmers(54%) apply insecticides with afrequency between 10-15 times percrop cycle. A significant proportion(16%) of the farmers interviewed applypesticides more than 20 times in asingle crop cycle. One hundred andforty-seven different commercialpesticides were mentioned during thissurvey. Sixty-two percent of thefarmers interviewed mentionedpreventive pesticide applications as arisk-averting practice.
Of the farmers interviewed, about97% recognizes the whitefly problembut only 36% associates this pest withviral diseases. Farmers believe thatwhitefly-related problems are moresevere during the dry months of theyear but they cannot predict theincidence of whiteflies and viraldiseases based on climatic factors.Over 60% of the respondent farmersconsidered that yield losses insusceptible crops range from25%-50% because of the whitefly/begomovirus problems.
Strengthened ResearchCapacity
El Salvador was undoubtedly thecountry where this project establishedmore collaborative activities withdifferent agricultural researchinstitutions. The local coordinatingentity was the national agriculturalresearch institution, CENTA, underthe co-ordination of Dr. PriscilaHenríquez (biotechnology) andIng. Carlos A. Pérez (Coordinator of
Programa Cooperativo Regional deFrijol para Centro América, México y elCaribe [PROFRIJOL] activities inEl Salvador). A major collaborator inthis project has been the University ofEl Salvador, under the guidance ofProf. Leopoldo Serrano. The Universityhas generated and collected aconsiderable amount of grey literatureon whitefly-related productionproblems in El Salvador and hasoffered students to conduct thesisresearch on whitefly-transmittedviruses affecting crops of socio-economic importance in the country.The University of El Salvador coveredthe San Vicente Valley, anotherimportant agricultural area. A secondinstitution of higher learningcollaborating in this project was theUniversidad Técnica Latinoamericana(UTLA), which also conducted somestudies in the Zapotitán Valley. ThePlant Health Division of the DirecciónGeneral de Salud Ambiental (DIGESA)also participated in this collaborativeeffort, covering the departments ofSanta Ana, Ahuachapán andSonsonate. Finally, anongovernmental organization, thePrograma Regional de ManejoIntegrado de Plagas en AméricaCentral (PROMIPAC), devoted to theproduction of non-traditional crops,also joined the project by providingsupport to the various activitiesundertaken by the different groups.The creation of a national network wasunique for Central America andrepresented a major local effort toaddress the important problem ofwhitefly-transmitted begomoviruses inEl Salvador.
All of these institutions wereprovided with technical guidelines forthe collection and processing ofbiological samples related to thewhitefly problem affecting crops ofsocio-economic importance.

203
El Salvador
Current Status ofWhitefly/Begomovirus
Problems
The whitefly/begomovirus problems inEl Salvador are widely dispersed in themain agricultural areas of the country.However, the incidence of these pestsis higher in the plains and mid-altitudevalleys. These agro-ecosystems range inaltitude, from sea level to 1000 m, withaverage temperatures of 27 ºC andabout 1600 mm of rainfall; all of thesebeing conditions that favour the biologyand reproduction of B. tabaci.
The displacement of traditionalagriculture (e.g., maize and commonbean) with non-traditional export crops(e.g., tomato, pepper and cucurbits)has aggravated the whitefly/begomovirus problem because of theavailability of new reproductive hostsfor B. tabaci and the intensive use ofpesticides to protect the new high-value crops against begomoviruses.
The current situation of thewhitefly/begomovirus problems in ElSalvador merits an effort to implementintegrated pest management strategiesin the main horticultural areas of thecountry, mainly in the departments ofSan Vicente, San Salvador, LaLibertad, Sonsonate and Santa Ana.
The use of virus-resistant commonbean cultivars has attenuated theproblem in this food crop but geneticresistance is not available in any of theother crops affected by begomovirusesin El Salvador.
References
Granillo, C. R.; Díaz, A.; Anaya, M. A.;Bermúdez, L. A. 1974.Enfermedades transmitidas porBemisia tabaci en El Salvador.Resumen de trabajo, seminario deleguminosas de grano, Río Piedras,PR. 2 p.
Parada, M. E.; Pérez, C. A. 1996.Situación actual del cultivo del frijolcomún en El Salvador con énfasisen bacteriosis común. InformeTécnico, Taller de bacteriosis,Mayagüez, PR. 12 p.
Pastor, R. 1988. Historia deCentroamérica. Editorial PiedraSanta, GT. 272 p.
Zaumeyer, W. J.; Smith, F. F. 1964.Report of a bean disease and insectsurvey in El Salvador. TechnicalReport, Agency for InternationalDevelopment (AID). TechnicalAssistance Agreement, AgriculturalResearch Service, United StatesDepartment of Agriculture (USDA),Beltsville, MD, USA.

204
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 3.5
HondurasFrancisco Morales*, María Mercedes Doyle**,
Rogerio Trabanino**, Carolina Nolasco**,Elsa Barrientos**, Luís Jara** and Norman Escoto***
Introduction
Geographical context
The territory of Honduras consists ofhighly mineralized soils and mountains,which favoured the development ofmining and livestock exploitations incolonial times. Unlike in other PacificCoast valleys of neighbouring countries,there are few volcanoes contributingtheir fertile ashes for the benefit of thesoils. The first region to be devoted toagriculture was the humid Caribbeanlowlands or “Costa”, which possessfertile alluvial soils planted primarily tobanana (Musa spp. L.). Thesegeographic characteristics leave only theinterior mid-altitude valleys(350-1000 m), for agriculturalproduction (West and Augelli, 1977).One of the most important valleys inthis mountainous region is theComayagua, with an area of about295 km2. Much of the mountainousinterior of Honduras supports one of themost extensive pine and oak forests inMiddle America. Figure 1 shows themain agricultural regions affected bywhitefly-transmitted begomoviruses.
The emergence of Bemisia tabacias a pest and virus vector
The isolated situation of the interiorvalleys and the absence of an extensivecotton (Gossypium hirsutum L.) industryin the Pacific coast of Hondurasprobably protected this country fromearly outbreaks of Bemisia tabaci(Gennadius) and begomovirusesrecorded in neighbouring countries tothe north, already in the 1970s. It wasnot until the mid-1980s that theintensification and diversification ofcropping systems in the highland centralvalleys coincided with the emergence ofbegomovirus problems in Honduras.Bean golden yellow mosaic virus(BGYMV) emerged around 1985. In1989, most common bean (Phaseolusvulgaris L.) producing regions inHonduras were already affected byBGYMV. The most affected areas werethose in the central and southernagricultural regions of the country,where annual rainfall does not surpassthe 1500 mm range, particularly fromMay through July (Rodríguez et al.,1994).
As in the rest of Middle America, inthe last decade Honduras hasexperienced a boom in the production ofnon-traditional crops such as tomato(Lycopersicon esculentum Mill.), pepper(Capsicum spp. L.) and melon (Cucumismelo L.). The tomato industry in theComayagua Valley (580 m) wasdeveloped as early as 1978, with a view
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** Escuela Agrícola Panamericana (EAP),Honduras.
*** Departamento de Investigación yCapacitación Agropecuaria (DICTA),Honduras.

205
Honduras
Sonora
to producing high-quality produce forexport (Standard Fruit Company, 1978).This industry has come practically to anend now because of the persistentattacks of whitefly-borne viruses. Asimilar phenomenon has been recordedfor the valleys of Jamastrán, El Paraísoand La Lima (El Paraíso Department),although the most affected crop in theseareas is pepper. The melon industry ofsouthern Honduras also has beenbrought to the point of extinction bywhitefly-related problems (Caballero,1995). These production problems haveforced many growers to abandon thecultivation of susceptible food cropsleaving as alternative crops such astobacco (Nicotiana tabacum L.). However,tobacco is currently under attack bywhiteflies and begomoviruses inneighbouring countries, particularly inGuatemala.
Advances in BiologicalResearch
Research on whitefly-transmittedbegomoviruses in Honduras has been
scant. During this project, commonbean plants exhibiting golden mosaic-like symptoms were tested with themonoclonal BGYMV antibodies. Inthese tests, the Honduran BGYMVisolate reacted with the broadspectrum monoclonal antibody (MAB-BS) but not with the Guatemalan(GA), as observed in this project forother BGYMV isolates fromGuatemala and El Salvador. In adifferent survey, eight samples takenin the Comayagua Valley fromdiseased common bean, tomato,pepper, cucumber (Cucumis sativus L.var. sativus) and a new vegetablegrown for export, cundeamor(Momordica charantia L.), wereassayed serologically. Table 1 showsthe results obtained with thesesamples.
Begomoviruses were detected incommon bean and tomato but not inthe two cucurbits tested, namelycucumber and cundeamor. The latterseemed to have a low incidence of aphytoplasma disease, and thecucumber sample was infected by a
Comayagua
Copan
El Paraíso
Yoro
Olancho
Honduras
Ocotepeque
Figure 1. The main agricultural regions affected by whitefly-transmitted begomoviruses, Honduras.
Islas de la Bahía
AtlántidaCortés
SantaBárbara
Colón
Gracias a Dios
Lempira
Intibuca
La Paz
FranciscoMorazán
ValleCholuteca
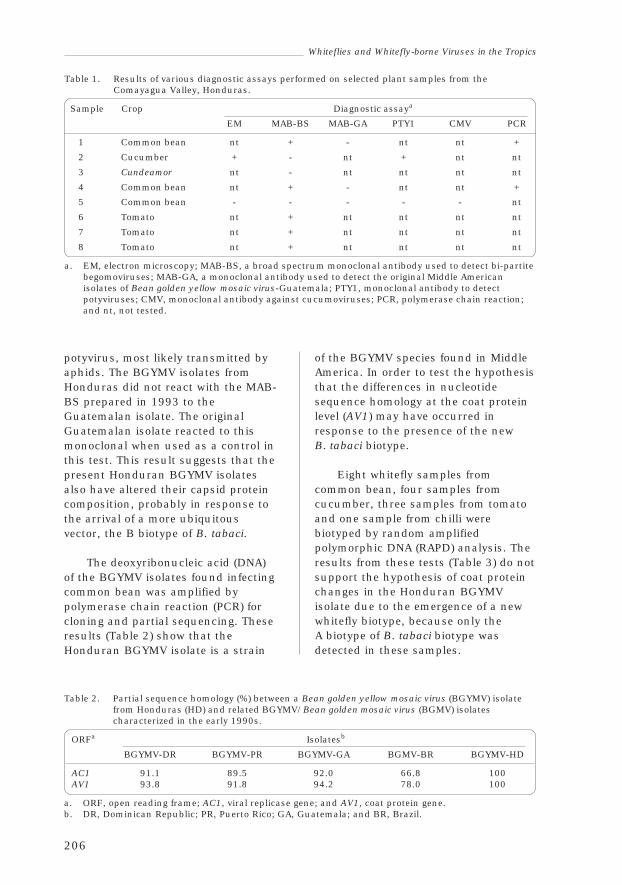
206
Whiteflies and Whitefly-borne Viruses in the Tropics
potyvirus, most likely transmitted byaphids. The BGYMV isolates fromHonduras did not react with the MAB-BS prepared in 1993 to theGuatemalan isolate. The originalGuatemalan isolate reacted to thismonoclonal when used as a control inthis test. This result suggests that thepresent Honduran BGYMV isolatesalso have altered their capsid proteincomposition, probably in response tothe arrival of a more ubiquitousvector, the B biotype of B. tabaci.
The deoxyribonucleic acid (DNA)of the BGYMV isolates found infectingcommon bean was amplified bypolymerase chain reaction (PCR) forcloning and partial sequencing. Theseresults (Table 2) show that theHonduran BGYMV isolate is a strain
of the BGYMV species found in MiddleAmerica. In order to test the hypothesisthat the differences in nucleotidesequence homology at the coat proteinlevel (AV1) may have occurred inresponse to the presence of the newB. tabaci biotype.
Eight whitefly samples fromcommon bean, four samples fromcucumber, three samples from tomatoand one sample from chilli werebiotyped by random amplifiedpolymorphic DNA (RAPD) analysis. Theresults from these tests (Table 3) do notsupport the hypothesis of coat proteinchanges in the Honduran BGYMVisolate due to the emergence of a newwhitefly biotype, because only theA biotype of B. tabaci biotype wasdetected in these samples.
Table 1. Results of various diagnostic assays performed on selected plant samples from theComayagua Valley, Honduras.
Sample Crop Diagnostic assaya
EM MAB-BS MAB-GA PTY1 CMV PCR
1 Common bean nt + - nt nt +
2 Cucumber + - nt + nt nt
3 Cundeamor nt - nt nt nt nt
4 Common bean nt + - nt nt +
5 Common bean - - - - - nt
6 Tomato nt + nt nt nt nt
7 Tomato nt + nt nt nt nt
8 Tomato nt + nt nt nt nt
a. EM, electron microscopy; MAB-BS, a broad spectrum monoclonal antibody used to detect bi-partitebegomoviruses; MAB-GA, a monoclonal antibody used to detect the original Middle Americanisolates of Bean golden yellow mosaic virus-Guatemala; PTY1, monoclonal antibody to detectpotyviruses; CMV, monoclonal antibody against cucumoviruses; PCR, polymerase chain reaction;and nt, not tested.
Table 2. Partial sequence homology (%) between a Bean golden yellow mosaic virus (BGYMV) isolatefrom Honduras (HD) and related BGYMV/Bean golden mosaic virus (BGMV) isolatescharacterized in the early 1990s.
ORFa Isolatesb
BGYMV-DR BGYMV-PR BGYMV-GA BGMV-BR BGYMV-HD
AC1 91.1 89.5 92.0 66.8 100AV1 93.8 91.8 94.2 78.0 100
a. ORF, open reading frame; AC1, viral replicase gene; and AV1, coat protein gene.b. DR, Dominican Republic; PR, Puerto Rico; GA, Guatemala; and BR, Brazil.

207
Honduras
Socio-economic Analysis
A case study was conducted in theComayagua Valley with thecollaboration of the Escuela AgrícolaPanamericana (EAP), Zamorano. Theresults of this study are published in athesis (Jara, 1998). In the ComayaguaValley, more than 70% of the100 producers surveyed had beencultivating the land longer than5 years. The number of literate farmersin the population surveyed was 75%, ofwhich 66% had received only primaryeducation. Only half or less (in someareas) of the farmers surveyed hadreceived technical assistance to controlwhitefly problems.
Regarding whitefly incidence, 87%of the farmers believed that whitefliesand whitefly-borne viruses increased inthe warmer months of the year. About80% of the farmers noted that whiteflypopulations increase in periods of lowrainfall. The worst whitefly/virusepidemics in the Comayagua Valleyoccurred in 1989, when drought andhigh temperatures struck the region.
These results demonstrate that mostfarmers are aware of the key climaticfactors that determine whiteflyepidemics.
Over 40% of the farmers interviewedapply insecticides on a calendar basisfor whitefly control. This practice canbe interpreted as a risk reductionmeasure, particularly in the case oftomato growers, who invest betweenUS$2100 and $3500 per manzana(0.764 ha). In tomato plantings, 63% ofthe growers apply insecticides againstwhiteflies as soon as the tomato plantsare transplanted, 31% apply 1 week aftertransplanting and only 6% wait untilthey see whiteflies in their fields. In thecase of common bean, 12% of thefarmers apply pesticides at plantingtime, 42% start controlling whiteflies1 week after germination of the plantsand 46% apply insecticides later onduring the vegetative phase of the crop.These results demonstrate farmers’perception regarding the investmentrequired to plant tomato (usually10 times higher than in the case ofcommon bean).
Table 3. Biotyping of whitefly (Bemisia tabaci) samples collected from different commercial cropsgrown in different municipalities of two departments in Honduras.
Department Municipality Crop Biotype
Comayagua V. San Antonio Common bean A
Comayagua V. San Antonio Common bean A
Comayagua V. San Antonio Common bean A
Comayagua V. San Antonio Common bean A
Comayagua V. San Antonio Common bean A
Comayagua Comayagua Common bean A
Comayagua Comayagua Cucumber A
Comayagua Comayagua Cucumber A
Comayagua San Nicolás Tomato A
Comayagua San Nicolás Tomato A
Comayagua Flores Tomato A
Comayagua V. San Antonio Common bean A
Comayagua Flores Common bean A
Comayagua Comayagua Cucumber A
Comayagua Comayagua Cucumber A
Francisco Morazán Cedros Chilli A

208
Whiteflies and Whitefly-borne Viruses in the Tropics
Regarding the likelihood ofintroducing non-chemical controlmeasures based on cultural practicessuch as changing planting dates toavoid whitefly population peaks, about65% of the growers were reluctant tochange. This finding suggests thatclimatic and market factors determineplanting times in Honduras.
In economic terms, 53% of theproduction costs for tomato and 47%for common bean are related to thechemical control of whitefly/begomoviruses. In cucumberproduction, only 16% of the total costsare related to chemical whitefly control.Total production costs for tomato areup to 20 times more than for commonbean or cucumber. It is evident fromthis study that the implementation ofan integrated whitefly managementpackage in the Comayagua Valleywould significantly contribute tolessening the environmental impact ofinsecticides and to a significantreduction in production costs of high-value crops such as tomato andpeppers.
Strengthened ResearchCapacity
The Tropical Whitefly Integrated PestManagement Project made possible thedevelopment of a thesis entitled“Characterization of the incidence andmanagement of whiteflies(Homoptera:Aleyrodidae) in the valley ofComayagua, Honduras” at the PlantProtection Department, EAP-Zamorano.The case study helped acquire a betterview of how small-scale farmersperceive the whitefly problem. Expertsin various fields such as agronomy,entomology, virology, socio-economicsand biometry developed thequestionnaire, which consequentlyconstitutes a guide for future surveyson similar field production problems.
The Plant Protection Department ofthe EAP sent Elsa Barrientos to theCentro Internacional de AgriculturaTropical (CIAT), Colombia, for trainingin molecular biotyping of B. tabaci andcharacterization of plant viruses duringa 2-week period.
Current Status ofWhitefly/Begomovirus
Problems
Although the whitefly/begomovirusproblems arrived relatively late inHonduras, most of the territory offerssuitable environmental conditions forB. tabaci to reproduce and attacksusceptible crops, either as a pest or asa virus vector. The exceptions are thenorthern Caribbean coast, whichusually receives more than 2000 mm ofrainfall, a deleterious climatic factor forB. tabaci. This region is the primebanana-producing zone of Honduras.The rest of the country has an averagerainfall of less than 1500 mm andaverage temperature of 26 ºC, andmost horticultural regions are locatedbetween 200 and 1000 m above sealevel—all favourable conditions forB. tabaci.
A good indicator of the presence ofwhitefly-transmitted viruses in a regionis the occurrence of BGYMV. This virusis distributed in the departments ofEl Paraíso, Francisco Morazán,Choluteca, Valle, Olancho, Comayaguaand Copán, marking a vast regionwhere whitefly/begomovirus problemscan emerge on susceptible crops.Undoubtedly, one of the most affectedareas is the Comayagua Valley, northof the capital, Tegucigalpa. This valleyis located in a drier zone (500-1000mm) and has been planted intensivelyto non-traditional export crops underconsiderable pesticide abuse.

209
Honduras
References
Caballero, R. 1995. Honduras. Reporte,IV Taller Latinoamericano sobremoscas blancas y geminivirus.Ceiba 36:21-23.
Jara, L. A. 1998. Caracterización de laincidencia y manejo de moscablanca (Homoptera:Aleyrodidae) enel Valle de Comayagua, Honduras.Thesis, Departamento de ProtecciónVegetal, Escuela AgrícolaPanamericana (EAP), El Zamorano,HN. 84 p.
Rodríguez, F.; Díaz, O.; Escoto, N. D.1994. Situación del mosaico doradoen Honduras. In: Morales, F. J.(ed.). Mosaico dorado del frijol:Avances de investigación 1994.Programa Cooperativo Regional deFrijol para Centro América, Méxicoy el Caribe (PROFRIJOL)-SwissAgency for Development andCooperation (SDC). CentroInternacional de AgriculturaTropical (CIAT), Cali, CO. p. 45-50.
Standard Fruit Company. 1978. Reportefinal de los cultivos semi-comerciales de tomate y pepinopara exportación en los valles deComayagua y la Entrada, Copán.La Ceiba, HN.
West, R. C.; Augelli, J. P. 1977. MiddleAmerica: Its lands and peoples.Second edition, Prentice-Hall, NJ,USA. 494 p.

210
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 3.6
NicaraguaFrancisco Morales*, Alberto Sediles**,
Aurelio Llano*** and Falguni Guharayψ
Introduction
Geographical context
Nicaragua is the largest country inCentral America and one of its richestnations in terms of agriculturalresources. Its agricultural economyfollows the natural divisions of thecountry: the Pacific region, thenorthern part of the Central Highlandsand the lowlands of the Caribbean. ThePacific region has been Nicaragua’scentre of commercial agriculture sincecolonial times, when livestock and theproduction of indigo (blue dye) weretwo of the main commodities. Livestockproduction still remains an importantactivity in this region. Cotton(Gossypium hirsutum L.) became ahighly important crop in the Pacificlowlands, between Lake Managua andthe Gulf of Fonseca (Chinandega toLeón) and along the eastern side ofLake Managua (Tipitapa) in the early1950s. By 1977, Nicaragua was thelargest (217,000 ha) producer of cottonin Central America (Gill, 1994).
As in the rest of Central America,there are a number of fertile mid-altitude valleys in Nicaragua such as
Boaco and Sébaco where a moreintensive agriculture eventuallydeveloped, consisting of non-traditionalcrops such as tomato (Lycopersiconesculentum Mill.), pepper (Capsicumspp. L.) and other horticultural crops.Common bean (Phaseolus vulgaris L.)remains a major staple andconsequently is grown throughout thecountry. However, the main regionsproducing common bean are the Pacificregion (30% of total production), theCentral Highlands (50%) and theCaribbean Plains (10%). In thehighlands, the main departmentsproducing common bean are Matagalpa,Jinotega, Estelí, Madriz and NuevaSegovia. Figure 1 shows the mainagricultural regions affected by whitefly-transmitted begomoviruses.
The emergence of Bemisiatabaci as a pest and virusvector
Bemisia tabaci (Gennadius) first becamea pest of cotton in the 1970s, whichtogether with adverse marketingcircumstances reduced the area plantedto cotton to a mere 2520 ha in 1993.Interestingly, B. tabaci was not aninsect of economic significance in theearly years of cotton production inNicaragua. The emergence of thiswhitefly species as a major pest ofcotton followed the introduction andintensive use of pesticides on this cropin the 1960s (Gill, 1994).
* Centro Internacional de AgriculturaTropical (CIAT), Cali, Colombia.
** Universidad Nacional Agraria (UNA),Managua, Nicaragua.
*** Centro Nacional de InvestigaciónAgropecuaria (CNIA), Nicaragua.
ψ Centro Agronómico Tropical deInvestigación y Enseñanza (CATIE)-ProyectoManejo Integrado de Plagas (MIP), ElZamorano, Nicaragua.

211
Nicaragua
Bean golden yellow mosaic virus(BGYMV) was first observed affectingcommon bean in the Pacific Coast ofNicaragua, around 1971. BGYMV isparticularly severe in Estelí, NuevaSegovia, León, Chinandega and thelowlands of Matagalpa (Llano et al.,1997). Large populations of B. tabacihave been reported on cotton in thePacific Coast of Nicaragua since 1952,and from tomato in the Atlantic Coastas early as 1953 (Hidalgo et al., 1975).This whitefly species was the third mostimportant pest problem in cotton inNicaragua in the late 1970s (Kramer,1966). The whitefly problem became soserious that, in 1975, Nicaraguacreated a special commission to studyB. tabaci and make recommendationson research and whitefly management(Hildago et al., 1975).
Mid-altitude valleys such as Sébaco(Matagalpa Department) witnessed theemergence of whitefly-transmittedviruses in 1986. By 1992, yield lossesreached 100% in several tomato fieldsaffected by what growers referred to as“crespo” (leaf curl). In 1991, largepopulations of whiteflies attackedcommon bean and melon (Cucumis melo
L.) but only bean was affected bybegomoviruses. Whitefly-transmittedbegomoviruses also attacked pepper andtomato plantings in the CentralHighlands. Tomato plantings wereaffected in the region of Boaco in 1991.The same year, B. tabaci attackedtobacco (Nicotiana tabacum L.) plantingsin the departments of Nueva Segovia,Jinotega, Estelí, Chinandega, Masayaand Rivas. The melon industry in thedepartments of León, Rivas, Managuaand Matagalpa was first affected bywhiteflies in 1991 but the incidence ofviruses was low (Sediles, 1998).
Advances in BiologicalResearch
No records exist of serologicalidentification of BGYMV in Nicaraguaprior to 1995. In 1993, a few commonbean and tomato samples were collectednear the town of Santa Lucía in theBoaco Valley. The common beansamples were taken from plantsshowing dwarfing symptoms, ratherthan mosaic. These samples reactedpositively with the broad spectrummonoclonal antibody that detectswhitefly-transmitted begomoviruses ingeneral (MAB-BS). The virus was lateridentified as Bean dwarf mosaic virus(BDMV) by nucleic acid hybridizationmethods (Zamora 1996). At that time,and from the same locality, samplescollected from tomato plants affected bycrespo disease also gave positive resultsin serological assays with the samemonoclonal antibody. The crespodisease of tomato was first observed inNicaragua in 1986. An in-depthinvestigation conducted in2000 showed that tomato in Nicaraguais infected by at least four distinctbegomoviruses (Rojas et al., 2000).
In 1995, three BGYMV-affectedcommon bean samples from the north-western department of Estelí were tested
Madriz
Chinandega
Nicaragua
Estelí
Nueva Segovia
Boaco
Carazo
Figure 1. The main agricultural regionsaffected by whitefly-transmittedbegomoviruses, Nicaragua.
León

212
Whiteflies and Whitefly-borne Viruses in the Tropics
with the monoclonal antibody used todetect the original Middle Americanisolates of BGYMV-Guatemala (MAB-GA), available at the CentroInternacional de Agricultura Tropical(CIAT). The three common beansamples reacted with the MAB-BS butonly one common bean sample wasrecognized by the specific MAB-GA.
A survey in the departments ofEstelí (Condega), Matagalpa (Apompuaand Sébaco) and Managua (Managua
and Pochocoape) was undertaken tocollect plant samples for virus assay.Table 1 gives the results of this survey,which show a rather low incidence ofwhitefly-transmitted geminiviruses (GV)in the samples tested. However, thesampling was done at the peak of therainy season, which, for the secondtime since Hurricane Mitch in theprevious year, brought considerableamounts of rain. Rainfall in 1999almost doubled the amount of rainrecorded in 1998 in the Pacific
Table 1. Results of survey of virus-affected plants in three departments of Nicaragua.
Sample Crop Locality Department Reactiona
MAB-BS PTY1
1-N5 Tomato Condega Estelí - -2-N5 Tomato Condega + / - -3-N5 Tomato Condega - -4-N5 Tomato Condega + -5-N5 Tomato Condega - -6-N5 Tomato Condega - -7-N6 Tomato Apompua Matagalpa - -8-N6 Tomato Apompua - -9-N6 Tomato Apompua - -
10-N6 Tomato Apompua - -11-N6 Tomato Apompua - -12-N6 Tomato Apompua - -13-N7 Tomato Managua Managua - -14-N7 Tomato Managua - -15-N7 Tomato Managua - -16-N7 Tomato Managua - -17-N8 Tomato Sébaco Matagalpa - -18-N8 Tomato Sébaco - -19-N9 Tomato Pochocoape Managua - -20-N9 Tomato Pochocoape - -21-N9 Tomato Pochocoape - -22-N9 Tomato Pochocoape - -23-N9 Tomato Pochocoape - -24-N9 Tomato Pochocoape - -
25-N10 Pipián Apompua Matagalpa + +26-N10 Pipián Apompua + +27-N10 Pipián Apompua + +28-N11 Tomato Sébaco Matagalpa - -29-N11 Tomato Sébaco - -30-N11 Tomato Sébaco - -31-N11 Tomato Sébaco - -32-N11 Tomato Sébaco - -33-N11 Tomato Sébaco - -34-N11 Tomato Sébaco - -35-N12 Tomato Apompua Matagalpa - -36-N12 Tomato Apompua - -
a. Positive or negative reactions to a broad spectrum monoclonal antibody used to detect bi-partitebegomoviruses (MAB-BS); reactions to a potyvirus-specific monoclonal antibody (PTY1).

213
Nicaragua
Lowlands of Nicaragua. The presence ofbegomoviruses in pipián (Cucurbitaargyrosperma C. Huber subsp.argyrosperma) is interesting,considering that this crop had not beenrecorded previously as an importanthost/reservoir of begomoviruses.However, the same observation wasmade in El Salvador and, furthermore,this cucurbit was also doubly infectedwith aphid-transmitted potyviruses inboth countries.
Because of the rainy conditionsthat affected Nicaragua in 1998 and1999, very few whitefly samples couldbe taken for analyses. Table 2summarizes some of the preliminaryresults obtained. Although representingonly a limited sample, results are takenfrom one of the main horticulturalvalleys of Nicaragua and, consequently,are interesting in that they show apredominance of the original biotype A.
traditional food crops from most of thefertile valleys in Central America inorder to make room for high-value,non-traditional export crops such astomato, pepper and melon. As a result,there is a higher demand for traditionalcrops such as common bean in theregion. Nicaragua could take advantageof its relatively larger agricultural areato capture the increased regionaldemand for food staples.
Horticultural crops such as tomatoand pepper have been cultivated inNicaragua mainly to meet localdemand. The area planted to tomatohas doubled in the past 30 years from350 to 750 ha, whereas in Guatemala,tomato production grew from 5000 hain 1960 to 12,000 ha in 1970. In 1986,tomato plantings in the Sébaco Valley,Matagalpa Department, sufferedunusual infestations of the whiteflyB. tabaci and, soon after, theemergence of viral diseases associatedwith the whitefly outbreak. By 1991,tomato production in the Sébaco Valleyhad been reduced 20%-50% because ofyield losses ranging between 30%-100% (Sediles, 1998). A similarsituation was observed for pepperplantings, which in 1991-92, reportedyield losses between 30%-50%. Melon,another non-traditional crop grownmostly for export, was beginning toexperience whitefly-related problems.However, there was no significantincidence of begomoviruses in thiscrop.
The whitefly problem on tomatobecame so severe that it prompted thecreation of an inter-institutional tomatogroup composed of CATIE/MAG-MIP–NORAD-ASDI (for full names, seeAcronyms list on page 345), whichorganized the first national meeting oftomato producers in April 1994.Tomato growers met to discuss whiteflymanagement strategies with thetechnical organizing group (Grupo
Table 2. Results of Bemisia tabaci biotypinganalyses performed on 15 samples ofwhiteflies from Apompua andSébaco, department of Matagalpa,Nicaragua.
Sample Biotype A Othercode
N2 4 1
N3 5 -
N4 5 -
Socio-economic Analysis
Common bean is one of the two mainfood staples, together with maize(Zea mays L.), being grown on about140,000 ha. However, productivity islow (about 450 kg/ha) because 80% ofthe national production takes place onfarms of less than 3 ha. The neweconomic policies that followed theeconomic crisis of the 1970s havegradually led to the displacement of

214
Whiteflies and Whitefly-borne Viruses in the Tropics
Interinstitucional e Interdisciplinario deTomate-GIIT). In June 1995, a secondmeeting took place in Santa Emilia,Matagalpa, with 24 producersrepresenting 16 tomato-growing regionsof Nicaragua, where the whitefly is alimiting factor to tomato production. Inthese meetings, tomato farmersbecame aware of the consequences ofabusing pesticides as well as of theexistence of integrated pestmanagement practices (live barriers,yellow traps and organic insecticides).An outcome of the meeting was thecreation of the Nicaraguan TomatoGrowers Association, which isresponsible for the sustainability oftomato production in Nicaragua.
Strengthened ResearchCapacity
The Universidad Nacional Agraria (UNA)located in Managua, the capital ofNicaragua, was selected as the maincollaborating institution. The nationalprogram and the Ministry ofAgriculture of Nicaragua, already havethe substantial financial support ofinternational organizations. Thefinancial support to UNA permitted ageneral evaluation of the whiteflysituation in the six administrativeregions of Nicaragua described below.
Region I includes the north-western departments of Nueva Segovia,Madriz and Estelí. In this region, thereare valleys and plateaus with altitudesranging between 500 and 1000 m anda variety of food crops such as bean,tomato, cucurbits, tobacco and pepper,which attract whiteflies. The whiteflypopulation peak occurs in the periodNovember-May.
Region II includes the south-western departments of León andChinandega and agricultural lowlandsbelow 500 m. The region is formed by
the hot Pacific Plains, where cottonproduction took place in past decades.The whitefly B. tabaci became a limitingbiotic problem for cotton production andthe region has moved on to new crops:tomato, watermelon (Citrullus lanatus[Thunb.] Matsum. & Nakai), squash(Cucurbita spp. L.), pepper, melon andtraditional ones as well such ascommon bean and tobacco. Soybean(Glycine max [L.] Merr.), peanut (Arachishypogaea L.) and cassava (Manihotesculenta Crantz) are other cropsinfested by whiteflies in this region.Whitefly population peaks occur inJanuary and February.
Region III consists of ManaguaDepartment and two vegetableproduction zones, the coastal area ofLake Managua located north of the cityof Managua and the zone ofPochocuape, south of Managua. Here,whiteflies attack tomato, squash,watermelon, common bean and tobaccoand most of these crops are attackedalso by begomoviruses.
Region IV includes the departmentsof Carazo, Granada, Masaya and Rivas,which form the southernmost portion ofthe Pacific Lowlands. At these lowaltitudes, the whitefly B. tabaci thrives,severely attacking tomato and tobaccoand, to a lesser extent, watermelon,squash, common bean, pepper andmelon. The main whitefly peak occurs inJanuary and February.
Region V is formed by thedepartments of Boaco and Chontales.Although this area has been devotedprimarily to livestock, there are someparts such as Santa Lucía, Boaco,where bean and tomato traditionallyhave been planted. These crops havebeen affected by whitefly-transmittedbegomoviruses since the mid 1980s,particularly by BGYMV (common bean)and by the crespo disease in the case oftomato.

215
Nicaragua
Region VI is a main horticulturalarea of Nicaragua, particularly theSébaco Valley, in the department ofMatagalpa. The second departmentthat forms this region, Jinotega, liesfurther north, bordering Honduras. Thetomato crop in this region has beenaffected severely by whitefly-bornebegomoviruses, to the point that somefarmers have abandoned this crop,particularly in the southern part of theSébaco Valley. Large whiteflypopulations and viral symptoms alsoaffected cucurbits such as squash,pumpkin (Cucurbita spp. L.) andcucumber (Cucumis sativus L. var.sativus).
It is evident that whiteflies andB. tabaci-transmitted viruses affect allof the agricultural regions of Nicaragua.Nicaraguan agricultural researchinstitutions have come together toreach affected farmers and manage thewhitefly problem in a highly exemplarymanner. The integrated pestmanagement projects developed inNicaragua with the collaboration ofinternational and national institutionshave recovered some of the mostaffected agricultural areas for theproduction of traditional (e.g., commonbean) and non-traditional (e.g., tomato)crops. It is expected that the scientificknowledge generated in these projectswill contribute to the implementation ofmore effective measures to controlwhiteflies and the viruses they transmitin Nicaragua and Middle America.
Current Status ofWhitefly/Begomovirus
Problems
Most of the whitefly/begomovirusproblems in Nicaragua occur in thewestern half of the country, mainlybecause this region concentrates mostof the country’s population. Theeastern half of Nicaragua (the
Caribbean Plains) is only cultivated to aminor extent with, for example, banana(Musa spp. L.), rice (Oryza sativa L.)and oil palm (Elaeis guineensis Jacq.),because it is one of the rainiest regionsin Central America. This climatic factoris a major deterrent for the whiteflyB. tabaci and for the establishment ofits main plant hosts. The PacificLowlands, on the other hand,experience several dry months withoutsignificant rainfall and thus possess allthe necessary conditions for thewhitefly B. tabaci to thrive, includingthe presence of good breeding hostssuch as cotton. Moreover, as in the restof Central America, most horticulturalcrops are located in mid-altitude(200-1000 m) valleys created by themountain ranges that make up thecentral and northern regions ofNicaragua. Valleys such as the Sébaco,Matagalpa Department (450 m)constitute an example of a locality witha high incidence of B. tabaci andbegomoviruses, because of thefavourable environmental conditions forthe pest and the presence of susceptiblehorticultural crops, some of which alsoact as suitable reproductive hosts forB. tabaci.
References
Gill, A. 1994. Problemática del complejomosca blanca-virus en algodón enCentroamérica. In: Mata, M.;Dardón, D. E.; Salguero, V. E. (eds.).Biología y manejo del complejomosca blanca-virosis. Memorias IIITaller Centroamericano y del Caribesobre mosca blanca, Antigua, GM.p. 23-38.
Hidalgo, O.; León, G.; Lindo, O.;Vaugham, M. 1975. Informe de lamisión de estudio de la moscablanca. Banco Nacional deNicaragua (BNN), Comisión Nacionaldel Algodón (CONAL), Ministerio deAgricultura y Ganadería (MAG),Managua, NI. 120 p.

216
Whiteflies and Whitefly-borne Viruses in the Tropics
Kramer, P. 1966. Serious increase ofcotton whitefly and virustransmission in Central America. J.Econ. Entomol. 59:1531.
Llano, A.; Munguía, R.; Viana, A. 1997.Perfil de la producción de frijol enNicaragua. Instituto Nacional deTecnología Agropecuária (INTA)-Programa Cooperativo Regional deFrijol para Centro América, Méxicoy el Caribe (PROFRIJOL). 33 p.(Documento de trabajo)
Rojas, A.; Kvarnheden, A.; Valkonen, J. P.T. 2000. Geminiviruses infectingtomato crops in Nicaragua. PlantDis. 84:843-846.
Sediles, A. 1998. La mosca blancaBemisia tabaci (Gennadius) enNicaragua: Historia y problemática.Informe Universidad NacionalAgraria (UNA), NI. 23 p.
Zamora, M. E. 1996. Identificación deplantas silvestres como reservoriosde los virus del mosaico dorado(BGMV) y del mosaico enano(BDMV) del frijol en Pueblo Nuevo,Nicaragua. M.Sc. thesis,Universidad Nacional Agraria, NI.84 p.

217
Costa Rica
CHAPTER 3.7
Costa RicaFrancisco Morales*, Luko Hilje**, Juan Vallejos**,
Guillermo Sibaja***, Carlos Arayaψ and Rodolfo Arayaψψ
Introduction
Geographical context
The Central Plateau, around the capitalcity of San José, is one of the mostintensively cultivated areas in CostaRica. North and south of this region,the broken and volcanic topography ofmountain ranges expands all the wayto the northern border with Nicaraguaand the southern border with Panama.East and west of the northernmountain ranges lie the Caribbean andPacific lowlands, the least developedand inhabited regions of Costa Rica.South-eastern Costa Rica receives over1500 mm of rain annually, whichprompted the development of a thrivingbanana (Musa spp. L.) productionindustry towards 1880. Coffee (Coffeaarabica L.) was introduced into CostaRica in the 1830s and soon became asource of wealth for many farmers inthe Central Plateau. Among the basicfood staples, maize (Zea mays L.) andcommon bean (Phaseolus vulgaris L.)are important components of the CostaRican diet, together with crops such aspotato (Solanum tuberosum L.) and
plantain (Musa × paradisiacal L.).Cotton (Gossypium hirsutum L.) isproduced only to a limited extent in thePacific lowlands of the Province ofGuanacaste (West and Augelli, 1977;Pastor, 1988). The Central Plateau isundoubtedly one of the most affectedareas in terms of whitefly-relateddamage. Figure 1 shows the mainagricultural regions affected bywhitefly-transmitted begomoviruses.
* Centro Internacional de AgriculturaTropical (CIAT), Cali, Colombia.
** Centro Agronómico Tropical deInvestigación y Enseñanza (CATIE), CostaRica.
*** Ministerio de Agricultura, Costa Rica.ψ Universidad Nacional, Costa Rica.ψψ Universidad de Costa Rica.
San José
Costa Rica
Cartago
Figure 1. The main agricultural regionsaffected by whitefly-transmittedbegomoviruses, Costa Rica.
The emergence of Bemisiatabaci as a pest and virusvector
The lack of suitable reproductive hostsfor the whitefly Bemisia tabaci(Gennadius) such as cotton or soybean(Glycine max [L.] Merr.), together withthe high rainfall over much of theCosta Rican territory, protected the
GuanacasteAlajuela
Heredia
Puntarenas
Puntarenas
Limón

218
Whiteflies and Whitefly-borne Viruses in the Tropics
Central Plateau’s agricultural regionagainst the early arrival of this pest.The first records of the presence ofB. tabaci in the country originated inthe 1970s, precisely in the drierlowlands of the Pacific province ofGuanacaste, where an incipient cottonindustry was developed in the 1960s.The first record of B. tabaci as a pestand vector of a plant virus Bean goldenyellow mosaic virus (BGYMV) incommon bean fields in the CentralValley, occurred in 1987. BGYMValready had been observed in thecountry in the 1960s (Gámez, 1970),albeit at a very low incidence. In 1988,B. tabaci moved on to tomato(Lycopersicon esculentum Mill.) in theCentral Valley (Hilje, 1997).Coincidentally, the attack to commonbean and tomato took place in thewestern areas of the Central Valley,which is the closest region to thecotton-growing areas of Costa Rica.B. tabaci is currently a pest and vectorof viruses in common bean and tomatofields in most of the horticulturalprovinces of the country.
Advances in BiologicalResearch
A BGYMV isolate from Alajuela (CentralValley) had been characterizedserologically at the Centro Internacionalde Agricultura Tropical (CIAT) in 1993as a member of the Central American/Caribbean group of BGYMV isolates.However, different begomovirusesisolated during the course of thisproject from BGYMV-affected beanplants in different regions of Costa Rica(Puriscal, Alajuela and Los Chiles) wereshown to be serologically distinct fromthe original Middle American BGYMVisolates characterized in the early1990s (Cancino et al., 1995).
A begomovirus of tomato had beenisolated previously in the locality of
Turrialba (Castillo, 1997) and identifiedas Tomato yellow mosaic virus (ToYMV).This virus is currently considered atentative new species referred to asTomato yellow mottle virus (ToYMoV).Another begomovirus, isolated fromdiseased tomato plants in Turrialba,was shown in 1998 to be related to atomato begomovirus tentativelydesignated as Tomato leaf curl Sinaloavirus, originally isolated in north-western Mexico (Idris et al., 1999).
The Tropical Whitefly Projectfinanced a survey of B. tabaci biotypesin Costa Rica, conducted under theguidance of Dr. Luko Hilje of theCentro Agronómico Tropical deInvestigación y Enseñanza (CATIE),Turrialba. Table 1 shows the results ofthe molecular characterization analysesconducted at CIAT for some of thesamples collected. These resultsconfirmed the presence of B. tabaci invarious crops of socio-economicimportance and showed thepenetration of biotype B of B. tabaci indifferent regions of Costa Rica.However, at the time of the survey, theoriginal A biotype still predominated inthe country. Some of the most heavilycolonized crops were cucurbits but,unlike the case of other CentralAmerican countries, B. tabaci wasfound to colonize pepper (Capsicumspp. L.) in Costa Rica (Hilje, 1997;Vallejos, 1997).
Socio-economic Analysis
At the beginning of this project, aprevious investigation on the economicimpact of the B. tabaci-begomoviruspest complex in Costa Rica, andparticularly on common bean, tomatoand pepper, was used as ex ante datafor the survey conducted during thecurrent project (Vallejos, 1997).According to the results obtained inthat investigation, the trend to replace

219
Costa Rica
basic food crops such as common beanfor high-value crops such as tomatoand pepper was quite apparent andstill continues in Costa Rica. As aresult, common bean and othertraditional crops are increasinglyplanted in marginal soils, with an
expected fall in the productivity ofthese crops.
Yield losses due to whitefly-transmitted viruses were 37% forcommon bean, 22% for tomato and 9%for pepper. The whitefly B. tabaci could
Table 1. Characterization of Bemisia tabaci biotypes in Costa Rica.
Province Locality Crop Biotype
Alajuela Orotina Watermelon BAlajuela Orotina Cucumber BPuntarenas Puntarenas Melon BPuntarenas Ticaral Watermelon BPuntarenas Ticaral Pumpkin APuntarenas Lepanto Common bean A/B
Guanacaste Carrillo Melon AGuanacaste Carrillo Watermelon AGuanacaste Bagaces Melon AGuanacaste Bagaces Spider flower AGuanacaste Bagaces Cucurbit AGuanacaste Tilarón Tomato A
Guanacaste Tilarón Chilli ASan José Pérez Zeledón Snap bean ASan José Pérez Zeledón Pepper ASan José Pérez Zeledón Chilli ASan José Pérez Zeledón Tomato ASan José Pérez Zeledón Tomato A
Heredia not cited Chilli AHeredia not cited Tomato AHeredia not cited Chilli AHeredia not cited Chilli ALimón Guácimo Chilli ALimón Guápiles Pumpkin A
Alajuela Gracia Tomato AAlajuela Gracia Chilli AAlajuela Naranjo Chilli AAlajuela Naranjo Cucumber AAlajuela S. Ramón Tomato AAlajuela Atenas Tomato AAlajuela Alajuela Sweetpotato A
Alajuela Alajuela Tomato AAlajuela Alajuela Chilli AAlajuela Alajuela Common bean AAlajuela Alajuela Cucumber ACartago Turrialba Chilli ACartago Cervante Tomato A
Cartago Turrialba Tomato BCartago Turrialba Chilli ACartago Cartago Tomato ACartago Cartago Chilli ACartago Paraíso Tomato BCartago Paraíso Chilli B

220
Whiteflies and Whitefly-borne Viruses in the Tropics
reproduce on pepper without causingapparent damage to this host plant.Regarding the economic feasibility ofthese crops, tomato and chilli(Capsicum sp.) were the most profitablecrops, producing over US$6300/ha.The net return for common bean wasUS$145/ha.
Strengthened ResearchCapacity
Costa Rica, and specifically CATIE, hasbeen a leader in the collection anddissemination of information onintegrated pest management (IPM)strategies to control whitefly pests.Dr. Luko Hilje of CATIE has co-ordinated these information exchangesthrough an international networkcreated in 1992. Annual workshopshave been organized in different LatinAmerican countries to update anddiscuss whitefly-related problems andthe most suitable IPM measuresimplemented to date. The SystemwideTropical Whitefly (TWF)-IPM Project hasgiven its support to this effort bypromoting complementary activities,particularly in the area of molecularcharacterization of whiteflies andbegomoviruses. With the fundsprovided by this project, Dr. Hilje wasable to attend the InternationalWorkshop on Bemisia andGeminiviruses held in San Juan,Puerto Rico, in June 1998. Hispresence in this meeting was importantfor maintaining the Latin Americanwhitefly network active and forplanning future activities. Additionally,the funds provided to CATIE allowedDr. Hilje to conduct a countrywidesurvey of the whitefly/begomovirus-affected regions in Costa Rica.
A short study fellowship wasawarded to Ing. Agr. Guillermo SibajaChinchilla of the Costa Rican Ministryof Agriculture and Livestock, Plant
Health Division. Mr. Sibaja came toCIAT, Colombia, for intensive trainingon the characterization of B. tabacibiotypes.
Current Status ofWhitefly/Begomovirus
Problems
Whiteflies and begomoviruses in CostaRica mainly affect common bean andtomato crops, particularly in theCentral Valley, where most of thehorticultural crops are grown. Anotherarea where the whitefly problem isgaining momentum is the “PacíficoSeco” (Dry Pacific Region) inGuanacaste Province. Rainfall in thisregion ranges from 1220 to 2000 mmannually, with an average temperatureof 20 to 25 ºC. These are not optimalconditions for B. tabaci but theoccurrence of a prolonged dry periodfrom December through April favoursthe build up of whitefly populationsand transmission of viruses tosusceptible horticultural crops.Fortunately, most of the Costa Ricanterritory receives over 1500 mm ofannual rainfall, which maintainswhitefly populations depressed most ofthe year. These climatic factors andvarious IPM measures implemented inCosta Rica, including the use ofresistant varieties in the case ofcommon bean, have contributedsignificantly to the attenuation of thewhitefly/begomovirus problems in thiscountry.
References
Cancino, M.; Abouzid, A. M; Morales, F. J;Purcifull, D. E; Polston, J. E.;Hiebert, E. 1995. Generation andcharacterization of threemonoclonal antibodies useful indetecting and distinguishing Beangolden mosaic virus isolates.Phytopathology 85:484-490.

221
Costa Rica
Castillo, J. 1997. Movimiento diario deBemisia tabaci en parcelas detomate, diseminación local delmosaico amarillo y fuentes deinoculo del ToYMV-CR en Guayabo,Costa Rica. M.Sc. thesis, CentroAgronómico Tropical deInvestigación y Enseñanza (CATIE),Turrialba, CR. 93 p.
Gámez, R. 1970. El virus del moteadoamarillo del frijol, plantashospederas y efecto en producción.In: XV Reunión Annual PCCMCA,Antigua Guatemala, GT. p. 44-48.
Hilje, L. 1997. Posibilidades para elmanejo integrado del complejoBemisia tabaci-geminivirus en CostaRica. Agron. Costarricense 21:139-142.
Idris, A. M.; Rivas-Platero, G.; Torres-Jerez, I.; Brown, J. K. 1999. Firstreport of Sinaloa tomato leaf curlgeminivirus in Costa Rica. PlantDis. 83:303.
Pastor, R. 1988. Historia deCentroamérica. Editorial PiedraSanta, GT. 272 p.
Vallejos, J. E. 1997. Sistema experto parala evaluación del impacto delcomplejo Bemisia tabaci-geminivirus, en frijol, tomate y chiledulce con fines de planificación.M.Sc. Thesis, Centro AgronómicoTropical de Investigación yEnseñanza (CATIE), Turrialba, CR.109 p.
West, R. C.; Augelli, J. P. 1977. MiddleAmerica: Its lands and peoples.Second edition. Prentice-Hall, NJ,USA. 494 p.

222
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 3.8
Panama
Francisco Morales*, Orencio Fernández**† andJosé Guerra**
Introduction
Geographical context
Most of the Panamanian territoryreceives tropical rains in excess of2500 mm/yr, a climatic factor knownto drastically reduce whiteflypopulations. Nevertheless, during thefirst (1992) workshop on whiteflies inCentral America and the Caribbean,held in Costa Rica, Panama wasalready present (Zachrisson andPoveda, 1992). According to the reportpresented by the Panamanian delegateat that meeting, the emergence ofwhiteflies as agricultural pests inPanama was associated with theexpansion of non-traditional crops,mainly melon (Cucumis melo L.), forexport to the United States. Threehorticultural areas were developed inthe late 1980s: the Pacific Coast of theprovince of Panama; the centralprovinces of Coclé, Herrera and LosSantos (the largest); and the westernprovince of Chiriquí, near the CostaRican border (Zachrisson and Poveda,1992; Delgado, 1994). These regionshave in common the presence of a well-defined dry period and below-averageprecipitation during the year. Figure 1
shows the main agricultural regionsaffected by whitefly-transmittedbegomoviruses.
The emergence of Bemisiatabaci as a pest and virusvector
The presence of Bemisia tabaci(Gennadius) on tomato (Lycopersiconesculentum Mill.) was first observed inthe Azuero Peninsula in 1983.However, it was not until 1991, whenunusually dry climatic conditionsfavoured the reproduction of B. tabaci,that this whitefly species became a pestin the central provinces of Panama.B. tabaci has been observed toreproduce in eggplant (Solanummelongena L.), sweet pepper (Capsicumannuum L. var. annuum) and chillipepper (Capsicum spp. L.). In 1992, thesilver leaf symptom was observed onsquash (Cucurbita spp. L.) attacked byB. tabaci, which suggested the presenceof biotype B in Panama. The westernhorticultural zone, located in ChiriquíProvince, is at a higher elevation andreceives more rainfall than the othertwo horticultural zones of Panama. Theproblem pest in this area is not somuch B. tabaci as Trialeurodesvaporariorum (Westwood), a whiteflyspecies that attacks horticultural cropsat altitudes above 1000 m (Zachrissonand Poveda, 1992).
In Panama, common bean(Phaseolus vulgaris L.) is cultivated in
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** Instituto de Investigación Agropecuaria dePanamá (IDIAP).
† R.I.P.

223
Panama
Chiriquí Province, at altitudes between550 and 1000 m. Bean golden yellowmosaic virus (BGYMV) was observedaffecting common bean in Panamaprior to 1970 (Gámez, 1970) but thisdisease never has been a majorproduction problem of common bean inPanama.
Advances in BiologicalResearch
A whitefly-borne geminivirus affectingtomato was described in Panama in1998 (Engel et al., 1998). The virus,named Tomato leaf curl virus, wasshown in this project to have a highdegree of nucleotide sequencesimilarity when compared to Tomatoyellow mosaic virus (ToYMV) fromVenezuela. The virus had beendescribed in Panama as a strain ofPotato yellow mosaic virus, a misnomerfor ToYMV. Although this virus may bea new species, the name “Tomato leafcurl virus” is not appropriate because itis already given to a distinctbegomovirus species that only exists inthe Old World (Asia). Therefore, thePanamanian virus should be renamed.
A molecular characterization ofB. tabaci biotypes from the central
horticultural zone of Azuero wasconducted at the Centro Internacionalde Agricultura Tropical (CIAT), withsamples provided by Ing. Agr. JoséAngel Guerra of the Instituto deInvestigación Agropecuaria de Panamá(IDIAP) (Table 1). Results demonstratethe coexistence of biotypes A and B ofB. tabaci in the central provinces of theAzuero Peninsula. This is aninteresting situation that needs to bemonitored in order to study theoutcome of the interaction betweenthese two biotypes in a horticulturalzone characterized by mixed croppingsystems.
Socio-economic Analysis
Because Panama was not originallyincluded in this project, no socio-economic information was collected forthis country. However, it is known thatwhitefly-transmitted geminiviruses donot affect common bean severely inChiriquí Province.
Between 1991 and 1994, about6000 tons of tomato were lost towhitefly problems in Panama. Duringthe 1997-98 tomato planting season,2800 tons were lost because of theeffect of B. tabaci, both as a pest and
PanamaLos Santos
Figure 1. The main agricultural regions affected by whitefly-transmitted begomoviruses, Panama.
Bocas del Toro
Chiriquí
Veraguas
Colón
Coclé
Herrera
Colón
Darién
PanamaComarca de
San Blas
Panama

224
Whiteflies and Whitefly-borne Viruses in the Tropics
vector of plant viruses. Whereas large-scale tomato growers absorbed most ofthe losses, these pests affected anumber of small-scale producers in theprovince of Los Santos.
Strengthened ResearchCapacity
Considering that Panama was notincluded officially in the project, anagreement was made with the RedColaborativa de Investigación yDesarrollo de Hortalizas para AméricaCentral, Panamá y RepúblicaDominicana (REDCAHOR), ahorticultural research networkoperating in the region, to trainPanamanian researchers. The networkfinanced the training at CIAT of twonational scientists: an entomologist(Ing. José Guerra) and a virologist(Dr. Orencio Fernández) from IDIAP.
These scientists were trained in themolecular characterization of B. tabacibiotypes and the use of monoclonalantibodies for the identification ofbegomoviruses.
Current Status ofWhitefly/Begomovirus
Problems
Although Panama is one of the rainiestcountries in the world, with an averageannual rainfall above 2000 mm, thereare less humid (1000-2000 mm)coastal regions around the Gulf ofPanama, where horticulturalproduction has increased significantlyin the last decades. The mainhorticultural area is located in theAzuero Peninsula, where most of thewhitefly/begomovirus problems haveemerged in past years. Whiteflypopulations increase in the driermonths of the year: January-March.Panama is in the process of expandingthe production of non-traditionalexport crops and consequently thiscountry should take measures tocontrol the whitefly problem before itbecomes unmanageable.
References
Delgado, A. 1994. Cultivos hortícolas enPanamá: Algunos datosestadísticos. Folleto informativo,Instituto de InvestigaciónAgropecuaria de Panamá (IDIAP),PA. 17 p.
Engel, M.; Fernández, O.; Jeske, H.;Frischmuth, T. 1998. Molecularcharacterization of a new whitefly-transmissible bipartite geminivirusinfecting tomato in Panama. J. Gen.Virol. 79:2313-2317.
Table 1. Molecular characterization of Bemisiatabaci biotypes from Azuero, Panama.
Sample Crop Biotype
1 Tomato A/B
2 Melon A
3 Melon B
4 Melon A/B
5 Melon A
6 Tomato B
7 Tomato B
8 Tomato B
9 Tomato B
10 Tomato A/B
11 Melon A/B
12 Melon B
13 Melon B
14 Melon B
15 Melon B
16 Melon B
17 Melon B
18 Melon A/B

225
Panama
Gámez, R. 1970. Los virus del frijol enCentroamérica. I. Transmisión pormoscas blancas (Bemisia tabaciGen.) y plantas hospedantes delvirus del mosaico dorado. Turrialba21:22-27.
Zachrisson, B.; Poveda, J. 1992. Lasmoscas blancas en Panama. In:Hilje, L.; Arboleda, O. (eds.). Lasmoscas blancas (Homoptera:Aleyrodidae) en América Central y elCaribe. Centro Agronómico Tropicalde Investigación y Enseñanza(CATIE), Turrialba, CR. p. 64-66.

226
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 3.9
HaitiFrancisco Morales*, Jackson Donis** and
Emmanuel Prophete**
Introduction
Geographical context
Haiti is a mountainous country; 40% ofits territory is located at altitudes above490 m. The mountain ranges alternatewith fertile valleys, the largest being thePlaine du Nord. Annual rainfall variesfrom 500 in the north-west to2500 mm in the eastern/southernhighlands. Agriculture accounts forone-third of the gross domestic productbut subsistence farming predominates.Maize (Zea mays L.; 250,000 ha),sorghum (Sorghum bicolor [L.] Moench;160,000 ha), common bean (Phaseolusvulgaris L.; 100,000 ha) are the mainstaples, with rice (Oryza sativa L.) andsweetpotato (Ipomoea batatas [L.] Lam.)also being important components ofthe Haitian diet. However, most of thefood consumed in Haiti has to beimported. Coffee (Coffea arabica L.) isthe main export crop (145,000 ha),followed by sugarcane (Saccharumofficinarum L.), sisal (Agave sisalanaPerrine ex Engelm.) and cacao(Theobroma cacao L.). French-Creole isspoken by 90% of the population andonly 10% speak French. Horticultural
crops have become increasinglyimportant in Haiti, mainly as anextension of the agricultural exportbusiness in the Dominican Republic.Figure 1 shows the main agriculturalregions affected by whitefly-transmittedbegomoviruses.
The emergence of Bemisiatabaci as a pest and virusvector
The whitefly Bemisia tabaci was firstregarded as a serious pest in Haiti, inthe early 1980s. Common bean, tomato(Lycopersicon esculentum Mill.),eggplant (Solanum melongena L.),pepper (Capsicum spp. L.), cowpea(Vigna unguiculata [L.] Walp.) and limabean (Phaseolus lunatus L. var.lunatus) are the main crops affected bythis insect pest (Donis and Prophete,1997). As a vector, B. tabaci alreadyhad been observed to transmit Beangolden yellow mosaic virus (BGYMV) in1978 (Balthazar, 1978).
Advances in BiologicalResearch
The main agricultural areas of Haitiwere surveyed and plant samplesanalysed to determine the importanceof whiteflies and whitefly-borne viruses.Table 1 shows the different regionsvisited and the results obtained fromthe tests practiced with samplescollected. Of whitefly-transmitted
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** Centre de recherche et de documentationagricoles-Ministère de l’agriculture desressources naturelles et du développementrural (CRDA-MARNDR), Haiti.

227
Haiti
begomoviruses, BGYMV was present incommon bean and Tomato yellow leafcurl virus (TYLCV) in tomato. As in thecase of the Dominican Republic, thespecific monoclonal antibody (MAB-GA), which recognizes BGYMV isolatesfrom Central America and southernMexico, detected the Haitian isolate ofBGYMV. This isolate was partially
sequenced and compared to the rest ofBGYMV isolates from the Caribbeanand Central America characterized todate. Table 2 shows the results of thesecomparative analyses.
As suggested by the serologicalassay using the Guatemalan (GA)specific MAB, the Haitian BGYMV
Haiti
Nord-Ouest
Nord-Est
Centre
Nord
Artibonite
Sud-Est
Grande-Anse
Figure 1. The main agricultural regions affected by whitefly-transmitted begomoviruses, Haiti.
Table 1. Results of serological and polymerase chain reaction (PCR) assays of selected samplescollected in Haiti.
Sample Locality Plant Reactiona
MAB-BS MAB-GA PTY1 TYLCV
1 Cul-de-Sac Cowpea - nt - nt
2 Passereine Tomato - nt - +
3 Passereine Tomato - nt - +
4 Passereine Tomato - nt - +
5 Passereine Cowpea - nt + nt
6 Passereine Common bean + + - nt
7 Terrier Rouge Tomato - nt + +
8 Terrier Rouge Tomato - nt - +
9 St Raphael Tomato - nt - +
10 St Raphael Tomato - nt - +
11 St Raphael Pepper - nt + nt
a. Positive or negative reactions to a broad spectrum monoclonal antibody used to detect bi-partitebegomoviruses (MAB-BS); a MAB used to detect the original Middle American isolates of Beangolden yellow mosaic virus-Guatemala (MAB-GA); a potyvirus-specific monoclonal antibody (PTY1);and PCR detection of Tomato yellow leaf curl virus; and nt, no test.
Ouest
Sud
Sud
Grande-Anse
Ouest
Nord-Ouest
Ouest

228
Whiteflies and Whitefly-borne Viruses in the Tropics
isolate assayed is similar to the CentralAmerican and Caribbean BGYMVisolates but different from the SouthAmerican bean begomoviruses, Beangolden mosaic virus and Bean dwarfmosaic virus. Few of the tomato, pepperand cowpea samples collected in Haiticontained aphid-transmittedpotyviruses. All tomato samples wereshown by polymerase chain reaction(PCR) to be infected by TYLCV. Thecomposite whitefly sample collectedfrom eggplant resulted in the detectionand identification of biotype B ofB. tabaci. Although preliminary, theseresults suggest that the whitefly andbegomovirus problems in theHispaniola Island (Dominican Republicand Haiti) are similar.
Socio-economic Analysis
Whitefly-transmitted begomoviruseshave been observed to cause significantor even total yield losses in commonbean and susceptible horticulturalcrops in Haiti. In the case of commonbean, the cultivation of BGYMV-resistant varieties (available through thePrograma Cooperativo Regional de Frijolpara Centro América, México y el Caribe[PROFRIJOL] project) and thecultivation of common bean at altitudesabove 1000 m should reduce theincidence of BGYMV. The same strategycould be adopted for begomovirus-susceptible horticultural crops, that is,growing vegetables above 1000 m. For
valleys below this altitude, whereB. tabaci thrives, cultural practicessuch as the implementation of a periodfree of certain crops that act asreproductive hosts for the whiteflyshould be adopted following the modelimplemented in the DominicanRepublic. Although there are tomatovarieties resistant to TYLCV, these arecommercial varieties that small-scalefarmers cannot afford. The productionof high-value crops (e.g., tomato,pepper and eggplant) by small-scalefarmers could greatly benefit from theadoption of integrated pestmanagement (IPM) practices such asthe use of micro-tunnels and biologicalinsecticides or mild soaps.
Strengthened ResearchCapacity
Two collaborating scientists from theCentre de recherche et dedocumentation agricoles-Ministère del’agriculture des ressources naturelleset du développement rural (CRDA-MARNDR, Damien), participated in thefield survey conducted in Haiti. Thesescientists were invited to attendinternational workshops in CentralAmerica and to acquire new whitefly/begomovirus characterizationtechniques at the Centro Internacionalde Agricultura Tropical (CIAT) butofficial and administrative problemsprevented their participation in theseactivities. Nevertheless, as
Table 2. Comparative nucleotide sequence homology (%) between a begomovirus isolated fromcommon bean in Haiti and previously sequenced common bean begomoviruses.
Region BGa Haiti Common bean begomovirusesb
BGYMV BGYMV BGYMV BGMV BDMVDominican Guatemala Puerto Rico Brazil ColombiaRepublic
Coat protein (AV1) 100 90.2 92.0 89.5 68.3 68.7
Replicase (AC1) 100 87.5 90.9 88.9 77.6 78.8
a. BG, begomovirus.b BGYMV, Bean golden yellow mosaic virus; BGMV, Bean golden mosaic virus; and BDMV, Bean
dwarf mosaic virus.

229
Haiti
collaborators in this project, theseresearchers have benefited from theinformation generated to date.
The visit to Haiti was also helpfulfor establishing closer links betweenthe PROFRIJOL project, CIAT andHaiti. As a result, two bean breedersare now actively working to deployBGYMV-resistant bean varieties inHaiti, and the United States Agency forInternational Development (USAID) hasfinanced a new project to work onsustainable agricultural practices,including hillside technologies and foodsecurity issues in Haiti.
Current Status ofWhitefly/Begomovirus
Problems
The importance of B. tabaci as a pestand vector of plant viruses is expectedto decrease in the near future as newbean cultivars possessing resistance toBGYMV are introduced by the USAID-funded project. The introduction ofTYLCV-resistant tomato varieties fromthe Dominican Republic should alsoreduce the impact of TYLCV in Haiti.However, poverty issues will limit theadoption of resistant cultivars,particularly in the case of tomato. The
low economic capacity of most small-scale farmers practicing subsistenceagriculture also precludes the use ofeffective but expensive insecticides(e.g., imidacloprid) to control whitefliesand whitefly-transmitted viruses. Thelow levels of insecticidal action ofcheaper pesticides encouragespesticide abuse, with the consequentdevelopment of insecticide resistance inB. tabaci populations. Finally, Haitimight benefit from its brokentopography to avoid the attack ofB. tabaci by planting susceptible cropsat altitudes above 1000 m.
References
Balthazar, S. 1978. Les viroses du haricotcommun en Haiti. FondoSalvadoreño de Estudios del’Agriculture, des RessourcesNaturelles et du DéveloppementRural (DARNDR)-Servicio Nacionalde Sanidad Agropecuária (SENASA),Damien, SV.
Donis, J.; Prophete, E. 1997. Las moscasblancas (Homoptera:Aleyrodidae) enHaití: Situación actual y manejo.In: Memoria del VI TallerLatinoamericano y del Caribe sobremoscas blancas y geminivirus,18-19 agosto 1997, Santo Domingo,DM.

230
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 3.10
CubaFrancisco Morales*, Gloria González**, Carlos Murguido**,
Ana Echemendía**, Yolanda Martínez***, Yenín Hernández***,Benito Faureψ and María Chaillouxψ
Introduction
Geographical context
About 25% of Cuba’s territory ismountainous, with three distinctmountain ranges running east-west:the eastern (Sierra Maestra), central(alturas) and western (Cordillera deGuaniguanico) ranges. The remainingland is composed of extensive plainsand basins. Cuba has a semi-tropicalclimate with two seasons: dry fromNovember through April and rainy fromMay through October. The meanannual temperature is 26 °C, with a23-28 °C range. The averageprecipitation is 1380 mm. All theseconditions favour the dissemination ofwhiteflies, particularly during the dryseason of the year, particularly in theregions marked in Figure 1.
Horticultural products have beentraditional commodities in Cuba,particularly those referred to asviandas and basic grains. These cropsinclude tomato (Lycopersiconesculentum Mill.), squash (Cucurbitaspp. L.), cucumber (Cucumis sativus L.
var. sativus), cabbage (Brassicaoleraceae L.), sweetpotato (Ipomoeabatatas [L.] Lam.), eggplant (Solanummelongena L.) and common bean(Phaseolus vulgaris L.). Vegetableproduction has become increasinglyimportant to support a rapidlyexpanding tourist industry in Cuba.
The emergence of Bemisiatabaci as a pest and virusvector
The emergence of Bemisia tabaci(Gennadius) as a pest is probablylinked to the advent and intensive useof agricultural pesticides soon afterWorld War II. B. tabaci was reported asa pest of tobacco (Nicotiana tabacum L.)and a vector of plant viruses ofcommon bean in the mid 1970s(Blanco and Bencomo, 1978). But itwas not until 1989 that B. tabacibecame a major production problem oftomato and common bean, as a vectorof begomoviruses (Murguido et al.,1997). Currently, this whitefly speciesattacks tomato, common bean, squash,cucumber, melon (Cucumis melo L.),cabbage and eggplant throughoutCuba.
By 1990, the new B biotype ofB. tabaci already had been introducedinto Cuba and surrounding Caribbeanislands (Brown, 1994). Between 1991and 1993, the silver leaf syndromeinduced by this new biotype wasobserved on squash (Murguido et al.,
* Centro Internacional de AgriculturaTropical (CIAT), Cali, Colombia.
** Instituto de Investigaciones de SanidadVegetal (INISAV), Cuba.
*** Centro Nacional de Sanidad Agropecuaria(CENSA), Cuba.
ψ Instituto de Investigaciones de Hortícolas“Liliana Dimitroya” (LILIANA), Cuba.

231
Cuba
1997). Another major productionproblem for Cuba and the Caribbeanregion occurred when and Old Worldvirus, Tomato yellow leaf curl virus(TYLCV) was introduced from Israelinto the Caribbean region, includingCuba (Polston and Anderson, 1997).This exotic virus has caused croplosses worth millions of dollars in theCaribbean region, and it is now knownto occur in southern USA and thePeninsula of Yucatán, Mexico.
Advances in BiologicalResearch
A main objective of the researchundertaken in Cuba was to determinethe composition of the whiteflypopulation. A Cuban trainee, Ms. YenínHernandez of the Centro Nacional deSanidad Agropecuaria (CENSA)conducted the tests at the CentroInternacional de Agricultura Tropical(CIAT) in Colombia, under thesupervision of CIAT technical staff.Ninety-nine samples were collectedfrom tomato plants in the localities ofQuivicán and Alquizar, HavanaProvince. The results of the randomamplified polymorphic DNA (RAPD)assay showed that 100% of the whiteflysamples tested corresponded to biotypeB of B. tabaci. Additionally, sevenwhitefly samples from potato (Solanumtuberosum L.) plants grown in Havana
Province were also shown to consist ofB. tabaci biotype B. These findingssuggest that biotype B has adaptedwell and now predominates overbiotype A of B. tabaci in HavanaProvince.
Bean golden yellow mosaic virus(BGYMV) has been an importantdisease of common bean in Cuba sincethe early 1970s (Blanco and Bencomo,1978). The epicentre of the problemwas the locality of Velasco in thewestern province of Holguín and it isnow widely disseminated in theprovinces of Holguín, Las Tunas,Guantánamo and Havana. In recentyears, the incidence of BGYMV hasincreased in the provinces of Ciego deAvila, Holguín, Las Tunas andCamagüey.
When the monoclonal antibodies(MABs) to BGYMV were developed in1990 (Cancino et al., 1995), the Cubanisolate of BGYMV reacted with amonoclonal antibody (MAB 2G5)produced to a Guatemalan isolate ofBGYMV, which recognized all of theMiddle American BGYMV isolates. By1993, the Cuban isolates of BGYMVwere not reacting with the specific MAB2G5. As can be observed in Table 1,none of the BGYMV samples from threedifferent provinces of Cuba reactedwith the specific MAB-GA (2G5),including a BGYMV isolate from lima
Cuba
Pinar del Río
Camagüey
Matanzas
Las Tunas
La Habana
Holguín
Ciego deAvila
Ciudad de la Habana
Figure 1. The main agricultural regions affected by whitefly-transmitted begomoviruses, Cuba.
Isla de laJuventud
Villa Clara
Cienfuegos
SanctiSpiritus
GranmaSantiago de Cuba
Guantánamo

232
Whiteflies and Whitefly-borne Viruses in the Tropics
bean (Phaseolus lunatus L.). Thehomologous control, BGYMV-GA,reacted with MAB 2G5 as expected.However, all of the diseased plantsamples that were assayed reactedwith the broad spectrum MAB (3F7) toBGYMV, which recognizes bipartitebegomoviruses.
These tests were repeated in 1997,including some BGYMV-infectedcommon bean plants from theDominican Republic, a neighbouringCaribbean island (Table 2). The resultsshow that the Cuban BGYMV isolateshad changed their antigenic properties,
whereas the Dominican BGYMVisolates had not. In order to investigatewhether the failure of the MAB-GA todetect the Cuban BGYMV isolates wasdue to a major change in the capsidprotein or putative replicase virusgenes, one of the BGYMV isolates fromQuivicán was partially sequenced.Table 3 shows the result of thisanalysis. These results suggest that theCuban BGYMV isolate has not changedsignificantly in relation to the originalBGYMV isolates from the region andthat the Cuban isolate still can beconsidered a strain of BGYMV.
Table 1. Assay of selected common bean samples with a broad spectrum and specific monoclonalantibodies prepared to Bean golden yellow mosaic virus (BGYMV), Guatemala, 1993.
Sample Plant Locality MAB-BSa MAB-GAb
1 Common bean Velasco, Holguín + -
2 Lima bean Velasco, Holguín + -
3 Common bean (Bonita 11) Tomeguín, Matanzas + -
4 Common bean (Chévere) Tomeguín, Matanzas + -
5 Common bean (Velasco Largo) Tomeguín, Matanzas + -
6 Common bean (BAT 304) Pulido, La Habana + -
7 Common bean CIATc + +
8 Common bean CIATd - -
a. MAB-BS, a broad spectrum monoclonal antibody used to detect bi-partite begomoviruses.b. MAB-GA, a monoclonal antibody used to detect the original Middle American isolates of BGYMV-
Guatemala.c. Sample 7, BGYMV-GA, Guatemala; CIAT, Centro Internacional de Agricultura Tropical.d. Sample 8, healthy common bean.
Table 2. Assay of selected common bean samples with a broad spectrum and specific monoclonalantibodies prepared to Bean golden yellow mosaic virus (BGYMV), Guatemala, 1997.
Sample Plant Locality MAB-BSa MAB-GAb
1 Common bean Quivicán + -
2 Common bean Quivicán + -
3 Common bean Quivicán + -
4 Common bean Dominican Republic + +
5 Common bean Dominican Republic + +
6 Common bean CIATc + +
7 Common bean CIATd - -
a MAB-BS, a broad spectrum monoclonal antibody used to detect bi-partite begomoviruses.b. MAB-GA, a monoclonal antibody used to detect the original Middle American isolates of BGYMV-
Guatemala.c. Sample 6, BGYMV-GA, Guatemala; CIAT, Centro Internacional de Agricultura Tropical.d. Sample 7, healthy common bean.

233
Cuba
With respect to tomato, one of themost affected crops in Cuba, threerandom samples were take in thelocality of Quivicán, Havana Province,and tested for different pathogens(Table 4). As shown, the broadspectrum monoclonal antibody (MAB-BS) used to detect bi-partitebegomoviruses did not detect anybipartite begomovirus in the tomatosamples, although the observation oftwo of the samples in the electronmicroscope revealed the presence ofbegomovirus-like particles. This resultsuggested the need to test for thepresence of TYLCV, a monopartitebegomovirus reported to attack tomatoin Cuba (Polston and Anderson, 1997).This introduced virus is best detectedby polymerase chain reaction (PCR),using specific primers, kindly providedby D. P. Maxwell and M. K. Nakhla, ofthe Department of Plant Pathology,University of Wisconsin. The PCR assaydetected the presence of TYLCV in twoof the tomato samples from Quivican,Cuba.
In 1998, Dr. Gloria González of theInstituto de Investigaciones de SanidadVegetal (INISAV), Havana, Cuba,collected samples from symptomaticpotato plants in Havana Province. Asreported above, potato in Cuba iscolonized also by the B biotype ofB. tabaci. For this test, the viral nucleicacid extracted from affected potatoplants was amplified by PCR using theprimers developed by Rojas (1992). Theamplified region (AV1 and AC1) wascloned and sequenced. Table 5 showsthe results of the comparativenucleotide sequence analysesconducted in reference to otherwhitefly-transmitted begomoviruses.
As can be observed from thiscomparative test, the begomovirusisolated from potato in Cuba is anisolate of Tomato mottle Taino virus(ToMoTV), a virus described in 1997attacking tomato in Cuba (Ramos etal., 1997). This virus was also closelyrelated to Bean dwarf mosaic virus fromColombia (93.3%), Tomato yellow
Table 3. Comparative homologies (%) between a Cuban Bean golden yellow mosaic virus (BGYMV)isolate and other common bean begomoviruses.
ORFa BGYMV BGMV-BRb BDMV-COc
Guatemala Dominican Republic Puerto Rico
AC1 91.5 91.5 89.8 71.9 69.9
AV1 91.3 91.6 91.3 81.6 78.1
a. ORF, open reading frame; AC1, replicase; and AV1, capsid protein.b. BGMV-BR, Bean golden mosaic virus-Brazil.c. BDMV-CO, Bean dwarf mosaic virus-Colombia.
Table 4. Diagnostic assays practiced with three diseased tomato samples from Quivicán, Havana,Cuba.
Sample EMa MAB-BSb PTY1c CMVd PCR-TYLCVe
1 Isometric - - - +
2 Isometric - - - +
3 - - - - -
a. EM, electron microscopy.b. MAB-BS, a broad spectrum monoclonal antibody used to detect bi-partite begomoviruses.c. PTY1, potyviruses.d. CMV, cucumoviruses.e. PCR-TYLCV, DNA amplification of Tomato yellow leaf curl virus.

234
Whiteflies and Whitefly-borne Viruses in the Tropics
mosaic virus (ex-Potato yellow mosaicvirus) from Venezuela, Tomato mottlevirus from Florida, USA, Sida goldenmosaic virus from Costa Rica andAbutilon mosaic virus (South Americanisolate and type species of the clusterin which the Cuban tomato virus istaxonomically placed).
Tabaquina, a concoction of tobaccoleaf residues from the intensive Cubantobacco industry, is another industrialsubproduct used in Cuba to controlwhiteflies in a highly effective manner.Obviously, the well-planned croppingsystems in Cuba also facilitate theimplementation of legal measuresregulating the time of planting forcertain crops which act as reproductivehost for the whitefly B. tabaci(Murguido et al., 1997).
The control of BGYMV in beans, onthe other hand, is effectively carriedout through the use of BGYMV-resistant bean cultivars developed incollaboration with the ProgramaCooperativo Regional de Frijol paraCentro América, México y el Caribe(PROFRIJOL) network and CIAT-Colombia.
Strengthened ResearchCapacity
The two main Cuban institutionscollaborating in the Tropical WhiteflyIntegrated Pest Management (TWF-IPM)Project were INISAV and the Institutode Investigaciones de Hortícolas“Liliana Dimitroya” (LILIANA). DuringPhase I, a third Cuban institute,CENSA, joined the project. Thesub-project’s coordinator for Cuba,Dr. Gloria González of INISAV, receivedfunds to attend the VII Latin Americanand Caribbean Workshop on whitefliesand begomoviruses held on26-30 October 1998, in Nicaragua.Dr. González also visited CIAT,Colombia, for a short but intensivetraining period on the characterizationof whitefly-transmitted viruses.
The TWF-IPM Project contributedto the organization of an InternationalWorkshop on Begomoviruses in theCaribbean Region, held in Quivicán,Cuba on 24-29 November 1997. The
Table 5. Comparative nucleotide sequencehomology (%) between a begomovirusisolated from potato in Cuba andpreviously sequenced whitefly-transmitted begomoviruses.
Region Virusa
ToMoTV ToYMV SiGMV
Coat protein (AV1) 98.5 84.2 85.1
Replicase (AC1) 97.7 83.7 82.4
a. ToMoTV, Tomato mottle Taino virus; ToYMV,Tomato yellow mosaic virus; and SiGMV,Sida golden mosaic virus.
Socio-economic Analysis
Cuba has a unique comparativeadvantage over the rest of the LatinAmerican countries affected bywhiteflies and whitefly-borne viruses,specifically, the limited use of agro-chemicals in this island. Moreover,Cuba has been able to develop andsuccessfully implement biologicalcontrol methods to combat the whiteflyB. tabaci. One of the most effectiveentomopathogens produced has beenVerticillium lecanii. This fungus is verypathogenic to the immature stages ofB. tabaci. The production of biologicalcontrol agents in Cuba constitutes anindustrial activity in at least15 provinces of Cuba, where it is beingapplied on a regular basis tohorticultural crops affected byB. tabaci. In 1996, 170 tons of thefungus compound were produced inCuba to treat 12,565 ha of affectedhorticultural crops. During the peak ofthe 1992 whitefly epidemics, 29,896 hawere treated in Cuba with this bio-control agent (Murguido et al., 1997).

235
Cuba
project provided three keynotespeakers: the coordinator, Dr.Francisco J. Morales; the past co-ordinator of the TWF-IPM Project,Dr. Pamela K. Anderson; and the mainsupport scientist for the Caribbeanregion, Dr. Jane Polston of theUniversity of Florida. Dr. Andersonexplained the nature and objectives ofthe TWF-IPM Project; Dr. Polstondelivered a talk on whitefly-transmittedviruses in the Caribbean Region anddisserted on advanced techniques forthe characterization of begomoviruses;and Dr. Morales gave a talk onbreeding for disease resistance towhitefly-transmitted viruses.
The Whitefly Project also providedfunds for the training at CIAT-Colombiaof a junior scientist, Ms. YenínHernández, of CENSA, Cuba, on themolecular characterization of B. tabacibiotypes. CENSA is one of the leadinginstitutes in Cuba in the area ofEntomology.
Current Status ofWhitefly/Begomovirus
Problems
The whitefly situation in Cubaworsened following the invasion of thenew B biotype of B. tabaci. However,the relatively low use of insecticidesand the production of biological controlagents in Cuba should maintain thewhitefly problem at a manageable level.The Cuban provinces most affected byB. tabaci and the viruses it transmitshave been Pinar del Río and Havana inthe western end of the island andHolguín and Las Tunas in the easternhalf of the country. The whiteflyproblem also has been severe in certainyears in the provinces of Santiago andGuantánamo in the eastern end ofCuba. Other provinces affected havebeen Matanzas, Ciego de Avila,
Granma, Camagüey, Villa Clara,Cienfuegos and Sancti Spiritus.
The provinces of Holguín andLas Tunas are located in one of theregions with less precipitation. Also,these provinces have high averagetemperatures (23.5 ºC) during thewinter season (January), conditionsthat favour the reproduction of B.tabaci. The eastern end of the islandconcentrates the bulk of the tobaccoand vegetable production of Cuba,which probably favours thedevelopment of large whiteflypopulations.
The use of entomopathogens,mainly the fungus Verticillium lecanii,constitutes a highly effective controlmeasure against the proliferation ofB. tabaci. Unfortunately, theproduction of this micro-organism doesnot meet the needs of the intensiveagriculture practiced in Cuba. In thecase of common bean, the use ofvarieties possessing high levels ofresistance to BGYMV remains the mainstrategy for virus control.
References
Blanco, N.; Bencomo, I. 1978. Afluenciade la mosca blanca (Bemisia tabaci)vector del virus del mosaico doradoen plantaciones de frijol. Cienciasde la Agricultura 2:39-45.
Brown, J. K. 1994. Current status ofBemisia tabaci as a plant pest andvirus vector in agro-ecosystemsworldwide. FAO Plant Protect. Bull.42:3-32.
Cancino, M.; Abouzid, A. M.; Morales, F.J.; Purcifull, D. E.; Polston J. E.;Hiebert, E. 1995. Generation andcharacterization of threemonoclonal antibodies useful indetecting and distinguishing Beangolden mosaic virus isolates.Phytopathology 85:484-490.

236
Whiteflies and Whitefly-borne Viruses in the Tropics
Murguido, C.; Vázquez, L.; Gómez, O.;Mateo, A. 1997. Informe sobre laproblemática mosca blanca-geminivirus. Instituto deInvestigaciones de Sanidad Vegetal(INISAV), Boletín Técnico no. 5,La Habana, CU. 13 p.
Polston, J. E.; Anderson, P. K. 1997. Theemergence of whitefly-transmittedgeminiviruses in tomato in thewestern hemisphere. Plant Dis. 81:1358-1369.
Ramos, P. L.; Guerra, O.; Peral, P.;Oramas, P.; Guevara, R. G.; Rivera-Bustamante, R. 1997. Taino tomatomottle virus, a new bipartitegeminivirus from Cuba. Plant Dis.81:1095.
Rojas, M. R. 1992. Detection andcharacterization of whitefly-transmitted geminiviruses by theuse of polymerase chain reaction.University of Wisconsin, Madison,USA.

237
Dominican Republic
CHAPTER 3.11
Dominican RepublicFrancisco Morales*, Emigdio Gómez**,
Augusto Villar** and Julio Nin***
Introduction
Geographical context
The Dominican Republic comprises theeastern two-thirds of the island ofHispaniola. The country is generallymountainous, with the most prominentrange being the Central Highlands(elevation up to 3175 m), the highestpoint in the West Indies. The ConstanzaValley is a highly developed agriculturalarea located in the Central Highlandsand is currently planted to high-valuecrops such as garlic (Allium sativum L.),onion (Allium cepa L.) and otherhorticultural crops. The Cibao Valley, inthe north-west, is one of the most fertileagricultural areas in the country, whererice (Oryza sativa L.), maize (Zea maysL.), common bean (Phaseolus vulgarisL.), tobacco (Nicotiana tabacum L.) andcoffee (Coffea arabica L.) are produced.The western part of the country is drybut rivers provide irrigation. San Juande la Maguana is the main bean areaand the Azua Valley is the mainhorticultural area in south-westernDominican Republic. The annual meantemperature is 25 °C and precipitationis 1346 mm. Tropical storms and
hurricanes are a major threat every year(West and Augelli, 1977). Figure 1 showsthe main agricultural regions affected bywhitefly-transmitted begomoviruses.
The emergence of Bemisia tabacias a pest and virus vector
Bemisia tabaci (Gennadius) was firstobserved attacking common bean in1975. However, the presence of Beangolden yellow mosaic virus (BGYMV) inthe Dominican Republic had beennoticed as early as the late 1960s(Schieber, 1970). By 1988, other cropssuch as tomato (Lycopersiconesculentum Mill.), melon (Cucumis meloL.), eggplant (Solanum melongena L.),cucumber (Cucumis sativus L. var.sativus) and watermelon (Citrulluslanatus [Thunb.] Matsum. & Nakai) wereunder attack from B. tabaci. In 1991,biotype B of B. tabaci made itsappearance in the main horticulturalregions of the Dominican Republic. Atthe same time, the first begomovirusfrom the Old World, Tomato yellow leafcurl virus (TYLCV) was irresponsiblyintroduced into the Dominican Republicand the Americas. Following theintroduction of a new and moreaggressive biotype of the whitefly vectorand an exotic begomovirus, crops suchas tomato and melon practicallydisappeared, and the tomato processingplants had to close down in certainareas.
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** Programa Nacional de Manejo Integrado dePlagas (MIP), Secretaría del Estado deAgricultura (SEA), Dominican Republic.
*** Programa de Mejoramiento, SEA, DominicanRepublic.

238
Whiteflies and Whitefly-borne Viruses in the Tropics
Advances in BiologicalResearch
The first begomovirus of economicimportance to be characterized at themolecular level in the DominicanRepublic was BGYMV. This virus wasisolated in the early 1990s by thesenior author and later shown to bemolecularly and serologically related tothe Guatemalan isolate of BGYMV(Faria et al., 1994; Cancino et al.,1995). The BGYMV isolate from theDominican Republic (San Juan de laMaguana) proved to be similar toBGYMV isolates from the Caribbean(Puerto Rica) and Central America(Guatemala). The BGYMV isolate fromthe Dominican Republic was shown tobe a different species when comparedwith Bean golden mosaic virus (BGMV)from Brazil. BGYMV affects over20,000 ha of common bean in theDominican Republic each year,particularly in the south-western valleyof San Juan de la Maguana. The
presence of a begomovirus infectingtomatoes in the Dominican Republicwas first noticed in 1992 andsubsequently identified as TYLCV fromIsrael (Polston et al., 1994; Polston andAnderson, 1997). The introduction ofan exotic begomovirus from the OldWorld created a major pandemicthroughout the Caribbean at a cost totomato growers and tomatoprocessing plants worth millions ofUS dollars in yield/export losses(Dupuy, 1998).
At the beginning of this project, inApril 1997, common bean sampleswere collected in San Juan de laMaguana and tomato samples in Azua,in south-western Dominican Republic.The common bean samples reactedwith the broad-spectrum monoclonalantibody 3F7 as well as with themonoclonal antibody prepared to theGuatemalan isolate of BGYMV.However, the tomato samples did notreact against the broad-spectrum
Valverde
Salcedo
Azua
Espaillat
Dominican Republic
Figure 1. The main agricultural regions affected by whitefly-transmitted begomoviruses, DominicanRepublic.
ElíasPina
Barahona La Romana
LaAltagracia
El Seibo
LaRomana
San Pedro deMacoris
DistritoNacional
SanCristóbal
Peravia
Monte Plata HatoMayor
MonseñorNoube
La Vega
San Juan
Santiago Duarte Samana
SánchezRamírez
MaríaTrinidadSánchez
Puerto PlataMonteCristi
Dajabon SantiagoRodríguez
Baoruco

239
Dominican Republic
monoclonal antibody (MAB) in thesetests. These tomato samples were thenassayed by polymerase chain reaction(PCR) using two sets of primers(PTYIRV21/PTYIRC287 and PTYIRV21/RTYC2C1814) to TYLCV provided byDr. M. Nakhla, Plant PathologyDepartment of the University ofWisconsin, Madison, WI. These primersamplified two fragments of Mr 287 and1793 pb, respectively, demonstratingthe presence of TYLCV in the tomatoplants sampled in Azua.
In February 1998, six more tomatosamples from Azua and one from thenorthern locality of Santiago wereassayed with similar results (Table 1),demonstrating the endemic nature ofTYLCV in the Dominican Republic. Thethree tomato samples (9-11) from theCentro de Investigaciones Aplicadas aZonas Áridas (CIAZA), Azua,corresponded to TYLCV-tolerant tomatovarieties (Gem-Pear, Gem-Star andUC82), which produce acceptably inthis locality.
The common bean samples fromthe valley of San Juan de la Maguanabehaved as expected for the BGYMVisolates previously detected in thislocality. The presence of BGYMV in theConstanza Valley is interesting,because it is located at altitudesbetween 1000 and 1100 m, above thealtitudinal range (0-950 m) at whichB. tabaci is usually a problem.However, the incidence of the virus wasmoderate in the Constanza Valley.
Socio-economic Analysis
The agriculture of the DominicanRepublic was described in the 1970s asa mixture of traditional and moderncropping systems. Traditionalagriculture was equivalent to“subsistence” agriculture and includedcrops such as common bean, cassava(Manihot esculenta Crantz), rice,sweetpotato (Ipomoea batatas [L.]Lam.), pigeon pea (Cajanus cajan [L.]Millsp.) and taro (Colocasia esculenta
Table 1. Analyses of selected plant samples from the Dominican Republic.
Sample Plant Localitya MAB-BSb MAB-GAc PTY1d TYLCVe
1 Common bean San Juan + + - nt
2 Common bean San Juan - - - -
3 Common bean Constanza + + - nt
4 Common bean Constanza + + - nt
5 Garlic Constanza nt nt + nt
6 Tomato Azua - nt nt +
7 Tomato Azua - nt nt +
8 Tomato Azua - nt nt +
9 Tomato CIAZA-Azua - nt nt +
10 Tomato CIAZA-Azua - nt nt +
11 Tomato CIAZA-Azua - nt nt +
12 Tomato Santiago - nt nt +
13 Tomato Santiago - nt nt +
a. CIAZA, Centro de Investigaciones Aplicadas a Zonas Áridas.b. MAB-BS, a broad spectrum monoclonal antibody used to detect bi-partite begomoviruses.c. MAB-GA, a monoclonal antibody used to detect the original Middle American isolates of Bean
golden yellow mosaic virus-Guatemala; and nt, no tests for these samples.d. PTYI, monoclonal antibodies to detect potyviruses; and nt, no tests for these samples.e. TYLCV, polymerase chain reaction detection of Tomato yellow leaf curl virus; and nt, no test for
these samples.

240
Whiteflies and Whitefly-borne Viruses in the Tropics
[L.] Schott). Modern agriculture wasequivalent to the “plantation”agriculture going back to colonialtimes, represented mostly by theextensive cultivation of sugarcane(Saccharum officinarum L.). In 1970,sugarcane was the main agro-industryof the country, occupying over150,000 ha.
In 1966, a Land ReformProgramme began to be implementedin the Dominican Republic. Large-scalefarming in the form of co-operativesbegan to take place with the support ofthe government, the financialcontribution of international agencies—for example the Agency forInternational Development (AID)—andthe technical support of thirdcountries. The Sisal Project, forinstance, was created at that time withthe help of the Government of Israel.The objective of the project was toproduce tomato under irrigation in thevalley of Azua. The irrigation systemwas initiated in 1970 with theconstruction of the Sabana Yegua Damto irrigate some 18,000 ha. Thus, theDominican Republic was one of thefirst countries in Latin America to lookinto the possibility of producing non-traditional, high-value crops for exportsuch as tomato, chilli (Capsicumannuum L. var. annuum), melon,watermelon and eggplant. In 1970,1600 ha of tomato were planted in theDominican Republic. In the late 1990s,the tomato industry covered 8000 ha,employed over 6500 small-scalefarmers (farms averaging less than2.5 ha) and created jobs for90,000 people per crop cycle (Dupuy,1998).
The arrival of TYLCV from Israeland biotype B of B. tabaci brought thethriving tomato industry to a halt,causing economic losses in excess of30 million US dollars. Table 2 shows
the yield losses in the production ofindustrial tomato for the two mainproduction areas, Azua and Cibao,between 1988 and 1995 (Alvarez andAbud-Antún, 1995). This crisis wasmanaged through the adoption of legalmeasures that included a 3-monthcrop-free period to break the cycle ofthe whitefly vector. Additionally, virus-resistant tomato cultivars wereintroduced.
Table 2. Yield losses (%) caused by whitefly/begomovirus damage to industrialtomatoes in the main tomatoproducing regions of the DominicanRepublic.
Cropping Regionseason Azua Cibao
1988-89 25 5
1989-90 45 10
1990-91 40 15
1991-92 30 15
1992-93 80 80
1993-94 95 50
1994-95 20 15
Following the implementation oflegal measures against B. tabaci, themelon industry made a comeback. Thecase of common bean was notpromising, because this crop was beingdisplaced from the traditional bean-growing areas such as the valley of SanJuan de la Maguana by other cropswithout whitefly problems and inhigher demand, including cassava andsweetpotato. In 1984, there were63,000 ha of red-seeded bean in thecountry and in 1994 the area plantedto common bean is only 36,000 ha,almost half of the area planted15 years ago. As a result, the countryhas had to import common bean,usually grain types of little demand inthe country, considering that red-seeded bean is the main food staple inthe diet of the Dominicans (Fundaciónde Desarrollo Agropecuario, 1995).

241
Dominican Republic
Strengthened ResearchCapacity
Considering that the two mainhorticultural regions affected bywhiteflies and whitefly-transmittedgeminiviruses in the DominicanRepublic are Azua and Cibao, twoAgricultural Engineers were contactedto collaborate in this project. They wereAugusto Villar of the Secretaría delEstado de Agricultura (SEA)-CIAZA,Azua, and Emigdio Gómez of SEA-Programa Nacional de ManejoIntegrado de Plagas (MIP), N-N.W.Cibao. These national programmescientists were brought to the CentroInternacional de Agricultura Tropical(CIAT) for training in the identificationof B. tabaci biotypes and diagnosis ofviruses affecting crops of socio-economic importance in theirrespective agricultural regions. Table 3shows the results obtained by thesetwo researchers at CIAT with thewhitefly samples they collected in theDominican Republic.
The two researchers were trainedalso in the identification ofgeminiviruses and other plant virusesusing the enzyme-linkedimmunosorbent assay (ELISA)technique in conjunction with the
utilization of monoclonal andpolyclonal antibodies and financed bythe project to attend the InternationalWorkshop on Bemisia andGeminiviruses, held in San Juan,Puerto Rico, June 1998. Here theypresented the novel approach adoptedin the Dominican Republic to controlwhitefly/begomovirus damage byregulatory measures designed to breakthe continuous populational cycle ofB. tabaci in tropical environments.
The project also supported theactivities of several researchers onhorticultural crops in the ConstanzaValley, in the central region of thecountry, in collaboration with thehorticultural research network, La RedColaborativa de Investigación yDesarrollo de Hortalizas para AméricaCentral, Panamá y RepúblicaDominicana (REDCAHOR). Twoconferences were delivered on theTropical Whitefly Integrated PestManagement (TWF-IPM) Project and onthe breeding and selection ofhorticultural crops for their resistanceto whitefly-transmitted begomoviruses.The CIAT Bean Project is collaboratingwith the Dominican Republic to breedDominican bean cultivars, mainly thePompadour types, for resistance toBGYMV.
Table 3. Results of the analyses of Bemisia spp. biotypes found in the Dominican Republic ondifferent economically important crops.
Locality Crop No. samples Whitefly
Azua-CIAZAa Eggplant 3 Bemisia tuberculata
Azua-Estebanía Tomato 9 Bemisia tabaci-B
Barahona Eggplant 3 Bemisia tuberculata
Azua-Arroyo Salado Eggplant 3 Bemisia tuberculata
San José de Ocoa Tomato 6 Bemisia tabaci-B
Santiago Okra 3 Bemisia tabaci-B
Santiago Cucumber 3 Bemisia tabaci-B
Santiago Eggplant 3 Bemisia tabaci-B
Montecristi Melon 6 Bemisia tabaci-B
Santiago Tomato 3 Bemisia tabaci-B
Santiago Okra 3 Bemisia tabaci-B
a. CIAZA, Centro de Investigaciones Aplicadas a Zonas Áridas.

242
Whiteflies and Whitefly-borne Viruses in the Tropics
Current Situation ofWhitefly/Geminivirus
Problems
The Dominican Republic constitutes aunique example of a country thatimplemented legal measures to minimizeyield losses due to the activity of B.tabaci both as a pest and virus vector.Nevertheless, pesticide use remainshigh in most agricultural regions of thecountry, and the existing croppingsystems favour the reproduction ofB. tabaci. Currently, the situation in thelowlands and highlands remains underpartial control, and some crops thathad been taken out of production suchas melon are being planted again.Another crop and good host to thewhitefly B. tabaci, tobacco, also hasbeen increasing its area in the country.A recent viral epidemic in tobacco in theCibao region was diagnosed at CIAT asbeing caused by Tobacco mosaic virus.
In the case of tomato, thecultivation of varieties resistant toTYLCV should contribute to therecovery of the production of industrialtomato plantings. In the case ofcommon bean, however, we havenoticed a marked reduction in the areaplanted, because of the susceptibility ofmost local varieties to BGYMV.Moreover, TYLCV has been reported toattack common bean in other parts ofthe world (Navas-Castillo et al., 1999).
References
Alvarez, P. A.; Abud-Antún, A. 1995.Reporte de la República Dominicana.CEIBA 36:39-47.
Cancino, M.; Abouzid, A. M.; Morales, F.J.; Purcifull, D. E.; Poston, J. E.;Hiebert, E. 1995. Generation andcharacterization of three monoclonalantibodies useful in detecting anddistinguishing Bean golden mosaicvirus isolates. Phytopathology85:484-490.
Dupuy, J. 1998. Dealing with whitefliesand geminiviruses in the DominicanRepublic. Special Report. BarcelóIndustries, Santo Domingo, DO.12 p.
Faria, J. C.; Gilbertson, R. L.; Hanson, S.F.; Morales, F. J.; Ahlquist, P.;Loniello, A. O.; Maxwell, D. P. 1994.Bean golden mosaic geminivirustype II isolates from the DominicanRepublic and Guatemala:Nucleotide sequences, infectiouspseudorecombinants, andphylogenetic relationships.Phytopathology 84:321-329.
Fundación de Desarrollo Agropecuario.1995. Cultivo de habichuela.Boletín Técnico no. 2. SantoDomingo, DO. 38 p.
Navas-Castillo, J.; Sánchez-Campos, S.;Díaz, J. A. 1999. Tomato yellow leafcurl virus-Is causes a novel diseaseof common bean and severeepidemics in tomato in Spain. PlantDis. 83:29-32.
Polston, J. E.; Bois, D.; Serra, C. A.;Concepción, S. 1994. First report ofa tomato yellow leaf curl-likegeminivirus in the westernhemisphere. Plant Dis. 78:831.
Polston, J. E.; Anderson, P. K. 1997. Theemergence of whitefly-transmittedgeminiviruses in tomato in thewestern hemisphere. Plant Dis.81:1358-1369.
Schieber, E. 1970. Enfermedades delfrijol (Phaseolus vulgaris L.) en laRepública Dominicana. Turrialba20:20-23.
West, R. C.; Augelli, J. P. 1977. MiddleAmerica: Its lands and peoples.Second edition. Prentice-Hall, NJ,USA. 494 p.

243
Reproductive Crop Hosts of Bemisia tabaci
CHAPTER 3.12
Reproductive Crop Hosts of Bemisiatabaci (Gennadius) in Latin Americaand the Caribbean
Pamela K. Anderson*, Avas Hamon**,Pilar Hernández*** and Jon Martinψ
Introduction
The sweetpotato whitefly Bemisia tabaci(Gennadius) is a vector of at least30 begomoviruses (Geminiviridae:Begomovirus) in Latin America and theCaribbean (Morales and Anderson,2001). As is often the case for insectvector-host relations, many of thecrops that begomoviruses affect are notreproductive hosts for B. tabaci. A firstcritical step in vector entomology, then,is to answer the question. “Where isB. tabaci reproducing?”
Crop biomass can provide excellentbreeding and feeding sources and thusgive rise to large populations ofB. tabaci (Byrne et al., 1991). We arguehere that the first step tounderstanding regional populationdynamics as the basis for area-widemanagement is to elucidate whichcrops B. tabaci utilizes as reproductivehosts and define their relativeimportance.
Crop Hosts of Bemisiatabaci
The most reliable sources of hostplant records for B. tabaci arepublished taxonomic reports andunpublished records from museumcollections. The first author hasreviewed the white and grey literatureas well as the arthropod collectionsfrom the United States NationalMuseum (USNM), the Florida StateCollection of Arthropods (FSCA), theBritish Museum of Natural History(BMNH), and the Escuela AgrícolaPanamericana (EAP) in Honduras. TheEAP (1992) references areunpublished records from LatinAmerica, collected and identified byR. Caballero as the basis fordevelopment of several taxonomickeys (Caballero, 1992). Table 1 alsorecords B. tabaci host recordsgenerated by the national program(national agricultural researchsystems [NARS]) teams as part of theextensive diagnostic surveys forPhase 1 of the Tropical WhiteflyIntegrated Pest Management Project(TWF-IPM). P. Hernández identifiedthese specimens and A. Hamonverified them. Verified specimens aredeposited in the insect museum atthe Centro Internacional deAgricultura Tropical (CIAT) in Cali,Colombia, and the FSCA inGainesville, Florida, USA.
* Former coordinator for Phase 1 of theTropical Whitefly IPM Project; currentlyResearch Director at the CentroInternacional de la Papa (CIP), Lima, Peru.
** Florida State Collection of Arthropods(FSCA), Gainesville, Florida, USA.
*** Centro Internacional de AgriculturaTropical (CIAT), Cali, Colombia.
ψ British Museum of Natural History, London,UK.

244
Whiteflies and Whitefly-borne Viruses in the Tropics
Based upon the reliable publishedtaxonomic reports, unpublishedmuseum records and the verifiedspecimen collected during Phase 1 ofthe TWF-IPM, we conclude that
Table 1. Known crop hosts of Bemisia tabaci in Latin America and the Caribbean.
Crop Country Referencea
Latin name Common name
Abelmoschus esculentus (L.) Moench Okra Cuba Vázquez et al. (1996)[= Hibiscus esculentus L.]
Dom. Rep. TWF-IPM ProjectEl Salvador USNM (1967), Lipes
(1968), WF-IPMProject
Guatemala TWF-IPM ProjectTrinidad BMNH (1989)Venezuela Arnal et al. (1993b)
Annona muricata L. Soursop Cuba Vázquez et al. (1996)Arachis hypogaea L. Peanut Cuba Vázquez et al. (1996)
Venezuela Arnal et al. (1993b)Beta vulgaris L. Beet Cuba Vázquez et al. (1996)Beta vulgaris subsp cicla (L.) W. Koch Swiss chard Cuba Vázquez et al. (1996)Brasica rapas L. subsp. rapa Turnip Cuba Vázquez et al. (1996)Brassica oleracea L. var. capitata L. Cabbage Cuba Vázquez et al. (1995)Brassica oleracea L. var. gemminifera DC Brussel sprouts Venezuela Arnal et al. (1993b)Brassica oleracea L. var. gongylodes L. Kohl-rabi Cuba Vázquez et al. (1995)Cajanas cajan (L.) Millsp. Pigeon pea Cuba Vázquez et al. (1996)Canavalia ensiformis (L.) DC. Jack bean, Cuba Vázquez et al. (1996)
sword beanCapsicum annuum L. var. annuum Bell pepper Belize EAP (1992)
Costa Rica TWF-IPM ProjectCuba Vázquez et al. (1996)Honduras EAP (1992)Panama TWF-IPM ProjectVenezuela Arnal et al. (1993b)
Capsicum spp. Chilli peppers Costa Rica TWF-IPM ProjectCuba Vázquez et al. (1996)El Salvador TWF-IPM ProjectGuatemala TWF-IPM ProjectHonduras TWF-IPM ProjectNicaragua EAP (1992)Venezuela Arnal et al. (1993b)
Carica papaya L. Papaya Cuba Vázquez et al. (1996)Guatemala TWF-IPM Project
Cicer arietinum L. Chick-pea Cuba Vázquez et al. (1996)Citrullus lanatus (Thunb.) Matsum. & Nakai Watermelon Belize EAP (1992)[= Citrullus vulgaris Schrad.] Colombia TWF-IPM Project
Costa Rica TWF-IPM ProjectCuba Vázquez et al. (1995)Ecuador TWF-IPM ProjectEl Salvador TWF-IPM ProjectGuatemala TWF-IPM ProjectHonduras EAP (1992)Panama TWF-IPM ProjectVenezuela Arnal et al. (1993b)
Cucumis melo L. Melon Belize EAP (1992)Colombia TWF-IPM ProjectCosta Rica TWF-IPM ProjectDom. Rep. TWF-IPM ProjectEcuador TWF-IPM ProjectEl Salvador TWF-IPM Project
(Continued)
B. tabaci is utilizing at least50 cultivated species as reproductivehosts in Latin America and theCaribbean (Table 1).

245
Reproductive Crop Hosts of Bemisia tabaci
Guatemala EAP (1992),TWF-IPM Project
Honduras EAP (1992)Venezuela Arnal et al. (1993a;
1993b), TWF-IPMProject
Cucumis sativus L. Cucumber Colombia TWF-IPM ProjectCosta Rica TWF-IPM ProjectCuba Vázquez et al. (1996)Dom. Rep. TWF-IPM ProjectEl Salvador TWF-IPM ProjectGuatemala TWF-IPM ProjectHonduras EAP (1992),
TWF-IPM ProjectVenezuela Arnal et al. (1993b),
TWF-IPM ProjectCucurbita argyrosperma C. Huber Pipián El Salvador TWF-IPM Project subsp. argyrospermaCucurbita maxima Duch. Ex Lam. Squash Cuba Vázquez et al. (1996)
Venezuela Arnal et al. (1993b),TWF-IPM Project
Cucurbita moschata (Duch. ex Lam.) Ayote Colombia TWF-IPM Project Duch. ex Poir. El Salvador TWF-IPM Project
Honduras EAP (1992)Panama TWF-IPM Project
Cucurbita pepo L. Squash Ecuador TWF-IPM ProjectNicaragua USNM (1958)Venezuela Arnal et al. (1993b)
Euphorbia pulcherrima Willd. Ex Klotzch Poinsettia El Salvador USNM (1967)Fernaldia pandurata (A. DC.) Woodson Loroco El Salvador TWF-IPM ProjectGlycine max (L.) Merr. Soybean Argentina TWF-IPM Project
Brazil BMNH (1974)Colombia EAP (1992),
TWF-IPM ProjectCuba Vázquez et al. (1996)Ecuador TWF-IPM ProjectHonduras EAP (1992)Venezuela Arnal et al. (1993a;
1993b)Gossypium hirsutum L. Cotton Argentina Viscarrret and Botto
(1996)Barbados BMNH (1982)Colombia EAP (1992),
TWF-IPM ProjectEl Salvador USNM (1964; 1965;
1967; 1969)Guatemala EAP (1992)Honduras USNM (1966)Nicaragua USNM (1951; 1965;
1978)Venezuela Arnal et al. (1993b)
Helianthus annuus L. Sunflower Cuba Vázquez et al. (1996)Venezuela Arnal et al. (1993b)
Ipomoea batatas (L.) Lam. Sweetpotato Cuba Vázquez et al. (1996)Venezuela Arnal et al. (1993b)
Lactuca sativa L. Lettuce Cuba Vázquez et al. (1996)Venezuela Arnal et al. (1993b)
Lycopersicon esculentum Mill. Tomato Belize EAP (1992)Colombia EAP (1992),
TWF-IPM Project
Table 1. (Continued.)
Crop Country Referencea
Latin name Common name
(Continued)

246
Whiteflies and Whitefly-borne Viruses in the Tropics
Costa Rica TWF-IPM ProjectCuba Vázquez et al. (1996)Dom. Rep. EAP (1992),
TWF-IPM ProjectEl Salvador USNM (1967),
TWF-IPM ProjectGuatemala EAP (1992),
TWF-IPM ProjectHonduras EAP (1992),
TWF-IPM ProjectMexico BMNH (1992)Nicaragua USNM (1952),
TWF-IPM ProjectPanama TWF-IPM ProjectPuerto Rico BMNH (1988)Trinidad BMNH (1990)Venezuela Arnal et al. (1993a;
1993b), TWF-IPMProject
Mangifera indica L. Mango Cuba Vázquez et al. (1996)Manihot esculenta Crantz Cassava Colombia EAP (1992), TWF-IPM
ProjectCuba Vázquez et al. (1996)Dom. Rep. TWF-IPM ProjectEcuador TWF-IPM ProjectVenezuela Arnal et al. (1993b)
Musa × paradisiacal L. Banana/plantain Cuba Vázquez et al. (1996)Venezuela Arnal et al. (1993b)
Nicotiana tabacum L. Tobacco Cuba Vázquez et al. (1996)Guatemala TWF-IPM ProjectNicaragua USNM (1951)Venezuela Arnal et al. (1993b)
Ocimum basilicum L. Basil Cuba Vázquez et al. (1996)Passiflora edulis Sims Passionfruit Venezuela Arnal et al. (1993b)Persea americana Mill. Avocado Cuba Vázquez et al. (1996)
Guatemala USNM (1954)Venezuela Arnal et al. (1993b)
Phaseolus acutifolius A. Gray Tepary bean El Salvador TWF-IPM Project var. acutifoliusPhaseolus lunatus L. Lima bean Brazil BMNH (1975)
Venezuela Arnal et al. (1993b)Phaseolus vulgaris L. Common bean Argentina USNM (1982),
TWF-IPM ProjectColombia USNM (1974),
TWF-IPM ProjectCosta Rica TWF-IPM ProjectCuba Vázquez et al. (1996)Dom. Rep. BMNH (1979);
TWF-IPM ProjectEcuador TWF-IPM ProjectEl Salvador USNM (1964);
TWF-IPM ProjectPhaseolus vulgaris L. Common bean Guatemala USNM (1973);
TWF-IPM ProjectHonduras EAP (1992);
TWF-IPM ProjectNicaragua USNM (1974);
TWF-IPM ProjectPanama TWF-IPM ProjectVenezuela Arnal et al. (1993a;
1993b)
Table 1. (Continued.)
Crop Country Referencea
Latin name Common name
(Continued)

247
Reproductive Crop Hosts of Bemisia tabaci
Psidium guajava L. Guava Cuba Vázquez et al. (1996)Nicaragua USNM (1978)
Raphanus sativus L. Radish Cuba Vázquez et al. (1995)Sesamum indicum L. Sesame Cuba Vázquez et al. (1996)
Venezuela Arnal et al. (1993b)Solanum melongena L. Eggplant Colombia TWF-IPM Project
Cuba Vázquez et al. (1996)Dom. Rep. EAP (1992),
TWF-IPM ProjectEcuador TWF-IPM ProjectGuatemala TWF-IPM ProjectMexico TWF-IPM ProjectPuerto Rico BMNH (1987)Trinidad BMNH (1988)Venezuela Arnal et al. (1993b)
Solanum tuberosum L. Potato Argentina USNM (1982)Cuba Vázquez et al. (1996)Honduras EAP (1992)Nicaragua EAP (1992)Venezuela Arnal et al. (1993a;
1993b)Spondias purpurea L. Spanish plum Cuba Vázquez et al. (1996)Vicia faba L. Broad bean Cuba Vázquez et al. (1996)Vigna unguiculata (L.) Walp. Cowpea Cuba Vázquez et al. (1996)
Venezuela Arnal et al. (1993b)Vigna unguiculata subsp. Yard-long bean Cuba Vázquez et al. (1996) sesquipedalis (L.) Verdc.Xanthosoma sagittifolium (L.) Schott Malanga Cuba Vázquez et al. (1996)
a. For references, see list at end of chapter.BMNH, British Museum of Natural History, data from various dates;EAP, Escuela Agrícola Panamericana, Honduras, unpublished data;USNM, United States National Museum, data from various dates;TWF-IPM Project, Tropical Whitefly-Integrated Pest Management Project.
Table 1. (Continued.)
Crop Country Referencea
Latin name Common name
Relative Importance ofBemisia tabaci on
Different Crop Hosts
B. tabaci is cosmopolitan indistribution, highly polyphagous andcharacterized by intercrop movement(Butler et al., 1986). Based onidentified museum specimens, Moundand Halsey (1978) listed 317 plantspecies as host plants for B. tabaci,worldwide. However, sufficientempirical and experimental evidenceexists to indicate B. tabaci populationsdo not reproduce equally among allpotential crop hosts in any givengeographical region. Rather, B. tabaciappears to develop strong hostassociations, resulting in host races. A
host race is a population of a speciesthat is partially reproductivelyisolated from other non-specific,sympatric sister populations as adirect consequence of adaptation to aspecific host or habitat (Diehl andBush, 1984; Bush, 1994). Host racesare one category of biotypes, that is,two or more morphologically similaror indistinguishable taxa that differfrom one another in a biologicallysignificant way such as hostpreference, emergence time or someother ecological or behavioral trait(Bush, 1994).
In any given geographical area,beyond more precisely defining therange of reproductive crop hosts, it isnecessary to define the relative

248
Whiteflies and Whitefly-borne Viruses in the Tropics
importance, or reproductive potential,of the host plants, that is, to quantifythe average number of B. tabaciproduced per unit area. For example,intensive surveys carried out inVenezuela (Arnal et al., 1993a; 1993b)and Cuba (Vázquez et al., 1995; 1996)each identified 27 and 39reproductivecrop hosts for B. tabaci respectively(Table 1). The next step is to identifythe relative importance of each cropas a reproductive host.
Caballero and Nolasco (1995)made 289 collections of B. tabaci fromeconomically important crop hosts inBelize, Costa Rica, El Salvador,Guatemala, Honduras, Nicaragua andPanama. They quantified B. tabaciabundance for each crop utilizingB. tabaci density/leaf as an indicatorof abundance (Table 2).
However, comparisons of whiteflyabundance on different crops isproblematic because of variation inarchitecture, growth habits, foliagedensity and distribution of B. tabaciwithin the plant, among differentcrops and among different varietieswithin the same crop. Density
measures on a per ground unit basisare necessary for meaningfulcomparisons. Recently S. Naranjo andL. Cañas (United States Department ofAgriculture [USDA]- AgriculturalResearch Service [ARS]) havedeveloped a protocol to quantifyB. tabaci abundance for a range ofcrop plants within the same area on aper ground unit basis. This protocol iscurrently being tested in Arizona(Cañas, Naranjo and Ellis, inpreparation) and in El Salvador(Serrano and Anderson, inpreparation). Such quantification willallow us to compare the relativeimportance of B. tabaci reproductivehosts within and among zones in acountry, and among differentcountries; to monitor fluctuations inthe B. tabaci populations on the samecrop in the same zone from year toyear; and to identify newly emergingreproductive hosts for B. tabaci.
We cannot effectively implementcrop protection tactics aimed atsource reduction within an area-widemanagement program if we cannotprioritize and target the principalreproductive sources for B. tabaci.
Table 2. Relative importance of Bemisia tabaci crop hosts in Central America, based on leaf samples.
Rankinga Crop Common name
5 Gossypium hirsutum L. Cotton
Citrillus lanatus (Thumb.) Matsum. & Nakai Watermelon
Cucurbita pepo L. Squash
4 Lycopersicon esculentum Mill. Tomato
Cucmus melo L. Melon
Capsicum annuum L. Sweet pepper
3 Nicotiana tabacum L. Tobacco
Solanum melongena L. Eggplant
Phaseolus vulgaris L. Common bean
2 Glycine max L. (Merr.) Soybean
a. Rank 5, up to 100 nymphs per leaf, in all countries surveyed; rank 4, up to 100 nymphs per leaf,in certain countries; rank 3, up to 50 nymphs per leaf, in all countries; rank 2, up to 20 nymphsper leaf, in all countries; and rank 1, less than 10 nymphs per leaf, in all countries.

249
Reproductive Crop Hosts of Bemisia tabaci
Conclusions andRecommendations
After more than a decade of epidemicevents in Mexico, Central America andthe Caribbean, we still do not have aclear enough understanding of therange of reproductive crop hosts forB. tabaci. Nor do we understand therelative importance of the B. tabacireproductive hosts within a zone, forpurposes of area-wide management.The question, “where is B. tabacireproducing?”, continues to be a basicresearch priority for vector entomologyin the region.
References
Arnal, E.; Russell, L. M.; Debrot, E.;Ramos, F.; Cermeli, M.; Marcano,R.; Montagne, A. 1993a. Lista demoscas blancas (Homoptera:Aleyrodidae) y sus plantashospederas en Venezuela. Fla.Entomol. 76:365-381.
Arnal, E.; Ramos, F.; Debrot, E. 1993b.Plantas hospederas de la moscablanca Bemisia tabaci (Gennadius)en Venezuela. Agron. Trop.(Venezuela) 43:267-286.
Bush, G. L. 1994. Bemisia tabaci: Biotypeor species complex? In: Rojas, A.;Varela, G.; Valle, N.; Chavarria, J.A. (eds.). Memoria, II TallerLatinoamericano y del Caribe sobremoscas blancas y geminivirus,20-22 octubre 1993, OrganismoInternacional Regional de SanidadAgropecuaria (OIRSA), Managua,NI. p. 17-24.
Butler, G. D. Jr.; Henneberry, T. J.;Hutchinson, W. D. 1986. Biology,sampling and population dynamicsof Bemisia tabaci. Agric. Zool. Rev.1:167-195.
Byrne, D. N.; Bellows, T. S.; Parrella, M.P. 1991. Whiteflies in agriculturalsystems. In: Gerling, D. (ed.).Whiteflies: Their bionomics, peststatus and management. Intercept,Andover, Hants, GB. p. 227-261.
Caballero, R. 1992. Whiteflies(Homoptera: Aleyrodidae) fromCentral America and Colombiaincluding slide-mounted pupal andfield keys for identification, fieldcharacteristics, hosts, disbribution,natural enemies, and economicimportance. M.Sc. thesis, KansasState University, USA. 201 p.
Caballero, R.; Nolasco, C. 1995.Determinación de especies demoscas blancas, biotipos deBemisia tabaci (Genn.), malezashospederas y geminivirus.Programa regional de reforzamientode la investigación agronómicasobre los granos básicos en CentroAmérica (ITR 9305-0793). 11 p.
Diehl, S. R.; Bush, G. 1984. Anevolutionary and appliedperspective of insect biotypes. Ann.Rev. Entomol. 29:471-504.
Lipes, J. E. 1968. El Salvador:Occurrence of the banded-wingwhitefly, Trialeurodes abutilonea.FAO Plant Prot. Bull. 16:32.
Morales, F. J.; Anderson, P. K. 2001. Theemergence and dissemination ofwhitefly-transmitted geminivirusesin Latin America. Arch. Virol.146 (3):415-441.
Mound, L. A.; Halsey, S. H. 1978.Whitefly of the world: A systematiccatalogue of the Aleyrodidae(Homoptera) with host plant andnatural enemy data. John Wiley,Chichester, GB. 340 p.
Vázquez, L. L.; de la Iglesia, M.; López, D.;Jiménez, R.; Mateo, A.; Vera, E. R.1995. Moscas blancas (Homoptera:Aleyrodidae) detectadas en losprincipales cultivos agrícolas deCuba. MIP (Costa Rica) 36:18-21.

250
Whiteflies and Whitefly-borne Viruses in the Tropics
Vázquez, L. L.; Jiménez, R.; de la Iglesia,M.; Mateo, A.; Borges, M. 1996.Plantas hospederas de Bemisiatabaci (Homoptera: Aleyrodidae) enCuba. Rev. Biol. Trop. 44(3):143-148.
Viscarret, M. M.; Botto, E. N. 1996.Descripción e identificación deTrialeurodes vaporariorum(Westwood) y Bemisia tabaci(Gennadius) (Hemiptera,Homoptera: Aleyrodidae). Rev.Chilena Ent. 23:51-58.

251
Biotypes of Bemisia tabaci
CHAPTER 3.13
Using Molecular Techniques toAnalyse Whitefly Species and Biotypesin Latin America
Lee Calvert*, Natalia Villarreal* andDonald Frohlich**
Introduction
When identifying and describing insecttaxa, morphology has been usedhistorically to separate species. Amongmany groups of insects, however,morphological characters can vary withrespect to environmental factors withina single species, or be so convergentand cryptic among closely relatedspecies as to be of limited usefulness.Under such conditions, studies of theirbiology and molecular profiles becomeessential to defining species andcharacterizing populations. At amolecular level, protein and DNApolymorphisms can be combined withstudies of biological characteristics byusing one of four experimental ortechnological approaches:electrophoresis of allozymes, analysisof randomly amplified polymorphicDNAs (RAPDs) and nucleic acidsequence comparisons of nuclear ormitochondrial DNA markers. Here, wereview the application of molecularapproaches to characterizing whiteflyBemisia tabaci (Gennadius) populationsand biotypes in Latin America.
Each method has its owncharacteristic advantages anddisadvantages. Isozyme analyses using
starch—or polyacrylamide—gelelectrophoresis have been in use forseveral decades and are useful forprocessing large numbers of samplesrelatively inexpensively compared toDNA sequencing. However, because thetechnique relies on detecting enzymes,samples must be kept live or frozen inorder to preserve activity. Thetechnique is also less sensitive thanDNA approaches because theunderlying nucleic acid variability isusually masked.
When, in the early 1980s, changesin whitefly populations and associatedbegomovirus infections were firstnoticed, protein polymorphisms wereemployed to investigate naturalpopulations of B. tabaci. Differences inesterase isozyme patterns were used todescribe two biotypes in the Americas:a now-known-to-be native form, orbiotype A, and a second form, biotypeB, which exhibited high populationdensity, wide host plant range,relatively high insecticide resistanceand was capable of inducing “silverleaf”symptoms on some plants (Brown etal., 1995). B. tabaci biotype A waspredominant in most regions of theAmericas but many of thesepopulations now have been displacedby B. tabaci biotype B. Other reportssuggested that there might beadditional biotypes in Latin America(Wool et al., 1994). Eighteen otherbiotypes from throughout the world
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** University of St. Thomas, Houston, TX, USA.

252
Whiteflies and Whitefly-borne Viruses in the Tropics
have been described based on esterasebanding patterns alone (Brown et al.,1995). Reliable morphological markers,which do not vary with ecologicalparameters (Mound, 1963; David andAnanthakrishnan, 1976; Mohanty andBasu, 1986) and which can distinguishbetween biotypes, are not known(Rosell et al., 1997).
DNA sequence comparisons can bemade between individuals orpopulations using polymerase chainreaction (PCR) generated sequencesfrom several markers in either thenuclear or mitochondrial genome. Thelatter offers some advantages becauseit is maternally inherited and non-recombining and contains known andpredictable gene sequences for whichgeneral insect primers have beendesigned. Some sequences vary enoughto be useful for population levelanalyses (e.g., the control region, partsof cytochrome oxidases I-III), whileothers evolve slowly enough to be moreappropriate to analyses above thespecies level (e.g., conserved parts ofcytochrome oxidase I [COI], 12S andparts of the 16S r-DNA). However, aswith some nuclear markers,comparisons between mitochondrialsequences may reflect local geneevolution and not population or speciesevolution. Nuclear data offer a widerange of neutral markers but can beparticularly difficult to design PCRprimers for specific targets in poorlyknown or unknown genomes (e.g.,introns). Thus, many studies have usedvariable regions surrounded byrelatively conserved sequences (e.g.,internal transcribed spacers or ITS ofthe rDNA genes) for population/speciesanalyses, or more conserved sequences(e.g., 18S rDNA) for higher orderstudies.
An alternative approach to specificnuclear markers involves the use ofshort, random sequence PCR primers
that anneal randomly in the genomeand produce characteristic bandingpatterns of collections of PCR productswhen run on an agarose gel (RAPDs).Advantages here include the low costand capability of processing largenumbers of samples without thenecessity of cloning or sequencing PCRproducts as well as no requirement fordetailed sequence knowledge about thegenome. Disadvantages include the factthat repeatability can be difficult, andthe polymorphisms may be hard todistinguish from PCR artefacts. That is,template preparation and amplificationconditions must be consistent andtightly controlled. Interpretation ofnegative data (absence of bands) alsomay have numerous explanations.
Perring et al. (1993) used RAPDpatterns in combination with allozymefrequency data and mating andbehavioural studies to elevate the Bbiotype to new species status, Bemisiaargentifolii Bellows & Perring. In adetailed study of 20 RAPD primers,however, Gawel and Bartlett (1993)concluded that the technique was notuseful for clarifying taxon status in thecase of whiteflies. Campbell et al.(1994) were the first to look at anuclear DNA sequence, albeit highlyconserved, and showed that only asingle unique nucleotide difference liesbetween B. tabaci biotypes A and B inthe 18S gene. More recently, Frohlichet al. (1999) used variable portions ofthe mitochondrial COI gene and16S rDNA sequence to evaluateB. tabaci populations from fourcontinents, and concluded that biotypeB is a recent introduction from the OldWorld to the Americas. De Barro et al.(2000), using ribosomal ITS 1, reachedthe same conclusion and cautionedthat if the status of the B biotype wereto remain species novum, a taxonomicreview of all of the biotypes of B. tabaciwould have to be made.

253
Biotypes of Bemisia tabaci
For this study of the whitefliesaffecting crops of economic importance,a variable portion of the 16Smitochondrial ribosomal DNA waschosen for evolutionary analysis. It ismore variable than the 18S mt DNA butless so than the COI gene. This allowsboth a comparison of distinct genera aswell as closely related biotypes.
Methods and Materials
PCR, cloning and sequenceanalysis of a region of the 16Smitochondrial DNA
The primers 4119(5’ CGCCTGTTTAACAAAAACAT) and4118 (5’ CCGGTCTGAACTCAGATCACGT 3’) were used to amplify aregion of the 16S mitochondrial DNA(Xiong and Kocher, 1991). The PCRreaction conditions were 30 cycles of1 min at 95 °C, 50 s at 50 °C and50 s at 72 °C, and in the last cycle the72 °C reaction was extended to 10 min.The products were purified using theWizardTM PCR purification columns(Promega, WI, USA) and were visualizedby agarose gel electrophoresis. The PCRproducts were cloned into the plasmidPCR script amp SK(+)TM (Stratagene,La Joya, CA, USA). Using the ABI dyeterminator kit, the sequences weredetermined in an automated sequencerusing the dideoxynucleotide chaintermination procedure (Sanger et al.,1977).
Phylogenetic analyses
Phylogenetic analyses were done withmultiple individuals within populations.DNA sequences were aligned using theClustalW algorithm (Thompson et al.,1994) by the ClustalW 1.7 program(BCM Search Launcher at the HumanGenome Centre, Baylor College ofMedicine, Houston, TX, USA). Becausedifferent tree building algorithms make
different evolutionary assumptions, datawere evaluated by parsimony, neighbourjoining and maximum-likelihood. Allanalyses were performed with PAUP,version 4.0b2, for Macintosh (Swofford,1999). For parsimony, the branch-and-bound method was used (charactersunordered, equal weight). Bootstrappingwas performed with the branch-and-bound option for 2000 replicates(stepwise sequence addition, tree-bisection-reconnection [TBR], MulTreesoption). For neighbour joining, distanceswere calculated using the Kimura 2parameter model. Maximum-likelihoodtrees were constructed with atransition/transversion ratio of 2.0 byheuristic search (100 replicates, randomaddition sequence, MulTrees, TBR)(Swofford, 1999).
Whitefly survey
The whiteflies that were identified byRAPD PCR analysis were collected andprocessed.1
RAPD PCR analysis
Total DNA was isolated from individualwhiteflies using a method developed forplants (Gilbertson et al., 1991). The DNAwas amplified in a PCR. The Operonprimers F2 (5’GAGGATCCCT3’),F12 (5’ACGGTACCAG3’), H9(5’TGTAGCTGGG3’) and H16(5’TCTCAGCTGG3’) (De Barro andDriver, 1997) were tested for theirefficacy to distinguish whiteflies in LatinAmerica. The reaction conditions for thefirst cycle were 5 min at 94 °C, 2 min at40 °C and 3 min at 72 °C. This wasfollowed with 39 cycles of 1 min at94 °C, 1.5 min at 40 °C, and
1. For further information on specificmethodologies used to produce the resultspublished in this book, please contact theinformation and communication assistant ofthe Project (www.tropicalwhiteflyipmproject.cgiar.org).

254
Whiteflies and Whitefly-borne Viruses in the Tropics
2 min at 72 °C. The PCR products wererun in agarose gels stained withethidium bromide and visualized usingUV light.
Results
Analysis of selected whitefliesin Latin America
A region of the 16S mitochondrial DNAof B. tabaci biotypes A and biotype B,B. tuberculata, Aleurotrachelus socialis(Bondar), Trialeurodes vaporariorum(Westwood) and T. variabilis(Quaintance) were compared byparsimony and distance analysis usingPhylip version 3.57 (Felsenstein, 1993).At least two independent clones fromeach whitefly population of the3’ region of the mitochondrial 16S genewere prepared and sequenced. Includedin the comparison were sequence dataof the mitochondrial 16S gene fromB. tabaci biotype A of Arizona (Genbankaccession: AF110722), Costa Rica(Genbank accession: AF110715) andPuerto Rico (Genbank accession:AF110719) (Frohlich et al., 1999). In the
analysis of various B. tabaci, those ofbiotype A were grouped together andthere was at least 97% identitywith a maximum mean distance of0.02 (Table 1) (Calvert et al., 2001). TheB. tabaci biotype A is widespreadthroughout the region of the whiteflysurvey.
In the distance analysis, the IsraelB. tabaci isolate (Genbank accession:AF110717) was 98.8% identical withthe B. tabaci biotype B isolates fromColombia (Table 1). Comparisons werealso made with individuals fromColombia, Arizona, Israel and Yemenand all were at least 98.2% identical(data not shown). Given the rapidspread of the B biotype, the lack ofdiversity between populations inArizona USA and Colombia wasexpected. This gives additional evidencethat B. tabaci biotype B was recentlyintroduced into the Americas from theregion of the Middle East.
A representative from each speciesor biotype was used in the analysis tofind the most parsimonious tree(Figure 1). B. tabaci biotypes A and
Table 1. Mean distances for a 3’ region of mitochondrial 16S ribosomal gene in fifteen individualwhiteflies representing different species and populations.
Whiteflya 1 2 3 4 5 6 7 8 9 10 1 1 12 13 14 15
1 B. tabaci B Sucre 0.00 0.01 0.01 0.01 0.10 0.10 0.09 0.10 0.10 0.22 0.35 0.35 0.37 0.37 0.36
2 B. tabaci B CT cass 0.00 0.00 0.00 0.08 0.09 0.08 0.09 0.09 0.21 0.34 0.34 0.36 0.36 0.35
3 B. tabaci B CT bn 0.00 0.00 0.09 0.10 0.09 0.09 0.09 0.21 0.34 0.35 0.37 0.36 0.35
4 B. tabaci Israel 0.00 0.08 0.09 0.08 0.09 0.09 0.21 0.34 0.34 0.36 0.36 0.35
5 B. tabaci A CT 1 0.00 0.01 0.01 0.02 0.00 0.22 0.34 0.35 0.36 0.35 0.38
6 B. tabaci A CR 0.00 0.01 0.02 0.01 0.23 0.35 0.36 0.37 0.36 0.39
7 B. tabaci A AZ 0.00 0.02 0.01 0.22 0.34 0.36 0.37 0.36 0.38
8 B. tabaci A PR 0.00 0.02 0.22 0.35 0.36 0.36 0.36 0.38
9 B. tabaci CT 2 0.00 0.22 0.34 0.35 0.36 0.36 0.38
10 B. tuberculata CT 0.00 0.28 0.28 0.37 0.36 0.38
11 A. socialis Mon 0.00 0.07 0.40 0.39 0.43
12 A. socialis CT 0.00 0.41 0.40 0.41
13 T. vaporariorum CT 0.00 0.00 0.39
14 T. vaporariorum AZ 0.00 0.38
15 T. variabilis CT 0.00
a. B, Bemisia; A, Aleurotrachelus; T, Trialeurodes.

255
Biotypes of Bemisia tabaci
B were always grouped together.B. tuberculata was grouped with theB. tabaci 92% of the time andT. vaporariorum and T. variabilis weregrouped together 86% of the time.Using parsimony analysis, therelationship between the three generaof Bemisia, Trialeurodes andAleurotrachelus was not clear. Theresults of the distance analysis(Table 1) show that Aleurotracheluswas closer to the genus Bemisia thanto Trialeurodes. An unexpected resultwas the relatively large meandistance between T. vaporariorum andT. variabilis.
Gene sequences andphylogenetic studies
The use of DNA sequences hasbecome increasingly more importantas a tool to study the evolution,populations and systematics ofinsects. Since the database forsequence information is expandingexponentially, it is certain that thesemethods will be even more importantin the future. The several advantagesto using DNA include the fact thatthe genotype, and not the phenotype,is examined directly. There is also an
expanding base of phylogeneticinformation on an increasing number ofgene sequences that exhibit differentrates of change.
This study generated additionalevidence that the B. tabaci biotype B ishighly conserved throughout theAmericas. This was expected because itis a recent introduction. Thepopulations of B. tabaci biotype A aremore conserved than we would havepredicted from the studies that usedesterase isozyme pattern as the meansto detect diversity. Because theexpression of esterases can be inducedby insecticide applications, theirphenotypes are probably not reliable fordetermining biotypes. Comparing theresults of the molecular and esteraseanalyses, many esterase isozymepatterns are associated with theB. tabaci biotype A based on DNAmarkers.
Even though, in the phylogeneticanalysis, T. vaporariorum andT. variabilis were in the same clade, theabsolute distance was fairly high. Sincethe distance is equally great betweenT. vaporariorum, T. variabilis and the
Bemisia tuberculata
B. tabaci A
B. tabaci B
Aleurotrachelus socialis
Trialeurodes vaporariorum
T. variabilis
Magicicada septendecula
Stenocladius sp.
Figure 1. Cladogram showing the relationship between the whiteflies in this study. The cladogram isbased on the most parsimonious tree inferred from the analysis of 485 base sites of a regionof the mitochondrial 16S gene. Numbers above the branches indicate the level of statisticalsupport for the corresponding node from 10,000 bootstrap replicates.
100
100
92
65
99
90

256
Whiteflies and Whitefly-borne Viruses in the Tropics
whiteflies in the other genera, there is aquestion of how closely related themembers of Trialeurodes may be.Further studies should determine ifthese two species are members of thesame genera, or whether they should beplaced into separate genera.
The application of RAPD foridentification of whiteflies
Whenever an increase in whitefliesoccurs or they begin to affect additionalcrops, the introduction of the B. tabaciB biotype is suspected. Often, this isthe case but whitefly populations areaffected by many environmentalfactors, including the cropping systemand the varieties grown. The molecularmethods that generate DNA sequencedata are time and labour intensive andare not suited to large scale monitoringof populations. Rapid and reliablemethods are needed to distinguishbetween the most common whiteflies.In Latin America, these are B. tabacibiotypes A and B, and T. vaporariorumon most crops.
Using RAPDs has severaladvantages. Foreknowledge about anyparticular gene in the target organismis not needed. More than one primercan be used to increase confidence inthe reliability of the method. Withwhiteflies and many other types ofsamples, the ability to store them formany months at room temperature in70% ethanol facilitates shipping themacross international borders and allowsthe analysis of large numbers ofsamples to be processed in an orderlymanner.
RAPD was used (De Barro andDriver, 1997) to distinguish indigenousAustralian populations of B. tabaci fromthe introduced B biotype. The authorsreported on four oligonucleotideprimers that they considered the mostuseful for identifying the native B.tabaci from the B biotype.
Analysis of molecular markersto identify whiteflies in LatinAmerica
Four oligonucleotide primers (De Barroand Driver, 1997) were tested for theirutility in distinguishing B. tabacibiotype A and biotype B, andT. vaporariorum. In the analysis of PCRproducts using the primer H9 (Operon,USA), there were differences for therange of whitefly species tested(Figure 2). For B. tabaci biotype B andT. vaporariorum, there are prominentPCR products ca. 600 and 800 bp thatcan sometimes make distinguishing thetwo species difficult. The unique PCRproduct in B. tabaci biotype B ca.950 bp and T. vaporariorum’s uniquePCR product at ca. 500 bp areimportant for distinguishing betweenthese two species. Although confusioncan occur in the interpretation betweensome species, the H9 primer was mostuseful in distinguishing betweenB. tabaci biotypes A and B. At 600 bp,some PCR products are similar in sizein both biotypes but the biotype A hasseveral unique PCR products, includingdoublet bands at 300-350 bp. The
Figure 2. Using Operon primer H9, these arethe RAPD-PCR DNA products fromindividual whiteflies. M:1kb markers(BRL), BA: Bemisia tabaci biotype A,BB: Bemisia tabaci biotype B.
M BA BB

257
Biotypes of Bemisia tabaci
biotype B has unique PCR products atca. 600, 700 and 900 bp comparedwith one product of ca. 850 bp in thebiotype A.
The primer H16 (Figure 2) wasmost useful in distinguishing betweenthe whitefly species. While there can besome common bands in the 500 to1000 bp range for both B. tabacibiotypes, three products in B. tabacibiotype B of ca. 350, 450 and 550 bpwere consistently useful foridentification of the B biotype.
When primers F2 and F12 wereused, there were larger numbers ofPCR products. These can be used fordistinguishing the whitefly species but,because of the large numbers of bands,were generally less useful than H9 andH16. When using RAPDs for theidentification of the whiteflies in thisstudy, analysing the individualwhiteflies with both the H9 andH16 primers is recommended.
Mapping the Distributionof Whitefly Species and
Biotypes in Latin America
This survey was undertaken to map thedistribution and range of whiteflies invarious countries in Latin America. Forthis purpose, RAPDs were used tocharacterize the principal whitefly pestsin many countries of Central Americaand the Caribbean region.
The survey using molecularmarkers complemented the activitieswhere whitefly specimens wereanalysed by light microscopy. For themolecular analysis, at least one andoften two primers were used and theresults were compared to themorphological identification. The use ofRAPD data was the only method todistinguish between the biotypes A andB in the Bemisia complex.
This survey was extensive but notexhaustive, therefore results should beinterpreted as a representation of thepredominate whiteflies populations invarious regions of Cuba, the DominicanRepublic, Guatemala, Honduras, CostaRica, Panama, Colombia and Venezuela.
Dominican Republic
In the departments of Azua, Barahona,Peravia, Santiago, Montecristo and SanJuan, the predominant whitefly wasB. tabaci biotype B. Plants testedinclude eggplant (Solanum melongenaL.), tomato (Lycopersicon esculentumMill.), okra (Abelmoschus esculentus [L.]Moench), melon (Cucumis melo L.),cucumber (Cucumis sativus L. var.sativus) and hibiscus (Hibiscus spp. L).Only one sample of tomato in Azua wasclassified a biotype A. In the lowlandtropics of the Dominican Republic, thebiotype B was introduced nearly adecade ago and has nearly excluded theindigenous biotype A. In the departmentof Vega, common bean (Phaseolusvulgaris L.), tomato and potato (Solanumtuberosum L.) were tested and T.vaporariorum was the only whiteflyfound.
Guatemala
In the department of Zacapa, thepredominant whitefly was B. tabaci. InZapcapa, the B biotype was present onall the hosts tested, which includedokra, melon, watermelon (Citrulluslanatus [Thunb.] Matsum. & Nakai) andcucumber. In Japala, both B. tabacibiotype B and T. vaporariorum werefound. The B biotype was found onokra, melon, watermelon, cucumber,tomato, eggplant, tobacco (Nicotianatabacum L.) and weed species.T. vaporariorum was the predominatewhitefly in several host plants includingpaw paw (Carica papaya L.), “tomate-manzana” (Lycopersicon esculentum var.beef tomato), cherry tomato(Lycopersicon esculentum var.

258
Whiteflies and Whitefly-borne Viruses in the Tropics
cerasiforme [Dunal] A. Gray), commonbean and some samples of tomato.
Honduras
In Honduras, the predominant whiteflywas B. tabaci biotype A. Most of thesamples were common bean, tomatoand cucumber from the ComayaguaDepartment. The paw paw inComayagua was host toT. vaporariorum. On chilli peppers(Capsicum spp. L.) in the FranciscoMorazan Department, the A biotypewas also the only whitefly identified.
Costa Rica
In the lowland tropics of Costa Rica,B. tabaci biotype A is still thepredominant whitefly. In thedepartments of Guanacaste, San José,Heredia and Alajuela, most of thesamples were biotype A. The A biotypewas found on common bean, melon,watermelon, tomato, chilli peppers,cucumber and others. In thedepartments of Arajuel andPuntarenas, the B biotype was present.It was the only whitefly found in thetwo samples from Arajuel but inPuntarenas both biotype A and B werepresent. T. vaporariorum was present inthe Alajuela Department and wascommon in Cartago where it was foundon chilli peppers and tomato.
El Salvador
Only a limited number of samples (15)have been tested and all were B. tabacibiotype B.
Panama
The principal whitefly in theDepartment of Chiriqui wasT. vaporariorum. In the other regions ofthe country, B. tabaci biotype B wasthe predominant whitefly. Populationsof B. tabaci biotype A are still foundand in some areas the populations aremixed. This suggests a relatively recent
introduction of B. tabaci biotype B inPanama.
Cuba
B. tabaci biotype B was the onlywhitefly found in the samples fromCuba. The B biotype has been in Cubafor nearly 10 years and clearly hasbecome the predominant whitefly.
Colombia
Most of the samples tested were fromthe north coastal region of Colombia,where the principal whitefly is B. tabacibiotype B. The B biotype is affectingtomato, col (Brassica oleracea L. var.capitata L.), eggplant, melon andcassava (Manihot esculenta Crantz). TheA biotype is still present but theintroduction of the B biotype isrelatively recent.
Venezuela
Samples were collected from thedepartments of Zamora and Jiménez,where the principal whitefly was theB biotype. In the UrdanetaDepartment, there was a mixture of theA and B biotypes. Only a few sampleswere analysed and more are neededbefore conclusions should be made.
A Regional View ofWhitefly Populations
Most of the samples that weredetermined to be in the B. tabacicomplex by morphological methodswere classified as either biotype A orB (Table 2). In general, about 15% ofthe samples could not be amplified;only 23% of the samples fromVenezuela could be amplified. This maydemonstrate the importance of theproper sample handling. About 5% ofthe samples also had RAPD PCRproducts that could not be identified.These may well be either different
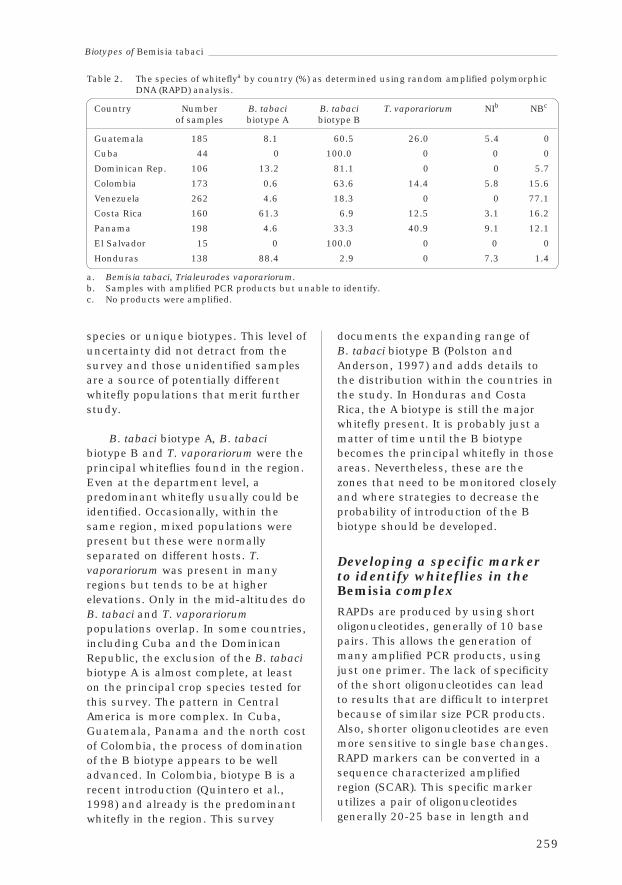
259
Biotypes of Bemisia tabaci
species or unique biotypes. This level ofuncertainty did not detract from thesurvey and those unidentified samplesare a source of potentially differentwhitefly populations that merit furtherstudy.
B. tabaci biotype A, B. tabacibiotype B and T. vaporariorum were theprincipal whiteflies found in the region.Even at the department level, apredominant whitefly usually could beidentified. Occasionally, within thesame region, mixed populations werepresent but these were normallyseparated on different hosts. T.vaporariorum was present in manyregions but tends to be at higherelevations. Only in the mid-altitudes doB. tabaci and T. vaporariorumpopulations overlap. In some countries,including Cuba and the DominicanRepublic, the exclusion of the B. tabacibiotype A is almost complete, at leaston the principal crop species tested forthis survey. The pattern in CentralAmerica is more complex. In Cuba,Guatemala, Panama and the north costof Colombia, the process of dominationof the B biotype appears to be welladvanced. In Colombia, biotype B is arecent introduction (Quintero et al.,1998) and already is the predominantwhitefly in the region. This survey
documents the expanding range ofB. tabaci biotype B (Polston andAnderson, 1997) and adds details tothe distribution within the countries inthe study. In Honduras and CostaRica, the A biotype is still the majorwhitefly present. It is probably just amatter of time until the B biotypebecomes the principal whitefly in thoseareas. Nevertheless, these are thezones that need to be monitored closelyand where strategies to decrease theprobability of introduction of the Bbiotype should be developed.
Developing a specific markerto identify whiteflies in theBemisia complex
RAPDs are produced by using shortoligonucleotides, generally of 10 basepairs. This allows the generation ofmany amplified PCR products, usingjust one primer. The lack of specificityof the short oligonucleotides can leadto results that are difficult to interpretbecause of similar size PCR products.Also, shorter oligonucleotides are evenmore sensitive to single base changes.RAPD markers can be converted in asequence characterized amplifiedregion (SCAR). This specific markerutilizes a pair of oligonucleotidesgenerally 20-25 base in length and
Table 2. The species of whiteflya by country (%) as determined using random amplified polymorphicDNA (RAPD) analysis.
Country Number B. tabaci B. tabaci T. vaporariorum NIb NBc
of samples biotype A biotype B
Guatemala 185 8.1 60.5 26.0 5.4 0
Cuba 44 0 100.0 0 0 0
Dominican Rep. 106 13.2 81.1 0 0 5.7
Colombia 173 0.6 63.6 14.4 5.8 15.6
Venezuela 262 4.6 18.3 0 0 77.1
Costa Rica 160 61.3 6.9 12.5 3.1 16.2
Panama 198 4.6 33.3 40.9 9.1 12.1
El Salvador 15 0 100.0 0 0 0
Honduras 138 88.4 2.9 0 7.3 1.4
a. Bemisia tabaci, Trialeurodes vaporariorum.b. Samples with amplified PCR products but unable to identify.c. No products were amplified.

260
Whiteflies and Whitefly-borne Viruses in the Tropics
amplifies a single region of a genome(Ohmori et al., 1996). This makesinterpretation of the results muchsimpler than with RAPDs. Instead of agroup of bands with a degree ofvariability, SCAR produces singlebands in the target organism.
The SCAR for the B. tabaci biotypeB was developed by cloning the RAPDPCR products of the primer H9. ThecDNA clones were analysed andsequenced. The primers were designedand tested. One set of primers onlyamplified B. tabaci biotype B (Figure 3).This set of primers did not amplify anytested population of B. tabaci biotype Aor T. vaporariorum. Further testing isneeded to confirm that these primersamplify this PCR product ingeographically separated populations ofB. tabaci biotype B. Given the lack ofdiversity in the evolutionary studies,this SCAR is expected to be useful forrapidly identifying all populations ofbiotype B.
The current research is directed todeveloping SCARs for B. tabaci biotype
A and T. vaporariorum. Each set ofprimers will amplify bands of differentsizes. This will allow the primers to bemixed together and will distinguishbetween the whiteflies in a single PCRreaction. Thus more extensive surveyspossible will be possible and thetechnique will be more consistent in allthe laboratories that need thediagnostic capability to rapidly identifywhitefly pests.
Conclusions
Our detailed molecular analysis ishelping define both the range and thevariability of B. tabaci biotypes A and Bin Latin America. Similar studies inother regions of the world have beenmade and will allow a global view of theBemisia complex. The molecularmethod of RAPDs proved essential indistinguishing between the B. tabacibiotype A and B but, because of thenature of the method, comparingresults between laboratories is difficult.Therefore we are developing SCARs tosimplify the identification of whiteflies.
Figure 3. Two SCARs were developed to distinguish Bemisia tabaci biotype A (1) and Bemisia tabacibiotype B (2). Lane 1 Bemisia tabaci biotype A, lane 2 Bemisia tabaci biotype B, lane 3Bemisia tuberculata, lane 4 Trialeurodes vaporariorum, lane 5 Aleurotrachelus socialis,6:DNA free reaction and M:1kb markers (BRL).
M 1 2 3 4 5 6 M M 1 2 3 4 5 M

261
Biotypes of Bemisia tabaci
Already we can identify B. tabacibiotype B using a SCAR, and we aremaking progress in the development ofSCARs for B. tabaci biotype A andT. vaporarorium. Specific unambiguousresults of one amplified band for eachspecies or biotype of whitefly willsimplify the interpretation of resultsand become a rapid and relativelyinexpensive method to use in surveysof whiteflies.
References
Brown, J. K.; Coats, S.; Bedford, I. D.;Markham, P. G.; Bird, J.; Frohlich,D. R. 1995. Characterization anddistribution of esteraseelectromorphs in the whitefly,Bemisia tabaci (Genn.)(Homoptera:Aleyrodidae). Biochem. Genet.33:205-214.
Calvert, L. A.; Cuervo, M.; Arroyave, J. A.;Constantino, L. M.; Bellotti, A.;Frohlich, D. 2001. Morphologicaland mitochondrial DNA markeranalyses of whiteflies (Homoptera:Aleyrodidae) colonizing cassava andbeans in Colombia. Ann. Entomol.Soc. Am. 94(4):512-519.
Campbell, B. C.; Steffen-Campbell, J. D.;Gill, R. J. 1994. Evolutionary originof whiteflies (Hemiptera:Sternorryncha: Aleyrodidae)inferred from 18S rDNA sequences.Insect Mol. Biol. 3:73-89.
David, B. V.; Ananthakrishnan T. N.1976. Host correlated variation inTrialeurodes rara (Singh) andBemisia tabaci (Gennadius)(Homoptera: Aleyrodidae). Curr. Sc.(Bangalove) 45:223-225.
De Barro, P. J.; Driver, F. 1997. Use ofrandom amplified polymorphic DNApolymerase chain reaction (RAPDPCR) to distinguish the B biotypefrom other biotypes of Bemisiatabaci (Gennadius) (Hemiptera:Aleyrodidae). Aust. J. Entomol.36:149-152.
De Barro, P. J.; Driver, F.; Trueman, J. W.H.; Curran, J. 2000. Phylogeneticrelationships of world populationsof Bemisia tabaci (Gennadius) usingribosomal ITS1. Molec. Phylo. Evol.16:29-36.
Felsenstein, J. 1993. PHYLIP (PhylogenyInference Package) version 3.5c.Distributed by author. Departmentof Genetics, University ofWashington, Seattle, USA.
Frohlich, D. R.; Torres-Jerez.; Bedford, I.D.; Markham, P. G.; Brown, J. K.1999. A phylogeographic analysis ofthe Bemisia tabaci species complexbased on mitochondrial DNAmarkers. Mol. Ecol. 8:1683-1691.
Gawell, N. J.; Bartlett, A. C. 1993.Characterization of differencesbetween whiteflies using RAPDPCR. Insect Mol. Biol. 2:33-38.
Gilbertson, R. L.; Rojas, M. R.; Russell, D.R.; Maxwell, D. P. 1991. Use of theasymmetric polymerase chainreaction and DNA sequencing todetermine genetic variability of beangolden mosaic geminivirus in theDominican Republic. J. Gen. Virol.72: 2843-2848.
Mohanty, A .K.; Basu, A. N. 1986. Effectof host plants and seasonal factorson intraspecific variations in pupamorphological of the whitefly vector,Bemisia tabaci (Genn.) (Homoptera:Aleyrodidae). J. Ent. Res. 10:19-26.
Mound, L. A. 1963. Host-correlatedvariation in Bemisia tabaci (Genn.)(Homoptera Aleyrodidae). Proc. R.Ent. Soc. London. 38:171-180.
Ohmori, T.; Murata, M.; Motoyoshi, F.1996. Molecular characterization ofrandom amplified polymorphic DNA(RAPD) and sequence characterizedamplified region molecular (SCAR)markers linked to the Tm-1 locus intomato. Theor. Appl. Genet. 92:151-156.

262
Whiteflies and Whitefly-borne Viruses in the Tropics
Perring, T. M.; Cooper, A. D.; Rodríguez,R. J.; Farrar, C. A.; Bellows, T. S.1993. Identification of a whiteflyspecies by genomic and behaviouralstudies. Science 259:74-77.
Polston, J. E.; Anderson, P. K. 1997. Theemergence of whitefly-transmittedgeminiviruses in tomato in thewestern hemisphere. Plant Dis.81:1358-1369.
Quintero, C.; Cardona, C.; Ramírez, D.;Jiménez, N. 1998. Primer registrodel biotipo B de Bemisia tabaci(Homoptera: Aleyrodidae) enColombia. Rev. Colomb. Entomol.24:23-28.
Rosell, R. C.; Bedford, I. D.; Frohlich, D.R.; Gill, R. J.; Brown, J. K.;Markham, P. G. 1997. Analysis ofmorphological variation in distinctpopulations of Bemisia tabaci(Homoptera: Aleyrodidae). Ann.Entomol. Soc. Am. 90:575-589.
Sanger, F.; Nicklen, S.; Coulson, A. R.1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl.Acad. Sci. USA. 74:5463-5467.
Swofford, D. L. 1999. PAUP*. PhylogeneticAnalysis Using Parsimony (*andother methods). Version 4. SinauerAssociates, Sunderland, MA, USA.
Thompson, J. D.; Higgins, D. G.; Gibson,T. L. 1994. CLUSTAL W: Improvingthe sensitivity of progressivemultiple sequence alignmentthrough sequence weighting,positions-specific gap penalties andweight matrix choice. Nucleic AcidsRes. 22:4673-4680.
Wool, L.; Calvert, L.; Constantino, L. M.;Bellotti, A. C.; Gerling, D. 1994.Differentiation of Bemisia tabaci(Genn.) populations in Colombia. J.Appl. Entomol. 117:122:134.
Xiong, B.; Kocher, T. D. 1991.Comparison of mitochondrial DNAsequences of seven morphospeciesof black flies (Diptera: Simuliidae).Genome 34:306-311.

263
Conclusions and Recommendations
CHAPTER 3.14
Conclusions and RecommendationsFrancisco Morales*
Despite over 40 years of research onBemisia tabaci (Gennadius)(Homoptera:Aleyrodidae) as a pest andvector of plant viruses, this whiteflyspecies continues to causeconsiderable damage to food andindustrial crops in Mexico, CentralAmerica and the Caribbean. B. tabacifirst became a pest of cotton(Gossypium hirsutum L.) in MiddleAmerica, and then a major vector ofplant viruses in common bean(Phaseolus vulgaris L.) and tomato(Lycopersicon esculentum Mill.)plantings throughout the region. Theexpansion of non-traditional crops inthe 1980s, mainly vegetables forexport, further aggravated the problemby providing more reproductive hostsfor the whitefly pest and a higherdemand for pesticides. Theintroduction in the early 1990s of amore aggressive and polyphagousbiotype (B) of B. tabaci in the Americasfurther jeopardized past efforts tocontrol this pest.
In this region, whitefly-bornegeminiviruses (begomoviruses) aremore prevalent during the dry monthsof the year (November through April),when B. tabaci populations encountermore favourable weather. The areaplanted to crops such as melon
(Cucumis melo L.), common bean,tomato, eggplant (Solanum melongenaL.), cucurbits and sweet and hotpeppers (Capsicum spp. L.) has beenreduced greatly or completelyabandoned during this period of theyear because of the frequent outbreaksof whiteflies and whitefly-borne viruses.The development of B. tabaci from aharmless insect to a major pest isclosely related to the introduction andintensive use of pesticides after theSecond World War. Pesticide abuseremains a major factor in B. tabaci andbegomovirus epidemics in this region.Whiteflies are known to developresistance to insecticides, the frequentuse of which eliminates naturalpredators of B. tabaci. It is notuncommon to see farmers sprayinginsecticides on susceptible crops everyother day and even daily, up untilharvest time, in open defiance of foodsafety regulations. The high levels ofpesticide residues in tomato, pepper,common bean, eggplant and othervegetables attacked by B. tabacidisqualifies these products for sale ininternational markets and constitutes aserious health hazard for the millionsof rural and urban consumers in theregion, where produce is not tested fortoxic residues.
But the main reason for thepersistence of these pest problems isthe lack of qualified technicalassistance for farmers, particularly
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.

264
Whiteflies and Whitefly-borne Viruses in the Tropics
those who have attempted to diversifytheir subsistence crops with moreprofitable cash crops such as tomatoand pepper. This situation is areflection of the increasinglydeteriorating economic situation of theregion since the mid 1970s, which hasnegatively impacted on the capacity ofnational agricultural researchinstitutions and internationalagricultural research centres toconduct research and provide thenecessary technical assistance tofarmers affected by severephytosanitary problems such aswhiteflies and begomoviruses.Moreover, the diversion of foreign aidfunds from crop improvement tonatural resource management hasfurther contributed to increased yieldlosses due to pest problems,abandonment of profitable crops grownby small-scale farmers, higherproduction costs and considerablymore environmental and foodcontamination.
Whitefly and begomovirus controlstrategies in the region differ accordingto countries, agricultural areas andcrops. In north-western Mexico, theuse of virus-resistant common beanvarieties and more efficient insecticidesfor vegetables has greatly reduced theimpact of these pests. In the YucatánPeninsula, Mexico, small-scale farmerswere using physical control methodsduring the first 3 weeks aftertransplant of tomato and pepper. In theDominican Republic, a crop-free periodwas legally imposed throughout thecountry to break the cycle of thewhitefly before the main plantingseason. And in most Central Americancountries, susceptible crops were onlyplanted during the rainy season, whenwhitefly populations are at their lowestlevel. These whitefly/virus controlstrategies have reduced, but noteliminated, the production lossesassociated with outbreaks of B. tabaci
due to various factors. The constantflow of improved cultivars developed inthe past through conventional plantbreeding methods has been greatlyreduced because of the radical changesthat have occurred in the past decadein favour of molecular breedingmethods. Second, technical assistanceto small-scale farmers is practically nil,which leaves plant protection in thehands of pesticide salespeople.Susceptible crop-free periods reducewhitefly incidence but do not increasethe productivity of prime agriculturalland that must be devoted to lessprofitable crops or remain idle duringseveral months of the year. The newinsecticides in the market are effectivefor whitefly control but are veryexpensive for resource-poor farmers.
The Tropical Whitefly IntegratedPest Management (TWF-IPM) Projecthas successfully identified, tested andvalidated the most efficient andsustainable whitefly control methodsused in over 10 countries in MiddleAmerica. So far, the most valuable IPMmeasure identified is genetic resistanceto control whitefly-transmitted viruses,particularly in the case of commonbean. Unfortunately, practically nocrop improvement work is being donein the case of vegetables such astomato and pepper despite theirtropical American origin. Mostvegetable varieties are bred intemperate countries and thus are notadapted to the tropical conditions andpests that exist in the region. The TWF-IPM Project has successfully usedphysical barriers (micro-tunnels) toprotect sensitive crops during the firstcritical month of their life cycle. Thisstrategy also protects susceptibleplants against other insect-borneviruses and pests, and greatlyincreases their productivity per unitarea with minimum inputs. Theidentification and use of bio-controlagents of B. tabaci has been advanced

265
Conclusions and Recommendations
greatly in the region, particularly incountries such as Cuba and theDominican Republic. Finally, anenhanced understanding of the whiteflyspecies, biotypes and plant virusespresent in the region, has helpedunderstand the ecology andepidemiology of these crop productionproblems and, more important, hascontributed to the implementation ofsustainable and efficient IPM methods.
It is imperative to re-orientagricultural research in the region byunderstanding that spending ourlimited resources on purelyenvironmental or sociological issues,without a food production component,only perpetuates misery and increasesenvironmental degradation and humanhealth problems. There is considerableneed to strengthen national andinternational research and extensionservices to provide timely and effectivetechnical assistance to small-scalefarmers. We must help resource-poorfarmers to diversify their cropping
systems with more profitable crops inorder to maximize the income derivedfrom their limited land resources.Traditional crops are the basis of foodsecurity but they are merelysubsistence crops. High-value crops,particularly vegetable and fruit crops,can help small-scale farmers improvetheir livelihoods but this can only beachieved with proper and timelytechnical assistance provided by multi-disciplinary groups of agriculturalspecialists. Finally, there is a vacuumin the area of agricultural economicsand marketing for small-scale farmers.Unless these farmers are incorporatedinto the market chains, they will be atthe mercy of the vagaries of the openmarkets and intermediaries who profitfrom the misery and lack of marketknowledge of the resource-poor farmer.The TWF-IPM Project expects toaddress these issues and scale up thedissemination of the IPM measuresvalidated to date in Central America,Mexico and the Caribbean region.

BLANCA 266

SECTION FOUR
Whiteflies as Pests of Annual Cropsin the Tropical Highlands of
Latin America
SECTION FOUR
Whiteflies as Pests of Annual Cropsin the Tropical Highlands of
Latin America

BLANCA 268

269
Introduction
CHAPTER 4.1
IntroductionCésar Cardona*
According to Strong et al. (1984),724 species of whiteflies (Homoptera:Aleyrodidae) have been described fromthe tropics but only 420 fromtemperate regions. Most of theseinsects do not represent an economicthreat to agriculture and only ahandful can be described as seriouspests. In a recent survey conducted inCentral America and Colombia,Caballero (1992) listed the30 commonest and economically mostimportant whitefly species recordedfrom 84 host plant species. Amongthese, the sweetpotato whitefly Bemisiatabaci (Gennadius), the greenhousewhitefly Trialeurodes vaporariorum(Westwood) and the citrus whiteflyAleurocanthus woglumi Ashby wereregarded as key pests. Of lesserimportance were Trialeurodesabutiloneus (Haldeman) and threespecies affecting cassava:Aleurotrachelus socialis Bondar,Bemisia tuberculata Bondar andTrialeurodes variabilis (Quaintance).
T. vaporariorum and B. tabaci areby far the most important whiteflyspecies affecting annual crops in theLatin American highlands and thesetwo species are the focus of the presentreview. While reference is made to
B. tabaci and T. vaporariorum as virusvectors, emphasis is on the importanceof these insects as direct pests causingmechanical damage. The whitefliesaffecting cassava (Manihot esculentaCrantz), a semi-perennial crop, are notdiscussed.
Legumes andHorticultural Crops in the
Andean Highlands
The Andes encompass a wide range oflatitudes and climates but in thetropics especially, altitude is the maindeterminant of temperature andbiological character. Areas between1000 and 2000 m above sea level areknown as highlands. Those above2000 m have a quasi-temperate climateand are collectively known as the “highAndes”. Transitions from tropicallowlands to temperate highlands maybe abrupt and the alpine zones formecological islands, large or small. Asindicated by Winograd et al. (1998), theAndean backbone and its lateral rangescontribute remarkable diversity to theecosystems of Colombia and Ecuador.In parts of Colombia, small ridges andspurs connect the three main ranges,circumscribing a series ofintermountain basins, each of whichhosts a distinctive local community ofnatural biodiversity. Agriculturalsystems and their ecology arecorrespondingly diverse.
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.

270
Whiteflies and Whitefly-borne Viruses in the Tropics
The tropical highlands of Colombiaand Ecuador include extensiveagricultural areas with meantemperatures of 12-20 °C, variablerainfall patterns (700-1600 mm) andrelative humidity ranging from 50% to85%. Small-scale farmers growcommon bean and snap bean(Phaseolus vulgaris L.), tomato(Lycopersicon esculentum Mill.), potato(Solanum tuberosum L.) and otherhorticultural crops, both for homeconsumption and for sale. Crops areusually grown on steep, erosion-proneslopes, with no irrigation and little orno use of fertilizers.
In contrast, the lowlands ofColombia and Ecuador experiencemean temperatures of 26-28 °C,similarly variable rainfall patterns(600-1600 mm) and prolonged dryseasons that favour the buildup ofwhitefly populations. Cotton(Gossypium hirsutum L.), soybean(Glycine max [L.] Merr), melon (Cucumismelo L.) and, to a lesser extent, tomatoare grown on large farms that occupy asignificant proportion of theagricultural land. Many small-scalefarmers also grow less than 1 hectareof legumes, vegetables or tobacco(Nicotiana tabacum L.).
Status of Research onTrialeurodes and Bemisia
The centre of origin for T. vaporariorumhas not been identified. According toVet et al. (1980), the species is mostprobably indigenous to tropical andsubtropical America. T. vaporariorum, awell-known pest of greenhouse crops inEurope, has long been recorded onannual field crops in highland areas ofColombia (Posada, 1976) and Ecuador(Merino and Vásquez, 1962). It isconfirmed as a competent vector ofnumerous plant viruses (Díaz et al.,1990; Duffus, 1987; 1996; Wisler et al.,
1998; Salazar et al., 2000) but it ismore important as a direct pest ofseveral crops. This whitefly was viewedas a minor pest until the early 1980s(Cardona, 1995). Since then,T. vaporariorum has become a key peston dry beans, snap bean, tomato,potato and ornamentals and isconsidered the most important whiteflyspecies in the tropical highlands ofSouth America. Damage is due to thecontinuous sucking of sap from thephloem by nymphs and adults and tothe abundant excretion of honeydewthat falls on leaves and fruits andserves as a substrate for fungi thatblock photosynthesis. Both the yieldand quality of crops are affected. Aseries of trials in Colombia (Cardona etal., 1993; Prada et al., 1993) showedthat high infestation levels (up to450 nymphs/leaf) can reduce the yieldsof common bean by 38% and of snapbean by 54%.
Whitefly problems in snap beanproduction in Colombia have increaseddramatically since the early 1980s andillustrate the need for integrated pestmanagement (IPM) work on whiteflies.In an attempt to reduce pesticide usein whitefly-affected areas, a pilotproject, funded by the InternationalDevelopment Research Centre (IDRC)of Canada, was initiated in theSumapaz region of central Colombia.Surveys showed that all of 286 farmersinterviewed were using insecticides asthe sole method to managewhiteflies. Insecticide use averaged11 applications in a 70-day croppingcycle. Most of the insecticides usedwere organophosphates andcarbamates belonging to toxicologicalcategory I (highly toxic) as defined byMetcalf (1994): those insecticides withacute oral LD50 values of less than50 mg/kg or dermal LD50 values of lessthan 200 mg/kg. Field tests indicatedthat seven of the 10 most widely usedinsecticides were ineffective for whitefly

271
Introduction
control, due to resistance (Cardona,1995). Research showed that simplesanitation measures such as leafroguing and destruction of cropresidues reduced overall whiteflyinfestation levels if implemented on acommunity-wide basis. In addition,selective use of granular insecticidesdelayed the need for the first foliarinsecticide application, enhancingnatural enemy activity. A relativelysimple IPM package based on culturalcontrol and sanitation practices, timelyapplication of effective insecticides andreliance on natural biological controlresulted in a 66% reduction ininsecticide use (Cardona, 1995).Farmers’ participation in the overallprocess was essential for widespreadadoption of the system proposed.However, it was concluded that, beforecontinuing with IPM training andscaling up, it was necessary to verifythat patterns of T. vaporariorumdistribution, reproductive hosts plants,perceptions of the problem andinsecticide use were similar throughoutthe Andean region.
B. tabaci also has beendocumented as a pest of several cropsin Colombia (Posada, 1976; Bolaño,1997) and Ecuador (Merino andVásquez, 1962). Except for sporadicoutbreaks on cotton in Colombia(Alcaraz et al., 1990), this insect wasgenerally regarded as a secondary pestuntil 1993 when serious outbreaksoccurred in the western provinces ofManabí, Guayas, and Los Ríos inEcuador. Mendoza et al. (1995)estimated that vegetable growers in theGuayas region lost US$400,000 peryear as a result of whitefly attacks. Upto 10,000 ha of soybean were destroyedin the Guayas and Los Ríos provinces(Mendoza, 1996). Similar outbreaksoccurred in 1996 in the north-westernDepartments of Sucre, Córdoba andAtlántico in Colombia (Quintero et al.,2001). The change in B. tabaci’s pest
status was attributed to theintroduction into the region of the Bbiotype (Quijije et al., 1995), which wasfirst detected in Colombia in 1997(Quintero et al., 1998).
A New Foundation forWhitefly IPM in the Andes
In view of the increasing importance ofwhiteflies in the Andean highlands, thelaunching in 1997 of a new project for“sustainable integrated management ofwhiteflies as pests and vectors of plantviruses in the tropics” by theSystemwide Programme on IntegratedPest Management was especiallytimely. A sub-project was established,under the co-ordination of the CentroInternacional de Agricultura Tropical(CIAT), to look specifically at “Whitefliesas pests in the tropical highlands ofLatin America”.
The purpose of the diagnosticphase of this sub-project (and of othersaddressing different whitefly problemsaround the world) was to provide asound basis for future IPM efforts bygathering and analysing baseline dataand so being able to characterizeproperly the nature of major whiteflyproblems. Specifically, in the Andeanhighlands, there was a need to describethe patterns of distribution ofT. vaporariorum and B. tabaci,especially in relation to altitude, as wellas the distribution of the newlyintroduced B biotype of B. tabaci.Information on the range ofreproductive host plants andidentification of natural enemies wasalso desired. Additional socio-economicinformation on crop losses, farmers’perception of the problems andpatterns of insecticide use, as well asbiological assessment of levels ofinsecticide resistance, were needed as afoundation for moving forward to morewidespread adoption of IPM.

272
Whiteflies and Whitefly-borne Viruses in the Tropics
Surveys were conducted betweenOctober 1997 and December 1998 inwhitefly-affected areas of Colombia andEcuador while insecticide resistancetesting continued through May 1999.The research was carried out in closecollaboration among staff of CIAT andpartner organizations, the InstitutoNacional Autónomo de InvestigacionesAgropecuarias (INIAP) in Ecuador andthe Corporación Colombiana deInvestigación Agropecuaria(CORPOICA) in Colombia. The methodsused and the results of the work arereported in the following two chapters.
References
Alcaraz, H.; Cardona, C.; Revelo, F.;Revelo, R.; Herrera, M.; Alvarez, A.;Siabato. A. 1990. Plagassecundarias o de incidenciaocasional. In: Bases técnicas para elcultivo del algodón en Colombia.Cuarta edición. FederaciónNacional de Algodoneros, Bogotá,CO. p. 519-524.
Bolaño, R. E. 1997. Determinación deniveles de daño económico deBemisia tabaci en tomate en el nortedel Cesar, Colombia. MIP 46:26-33.
Caballero, R. 1992. Whiteflies(Homoptera: Aleyrodidae) fromCentral America and Colombiaincluding slide-mounted pupal andfield keys for identification, fieldcharacteristics, hosts, distribution,natural enemies, and economicimportance. M.Sc. thesis, KansasState University, USA. 201 p.
Cardona, C. 1995. Manejo de Trialeurodesvaporariorum (Westwood) en frijol enla zona Andina: Aspectos técnicos,actitudes del agricultor ytransferencia de tecnología.Memoria IV Taller Latinoamericanosobre moscas blancas ygeminivirus, 16-18 octubre 1995,Zamorano, HN. CEIBA 36(1):53-64.
Cardona, C.; Rodríguez, A.; Prada, P. C.1993. Umbral de acción para elcontrol de la mosca blanca de losinvernaderos, Trialeurodesvaporariorum (Westwood)(Homoptera: Aleyrodidae), enhabichuela. Rev. Colomb. Entomol.19(1):27-33.
Díaz, M. C.; Pulgarín, J. M.; Saldarriaga,A. V. 1990. Relaciones insecto-patógeno en el problema delamarillamiento de las venas de lapapa. Rev. Colomb. Entomol.16(2):3-14.
Duffus, J. E. 1987. Whitefly transmissionof plant viruses. Curr. Topics VectorRes. 4:73-94.
Duffus, J. E. 1996. Whitefly-borneviruses. In: Gerling, D.; Mayer, R.T.(eds.). Bemisia: 1995. Taxonomy,biology, damage, control andmanagement. Intercept, Andover,GB. p. 252-263.
Mendoza, J. 1996. Qué está pasando conla mosca blanca en el Ecuador?Rev. INIAP 8:8-10.
Mendoza, J.; Valarezo, O.; de López, M.A.; Quijije, R.; Cañarte, E.; Alcaraz,V. 1995. Reporte de Ecuador.Memoria IV Taller Latinoamericanosobre moscas blancas ygeminivirus, 16-18 octubre 1995,Zamorano, HN. CEIBA 36(1):13-15.
Merino, G.; Vásquez, V. 1962.Identificación de algunas de lasnuevas especies de insectoscoleccionadas en Ecuador. Bol.Técnico no. 7, Dirección General deAgricultura y Servicio CooperativoInteramericano de Agricultura,Quito, EC. 35 p.
Metcalf, R. L. 1994. Insecticides in pestmanagement. In: Metcalf, R. L.;Luckmann, W. H. (eds.).Introduction to insect pestmanagement. 3rd edition. JohnWiley, NY, USA. p. 245-284.

273
Introduction
Posada, L. 1976. Lista de insectosdañinos y otras plagas en Colombia.Bol. Técnico no. 43, InstitutoColombiano Agropecuario (ICA).Bogotá, CO. 662 p.
Prada, P. C.; Rodríguez, A.; Cardona, C.1993. Evaluación de un sistema demanejo integrado de la habichuelaen la provincia de Sumapaz(Cundinamarca). Rev. Colomb.Entomol. 19(2):58-63.
Quijije, R.; Mendoza, J.; Gómez, A. 1995.Ciclo biológico de Bemisiaargentifolii en condiciones delaboratorio. Memoria IV TallerLatinoamericano sobre moscasblancas y geminivirus, 16-18octubre 1995, Zamorano, HN.CEIBA 36(1):84.
Quintero, C.; Cardona, C.; Ramírez, D.;Jiménez, N. 1998. Primer registrodel biotipo B de Bemisia tabaci(Homoptera: Aleyrodidae) enColombia. Rev. Colomb. Entomol.24(1-2):23-28.
Quintero, C.; Rendón, F.; García, J.;Cardona, C.; López-Avila, A.;Hernández, P. 2001. Especies ybiotipos de moscas blancas(Homoptera: Aleyrodidae) encultivos semestrales de Colombia yEcuador. Rev. Colomb. Entomol.27(1-2):27-31.
Salazar, L. F.; Muller, G.; Querci, M.;Zapata, J. L.; Owens, R. A. 2000.Potato yellow vein virus: Its hostrange, distribution in SouthAmerica and identification as acrinivirus by Trialeurodesvaporariorum. Ann. Appl. Biol.137:7-19.
Strong, D.; Lawton, J.; Southwood, R.1984. Insects on plants:Community patterns andmechanisms. Blackwell Scientific,Oxford, GB.
Vet, L. E.; van Lenteren, J. C.; Woets J.1980. The parasite-hostrelationship between Encarsiaformosa (Hymenoptera: Aphelinidae)and Trialeurodes vaporariorum(Homoptera: Aleyrodidae). Z. Angew.Ent. 90:26-51.
Winograd, M.; Farrow, A.; Eade, J. 1998.Atlas de indicadores ambientales yde sustentabilidad para AméricaLatina y el Caribe. Versión 1.Centro Internacional de AgriculturaTropical (CIAT)-Progama de lasNaciones Unidas para el MedioAmbiente (PNUMA), Cali, CO.1 CD-ROM.
Wisler, G.; Duffus, J. E.; Liu, H-Y.; Li, R.H. 1998. Ecology and epidemiologyof whitefly-transmittedclosteroviruses. Plant Dis. 82:270-275.

274
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 4.2
Colombia and EcuadorCésar Cardona*, Aristóbulo López-Avila** and
Oswaldo Valarezo***
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** Corporación Colombiana de InvestigaciónAgropecuaria (CORPOICA), Bogotá,Colombia.
*** Instituto Nacional Autónomo deInvestigaciones Agropecuarias (INIAP),Manabí, Ecuador.
As part of the diagnostic phase of theTropical Whitefly Integrated PestManagement (TWF-IPM) Project (seeIntroduction and Chapter 4.1, thisvolume), partners in the sub-projectconcerned with “Whiteflies as pests inthe tropical highlands of Latin America”conducted extensive field surveys inColombia and Ecuador from October1997 to December 1998. The sub-project was co-ordinated by the CentroInternacional de Agricultura Tropical(CIAT) and carried out in collaborationwith the Ecuadorian national researchorganization: the Instituto NacionalAutónomo de InvestigaciónesAgropecuarias (INIAP), and theColombian national researchorganization: the CorporaciónColombiana de InvestigaciónAgropecuaria (CORPOICA). The presentchapter summarizes the diagnosis andcharacterization of problems caused bythe greenhouse whitefly Trialeurodesvaporariorum (Westwood) as a pest ofannual field crops in Colombia andEcuador, and its relationship to thesweetpotato whitefly Bemisia tabaci(Gennadius).
The surveys focused on the tropicalhighlands (1000-3000 m above sealevel) along the Andean corridor ofColombia and Ecuador (0° 30'-11°N,73°-78° 30'W). Within this zone,surveys covered areas representingabout 97% of whitefly-affected areas(Figure 1). Representative samplingalso was carried out in the lowlandsand mid-altitudes in order to constructan altitudinal gradient from sea levelup to 3000 m. Samples were collectedin mid-altitude valleys within thehighland tropics of Colombia(400-1000 m above sea level). And, inthe tropical lowlands (less than 400 mabove sea level), surveys wereconducted along the northern (Atlantic)coast of Colombia (7° 30'-11°N,72°-77°W) and the Pacific coast ofEcuador (0°-1° 30'S, 79°-80° 30'W),including the Galápagos Islands ofEcuador (2°-3°S, 90°-90° 30'W) in thePacific Ocean. INIAP staff conductedfield surveys and interviews withfarmers along the coast of Ecuador.CIAT personnel sampled the northernhighlands of Ecuador. CORPOICA andCIAT staff conducted fieldwork inColombia.
Surveys followed the standardizedmethodology agreed among the projectpartners. Twenty survey areas weredefined in Colombia and Ecuadorbased on the importance of annualcrops and previous knowledge of

275
Colombia and Ecuador
Figure 1. Survey area and distribution of whitefly species (Bemisia tabaci and Trialeurodesvaporariorum) in Colombia and Ecuador. Uncoloured areas were not sampled.
Caribbean Sea
Pacific OceanColombia
Ecuador
B biotype B. tabaci
A and B biotypes B. tabaci andT. vaporariorum
B biotype B. tabaci andT. vaporariorum
T. vaporariorum
A and B biotypesB. tabaci
whitefly incidence. Nine areas werechosen to represent the lowlands alongthe Pacific coast of Ecuador and theAtlantic coast of Colombia where cotton(Gossypium hirsutum L.), soybean(Glycine max [L.] Merr.) and tomato(Lycopersicon esculentum Mill.) aregrown on large farms that occupy asignificant proportion of theagricultural land. Eleven areas werechosen to represent the highlands ofnorthern Ecuador and the main
agricultural areas along the threemountain ranges in Colombia.Highland areas in both countries areoccupied by small-scale farmers whogrow common and snap bean(Phaseolus vulgaris L.), tomato, potato(Solanum tuberosum L.) and otherhorticultural crops for consumptionand sale. Between 10 and 20 data setswere collected and processed in eachsurvey area. A complete data setconsisted of a producer questionnaire
N

276
Whiteflies and Whitefly-borne Viruses in the Tropics
Figure 2. Composition of whitefly populationsin three major ecological zones, asdefined by altitude (masl = metresabove sea level), in Colombia andEcuador.
100
80
60
40
20
0
8.5
91.5
13.7
25.5
60.8
0 0
100
Tropicallowlands
(<400 masl)
Mid-altitudevalleys (400-1000 masl)
0
Tropicalhighlands
(>1000masl)
Bemisiatabaci A
Bemisiatabaci B
Trialeurodesvaporariorum
and biological samples of whiteflies andnatural enemies from one of thatproducer’s fields. Three hundred andtwenty-five data sets were processed,based on visits and interviews with 274farmers and 168 extension agents,private technical assistants, insecticidesalesmen and government officers incharge of rural development projects.Insecticide resistance was alsoassessed in whiteflies from 40 sites, asdetailed in Chapter 4.3 (this volume).
Whiteflies and natural enemieswere identified at CIAT and whiteflybiotypes determined using esterasebanding patterns in polyacrylamide gelelectrophoresis (PAGE) and randomamplified polymorphic DNA/polymerase chain reaction (RAPD-PCR)analysis (Quintero et al., 1998).Biological samples for whitefly speciesand biotype identification were taken at215 sites; 74 samples for identificationof natural enemies were collected.
Whitefly populations were sampledin 14 crops. In the tropical highlands,emphasis was placed on common bean(73% of samples taken), tomato (18%)and potato (7%). In mid-altitudevalleys, most samples were taken ontomato (58%) and common bean (18%).Cucurbits (38%), tomato (31%), pepper(Capsicum annuum L.) (10%) andeggplant (Solanum melongena L.) (5%)were the crops most intensivelysurveyed in the tropical lowlands. Yieldlosses due to the greenhouse whiteflyon snap beans were measured.
Increased BiologicalUnderstanding
Species
An analysis of whitefly speciescomposition along an altitudinalgradient suggests that T. vaporariorumand B. tabaci occupy well-defined
niches and habitats (Figure 2). In thetropical lowlands, 91.5% of samplescollected were identified as B. tabaci,whereas 100% of the whitefliescollected in the tropical highlands wereT. vaporariorum. The species co-exist inall mid-altitude valleys where sampleswere taken, with T. vaporariorumpredominating. Thus, in Colombia andEcuador, T. vaporariorum occursbetween 780 and 2830 m above sealevel and is the only species presentabove 1600 m, whereas B. tabaci is theonly species present at lower elevations(500 m or less) (Quintero et al., 2001).
This distribution of the two majorwhitefly species may be related toclimatic conditions, especiallytemperature regimes, which aredetermined in the tropics principally byelevation. As much of the agriculturalproduction in the Andes isconcentrated in mid-altitude valleys,our findings clearly show the need for acorrect identification of the whiteflyspecies concerned. If it is assumed, forexample, that B. tabaci is the majorpest in mid-altitude valleys andhighlands, IPM packages will be illtargeted. Our findings also dispel thenotion, prevalent among technical
Wh
itef
ly s
pec
ies
(%)

277
Colombia and Ecuador
B. tabaci biotype BTv A B
h k Ch k
TvAB 100
M1 2 3 4 5 6 7 8 9 151210 11 13 14
B. tabaci biotype BTv A B
h k Ch k
TvAB 100
M1 2 3 4 5 6 7 8 9 151210 11 13 14
Figure 3. RAPD patterns obtained with primer OPA-04 for a sample of whiteflies collected in thenorthern coastal zone of Colombia. Lane 1: Trialeurodes vaporariorum (control);lane 2: Bemisia tabaci biotype A (control); lane 3: B. tabaci biotype B (control); lanes 4-12:B. tabaci biotype B (field sample); lane 13: B. tabaci biotype B (control); lane 14: B. tabacibiotype A (control); lane 15: T. vaporariorum (control); lane M: marker (1 kb ladder).
assistants and extension personnel,that B. tabaci is the key pest in thehighlands and mid-altitude valleys ofthe Andean zone. T. vaporariorum is thekey pest in this region.
Biotypes
No evidence for the existence ofdifferent biotypes of T. vaporariorumwas found in more than 400 samplesexamined by RAPD-PCR. Thus it isconcluded that there is no basis forcontinued biotype analysis of thegreenhouse whitefly within the contextof the TWF-IPM Project.
Both A and B biotypes of B. tabaciwere detected among the 214 samplesevaluated by RAPD-PCR (Figure 3). TheB biotype is the only one of importancealong the coast of Ecuador. It ispresent also throughout the northern(Atlantic) coast of Colombia and in theDepartments of Valle, Huila, Tolimaand Cundinamarca. The A biotype of
B. tabaci was detected only in fourdepartments in Colombia: Córdoba,Sucre, Valle and Huila (Figure 2). Theprevalence of the B biotype in thecoastal areas of Ecuador andColombia represents a serious threatto agriculture in the region. Recentwork (CIAT, unpublished results) hasshown that the presence of the Bbiotype can also be a major threat toagriculture in mid-altitude valleyssuch as the Cauca Valley inColombia. In a survey conducted in2003, the B biotype was present in63% of samples taken (as opposed to11.5% in 1997). The increasedincidence of the B biotype was alsoevidenced by the commonoccurrence of such physiologicaldisorders as irregular ripening oftomato, and silver leaf symptoms onsquash (Cucurbita moschataDuchesne). This underlines the needfor continuous monitoring of whiteflyspecies composition.

278
Whiteflies and Whitefly-borne Viruses in the Tropics
Reproductive hosts
In Colombia, T. vaporariorum wasfound breeding on common bean andsnap bean, tomato, potato, squash,cucumber (Cucumis sativus L. var.sativus), eggplant, pepper, pea (Pisumsativum L.), tobacco (Nicotiana tabacumL.), cabbage (Brassica oleracea L.),coriander (Coriandrum sativum L.) andseveral species of ornamentals. In mid-altitude areas, T. vaporariorum alsoreproduces on cotton and soybean. AllT. vaporariorum samples in Ecuadorwere taken on common bean andtomato. Snap bean and common beanare by far the most importantreproductive hosts for T. vaporariorumin Colombia and Ecuador, followed inorder of decreasing importance bytomato and potato (Table 1).
In Colombia, the A biotype ofB. tabaci was recorded breeding onsoybean, tobacco, tomato, cotton,poinsettia (Euphorbia pulcherrima Willd.ex Klotzch), broccoli (Brassica oleraceaL. var. italica Plenck) and eggplant. Inboth Colombia and Ecuador, theB biotype was found reproducing oncotton, tomato, eggplant, squash,pepper, cucumber, common bean,snap bean, soybean, tobacco, cabbage,lettuce (Lactuca sativa L. var. capitataL.), sweetpotato (Ipomoea batatas [L.]Lam.), melon (Cucumis melo L.),watermelon (Citrullus lanatus [Thunb.]
Matsum. & Nakai), pumpkin (Cucurbitaficifolia Bouché), lima bean (Phaseoluslunatus L. var. lunatus), forage Arachis(Arachis pintoi Krapov. & W. Gregory),grape (Vitis vinifera L.), poinsettia andadditional Brassica species. Table 1shows the main crops affected byB. tabaci.
In greenhouse studies on thecomparative biology of biotypes A andB on 20 cultivated host plants, theB biotype bred faster than theA biotype on cauliflower (Brassicaoleracea L. var. botrytis L.), cabbage,squash and tomato. Percentageemergence of the B biotype wassignificantly higher on 13 out of the20 host plants studied; the A biotypedid not breed on squash or beet (Betavulgaris L. subsp. vulgaris). The Bbiotype did more damage to lettuce,squash, cabbage, cauliflower,cucumber, Brassica sp., soybean andwatermelon than did the A biotype.Snap bean was affected also andsuffered significant reductions inbiomass as a result of damage by the Bbiotype. Common bean, on the otherhand, was not as severely affected asother host plants.
The B biotype successfullycolonized plants of four Cucurbitaspecies tested. The A biotype did notsurvive to the adult stage on pumpkin
Table 1. Main crops serving as reproductive hosts of whiteflies in major agro-ecological zones ofColombia and Ecuador.
Whitefly species Tropical lowlands Mid-altitude valleys Tropical highlands
A biotype of Bemisia tabaci Eggplant Tobacco -Tomato Soybean -
B biotype of B. tabaci Squash Squash -Cotton Cotton -Eggplant Tobacco -Brassica sp. - -Tomato - -
Trialeurodes vaporariorum - Common bean Common bean - Snap bean Snap bean - Tomato Tomato - - Potato

279
Colombia and Ecuador
and squash. On Cucurbita mixtaPangalo (= Cucurbita argyrosperma C.Huber subsp. argyrosperma) andCucurbita pepo L., the B biotypereached the adult stage in ca. 20 days,33% faster than the A biotype. Theability of the B biotype to successfullycolonize hosts that are not attacked bythe A biotype and its ability toreproduce faster on certain host plantsare important diagnostic tools (Brownet al., 1995; Schuster et al., 1996).When C. moschata, C. ficifolia andC. pepo plants were infested with eitherA or B biotype adults, full expression ofsilverleaf symptoms, as described byYokomi et al. (1990) and Costa andBrown (1991), were obtained 21 daysafter infestation with the B biotype. Nosilvering was observed on any plantinfested with the A biotype.
Apart from silverleaf, othersymptoms of B biotype attack thathave been described in the literaturewere observed in numerous differentfields: uneven ripening of tomato(Schuster et al., 1996; Polston andAnderson, 1997), chlorotic pods andpetioles on snap beans (Hassan andSayed, 1999), white stem streaking ofBrassica spp. and phytotoxic disorderson poinsettia and lettuce (Brown et al.,1995).
Natural enemies
Survey work confirmed that Amitusfuscipennis MacGown and Nebeker isthe most common parasitoid ofT. vaporariorum in the highlands ofColombia and Ecuador. Other naturalenemies of T. vaporariorum recordedwere the parasitoids Encarsia hispidaDeSantis, Encarsia pergandiellaHoward, Encarsia nigricephala Dozierand Encarsia sp., and the predatorsDelphastus pusillus Leconte,Hippodamia convergens (Guérin-Ménenville), Cycloneda sanguinea (L.),Coleomegilla maculata (DeGeer) andChrysoperla sp. The fungi Verticillium
lecanii (Zimm.) and Paecilomycesfumosoroseus (Wize) were found in theDepartments of Antioquia, CaucaValley and Cundinamarca in Colombia(López-Avila et al., 2001).
Additional studies by Manzano etal. (2000) demonstrated thatA. fuscipennis is abundant on commonbean crops where no chemicaltreatments have been made, andremains active even after heavypesticide use. Some studies have beenconducted on the biology or efficacy ofthis species as a natural enemy ofT. vaporariorum in the Andean zone(Márquez and Valencia, 1991; Medinaet al., 1994; Manzano et al., 1999).Recent results (Manzano et al., 2000)suggest that A. fuscipennis couldpotentially be a good biological controlagent of T. vaporariorum in Colombia,in environments that are not overly dryor warm. However, as the parasitoidseems to be negatively affected bycertain combinations of climaticconditions, its limitations need to befurther studied in order to ascertain itstrue potential as a natural enemy.
The entomopathogen V. lecanii hasbeen tested under laboratory and fieldconditions in Colombia (González andLópez-Avila, 1997) with variable,usually poor, results.
The natural enemies of B. tabaci,found at many different sites inColombia and Ecuador, included theaphelinid parasitoids E. hispida,E. pergandiella, E. nigricephala,Encarsia sp. and Eretmocerus sp., aswell as the predators D. pusillus andChrysoperla sp. The relative importanceand efficiency of these natural enemieshas not been assessed.
Yield losses
Three trials aimed at measuring lossesdue to T. vaporariorum on snap beanwere conducted during 1998 and 1999.

280
Whiteflies and Whitefly-borne Viruses in the Tropics
The establishment of these trialscoincided with the appearance inColombia of another serious pest ofcommon bean, the melon thrips, Thripspalmi Karny. As a result, the trials hadto be redesigned to account for theincidence of the new pest. In largereplicated plots, different insecticideregimes were established to measureyield responses and to partition lossesdue to thrips and whiteflies. The latterwas achieved by using spinosad(Tracer®) Dow Chemical Co.) as aselective insecticide for T. palmi controland buprofezin (Applaud®, ICI) inmixture with monocrotophos(Azodrin®, Shell Co.) for whiteflyexclusion. The mean loss due to T.palmi was 4.1 t/ha or 46% of thepotential yield. T. vaporariorum causedlosses of 2.1 t/ha or 23% of potentialyield (Rendón et al., 2001).
Increased Socio-EconomicUnderstanding
Farmers’ perceptions of thewhitefly problem
From 88% to 100% of farmerssurveyed were male; 41% to 63% wereowners who had been on their farmsfor over 5 years, planting an average of1.2 ha with three or more cropsplanted in succession through the year.Table 2 shows that most farmers knewthe insects; they were able to identifythem as whiteflies and seemed to beaware of whitefly-related problems. Thewhitefly was regarded as an endemicpest by about 49% of farmers in thetropical highlands of Colombia andEcuador, by 23% in mid-altitudevalleys (400-1000 m) in Colombia andby 82% in the tropical lowlands ofColombia and Ecuador. A minorityclaimed to be able to predict whiteflyepidemics, which they usuallyassociated with dry weather. Whiteflyproblems in the survey area are
distributed extensively, the insectsbeing present in 72% of farms visited.The lowest incidence was in theDepartments of Cauca and CaucaValley, Colombia, where the insectswere present in 46% of farms visited.
Whiteflies ranked as the mostimportant pest of common bean in50% of the sites visited, and as the keypest in 45% of the tomato farms thatwere surveyed. A relatively smallpercentage of farmers gave whitefliesthe highest rating as pests whencompared with other insects on thefarm (for example, T. palmi on commonbean or fruit borers on tomato) but asignificant proportion thought thatwhiteflies could reduce yields by 50%or more. Up to 30% of melon growersand 34% of those planting tomatoindicated that they have abandonedcrops as a result of devastating whiteflyattacks. Most farmers (76% of thosegrowing cucurbits, 80% of thosegrowing tomato) could identify theinsects as pests but very few associatedthe presence of whiteflies with virusdiseases (Table 2).
Technicians’ perceptions of thewhitefly problem
Virtually all extension agents andtechnical assistants interviewedconsidered that whiteflies were a majorproblem on tomato and common bean.Most (80%-93%) regarded whiteflies aspests, while 27%-39% considered thatwhiteflies act as both pests andvectors. Up to 70% thought that thesituation in most crops had changed asa result of whitefly incidence. Very fewwere aware of new technologies to solvethe problem, considered that the areaplanted to several crops had beenreduced or that insecticide use hadincreased as a result of whiteflyincidence. All acknowledged that usinginsecticides was the onlyrecommendation they could make tofarmers.
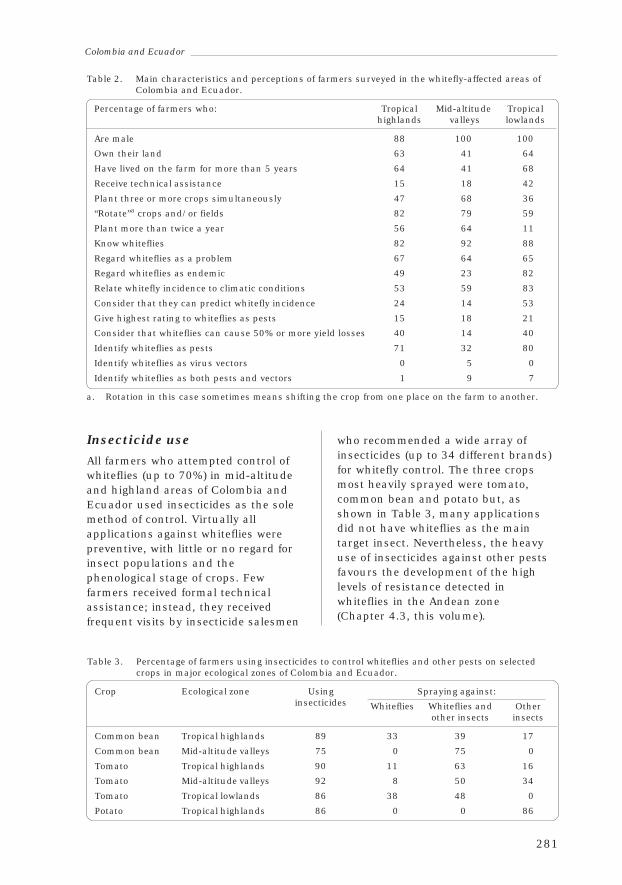
281
Colombia and Ecuador
Insecticide use
All farmers who attempted control ofwhiteflies (up to 70%) in mid-altitudeand highland areas of Colombia andEcuador used insecticides as the solemethod of control. Virtually allapplications against whiteflies werepreventive, with little or no regard forinsect populations and thephenological stage of crops. Fewfarmers received formal technicalassistance; instead, they receivedfrequent visits by insecticide salesmen
who recommended a wide array ofinsecticides (up to 34 different brands)for whitefly control. The three cropsmost heavily sprayed were tomato,common bean and potato but, asshown in Table 3, many applicationsdid not have whiteflies as the maintarget insect. Nevertheless, the heavyuse of insecticides against other pestsfavours the development of the highlevels of resistance detected inwhiteflies in the Andean zone(Chapter 4.3, this volume).
Table 2. Main characteristics and perceptions of farmers surveyed in the whitefly-affected areas ofColombia and Ecuador.
Percentage of farmers who: Tropical Mid-altitude Tropicalhighlands valleys lowlands
Are male 88 100 100
Own their land 63 41 64
Have lived on the farm for more than 5 years 64 41 68
Receive technical assistance 15 18 42
Plant three or more crops simultaneously 47 68 36
“Rotate”a crops and/or fields 82 79 59
Plant more than twice a year 56 64 11
Know whiteflies 82 92 88
Regard whiteflies as a problem 67 64 65
Regard whiteflies as endemic 49 23 82
Relate whitefly incidence to climatic conditions 53 59 83
Consider that they can predict whitefly incidence 24 14 53
Give highest rating to whiteflies as pests 15 18 21
Consider that whiteflies can cause 50% or more yield losses 40 14 40
Identify whiteflies as pests 71 32 80
Identify whiteflies as virus vectors 0 5 0
Identify whiteflies as both pests and vectors 1 9 7
a. Rotation in this case sometimes means shifting the crop from one place on the farm to another.
Table 3. Percentage of farmers using insecticides to control whiteflies and other pests on selectedcrops in major ecological zones of Colombia and Ecuador.
Crop Ecological zone Using Spraying against:insecticides Whiteflies Whiteflies and Other
other insects insects
Common bean Tropical highlands 89 33 39 17
Common bean Mid-altitude valleys 75 0 75 0
Tomato Tropical highlands 90 11 63 16
Tomato Mid-altitude valleys 92 8 50 34
Tomato Tropical lowlands 86 38 48 0
Potato Tropical highlands 86 0 0 86

282
Whiteflies and Whitefly-borne Viruses in the Tropics
Up to 57% of insecticideapplications were reportedly made withorganophosphate insecticides (OPs)alone. However, OPs also were reportedas being used in mixtures withcarbamates and pyrethroids (Table 4)to the extent that 80%-100% of theapplications had OPs as ingredients.Given the high levels of insecticideresistance to OPs and pyrethroids thatwere detected in our studies, it is notsurprising that a significant proportionof farmers (up to 60%) were beginningto use novel insecticides. This wasmost evident in tomato-growing areasin the tropical lowlands and in the mid-altitude valleys of Colombia.
Farmers had easy access to at least36 different active ingredients and40 commercial brands. Mostinsecticides used belong to the
toxicological category I (extremelydangerous), as defined by Metcalf(1994) and are used with minimalsafety precautions for handling andapplication. In the survey, the mostwidely used group of pesticides wereOPs, with 40% to 50% share of theuse, followed by carbamates andpyrethroids (Table 5). Novel productssuch as insect growth regulators,juvenoids and nicotinoids werereported as becoming popular(Rodríguez and Cardona, 2001). Alsoused were soaps, oils and botanicals.
Insecticides were reportedly appliedto tomato and snap bean as often asthree times a week (Rodríguez andCardona, 2001). The number ofapplications per cropping season variedamong regions but it is interesting tonote that 24% of farmers surveyedmade 10 or more applications (Table 6).
Most farmers used very low(0.2 cc/L) or very high (4.8 cc/L)dosages of concentrate, well below orabove the 2 cc/L average dosageusually recommended bymanufacturers. Up to 71% of farmerssurveyed in Colombia and 55% inEcuador took their own decisions ontiming of applications as opposed tothose who have access to technicalassistance. More than half reportedspraying on a calendar basis,regardless of infestation levels, andrelatively few had received any kind oftechnical assistance.
Table 4. Percentage of farmers usingorganophosphates (OP), carbamates(CAR) and pyrethroids (PYR) tocontrol whiteflies in major ecologicalzones of Colombia and Ecuador.
Insecticide Tropical Mid- Tropicalgroup highlands altitude lowlands
valleys
OP 55 33 57
CAR 4 0 2
PYR 4 0 8
OP + CAR 5 45 5
OP + PYR 24 22 25
CAR + PYR 0 0 0
OP + CAR + PYR 8 0 3
Table 5. Percentage participation of organophosphate, carbamate and pyrethroid insecticides inchemical control of whiteflies in selected tropical highland areas of Ecuador and Colombia.
Insecticide category Northern Ecuador– Departments of Department ofsouthern Colombia Cauca and Cauca Valley, Antioquia,
Colombia Colombia
Organophosphates 40 50 50
Carbamates 22 14 25
Pyrethroids 22 9 13
Novel productsa 16 23 12
Others (soap, oils, etc.) 0 4 0
a. Includes insect growth regulators, juvenoids and nicotinoids.

283
Colombia and Ecuador
Conclusions
In general, the results of both thebiological surveys and the interviewswith producers and their technicaladvisors underline the need for IPMresearch and implementation. A highlydangerous situation has developed forsmall-scale farmers in Colombia andEcuador over the past 15 years:commercial pressures and aninadequate knowledge base haveinfluenced farmers to adopt a“chemical culture”, involving excessivedependence on pesticides. Thesituation has been aggravated by theintroduction into the region of the Bbiotype of B. tabaci, a major pest ofannual crops. The results, theirconsequences and recommendationsfor further action are discussed inmore detail in Chapter 4.4.
References
Brown, J. K.; Frohlich, D. R.; Rosell, R. C.1995. The sweetpotato or silverleafwhiteflies: Biotypes of Bemisiatabaci or a species complex? Ann.Rev. Entomol. 40:511-534.
Costa, H. S.; Brown, J. K. 1991. Variationin biological characteristics andesterase patterns amongpopulations of Bemisia tabaci, andthe association of one populationwith silverleaf symptom induction.Entomol. Exp. Appl. 61:211-219.
González, J. G.; López-Avila, A. 1997.Evaluación de cepas nativas deVerticillium lecanii (Zimm.) en elcontrol de la mosca blanca de losinvernaderos Trialeurodesvaporariorum (Westwood). Rev.Colomb. Entomol. 23(1-2):25-30.
Hassan, A. A.; Sayed S. F. 1999. Chloroticpod: A new physiological disorder ofgreen-podded snap beans(Phaseolus vulgaris L.) associatedwith silverleaf whitefly infestation.Egypt. J. Hortic. 26:213-228.
López-Avila, A.; Cardona, C.; García, J.;Rendón, F.; Hernández, P. 2001.Reconocimiento e identificación deenemigos naturales de moscasblancas (Homoptera: Aleyrodidae)en Colombia y Ecuador. Rev.Colomb. Entomol. 27(3-4):137-141.
Manzano, M. R.; van Lenteren, J. C.;Cardona, C. 1999. Some biologicalcharacterisitics of Amitusfuscipennis MacGown and Nebeker(Hymenoptera: Platygasteridae),parasitoid of the greenhousewhitefly. IOBC Bull. 23(1):169-172.
Manzano, M. R.; van Lenteren, J. C.;Cardona, C.; Drost, Y. C. 2000.Developmental time, sex ratio, andlongevity of Amitus fuscipennisMacGown and Nebeker(Hymenoptera: Platygasteridae) onthe greenhouse whitefly. Biol.Control 18(2):94-100.
Márquez, M. L.; Valencia, S. A. 1991.Evaluación de Encarsia formosaGahan y Amitus fuscipennisMacGown and Nebeker, en elcontrol de Trialeurodesvaporariorum (Westwood) encrisantemo (Chrysanthemummorifolium Rainat.). B.Sc. thesis,Universidad Nacional de Colombia,Medellín, CO. 132 p.
Table 6. Patterns of insecticide use forwhitefly control in Colombia andEcuador.
Number of insecticide Percentageapplications per cropping of farmers
seasona
1-3 34.8
4-6 25.0
7-9 8.8
10 7.4
> 10 24.0
a. The mean number of applications perseason against the B biotype of Bemisiatabaci in the tropical lowlands of Colombiaand Ecuador is 6.5.

284
Whiteflies and Whitefly-borne Viruses in the Tropics
Medina, P. S.; Saladarriaga, A. V.; Pérez,G. 1994. Biología de Amitusfuscipennis MacGown and Nebeker,bajo tres condiciones ecológicas, enRionegro (Antioquia). Rev. Colomb.Entomol. 20(3):143-148.
Metcalf, R. L. 1994. Insecticides in pestmanagement. In: Metcalf, R. L.;Luckmann, W. H. (eds.).Introduction to insect pestmanagement. 3rd edition. JohnWiley, NY, USA. p. 245-284.
Polston, J. E.; Anderson P. K. 1997. Theemergence of whitefly-transmittedgeminiviruses in tomato in thewestern hemisphere. Plant Dis.81:1358-1369.
Quintero, C.; Cardona, C.; Ramírez, D.;Jiménez, N. 1998. Primer registrodel biotipo B de Bemisia tabaci(Homoptera: Aleyrodidae) enColombia. Rev. Colomb. Entomol.24(1):23-28.
Quintero, C.; Rendón, F.; García, J.;Cardona, C.; López-Avila, A.;Hernández, P. 2001. Especies ybiotipos de moscas blancas(Homoptera: Aleyrodidae) encultivos semestrales de Colombia yEcuador. Rev. Colomb. Entomol.27(1-2):27-31.
Rendón, F.; Cardona, C.; Bueno, J. 2001.Pérdidas causadas por Trialeurodesvaporariorum (Homoptera:Aleyrodidae) y Thrips palmi(Thysanoptera: Thripidae) enhabichuela en el Valle del Cauca.Rev. Colomb. Entomol. 27(1-2):39-43.
Rodríguez, I.; Cardona, C. 2001.Problemática de Trialeurodesvaporariorum y Bemisia tabaci(Homoptera: Aleyrodidae) comoplagas de cultivos semestrales en elValle del Cauca. Rev. Colomb.Entomol. 27(1-2):21-26.
Schuster, D. J.; Stansly, P. A.; Polston J.E. 1996. Expressions of plantdamage by Bemisia. In: Gerling, D.;Mayer, R.T. (eds.). Bemisia: 1995.Taxonomy, biology, damage, controland management. Intercept,Andover, GB. p. 153-165.
Yokomi, R. K.; Hoelmer, K. A.; Osborne, L.S. 1990. Relationships between thesweetpotato whitefly and thesquash silverleaf disorder.Phytopathology 80:895-900.

285
Insecticide Resistance in Colombia and Ecuador
CHAPTER 4.3
Insecticide Resistance in Colombiaand Ecuador
César Cardona*, Francisco Rendón*,Isaura Rodríguez* and Aristóbulo López-Avila**
Farmer interviews conducted as part ofextensive on-farm surveys for thediagnostic phase of the TropicalWhitefly Integrated Pest Management(TWF-IPM) Project (Rodríguez andCardona, 2001; Chapter 4.1, thisvolume) showed that insecticide useagainst whiteflies in Colombia andEcuador is excessive. In the tropicalhighlands and mid-altitude valleys,farmers spray their crops 5 to 6 timeson average to control Trialeurodesvaporariorum (Westwood). The meannumber of applications against the Bbiotype of Bemisia tabaci (Gennadius)in the tropical lowlands of Colombiaand Ecuador was estimated at6.5. Over-reliance on insecticides forwhitefly control is so widespread that30% of 325 farmers interviewedreported that they make more than10 applications per cropping season.The frequency of applications in manycases is as high as two to three timesper week. Most farmers complainedabout the limited control achieved byconventional insecticides and manyfarmers are now using novelinsecticides such as buprofezin,pyriproxyfen, diafenthiuron andimidacloprid reportedly with better
results. However, the 10 most widelyused insecticides identified in thesurveys comprised nine conventionalproducts—dimethoate, carbofuran,chlorpyriphos, methamidophos,methomyl, profenofos, monocrotophos,cypermethrin and malathion—and onlyone of these novel insecticides,imidacloprid. The high toxicity ofseveral of the conventional products inwidespread use raises concerns overboth human and environmental health,underlining the need for alternativeapproaches based on IPM.
Insecticide resistance in whitefliesis well documented as a widespreadphenomenon in those countries inwhich monitoring of resistance hasbeen conducted (Horowitz and Ishaaya,1996). Recent reviews on the subjectare those of Dittrich et al. (1990), Cahillet al. (1996b), Denholm et al. (1996)and Palumbo et al. (2001). In theAndean zone, Buitrago et al. (1994)detected high levels of resistance tocypermethrin and deltamethrin inpopulations of T. vaporariorum from thecentral highlands of Colombia.Resistance to organophosphates andcarbamates was not as high, andranged from low to moderate. Cardonaet al. (1998; 2001) reported that the Bbiotype of B. tabaci, the main whiteflyspecies in the lowlands of Colombia,was highly resistant to methamidophosand methomyl.
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** Corporación Colombiana de InvestigaciónAgropecuaria (CORPOICA), Bogotá,Colombia.

286
Whiteflies and Whitefly-borne Viruses in the Tropics
A more general account of whiteflyproblems in the high Andes, mid-altitude valleys and lowlands ofColombia and Ecuador is presentedelsewhere (Chapter 4.2, this volume),as are the results of biological surveysand farmer questionnaires, intended tocharacterize whitefly problems in theseareas. Here, a summary is presented offindings on the resistance of whitefliesto conventional insecticides in thesurvey areas. Also included arebaseline responses of T. vaporariorumadults to monocrotophos, lamda-cyhalothrin, bifenthrin, carbosulfan,carbofuran, thiamethoxam andimidacloprid. Baseline responses of T.vaporariorum nymphs to buprofezin,diafenthiuron and imidacloprid areincluded. The knowledge of whiteflyproblems synthesized in these studiesis intended to provide a sound basis forpursuing IPM approaches in theAndean region of Latin America.
Methods and Materials
Experimental approach andlocations
Field surveys and insecticide resistancetesting were carried out betweenOctober 1997 and May 1999. Inaccordance with the common researchmethods agreed among the projectpartners, the vial technique wasadopted to establish baseline data inthe laboratory and for field assessmentof resistance to conventionalinsecticides. The technique involvesassessing the rate of mortality of asample of whiteflies within a vial coatedwith a known concentration ofinsecticide (Plapp et al., 1990; Cahilland Hackett, 1992). Baseline data forimidacloprid were obtained followingthe methodology developed by Cahill etal. (1996a).
Laboratory work to establishbaseline data and select diagnosticdosages was conducted at theheadquarters of the CentroInternacional de Agricultura Tropical(CIAT) in Cali, Colombia. Averagelaboratory conditions were 24 °C and75%-80% relative humidity (RH).
Field monitoring extended fromnorthern Ecuador (Imbabura andCarchi provinces) to the north-easterndepartment of Guajira in Colombia.With the exception of coastal areas inEcuador, all major areas affected bywhiteflies were covered. Sites werechosen according to previous data onheavy insecticide use but care wastaken not to perform tests in areas withmixed populations of whitefly species,as indicated in biological surveyscarried out in parallel (Chapter 4.2,this volume). Thus, no insecticideresistance work was conducted in mid-altitude valleys where mixedpopulations of B. tabaci andT. vaporariorum had been recorded. Intotal, insecticide resistance wasmeasured at 40 sites in Colombia andEcuador. Between two and fourinsecticide resistance tests wereconducted in each target area.Resistance of T. vaporariorum collectedfrom common bean (Phaseolus vulgarisL.), tomato (Lycopersicon esculentum L.)and potato (Solanum tuberosum L.) wasmeasured in highland areas along theAndean corridor from northernEcuador to northern Colombia.Resistance of B. tabaci biotype B fromtomato, eggplant (Solanum melongenaL.), squash (Cucurbita moschataDuchesne) and cotton (Gossypiumhirsutum L.) was measured in lowlandareas of the region known as the“northern coast” of Colombia and inthe Cauca Valley (a mid-altitude valley).

287
Insecticide Resistance in Colombia and Ecuador
Baseline data and diagnosticdoses
Adult whiteflies obtained from separatemass rearings of T. vaporariorum andB. tabaci biotype A, maintained at CIATfor over 10 years (without exposure toinsecticides) were used to obtainbaseline data. Technical gradeinsecticides were dissolved in acetoneand 250 µL samples of the solutionwere pipetted into each 20-mL vial. Theacetone was allowed to evaporatecompletely before introducing the testinsects. Insecticides and dosages testedwith T. vaporariorum weremethamidophos (16.0, 8.0, 4.0, 2.0,1.0 and 0.5 µg/vial), methomyl(2.5, 0.5, 0.1 and 0.02 µg/vial),cypermethrin (100.0, 50.0, 25.0,12.5 and 6.25 µg/vial) and carbofuran(10.0, 3.3, 1.1, 0.33 and 0.11 µg/vial).Insecticides and dosages tested withthe A biotype of B. tabaci weremethomyl (25.0, 6.25, 1.56, 0.39 and0.10 µg/vial), methamidophos (16.0,4.0, 2.0, 1.0 and 0.5 µg/vial) andcypermethrin (450.0, 150.0, 50.0,16.7 and 5.6 µg/vial). Five replicates of20 insects were tested for each dosage.Acetone-coated vials were used aschecks (controls). Mortality wasrecorded 6 hours after the adults wereplaced in the vials.
To generate baseline data forimidacloprid and thiamethoxam(Rodríguez et al., 2003), petioles ofexcised common bean leaves wereimmersed in aqueous solutions ofcommercial grade imidacloprid (10.0,5.0, 2.5, 1.25, 0.625 and 0.312 ppm)for 48 hours to allow the leaves to takeup the insecticide solution. Leaf discswere then cut and placed on agar inpetri dishes, one leaf disk per petri dish(Cahill et al., 1996a) and infested with20 T. vaporariorum adults per unit.Discs from leaves whose petioles hadbeen immersed in distilled water wereused as checks. Five replicates (units)per dosage were used. Mortality was
recorded 6 hours after introduction ofthe adults into the units.
To generate baseline data withnymphs, we collected common beanleaves infested with pupae in each ofthe test sites. The infested material wasthen confined to cages. Adultsemerging from these pupae werecollected with an aspirator andintroduced in clip cages attached toleaves of common bean seedlings. Theadults (10 per clip cage) were allowedto oviposit for 24 hours. After hatchingof the eggs, the area of the leaf infestedwith first instar nymphs was markedand the number of individuals thusobtained was recorded. Infestedseedling leaves were then dipped for5 s in 100 mL of the desiredconcentration of each of theinsecticides tested. Formulatedmaterial was used in all tests and thedesired concentrations were preparedby dilution with distilled water.Controls were dipped in distilled water.Treated plants were kept in a rearingchamber at 24 oC, 75%-80% RH andleft undisturbed until adult emergenceoccurred. The number of survivingindividuals in each treatment wasrecorded. Insecticides and dosagestested were buprofezin (100, 25, 6.2,1.5, 0.3, 0.09 and 0.02 ppm),diafenthiuron (1000, 300, 100, 30, 10,3, 1 and 0.3 ppm) and imidacloprid,(1000, 300, 100, 30, 10, 3 and 1 ppm).Four replicates were tested for eachdosage.
Percentage mortality was correctedusing the Abbott formula (Busvine,1971). Tests in which check mortalitywas higher than 10% were notincluded in the analysis. Results weresubjected to probit analysis (SASInstitute, 1988). Slopes, LC50 and LC90
values, as well as 95% confidencelimits were tabulated. Baseline datawere then used to choose diagnosticdoses. Four doses were chosen

288
Whiteflies and Whitefly-borne Viruses in the Tropics
empirically so as to obtain mortalityranging from 5% to 95%. After severaltests, a single dose for each insectspecies and insecticide was chosen asthe diagnostic dose (as defined byHalliday and Burnham, 1990) to beused in extensive monitoring ofresistance under field conditions.
Field assessment of resistance
Resistance was assessed in the fieldusing vials coated in the laboratorywith diagnostic dosages of methomyl,methamidophos and cypermethrin, andtransported to the field in ice-chestsmaintained at about 20 °C. Theseinsecticides are representative of theconventional insecticides most widelyused in the study area. At each site,adult whiteflies were collected with amouth aspirator from the foliage ofinfested plants and transferred to thevials. Twenty adults per vial, replicatedfive times, were used for each test.Acetone-coated vials were used ascontrols. Infested vials were kept in the
ice-chests for 6 hours before mortalitywas recorded. Any test in whichmortality in control vials was higherthan 10% was discarded. Percentagemortality was corrected using theAbbott formula (Busvine, 1971) andtransformed to arcsine square root ofproportion. Data were then analysed bya 1-way analysis of variance (ANOVA)(SAS Institute, 1988). When the F testwas significant, comparison of meanswas by the least significance difference(LSD). Untransformed means arepresented.
Results and Discussion
Baseline data and diagnosticdoses
Table 1 presents baseline toxicologydata for reference strains ofT. vaporariorum and Table 2, the Abiotype of B. tabaci. The LC50 and LC90
values reflect toxicities of the test
Table 1. Toxicological responses of laboratory strains of Trialeurodes vaporariorum to fiveinsecticides.a
Insecticide n LC50
(95% FL)b LC90
(95% FL)b Slope ± SEM X2
Adults
Methomyl 457 0.25 (0.15-2.6) 0.95 (0.76-14.7) 2.19 ± 0.55 4.05
Methamidophos 600 5.30 (2.5-7.6) 22.50 (15.6-48.7) 2.05 ± 0.49 0.08
Monocrotophos 710 9.70 (6.7-13.4) 175.40 (115.5-299.8) 1.00 ± 0.08 4.80
Cypermethrin 480 37.00 (22.0-55.7) 400.00 (232.7-953.5) 1.24 ± 0.18 2.95
Lambda-cyhalothrin 605 9.40 (6.1-13.5) 264.90 (170.9-455.3) 0.90 ± 0.06 0.10
Bifenthrin 380 2.40 (1.6-3.1) 6.70 (5.2-9.8) 2.90 ± 0.49 0.90
Carbofuran 504 1.97 (1.5-2.5) 6.80 (5.4-9.7) 2.37 ± 0.31 4.31
Carbosulfan 712 1.80 (1.5-2.1) 19.90 (16.3-24.9) 1.20 ± 0.05 3.30
Imidacloprid 921 5.70 (4.6-6.9) 28.40 (20.9-44.9) 1.90 ± 0.22 7.80
Thiamethoxam 670 8.60 (6.0-11.7) 101.00 (68.8-170.4) 1.20 ± 0.12 5.90
First instar nymphs
Buprofezin 907 0.80 (0.6-1.1) 9.20 (6.9-13.0) 1.20 ± 0.09 7.30
Diafenthiuron 904 3.20 (2.3-4.4) 60.10 (41.8-92.8) 1.00 ± 0.06 10.50
Imidacloprid 483 16.50 (10.7-23.4) 171.50 (115.6-290.9) 1.21 ± 0.10 3.60
a. Conventional insecticides were tested using insecticide-coated glass vials. Imidacloprid tests wereconducted using the technique developed by Cahill et al. (1996a). Nymphs were tested using amodification of the methodology described by Prabhaker et al. (1985).
b. Imidacloprid, thiamethoxam, buprofezin and diafenthiuron concentrations in ppm. All others inµg/vial. LC, lethal concentration; FL, fiducial limits.

289
Insecticide Resistance in Colombia and Ecuador
insecticides to susceptible strains ofwhiteflies that have not been exposed toinsecticides for at least 10 years. Asdiscussed by ffrench-Constant andRoush (1990), establishing baselinedata for different insecticides is afundamental step in resistance studiesbecause these data will serve as a basisfor future comparisons, which will allowresearchers to detect any changes ininsecticide resistance levels. In addition,as explained by Sanderson and Roush(1992) and Denholm et al. (1996),calculation of baseline data permits theselection of diagnostic doses, which canbe used more conveniently in extensiveefforts to detect and monitor resistancesuch as the one reported here that wascarried out over numerous sites in theAndean zone of Ecuador and Colombia.Carbofuran was included becausegranular formulations of this insecticideare still effective againstT. vaporariorum in Colombia (Cardona,1995). Imidacloprid is a highly efficientinsecticide that is becoming popular inthe region. Careful monitoring of theefficiency of this product will be neededfor future resistance management andIPM work to be undertaken by thesecond phase of the TWF-IPM Project. Itis interesting to note that the LC50 valuefor imidacloprid for our reference strain
of T. vaporariorum (2.4 ppm) wassimilar to that obtained by Cahill et al.(1996a) using susceptible strains ofB. tabaci from Sudan and Pakistan.
The main purpose of thepreliminary tests was to selectdiagnostic doses for methomyl,methamidophos and cypermethrin(Table 3) that were then used tocompare the insecticide resistance oftest samples of whiteflies fromColombia and Ecuador with that of thefully susceptible laboratory strains. Wedid not calculate diagnostic doses bystatistical means. Instead, diagnosticdoses were chosen empirically toproduce >95% mortality.
Field monitoring of resistancein biotype B of B. tabaci
Responses to insecticides in ourstudies were arbitrarily classified asfollows on the basis of meanpercentage mortality:
· 0%-50% mortality: high resistance;· 50%-80% mortality: intermediate
resistance; and· 80% mortality: low resistance.
According to this scale, the Bbiotype of B. tabaci recently introduced
Table 2. Toxicological responses of laboratory strains of Bemisia tabaci biotype A to three insecticides,using insecticide-coated glass vials.
Insecticide n LC50
(95% FL)a LC90
(95% FL)a Slope ± SEM X2
Methomyl 500 1.7 (1.1-2.3) 9.1 (6.7-13.7) 1.76 ± 0.21 1.73
Methamidophos 517 1.4 (0.9-1.6) 6.6 (5.3-14.7) 1.86 ± 0.47 14.89
Cypermethrin 502 14.4 (5.8-27.2) 202.9 (122.5-352.4) 1.12 ± 0.14 3.27
a. Insecticide concentration: µg/vial. LC, lethal concentration; FL, fiducial limits.
Table 3. Response (corrected percentage mortality) of Trialeurodes vaporariorum and Bemisia tabaciadults to three insecticides.a
Whitefly species Methomyl Methamidophos Cypermethrin(2.5 µg/vial) (32 µg/vial) (500 µg/vial)
T. vaporariorum 97 99 88
B. tabaci biotype A 99 100 98
a. Diagnostic dosages were tested using insecticide-coated glass vials.

290
Whiteflies and Whitefly-borne Viruses in the Tropics
into Colombia showed high levels ofresistance to methomyl andmethamidophos and, in some places,moderate levels of resistance tocypermethrin (Table 4). Considerableinter-population variation was detected.Variability in the response of the Bbiotype to insecticides has beendetected also in Arizona(Sivasupramaniam et al., 1997), inHawaii (Omer et al., 1993) and inCalifornia (Prabhaker et al., 1996). Ingeneral, insecticide resistance levelsreflected insecticide use patterns. Forexample, the very high levels ofresistance to acetylcholine esteraseinhibitors (exemplified here bymethomyl and methamidophos) indifferent sites in the departments ofAtlántico, Córdoba and Sucre mayreflect the extensive use oforganophosphates on cotton, tomatoand vegetables in the region. Lowerlevels of resistance to the testpyrethroid (cypermethrin) may be due tothe less frequent use of this type ofinsecticide for whitefly control (Table 4in Chapter 4.2, this volume). However,further interpretation of the data ishampered by the lack of knowledge of
exactly where the B biotype whiteflythat invaded Colombia originated and ofthe pattern of insecticide use in thearea of origin. It is likely that thefrequency of insecticide resistance wasalready high in the pest population atthe time it was introduced.
Field monitoring of resistancein T. vaporariorum
The responses of T. vaporariorum toinsecticides were arbitrarily classifiedon the same basis as those of B. tabaci(0%-50% mortality, high resistance;50%-80% mortality, intermediateresistance; and > 80% mortality, lowresistance). In the northern Ecuador-southern Colombia region (Table 5),T. vaporariorum populations weresusceptible to methomyl in seven out ofeight sites tested. Generalizedresistance to methamidophos (the mostwidely used insecticide) was found.Extreme cases were those in northernEcuador (Turquisal, Ibarra) and insouthern Colombia (El Tambo,Gualmatán, Funes). Intermediate to lowresistance to cypermethrin was detectedin west-central Colombia (Table 6).
Table 4. Response (corrected percentage mortality) of Bemisia tabaci biotype B adults to threeinsecticides at sites on tropical lowland areas of the northern coast of Colombia.a
Site (department)b Methomyl Methamidophos Cypermethrin(2.5 µg/vial) (32 µg/vial) (500 µg/vial)
Repelón (Atlántico) 42.8 c 2.0 d 92.0 ab
Cereté 1 (Córdoba) 54.0 b 1.5 d 87.8 abc
Cereté 2 (Córdoba) 12.4 de 0.8 d 68.8 c
Ciénaga de Oro (Córdoba) 41.8 bcd 3.0 d 94.6 a
Cotorra (Córdoba) 6.8 e 4.2 d 78.2 bc
Montería (Córdoba) 5.4 e 2.4 d 87.2 abc
Manaure (Guajira) 100.0 a 25.4 b 91.5 ab
Corozal 1 (Sucre) 9.8 e 14.0 bc 85.4 abc
Corozal 2 (Sucre) 30.0 bcde 4.8 cd 65.2 c
Sampués (Sucre) 15.4 cde 16.2 bc 88.8 abc
“CIAT” (reference strain) 100.0 a 100.0 a 100.0 a
a. Diagnostic dosages were tested under field conditions using insecticide-coated glass vials. Twentyadults per vial replicated five times were used for each test. Means within a column followed by thesame letter are not significantly different at the 5% level (LSD test).
b. “CIAT” indicates a laboratory population of B. tabaci biotype A maintained for at least 10 yearswithout exposure to insecticides.

291
Insecticide Resistance in Colombia and Ecuador
Table 5. Response (corrected percentage mortality) of Trialeurodes vaporariorum adults to threeinsecticides in the southern Colombia-northern Ecuador tropical highland region.a
Site (country)b Methomyl Methamidophos Cypermethrin(2.5 µg/vial) (32 µg/vial) (500 µg/vial)
El Tambo (Colombia) 100.0 a 1.8 d 95.2 a
Gualmatán (Colombia) 100.0 a 1.8 d 35.2 c
Funes (Colombia) 84.2 a 9.0 d 78.0 b
Ibarra (Ecuador) 100.0 a 4.0 d 100.0 a
Pimampiro (Ecuador) 87.4 a 46.4 bc 100.0 a
San Vicente ( Ecuador) 55.6 b 59.0 b 100.0 a
Turquisal (Ecuador) 100.0 a 14.0 cd 100.0 a
“CIAT” (reference strain) 96.6 a 99.0 a 87.7 b
a. Diagnostic dosages were tested under field conditions using insecticide-coated glass vials. Twentyadults per vial replicated five times were used for each test. Means within a column followed by thesame letter are not significantly different at the 5% level (LSD test).
b. “CIAT” indicates a laboratory population of whiteflies of the same species as the test populationsmaintained for at least 10 years without exposure to insecticides and used here as a susceptiblecheck for toxicological studies.
Table 6. Response (corrected percentage mortality) of Trialeurodes vaporariorum adults to threeinsecticides at sites in the Antioquia-Cauca-Valle tropical highland region of west-centralColombia.a
Site (department)b Methomyl Methamidophos Cypermethrin(2.5 µg/vial) (32 µg/vial) (500 µg/vial)
Carmen de Viboral (Antioquia) 96.0 a 55.0 c 69.8 d
El Peñol (Antioquia) 90.0 ab 76.4 b 80.2 cd
Rionegro (Antioquia) 100.0 a 43.4 c 84.2 bcd
La Unión (Cauca) 100.0 a 99.0 a 98.2 a
Pescador (Cauca) 62.0 c 2.6 e 67.0 d
Cerrito (Valle) 100.0 a 8.2 e 89.0 abc
La Cumbre (Valle) 100.0 a 100.0 a 98.0 a
Pradera (Valle) 79.4 bc 2.8 e 64.8 d
Tenerife (Valle) 99.6 a 3.0 e 72.4 cd
“CIAT” (reference strain) 96.6 a 99.0 a 87.8 bcd
a. Diagnostic dosages were tested under field conditions using insecticide-coated glass vials. Twentyadults per vial replicated five times were used for each test. Means within a column followed by thesame letter are not significantly different at the 5% level (LSD test).
b. “CIAT” indicates a laboratory population of whiteflies of the same species as the test populationsmaintained for at least 10 years without exposure to insecticides and used here as a susceptiblecheck for toxicological studies.
Whiteflies were susceptible tomethomyl in all but two of the sitesstudied. Extreme resistance tomethamidophos occurred in the CaucaValley (Cerrito, Pradera, Tenerife) andin Cauca (Pescador). These are placeswhere insecticide use on common beanand tomato is heaviest. Again, low tointermediate resistance to
cypermethrin was detected. In theeastern mountain ranges of Colombia(Table 7), T. vaporariorum wassusceptible to methomyl and lessresistant to methamidophos than inother parts of the country, possiblybecause of lower insecticide use in thatarea.

292
Whiteflies and Whitefly-borne Viruses in the Tropics
Conclusions
In summary, the B biotype of B. tabaci,recently introduced into Colombia,showed high levels of resistance tomethomyl and methamidophos and, insome places, moderate levels ofresistance to cypermethrin. In highlandand mid-altitude areas of Colombiaand Ecuador, T. vaporariorum showedhigh resistance to methamidophos,intermediate resistance tocypermethrin and low resistance tomethomyl. In general, levels ofinsecticide resistance in whitefliesseem to be related to the intensity ofuse of the particular kind ofinsecticide. Baseline responses ofT. vaporariorum to carbofuran andimidacloprid are presented.
These results provide a basis forfuture monitoring of resistance and forthe development of insecticideresistance management strategies.Novel insecticides such asdiafenthiuron, buprofezin, pyriproxyfenand imidacloprid are still effective forwhitefly control in Colombia and
Table 7. Response (corrected percentage mortality) of Trialeurodes vaporariorum adults to threeinsecticides at sites in the central-eastern tropical highland region of Colombia.a
Site (department)b Methomyl Methamidophosc Cypermethrin(2.5 µg/vial) (32 µg/vial) (500 µg/vial)
Boyacá (Boyacá) 100.0 a 95.3 a 89.0 a
Tinjacá (Boyacá) 100.0 a 85.6 ab 75.2 abc
Bojacá (Cundinamarca) 100.0 a 40.0 c 57.9 cd
Pasca (Cundinamarca) 100.0 a - 81.0 ab
Fosca (Cundinamarca) 100.0 a 71.6 bc 41.4 d
Garzón (Huila) 100.0 a 52.2 bc 88.8 a
Rivera (Huila) 100.0 a 40.0 c 88.7 a
Abrego (Norte de Santander) 98.4 a 60.3 bc 71.3 abc
Lebrija (Santander) 91.5 b 3.0 d 64.4 bc
“CIAT” (reference strain) 96.6 a 98.8 a 87.8 a
a. Diagnostic dosages were tested under field conditions using insecticide-coated glass vials. Twentyadults per vial replicated five times were used for each test. Means within a column followed by thesame letter are not significantly different at the 5% level (LSD test).
b. “CIAT” indicates a laboratory population of whiteflies of the same species as the test populationsmaintained for at least 10 years without exposure to insecticides and used here as a susceptiblecheck for toxicological studies.
c. Test was not conducted at Pasca location.
Ecuador (Rodríguez et al., 2003).Because whiteflies can also developresistance to novel insecticides(Denholm et al., 1996; Horowitz andIshaaya, 1996; Elbert and Nauen,2000; Palumbo et al., 2001) monitoringof resistance and development ofbaseline data should continue in orderto facilitate management of whiteflypopulations. The results are furtherdiscussed and proposals for furtheraction based on these results arepresented in Chapter 4.4 (this volume).
References
Buitrago, N. A.; Cardona, C.; Acosta, A.1994. Niveles de resistencia ainsecticidas en Trialeurodesvaporariorum (Westwood)(Homoptera: Aleyrodidae), plaga delfrijol común. Rev. Colomb. Entomol.20(2):109-114.
Busvine, J. R. 1971. A critical review ofthe techniques for testinginsecticides. The CommonwealthInstitute of Entomology,Commonwealth AgriculturalBureaux, London, GB. 345 p.

293
Insecticide Resistance in Colombia and Ecuador
Cahill, M.; Hackett, B. 1992. Insecticidalactivity and expression of pyrethroidresistance in adult Bemisia tabaciusing a glass vial bioassay. In:Procs. 1992 Brighton Crop Prot.Conf. Pests and Diseases.p. 251-256.
Cahill, M.; Gorman, K.; Day, S.; DenholmI. 1996a. Baseline determinationand detection of resistance toimidacloprid in Bemisia tabaci(Homoptera: Aleyrodidae). Bull.Entomol. Res. 86:343-349.
Cahill, M.; Denholm, I.; Byrne, F. J.;Devonshire, A. L. 1996b. Insecticideresistance in Bemisia tabaci: Currentstatus and implications formanagement. In: Procs. 1996Brighton Crop Prot. Conf. p. 75-80.
Cardona, C. 1995. Manejo de Trialeurodesvaporariorum (Westwood) en frijol enla zona Andina: Aspectos técnicos,actitudes del agricultor ytransferencia de tecnología. MemoriaIV Taller Latinoamericano sobremoscas blancas y geminivirus,16-18 octubre 1995, Zamorano, HN.CEIBA 36(1):53-64.
Cardona, C.; Rendón, F.; Rodríguez, I.1998. Chemical control andinsecticide resistance of whiteflies inthe Andean zone: A progress report.In: Procs. International Workshop onBemisia and geminivirus, 7-12 June1998, San Juan, PR. p. 1-70.
Cardona, C.; Rendón, F.; García, J.;López-Avila, A.; Bueno, J.; Ramírez,J. 2001. Resistencia a insecticidasen Bemisia tabaci y Trialeurodesvaporariorum (Homoptera:Aleyrodidae) en Colombia yEcuador. Rev. Colomb. Entomol.27(1-2):33-38.
Denholm, I.; Cahill, M.; Byrne, F. J.;Devonshire, A. L. 1996. Progresswith documenting and combatinginsecticide resistance in Bemisia. In:Gerling, D.; Mayer, R. T. (eds.).Bemisia: 1995. Taxonomy, biology,damage, control and management.Intercept Ltd. Andover, Hants, GB.p. 577-603.
Dittrich, V.; Uk, S.; Ernst, H. 1990.Chemical control and insecticideresistance of whiteflies. In: Gerling,D. (ed.). Whiteflies: Their bionomics,pest status and management.Intercept Ltd., Andover, Hants, GB.p. 263-285.
Elbert, A.; Nauen, R. 2000. Resistance ofBemisia tabaci (Homoptera:Aleyrodidae) to insecticides insouthern Spain with specialreference to neonicotinoids. PestManag. Sci. 56(1):60-64.
ffrench-Constant, R. H.; Roush, R.T .1990. Resistance detection anddocumentation: The relative roles ofpesticidal and biochemical assays.In: Roush, R. T.; Tabashnik, B. E.(eds.). Pesticide resistance inarthropods. Chapman and Hall, NY,USA. p. 4-38.
Halliday, W.; Burnham, K. P. 1990.Choosing the optimal diagnosticdose for monitoring insectresistance. J. Econ. Entomol.83(4):1151-1159.
Horowitz, A. R.; Ishaaya, I. 1996.Chemical control of Bemisia–Management and application. In:Gerling, D.; Mayer, R. T. (eds.).Bemisia: 1995. Taxonomy, biology,damage, control and management.Intercept Ltd. Andover, Hants, GB.p. 537-556.
Omer, A. D.; Johnson, M. W.; Tabashnik,B. E.; Costa, H. S.; Ullman, D. E.1993. Sweetpotato whiteflyresistance to insecticides in Hawaii:Intra-island variation is related toinsecticide use. Entomol. Exp. Appl.67:173-182.
Palumbo, J.; Horowitz, A.; Prabhaker, N.2001. Insecticidal control andresistance management for Bemisiatabaci. Crop Prot. 20(9):739-765.

294
Whiteflies and Whitefly-borne Viruses in the Tropics
Plapp, F. W.; Jackman, J. A.;Campanhola, C.; Frisbie, R. E.;Graves, J. B.; Lutrell, R. G.; Kitten,W. F.; Wall, M. 1990. Monitoringand management of pyrethroidresistance in the tobacco budworm(Lepidoptera: Noctuidae) in Texas,Mississippi, Louisiana, Arkansasand Oklahoma. J. Econ. Entomol.83:335-341.
Prabhaker, N.; Coudriet, D.; Meyerdirk,D. 1985. Insecticide resistance inthe sweetpotato whitefly Bemisiatabaci (Homoptera: Aleyrodidae). J.Econ. Entomol. 78:748-752.
Prabhaker, N.; Toscano, N. C.;Henneberry, T. J.; Castle, S. J.;Weddle, D. 1996. Assessment of twobioassay techniques for monitoringof silverleaf whitefly (Homoptera:Aleyrodidae) in California. J. Econ.Entomol. 89(4):805-815.
Rodríguez, I.; Cardona, C. 2001.Problemática de Trialeurodesvaporariorum y Bemisia tabaci(Homoptera: Aleyrodidae) comoplagas de cultivos semestrales en elValle del Cauca. Rev. Colomb.Entomol. 27(1-2):21-26.
Rodríguez, I.; Morales, H.; Cardona, C.2003. Líneas base, dosisdiagnóstico y medición periódica deresistencia a insecticidas enpoblaciones de adultos e inmadurosde Trialeurodes vaporariorum(Homoptera: Aleyrodidae) en el Valledel Cauca, Colombia. Rev. Colomb.Entomol. 29(1):21-27.
Sanderson, J. P.; Roush, R. T. 1992.Monitoring resistance ingreenhouse whitefly (Homoptera:Aleyrodidae) with yellow stickycards. J. Econ. Entomol. 85(3):634-641.
SAS Institute. 1988. 1998 SAS user’sguide. SAS Institute, Cary, NC,USA.
Sivasupramaniam, S.; Johnson, S.;Watson, T. F.; Osman, A. A.;Jassim, R. 1997. A glass-vialtechnique for monitoring toleranceof Bemisia argentifolii (Homoptera:Aleyrodidae) to selected insecticidesin Arizona. J. Econ. Entomol.90(1):66-74.

295
Conclusions and Recommendations
CHAPTER 4.4
Conclusions and RecommendationsCésar Cardona*, Aristóbulo López-Avila** and
Oswaldo Valarezo***
When the Systemwide Program onIntegrated Pest Management (SP-IPM)launched its project for “Sustainableintegrated management of whiteflies aspests and vectors of plant viruses inthe tropics” (The Tropical Whitefly IPMProject, TWF-IPM), the nature ofwhitefly problems in the high Andes ofColombia and Ecuador was poorlyunderstood. Anecdotal reports andfragmentary scientific studies, mostlypublished only in the “grey” literature,suggested that whitefly problems weresevere and insecticide misusewidespread in this socially andeconomically important horticulturalarea (Chapter 4.1, this volume). Theseriousness of the reported situationled partners in the project planningprocess to establish a sub-projectspecifically to look at “Whiteflies aspests of annual crops in the highlandsof Latin America”. However, at theoutset of the project, the species ofwhiteflies responsible for the problemswas uncertain and the role ofinsecticides and insecticide-resistancein aggravating whitefly-relatedproblems was unclear. The first phaseof this project provided, for the first
time, an opportunity to surveysystematically the main cropproduction areas of this ecologicallycomplex region and characterize theirwhitefly problems, both from abiological point of view and in terms offarmers’ perceptions of these problems.
Scientists and technicians of theparticipating national researchorganizations, the Instituto NacionalAutónomo de InvestigacionesAgropecuarias (INIAP) in Ecuador andthe Corporación Colombiana deInvestigación Agropecuaria(CORPOICA) in Colombia, conductedfield surveys and farmer interviewsbetween October 1997 and December1998, in close collaboration with theircounterparts from the project’scoordinating organization, the CentroInternacional de Agricultura Tropical(CIAT) (Chapter 4.2, this volume).Researchers at CIAT continued untilMay 1999 to complete parallel studiesof insecticide resistance in whitefliesfrom selected sites within the studyarea (Chapter 4.3, this volume).
The results of these studies(Chapters 4.2 and 4.3, this volume)provide a unique description of whiteflyproblems in Andean agriculture, whichis not only of inherent scientific valuebut also will now serve as a soundfoundation for action to address thevery considerable agriculturalproduction, human health and
* Centro Internacional de AgriculturaTropical (CIAT), Cali, Colombia.
** Corporación Colombiana de InvestigaciónAgropecuaria (CORPOICA), Bogotá,Colombia.
*** Instituto Nacional Autónomo deInvestigaciones Agropecuarias (INIAP),Manabí, Ecuador.

296
Whiteflies and Whitefly-borne Viruses in the Tropics
environmental problems that the studyrevealed. Integrated pest managementapproaches are likely to play a key rolein tackling these problems, both withinthe framework of future phases of thecurrent project and beyond. Keylessons learned in this study andproposals for further action arediscussed here and in the final chapteron Conclusions (this volume).
Conclusions
Whitefly identification andhost crop importance
The greenhouse whitefly Trialeurodesvaporariorum (Westwood) is the mostimportant whitefly species in highlandand mid-altitude areas of Colombia andEcuador. Although it affects numeroushorticultural crops and ornamentals,T. vaporariorum is most important as apest of common bean (Phaseolusvulgaris L.) and tomato (Lycopersiconesculentum Mill.). This confirmsprevious work (Cardona, 1995;Corredor et al., 1999) on the increasingimportance of T. vaporariorum as amajor pest in the highlands of theAndes. However, it is now recognizedthat whitefly problems in the area aremore widespread than initially thought.
Altitudinal distribution
The results of extensive surveysshowed that T. vaporariorum occurs inmid-altitude and highland areasranging from 700 to 2800 m above sealevel and that Bemisia tabaci(Gennadius), the tobacco whitefly,ranges from sea level to 900 m abovesea level. Thus, these two speciesoverlap in many of the mid-altitudevalleys. This is important becauseproper identification of whitefly speciesis a critical step in the formulation ofresearch strategies. If, for example,B. tabaci is erroneously assumed to be
the main pest species in mid-altitudevalleys or highland areas, then IPMprograms will be ill targeted and maynot be effective.
Farmer perceptions
Whiteflies, in this case T. vaporariorum,were considered to be the key pest incommon bean at 50% of the sitesvisited and the key pest in 45% of thetomato farms that were surveyed. Asignificant proportion of farmers thinkthat whiteflies can reduce yields by50% or more. This perception may wellbe justified. Previous studies (Cardonaet al., 1993; Cardona, 1995) and workconducted in the first phase of theTWF-IPM Project showed thatT. vaporariorum may cause lossesranging from 23% to 54% of thepotential yield in snap bean (alsoPhaseolus vulgaris L.). There is nodoubt that whiteflies have a highdamage potential and that farmers arecorrect in believing that whiteflyattacks have created numerousproblems and have become a majorconstraint to agricultural production inthe region.
Insecticide use
Whiteflies in Colombia and Ecuadorhave become the target of intensive, inmost cases excessive, use ofinsecticides. This in turn has resultedin the destruction of natural enemiesand the creation of new secondary pestproblems such as the leafminerLiriomyza huidobrensis (Blanchard) andpod borers such as Epinotia aporema(Walsingham) on common bean and theNeoleucinodes elegantalis (Gueneé) ontomato. As shown in the previouschapter, excessive insecticide use hasresulted also in the development ofhigh levels of resistance of whiteflies toconventional insecticides. Somefarmers are now using costly novelinsecticides that may become uselessin a few years because of insecticide

297
Conclusions and Recommendations
resistance. Knowledge on insecticideuse patterns and insecticide resistancelevels obtained in this first, diagnostic,phase of the TWF-IPM Project will beimportant in the formulation ofinsecticide resistance managementstrategies and in the development ofIPM options aimed at reducingpesticide use.
Insecticide resistance
In highland and mid-altitude areas ofColombia and Ecuador, T. vaporariorumshowed high resistance tomethamidophos (an organophosphate),intermediate resistance tocypermethrin (a pyrethroid) and lowresistance to methomyl (a carbamate),three test products chosen asexemplifying the major groups ofconventional insecticides currently inuse. In general, resistance levels seemto be related to intensity of insecticideuse. Responses of T. vaporariorum tocarbofuran and imidacloprid weredetermined, to provide a baseline forfuture monitoring of resistance againstthese products, both of which are inwidespread use. The B biotype ofB. tabaci, recently introduced intoColombia, showed high levels ofresistance to methomyl andmethamidophos and, in some places,moderate levels of resistance tocypermethrin.
Development of resistance torepresentative conventionalinsecticides was higher than thosereported in a previous study (Buitragoet al., 1994). The diagnostic surveyshowed that insecticide resistance iswidespread, affecting common beanand tomato growers alike. These areimportant findings that help explainwhy conventional insecticides are nolonger effective and why farmers arenow using excessive numbers ofapplications of conventional pesticidesand new generation pesticides that are
costly. In the long-term, the need touse more expensive insecticides maythreaten the economic viability ofcommon bean and tomato productionin mid-altitude and highland areaswhere small-scale, usually poor,farmers predominate.
Recommendations
There is an urgent need to develop andimplement IPM systems that will helpre-establish the ecological equilibrium.Experience in the Sumapaz region ofColombia has shown that this isfeasible: implementation of a relativelysimple IPM package based on culturalcontrol and sanitation practices, timelyapplication of effective insecticides andreliance on natural control in certainareas resulted in a 66% reduction ininsecticide use (Cardona, 1995).Although the system was economicallyand technically viable, adoption ratesvaried with geographical area withinthe region and were not as high asresearchers expected. A main obstacleencountered was the high level of riskaversion (as defined by Tisdell, 1986)among small-scale farmers. However,overall adoption rates of 30%-35%were regarded as high by socialscientists linked to the project (NohraRuiz de Londoño [CIAT Impact Unit],personal communication, 1996) andsuggest that careful planning, activeparticipation of farmers in decisionmaking, and participatory researchapproaches may help overcomebarriers to adoption.
Several promising IPM tactics havebeen identified (Cardona, 1995; CIAT,2000). These include the replacementof broad-spectrum insecticides withmore selective ones, and the timing ofapplications according to pre-established action thresholds. Inaddition, cultural control measuressuch as the incorporation of crop

298
Whiteflies and Whitefly-borne Viruses in the Tropics
residues, leaf roguing andmanipulation of planting dates may beeffective in some circumstances.Advances have been made also in theevaluation of the parasitoid Amitusfuscipennis MacGown and Nebeker(Manzano et al., 2000) and theentomopathogen Verticillium lecanii(Zimm.) (González and López-Avila,1997) but more work needs to be donein order to measure the effectivenessof these and other natural enemies.These preliminary efforts to developand test IPM strategies locally,coupled with findings from thediagnostic phase of the TWF-IPMProject, can be used as a basis fordeveloping more widely applicable IPMstrategies. The overall results of theseefforts should be the development ofsystems that will: (1) reduce levels ofpesticide used by common beanfarmers; (2) reduce health risks tofarmers and consumers; and(3) re-establish the ecologicalequilibrium in areas that have beendisturbed by excessive pesticide use.
The immediate objective of anyfuture IPM initiative should be toreduce pesticide use. Farmerscurrently depend so heavily onchemical pesticides, andenvironmental disruption is of suchmagnitude, that for the foreseeablefuture it is most unlikely that any IPMsystem could be effective without achemical component. In the longerterm, as more farmers adopt IPMapproaches and environmentalequilibrium is re-established, moreambitious, pesticide-free, strategiesmight become feasible; however, forthe moment at least, monitoring ofresistance should continue and soundstrategies to manage insecticideresistance will have to be developed.Testing of new chemicals that areeffective but less detrimental to theenvironment will also be important.
Based on the diagnostic phasesurvey work, IPM pilot studies shouldbe continued in the following hot spots:Sumapaz, Oriente Antioqueño,Tenerife, Pradera and Nariño inColombia, and in the Chota valley innorthern Ecuador. However, problemsassociated with T. vaporariorum extendfar beyond the Colombian andEcuadorian highlands and commonbean. This whitefly also has beenreported as a significant field pestaffecting melon (Cucumis melo L.) andtomato in Mexico (Flores et al., 1995),tomato in Costa Rica (Alpizar, 1993),tomato and common bean in theDominican Republic (FUNDESA, 1996),as well as tomato, common bean andsnap bean in Venezuela (Arnal et al.,1993), Ecuador (Mendoza, 1996),Colombia (Cardona, 1995), Bolivia(CIAT, unpublished records), Brazil(Gerk et al., 1995), Peru (Núñez 1995)and Argentina (L’Argentier et al., 1996;Giganti et al., 1997). T. vaporariorum isalso a major pest in greenhouses inColombia (Corredor et al., 1999) andArgentina (Viscarret and Botto, 1996;Salas et al., 1999). The benefit ofdeveloping effective IPM approaches forT. vaporariorum is therefore likely to befelt beyond the highlands of Colombiaand Ecuador.
In order to overcome existingbarriers to adoption (Cardona, 1995),IPM components should be tested withlocal communities using participatoryresearch methods (Ashby, 1990). Apossible approach that already hasshown promise in Latin America(Brown and Hocdé, 2000) is to involveComités de Investigación Agrícola Local(CIALs) in the testing (and, in somecases, validation) of individual IPMcomponents and of promising IPMstrategies. If major changes in existingpractices are found to be necessarysuch as the general adoption ofparticular cropping dates or crop

299
Conclusions and Recommendations
rotations, then the implementation ofIPM will have to be discussed with localcommunities and government. Thesuccess of IPM approaches will dependultimately on establishing consensusamong IPM stakeholders. Givenfarmers’ attitudes to chemicalpesticides and their current reliance onthese products as the sole method ofaddressing whitefly problems, farmereducation (Whitaker, 1993) will bemost important; however, if a majorshift towards IPM approaches is to beachieved, advocacy efforts directed atconsumers and the variousstakeholders involved in agriculturalproduction and processing is likely tobe necessary as well.
Whitefly problems in the Andeanhighlands are evidently serious,extensive and deep-rooted, soimmediate success cannot beanticipated. However, a phasedapproach, as conceived within theframework of the TWF-IPM Project,promises tremendous benefits toproducers, consumers and theenvironment. The necessaryinvestment in research, education andpolicy change should therefore beundertaken without delay.
References
Alpizar, M. D. 1993. Aspectos básicossobre las moscas blancas conénfasis en Bemisia tabaci yTrialeurodes vaporariorum. Bol.Divulgativo no. 112, Ministerio deAgricultura y Ganadería, San José,CR. 22 p.
Arnal, E.; Russell, L. M.; Debrot, E.;Ramos, F.; Cermeli, M.; Marcano,R.; Montagne A. 1993. Lista demoscas blancas (Homoptera:Aleyrodidae) y sus plantashospederas en Venezuela. Fla.Entomol. 76(2):365-381.
Ashby, J. A. 1990. Evaluating technologywith farmers. A handbook. CentroInternacional de AgriculturaTropical (CIAT), Cali, CO. 95 p.
Brown, A. R.; Hocdé, H. 2000. Farmerparticipatory research in LatinAmerica: Four cases. In: Workingwith farmers: The key to adoption offorage technologies. Procs.International Workshop,12-15 October 1999, Cagayan deOro City, Mindanao, PH. AustralianCentre for International AgriculturalResearch (ACIAR), Canberra, AU.p. 32-53.
Buitrago, N. A.; Cardona, C.; Acosta, A.1994. Niveles de resistencia ainsecticidas en Trialeurodesvaporariorum (Westwood)(Homoptera: Aleyrodidae), plaga delfríjol común. Rev. Colomb. Entomol.20(2):109-114.
Cardona, C. 1995. Manejo de Trialeurodesvaporariorum (Westwood) en frijol enla zona Andina: Aspectos técnicos,actitudes del agricultor ytransferencia de tecnología.Memoria IV Taller Latinoamericanosobre moscas blancas ygeminivirus, 16-18 octubre 1995,Zamorano, HN. CEIBA 36(1):53-64.
Cardona, C.; Rodríguez, A.; Prada, P. C.1993. Umbral de acción para elcontrol de la mosca blanca de losinvernaderos, Trialeurodesvaporariorum (Westwood)(Homoptera: Aleyrodidae), enhabichuela. Rev. Colomb. Entomol.19(1):27-33.
CIAT (Centro Internacional de AgriculturaTropical). 2000. Annual report,Project IP-1. Bean Improvement forSustainable Productivity, Input UseEfficiency, and Poverty Alleviation.Cali, CO. p. 143-147.
Corredor, D.; Fischer, G.; Angarita, A.1999. Integrated pest managementin cut flower crops grown in plastichouses at the Bogotá Plateau.Procs. Int. Symp. on cut flowers inthe tropics, 14-17 October 1997,Bogotá, CO. Acta Horticulturae482:241-246.

300
Whiteflies and Whitefly-borne Viruses in the Tropics
Flores, A.; González, M. C.; Alarcón, A.;Ferrera R. 1995. Bioproducción demelón en campo. Rev. ChapingoSerie Hortic. 1(4):83-88.
FUNDESA (Fundación para el DesarrolloAgropecuario, Santo Domingo).1996. Proyecto de evaluación devariedades e híbridos de tomate portolerancia a geminivirosistransmitida por moscas blancas.Boletín FDA no. 9:2, 8. SantoDomingo, DO.
Gerk, A. O.; Vilela, E. F.; Pires, C. S.;Eiras, A. E. 1995. Biometría e ciclode vida da mosca branca,Trialeurodes vaporariorum(Westwood) e aspectos daorientacao de seu parasitoideEncarsia formosa Gahan. An. Soc.Entomol. Bras. 24(1):167-171.
Giganti, H.; Dapoto, G.; González, J. R.;Junyent, R. 1997. Insectos y ácarosasociados a los principales cultivoshortícolas del Alto Valle del RíoNegro y Neuquén, Argentina. Hortic.Argentina 16:40-41.
González, J. G.; López-Avila, A. 1997.Evaluación de cepas nativas deVerticillium lecanii (Zimm.) en elcontrol de la mosca blanca de losinvernaderos Trialeurodesvaporariorum (Westwood). Rev.Colomb. Entomol. 23(1-2):25-30.
L’Argentier, S. M.; de Manero, E. A.;Liacono A. 1996. Especies deAleyrodidae (Homoptera) presentesen poroto (Phaseolus vulgaris L.) yen las malezas asociadas al cultivoen las provincias de Jujuy y Salta,República Argentina. Rev. Investig.Centro de Investigaciones para laRegulación de Poblaciones deOrganismos Vivos 10:1-4.
Manzano, M. R.; van Lenteren, J. C.;Cardona, C.; Drost, Y. C. 2000.Developmental time, sex ratio, andlongevity of Amitus fuscipennisMacGown and Nebeker(Hymenoptera: Platygasteridae) onthe greenhouse whitefly. Biol.Control 18(2):94-100.
Mendoza, J. 1996. Qué está pasando conla mosca blanca en el Ecuador?Rev. INIAP 8: 8-10.
Núñez, E. Y. 1995. Reporte de Perú.Memoria IV Taller Latinoamericanosobre moscas blancas ygeminivirus, 16-18 octubre 1995,Zamorano, HN. CEIBA 36(1):157-162.
Salas, H.; Fernández, M.; Willink, E.1999. La mosca blanca de losinvernáculos en cultivos hortícolasde Tucumán. Avance Agroind.(Argentina) 19:39-41.
Tisdell, C. 1986. Levels of pest controland uncertainty of benefits. Aust. J.Agric. Econ. 30:157-161.
Viscarret, M. M.; Botto, E. N. 1996.Descripción e identificación deTrialeurodes vaporariorum(Westwood) y Bemisia tabaci(Gennadius) (Hemiptera,Homoptera: Aleyrodidae). Rev. Chil.Ent. 23:51-58.
Whitaker, M .J. 1993. The challenge ofpesticide education and training fortropical smallholders. Int. J. PestManag. 39(2):117-125.

SECTION FIVE
Special Topics on Pest andDisease Management
SECTION FIVE
Special Topics on Pest andDisease Management

BLANCA 302

303
Sustainable IPM through Host Plant Resistance
CHAPTER 5.1
Sustainable Integrated Management ofWhiteflies through Host PlantResistance
Anthony Bellotti*, Joseph Tohme*,Michael Dunbier** and Gail Timmerman**
Introduction
As direct feeding pests and vectors ofplant viruses, whiteflies constitute amajor problem in the production ofcassava (Manihot esculenta Crantz) inAfrica, the neotropics and, to a lesserdegree, Asia. The largest complex in theneotropics includes Bemisia tabaci(Gennadius)1, B. tuberculata (Bondar)and Trialeurodes variabilis(Quaintance). B. tabaci has apantropical distribution, feeding oncassava throughout Africa, where ittransmits African Cassava MosaicDisease, and several countries in Asia,including India and Malaysia.
Whitefly feeding affects cassava inthree ways. Direct damage is caused byfeeding in the phloem of the leaves,inducing chlorosis and leaf fall, whichresults in considerable reduction inroot yield if prolonged feeding occurs.Yield losses of this type are common inthe neotropics. Whiteflies also producea honeydew, which provides a mediumfor sooty mould growth that can reduceyields. Most importantly, B. tabaci is amajor vector of cassava viruses.
Research on whitefly control inthe neotropics has emphasized hostplant resistant (HPR) and biologicalcontrol. The bulk of this report willcover HPR research on whiteflies(Aleurotrachelus socialis Bondar) as acomponent of the Cassava WhiteflyIntegrated Pest Management (IPM)Project.
In traditional productionsystems, resource-limited farmershave few options available forcontrolling pests. The cassavagermplasm bank at the CentroInternacional de Agricultura Tropical(CIAT) has nearly 6000 accessionsand locally selected cultivars (landraces) collected primarily in theneotropics. These traditional cultivarsrepresent centuries of cassavacultivation in diverse habitats, havingbeen selected by farmers over a longperiod in the presence of a highdiversity of herbivores. These landraces often possess traits that conferlow to moderate levels of resistance tomultiple pests. This germplasm bankis constantly being evaluated forresistance to several arthropod peststhat can cause yield losses incassava. Evaluations often are donein more than one ecosystem.
The general purpose of thisproject is “to reduce crop losses dueto whitefly feeding damage andwhitefly-transmitted viruses, and
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** Crop and Food Research, Levin, NewZealand.
1. Evidence suggests that Bemisia tabacirepresents a species complex with numerousbiotypes.

304
Whiteflies and Whitefly-borne Viruses in the Tropics
prevent further environmentaldegradation and food contaminationdue to excessive pesticide use, leadingto more productive and sustainableagricultural systems”.
The project has three majorobjectives, which are to:
(1) Identify and access exotic or novelgenes and gene combinations thatcan contribute to germplasmenhancement for whiteflyresistance in cassava;
(2) Study the genetics of resistanceand to map genes for whiteflyresistance in cassava and developmolecular markers for theirincorporation into improvedAfrican, Latin American and Asiangermplasm; and
(3) Develop crop management optionsfor reducing whitefly populationsand the transmission of whitefly-transmitted viruses.
The research presented in thischapter is being funded by the Ministryof Foreign Affairs and Trade (MFAT)Host Plant Resistance (HPR) Projectand consists of four major areas ofactivity:
(1) Cassava germplasm evaluation toidentify sources of whiteflyresistance in land race varieties inthe CIAT cassava germplasmcollection;
(2) Identification of genomic regionsresponsible for the determinationof whitefly resistance in cassava;
(3) Identification of resistancemechanisms in cassava; and
(4) Tritrophic relationships todetermine the effect of HPR onwhitefly parasitism.
Cassava GermplasmEvaluation
Field screening of cassava germplasmcan be done at several sites inColombia. Ideally, field populations ofwhiteflies should be high and damagelevels significant so as to distinguishsusceptible cultivars. Field evaluationsof cassava germplasm use a population(nymphs) scale combined with a leafdamage scale (Table 1). Evaluations aremade periodically throughout thegrowing cycle; four to five evaluationsare done, 1.5 to 2.0 months apart.During the 1998 growing cycle, cassavaclones were evaluated at CIAT in theCauca Valley and Nataima in theTolima valley of Colombia.
From 1994 through to 1996,whitefly (A. socialis) populations atNataima, Tolima, were lower thannormal. Low whitefly populationsprovide inadequate selection pressureto ensure reliable resistanceevaluation. All evaluations done atNataima are with natural fieldpopulations of whiteflies.
During 1997-98, whiteflypopulations increased significantly,allowing for more accurate germplasmevaluation. During 1999, some1651 cassava clones were planted inobservational fields, and 1418 wereevaluated. These clones were the F1
progeny from crosses of cassava clonespreviously selected for their resistanceto whiteflies, with clones with desirableagronomic characteristics. Thesecrosses resulted in 17 families(Table 1). Four field evaluations weredone over a period of several months,using a whitefly damage andpopulations scale, as previouslydescribed.
Results indicate that whiteflypopulations were high and selection

305
Sustainable IPM through Host Plant Resistance
pressure was good. Of the 1418clones evaluated, 938 (66%) haddamage ratings above 3.6 and wereeliminated as susceptible (Figure 1);308 (22%) clones had an intermediatedamage evaluation (2.6 to 3.5). Theremaining 172 (12%) clones haddamage ratings below 2.6 and areconsidered promising for resistance;56 (3.9%) clones had damage ratingsof 1 to 1.5, indicating possible highlevels of resistance. However, onecycle (year) of field evaluation is notadequate to identify resistanceconfidently and these clones willcontinue to be evaluated for at least3 cycles (years). A 1.0 to 1.5 ratingsignifies no damage symptoms andlow whitefly population levels. Thefamilies that most frequently occurredin this group were CM 8984, withnine selections, and CM 8893, withsix. In both these families, theresistant parent was CG 489-34,indicating that this clone may containgood heritable resistant traits.
Table 1. Families of cassava clones formed from crosses of whitefly (Aleurotrachelus socialis)-resistantand -susceptible cultivars.
Family Crossesa Observation
Female Male
CM 3317 MBra 12 (T) MCol 1468 (S) MCol 1468 = CMC-40CM 5438 MBra 12 (T) MCol 1505 (S)CM 7559 MNGua 2 MBra 12 (T)CM 8884 CG 489-4 (R) MCol 1468 (S) CG 489-4 = MEcu 72 X MBra 12CM 8885 CG 489-4 (R) MCol 1505 (S) CG 489-4 = MEcu 72 X MBra 12CM 8887 CG 489-4 (R) MCol 2256 CG 489-4 = MEcu 72 X MBra 12CM 8889 CG 489-23 (R) MCol 1468 (S) CG 489-23 = MEcu 72 X MBra 12CM 8891 MCol 1468 (S) CG 489-34 (R) CG 489-34 = MEcu 72 X MBra 12CM 8892 MCol 2246 CG 489-34 (R) CG 489-34 = MEcu 72 X MBra 12CM 8893 MCol 2256 CG 489-34 (R) CG 489-34 = MEcu 72 X MBra 12CM 8960 MCol 2246 MBra 12 (T)CM 8961 MCol 2256 MBra 12 (T)CM 8984 MCol 1505 (S) CG 489-34 (R) CG 489-34 = MEcu 72 X MBra 12CM 8990 MCol 2026 CG 489-34 (R) CG 489-34 = MEcu 72 X MBra 12CM 8991 MCol 2026 MBra 12 (T)CM 8995 MEcu 72 (R) MCol 1468 (S)CM 8996 MEcu 72 (R) MCol 2246
a. R, whitefly resistant cultivar; T, whitefly tolerant cultivar; and S, whitefly susceptible cultivar.
Figure 1. Evaluations of F1 cassava clones
from crosses of whitefly(Aleurotrachelus socialis)-resistantand -susceptible cultivars at theCorporación Colombiana deInvestigación Agropecuaria(CORPOICA), Nataima, Tolima(1997-98).
800
700
600
500
400
300
200
100
0
No.
clo
nes
56
116
308
703
235
1.5 2.5 3.5 4.5 6
Resistant Intermediate Susceptible
Damage grade

306
Whiteflies and Whitefly-borne Viruses in the Tropics
Whitefly populations at CIAT weremoderate to high for the fourthconsecutive year. Evaluations at CIATduring 1998 concentrated on cultivarsthat had been selected as promising forresistance during previous years at theInstituto Colombiano Agropecuário(ICA)-Nataima.
Thirty-two clones sown in 50-plantplots were evaluated using a 1 to 6 (lowto severe damage or low to highwhitefly population level) scale.Whitefly population scales are based oncounts of nymphs, pupae and adults.The 32 clones evaluated consisted ofland race varieties, hybrids andbackcrosses, and previously had showngood levels of resistance in screeningtrials in Tolima. Five regional or farmervarieties from the Tolima area also wereincluded.
Results showed that nine cultivars,or 26.5%, presented very low damagelevels (1.0 to 1.5) and three cultivarshad damage levels between 1.6 and2.5. The remaining 20 cultivars haddamage levels between 2.6 and 5. Theregional cultivars from Tolima,Azucena, Ceiba Blanca, Almidona,Llanera Precoz and Cuero de Marranohad damage and population ratingsbetween 4 and 4.5, indicating thatfarmer varieties in the regions aresusceptible to whiteflies and probablyexperiencing significant yield losses.
The nine best cultivars wereMEcu 64, MPer 335, MPer 415 andMEcu 72 (all land race varieties), andCM 8424-6, CM 8424-33, CM 8424-4,CG 489-34 and CG 489-4 (all hybrids).All had damage ratings of 1.0, exceptMPer 415 and CG 489-34 with ratingsof 1.5. MEcu 72 and CG 489-34 hadmaintained low damage ratingsconsistently, over several years. Thevarieties MEcu 64 and MPer 335 hadexcellent growth habits as well as lowdamage ratings and low whitefly
populations. These two varieties willcontinue to be evaluated at CIAT andTolima and should enter into breedingschemes for improved whitefly-resistant clones.
Identification of GenomicRegions Responsible for
the Determination ofWhitefly Resistance in
Cassava
Previous research at CIAT determinedthe existence of different sources ofresistance to whitefly. The mostimportant genotypes are MBra 12 andMEcu 72, which were used asparentals in the generation of newgenotypes. CG489-34 has shown thebest resistance behaviour. The moresusceptible genotypes, MCol 2026 andMCol 1505, also were selected andevaluated.
The goal of this project is to studyand screen plants with resistance towhitefly in cassava, using moleculartechniques.
Different breeding populationswere obtained from the resistant andsusceptible genotypes as parentals:
CG 489-34 X MCol 2026 =131 individualsCG 489-34 X MCol 1505 =108 individualsMBra 12 X MCol 2026 =135 individuals
We selected the more contrastingindividuals (resistant and susceptible)in the field for each family. DNA wasextracted from the different individualsof each group and, with the parental,was mixed in a bulk. We usedmolecular screening (DNA restrictionfragment length polymorphisms[RFLPs] and random amplifiedpolymorphic DNA [RAPDs]) intended to

307
Sustainable IPM through Host Plant Resistance
find markers associated to resistance,and later it is intended to isolate theresponsible genes. The parental linesand the bulk will be screened withamplified fragment lengthpolymorphism (AFLP) markers.
Parentals from each family wereevaluated with RFLP markers from thecassava map (Fregene et al., 1997).Seventy-two polymorphic probes wereobtained. Sixty-two RAPD markers havebeen screened with the bulks of thesame families; the primer OP.P3 showeda clear polymorphism between thesusceptible and resistant group. Thismarker is being evaluated with thewhole population of each cross toconfirm its association with theresistance (Figure 2).
We intend to isolate and sequencethe polymorphic bands and generate asequence characterized amplified region(SCAR). This can be used to make adiagnosis of resistant materials anddetermine the most promisinggenotypes for breeding programs andfarmers.
With the sequence of the genes ofresistance, homologies will be
established with those reported onother crops, to understand expressionpatterns. The RFLP, RAPD,microsatellites and AFLP markers willbe used to generate a framework mapand for the quantitative trait locus(QTL) analysis.
Identification ofResistance Mechanisms
Whitefly feeding behaviour
Cassava genotypes with resistance towhiteflies have been identified at CIAT.Resistant clone MEcu 72 shows highmortality for both adult and immaturewhiteflies, which may suggest lessfeeding on this genotype under naturalconditions. It is necessary to identifywhitefly feeding behaviour onsusceptible genotypes and to makecomparisons with MEcu 72 in order tobetter understand the mechanisms ofresistance.
Electronic monitoring of insectfeeding (EMIF) is a technique thatpermits the identification andquantification of the feeding behaviours
Figure 2. Random amplified polymorphic DNA (RAPD) bulk analysis of whitefly (Aleurotrachelussocialis)-resistant and -susceptible cassava clones.
R1 and S1: CG 489-34 x MCol 2026 R2 and
S2: MBra 12 x MCol 2026R3 and S3: CG 489-34 x MCol 1505 R4:
Ecu 72; R5: Ecu 72 progeny

308
Whiteflies and Whitefly-borne Viruses in the Tropics
of hemipteran insects. By passing anelectrical signal to a test plant, andtethering an insect with a fine goldwire, modifications caused by styletmovements and feeding behaviours canbe observed as waveforms. The EMIFtechnique has been used extensivelyfor the study of mechanisms of plantresistance to insects. CIAT presentlyowns two AC-Electronic feedingmonitors, and has access to a DCversion of the system (electricalpenetration graph [EPG]). The ACsystems have been used withleafhoppers (Empoasca kraemeri Ross &Moore) on common bean (Phaseolusvulgaris L.), and the DC systems withcassava mealybugs (Phenacoccusmanihoti Matile-Ferrero). Preliminaryobservations with both systemssuggested that an easy protocol couldbe developed for monitoring thewhitefly. In addition, CIAT techniciansneeded to be trained in wiringtechniques, operation of the system,computer display and data acquisition.
Electronic monitoringmethodology
The methodology devised for electronicmonitoring of whitefly feedingbehaviour includes two major steps: awiring technique to attach the thin wireto the mesonotum of an adult whiteflyand the proper settings (groundvoltages, signal frequency) of theelectronic monitoring system.
The “normal” gold wire used withleafhoppers (12.7 µm) is too thick andstiff for the small whitefly adults. Athinner wire needed to be obtained.Since purchase of a thinner wire wasdifficult in the short period of timeallowed for this project, thinning of theexisting gold wire was necessary. To dothis, a 10- to 20-cm piece of the thickwire is placed for 45 min in a solutionof 3 mol nitric acid and 9 molhydrochloric acid. This is a potent
oxidizer that dissolves away part of thegold, leaving a thinner wire whileconserving its electricity conductingproperties. Pieces of this thin gold wireare attached with silver paint to acopper stub.
Female whiteflies from agreenhouse colony are placed by mouthaspirator on a vacuum stand and heldin place by a gentle vacuum. Severalindividuals can be placedsimultaneously. With the help of thecopper stub, the thin gold wire isplaced on the mesonotum of a female,and a small drop of electricallyconducting silver paint is used toattach the wire to the insect. Careneeds to be taken to not get silver painton the whitefly’s eyes or wings. Thisaffects their behaviour even more thanthe tethered condition. New insectsneed to be used should silver paint geton their wings or eyes.
Tethered whiteflies can beacclimated to the wire by placing themon a cassava leaf for 1 hour. Before theelectronic monitoring session begins,whiteflies are starved for 30 minutes.The copper stub holding the tetheredwhitefly is attached to the alligator clipon one of the input electrodes of theelectronic monitor.
A constant signal, 250 mV at250 Hz, was established as the bestsetting for A. socialis. This signal istransmitted via an electrode inserted inthe substrate of a potted plant (4-8weeks old). Detached leaves placed ona container filled with water and sealedwith the output electrode of theelectronic monitor also worked andshowed better conductivity than thepotted plants. However, the effects ofdetached leaves vs. whole plants onwhitefly feeding behaviour need to beevaluated further.

309
Sustainable IPM through Host Plant Resistance
Feeding behaviour
During probing, stylet movements andother feeding behaviours inducechanges to the constant signal. Thesemodifications, known as waveforms,are captured in a computer using ananalogue to digital converter board,displayed on the screen on a timescale, and stored for post-acquisitionmeasurement and analysis. Oncewaveforms have been correlated withspecific behaviours, quantification canbe made on their frequency andduration.
Waveforms produced by probingA. socialis on CMC 40 are shown inFigure 3. Because of time limitations,waveforms cannot be correlated withspecific behaviours for this whiteflyspecies. However, it has been reportedthat several species of whiteflies sharevery stereotypical styles of feeding and
that waveform patterns are very similaramong them. Since waveforms shownare similar in appearance to thosereported in the literature, we willdescribe them. All probes observedstarted with salivation waveformscorresponding to the intercellular paththat the stylets follow in their way tothe phloem. These waveforms areshown in Figures 3A and 3C as“intercellular stylet path”. They havebeen correlated also with thedeposition of a salivary sheath. In someinstances, the stylets come acrosstreachery elements in the xylem andthe whitefly ingests from them. Thisparticular waveform has not beencorrelated with completely, but there isevidence of ingestion. After reachingthe phloem, normally whiteflies go intocontinued ingestion, producing a flatwaveform whose beginning isillustrated toward the end of the trace(Figure 3).
Figure 3. Waveforms produced by probing Aleurotrachelus socialis on cassava cv. CMC 40.A and B: Single probe (stylet penetration). B and C: Single probe forms a second whitefly.Note that waveform identified as possible xylem ingestion and an unknown waveform thathas yet to be correlated with a feeding behaviour.
Intercellular Stylet Path (Salivation)
Intercellular Stylet Path (Salivation)
Intercellular Stylet Path (Salivation)
Phloem
Phloem
Possib. Xylem ingestion
Unknown
A
B
C
D

310
Whiteflies and Whitefly-borne Viruses in the Tropics
Figure 3D shows a new waveformthat previously has not been identifiedor associated with any behaviour. Itwould be expected that, with moreelectronic monitoring of whiteflyfeeding, more patterns would beidentified and more correlational workwould be needed. Now that amethodology exists for the study ofwhitefly feeding behaviour at CIAT,comparisons can be made amongresistant and susceptible genotypes.
Tritrophic Relationships:Determining Effect of
HPR on WhiteflyParasitism
It has been observed that farmers willuse pesticides to control highpopulations of cassava whiteflies.Pesticide applications will reduce theeffectiveness of biological control aswell as cause environmentalcontamination. Several years ofresearch at CIAT has identifiedcultivars with varying levels ofresistance to whiteflies, especially thespecies A. socialis.
Present research is investigatingthe compatibility between HPR andbiological control. The combination ofthese two methods could reduce pestpopulations below economic injurylevels or extend the usefulness of HPR.In addition, it is important to know theeffect that plant resistance, especiallyantibiosis, might have on parasitoidpopulations.
The compatibility of the parasitoidEncarsia hispida DeSantis wasevaluated on whitefly immaturesfeeding on whitefly resistancegenotypes of cassava. The resistantvarieties tested were MEcu 72, andCG 489-4; the susceptible check wasCMC 40. Parasite development time,fecundity and survival were measured.
Plants of the abovementionedvarieties at 1 month of age wereinfested with A. socialis eggs byexposing the leaf undersurface toovipositing adults for 36 hours. After15 days, when whitefly immatures werein the second instar, they were exposedto E. hispida parasites. E. hispida wasobtained by collecting cassava leaveswith parasitized whitefly pupae in thefield at CIAT, placing the leaves inblackened plastic traps and capturingemerging parasites. The results at thispoint are preliminary.
Results to date indicate that thefemale longevity of E. hispida wassimilar on varieties CMC 40 (17.8 days)and CG 489-34 in experiments 1 and2 (12.2 and 13.8 days, respectively)(Figure 4). E. hispida longevity onMEcu 72 was greatly reduced, only7.7 days, indicating that this varietyhas factors or characteristics thataffect the longevity of the parasite.These factors could be the number oftricomes or a chemical component thatis interfering with parasitoid feeding(Table 2).
20
15
10
5
0CMC40
Figure 4. Longevity of the parasitoid Encarsiahispida on the whiteflyAleurotrachelus socialis on threecassava varieties, CMC 40,CG 489-34 (experiments #1 and #2)and MEcu72.
Tim
e (d
ays
)
CG 489-34 #1
CG 489-34 #2
MEcu72
Variety
17.8
12.213.83
7.77
E. hispida emergence from A.socialis pupae was measured on thevarieties CG 489-34 and MEcu 72.Results show a low rate of emergenceof the parasitoid, 0.43 for CG 489-34and 0.86 for MEcu 72. These

311
Sustainable IPM through Host Plant Resistance
preliminary results indicate that hostplant resistance in cassava may affectthe development or emergence ofparasitoids. Experiments usingsusceptible varieties (CMC 40) stillneed to be completed. Since E. hispidais the most frequently collectedparasitoid in cassava fields, a muchhigher emergence is expected.
The survival of E. hispida whenassociated with A. socialis on thecultivar MEcu 72 decreased rapidlywhen compared to the other cultivars.One hundred percent mortalityoccurred within 19 days (Figure 5).Longest survival was on CMC 40, withindividuals living for 40 days. VarietyCG 489-34 was intermediate; in bothexperiments with this cultivar,
Table 2. Whitefly (Aleurotrachelus socialis) survival, development time and fecundity on fourcassava varieties.
Variety Percentage survival Development Number Tricomes/cm2
of immaturesa time/daysa eggs/female on second leaf
MBra 12 75.0 a 32.18 c 107 4.653
CMC 40 66.0 ab 32.07 c 64 189
CG 489-34 65.5 ab 33.13 b 46 15.290
MEcu 72 27.0 c 34.45 a 15 33.048
a. Means within a column followed by the same letter are not significantly different at the P = 0.05level, using LSD test.
individuals survived until 34 days.These data indicate that there may bevarietal influence associated withE. hispida survival. Since MEcu 72 andCG 489-34 are resistant varieties, thisresistance factor or mechanism may beaffecting parasitoid survival.
Host plant resistance andbiological control agents increasinglyare accepted as complementary pest-control tactics that can reduce the useof chemical pesticide, especially withdifficult-to-control pests such aswhiteflies. Cassava is most often grownby resource-limited, small-scalefarmers in the tropical and sub-tropicalregions of the world. HPR andbiological control offers a low-cost,practical, long-term solution for
100
80
60
40
20
01 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39
Time (days)
CG 489-34 #2MEcu 72 CMC 40 CG 489-34 #1
% s
urv
ival
Figure 5. Survival of Encarsia hispida, parasitoid of Aleurotrachelus socialis, on three varieties ofcassava, MEcu 72, CMC 40 and CG 489-34 (experiments #1 and #2).

312
Whiteflies and Whitefly-borne Viruses in the Tropics
maintaining lower whitefly populations,reducing crop losses, and limiting oreliminating the need for applications ofcostly insecticides. This is especiallyimportant in crops such as cassava,which has a long growing cycle (1 yearor more) that would require numerouspesticide applications to achieveadequate whitefly control. Research oncassava whitefly control at CIATinitially emphasized HPR. Morerecently, a concentrated effort is being
made to identify and use naturalenemies in an IPM context (seeChapter 5.2).
Reference
Fregene, M.; Angel, F.; Gómez, R.;Rodríguez, F.; Chavarriaga, P.;Roca, W.; Tohme, J.; Bonierbale, M.1997. A molecular genetic map ofcassava (Manihot esculenta Crantz).Theor. Appl. Gen. 95:431-441.

313
Biological Control of Whiteflies by Indigenous Natural Enemies
CHAPTER 5.2
Biological Control of Whiteflies byIndigenous Natural Enemies for MajorFood Crops in the Neotropics
Anthony Bellotti*, Jorge Peña**, Bernardo Arias*,José María Guerrero*, Harold Trujillo*,Claudia Holguín* and Adriana Ortega*
Introduction
Whiteflies as direct feeding pests andvirus vectors constitute a majorproblem in cassava (Manihot esculentaCrantz) and associated crops in Centraland South America and the Caribbeanregion. There is a large complex in theneotropics, where 11 species arereported on cassava alone (Bellotti etal., 1999). In the northern region ofSouth America (Colombia, Venezuela,and Ecuador) the major species isAleurotrachelus socialis (Bondar)(Castillo, 1996). Two additional species,of lesser importance, but frequentlyfound on cassava, are Bemisiatuberculata (Bondar) and Trialeurodesvariabilis (Quaintance). Highpopulations of A. socialis, frequentlyobserved in the region, can causeserious yield reductions in cassava.There is a correlation between durationof attack and yield loss: infestations of1 month resulted in a 5% yieldreduction, of 6 months in a 42%reduction, and of 11 months in a 79%reduction (Vargas and Bellotti, 1981).
Until recently, the Bemisia tabaci(Gennadius) biotypes found in theAmericas did not feed on cassava. Ithas been speculated that the absence
of African cassava mosaic disease(ACMD) in the Americas may be relatedto the inability of its vector, B. tabaci, tocolonize cassava (Costa and Russell,1975). Since the early 1990s, a newbiotype (B) of B. tabaci, considered bysome to be a separate species (Bemisiaargentifolii Bellows & Perring) (Perring etal., 1993) has been found feeding oncassava in the neotropics (França et al.,1996; Quintero et al., 1998). Morerecently, in greenhouse studies done atthe Centro Internacional de AgriculturaTropical (CIAT), it has been shown thatB. tabaci females will oviposit oncassava, and immatures will feed andpupate; however, few adults emerged(CIAT, 1999). It is considered thatACMD now poses a more serious threatto cassava production, because mosttraditional varieties in the neotropicsare highly susceptible to the disease. Inaddition, the B biotype of B. tabaci as avirus vector causes heavy crop losseson numerous other crops in theneotropics and these often are grown inassociation with cassava, or in thesame area. The possibility of virusdiseases moving between these crops,or the appearance of previouslyunrecorded viruses, has become apotential threat.
Host plant resistance and biologicalcontrol agents (e.g., parasitoids,predators and entomopathogens)increasingly are accepted as a means ofpest control that reduces environmental
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.
** University of Florida, USA.

314
Whiteflies and Whitefly-borne Viruses in the Tropics
contamination and other disadvantagesarising from excessive use of chemicalpesticides. Many natural enemies arefound associated with species ofwhiteflies on cassava in the neotropics,especially in Colombia and Venezuela(Gerling, 1986; Gold et al., 1989;Castillo, 1996; Evans and Castillo,1998). Although a large complex ofparasitoids have been collected fromcassava whiteflies, little is known aboutthe levels of parasitism they impose onthe whitefly population, either asindividual species or collectively.
For instance, during 1994 and1995, CIAT personnel carried outsurveys for cassava whitefly naturalenemies in three regions of Colombia:the Andes, the Atlantic Coast and theEastern Plains (Castillo, 1996). Onehundred sites were visited in20 departments (states). The threeregions differ in altitude, temperatureand precipitation, as well as in theircassava cultivation patterns. TheAndean region contains many smallplantations dispersed over a wide area;in the Atlantic Coast, cassavacultivation is generally in extensive anduniform areas; while in the EasternPlains, cassava is cultivated in small“patchy” areas near the foot of theeasternmost range of the Andes.
The whitefly species collected oncassava during these surveys wereA. socialis, B. tuberculata, T. variabilis,Tetraleurodes sp. and Aleuroglandulusmalangae Russell. The latter twospecies were collected for the first timein very low numbers in the AtlanticCoast and Andean region. A. socialisrepresented 64.5% of the adults and86% of the immature (eggs, nymphs,and pupae) collected across the threeregions. T. variabilis was representedby 27.5% of the adults and 11.0% ofthe immatures, while B. tuberculatarepresented 8.0% of the adults and3.0% of the immatures (Castillo, 1996).
A. socialis and B. tuberculata had awide distribution, both being collectedin almost all sites surveyed.T. variabilis was collected from theAtlantic Coast and the Andean region,but not from the Eastern Plains.A. socialis populations were highest inthe Eastern Plains and lowest in theAndean region. B. tuberculata numberswere highest on the Atlantic Coast andEastern Plains, and almost allT. variabilis were collected from theAndean region. The data from thisstudy showed that A. socialis andB. tuberculata populations decreasedwith increasing altitude, and were mostabundant below 900 m. T. variabiliswas most abundant at higherelevations, above 1400 m. A. socialispopulations were highest whentemperatures were above 23 °C, whileT. variabilis populations were higher atlower temperature (<19 °C).B. tuberculata were most abundant inthose areas with higher temperatures.Annual precipitation did not have asignificant effect on distribution of anyone of the three species.
A species richness of whiteflyparasitoids was collected during thesesurveys. Ten species were collectedfrom A. socialis, B. tuberculata andT. variabilis. Several of these wereunrecorded species (Castillo, 1996;Evans and Castillo, 1998). Theparasitoids collected were representedby three genera, Encarsia (fourspecies), Eretmocerus (four species)and Amitus (two species). Three of theEncarsia species were E. hispidaDeSantis, E. pergandiella Howard andE. bellotti. None of the Eretmocerusgenus was identified to species and oneAmitus was identified to species,A. macgowni Evans and Castillo.Eretmocerus sp. “b”, E. hispida,A. macgowni and E. bellotti werecollected parasitizing A. socialis.E. bellotti, E. hispida, E. pergandiellaand Eretmocerus sp. “a” were collected

315
Biological Control of Whiteflies by Indigenous Natural Enemies
from T. variabilis. Only Eretmocerus sp.“c” was collected parasitizingB. tuberculata (Castillo, 1996).
The highest level of parasitism wasaround 15% measured on A. socialis inthe Eastern Plains; almost 14% wasmeasured on B. tuberculata and 12%on T. variabilis, both in the Andeanregion. E. hispida represented thegreatest number of individualscollected, 1845 or about 64% of allparasitoids collected. Eretmocerus sp.“b” was the second most commonlycollected (485 individuals), followed byA. macgowni with 159 individuals.E. hispida and Eretmocerus sp. “b” werecollected in all three regions. Thegreatest complex of species was foundin the Andean region, whereas onlyE. hispida and Eretmocerus sp. “b” wereobserved in the Coastal and EasternPlains regions.
The results of this initial phase ofthe surveys indicated a rich complex ofparasitoids associated with cassavawhiteflies, many of which areunrecorded species that could proveuseful in biological control programs. Itwas further realized that an expandedsurvey would prove beneficial andincrease our knowledge of the naturalenemy complex associated withwhiteflies.
A second phase to these studieswas undertaken with the objectives to:
(1) Further define the complex ofnatural enemies, especiallyparasitoids, and quantify theirassociation with whitefly species;
(2) Expand these surveys to includeother crops in addition to cassava;
(3) Determine which natural enemiesregulate whitefly populations byevaluating parasitoid species undercontrolled conditions (laboratorystudies with E. hispida, a parasiteof A. socialis, were initiated); and
(4) Determine if the B. biotype ofB. tabaci is infesting cassava fields.
Methods
Survey methodology
Three geographic areas were selected forexploration for whitefly species and theirparasitoid natural enemies. These werethe Atlantic Coast (the Departments ofAtlántico, Córdoba, Bolívar andMagdalena), and the mid altitudeCentral Highlands (Departments ofCauca, Valle del Cauca, Caldas,Quindío and Risaralda) of Colombia.The surveyed Ecuadorian regions werethe coastal area and the highlands(Sierra).
The Colombian departmentssurveyed represented two distinctecological zones. The Atlantic Coast,which is hot and fairly dry, hastemperatures ranging from 27 to 36 °Cand an average relative humidity (RH)from 25% to 70%. The Andean region ismuch cooler, with temperatures rangingfrom 22 to 33 °C, and RH ranging from7% to 100%. The Atlantic Coast is alsocharacterized by a 4- to 6-month dryseason. Dry periods in the Andeanregion are usually 2 to 3 months.Altitude was 0 to 200 m for thecollection sites on the Atlantic Coast,and 25 to 1750 m in the Andean region.
Several crop hosts, includingcassava, common bean (Phaseolusvulgaris L.), tomato (Lycopersiconesculentum Mill.), eggplant (Solanummelongena L.), cotton (Gossypiumhirsutum L.), cucumber (Cucumis sativusL. var. sativus) and snap bean(P. vulgaris) were sampled. From eachcollection site, 100 leaves were collectedrandomly and 1 square inch leaf areawas examined to determine the whiteflyspecies present and their respectivedensities.

316
Whiteflies and Whitefly-borne Viruses in the Tropics
The rate of parasitism wasdetermined by collecting 40 leavesrandomly from the field and removing a1-inch square from each leaf sampled.Only one whitefly species was allowed toremain on each leaf square and theemergence of parasitoids is recorded foreach whitefly species. This methodologyallowed us to accurately determine theparasitoid species associated with eachwhitefly species. Identifications remainpending for some whitefly andparasitoid species.
Methodology for parasitismstudies with Encarsia hispida
Field surveys carried out over severalyears have shown that the parasitoidE. hispida is frequently foundparasitizing cassava whiteflies. A colonyof this species was established on A.socialis on cassava in the greenhouse,and activities were initiated to study E.hispida biology and behaviour.
To ensure successful rearing ofwhitefly parasitoids, a colony of thewhitefly A. socialis was maintained oncassava (var. CMC-40) in thegreenhouse. Potted 5-week old cassavaplants were infested weekly by exposingthem to A. socialis adults for ovipositionfor 48 hours. The parasitoid colony wasinitiated by collecting cassava leaveswith parasitized A. socialis pupae fromthe field. These were placed in a blackplastic box (emergence chambers) witha clear glass bottle attached to anopening in the lid where emergingparasites are drawn to the light andcollected. The parasitoids were removedfrom the bottle with an aspirator andidentified.
The desired parasitoid species(E. hispida) was collected in sufficientnumbers and released into nylon-meshcages (50 x 50 x 90 cm) in thegreenhouse (25 to 32 °C, 60%-80% RH).Each cage contained potted cassavaplants infested with second to third
instar A. socialis nymphs; 16 to 18 daysafter release, adult parasitoids began toemerge. Once emergence was detected,cassava leaves (now with parasitizedpupae) were collected and placed in the“emergence chamber” where parasitoidswere collected and species identificationconfirmed. Collected parasitoids wereused to maintain the parasite colony, orutilized in the various studies describedin this report.
E. hispida parasitism studies werecarried out in the greenhouse (25 to32 °C; 60%-80% RH), using A. socialisas host species. Three methodologieswere evaluated to determine parasitismefficiency:
(1) Four nylon-mesh cages (50 x 50 x90 cm) were used, containing twopotted cassava plants infested withwhitefly (A. socialis) nymphs. Sixtyparasitoids were released into eachcage with four repetitions for eachinstar.
(2) Four whitefly-infested cassavaplants were used; four infestedleaves were placed in a petri dish(100 x 15 mm), and 30 parasitoidswere released into each petri dishwith 16 repetitions for each instar.
(3) A muslin “bag” was placed over anentire cassava leaf and sealed atthe petiole. Each leaf wasinfested with 80 nymphs, and20 parasitoids were released intoeach bag with four repetitions perinstar.
Results
Surveys on the Colombian AtlanticCoast and the departments of Caucaand Valle del Cauca resulted in fourspecies of whiteflies being confirmed asfeeding on cassava: Aleurotrachelussocialis, Bemisia tuberculata,Trialeurodes sp. (probably T. variabilis)and Tetraleurodes sp. (Table 1).A. socialis was the predominant species

317
Biological Control of Whiteflies by Indigenous Natural Enemies
found in all three areas; however, itspopulation was considerably higher inValle del Cauca and lowest in Cauca(Figure 1). All four species werecollected from the Atlantic Coast,whereas Tetraleurodes sp. was not
Table 1. Whitefly species and their associated parasitoid complex collected on cassava from threegeographical regions of Colombia.
Region Whitefly species Parasitoid species
Atlantic Coast Aleurotrachelus socialis Encarsia sp.Eretmocerus sp.
Bemisia tuberculata Encarsia sp.Eretmocerus sp.Metaphycus sp.
Trialeurodes sp. Encarsia sp.Eretmocerus sp.
Tetraleurodes sp.
Valle del Cauca A. socialis Encarsia sp.Eretmocerus sp.
B. tuberculata
Cauca A. socialis Encarsia bellottiEretmocerus sp.Signiphora aleyrodis
B. tuberculata Encarsia pergandiellaEretmocerus sp.Euderomphale sp.Signiphora aleyrodis
Trialeurodes sp. Encarsia hispidaEncarsia pergandiellaEretmocerus sp.
6000
5000
4000
3000
2000
1000
0
No.
of pu
pae/
100 s
am
ple
s
AtlanticCoast
Cauca Valle delCauca
Aleurotrachelus socialis
Bemisia tuberculata
Trialeurodes sp.
Tetraleurodes sp.
Figure 1. Whitefly population densities oncassava in three geographic zones ofColombia.
collected in either Cauca or Valle delCauca.
Pupal populations of A. socialisranged from an average of about 55 persquare inch on leaf samples from Valledel Cauca to about 5 on the AtlanticCoast and 2 in Cauca (Figure 1). Thedata also showed that whenever thereare high whitefly densities orpopulations, A. socialis is thepredominant species regardless ofgeographic area. However, whendensities are low at sampling sites, otherspecies may predominate (Figure 2). Onthe Atlantic Coast, under low populationdensities, B. tuberculata and A. socialishad similar populations, whereas inValle del Cauca, B. tuberculatapredominated. In Cauca, Trialeurodeshad the highest population.
Numerous whitefly parasitoids werecollected from the three areas (Table 1).These parasitoids belong to the generaEncarsia, Eretmocerus, Metaphycus andEuderomphale, and indications are thatthe collection contains severalunrecorded species. The hyperparasitoid

318
Whiteflies and Whitefly-borne Viruses in the Tropics
Signiphora aleyrodis was collected onlyin Cauca. Encarsia was the genera mostfrequently collected in Cauca and Valledel Cauca (Figure 3), with the highestpercentage in the latter. On the AtlanticCoast, Eretmocerus was the mostfrequent genus. Metaphycus was onlycollected on the Atlantic Coast andEuderomphale only from Valle delCauca.
Levels of infestations were high forAleurodicus sp., A. socialis,B. tuberculata and Tetraleurodes sp. forthe coastal region of Ecuador.T. vaporariorum, which is foundinfesting common bean, was found forthe first time at high infestation levelsin cassava in the coastal and highlandsregions of Ecuador.
The association between whiteflyspecies and parasitoid species wasevaluated also. Results indicate asizable parasitoid species complexassociated with each whitefly speciesand that this can be influenced bygeographic area. For example, with
A. socialis, the predominant parasitoidgenus in the Atlantic Coast wasEretmocerus, while in Cauca and Valledel Cauca, Encarsia predominated(Figure 4). In Valle del Cauca, 99.6% ofthe parasitism of A. socialis was byEncarsia and 0.4% by Eretmocerus. Themost numerous complex of parasitoidswas found associated with B. tuberculata(Table 1). Eretmocerus and Metaphycuswere the predominant genera on theAtlantic Coast and Eretmocerus in Cauca.The hyperparasite S. aleyrodis wascollected from both A. socialis andB. tuberculata pupae.
Four species of parasitoids werecollected from Ecuador. The complex ofparasitoids collected from whiteflies incassava in Ecuador has been identifiedonly to the generic level (Greg Evans,Mike Rose, personal communication,1999). In the coastal region, A. socialiswas parasitized by Encarsia, followed byAmitus and Eretmocerus; Aleurodicus wasparasitized by Euderomphale sp.Tetraleurodes and Trialeurodes weremostly parasitized by Eretmocerus sp.
100
80
60
40
20
0
No.
of pu
pae/
100 in
sq.
AtlanticCoast
Cauca Valle delCauca
Aleurotrachelus socialis
Bemisia tuberculata
Trialeurodes sp.
Tetraleurodes sp.
Figure 2. The whitefly species complex atcollecting sites with low populationdensities in three geographic zonesof Colombia.
100
80
60
40
20
0
Par
asit
oids
AtlanticCoast
Cauca Valle delCauca
Encarsia sp. Euderomphale sp.
Eretmocerus sp. Signiphora aleurodis
Metaphycus sp. Not identified
Figure 3. Parasitoid species (%) collected fromwhiteflies on cassava in threegeographic zones of Colombia.
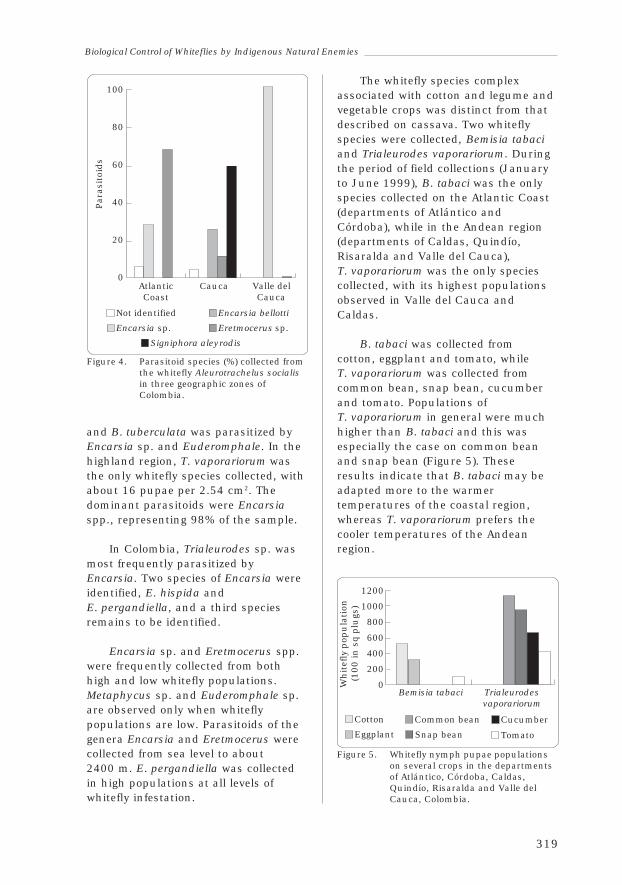
319
Biological Control of Whiteflies by Indigenous Natural Enemies
and B. tuberculata was parasitized byEncarsia sp. and Euderomphale. In thehighland region, T. vaporariorum wasthe only whitefly species collected, withabout 16 pupae per 2.54 cm2. Thedominant parasitoids were Encarsiaspp., representing 98% of the sample.
In Colombia, Trialeurodes sp. wasmost frequently parasitized byEncarsia. Two species of Encarsia wereidentified, E. hispida andE. pergandiella, and a third speciesremains to be identified.
Encarsia sp. and Eretmocerus spp.were frequently collected from bothhigh and low whitefly populations.Metaphycus sp. and Euderomphale sp.are observed only when whiteflypopulations are low. Parasitoids of thegenera Encarsia and Eretmocerus werecollected from sea level to about2400 m. E. pergandiella was collectedin high populations at all levels ofwhitefly infestation.
The whitefly species complexassociated with cotton and legume andvegetable crops was distinct from thatdescribed on cassava. Two whiteflyspecies were collected, Bemisia tabaciand Trialeurodes vaporariorum. Duringthe period of field collections (Januaryto June 1999), B. tabaci was the onlyspecies collected on the Atlantic Coast(departments of Atlántico andCórdoba), while in the Andean region(departments of Caldas, Quindío,Risaralda and Valle del Cauca),T. vaporariorum was the only speciescollected, with its highest populationsobserved in Valle del Cauca andCaldas.
B. tabaci was collected fromcotton, eggplant and tomato, whileT. vaporariorum was collected fromcommon bean, snap bean, cucumberand tomato. Populations ofT. vaporariorum in general were muchhigher than B. tabaci and this wasespecially the case on common beanand snap bean (Figure 5). Theseresults indicate that B. tabaci may beadapted more to the warmertemperatures of the coastal region,whereas T. vaporariorum prefers thecooler temperatures of the Andeanregion.
1200
1000
800
600
400
200
0Wh
itef
ly p
opu
lati
on(1
00 in
sq p
lugs
)
Bemisia tabaci Trialeurodesvaporariorum
Cotton
Eggplant
Common bean
Snap bean
Cucumber
Tomato
Figure 5. Whitefly nymph pupae populationson several crops in the departmentsof Atlántico, Córdoba, Caldas,Quindío, Risaralda and Valle delCauca, Colombia.
100
80
60
40
20
0
Par
asit
oids
AtlanticCoast
Cauca Valle delCauca
Not identified
Encarsia sp.
Encarsia bellotti
Eretmocerus sp.
Signiphora aleyrodis
Figure 4. Parasitoid species (%) collected fromthe whitefly Aleurotrachelus socialisin three geographic zones ofColombia.

320
Whiteflies and Whitefly-borne Viruses in the Tropics
Parasitoids from four genera,Encarsia and Eretmocerus(Aphelinidae), Amitus (Platygastridae)and Metaphycus (Encyrtidae) werecollected. These collections have beensent to taxonomists and speciesidentifications are pending. Therefore,in this report, they are referred to onlyby genera.
Encarsia was collected in all thedepartments except Risaralda (whereonly two sites were surveyed) butAmitus presented the highestpopulations, especially in Valle delCauca and Caldas (Figure 6). Thiscorresponded to the two departmentswith the highest whitefly populations.Eretmocerus was the predominantspecies in Córdoba. Parasitoidpopulations were highest in Valle delCauca and lowest in Risaralda andQuindío. Only one specimen ofMetaphycus was collected across thesites.
Encarsia parasites were collectedfrom all the plant hosts sampled andhad the overall highest densities amongall collected parasitoids (Figure 7).Amitus, although not collected from allhosts, also presented high populations,especially on common bean and snapbean. Eretmocerus was associatedalmost exclusively with cotton (only oneindividual was collected from tomato).
70
60
50
40
30
20
10
0
Para
sito
ids/
40 p
lugs
Atl
ánti
co
Amitus Encarsia Eretmocerus
Metaphycus Signiphora
Cal
das
Cór
dob
a
Qu
indío
Ris
aral
da
Val
le
Figure 6. Whitefly parasitoids collected fromseveral crops in the departments ofAtlántico, Córdoba, Caldas, Quindío,Risaralda and Valle del Cauca,Colombia.
70
60
50
40
30
20
10
0
Para
sito
ids/
40 p
lugs
Cot
ton
Eretmocerus
Metaphycus Signiphora
Egg
pla
nt
Com
mon
bea
n
Sn
apbea
n
Cucu
mbe
r
Tom
ato
Encarsia sp. Amitus
Figure 7. Whitefly parasitoid frequencycollected from several crops in thedepartments of Atlántico, Córdoba,Caldas, Quindío, Risaralda andValle del Cauca, Colombia.
Encarsia and Eretmocerus werecollected from both whitefly species(Figure 8), while Amitus was collectedonly on T. vaporariorum andMetaphycus and Signiphora werecollected only from B. tabaci.
During surveys, data on pesticideapplications were recorded duringfarmer interviews. In general,parasitoid populations were higher infields where pesticides were notapplied, regardless of the whiteflyspecies. Results show that populationsof Encarsia are less affected bypesticide applications than populations

321
Biological Control of Whiteflies by Indigenous Natural Enemies
of Amitus, indicating that Encarsia mayhave acquired a degree of resistance tosome of the pesticides applied.
Parasitism studies withE. hispida
Of the three methodologies evaluated,the highest rate of parasitism wasobtained using the third method.Parasitism levels using the first twomethods were low. The highest rate ofparasitism with method 1, the use ofnylon-mesh cages, only reached 9% inthe third instar. This improved with thesecond method (the use of leavesenclosed in petri dishes) to 30% in the2nd instar and to 20% in the 3rd.
The employment of muslin bags, asdescribed in the third methodology,gave the best results. In the first of twoexperiments using this methodology,parasitism rates reached about 75% inthe third instar and 16% in the1st instar, 45% in the 2nd instar and43% in the 4th instar (Figure 9). Resultsof a second experiment were similar,
with about 75% parasitism in the thirdinstar and 19% in the 1st instar, 61%in the 2nd instar and 25% in the4th instar (Figure 9). The averageparasitism rate for these twoexperiments were about 45%, whereasaverage parasitism rates were only 6%using methodology 1 and were 20%using methodology 2.
Figure 8. Relationship between parasitoidcomplex and whitefly speciescollected from several crops in thedepartments of Atlántico, Córdoba,Caldas, Quindío, Risaralda andValle del Cauca, Colombia.
Eretmocerus
Bemisiatabaci
Trialeurodesvaporariorum
Encarsia
Metaphycus Signiphora
Amitus
60
50
40
30
20
10
0
Para
sito
ids
(40 p
lugs
)
80706050403020100
% p
aras
itis
mI II III IV
Exp. 1 Exp. 2
Figure 9. Cassava whitefly (Aleurotrachelussocialis) instar preference by theparasitoid Encarsia hispida in twogreenhouse experiments.
These results also show that thethird whitefly instar is preferred forparasitism by E. hispida. An average ofall four experiments resulted in aparasitism rate of 21% for the1st instar, 35% for the 2nd instar, 46%for the 3rd instar and 22% for the4th instar. However, the average of thetwo experiments using methodology 3(muslin bags) is 17% for the 1st instar,53% for the 2nd, 75% for the 3rd and34% for the 4th instar. The highestparasitism rate is in the 3rd instar,followed by the 2nd, 4th and 1st instar.
The time period for optimalparasitoid activity was evaluated usingthe third methodology described.Percent parasitism evaluations weremade at 48, 72, 96 and 216 hours afterparasitoid release. Peak parasitismactivity occurred between 72 (35%parasitism) and 96 hours (33%parasitism) (Figure 10). However, even
Instars

322
Whiteflies and Whitefly-borne Viruses in the Tropics
at 216 hours, parasitism rates stillremained relatively high (nearly 29%)indicating a relatively lengthyparasitoid activity and the need to dofurther evaluations at time periodsbetween 96 and 216 hours.
parasitized. However, whiteflypopulations remain high and causeconsiderable plant damage and yieldloss, indicating that the actual activityof parasitoids is not sufficient toreduce whitefly populations beloweconomic injury levels that reducecassava yields.
Discussion
In future studies, additional parasitessuch as Eretmocerus and Amitus specieswill be evaluated in similar studies tothose with E. hispida. Continued surveyactivities to identify additional naturalenemies also are underway. During thisphase of exploration, more emphasiswill be given to regions or fields wherethere are low whitefly populations withthe hope of identifying key parasitoids,predators or entomopathogens that arepreventing eruptions of whiteflypopulations.
This is a collaborative projectbetween CIAT and the University ofFlorida, and is funded by the UnitedStates Agency for InternationalDevelopment (USAID). Thiscollaboration will provide training andimproved in-country capacity forresearch, production, delivery andmanagement of biological controlagents. The University of Florida willprovide expertise and input onparasitoid taxonomy, biology,behaviour, collecting, rearing,identification and data analysis. Since1998, University of Florida researchers(Dr. Jorge Peña and R. Nguyen) havevisited CIAT and collaborated in trainingCIAT personnel in some of the aboveareas. Whitefly parasitoids that arecollected from sampling are sent toUniversity of Florida taxonomists(G. Evans, M. Rose and A. Hammonds)for identification.
100
80
60
40
20
0
% p
aras
itis
m
48 72 96 216Time (hours)
Figure 10. Parasitoid (Encarsia hispida) activityin relation to time of exposure towhitefly host Aleurotrachelus socialison cassava in greenhouse studies.
The high rates of parasitism,especially using the muslin cagemethodology, indicate that E. hispidacould be an effective parasite in abiological control program forA. socialis. However, whiteflypopulations, especially of A. socialis,have been extremely high at CIAT forthe past 4 or 5 years. During thisperiod, E. hispida is the mostfrequently observed parasite. Thesepreviously described experimentswere carried out under controlledconditions where the parasitoid hadeasy access to the whitefly prey and theparasitoid probably is not adverselyinfluenced by environmental factors.Under natural field conditions,parasitoid activity may not be asefficient as indicated by thesegreenhouse studies.
Observations of pupal populationsin the field at CIAT and other selectedsites (i.e., Tolima) have shown that ahigh percentage of pupae are

323
Biological Control of Whiteflies by Indigenous Natural Enemies
References
Bellotti, A. C.; Smith, L.; Lapointe, S. L.1999. Recent advances in cassavapest management. Annu. Rev.Entomol. 44:343-70.
Castillo, J. A. 1996. Moscas blancas(Homoptera: Aleyrodidae) y susenemigos naturales sobre cultivosde yuca (Manihot esculenta Crantz)en Colombia. M.Sc. thesis,Universidad del Valle, Cali, CO.173 p.
CIAT (Centro Internacional de AgriculturaTropical). 1999. Annual Report,Project PE1. Cali, CO.
Costa, A. S.; Russell, L. M. 1975. Failureof Bemisia tabaci to breed oncassava plants in Brazil(Homoptera, Aleyrodidae). Cienc.Cult. 27:388-90.
Evans, G. A.; Castillo, J. A. 1998.Parasites of Aleurotrachelus socialis(Homoptera: Aleyrodidae) fromColombia including descriptions oftwo new species (Hymenoptera:Aphelinidae: Platygasteridae). Fla.Entomol. 81(2):171-178.
França, F. M.; Villas-Bôas, G. L.; CasteloBranco, M. 1996. Ocorrência deBemisia argentifolii Bellows andPerring (Homoptera: Aleyrodidae) noDistrito Federal. Anais Soc.Entomol. Bras. 25:369-372.
Gerling, D. 1986. Natural enemies ofBemisia tabaci, biologicalcharacteristics and potential asbiological control agents: A review.Agric. Ecosyst. Environ. 17:99-110.
Gold, C. S.; Altiere, M. A.; Bellotti, A. C.1989. The effects of intercroppingand mixed varieties on predatorsand parasitoids of cassavawhiteflies in Colombia: Anexamination of the “natural enemyhypothesis.” Bull. Entomol. Res.79:115-21.
Perring, T. M.; Cooper, A. D.; Rodriguez,R. J.; Farrar, C. A.; Bellows, T. S.1993. Identification of a whiteflyspecies by genomic and behaviouralstudies. Science 259:74-77.
Quintero, C.; Cardona, C.; Ramírez, D.;Jiménez, N. 1998. Primer registrodel biotipo B de Bemisia tabaci(Homoptera: Aleyrodidae) enColombia. Rev. Colomb. Entomol.24(1-2):23-28.
Vargas, O.; Bellotti, A. C. 1981. Pérdidasen rendimiento causadas pormoscas blancas en el cultivo de layuca. Rev. Colomb. Entomol.71(1-2):13-20.

324
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 5.3
Progress of Cassava Mosaic Disease inUgandan Cassava Varieties and inVarietal Mixtures
William Sserubombwe*, Michael Thresh**,James Legg*** and William Otim-Nape*
A feature of the production of cassava(Manihot esculenta Crantz) in Uganda,as in many other countries of sub-Saharan Africa, is that many differentvarieties are grown and it is usual tofind several varieties within individualplantings. Moreover, cassava is usuallygrown together with one or moreintercrops, including legumes andcereals. The great diversity in thevarieties grown and in the croppingsystems adopted can be expected toinfluence the incidence and severity ofcassava pests and diseases. However,this possibility has received onlylimited attention in relation to cassavamosaic disease (CMD), despiteindications that the current pandemicin Uganda and in adjacent parts ofnorth-west Tanzania and westernKenya is closely associated with thevarieties adopted and is least damagingin areas where many varieties aregrown.
The cassava component of theNatural Resources Institute (NRI)contribution to the Tropical Whitefly
Integrated Pest Management (TWF-IPM)Project has considered the role ofvarietal diversity in relation to CMDunder epidemic conditions in Uganda.It is proposed to consider furtheraspects of varietal diversity and theimplications of intercropping in anyadditional phase of the project.
Response of LocalVarieties Aladu and Bao
to Cassava Mosaic Disease
Ugandan farmers selected the varietiesAladu and Bao and originally grewthem mainly in Apac District, wherethey appeared to have at least somedegree of resistance to CMD. For thisreason, and in the absence of moreresistant varieties, they wereintroduced to Kumi, Soroti and otherdistricts of Uganda in the early 1990sin attempts to rehabilitate cassavaproduction following the devastatingepidemic of CMD.
This approach was partiallysuccessful and farmers have retainedboth Bao and Aladu in some areas,although they are less resistant thansome of the other improved varietiesnow available. Bao became heavilyinfected under the epidemic conditionsencountered originally. However, as theepidemic abated and the symptomsbecame less severe, some farmers haveattempted to return to Bao because of
* Namulonge Agricultural and AnimalProduction Research Institute (NAARI),Kampala, Uganda.
** Natural Resources Institute (NRI), GreenwichUniversity, Chatham Maritime, Kent, UK.
*** International Institute of TropicalAgriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda, and NRI, University of Greenwich,Chatham Maritime, Kent, UK.

325
Progress of Cassava Mosaic Disease in Single and Mixed Stands
its attractive eating qualities and otherdesirable attributes.
Replicated plots of Bao and Aladuwere established in May 1998 andrecorded monthly to assess theincidence and severity of CMD.Comparable plots of the highlyresistant SS4 also were established asa standard. All plants of Bao and Aladuwere harvested individually in June1999 and the data for individual plants(each of which had a full set of fourneighbours) were used for analysis.
There was rapid spread of CMD toBao and all plants were affected within6 months after planting (MAP)(Figure 1). CMD spread less rapidly inAladu and 15% of the plants wereunaffected at the final observation12 MAP. On average, symptomsdeveloped in Aladu 5.1 MAP comparedwith 3.7 MAP for Bao. Few plants ofSS4 were affected and the symptomswere less conspicuous than those ofBao and Aladu. Yields of tuberousroots were related to the stage ofgrowth when symptoms were firstexpressed and Bao was more severelyaffected than Aladu (Figure 2). Littleyield was obtained from plants of eithervariety that were grown from infectedcuttings. Plants infected by whiteflieswere affected less severely, especiallythose that developed symptoms at alate stage of growth.
The incidence and yield data wereused to calculate the losses caused byCMD in relation to fully healthy standsand assuming that all plants weregrown from healthy cuttings. Theoverall loss was 75% for Bao comparedwith 31% for Aladu. This confirms thevulnerability of Bao and the relativetolerance of Aladu, as farmers reported
earlier. Moreover, sufficientsymptomless plants of Aladu remainedto provide cuttings for a furtherplanting. This suggests that Aladucould be sustained even underepidemic conditions, provided thatfarmers select healthy cuttings at eachcycle of propagation. Additional studiesof this type are justified on thesustainability of a wider range of localvarieties and under less extremeconditions of inoculum pressure thanthose encountered at Namulongebetween 1998 and 1999.
120
100
80
60
40
20
0
600
500
400
300
200
100
0
Inci
den
ce o
f C
MD
(MIU
)In
ciden
ce o
f C
MD
(%
)
Months after planting
0 1 2 3 4 5 6 7 8 9 10 11 12
A
B
Figure 1. Mean monthly incidence of cassavamosaic disease (CMD) in localcassava varieties SS4, Aladu andBao at Namulonge, 1998-99experiment. Incidence data (A) aspercentages and (B) transformed tomultiple infection units (MIU).
Aladu SS4 Bao

326
Whiteflies and Whitefly-borne Viruses in the Tropics
Ability of UgandanVarieties to Withstand
Effects of Cassava MosaicDisease
Experience over the last decade hasshown that farmers in Uganda haveovercome the effects of the CMDepidemic by adopting resistant varietiesintroduced from Nigeria or thoseselected by the National AgriculturalResearch Organisation (NARO) and by
selecting from the local varieties alreadyavailable within the country. Muchinformation has been obtained on theperformance of the resistant materialthat has been released to farmersthrough official rehabilitation schemesbut little information is available on thefeatures of the local varieties that havebeen adopted widely following theepidemic. Accordingly, a study has beenmade of the 18 local varieties listed inTable 1. They were collected fromdifferent parts of Uganda and selectedfor their apparent ability to withstandthe most damaging effects of CMD.
Wherever possible, cuttings werecollected from symptomless plants ofeach variety selected for study and fromplants expressing mild and severesymptoms. The cuttings were planted atNamulonge in May 1998, together withthe varieties Ebwanateraka, Bao,Migyera and SS4, which were includedas standards. All plants were assessedmonthly to record the incidence andseverity of CMD. Vegetative growth andpopulations of the whitefly vectorBemisia tabaci (Gennadius) were alsoassessed and the plants were rated foroverall vigour 12 MAP.
Populations of adult whitefly wereunusually high throughout the period ofthe experiment and there was rapidspread of CMD both within the trial andin adjacent experiments. All the initiallyhealthy plants of the local varietiesbecame infected but there was relativelylittle spread to the resistant standards.There was wide variation in symptomseverity depending on the variety andthe initial health status of the plantingmaterial. Symptoms were generallyhighly conspicuous except in the localvarieties Buhimba, Njule and Tongoloand in the resistant standards SS4 andMigyera. Nevertheless, many of the localvarieties grew vigorously, especiallywhen grown from cuttings collected fromsymptomless or mildly affected plants.
5
4
3
2
1
0Tu
ber
ous
root
yie
ld p
er p
lan
t(k
g)A
100
90
80
70
60
50
40
30
20
10
0
Tu
ber
ous
root
yie
ld p
er p
lan
t(%
of
hea
lth
y)
B
Aladu Bao
Aladu Bao
Stage of virus infection
C E I L H C E I L H
Figure 2. Relationship between time of firstsymptom expression of cassavamosaic disease and tuberous rootyield per plant in local cassavavarieties Aladu (left) and Bao (right).Data presented as (A) mean yield perplant and (B) yields expressed aspercentage of healthy controls. Datafor 10 plants each having a full setof four immediate neighbours.C = infected as cutting, E = early,I = intermediate, L = late infectedand H = healthy (symptomless)plants. (The healthy Bao werenominal controls sampled in anadjacent plot in the absence ofuninfected plants.)
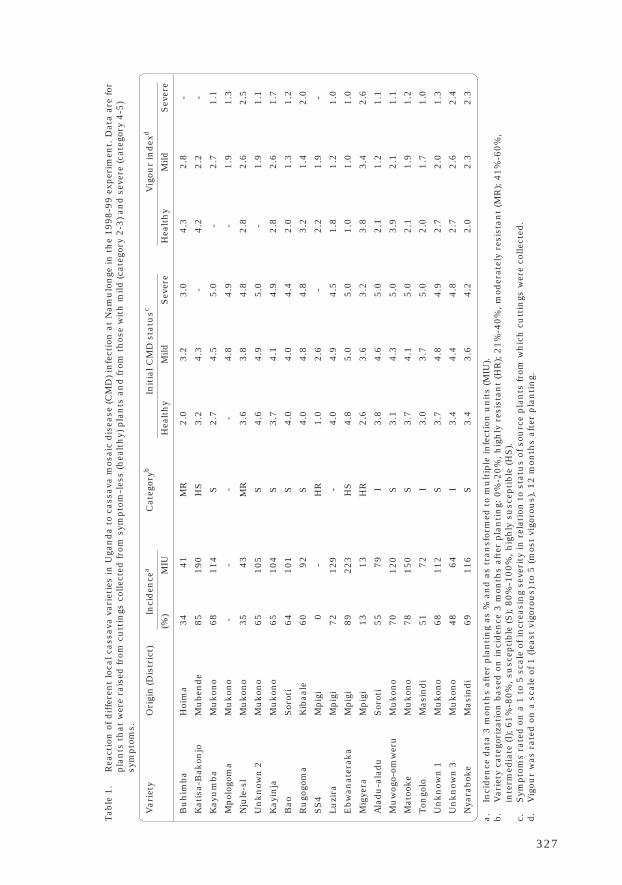
327
Progress of Cassava Mosaic Disease in Single and Mixed Stands
Table
1.
Rea
ctio
n o
f diffe
ren
t lo
cal ca
ssava
vari
etie
s in
Uga
nda t
o ca
ssava
mos
aic
dis
ease
(C
MD
) in
fect
ion
at
Nam
ulo
nge
in
th
e 1998-9
9 e
xper
imen
t. D
ata
are
for
pla
nts
th
at
wer
e ra
ised
fro
m c
utt
ings
col
lect
ed fro
m s
ympto
m-l
ess
(hea
lth
y) p
lan
ts a
nd fro
m t
hos
e w
ith
mild (ca
tego
ry 2
-3) an
d s
ever
e (c
ate
gory
4-5
)sy
mpto
ms.
Var
iety
Ori
gin
(Dis
tric
t)In
ciden
cea
Cat
egor
ybIn
itia
l C
MD
sta
tusc
Vig
our
index
d
(%)
MIU
Hea
lth
yM
ild
Sev
ere
Hea
lth
yM
ildS
ever
e
Bu
him
ba
Hoi
ma
34
41
MR
2.0
3.2
3.0
4.3
2.8
-
Kat
isa-
Bak
onjo
Mu
ben
de
85
190
HS
3.2
4.3
-4.2
2.2
-
Kay
um
ba
Mu
kon
o68
114
S2.7
4.5
5.0
-2.7
1.1
Mpol
ogom
aM
ukon
o-
--
-4.8
4.9
-1.9
1.3
Nju
le-s
lM
ukon
o35
43
MR
3.6
3.8
4.8
2.8
2.6
2.5
Un
kn
own
2M
ukon
o65
105
S4.6
4.9
5.0
-1.9
1.1
Kay
inja
Mu
kon
o65
104
S3.7
4.1
4.9
2.8
2.6
1.7
Bao
Sor
oti
64
101
S4.0
4.0
4.4
2.0
1.3
1.2
Ru
gogo
ma
Kib
aale
60
92
S4.0
4.8
4.8
3.2
1.4
2.0
SS
4M
pig
i
0-
HR
1.0
2.6
-2.2
1.9
-
Lu
zira
Mpig
i72
129
-4.0
4.9
4.5
1.8
1.2
1.0
Ebw
anat
erak
aM
pig
i89
223
HS
4.8
5.0
5.0
1.0
1.0
1.0
Mig
yera
Mpig
i13
13
HR
2.6
3.6
3.2
3.8
3.4
2.6
Ala
du
-ala
du
Sor
oti
55
79
I3.8
4.6
5.0
2.1
1.2
1.1
Mu
wog
o-om
wer
uM
ukon
o70
120
S3.1
4.3
5.0
3.9
2.1
1.1
Mat
ooke
Mu
kon
o78
150
S3.7
4.1
5.0
2.1
1.9
1.2
Ton
golo
Mas
indi
51
72
I3.0
3.7
5.0
2.0
1.7
1.0
Un
kn
own
1M
ukon
o68
112
S3.7
4.8
4.9
2.7
2.0
1.3
Un
kn
own
3M
ukon
o48
64
I3.4
4.4
4.8
2.7
2.6
2.4
Nya
rabok
eM
asin
di
69
116
S3.4
3.6
4.2
2.0
2.3
2.3
a.
Inci
den
ce d
ata
3 m
onth
s aft
er p
lan
tin
g as
% a
nd a
s tr
an
sfor
med
to
mu
ltip
le in
fect
ion
un
its
(MIU
).b.
Vari
ety
cate
gori
zati
on b
ase
d o
n in
ciden
ce 3
mon
ths
aft
er p
lan
tin
g: 0
%-2
0%
, h
igh
ly r
esis
tan
t (H
R); 2
1%
-40%
, m
oder
ate
ly r
esis
tan
t (M
R); 4
1%
-60%
,in
term
edia
te (I)
; 61%
-80%
, su
scep
tible
(S
); 8
0%
-100%
, h
igh
ly s
usc
epti
ble
(H
S).
c.S
ympto
ms
rate
d o
n a
1 t
o 5 s
cale
of in
crea
sin
g se
veri
ty in
rel
ati
on t
o st
atu
s of
sou
rce
pla
nts
fro
m w
hic
h c
utt
ings
wer
e co
llec
ted.
d.
Vig
our
was
rate
d o
n a
sca
le o
f 1 (le
ast
vig
orou
s) t
o 5 (m
ost
vigo
rou
s), 12 m
onth
s aft
er p
lan
tin
g.

328
Whiteflies and Whitefly-borne Viruses in the Tropics
Samples were collected fromrepresentative plants of all varieties forpolymerase chain reaction (PCR) andenzyme-linked immunosorbent assay(ELISA) analysis to determine thecassava mosaic virus(es) present andtheir concentration and associationwith the symptoms expressed.Preliminary results showed that theepidemic-associated virus (nowdesignated East African cassava mosaicvirus-Uganda [EACMV-Ug]) and Africancassava mosaic virus (ACMV) occurredalone or as a mixture. The fewsymptomless plants that reactedpositively contained mixed infectionsand so provided evidence of viruslatency.
These initial studies confirm theability of some local varieties topartially withstand infection. They alsoindicate the scope for further studieson the behaviour of local varieties andon their sustainability, especially underthe less extreme conditions ofinoculum pressure encountered inpost-epidemic situations.
Spread of Cassava MosaicDisease and Whitefly
Vector Populations onVarieties Grown Singly
and as a Mixture
Detailed analyses have been done offield data collected during an earlierM.Sc. project funded by the RockefellerFoundation. This project assessed thespread of CMD and whitefly vectorpopulations in four contrastingvarieties grown singly and as a mixtureunder epidemic conditions atNamulonge in 1995-96 and 1996-97.There was much spread of CMD to thesusceptible variety Ebwanateraka, littleto the resistant SS4 and intermediatelevels in TMS 30337 (Nase 2) and TMS30572 (Nase 3, also known as Migyera)(Figure 3). The amount of spread to the
resistant and partially resistant varietieswas similar in the sole plots and in themixture. In contrast, there was lessspread to the susceptible Ebwanaterakain the mixture than when it was grownalone. The effect was significant and islikely to have been even greater underlower levels of inoculum pressure thanthose experienced in the twoexperiments. This suggests thatresistant varieties may be used toprovide at least some degree ofprotection to susceptible ones. Indeed,the effect could be of considerablepractical importance because farmersmay be reluctant to discard susceptiblevarieties that are valued because ofquality or other favourable attributes.
A new varietal mixtures experimentwas planted at Namulonge in Septemberin 1998. There were four treatmentseach replicated four times in a completerandomized block design:
(1) Variety Bao (susceptible): alone.(2) Variety SS4 (resistant): alone.(3) Bao and SS4 planted alternately in
50:50 mixture.(4) SS4 (50%) grown as an outside
barrier around Bao (50%).
Monthly assessments were made ofadult whitefly populations, CMDincidence and CMD severity. Theassessment of whiteflies was done sothat any edge or barrier effects would bedetected.
The health status of the Baoplanting material was unsatisfactoryand there was substantial infectionarising from the cuttings used. Thisreflects the difficulty now beingexperienced in Uganda in obtaininghealthy material of susceptible varieties.Nevertheless, preliminary analyses showbig differences between SS4 and Bao inthe incidence and severity of CMD(Figure 4). There was also evidence thatthe incidence of CMD in Bao was less in

329
Progress of Cassava Mosaic Disease in Single and Mixed Stands
Figure 3. Mean monthly incidence of cassava mosaic disease (CMD, %) in successive months afterplanting in varieties Migyera, Nase 2 and Ebwanateraka when grown alone and as amixture at Namulonge in (left) the 1995-96 and (right) the 1996-97 experiments. Incidencedata for Ebwanateraka were transformed to multiple infection units (MIU) in the bottom twofigures.
100
80
60
40
20
0
100
80
60
40
20
0
100
80
60
40
20
0
800
600
400
200
0
8(J
a)
0(M
)
2(J
y)
1(J
u)
3(A
)
4(S
)
5(O
)
6(N
)
7(D
)
8(J
u)
0(O
)
2(D
)
1(N
)
3(J
a)
4(F
)
5(M
)
6(A
)
7(M
)
Inci
den
ce o
f C
MD
(%
)In
ciden
ce o
f C
MD
(MIU
)
Migyera Migyera
Nase 2 Nase 2
Ebwanateraka Ebwanateraka
Ebwanateraka Ebwanateraka
MAP MAP
Grown alone In varietal mixture
the mixture than in the pure stand.However, there was no evidence of abarrier or edge effect using SS4 and thecomparison may have been vitiated bythe extent of cutting infection with Bao.A further difficulty is that Bao is a tall
erect variety that outgrows SS4, which isa relatively low spreading type. Thissuggests that varieties of similar habitand conformation should be selected forany further evaluation of mixtures.
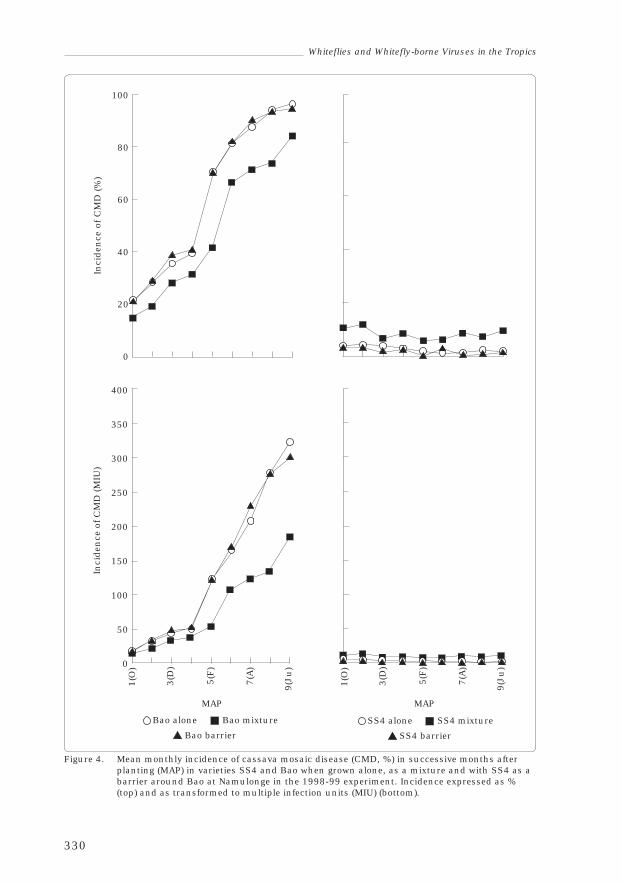
330
Whiteflies and Whitefly-borne Viruses in the Tropics
Figure 4. Mean monthly incidence of cassava mosaic disease (CMD, %) in successive months afterplanting (MAP) in varieties SS4 and Bao when grown alone, as a mixture and with SS4 as abarrier around Bao at Namulonge in the 1998-99 experiment. Incidence expressed as %(top) and as transformed to multiple infection units (MIU) (bottom).
100
80
60
40
20
0
Inci
den
ce o
f C
MD
(%
)
400
350
300
250
200
150
100
50
0
Inci
den
ce o
f C
MD
(MIU
)
9(J
u)
1(O
)
3(D
)
5(F
)
7(A
)
MAP
9(J
u)
1(O
)
3(D
)
5(F
)
7(A
)
MAP
Bao alone Bao mixture
Bao barrier
SS4 alone SS4 mixture
SS4 barrier

331
Progress of Cassava Mosaic Disease in Single and Mixed Stands
The experiment was harvested12 MAP to consider the yields ofhealthy and infected plants of eachvariety in relation to the type andhealthy status of their immediateneighbours. It is particularly importantto assess the ability of SS4 tocompensate for the impaired growth ofBao in the mixture of the two varieties.
Conclusions
The results obtained are consistentwith previous observations on thebehaviour of local varieties and on thescope for adopting somewhat tolerantvarieties and varietal mixtures as ameans of alleviating the effects of CMD(Sserubombwe et al., 2001). Furtherstudies on the behaviour andsustainability of local varieties,especially under the less extremeconditions of inoculum pressureencountered in post-epidemicsituations, need to be made.
Acknowledgments
This study was carried out incollaboration with the NARO CassavaProgramme, Uganda. The study willcontinue and results will be analysedand written up with short-term fundsprovided by the Crop ProtectionProgramme of the UK Department forInternational Development (DFID).
Reference
Sserubombwe, W. S.; Thresh, J. M.;Otim-Nape, G. W.; Osiru, D. O. S.2001. Progress of cassava mosaicvirus disease and whitefly vectorpopulations in single and mixedstands of four cassava varietiesgrown under epidemic conditions inUganda. Ann. Appl. Biol. 138:161-170.

332
Whiteflies and Whitefly-borne Viruses in the Tropics
CHAPTER 5.4
Management of the Cassava MosaicDisease Pandemic in East Africa
James Legg*, James Whyte**,Regina Kapinga*** and James Teriψ
Introduction
During the 1990s, a pandemic of anunusually severe form of cassavamosaic disease (CMD) expanded tocover a large part of East Africa,including virtually the whole of Ugandaand parts of western Kenya, southernSudan, north-western Tanzania andeastern Democratic Republic of Congo(DRC) (Otim-Nape et al., 1997;ASARECA, 1998; Legg et al., 1999b).This has been associated with theoccurrence of a novel and highlyvirulent cassava mosaic begomovirus(Deng et al., 1997; Harrison et al.,1997; Zhou et al., 1997). Surveys toassess the prevalence and severity ofCMD were conducted in Uganda, Kenyaand Tanzania as part of the diagnosticphase of the Tropical WhiteflyIntegrated Pest Management (TWF-IPM)Project (Chapters 1.6, 1.7 and 1.8, thisvolume). A principal outcome of thesesurveys was the identification ofregions that were either currentlyaffected or threatened by pandemic
expansion, based on epidemiologicaldata (Legg et al., 1999b).
In view of the acute effects of thesevere CMD associated with thepandemic, the co-ordination team ofthe TWF-IPM Project considered thatthere was an immediate need toidentify sources of funding to supportCMD control activities in recentlyaffected/threatened areas. Dialoguewas initiated with staff of the UnitedStates Agency for InternationalDevelopment (USAID) by the ProjectCo-ordinator and subsequentlyfollowed up in Uganda by theco-ordinator of the Africa-basedSub-Project 4. A concept note wassubmitted to the Office for ForeignDisaster Assistance (OFDA) of USAIDin April 1998 and funding for a 1-yearproject was approved the followingmonth. The project, entitled“Emergency programme to combat thecassava mosaic disease pandemic inEast Africa” began in October 1998 andthis first phase was completed inSeptember 1999.
Background to Control ofthe CMD Pandemic
After a decade of major losses tocassava production in Uganda due tothe CMD pandemic (Otim-Nape et al.,1997), substantial experience has beengained on control strategies. CMD-
* International Institute of TropicalAgriculture-Eastern and Southern AfricaRegional Center (IITA-ESARC), Kampala,Uganda, and Natural Resources Institute(NRI), University of Greenwich, ChathamMaritime, Kent, UK.
** IITA, Gabarone, Botswana.*** Centro Internacional de la Papa (CIP),
Kampala, Uganda.ψ Tanzania Coffee Research Institute, Moshi,
Tanzania.

333
Management of the Cassava Mosaic Disease Pandemic in E. Africa
resistant cassava germplasmintroduced from the InternationalInstitute of Tropical Agriculture (IITA)in the early 1980s was evaluated inmulti-locational and on-farm trialsduring the early 1990s and, in 1993,three cultivars (TMS 60142,TMS 30337 and TMS 30572) wereofficially released under the namesNase 1, Nase 2 and Nase 3 (or Migyera).In addition to being resistant toinfection, they were tolerant also of theeffects of cassava mosaicbegomoviruses (CMBs) once infected(Thresh et al., 1994). Multiplicationefforts were initiated during the mid-1990s (Otim-Nape et al., 1994; GCF,1997) and areas initially targetedincluded some of the first areas to beaffected by the pandemic in northernand eastern Uganda (Otim-Nape et al.,1994).
In more recent years, as thepandemic has expanded southwards,the emphasis of control efforts hasshifted to central and southerndistricts. At the end of 1997, a majorproject funded by the PL 480programme of USAID and entitled“Dissemination and utilization ofmosaic resistant cassava in Uganda”was initiated in the central-southerntarget districts of Masindi, Luwero,Mukono, Kamuli and Iganga. PrincipalCMD management themes within thisprogramme were monitoring anddiagnostics, plant health managementwithin the multiplication scheme andparticipatory evaluation of newCMD-resistant germplasm. The projecttargeted districts that had beenaffected by the CMD pandemic formore than 1 year and therefore alreadywere experiencing significant shortagesof planting material. This project didnot address districts affected morerecently, in 1996 and 1997, asidentified by the TWF-IPM Projectdiagnostic surveys.
Pandemic control in westernKenya and open quarantine
The expansion of the pandemic intowestern Kenya was first noted in 1995(Gibson, 1996; Legg et al., 1999b)although it was not until 2 years lateras the increasing seriousness of theproblem was recognized that initiativesbegan to control it. The key constraintto CMD management in western Kenyawas the virtual absence of any CMDresistant germplasm in the country. Inorder to address this problem,scientists of the East African RootCrops Research Network (EARRNET)worked with plant quarantine officialsof both Uganda and Kenya to developguidelines for the establishment of an“open quarantine” facility. This wassited at Busia, on the Kenyan side ofthe Kenya/Uganda border, and allowedthe introduction of stem cuttings of twoCMD resistant cultivars, SS4 andTMS 30572 (Nase 3), in addition to awide range of selected clones from theEARRNET germplasm developmentprogramme, based at Serere, Uganda.It was also at this time that thepandemic control programme inwestern Kenya received its first majoroffer of financial support, from theGatsby Charitable Foundation (GCF) inthe UK. EARRNET co-ordinated thedevelopment of a project, funded byGCF, which provided for theintroduction, evaluation andmultiplication of CMD-resistantcultivars in western Kenya and wasinitially targeted to run for 3 years.
Regional pandemic control andthe OFDA CMD Project
Whilst USAID and Gatsby-fundedinitiatives were providing substantialsupport for the multiplication anddissemination of CMD resistantcultivars in Uganda and Kenya, it wasclear that the increasing regionalizationof the CMD pandemic needed to beaddressed by taking a more pro-active

334
Whiteflies and Whitefly-borne Viruses in the Tropics
and co-ordinated approach. It wasenvisaged that such an approachwould involve establishing linksbetween control efforts in each of theEast African countries affected orabout to be affected by the pandemic,and targeting monitoring and controlinitiatives towards “threatened” zonesin addition to areas already affected.This was the rationale behind thedevelopment of the OFDA-fundedproject, which began in October 1998.
The purpose of the OFDA CMDProject was to “boost production ofcassava in Uganda, Kenya andTanzania and enhance both short andlonger term food security, through theimplementation of an emergencyprogramme to multiply anddisseminate mosaic resistant cassava”.Principal partners included IITA-Eastern and Southern Africa RegionalCenter (ESARC), the two regional rootcrops networks of EARRNET and theSouthern African Root Crops ResearchNetwork (SARRNET) and national rootcrops programmes in Kenya, Tanzaniaand Uganda. Principal project activitieswere organized under five themes:
(1) Monitoring and diagnostics:conduct focused surveys in south-western Uganda, western Kenyaand north-western Tanzania toprovide detailed distribution mapsof CMGs in the project target areasand baseline data for subsequentimpact analysis.
(2) Multiplication: multiply anddistribute in collaboration withproject partners, elite CMD-resistant materials.
(3) Germplasm diversification:increase the range of cassavamaterials available to farmers inareas targeted by the project,thereby reducing future risk ofproduction collapse.
(4) Stakeholder linkages: identify andstrengthen links between key
stakeholders with roles inenhancing cassava production intarget areas.
(5) Farmer training: develop producerskills in identification andmanagement of cassava pests anddiseases with special focus onCMD, in addition to basicproduction and multiplicationskills.
ImplementationHighlights—The OFDA
CMD Project
Significant progress was made in theattainment of project targets in each ofthe main themes and the project hasbeen successful in fostering a morecomprehensive regional approach totackling the CMD problem.
Monitoring and diagnostics
An important feature of the project hasbeen the use of regular monitoring anddiagnostic surveys to assess the statusof CMD in threatened zones anddevelop forecasts for the likely patternof development of the pandemic.Surveys focused on the Kagera region innorth-west Tanzania and Western andNyanza Provinces in western Kenya.During the first quarter of the project,the first report was made of severe CMDin Kagera and subsequent virusdiagnoses using specific polymerasechain reaction (PCR) primers confirmedthe association of severe CMD in thisregion with the presence in diseasedplants of mixed infections of Africancassava mosaic virus (ACMV) and theUganda variant of East African cassavamosaic virus (EACMV-Ug) (Legg andOkao-Okuja, 1999). The occurrence ofmixed ACMV/EACMV-Ug infections ischaracteristic of the “front” of the CMDpandemic (Harrison et al., 1997) andthe associated severe symptoms are themain reason for its acute impact oncassava cultivation in affected areas.

335
Management of the Cassava Mosaic Disease Pandemic in E. Africa
In western Kenya, the distributionof CMGs appears to be complex andACMV, EACMV and EACMV-Ug havebeen reported (Ogbe et al., 1996; Leggand Okao-Okuja, 1999). Monitoringsurveys in late 1998 demonstrated theapparent slowing of the spread of thepandemic. This was possibly as a resultof the presence of major naturalbarriers between north and southNyanza Province (Kisumu area), whichinclude the Winam Gulf of LakeVictoria and the Kano Plains, anexpanse of flood plain in which cassavacultivation is virtually absent (Legg etal., 1999a). Although high incidences ofsevere CMD have yet to be reportedfrom south Nyanza, the pandemic-associated EACMV-Ug has beendetected from Migori, near theTanzania border.
Data obtained from monitoring anddiagnostic surveys have been used todevelop regional CMD maps and theirpresentation has been an importanttool in raising awareness amongstagricultural workers based inthreatened zones. It is considered alsothat presentation of these graphics hasbeen a key component in persuadingboth national agricultural institutionsand donor agencies to take seriouslythe threat posed by the pandemic toregional cassava production and, withit, food security.
Multiplication
The multiplication and disseminationof CMD-resistant cultivars has beenthe main control technique used intackling the CMD pandemic in Ugandaand it has fulfilled the same role in theOFDA CMD Project in the wider region.Approaches to multiplication differed ineach of the three participatingcountries, however, since theavailability of planting material differedsubstantially in each. In Uganda,stems of cultivar SS4 were obtainedfrom the USAID-funded PL 480 Project
and used to plant 15 ha in each ofRakai and Masaka Districts.
In western Kenya, by contrast, lessthan 4 ha of CMD-resistant cultivarswere present at the outset of theproject, at the open quarantine site.The OFDA CMD Project joined GCF andthe Rockefeller Foundation insupporting the multiplication of thismaterial and by June 1999 the area ofCMD-resistant material had increasedto more than 35 ha.
In north-western Tanzania, the lastof the three countries to be affected bythe pandemic, only a small number ofplants of CMD-resistant varieties werepresent by the end of 1998 andtherefore more vigorous action had tobe taken to obtain resistant material.As a consequence, whilst lobbying theTanzania Ministry of Agriculture andCo-operatives for permission toestablish open quarantine in Kagera,tissue culture plantlets were requestedfrom IITA, Ibadan, Nigeria, and anintensive programme of true irrigatedrapid multiplication (IITA, 1998) ofavailable CMD-resistant cultivars wasexpanded at Ukiriguru AgriculturalResearch Institute (ARI), Mwanza. ByJune 1999, cassava planting materialequivalent to almost half a millioncuttings had been multiplied, morethan 10,000 tissue culture plantlets ofCMD-resistant cultivars had beenimported from IITA, Nigeria andproposals for the establishment of openquarantine had been approved.
Germplasm diversification
Broadening the genetic base of cassavais seen as a key aspect of a balancedapproach to the management ofcassava pests and diseases. This isparticularly important with respect tothe CMD pandemic, since a heavyemphasis on the development ofgermplasm with high levels ofresistance to CMD has resulted in less

336
Whiteflies and Whitefly-borne Viruses in the Tropics
attention being given to otherimportant pest and diseaseconstraints. A practical result of this isthat the two varieties most widelymultiplied for CMD control in EastAfrica, SS4 and TMS 30572, arerelatively susceptible to cassava greenmite damage. In more recentgermplasm development work, greateremphasis has been placed on multiplepest and disease resistance andgermplasm with these characteristics isnow becoming available both throughnational breeding programmes andthrough the EARRNET regionalgermplasm development programme.
In Uganda, two demonstration siteshave been established in Rakai andMasaka and include nine cultivars thatare currently being evaluated innational on-farm trials. In Kenya, morethan 500 clones were introduced to theopen quarantine site from theEARRNET programme based at Serereand the best 15 of these weremultiplied for “fast-track” multi-locational testing in 2000. In Tanzania,more than 20 elite cultivars from themultiplication programme are underevaluation in areas of the Kagera regionmost severely affected by thepandemic, and new cultivars with novelWest African landrace-derived sourcesof CMD resistance formed part of therecently imported tissue cultureconsignment from IITA, Nigeria. Eachof these initiatives should help ensurethat a diversity of cultivars is ultimatelymade available to farmers that shouldminimize losses likely to result fromfuture pest or disease outbreaks.
Training
Changes resulting from the CMDpandemic are dramatic, and experienceobtained from Uganda shows thatfarmers are typically unaware of thelikely impact at the outset andfrequently continue with cassavacultivation for a number of years,
suffering substantial yield losses, beforefinally abandoning the crop. Raisingawareness and the provision of trainingin CMD management are keyrequirements if this type of scenario isto be avoided. Training for bothextension workers and farmerstherefore has been an important part ofthe OFDA CMD Project. Various forahave been used for training exercisesincluding:
(1) Stakeholder workshops: held at theoutset of the project in each of thethree countries. Informationprovided on the cause, spread,impact and strategy for themanagement of the CMD pandemic.
(2) In-situ training: farmers andextension workers have beentrained in cassava pestmanagement and multiplicationtechniques at multiplication sites.
(3) Individual training: researchersfrom participating countries havebeen trained in virus diagnostictechniques and have beensponsored to participate in regionalCMD scientific meetings.
(4) Community training: village leadershave been trained in north-westernTanzania to facilitate the transfer ofinformation down to grassrootslevel.
(5) Formal course training: the projecthas helped sponsor courses on seedmultiplication in Tanzania and oncassava production, protection andutilization in Kenya, principallytargeted at local agriculturalworkers.
In addition to the provision oftraining through courses ordemonstration, use has been made ofthe media, particularly in Tanzania,where local radio communications haveprovided information on the CMDpandemic to a much greater number ofhouseholds than could have beenachieved using other methods.

337
Management of the Cassava Mosaic Disease Pandemic in E. Africa
Stakeholder linkages
The key to the success the project hasachieved to date is largely attributableto the active participation of manycassava stakeholders in target areas.These include non-governmentalorganizations, government districtagriculture offices, farmer trainingcentres and projects funded by multi-lateral donors. The inclusion of a broadrange of partners has the twinadvantages of fostering ownership ofthe project at local level and enhancinglocal co-ordination. The development ofa local steering committee for westernKenya, which was charged with theresponsibility for co-ordinating cassavadevelopment and multiplicationactivities in that region, was animportant development in which theOFDA Project participated (ASARECA,1999).
Conclusions
In a relatively short period of time theOFDA CMD Project, affiliated to theTWF-IPM Project, has made significantachievements in both establishing aregional collaborative mechanism forthe monitoring and management of theCMD pandemic and actuallyimplementing control measures. A keyadvantage of the pro-active approachthe project is taking, with its emphasison targeting threatened zones, is thatlead times for the implementation ofcontrol measures may be significantlyreduced compared with previousexperience in Uganda. Given themagnitude of losses associated with theCMD pandemic (Legg et al., 1999b;Otim-Nape et al., 1997; Thresh et al.,1997), this is an importantdevelopment. The multi-facetedapproach being pursued by theTanzania Root and Tuber CropsProgramme with its partners, andsupported by the Project, also isconsidered to be a useful model for
other countries threatened by thepandemic in the near future. Given thelikely continued expansion of thepandemic into other neighbouringcountries, similar such vigorousresponses will be essential if majorlosses are not to continue across thecassava-producing areas of East andCentral Africa.
Acknowledgements
The OFDA CMD Project was fundedthrough a grant from the Office forForeign Disaster Assistance of theUnited States Agency for InternationalDevelopment.
References
ASARECA (Association for theStrengthening of AgriculturalResearch in Eastern and CentralAfrica). 1998. Cassava mosaicpandemic threatens food security.AgriForum 3:1 and 8.
ASARECA (Association for theStrengthening of AgriculturalResearch in Eastern and CentralAfrica). 1999. Fighting cassavamosaic pandemic: Networkingcritical. AgriForum 7:1 and 12.
Deng, D.; Otim-Nape, G. W.; Sangare, A.;Ogwal, S.; Beachy, R. N.; Fauquet,C. M. 1997. Presence of a new virusclosely associated with cassavamosaic outbreak in Uganda. Afr. J.Root Tuber Crops 2:23-28.
GCF (Gatsby Charitable Foundation).1997. Mastering mosaic: The fightfor cassava production in Uganda.Gatsby Occasional Paper, London,GB. 24 p.
Gibson, R. W. 1996. The report of asurvey monitoring the spread of theepidemic of African cassava mosaicvirus from Uganda into westernKenya. Internal report, NaturalResources Institute (NRI), Chatham,GB. 17 p.

338
Whiteflies and Whitefly-borne Viruses in the Tropics
Harrison, B. D; Zhou, X.; Otim-Nape, G.W.; Liu, Y.; Robinson, D. J. 1997.Role of a novel type of doubleinfection in the geminivirus-inducedepidemic of severe cassava mosaicin Uganda. Ann. Appl. Biol 131:437-448.
IITA (International Institute of TropicalAgriculture). 1998. Rapidmultiplication of cassava. A colourpocketbook. Training and Extensionpubl. no. 122, Ibadan, NG. 61 p.
Legg, J. P.; Okao-Okuja, G. 1999.Progress in the diagnosis andepidemiological characterisation ofcassava mosaic geminiviruses inEast Africa. Procs. VIIthInternational Plant VirusEpidemiology Symposium,11-16 April 1999. Aguadulce(Almeria), ES. Abstract: 74-75.
Legg, J. P.; Kapinga, R.; Teri, J.; Whyte, J.B. A. 1999a. The pandemic ofcassava mosaic virus disease inEast Africa: Control strategies andregional partnerships. Roots6(1):10-19.
Legg, J. P.; Sseruwagi, P.; Kamau, J.;Ajanga, S.; Jeremiah, S. C.; Aritua,V.; Otim-Nape, G. W.; Muimba-Kankolongo, A.; Gibson, R. W.;Thresh, J. M. 1999b. The pandemicof severe cassava mosaic disease inEast Africa: Current status andfuture threats. Akoroda, M. O.; Teri,J. M. (eds.). Procs. ScientificWorkshop of the Southern AfricanRoot Crops Research Network(SARRNET), 17-19 August 1998,Lusaka, ZM. p. 236-251.
Ogbe, F. O.; Songa, W.; Kamau, J. W.1996. Survey of the incidence ofAfrican cassava mosaic and EastAfrican cassava mosaic viruses inKenya and Uganda using amonoclonal antibody baseddiagnostic test. Roots 3(1):10-13.
Otim-Nape, G. W.; Bua, A.; Baguma, Y.1994. Accelerating the transfer ofimproved production technologies:Controlling African cassava mosaicvirus disease epidemics in Uganda.Afr. Crop Sci. J. 2:479-495.
Otim-Nape, G. W.; Bua, A.; Thresh, J. M.;Baguma, Y.; Ogwal, S.; Semakula,G. N.; Acola, G.; Byabakama, B.;Martin, A. 1997. Cassava mosaicvirus disease in Uganda: Thecurrent pandemic and approachesto control. Natural ResourcesInstitute (NRI), Chatham, GB. 65 p.
Thresh, J. M.; Otim-Nape, G. W.;Jennings, D. L. 1994. Exploitingresistance to African cassava mosaicvirus. Asp. Appl. Biol. 39:51-60.
Thresh, J. M.; Otim-Nape, G. W.; Legg, J.P.; Fargette, D. 1997. Africancassava mosaic disease: Themagnitude of the problem? Afr. J.Root Tuber Crops. 2:13-19.
Zhou, X.; Liu, Y.; Calvert, L.; Muñoz, C.;Otim-Nape, G. W.; Robinson, D. J.;Harrison, B. D. 1997. Evidence thatDNA-A of a geminivirus associatedwith severe cassava mosaic diseasein Uganda has arisen byinterspecific recombination. J. Gen.Virol. 78:2101-2111.

339
Conclusions
ConclusionsFrancisco Morales*
The Tropical Whitefly Integrated PestManagement (TWF-IPM) Project is one ofthe most ambitious projects everundertaken in the IPM area. Themagnitude of the project responds to theglobal distribution of the pest and thesevere damage caused by whiteflies tomajor food and industrial crops intropical and subtropical agriculturalregions of the world (Jones, 2003). Theemergence of a more prolific andpolyphagous biotype (B) of Bemisiatabaci (Gennadius) has furthercomplicated the control of this pest onalfalfa (Medicago sativa [L.] subsp.sativa), cotton (Gossypium hirsutum L.),common bean (Phaseolus vulgaris L.),tomato (Lycopersicon esculentum Mill.),soybean (Glycine max [L.] Merr.), lettuce(Lactuca sativa L. var. capitata L.), peas(Pisum sativum L.), broccoli (Brassicaoleracea L. var. italica Plenck), collard(Brassica oleracea L. var. viridis L.),cabbage (Brassica oleracea L.) and okra(Abelmoschus esculentus [L.] Moench)(Gruenhagen et al., 1993; Godfrey et al.,1995; Morales and Anderson, 2001). Indeveloping countries of Latin America,sub-Saharan Africa, India and south-east Asia, B. tabaci is an efficient vectorof plant viruses affecting important foodcrops, particularly common bean(Morales and Anderson, 2001), cassava(Manihot esculenta Crantz) (Fargette et
al., 1990; Gibson et al., 1996), tomato(Polston and Anderson, 1997) andpeppers (Capsicum spp. L.) (Morales andAnderson, 2001). In subtropical andmountainous regions, the whiteflyspecies Trialeurodes vaporariorum(Westwood) attacks crops such aspotato (Solanum tuberosum L.), tomato,common bean, several cucurbits anddifferent vegetables (Boiteau and Singh,1988; Cardona, 1995). Until recently,cultivated grasses (Gramineae) wereamong the few plant species spared bywhitefly pests. Within the last decade,devastating whitefly attacks have takenplace in important food and industrialcrops such as rice (Oryza sativa L.),sorghum (Sorghum bicolor [L.] Moench),sugarcane (Saccharum officinarum L.)and pastures (author’s observation). Notsurprisingly, whiteflies were consideredby popular news media (e.g., CNN andNewsweek) as the “pest of the 20thcentury” and their importance as amajor agricultural pest remains high inthe new millennium.
Whiteflies harm plants in differentways by extracting sap, covering plantswith a sticky sugary substance(honeydew) that affects the quality(e.g., sticky cotton) or promotes thegrowth of fungi (sooty mould) on plantsurfaces blocking photosynthesis(Henneberry et al., 1996) and bytransmitting viruses (Markham et al.,1994; Jones, 2003). In heavily infestedplants, hundreds of immature and adult
* Centro Internacional de Agricultura Tropical(CIAT), Cali, Colombia.

340
Whiteflies and Whitefly-borne Viruses in the Tropics
whiteflies can be found feeding on asingle leaf, resulting in rapid plantdeath from the massive loss ofnutrients. Sooty mould can alsoprovoke plant death. And, finally,whitefly-transmitted viruses are amongthe most damaging viral pathogensknown, often causing total yield losses.Genetic immunity to these viruses israre in most of the plant speciesattacked by begomoviruses.
Although whiteflies have beenassociated with agriculture forcenturies, these insects were onlyrecognized as pests in the 1950s (Ernst,1994), coinciding with the developmentand intensive use of agriculturalpesticides following World War II. Half acentury later, most farmers are still notaware of the taxonomic or biologicaldifferences that characterize differentspecies and their physiological variants(biotypes). Nor are they informed aboutthe role of these species as pests and/orvectors of plant viruses. Therefore,farmers apply insecticides wheneverthey see whiteflies on their crops. Inrare instances, farmers who havereceived some technical assistanceapply insecticides when the populationof the pests has reached a pre-determined level, known as the “damagethreshold”. Whereas these “target” or“threshold” applications may beeffective in controlling whiteflies aspests (Chu et al., 1995; Riley andPalumbo, 1995), they are totallyineffective in the case of whitefly-transmitted viruses. Basically, few adultindividuals of a whitefly vector cantransmit a virus long before itspopulation is noticed in the field orreaches a particular density onsusceptible plants.
The introduction of the “B biotype”of Bemisia tabaci in the Americas(Brown and Bird, 1995) has drasticallyincreased the capacity of this whiteflyspecies to cause damage to a larger
number of different cultivated plantspecies and adapt to newenvironments. Farmers that try toescape B. tabaci in the tropical lowlandsand mid-altitude (500-900 m) regions bygrowing their crops at higher altitudesare often disappointed to find yetanother whitefly pest, Trialeurodesvaporariorum, in the highlands. UnlikeB. tabaci, the main whitefly pest andvector of plant viruses in the tropics,T. vaporariorum was considered a merenuisance in crops grown undercontrolled conditions (e.g., glasshouse,screen-house conditions). Currently,T. vaporariorum is also a major pest offield crops in the highlands andtemperate regions of the world and thenumber of plant viruses transmitted bythis species is steadily growing (Jones,2003).
Pesticide abuse is a common factorin the case of the main whitefly pests,regardless of the crops and ecosystemsaffected. The excessive application ofinsecticides, often using inadequatechemicals, alters the delicate balancebetween insect pests and their biologicalcontrol agents (i.e., predators,parasitoids and entomopathogens). Inthe absence of natural enemies, whiteflypopulations increase freely onsusceptible crops and eventuallydevelop resistance to the mostfrequently used insecticides (Omer etal., 1993; Dennehy and Antilla, 1996).Modern agricultural practices, such asthe intensive cropping of diverse plantspecies that act either as suitablefeeding or reproductive hosts, furthercontribute to the exponential increase ofwhitefly pests in disturbedenvironments (Godfrey et al., 1995).Last but not least, climate change hasplayed a major role in the increasingoutbreaks of whitefly pests throughoutthe world. As more forest and wild landsare cleared for agricultural purposes,the climate becomes drier and warmer,conditions that shorten the life cycle of

341
Conclusions
whitefly pests and thus increase theirpopulations (Chu et al., 1995).
Small-scale farmers are trying togrow more profitable crops in order tomaximize the output of their limitedlandholdings. In the absence of propertechnical assistance, farmers faceseveral problems: first, they are notfamiliar with new crops, let alone withtheir phytosanitary problems. Exoticcrops can and will encounter differentpests in their new environments. Often,exotic pathogens and pests areintroduced with imported plantgermplasm. In the case of horticulturalcrops, and particularly vegetables, thenew cultivars have been bred intemperate countries, even though theorigin of some of these plant species isthe tropics (e.g., tomato and peppers).This means that new crops may not bewell adapted to tropical conditions,including their lack of resistance tolocal pests. Although technicalassistance to small- and medium-scalefarmers is not forthcoming, pesticidesalesmen manage to reach every cornerof the rural world. This explains thereliance of farmers on pesticides toprotect their crops.
The main problem of most IPMpractices is their failure to control pestsbefore they cause economic damage.IPM is often wrongly associated with“organic agriculture”, in which chemicalinsecticides do not have a place.Unfortunately, once an agriculturalregion has been significantly modifiedby the introduction of new crops, andits biological equilibrium broken by theintensive use of pesticides, whiteflypopulations are usually too large to becontrolled by non-chemical IPMpractices, such as biological controlagents or yellow traps. Thus, the TWF-IPM Project has been following adifferent approach based on therationalization of chemical control,using the most effective yet specific and
safe systemic insecticides at the propertime. This strategy rapidly reduces thenumber of insecticide applications to aminimum, allowing the recovery of thebeneficial fauna over time. Once thiscondition is met, other IPM strategiescan be incorporated, such as culturalpractices, use of entomopathogens,physical barriers and legal measures,among others. A significant reduction inthe use of pesticides also contributes tolower production costs and higherprofits for farmers, which will becomecritical factors for emerging economiesin the age of free trade agreements. Lastbut not least, men, women and childrenin developing countries areinadvertently consuming vegetablescontaminated with highly toxicpesticides. The medium- and long-termeffects of these toxic residues aredifficult to determine, but they mayrange from chronic illness to life-threatening ailments, including cancer.Reducing pesticide use in vegetablecrops to safe levels should greatlycontribute to vegetables free of toxicpesticide residues, and improved healthstandards in rural and urbanpopulations in developing countries.
The history of cassava mosaicdisease (CMD) is over a century old(Fauquet and Fargette, 1990) and it hasbeen almost half a century sincewhiteflies emerged as pests and vectorsof plant viruses of economic importance.New viruses have been appearingthroughout the tropics and subtropicalregions of the world with a highfrequency (Polston and Anderson,1997). With the advent of moleculartechniques, the list of whitefly-borneviruses has increased exponentially inthe last 3 decades. Altogether,considerable research has beenconducted on these pests, and yet, theirimpact seems to increase every year indeveloping countries. We have alreadydiscussed some of the factors that havecontributed to the unexpected

342
Whiteflies and Whitefly-borne Viruses in the Tropics
dissemination of whiteflies and whitefly-borne viruses. Undoubtedly, the adventof chemical insecticides and theirintensive use in crops, such as cotton,elevated whiteflies to the category ofpests. Another importantepidemiological factor, the diversificationof crops in developing countries, both asexport commodities and cash crops, hasalso played a major role in the increaseof these pests.
The importance of some of the newcrops lies in their role as reproductivehosts to whiteflies (Tsai et al., 1996).These crops need not support very largepopulations of whiteflies, but ratheroccupy large areas or regions. The bestexample is perhaps soybean, which wasrediscovered in Latin America in the1970s as a potential export crop,increasing the total area planted to over10 million hectares. Because individualsoybean plants do not support largepopulations of whiteflies, primarily B.tabaci, soybean growers do not controlthis insect in the soybean crop. Whenthe soybean crops mature at the end ofthe year, common beans are planted,provoking the migration of whitefliesfrom the soybean fields to the youngcommon bean plantings. The search formore profitable crops has led small- andmedium-scale farmers to diversify andintensify their cropping systems,providing an opportunity for whitefliesto dispose of different food sourcesthroughout the year.
Crop improvement has played animportant role in both the onset andcontrol of whitefly-related problems. Inthe case of cassava, the lack of CMD-resistant cultivars and/or the limitedadoption of mosaic-resistant germplasmhas perpetuated the cultivation of CMD-susceptible cassava clones (Morales,2001). Consequently, the causal virusesfind hosts that act as virus reservoirsand permanent sources of inoculum forthe whitefly vector. In sub-Saharan
Africa, this situation has made possiblethe occurrence of multiple infectionsand subsequent recombination ofdifferent CMD-inducing virus species(Pita et al., 2001). The recombinantwhitefly-transmitted begomovirus hascaused severe yield losses in Ugandaand some neighbouring countries,necessitating the implementation of afamine relief project. In the case ofcommon bean, the downsizing of thenational and international programsthat worked on the development ofbegomovirus-resistant bean germplasmin Latin America has resulted in theabandonment of prime agriculturalregions where common beans used tobe produced. This happened because ofthe emergence of more pathogenicbegomoviruses and the breakdown ofthe resistance available in the earliergenerations of improved common beancultivars (Morales, 2001).
The economic crisis that affectsmost national agricultural researchprograms in developing nations hasgreatly affected the flow of technicalinformation to farmers and,consequently, their capacity to managecomplex and severe crop productionconstraints such as the whiteflyproblem. Industrialized nations arecurrently controlling whiteflies with anew generation of systemic insecticidesthat are more efficient and selective(Chao et al., 1997). They are available indeveloping countries, but their price isvery high for most small-scale farmers,who continue to use ineffective andhighly toxic contact insecticides, oftenon a daily basis. These contactinsecticides have practically annihilatedthe beneficial fauna and inducedresistance in whitefly pests, giving riseto very large populations of whitefliesthat can overcome most controlmethods, including systemicinsecticides. Fortunately, some of thenew systemic insecticides are beingproduced as “generic” compounds in

343
Conclusions
developing countries, at lower prices.These problems have forced manyfarmers to abandon the cultivation ofsusceptible crops in prime agriculturalregions.
The TWF-IPM Project has gatheredenough evidence to suggest that thewhitefly problem can be managed withthe collaboration of farmers, byproviding proper and timely technicalassistance. The challenge for the projectis to find the most appropriatecommunication means to disseminatethe IPM technology that has been foundto be effective and economically viablefor small-scale farmers. To this end, theTWF-IPM Project is about to initiate itsthird and final phase to disseminateand scale out suitable IPM technologyusing farmer participatory research anddifferent communication channels toreach affected farmers throughout thetropics, wherever whiteflies are a foodproduction constraint. We are confidentthat well-informed farmers will be ableto reduce the number of application ofinsecticides, lower production costs,increase profits, and reduceenvironmental contamination andhuman health hazards. Ultimately,farmers will be able to recover thebiological equilibrium of theiragricultural ecosystems, and thusprevent future whitefly outbreaks.
The information presented in thisbook represents a major achievement interms of compiling the international andnational (grey) literature and knowledgeavailable on the extent and socio-economic importance of whiteflies aspests and vectors of plant viruses. Thisinformation has been essential inselecting the most promising and viableIPM strategies to be tested in Phase II ofthe TWF-IPM Project. The informationcontained in this book also reflects theinterest and hard work of manycolleagues who now form one of themost extensive research networks in thetropics. This joint effort has already
yielded visible results in terms of raisingawareness about the nature of thewhitefly problem, particularly in relationto the key issue of pesticide abuse, andthe use of appropriate methods tocharacterize whitefly pests and diagnosewhitefly-borne viruses. The design,validation, implementation and successof IPM methods recommended to controlwhiteflies and whitefly-transmittedviruses largely depends upon a solidknowledge base.
References
Boiteau, G.; Singh, P. 1988. Resistance tothe greenhouse whitefly, Trialeurodesvaporariorum (Westwood)(Homoptera: Aleyrodidae), in a cloneof the wild potato Solanum berthaultiiHawkes. Ann. Entomol. Soc. Am.81:428-431.
Brown, J. K.; Bird, J. 1995. Variabilitywithin the Bemisia tabaci speciescomplex and its relation to newepidemics caused by geminiviruses.CEIBA 36:73-80.
Cardona, C. 1995. Manejo de Trialeurodesvaporariorum (Westwood) en frijol enla zona Andina: Aspectos técnicos,actitudes del agricultor ytransferencia de tecnología. MemoriaIV Taller Latinoamericano sobremoscas blancas y geminivirus,16-18 octubre 1995, Zamorano, HN.CEIBA 36(1):53-64.
Chao, S. L.; Dennehy, T. J.; Casida, J. E.1997. Whitefly (Homoptera:Aleyrodidae) binding site forimidacloprid and relatedinsecticides: a putative nicotinicacetylcholine receptor. J. Econ.Entomol. 90:879-882.
Chu, C. C.; Henneberry, T.; Akey, D. H.;Naranjo, S. E.; Perkins, H. H.;Prabhaker, N.; Mackey, B. E. 1995.Silverleaf whitefly: development of anaction threshold for chemical controlon cotton. In: Procs. Beltwide CottonConferences, 4-7 January 1995,San Antonio, TX, USA. Volume 2,p. 873-874.

344
Whiteflies and Whitefly-borne Viruses in the Tropics
Dennehy, T. J.; Antilla, L. 1996. Whiteflyresistance to insecticides inArizona. In: Procs. Beltwide CottonConferences, 9-12 January 1996Nashville, TN, USA. Volume 1,p. 144-145.
Ernst, G. H. 1994. Whiteflies (Bemisiatabaci) in cotton: Possible origin ofthe problem and today’s chemicalcontrol opportunities. Pestic. Sci.42:139-141.
Fargette, D.; Fauquet, C.; Grenier, E.;Thresh, J. M. 1990. The spread ofAfrican cassava mosaic virus intoand within cassava fields. J.Phytopathol. 130: 289-302.
Fauquet, C.; Fargette, D. 1990. Africancassava mosaic virus: Etiologyepidemiology, and control. PlantDis. 74:404-411.
Gibson, R. W.; Legg, J. P.; Otim-Nape,G.W. 1996. Unusually severesymptoms are a characteristic ofthe current epidemic of mosaicvirus disease of cassava in Uganda.Ann. Appl. Biol. 128:479-490.
Godfrey, L. D.; Goodell, P. B.; Summers,C. G.; Bentley, W. J.; Perring, T. M.1995. Seasonal development ofsilverleaf whitefly populations incotton and other crops in theSan Joaquin valley. In: Procs.Beltwide Cotton Conferences,4-7 January 1995, San Antonio, TX,USA. Volume 2, p. 831-834.
Gruenhagen, N. M.; Perring, T. M.;Bezark, L. G.; Daoud, D. M.; Leigh,T. F. 1993. Silverleaf whiteflypresent in the San Joaquin Valley.Calif. Agric. 47:4-6.
Henneberry, T. J.; Hendrix, D. L.;Perkins, H. H.; Jech, L. F.; Burke,R. A. 1996. Bemisia argentifolii(Homoptera: Aleyrodidae) honeydewsugars and relationships to stickycotton. Environ. Entomol.25:551-558.
Jones, D. R. 2003. Plant virusestransmitted by whiteflies. Eur. J.Plant Pathol. 109:195-219.
Markham, P. G.; Bedford, I. D.; Liu, S. J.;Pinner, M. S. 1994. The transmissionof geminiviruses by Bemisia tabaci.Pestic. Sci. 42:123-128.
Morales, F. J. 2001. Conventional breedingfor resistance to Bemisia tabaci-transmitted geminiviruses. Crop Prot.20:825-834.
Morales, F. J.; Anderson, P. K. 2001. Theemergence and dissemination ofwhitefly-transmitted geminiviruses inLatin America. Arch. Virol. 146(3):415-441.
Omer, A. D.; Johnson, M. W.; Tabashnik,B. E.; Ullman, D.E . 1993.Association between insecticide useand greenhouse whitefly (Trialeurodesvaporariorum Westwood) resistance toinsecticides in Hawaii. Pestic. Sci.37:253-259.
Pita, J. S.; Fondong, V. N.; Sangaré, A.;Otim-Nape, G. W.; Ogwal, S.;Fauquet, C. M. 2001. Recombination,pseudorecombination and synergismof geminiviruses are determinantkeys to the epidemic of severecassava mosaic disease in Uganda. J.Gen. Virol. 82:655-665.
Polston, J. E.; Anderson, P. K. 1997. Theemergence of whitefly-transmittedgeminiviruses in tomato in thewestern hemisphere. Plant Dis.81:1358-1369.
Riley, D. G.; Palumbo, P. J. 1995.Abundance of two whitefly species(Homoptera: Aleyrodidae) onGeorgia soybean. J. Entomol. Sci.30:527-533.
Tsai, J. H.; Kaihong, W.; Wang, K. H. 1996.Development and reproduction ofBemisia argentifolii (Homoptera:Aleyrodidae) on five host plants.Environ. Entomol. 25:810-816.

345
Acronyms and Abbreviations
Acronyms and Abbreviations
Acronyms
ACIAR Australian Centre forInternational AgriculturalResearch
AID Agency for InternationalDevelopment
ANPP Association Nationalepour la Protection desPlantes (NationalAssociation for PlantProtection), France
ARI Agricultural ResearchInstitute
ARP African RegionalProgram of AVRD
ARS Agricultural ResearchService of USDA, USA
ASARECA Association for theStrengthening ofAgricultural Research inEastern and CentralAfrica
ASDI Agencia Sueca para elDesarrollo Internacional(Swedish Agency forInternationalDevelopment)
ATC Agriculture TradingCompany, Malawi
AVRDC Asian VegetableResearch andDevelopment Center
BBA BiologischeBundesanstalt für Landund Fortwirtschaft(Federal BiologicalResearch Centre forAgriculture and Forestry),Germany
BMNH British Museum ofNatural History
BNN Banco Nacional deNicaragua (National Bankof Nicaragua)
C&F New Zealand Crop andFood Research
CATIE Centro AgronómicoTropical de Investigacióny Enseñanza (TropicalAgricultural Research andHigher Education Centre),Costa Rica
CENSA Centro Nacional deSanidad Agropecuaria(National Centre for Plantand Animal Health), Cuba
CENTA Centro Nacional deTecnificación Agrícola(National Centre ofAgricultural Technification),El Salvador
CGIAR Consultative Group onInternational AgriculturalResearch
CIAL Comité de InvestigaciónAgrícola Local (LocalAgricultural ResearchCommittee)
CIAS Centro de InvestigacionesAgrícolas del Sureste(South-eastern AgriculturalResearch Centre), Mexico
CIAT Centro Internacional deAgricultura Tropical(International Center forTropical Agriculture), Cali,Colombia

346
Whiteflies and Whitefly-borne Viruses in the Tropics
CIAZA Centro deInvestigacionesAplicadas a Zonas Áridas(Research Centre for AridZones), DominicanRepublic
CINVESTAV Centro deInvestigaciones yEstudios Avanzados(Centre for Research andAdvanced Studies),Irapuato, Mexico
CIP Centro Internacional dela Papa (InternationalPotato Center), Peru
CNIA Centro Nacional deInvestigaciónAgropecuaria (NationalCentre for AgriculturalResearch), Nicaragua
CNRE Centre National deRecherche surl’Environnement(National Centre forEnvironmental Research),Madagascar
CONAL Comisión Nacional delAlgodón (National CottonCommission), Nicaragua
CONVERDS Collaborative Networkfor Vegetable Researchand Development inSouthern Africa
CORPOICA Corporación Colombianade InvestigaciónAgropecuaria (ColombianCorporation ofAgricultural Research)
COSCA Collaborative Study ofCassava in Africa
CRDA Centre de recherche etde documentationagricoles (Centre forAgricultural Research andDocumentation), Haiti
CRS Chitedze ResearchStation, Malawi
CSIRO Commonwealth Scientificand Industrial ResearchOrganisation, Australia
DANIDA Danish InternationalDevelopment Agency
DARNDR Fondo Salvadoreño deEstudios de l’Agriculture,des RessourcesNaturelles et duDéveloppement Rural(Salvadoran Fund forstudies in Agriculture,Natural Resources andRural Development)
DDR German DemocraticRepublic
DFID Department forInternationalDevelopment, UK
DICTA Departamento deInvestigación yCapacitaciónAgropecuaria(Agricultural Researchand TrainingDepartment), Honduras
DIGESA Dirección General deSalud Ambiental (StateEnvironmental HealthOffice), El Salvador
DRC Democratic Republic ofCongo
EAP Escuela AgrícolaPanamericana (Pan-American AgriculturalSchool), Honduras
EARRNET East Africa Root CropsResearch Network
ESARC Eastern and SouthernAfrica Regional Center ofIITA
ESCaPP Ecologically SustainableCassava PlantProtection, regionalproject, Ghana
EXTOXNET Extension ToxicologyNetwork
FAO Food and AgricultureOrganization of theUnited Nations, Italy
FCA Florida Collection ofArthropods

347
Acronyms and Abbreviations
FIRA Fideicomisos Instituidosen Relación con laAgricultura (AgriculturallyRelated Trusts), Mexico
FOFIFA Centre National deRecherche Appliquée auDéveloppement Rural(National Centre forApplied Research in RuralDevelopment), Madagascar
FSCA Florida State Collection ofArthropods
FUNDESA Fundación para elDesarrollo Agropecuario(Foundation forAgricultural Development),Santo Domingo,Dominican Republic
GCF Gatsby CharitableFoundation, UK
GCRIO Global Change ResearchInformation Office, USA
GIIT Grupo Interinstitucionale Interdisciplinario deTomate (Interinstitutionaland InterdisciplinaryTomato Group), Nicaragua
GTZ Deutsche Gesellschaftfür TechnischeZusammenarbeit(German Agency forTechnical Cooperation)
HORTI Horticultural Researchand Training Institute,Tanzania
HPR Host Plant ResistanceProject of MFAT
IARCs International AgriculturalResearch Centres
ICA Instituto ColombianoAgropecuario (ColombianInstitute of Agricultureand Livestock)
ICIPE International Centre ofInsect Physiology andEcology, Kenya
ICTA Instituto de Ciencia yTecnología Agrícolas(Institute of AgriculturalScience and Technology),Guatemala City,Guatemala
IDIAP Instituto de InvestigaciónAgropecuaria de Panamá(Panamanian AgriculturalResearch Institute)
IDRC International DevelopmentResearch Centre, Canada
IICA Instituto Interamericanode Cooperación para laAgricultura (Inter-AmericanInstitute for Cooperation onAgriculture)
IIE Institution ofInternational Education,UK
IITA International Institute ofTropical Agriculture,Nigeria
IITA-ESARC International Instituteof Tropical Agriculture-Eastern and SouthernRegional Center, Nigeria
INIA Instituto Nacional deInvestigación Agraria(National Institute ofAgricultural Research),Mexico
INIAP Instituto NacionalAutónomo deInvestigacionesAgropecuarias (NationalInstitute for AgriculturalResearch), Ecuador
INIFAP Instituto Nacional deInvestigacionesForestales y Agropecuarias(National Institute forAgricultural and ForestryResearch), Sinaloa, Mexico
INISAV Instituto deInvestigaciones deSanidad Vegetal (NationalInstitute for Research onPlant Health), Havana,Cuba
INRAB Institut National deRecherche Agronomiquedu Bénin (Benin NationalInstitute for AgriculturalResearch)

348
Whiteflies and Whitefly-borne Viruses in the Tropics
IOBC InternationalOrganisation forBiological and IntegratedControl for NoxiousAnimals and Plants
IRA Institut de RechercheAgronomique(Agricultural ResearchInstitute), Cameroon
ISA Instituto Superior deAgricultura (HigherInstitute for Agriculture),Dominican Republic
ISTRC-AB International Society forTropical Root Crops-Africa Branch
JIC John Innes Centre, UKKARI Kenya Agricultural
Research InstituteLILIANA Instituto de
Investigaciones deHortícolas “LilianaDimitroya” (HorticulturalResearch Institute“Liliana Dimitroya”), Cuba
LZARDI Lake Zone AgriculturalResearch andDevelopment Institute,Tanzania
MAG Ministerio de Agriculturay Ganadería (Ministry ofAgriculture), Nicaragua
MARNDR Ministère de l’agriculture,des ressources naturelleset du développement rural(Ministry of Agriculture,Natural Resources andRural Development), Haiti
MFAT Ministry of ForeignAffairs and Trade, NewZealand
MIP Manejo Integrado dePlagas (Integrated PestManagement)
MSU Montana State University,USA
NAARI Namulonge Agriculturaland Animal ProductionResearch Institute,Uganda
NARES National AgriculturalResearch and ExtensionSystems
NARO National AgriculturalResearch Organisation,Uganda
NORAD Norwegian Agency forCo-operation forDevelopment
NRCRI National Root CropsResearch Institute,Nigeria
NRI Natural ResourcesInstitute, UK
NRTCIP National Root and TuberCrops ImprovementProject, Ghana
OFDA Office for ForeignDisaster Assistance ofUSAID
OIRSA Organismo InternacionalRegional de SanidadAgropecuaria (RegionalInternational Organizationfor Plant Protection andAnimal Health),Nicaragua
PAUP Phylogenetic AnalysisUsing Parsimony
PCCMCA Programa CooperativoCentroamericano para elMejoramiento de CultivosAlimenticios(CentroamericanCooperative Programme forthe Improvement of FoodCrops), Guatemala
PHYLIP Phylogeny InferencePackage
PNUMA Programa de las NacionesUnidas para el MedioAmbiente (United NationsEnvironment Programme),Geneva
PPRI Plant Protection ResearchInstitute, South Africa
PPRSD Plant Protection andRegulatory ServicesDirectorate, Ghana

349
Acronyms and Abbreviations
PROFRIJOL Programa CooperativoRegional de Frijol paraCentro América, Méxicoy el Caribe (Regional Co-operative BeanProgramme for CentralAmerica, Mexico and theCaribbean)
PROMIPAC Programa Regional deManejo Integrado dePlagas en AméricaCentral (RegionalIntegrated PestManagement Program forCentral America)
REDCAHOR Red Colaborativa deInvestigación yDesarrollo de Hortalizaspara América Central,Panamá y RepúblicaDominicana(Collaborative Network forVegetable Research andDevelopment for CentralAmerica, Panama andDominican Republic)
SADC Southern AfricaDevelopment Community
SARH Secretaría de Agriculturay Recursos Hidráulicos(Secretariat of Agricultureand HydraulicResources), Mexico
SARI Selian AgriculturalResearch Institute,Tanzania
SARRNET Southern African RootCrops Research Network
SDC Swiss Agency forDevelopment andCooperation
SEA Secretaría del Estado deAgricultura (StateAgricultural Secretariat),Dominican Republic
SENASA Servicio Nacional deSalud Animal (NationalService for AnimalHealth), El Salvador
SP-IPM Systemwide Programmeon Integrated PestManagement
TRTCP Tanzania Root and TuberCrops Programme
TWF-IPM Tropical WhiteflyIntegrated PestManagement Project
UA University of Arizona-Tucson, USA
UFL University of Florida-Gainesville, USA
UNA Universidad NacionalAgraria (NationalAgricultural University),Nicaragua
UNDP United NationsDevelopment Programme
USAID United States Agency forInternationalDevelopment
USDA United StatesDepartment ofAgriculture
USNM United States NationalMuseum
UTLA Universidad TécnicaLatinoamericana (LatinAmerican TechnicalUniversity)
UW University of Wisconsin-Madison, USA
Abbreviations
AC1 replicaseACMD African cassava mosaic
diseaseACMV African cassava mosaic
virusA.f. Aleurothrixus floccosusAFLP amplified fragment
length polymorphismANOVA analysis of varianceA.p. Aleyrodes proletellaAV1 capsid proteinB.a. Bemisia aferBCaMV Bean calico mosaic virusBDMV Bean dwarf mosaic virusBG begomovirusBGMV Bean golden mosaic virusBGYMV Bean golden yellow
mosaic virusB.h. Bemisia hirtaBMMV Bean mild mosaic virus

350
Whiteflies and Whitefly-borne Viruses in the Tropics
BR BrazilB.t. Bemisia tabaciB.t.-A Bemisia tabaci A biotypeB.t.-B Bemisia tabaci B biotypeC plant infected as cuttingCAR carbamatesCLCuV Cotton leaf curl virusCMD cassava mosaic diseaseCMG cassava mosaic
geminivirusCMV cucumoviruses,
Cucumber mosaic virusCO ColombiaCOI cytochrome oxidase ICPMMV Cowpea mild mottle virusCVYV Cucumber vein yellowing
virusDAS double antibody
sandwichDNA deoxyribonucleic acidDR Dominican RepublicDSM Dar es SalaamE early infected plantEACMV East African cassava
mosaic virusEg EgyptEIL economic injury levelELISA enzyme-linked
immunosorbent assayEM electron microscopyEMIF electronic monitoring of
insect feedingEPG electrical penetration
graphES El SalvadorFFS farmer field schoolFL fiducial limitsGA GuatemalaGIS geographic information
systemsGV geminivirusesH healthy, symptomless
plantHD HondurasHPR host plant resistantHR highly resistantHS highly susceptibleI intermediate infected
plantI intermediate resistance
ICMV Indian cassava mosaicvirus
IPM integrated pestmanagement
Is IsraelITS internal transcribed
spacersL late infected plantLC lethal concentrationLSD least significant
differenceMABs monoclonal antibodiesMAB-BS a broad spectrum
monoclonal antibodyused to detect bi-partitebegomoviruses
MAB-GA a monoclonal antibodyused to detect theoriginal Middle Americanisolates of Beangolden yellow mosaicvirus-Guatemala
MAP months after plantingMIU multiple infection unitsMo MochisMR moderately resistantMYMV Mungbean yellow mosaic
virusNARS national agricultural
research systemsNCM nitrocellulose membranent not testedO.c. Orchamoplatus citriOLCV Okra leaf curl virusOPs organophosphate
insecticidesORF open reading framePAGE polyacrylamide gel
electrophoresisPCR polymerase chain
reactionPepGMV Pepper golden mosaic
virusPepLCV Pepper leaf curl virusPHYVV Pepper Huasteco yellow
vein virusPR Puerto RicoPTY1 monoclonal antibody to
detect potyvirusesPVY Potato virus Y

351
Acronyms and Abbreviations
PYR pyrethroidsQTL quantitative trait locusR whitefly resistant cultivarRAPD random amplified
polymorphic DNARF resistance factorRFLPs restriction fragment
length polymorphismsRH relative humidityS susceptibleS whitefly susceptible
cultivarSACMV South African cassava
mosaic virusSCAR sequence characterized
amplified regionSEA East African strainSiGMV Sida golden mosaic virusSLCV Squash leaf curl virusS.p. Siphoninus phillyreaeSPCFV Sweetpotato chlorotic
fleck virusSPCSV Sweetpotato chlorotic
stunt virusSPFMV Sweetpotato feathery
mottle virusSPLV Sweetpotato latent virusSPMMV Sweetpotato mild mottle
virusSPSV-SEA East African strain of
SPCSVSPVD sweetpotato virus
diseaseT whitefly tolerant cultivarT.a. Tetraleurodes andropogon
Tan TanzaniaTAS triple antibody sandwichTBR tree-bisection
reconnectionTEV Tobacco etch virus
(Potyviridae: Potyvirus)TLC tomato leaf curl diseaseTLCV Tomato leaf curl virus,
now ToLCVTMV Tobacco mosaic virus
(Tobamovirus)ToLCV Tomato leaf curl virusToMoTV Tomato mottle Taino virusToMoV Tomato mottle virusToYMoV Tomato yellow mottle
virusToYMV Tomato yellow mosaic
virusTPV Texas pepper virus
(= Pepper golden mosaicvirus)
T.r. Trialeurodes riciniT.v. Trialeurodes vaporariorumTVTV Tomato vein thickening
virusTYLCV Tomato yellow leaf curl
virusUg UgandaWmCSV Watermelon chlorotic
stunt virusWTVs whitefly-transmitted
virusesXam Xanthomonas axonopodis
pv. manihotis


















