
6.4 Regulation at the Level of Translation - uni … · 6.4 Regulation at the Level of Translation...
103
6.4 Regulation at the Level of Translation 6.4.1 The components 6.4.2 Initiation 6.4.3 Elongation 6.4.4 Termination 6.4.5 Alternative translation events 6.4.6 Translational control of gene expression 6.4.7 Additional amino acids 6.4.8 Posttranslational regulation
Transcript of 6.4 Regulation at the Level of Translation - uni … · 6.4 Regulation at the Level of Translation...

6.4 Regulation at the Level of Translation
6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation

6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation

The Components
1. The tRNAs 2. The aminoacyl-tRNA synthetases3. The 70S ribosomes 4. The mRNAs

The tRNAs
1. They translate individual codons of the mRNA into the corresponding amino acid
2. They are recognized and aminoacylated specifically by their cognate aminoacyl-tRNA synthetases
3. They show a conserved cloverleaf-shaped secondary structure that folds into an L-shaped tertiary structure
4. Average length: 76 nucleotides

The Aminoacylation Reaction
First step: Activation at the active site with ATPSecond step: Transfer to the 3‘ end of the tRNA
J Ling (2009) Annu Rev Microbiol 63: 61
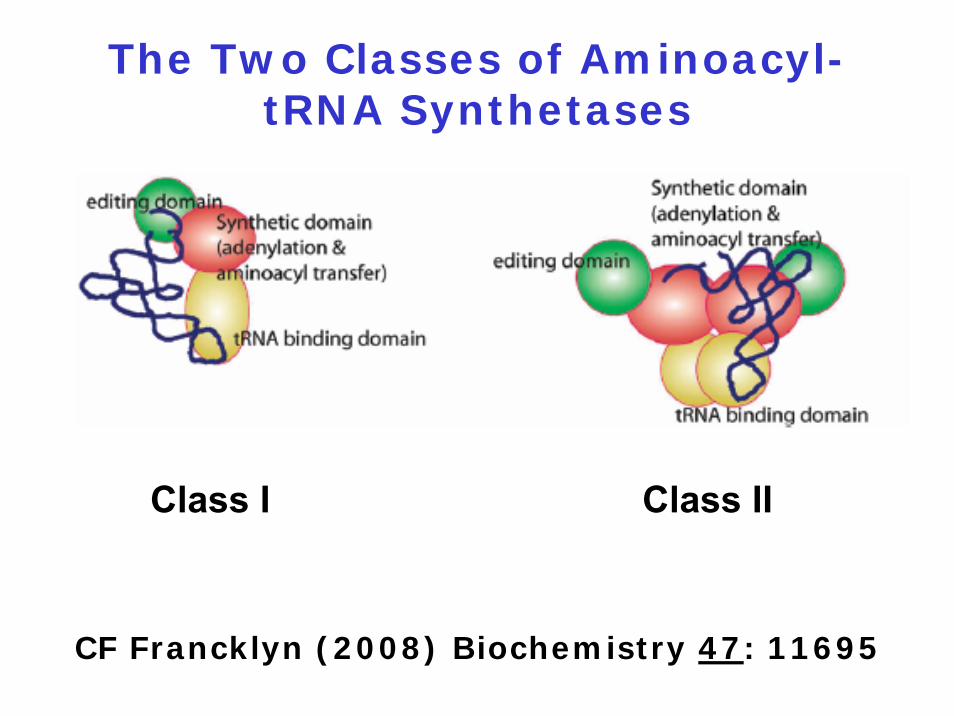
The Two Classes of Aminoacyl-tRNA Synthetases
Class I Class II
CF Francklyn (2008) Biochemistry 47: 11695

The 70S Ribosome
30S ribosomal subunit:- 16S RNA- 21 proteins
50S ribosomal subunit:- 5S and 23S RNA- 34 proteins
Three sites called A, P and E
Peptidyltransferase center: Start of an 100 Å long tunnel, 15 Å diameter

The mRNA
Two types of mRNAs:CanonicalLeaderless
Canonical mRNA: Contains an 5' untranslated region (5' UTR) with
Shine-Dalgarno (SD) sequenceStart codonS1 binding site (pyrimidine-rich)Sometimes regulatory sequences
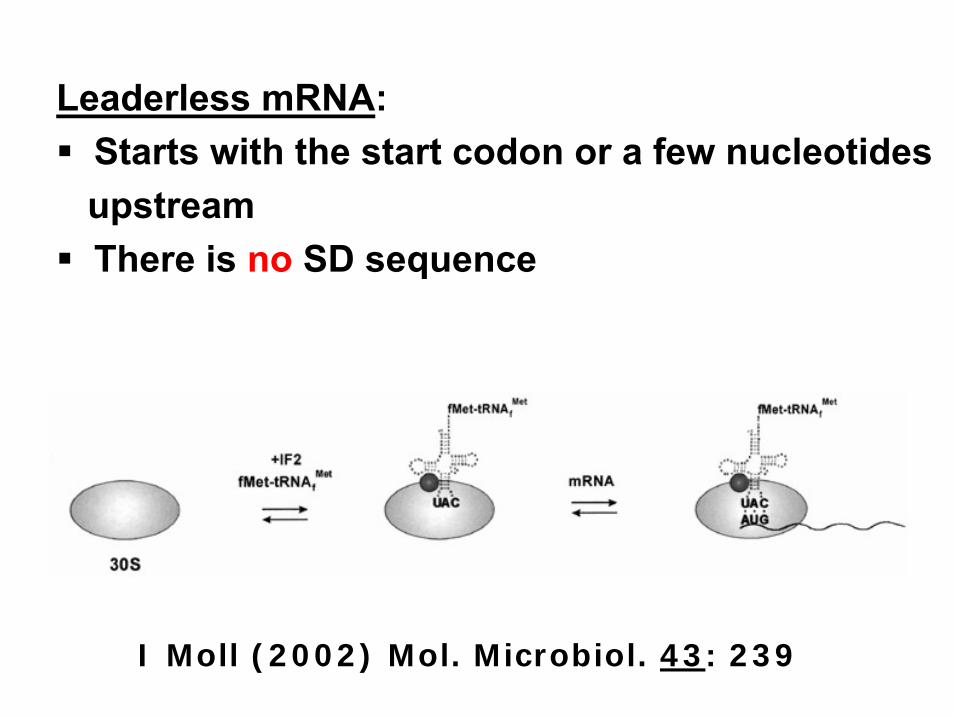
Leaderless mRNA:Starts with the start codon or a few nucleotidesupstreamThere is no SD sequence
I Moll (2002) Mol. Microbiol. 43: 239

Downstream Box (DB):
Occurs downstream from the initiation codonCan potentially base-pair with a region within the16S rRNANo genetic and biochemical evidence for thisinteraction

6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation
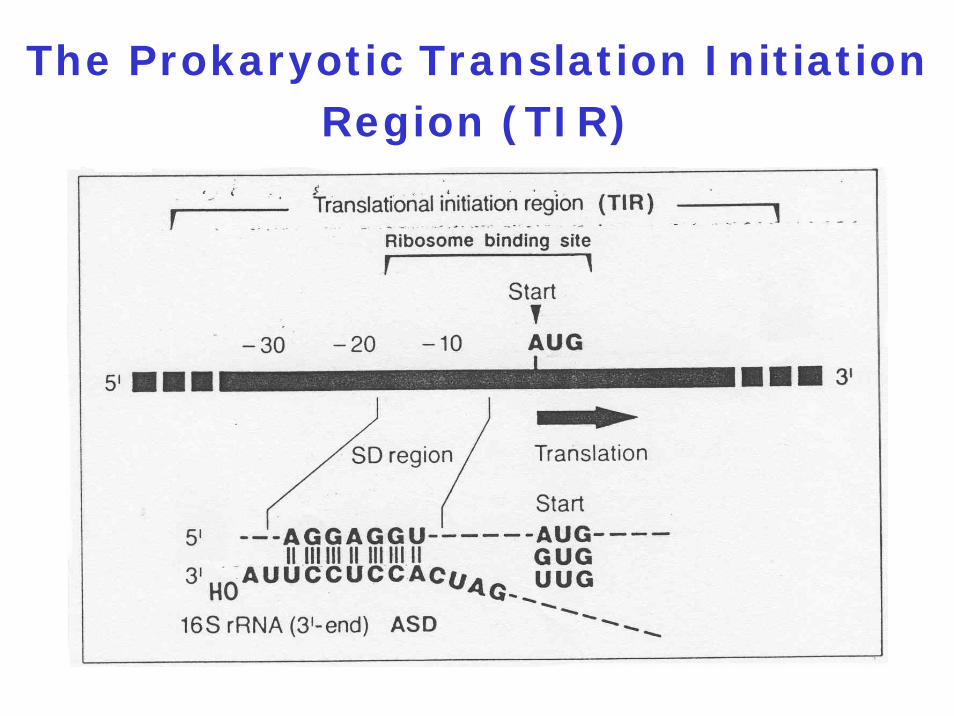
The Prokaryotic Translation Initiation Region (TIR)

6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation

Two mechanisms compete with
unimpaired elongation:
1. Drop-off
2. SsrA-tagging


Minigenes in Phage Lambda DNA

What happens when translating ribosomes arrive at the 3' end of a mRNA molecule devoid of its stop
codon ?
Will they dissociate spontaneously ? Will they stay stalled forever ? Is there a mechanism to release them from the mRNA ?

By which mechanisms mRNA molecules without a stop codon can
arise ?
Premature transcription termination
Endoribonucleolytic cleavage

The SsrA-Tagging Mechanism

Physiological roles of the tmRNA-
dependent peptide-tagging system:
1. Clearance and recycling of stalled ribosomes
2. Degradation of abortive proteins by cellular ATP-dependent proteases
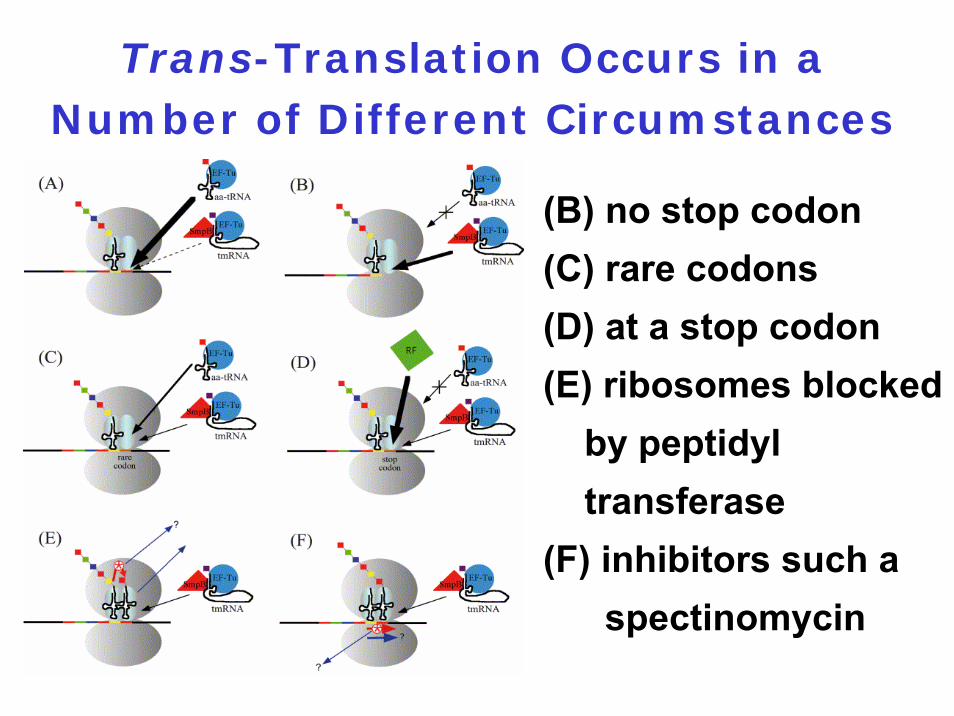
Trans-Translation Occurs in a Number of Different Circumstances
(B) no stop codon(C) rare codons(D) at a stop codon(E) ribosomes blocked
by peptidyl transferase
(F) inhibitors such a spectinomycin

Model for Trans-Translation Acting on the lacI mRNA
T Abo (2000) EMBO 19: 3762

Translational Coupling
Definition:Translation of the downstream genes needs translation of the upstream gene

Direct coupling:SD or/and initiation codon of the downstream gene is part of a secondary structure of the mRNA which is destroyed by translating ribosomes
Mechanisms of Translational Coupling
Indirect coupling:If the upstream gene cannot become completely translated, the downstream gene will be inactivated, e.g., by an endonuclease

Biological Function of Translational
Coupling
1. Ribosomes will not dissociate at the first stop codon
2. Ensures synthesis of equimolar amounts of the two proteins

6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation

Components of Translation
Termination
1. Three stop codons: UAG, UGA, UAA
2. Four release factors: RF1, RF2, RF3, RRF (RF4)

Function of the Four Release Factors
RF1: decodes UAG and UAA
RF2: decodes UGA and UAA
RF3: catalyzes dissociation of RF1 and RF2 from the ribosome
RF4: catalyzes dissociation of the ribosomal subunits
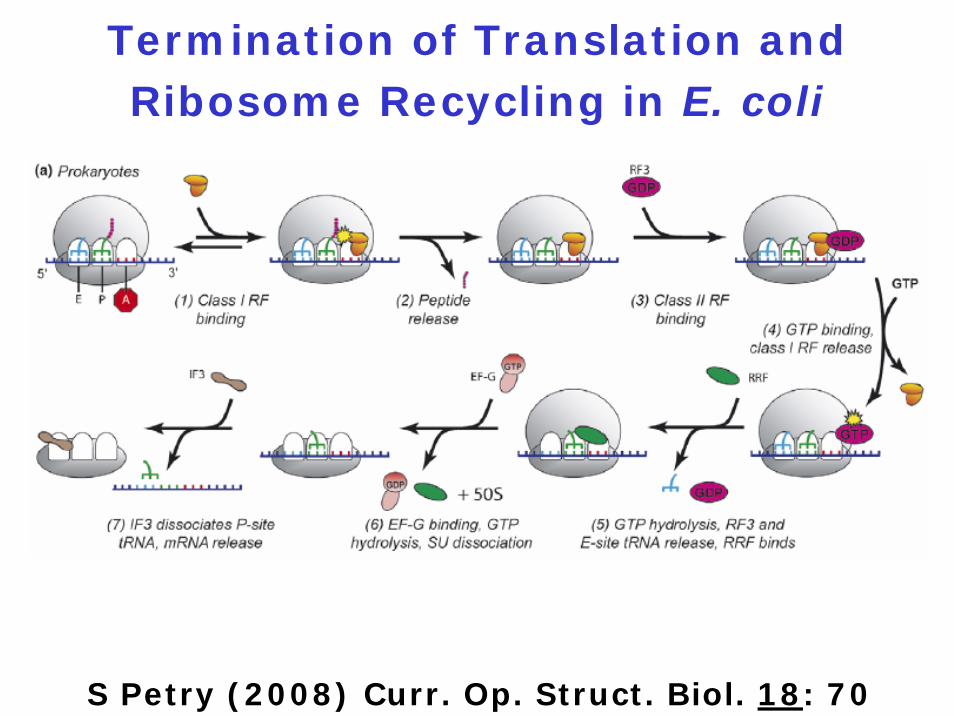
Termination of Translation and Ribosome Recycling in E. coli
S Petry (2008) Curr. Op. Struct. Biol. 18: 70

6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation

Read-through:1. A tRNA decodes the stop codon2. Translation continues until the next stop codon
Frameshifting:Ribosome switches to one of the two alternative frames
Bypassing:Extensive peptidyl-tRNA slippage and re-pairing with the mRNA at a non-overlapping codon
Alternative Translation Events

1. Autoregulation: To limit the amount of a protein, e.g., RF2
2. To yield two different proteins, e.g, dnaXgene codes for the two proteins τ and γ
Biological Functions of Frameshifting

Pausing site
Slippery (shifty) sequence
Components of Programmed
Frameshifting

1. Termination codon
2. Stem-loop structure
3. Pseudoknot
4. Hungry codon
Pausing site can be
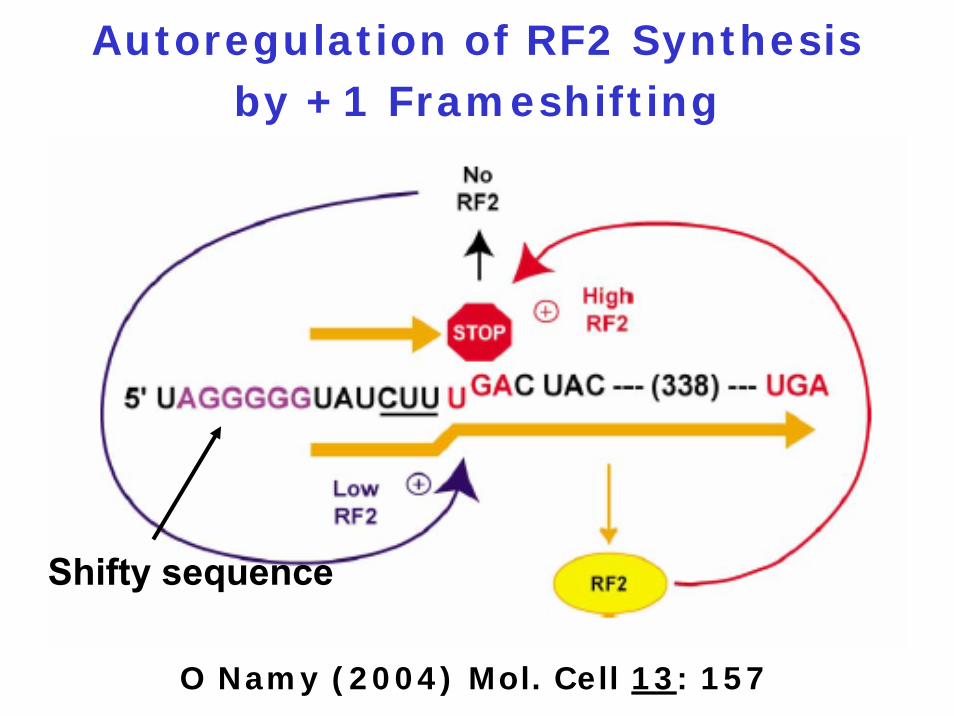
Autoregulation of RF2 Synthesis by +1 Frameshifting
O Namy (2004) Mol. Cell 13: 157
Shifty sequence
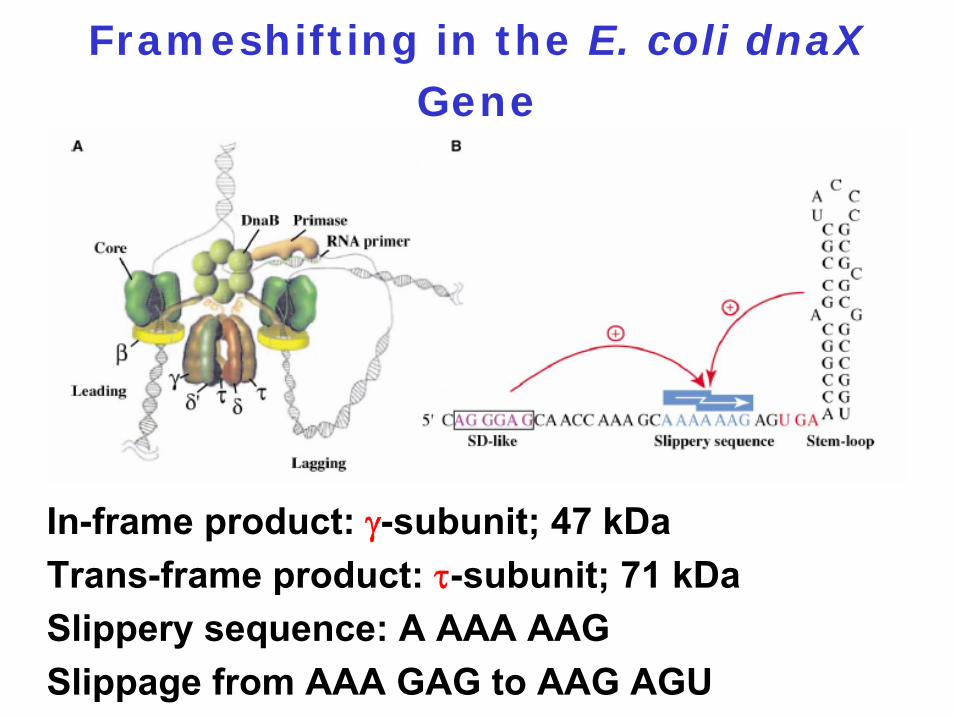
Frameshifting in the E. coli dnaXGene
In-frame product: γ-subunit; 47 kDaTrans-frame product: τ-subunit; 71 kDaSlippery sequence: A AAA AAGSlippage from AAA GAG to AAG AGU

Definition:
Joins the information found within two disparate open reading frames into a single polypeptide chain
Translational Bypassing
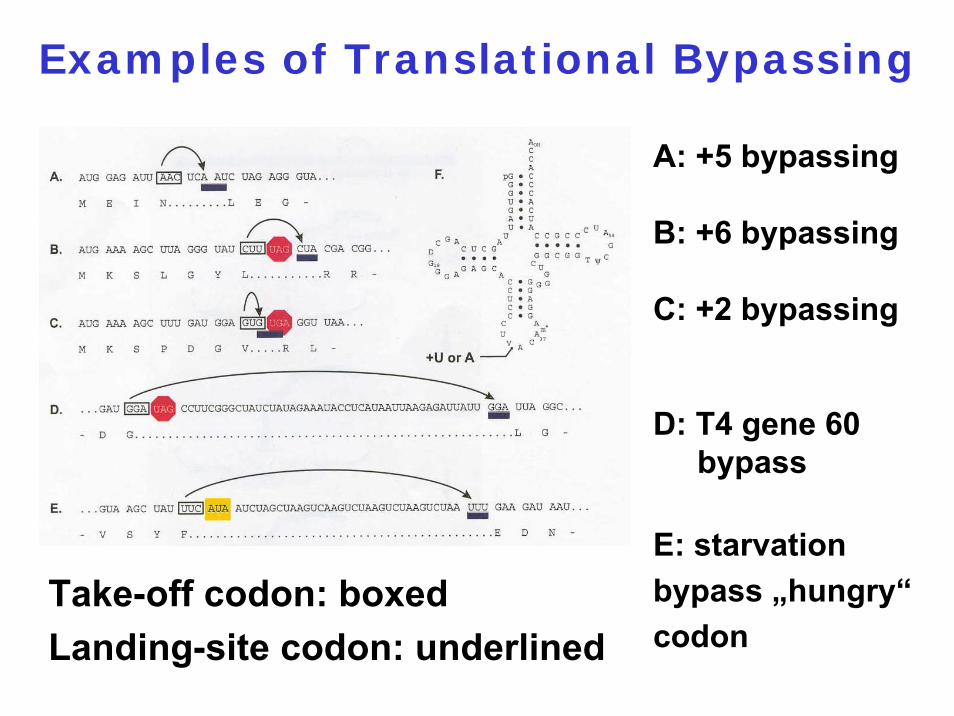
Examples of Translational Bypassing
Take-off codon: boxedLanding-site codon: underlined
A: +5 bypassing
B: +6 bypassing
C: +2 bypassing
D: T4 gene 60 bypass
E: starvation bypass „hungry“codon
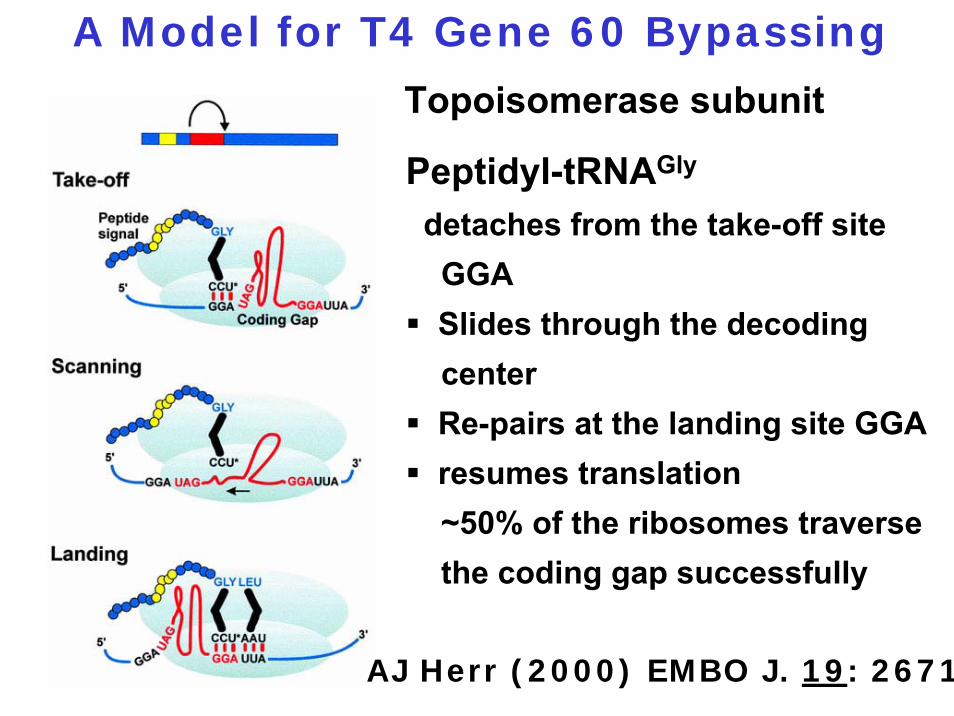
A Model for T4 Gene 60 Bypassing
Peptidyl-tRNAGly
detaches from the take-off site GGASlides through the decoding centerRe-pairs at the landing site GGAresumes translation~50% of the ribosomes traverse the coding gap successfully
Topoisomerase subunit
AJ Herr (2000) EMBO J. 19: 2671

1. The matching GGA codons2. An optimal spacing of 47 nucleotides
between the two GGA codons3. A nascent peptide signal4. A stem-loop structure just downstream of
the take-off site5. A UAG stop codon6. An SD-like interaction
Signals involved in bypassing:

6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation
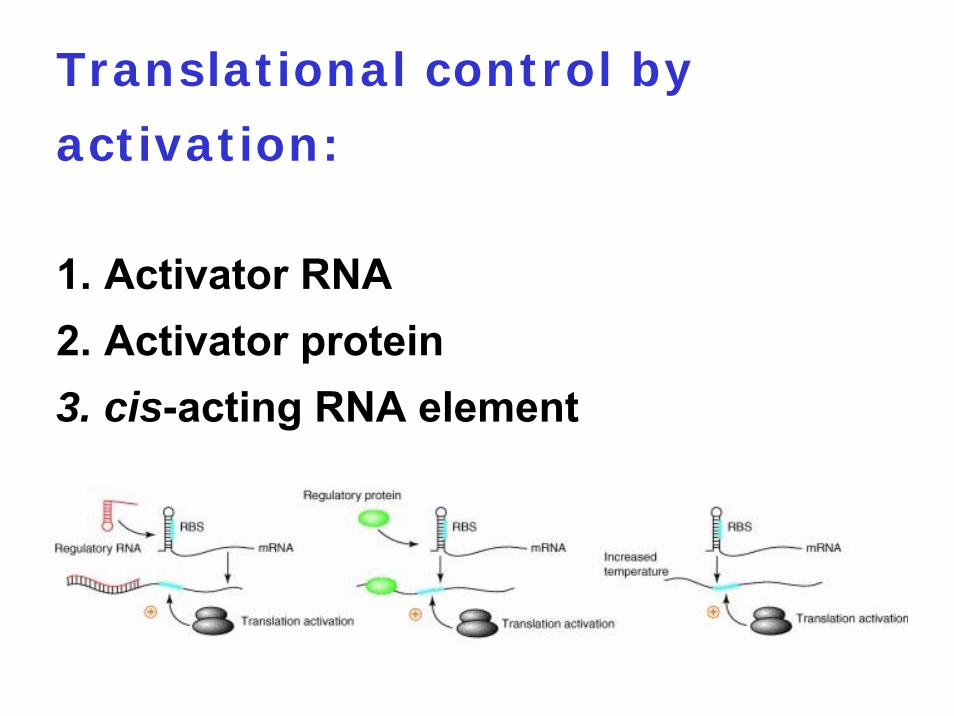
Translational control by
activation:
1. Activator RNA2. Activator protein3. cis-acting RNA element
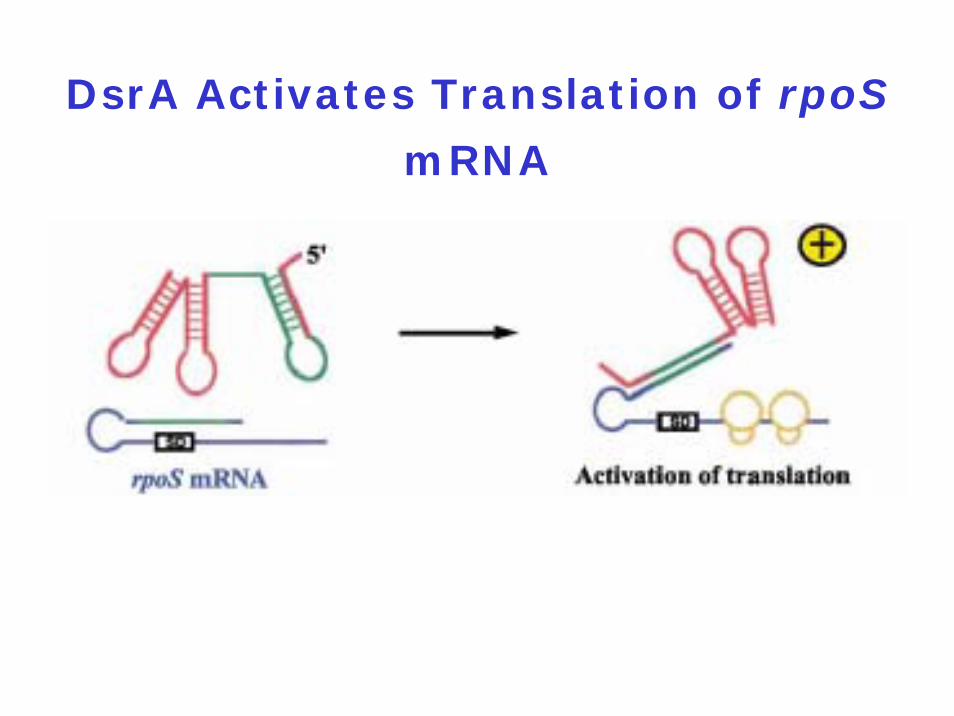
DsrA Activates Translation of rpoS
mRNA
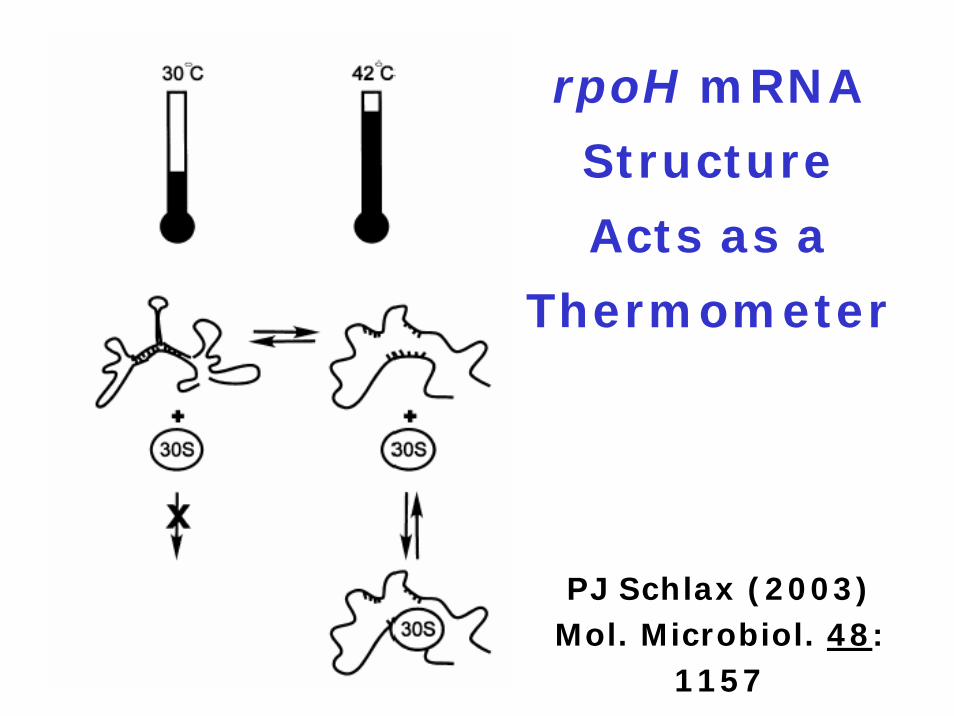
rpoH mRNA
Structure
Acts as a
Thermometer
PJ Schlax (2003) Mol. Microbiol. 48:
1157
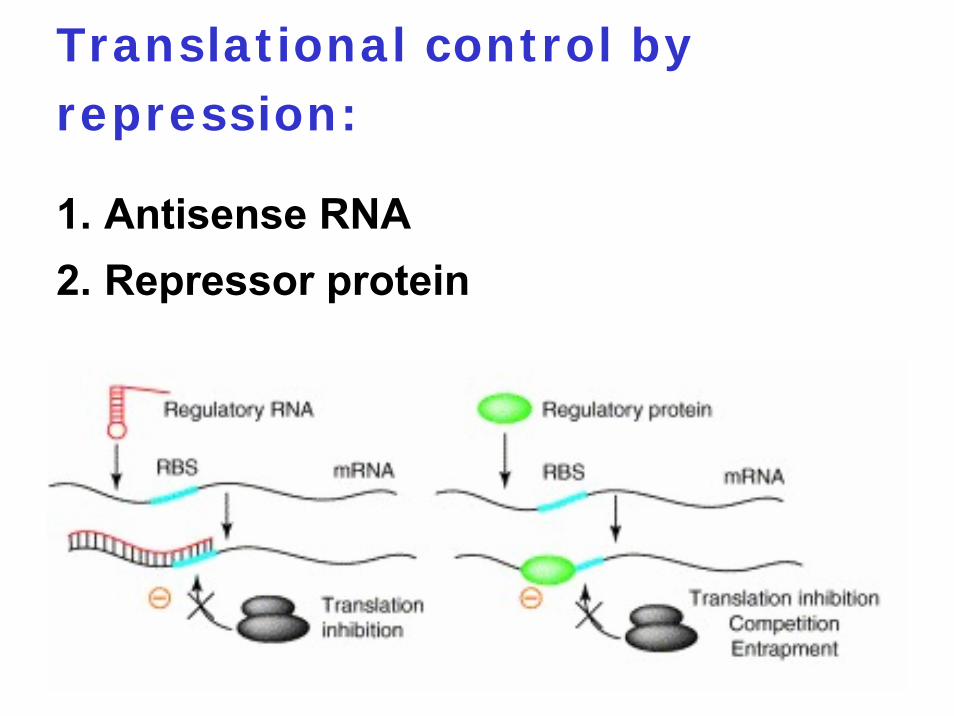
Translational control by repression:
1. Antisense RNA2. Repressor protein
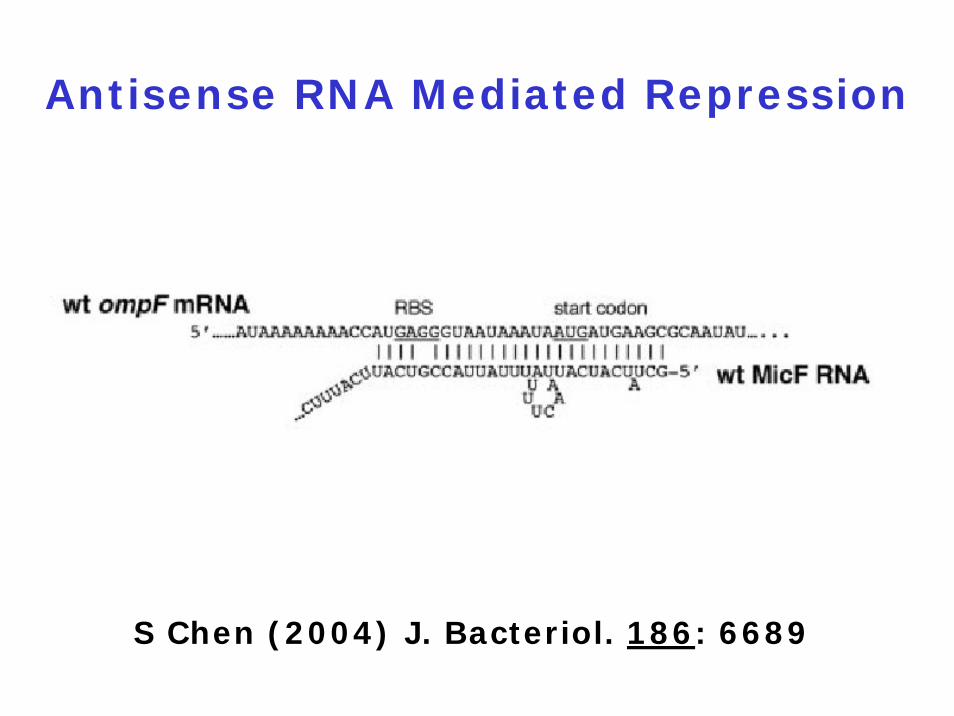
Antisense RNA Mediated Repression
S Chen (2004) J. Bacteriol. 186: 6689
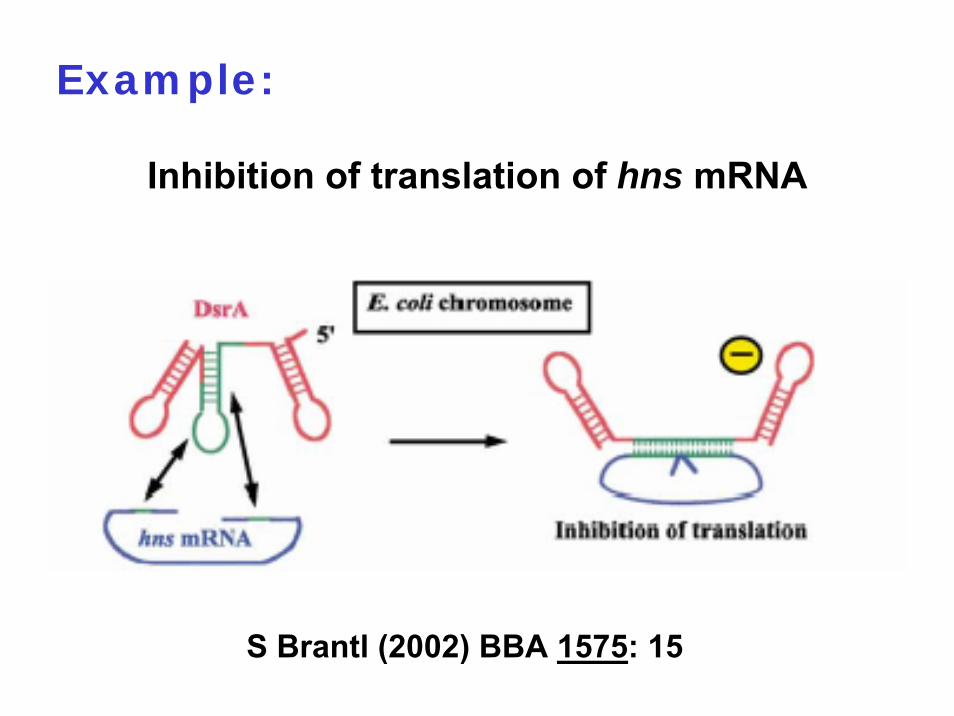
Example:
Inhibition of translation of hns mRNA
S Brantl (2002) BBA 1575: 15

Protein-Mediated Translational
Repression Mechanisms
1. Direct competition model
2. Entrapment model
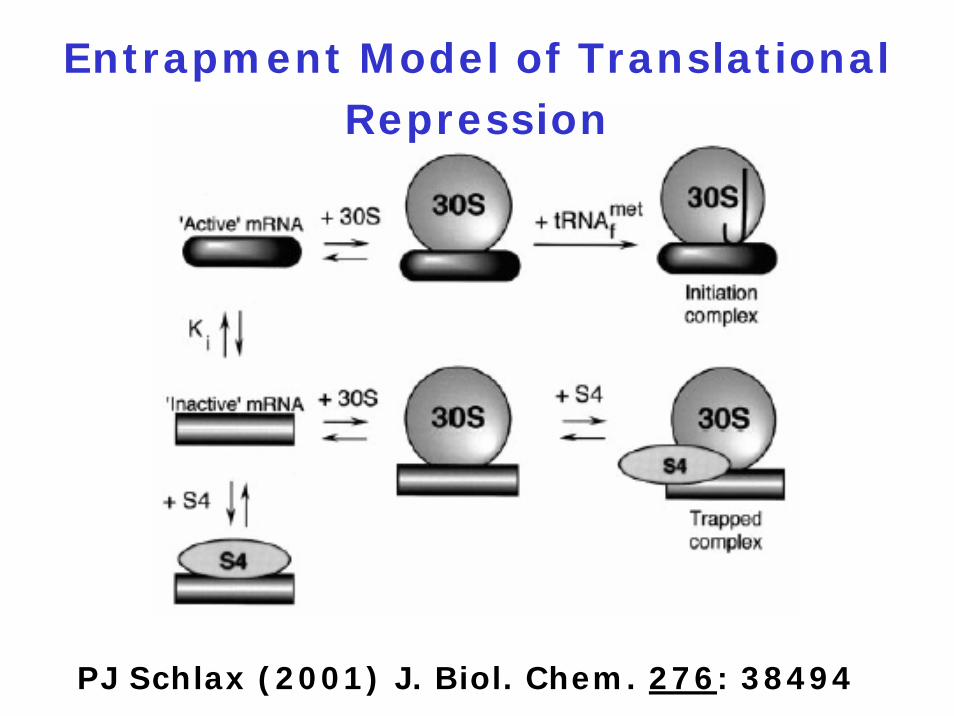
Entrapment Model of Translational Repression
PJ Schlax (2001) J. Biol. Chem. 276: 38494

6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation

Summary of Genetic Code Changes
A Ambrogelly(2007) Nature
Chem Biol 3: 29

Is a stop codon always a stop codon ?
Classical stop codons: UAA, UAG, UGA
1. Mitochondrial genomes UGA = tryptophan codon tRNATrp recognizes both UGG and UGA
2. Mycoplasma capricolumUGA = tryptophan codon
3. Many eubacteria and archaeaUGA = selenocystein (Sel) UGA = pyrrolysine (Pyl)
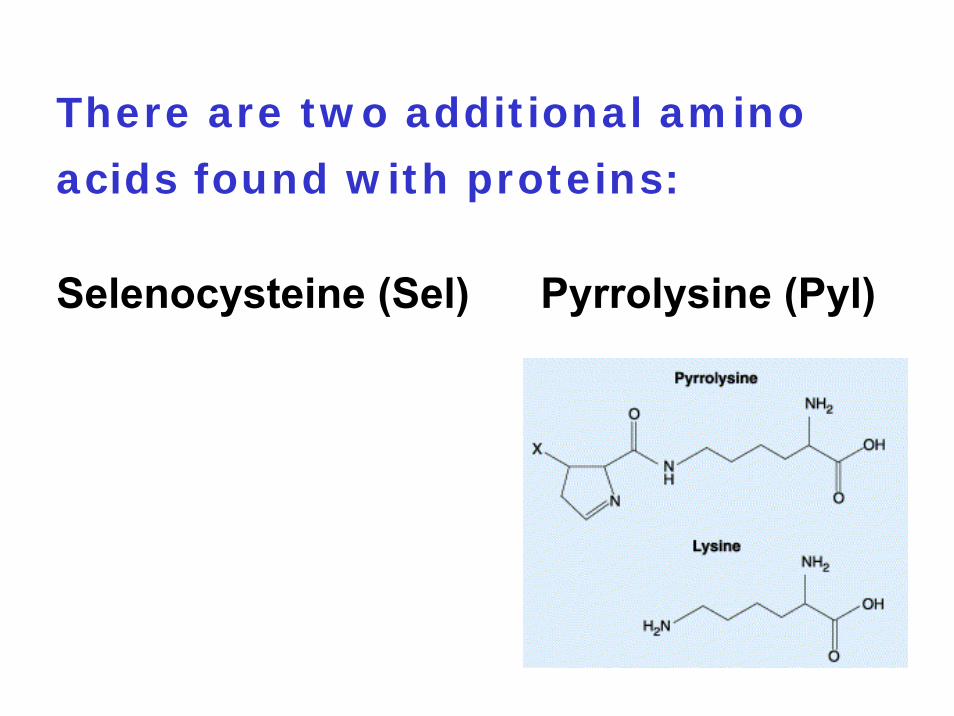
There are two additional amino
acids found with proteins:
Selenocysteine (Sel) Pyrrolysine (Pyl)

Properties of the Amino Acids
Seleno-cysteine and Cysteine
L Johansson (2005) BBA, in press

1. Selenium is a trace element incorporated into proteins in the form of selenocysteine (Sec) in all three kingdoms
2. Found mainly in the active site of oxidoreductases; negatively charged at physiological pH and highly reactive
3. Incorporated into proteins during translation elongation
4. Encoded by a combination of internal UGA stop codon and a specific mRNA hairpin structure = SECIS (selenocysteine insertion element)
Selenocysteine

The Four sel Genes of E. coli
selA: Selenocysteine synthetase; converts the seryl moiety into the selenocysteyl derivative
selB: specific elongation factor (EF-Tu)selC: tRNASec; charged with L-serine and serves
as an adapter at which the seryl moiety is converted into the selenocysteyl derivative
selD: Selenophosphate synthetase; synthesizes monoselenophosphate from selenide and ATP

The specialized SelB elongation factor binds three different ligands:
1. GTP2. Sec-tRNASec
3. The SECIS element (in bacteria located immediately 3' of the UGA codon)

In E. coli, three formate
dehydrogenases contain selenium:
1. fdoG: expressed constitutively2. fdhf: expressed under anaerobic conditions3. fdnG: expressed under anaerobic conditions
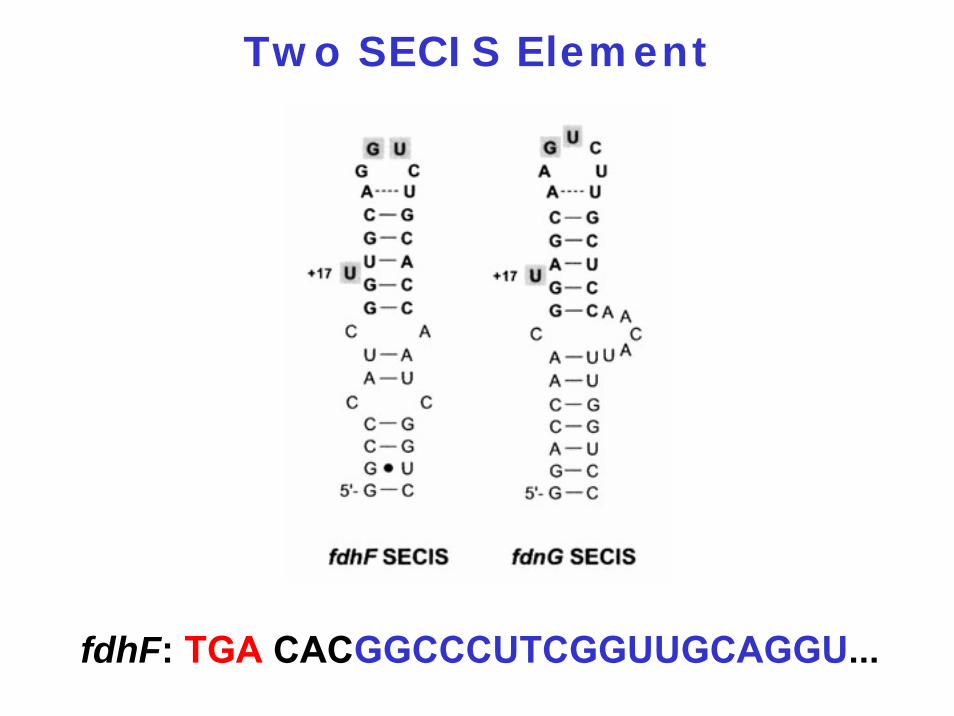
Two SECIS Element
fdhF: TGA CACGGCCCUTCGGUUGCAGGU...

M Thanbichler (2002) EMBO J. 21: 6925
Proposed Model of the Reactions Involved in the Regulation of selAB
Expression
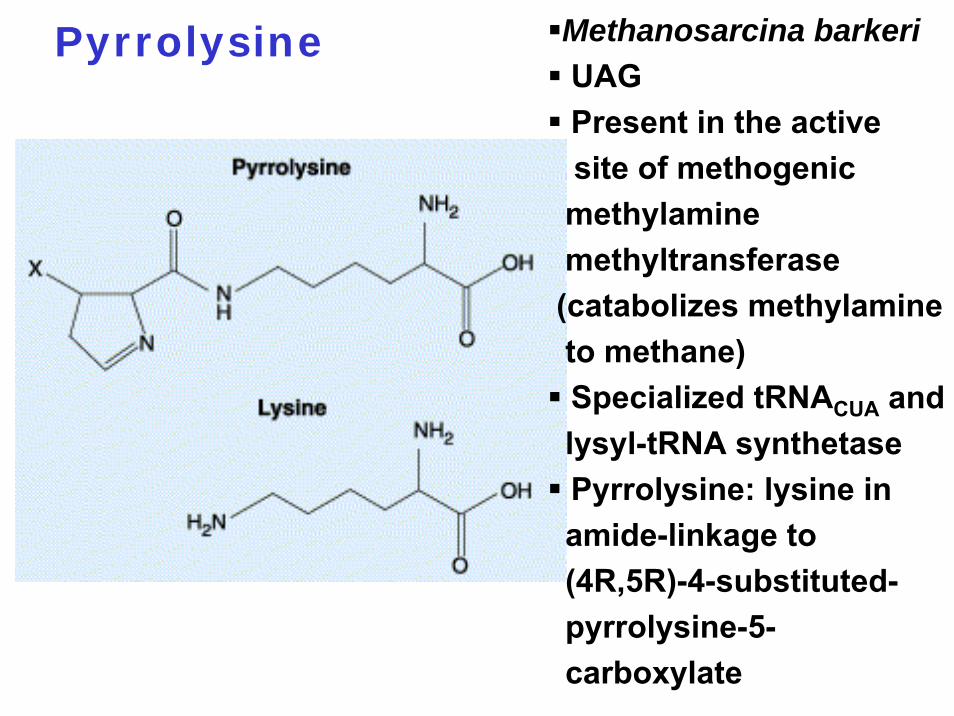
Pyrrolysine Methanosarcina barkeriUAGPresent in the active site of methogenicmethylaminemethyltransferase (catabolizes methylamineto methane)Specialized tRNACUA and lysyl-tRNA synthetasePyrrolysine: lysine in amide-linkage to (4R,5R)-4-substituted-pyrrolysine-5-carboxylate
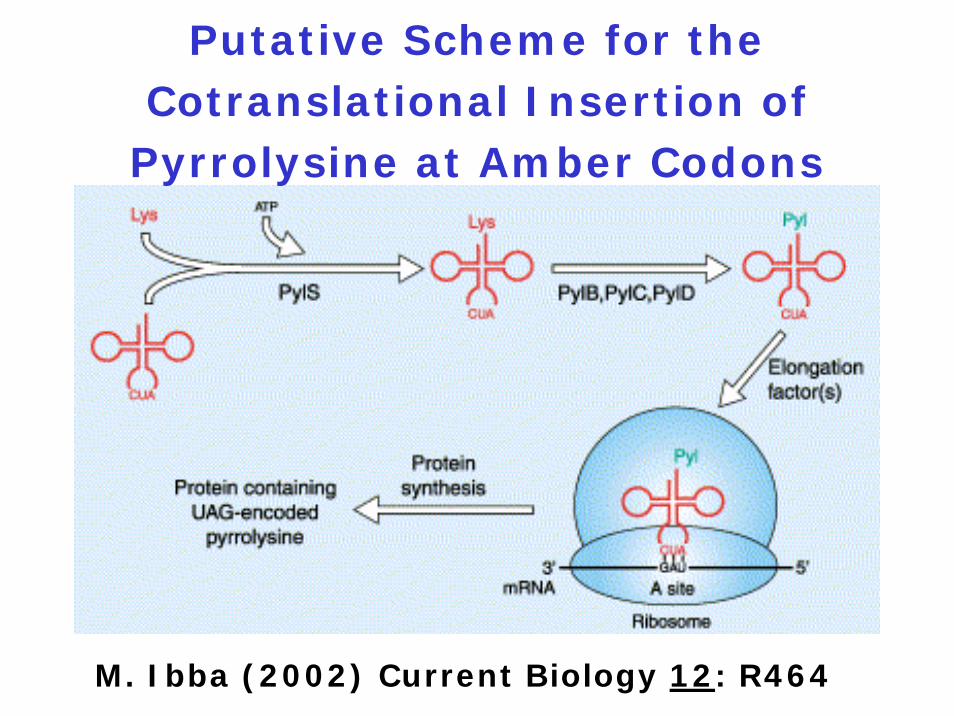
Putative Scheme for the Cotranslational Insertion of
Pyrrolysine at Amber Codons
M. Ibba (2002) Current Biology 12: R464

Distribution of Sel and Pyl
Deltaproteobacterium δ1:57 selenoprotein genes several Pyl-containing proteins

6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation
6.4.9 Collaborative behaviour in bacteria

1941: G. Beadle and E. Tatum
„ONE GENE - ONE ENZYME“ Hypothesis
George Beadle1903 - 1989
Edward Tatum1909 - 1975
Nobel Price 1958
Medicine

1. Overlapping genes
2. Ribosomal frameshifting
3. Bypassing
4. Gene within a gene
5. Polyprotein processing
Mechanisms Involved in Creating
Multiple Protein Products from a
Single mRNA

Overlapping Genes
There are two classes:
In-frame overlapping genes: permit the synthesis of more than one form of the protein from the same gene
Out-of-frame overlapping genes
ZI Johnson (2004) Genome Res. 14: 2268

In-Frame Overlapping Genes
Overlap may beLimited Complete
Reasons for overlapping genes:Economy strategy (genome compactness) Transcriptional or functional coupling
First in-frame gene pair: genes A and A* of phage ΦX174M. genitalium: 162 overlapping genes M. pneumoniae: 203 overlapping genes

Polyproteins
Definition:A precursor protein is proteolytically cleaved in two or more proteins

E. coli Penicillin G Acylase
Enzyme catalyses the conversion of penicillin G to phenylacetic acid and 6-aminopenicillanic acid (precursor for semisynthetic penicillins)
1 3 2
L Thöny-Meyer (1992) FEBS Lett. 307: 62
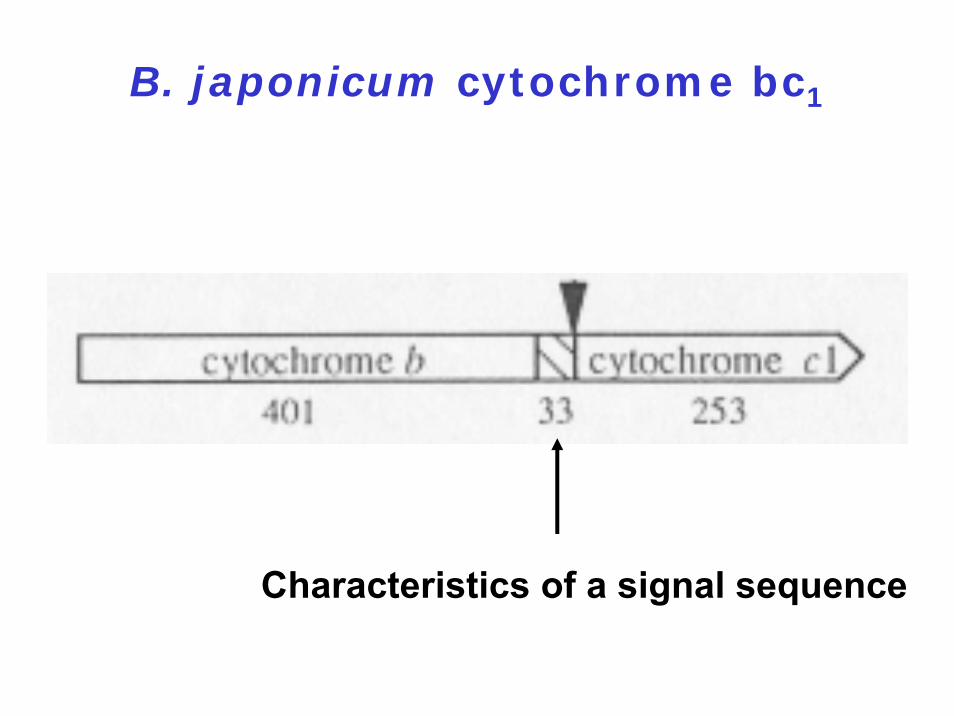
B. japonicum cytochrome bc1
Characteristics of a signal sequence

B. subtilis spore coat proteins
S Cutting (1991) J. Bacteriol. 173: 2915

B. polymyxa Amylases
N Uozumi (1989) J. Bacteriol. 171: 375
Three different β-amylases: 70, 65 and 42 kDa
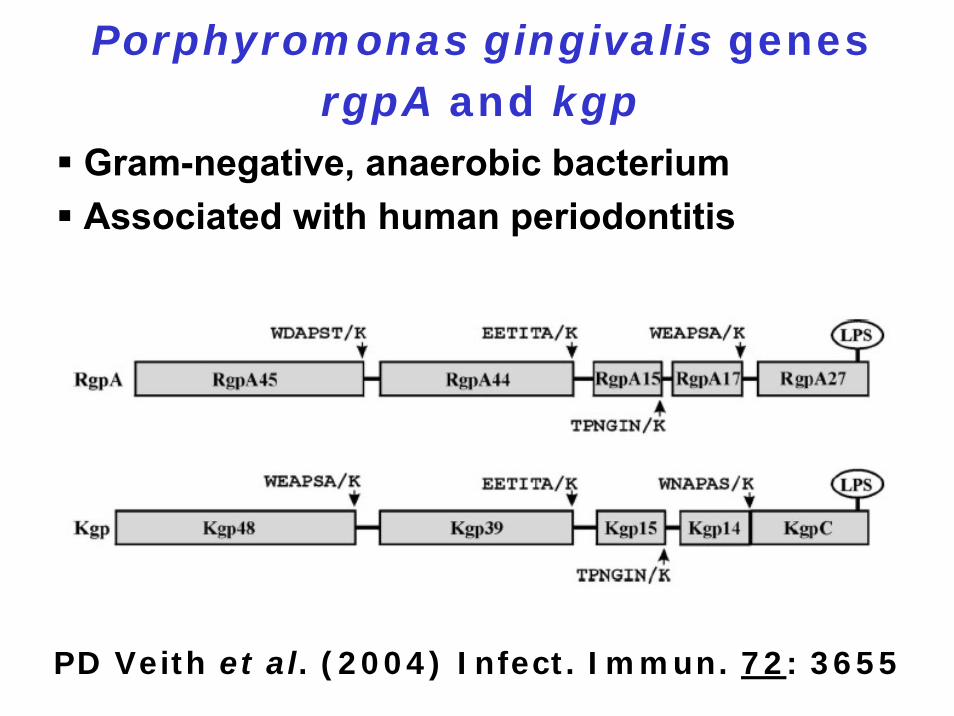
Porphyromonas gingivalis genes rgpA and kgp
Gram-negative, anaerobic bacteriumAssociated with human periodontitis
PD Veith et al. (2004) Infect. Immun. 72: 3655

What selective pressure forced evolution and maintenance of
polyproteins ?
Strict stoichiometry of the processed proteins
The different domains may fold interdependently (no chaperones needed)
Increase the local concentration of the different components (less dependent on diffusion)

There are two classes of circular proteins:
1. Those involved in host defense
2. Pilins
Circular Proteins
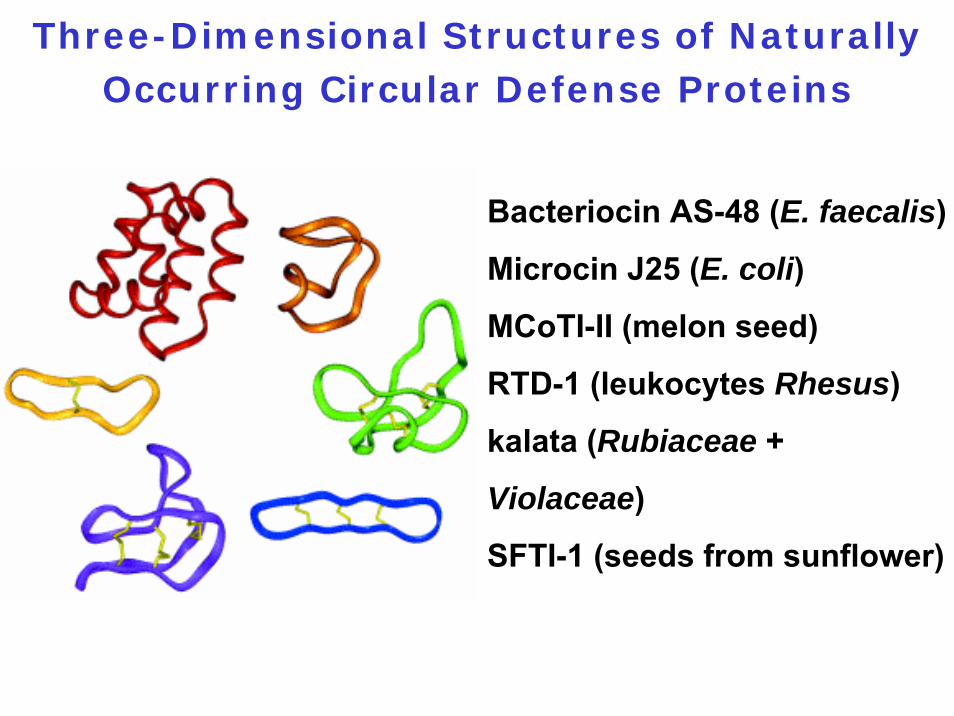
Three-Dimensional Structures of Naturally Occurring Circular Defense Proteins
Bacteriocin AS-48 (E. faecalis)
Microcin J25 (E. coli)
MCoTI-II (melon seed)
RTD-1 (leukocytes Rhesus)
kalata (Rubiaceae +
Violaceae)
SFTI-1 (seeds from sunflower)

Circular Defense Proteins
1. Range in size from 14 to 70 amino acids2. Involved in host defense3. Bacteriocin AS-48: Highly basic 70-amino acid
protein; forms pores in the cytoplasmic membrane of sensitive cells; thermal denaturation temperature = 93°C
4. They all appear to be derived from longer precursor proteins
5. Cycling enzymes or mechanisms of cycling unknown
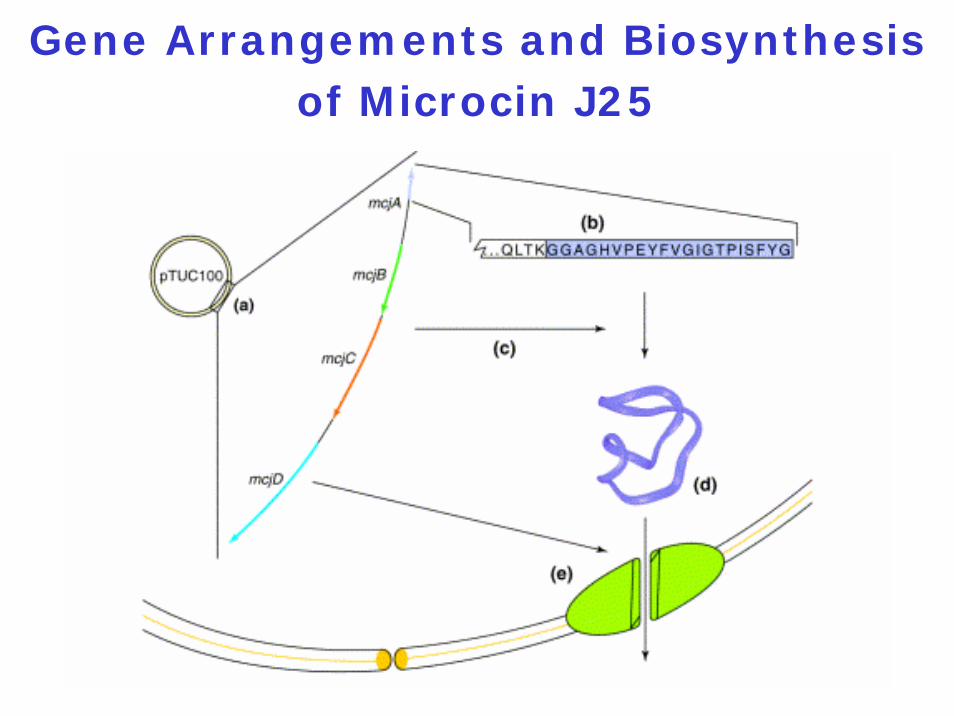
Gene Arrangements and Biosynthesis of Microcin J25

Moonlighting Proteins
Definition:
Proteins with more than one function
Examples:
PepA: - endopeptidase- DNA-binding protein
BirA: - biotin ligase- DNA-binding protein

6.4.1 The components
6.4.2 Initiation
6.4.3 Elongation
6.4.4 Termination
6.4.5 Alternative translation events
6.4.6 Translational control of gene expression
6.4.7 Additional amino acids
6.4.8 Posttranslational regulation
6.4.9 Collaborative behaviour in bacteria

Collaborative Behaviour in Bacteria
Definition:Groups of genetically identical organisms synchronize their patterns of gene expression to achieve specific goals that are unattainable for single cells acting on their own
Depends on cell-cell communication with secreted signaling molecules

Cannibalism and Fratricide
Definition:Some cells of a population produce a toxin which kills other cells of the population which do not produce the toxin (fratricide) and feed on them (cannibalism).

Two examples:
Sporulating Bacillus subtilis cells = cannibalism
Competent Streptococcus pneumoniae cells = fratricide

Cannibalism
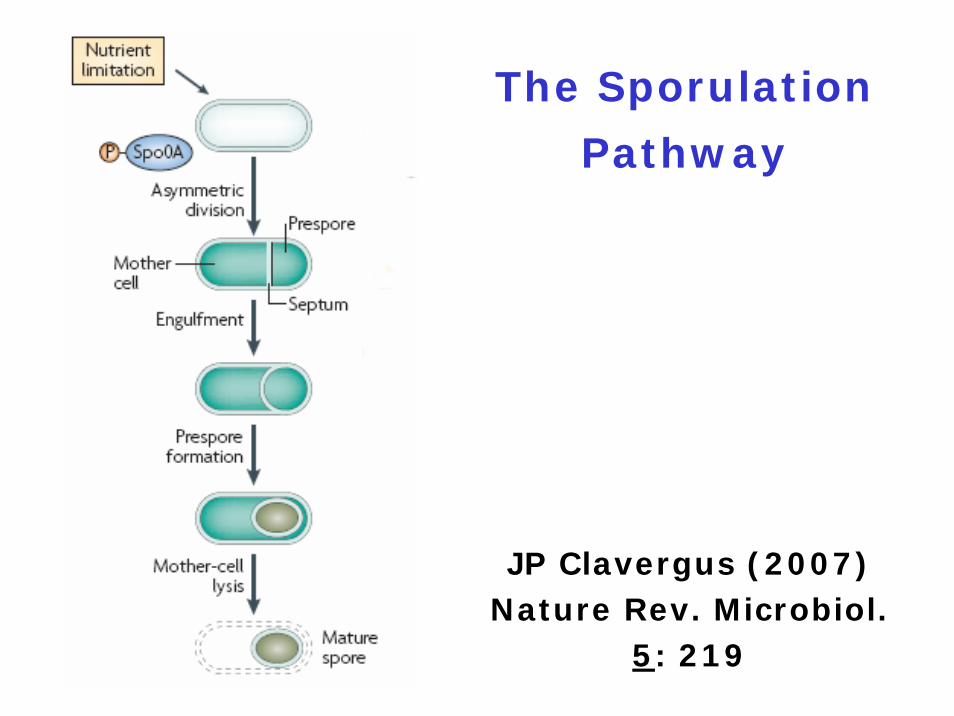
The Sporulation
Pathway
JP Clavergus (2007) Nature Rev. Microbiol.
5: 219

Sporulation is Induced by
Nutritional Stress
kinase Akinase Bkinase C ⇒ Spo0F ⇒ Spo0B ⇒ Spo0A~Pkinase D | |kinase E RapA PhrA Spo0E
RapB PhrCRapE PhrE
phosphorelay

Molecular Mechanism of Cannibalism
The skf (sporulation killing factor) and sdp(sporulating delay protein) operons are turned on by Spo0A~P
skfA-H: eight genes

The skf Operon
skfA-H: eight genesskfA: 55 amino-acid bacteriocin-like peptide = killing factor ?skfEF: ABC transporter = export of SkfAskfD: integral membrane protein = involved in maturation of SkfA ?skfB: involved in post-translationally modification of SkfA ?

The sdp Operon
sdpABCsdpC: codes for the toxin; derived from the 63 C-terminal amino acids of SdpC
Both SkfA and SdpC belong to the group of antimicrobial peptides

Spo0A
DNA-binding protein acting both as a repressor and as an activator
Present in an inactive (Spo0A) and in an active form (Spo0A~P)
60-70% of the cells produce a high amount of Spo0A~P = Spo0A-ON30-40% of the cells produce a low amountof Spo0A~P = Spo0A-OFF
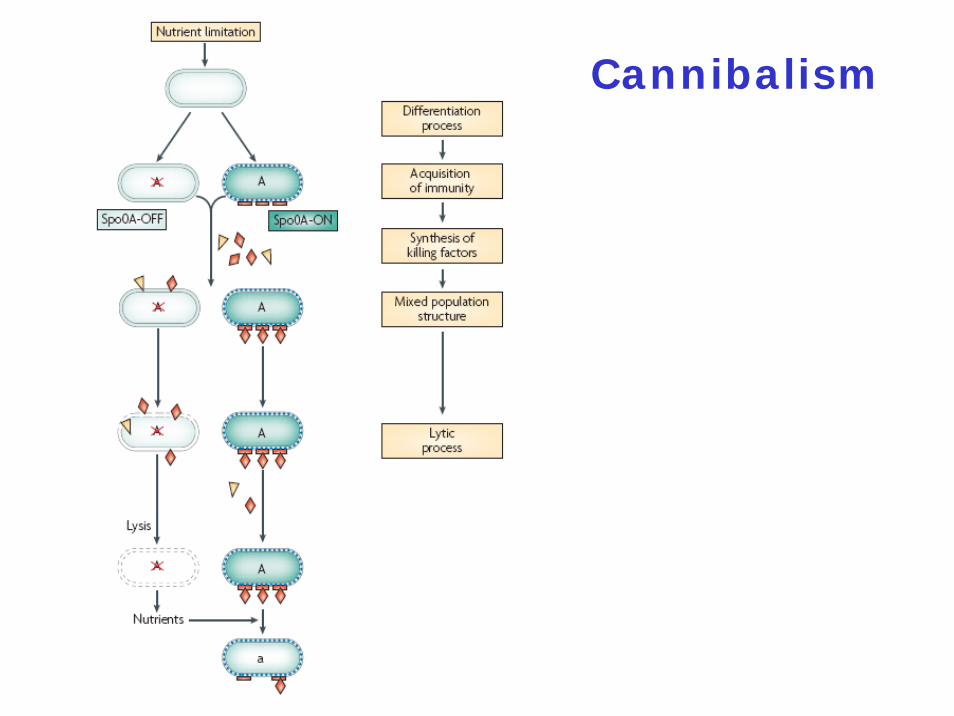
Cannibalism

Fratricide

Competence
JP Clavergus(2007) Nature
Rev. Microbiol. 5: 219

Induction of Competence
Competence develops in response to environmental signals such as- a change in external pH- antibiotic-induced stress:
Aminoglycoside antibiotics: Block ribosomefunction
Fluoroquinolone: Inhibits topoisomeraseMitomycin C: DNA damaging agent
Induction of competence = general stress response

Transformation of the Gram-
positive bacterium Streptococcus
pneumoniae
Induction of competence involves cell-cell communication mediated by the
competence stimulating peptide = CSP

Activation of Early com Genes
1. The secretion apparatus ComA/ComB isinvolved in secretion of the 17-residues CSP
2. If the cell concentration is high enough, CSPbinds to the ComD histidine kinase
3. Autophosphorylated ComD~P transfers thephosphate group to the response regulatorComE
4. ComE~P activates expression of ~20 early com genes among them comX

Activation of Late com Genes
1. comX codes for an alternative sigma factor2. ComX controls expression of ~60 late com
genes3. Four of these genes code for killing factors
- CbpD: murein hydrolyse- LytA: murein hydrolase - LytC: hydrolytic enzyme (not ComX-dependent)
- CibA, CibB: two-peptide bacteriocin
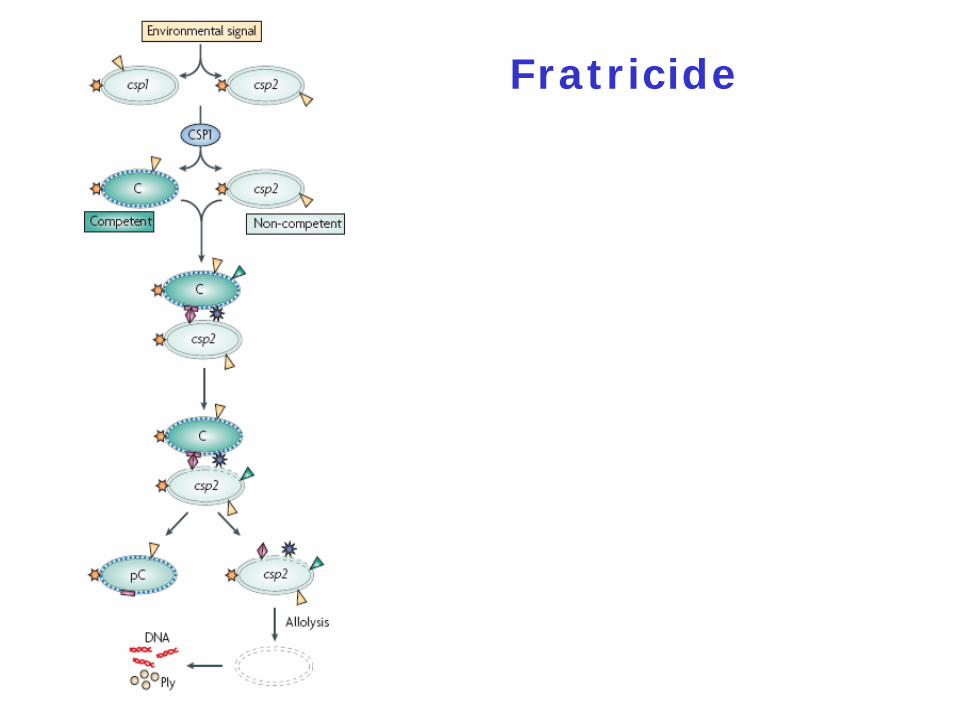
Fratricide

Tentative Model for Fratricide
Fratricide needs the three lytic enzymes and the two peptide bacteriocins
Fratricide is involved in cell lysis in trans = allolysis
Fratricide can still occur even if lytA and lytCare both inactivated provided they are intact in the targeted cells

How do cells prevent suicide ?
Two immunity proteins have been identifiedBoth are integral membrane proteinsCibC: 65 residues; two transmembrane segments; mechanism unknownComM: 206 residues

Who is killed ?
Competent cells kill non-competent cells
How could mixed pneumococcal populations arise ?
unknown

What is the biological function of
fratricide ?
Release of virulence factors and inflammatory
mediators
Access to DNA


















