
© 2011 Aabir Banerji ALL RIGHTS RESERVED
106
© 2011 Aabir Banerji ALL RIGHTS RESERVED
Transcript of © 2011 Aabir Banerji ALL RIGHTS RESERVED

© 2011
Aabir Banerji
ALL RIGHTS RESERVED

DYNAMICS AND ADAPTIVE SIGNIFICANCE OF THE INDUCIBLE
TROPHIC POLYMORPHISM OF TETRAHYMENA VORAX
By
AABIR BANERJI
A Dissertation Submitted to:
Graduate School – New Brunswick
Rutgers, The State University of New Jersey
in partial fulfillment of the requirements for the degree of:
Doctor of Philosophy
Graduate Program in Ecology & Evolution
Written under the direction of:
Dr. Peter J. Morin
and approved by:
_____________________________________
_____________________________________
_____________________________________
_____________________________________
_____________________________________
New Brunswick, New Jersey
October, 2011

ii
ABSTRACT OF THE DISSERTATION:
Dynamics and Adaptive Significance of the Inducible Trophic Polymorphism of
Tetrahymena vorax
By Aabir Banerji
Dissertation Director:
Dr. Peter J. Morin
This dissertation provides a four-part synopsis of the methodology, results, and
conclusions of an integrated set of experiments designed to explore the ecology of
inducible trophic polymorphisms (ITPs). These experiments were conducted using
isogenic populations of the polymorphic freshwater hymenostome ciliate, Tetrahymena
vorax Kidder, in combination with other protozoa, bacteria, algae, and micro-
invertebrates. Chapter 1 addresses the autecological significance of ITPs, revealing some
of their potential costs and benefits to individuals in the context of competition under
varying resource regimes. Chapter 2 demonstrates how ITPs can give rise to novel trait-
mediated indirect interactions that affect community structure and population dynamics.
Chapter 3 elucidates the extent to which the population dynamics of species exhibiting
ITPs are affected by the trophic complexity of their habitat. Chapter 4 presents a detailed
review of indirect offenses, a versatile adaptive strategy that ITPs can give rise to where
inducing agents are symbionts. The contents of these chapters provide unique insights
into the link between ITPs and related phenomena – such as cannibalism, intraguild
predation, omnivory, and expanded diet breadth – and open new avenues of inquiry
regarding the importance of the genetic and functional diversity of natural communities.

iii
ACKNOWLEDGEMENTS
First and foremost, I must thank Peter Morin, my academic advisor. I cannot
begin to describe the extent to which I am indebted to him for the countless ways in
which he has nurtured my personal growth and professional development since the day I
came to Rutgers. Rather than try to mold me into a re-creation of himself or persuade me
to become a mindless automaton to be exploited for his own agendas, Peter chose to
show respect for me both as his junior colleague and as an individual, which, I must add,
he does with all of his students, irrespective of where they are on the academic totem
pole. He did as much as he could to look out for me and was always willing to not only
hear me out, but also draw upon his vast, encyclopedic knowledge and years of
experience to help me achieve my goals. Time and again, I would come to him with
what I thought was an insurmountable obstacle, muttering profusely in my frustration
with it, only to have him take a quick look at it and systematically turn it into something
that I knew I could handle. Peter encouraged me when I needed it, straightened me out
when I needed it, left me to my own devices when I needed it, and did everything else
that was required to ensure that I would become a competent, self-respecting scientist
who was capable of finding his own way in the world. I could never have asked for a
better mentor, role-model, or friend than what Peter has been to me these past several
years.
Next, I must thank Marsha Morin, without whose intelligence, resourcefulness,
integrity, diligence, awareness, reliability, compassion, cat-herding ability, and sheer
tolerance for pain, my dissertation (along with our entire department) would have most
certainly burst into flames and/or collapsed into some kind of gravitational point

iv
singularity long ago. Her warmth and supportiveness have been a great source of
strength and joy for me from the very first day we met. To my gratitude for all of this, I
must also add my gratitude for the fact that she at some point forgave me for having
addressed her as “ma’am” for the first year or so of our acquaintance. Especially in light
of the fact that my parents had given both her and Peter express permission to beat me, I
consider this to have been most kind.
I thank my committee members (Tim Casey, Rebecca Jordan, Mike Pace, and
Cesar Rodriguez-Saona) for their always meaningful and prompt feedback on my various
drafts and progress reports, their willingness to work around their respective schedules to
accommodate my needs, their support of my research by way of resources and ideas, and
their having made every major milestone toward my degree fun and fulfilling for me.
I thank Martha Haviland, Doreen Glodowski, and Jana Curry (along with Jana’s
predecessor, Barbara Sena and the Busch MSLC’s administrative coordinator, Margie
Eickhorst) for having provided me with the opportunity and the means to take up
teaching, for having stood by me and come through for me whenever I needed help
solving problems and making things happen, for tolerating (and often even encouraging)
my various teaching-related shenanigans, and for being down-right awesome in general.
I thank the following people for helping me get started on the path to becoming a
full-fledged ecologist: Jim “Doc” Applegate, Barbara Goff, Colleen Hatfield, Bruce
Hamilton, Ted Stiles, Charles Harman, Fred Westcott, Ralph Caiazzo, Carol VanDalen,
Joyce Hartmann, and Kathy Parsell. Each of them in their own unique ways gave me
guidance, encouraged me to follow my heart, and enabled me to catch a glimpse of my
true potential. Their wisdom and their friendship will forever be a part of who I am.

v
I thank the following fellow participants in the 2005 Community Ecology course
(S211) at the Research School for Socio-Economic and Natural Sciences of the
Environment for the stimulating discussions and fun we had back then and the
stimulating discussions and fun we have had since: Anja Rubach, Karin Nilsson, Irene
van der Stap, Wade Ryberg, Tim Engelkes, Björn Christian Rall, Magnus Huss, Órla
McLaughlin, Euridice Leyequien, Geraldine Thiere, Sonia Kéfi, Martin Pedersen, Elly
Morrien, Jordie Netten, Maarten Eppinga, Roy van Grunsven, Mariska te Beest, Cleo
Gosling, Yuki Fujita, Ciska Veen, Marleen Riemens, Marleen Pierik, Tessa van der
Hammen, Charlotte Borvall, Johan Ahnström, Frans Kuenen, Ellen Weerman, and
Matthijs Schouten. Being together and working together with all of these fellow hard-
core community ecologists / citizens of the world will remain one of my fondest
memories in the years to come. Soon it will be our turn to do what our supervisors did
for us.
I thank my former lab mates (Lin Jiang, Christina Kaunzinger, Zac Long, Timon
McPhearson, Jen Price, Jenn Adams Krumins, Henry Stevens, Irene Karel, and Alex
Kulczycki) for showing me the ropes, helping me to imbibe the principle of K.I.S.S.
(Keep It Simple, Stupid!), and being a pleasure to hang out with.
I thank my current lab mates (Kevin Aagaard, Laura Chen, Jean Deo, Sean Hayes,
Ruchi Patel, Jack Siegrist, Maria Stanko, Holly Vuong, and Talia Young) for suffering
through my roughest drafts; putting up with my various quirks and ineptitudes within our
common, confined space; and sharing with me their ideas, enthusiasm, and friendship.
I thank the following faculty members and loose affiliates for profound, timely
advice and countless fascinating discussions (including everything from the need for

vi
applied ecology in modern medicine to the trouble with tribbles): Tamar Barkay, Mary
Cadenasso, John Dighton, David Drake, Siobain Duffy, David Ehrenfeld, Joan Ehrenfeld,
Nina Fefferman, Dina Fonseca, Jeremy Fox, Jason Grabosky, Ed Green, Steven Handel,
Brad Hillman, Claus Holzapfel, Henry John-Alder, Karl Kjer, Sharron Lawler, Julie
Lockwood, Bonnie McCay, Scott Meiners, Owen Petchey, Steward Pickett, Beth Ravit,
John Reinfelder, Peter Smouse, Lena Struwe, and Mike Sukhdeo.
While I owe every single one of my friends from Rutgers my gratitude and
acknowledgement for their empathy, encouragement, and collaboration, the following
deserve special recognition for having gone above and beyond the call of duty for me on
numerous occasions: Carrie Norin, Elena Tartaglia, Blake Mathys (and his lovely wife,
Dimitria), Wayne Rossiter (and his lovely wife, Melissa), Ai Wen (and her lovely
husband, Kenneth Elgersma), Dave Smith, Andrea Egizi, Ari Novy, Charlie Kontos,
Aspa Chatziefthimiou, Diana Johnson, Tricia Ramey-Balci, Kerri-Ann Norton, Rob
Hamilton IV, Kristen Ross, Em Stander, Jenni Momsen, Sharron Crane, Alison Seigel,
Alex Hernandez, Faye Benjamin, Wes Brooks, Brian Clough, Jeremy Feinberg, Zac
Freedman, Sona Mason, Christine Minerowicz, Mat Pompliano, and Stephanie
Baccarella.
Most of all, I thank my family. They have understood the formula.

vii
TABLE OF CONTENTS
Abstract of Dissertation …………………………….………………….… ii
Acknowledgements ……..……………………………………………..… iii
Table of Contents …………….………………………………………..… vii
Introduction ……………………………..……………………………..… 1
Chapter 1: Phenotypic Plasticity, Intraguild Predation, and Anti-
Cannibal Defenses in an Enigmatic Polymorphic Ciliate …… 5
Chapter 2: Prey-Induced Gape Expansion Mediates Apparent
Competition …………………………………………………. 32
Chapter 3: Dynamics between Protists and Crustacean Zooplankton: Do
Copepods Show Preference for the Predatory Morph of
Tetrahymena vorax? ………………………………………… 52
Chapter 4: On the Strategy of Mounting an Indirect Offense: What If My
Enemy Has Allies, Too? …………………………………….. 71
Conclusion ………………………………………………………….….…. 97
Curriculum Vitae …………….….….…………………………………..… 98

1
INTRODUCTION
Phenotypic plasticity is the ability of an organism to alter its morphology or
physiology in response to environmental cues. It is distinct from evolutionary adaptation
in that it occurs at the individual level, rather than at the population level, and generally
involves no change in the organism’s genetic material (DeWitt and Scheiner 2004).
Many examples of phenotypic plasticity are conspicuous and familiar, such as the color
changes of chameleons (Stuart-Fox et al. 2008) and the phototropism of plants (Correll
and Kiss 2002). Others, including the various internal responses that allow for immunity
(Frost 1999) and homeostasis (Woods 2009), are more subtle and easy to overlook. In
population studies, phenotypic plasticity is fascinating not only as a versatile adaptive
trait (Via et al. 1995), but also as a potential driver of diversification and speciation
(Adams and Huntingford 2004, Pfennig et al. 2010).
Phenotypic plasticity has been shown to influence the dynamics and stability of
ecological communities (Agrawal 2001, Garay-Narváez and Ramos-Jiliberto 2009). A
particularly intriguing way that it may do so is by generating inducible trophic
polymorphisms. Inducible trophic polymorphisms (hereafter abbreviated as ITPs) occur
when some individuals in a population alter their morphology or physiology to expand
the number of trophic levels on which they feed. Induction of alternative morphs occurs
in response to environmental factors, such as specific chemical cues released by
competitors and/or potential prey. ITPs are functionally complementary to inducible
defenses among prey (Tollrian and Harvell 1999). As is true for inducible defenses, ITPs
may initially seem like unusual adaptations that occur infrequently, but are in reality
widely distributed across taxa.

2
The consequences of ITPs for long-term population and community dynamics are
likely significant, but difficult to assess. For most long-lived organisms, the amount of
time and effort required to monitor populations through two or more generations is
substantial, and opportunities for genetic recombination can make it nearly impossible to
distinguish induced variation from constitutive variation. To approach the challenge of
addressing these consequences, I have conducted my research using protists in
experimental microcosms. This approach has been shown to be valuable in testing
ecological and evolutionary theory and unveiling the mechanisms that underlie general
patterns (Cadotte et al. 2005, Holyoak and Lawler 2005). Experimental microcosms
make for highly controlled and replicable systems. The short generation times of protists,
together with their large population sizes, facilitate the study of population dynamics
(Morin and Lawler 1996, Price and Morin 2004). In the microcosms I made use of in my
research, the focal species was the freshwater hymenostome ciliate, Tetrahymena vorax
Kidder.
The natural history of T. vorax makes this a fascinating and powerful model
system for the study of ITPs. T. vorax exhibits a conspicuous ITP. Rapid morphological
differentiation allows different individuals in a single isogenic population to feed lower
(on bacteria) or higher (on ciliates that consume bacteria) in the same food chain (Kidder
et al. 1940, Ryals et al. 2002). The different morphs of T. vorax are easily distinguished
and counted via microscopy. Populations in the early logarithmic stage of growth
typically consist entirely of small (~ 45-75 m long), pear-shaped morphs, called
“microstomes,” which feed on bacteria. In response to known chemical cues that are
released by competing bacterivores, microstomes may transform into larger (~ 250 m

3
long), oval-shaped morphs, called “macrostomes,” which feed on other ciliates (Smith-
Somerville et al. 2000, Ryals et al. 2002). Once a microstome has transformed into a
macrostome, it can give rise to additional macrostomes, or it can revert into a microstome
at the time of cell division (~ every 6 hours).
My use of T. vorax for my experimental investigations has enabled me to explore
the autecological significance of ITPs, evaluate the potential of ITPs to generate complex
trait-mediated indirect effects, and elucidate the extent to which population and
community dynamics associated with ITPs are affected by trophic complexity. In so
doing, I have: (1) uncovered potential costs and benefits of different inducible
morphologies in the context of competition, (2) demonstrated how ITPs can potentially
mediate interactions such as “apparent competition” (Holt 1977), and (3) identified
possible roles of major groups of aquatic consumers in limiting the distribution of
polymorphic ciliates in nature. A portion of my work – Chapter 1 of this dissertation –
has already been published in the journal Functional Ecology (Banerji and Morin 2009).
References
Adams, C. E., and F. A. Huntingford. 2004. Incipient speciation driven by phenotypic
plasticity? Evidence from sympatric populations of Arctic charr. Biological
Journal of the Linnean Society 81(4): 611-618.
Agrawal, A. A. 2001. Phenotypic plasticity in the interaction and evolution of species.
Science 294: 321-326.
Banerji, A., and P. J. Morin. 2009. Phenotypic plasticity, intraguild predation and anti-
cannibal defences in an enigmatic polymorphic ciliate. Functional Ecology 23:
427-434.
Cadotte, M. W., J. A. Drake, and T. Fukami. 2005. Constructing nature: laboratory
models as necessary tools for investigating complex ecological communities.
Advances in Ecological Research 37: 333-353.
Correll, M. J., and J. Z. Kiss. 2002. Interactions between gravitropism and phototropism
in plants. Journal of Plant Growth Regulation 21(2): 89-101.
DeWitt, T. J., and S. M. Scheiner. 2004. Phenotypic variation from single genotypes: a
primer. Pages 1-9 in T. J. DeWitt and S. M. Scheiner, editors. Phenotypic

4
Plasticity: Functional and Conceptual Approaches. Oxford University Press,
Oxford.
Frost, S. D. W. 1999. The immune system as an inducible defense. Pages 104-126 in R.
Tollrian and C. D. Harvell, editors. The Ecology and Evolution of Inducible
Defenses. Princeton University Press, Princeton.
Garay-Narváez, L., and R. Ramos-Jiliberto. 2009. Induced defenses within food webs:
the role of community trade-offs, delayed responses, and defense specificity.
Ecological Complexity 6(3): 383-391.
Holyoak, M., and S. P. Lawler. 2005. The contribution of laboratory experiments on
protists to understand population and metapopulation dynamics. Advances in
Ecological Research 37: 245-271.
Kidder, G. W., D. M. Lilly, and S. L. Claff. 1940. Growth studies on ciliates. IV. The
influence of food on the structure and growth of Glaucoma vorax sp. nov.
Biological Bulletin 78: 9-23.
Morin, P. J. and S. P. Lawler. 1996. Effects of food chain length and omnivory on
population dynamics in experimental microcosms. Pages 218-230 in G. Polis and
K. Winemiller, editors. Food Webs: Integration of Patterns & Dynamics.
Chapman and Hall.
Pfennig, D. W., M. A. Wund, E. C. Snell-Rood, T. Cruickshank, C. D. Schlichting, and
A. P. Moczek. 2010. Phenotypic plasticity’s impacts on diversification and
speciation. Trends in Ecology and Evolution 25: 459-467.
Price, J. E., and P. J. Morin. 2004. Colonization history determines alternate community
states in a food web of intraguild predators. Ecology 85:1017-1028.
Ryals, P. E., H. E. Smith-Somerville, and H. E. Buhse Jr. 2002. Phenotype switching in
polymorphic Tetrahymena: a single-cell Jekyll and Hyde. International Review of
Cytology 212: 209-238.
Smith-Somerville, H. E., J. K. Hardman, R. Timkovich, W. J. Ray, K. E. Rose, P. E.
Ryals, S. H. Gibbons, and H. E. Buhse Jr. 2000. A complex of iron and nucleic
acid catabolites is a signal that triggers differentiation in a freshwater protozoan.
Proceedings of the National Academy of Sciences 97: 7325-7330.
Stuart-Fox, D., A. Moussalli, and M. J. Whiting. 2008. Predator-specific camouflage in
chameleons. Biology Letters 4(4): 326-329.
Tollrian, R., and C. D. Harvell, editors. 1999. The ecology and evolution of inducible
defenses. Princeton Univerity Press, Princeton.
Via, S., R. Gomulkiewicz, G. De Jong, S. M. Scheiner, C. D. Schlichting, and P. H. Van
Tienderen. 1995. Adaptive phenotypic plasticity: consensus and controversy.
Trends in Ecology and Evolution 10(5): 212-217.
Woods, H. A. 2009. Evolution of homeostatic physiological systems. Pages 591-610 in
D. Whitman and T. N. Ananthakrishnan, editors. Phenotypic Plasticity of Insects.
Science Publishers, Plymouth.

5
CHAPTER 1
Phenotypic Plasticity, Intraguild Predation, and Anti-Cannibal Defenses in
an Enigmatic Polymorphic Ciliate
Summary
1. Inducible trophic polymorphisms are greatly underappreciated forms of
phenotypic plasticity that allow organisms to respond dynamically to
environmental variation by enabling them to change the trophic level upon
which they feed. Although inducible trophic polymorphisms occur in a
diverse array of organisms, their costs and benefits and their consequences for
long-term population and community dynamics are poorly understood.
2. I studied the inducible trophic polymorphism of the freshwater hymenostome
ciliate Tetrahymena vorax, whose isogenic populations can contain three
distinct morphs: pyriform, bacterivorous microstomes; larger, carnivorous
macrostomes; and elongate, “tailed” microstomes. I tested whether: (1) the
tailed microstome constitutes an inducible defense against macrostomes and
(2) the transformation of microstomes into macrostomes is size-dependent. I
also recorded the dynamics of the three morphs in the presence and absence of
an intraguild prey (Colpidium) across a gradient of growth medium
concentrations to infer potential tradeoffs in the success of different morphs at
different productivity levels.
3. Macrostomes do not discriminate between pyriform microstomes and readily-
consumed heterospecific prey (Colpidium) when provided only one of these

6
alternative prey items. Tailed microstomes display greatly reduced
susceptibility to consumption by macrostomes as compared to undefended,
pyriform microstomes. Morph dynamics are consistent with the hypothesis
that tailed microstomes serve as an inducible defense against cannibalism;
tailed microstomes and macrostomes appear simultaneously, in both the
presence and absence of Colpidium. At low productivity, T. vorax achieves
higher rates of growth when able to feed on Colpidium than when feeding on
bacteria alone. At higher productivity, this pattern is reversed, with growth
rates maximized in the absence of Colpidium.
4. The reduced consumption rate of tailed microstomes by cannibalistic
macrostomes, together with the simultaneous induction of tailed microstomes
and macrostomes, suggests that both morphs comprise a coordinated adaptive
response to the presence of intraguild prey.
Key-words: Induced offenses, Tetrahymena vorax, inducible trophic polymorphism,
omnivory

7
Introduction
Phenotypic plasticity can potentially have profound effects on the population
dynamics of interacting species. For instance, numerous examples of inducible defenses
have been shown to completely alter the predicted outcome of predator-prey interactions
(Tollrian and Harvell 1999). These inducible defenses can involve an array of
behavioral, physiological, metabolic, or life-history traits and can be direct or indirect
(Porter and Porter 1979; Abrahams 1993; Tollrian and Harvell 1999; Altwegg 2002).
While inducible defenses are the subject of extensive study (Tollrian and Harvell 1999),
their logical counterpart, inducible offenses (Padilla 2001), remain little-studied and
greatly underappreciated. Inducible offenses often involve inducible trophic
polymorphisms. In these situations, certain organisms alter the trophic level upon which
they feed via changes in morphology, physiology, or behavior, modifying the number and
types of interactions in their food web.
Inducible trophic polymorphisms take many forms. In some organisms,
reversible, morphological transformations provide flexible responses to environmental
variation within a single generation. In others, transformations constitute irreversible,
developmental trajectories that commit individuals to particular trophic roles. Some
inducible trophic polymorphisms produce changes in diet without changing the mode of
acquisition, such as where a predator turns into a higher-level predator or a cannibal
(Gilbert 1973, Wainwright et al. 1991). Others represent both a change in diet and in the
mode of acquisition, such as where an herbivore turns into a carnivore (Pfennig 1992,
Afik and Karasov 1995) or where an autotroph becomes a parasite (Baird and Riopel
1984).

8
Inducible trophic polymorphisms occur within a wide range of taxa that includes
birds (Afik and Karasov 1995), amphibians (Pfennig 1992), fish (Wainwright et al.
1991), gastropods (Padilla 2001), plants (Baird and Riopel 1984), fungi (Barron 1977,
Barron 1992, Kerry and Jaffee 1997), rotifers (Gilbert 1973), protists (Washburn 1988),
and bacteria (Berleman and Kirby 2007). The diversity and prevalence of inducible
trophic polymorphisms provide a compelling motivation to investigate what effects
inducible trophic polymorphisms have on long-term population dynamics. The study of
inducible trophic polymorphisms can also provide insights into related phenomena, such
as cannibalism (Fox 1975, Polis 1981), intraguild predation (Polis et al. 1989, Holt and
Polis 1997), omnivory (Polis 1991, Morin 1999, Diehl and Feissel 2000), variation in diet
breadth (Wainwright et al. 1991), and phenotypic plasticity in general (DeWitt and
Scheiner 2004). Inducible trophic polymorphisms may even shed light on certain
evolutionary processes, due to the fact that they potentially allow reciprocal induced
offenses and defenses to occur rapidly over short ecological time frames, changes that are
comparable to the shifting phenotypes seen over much longer time frames as a
consequence of reciprocal genetic changes in co-evolving species (Agrawal 2001).
The inducible trophic polymorphism of the freshwater protozoan Tetrahymena
vorax Kidder (Ciliatea: Hymenostomatida) offers five main advantages as a model
system for the experimental study of inducible offenses: (1) its small size (60-250 µm)
and ease of culture makes experimental studies relatively convenient; (2) its short
generation time (~ 6 hours) makes possible the collection of long-term population
dynamics data in a matter of days or weeks; (3) its clonal mode of reproduction (the cells
are amicronucleate and unable to conjugate) ensures that any short-term phenotypic

9
variation does not reflect underlying genetic variation; (4) its discrete polymorphism
facilitates distinguishing between morphs and quantifying the transformation; and (5) the
proximal cues triggering transformations are known and are easily manipulated (Ryals et
al. 2002).
Isogenic populations of T. vorax contain three distinct morphs: microstomes,
macrostomes, and tailed microstomes. Microstomes (~ 60 µm in length) are pyriform,
with a small cytostome that restricts their diet to bacteria. Macrostomes (~ 200 µm) are
ovoid, with a larger cytostome that allows them to prey on other smaller ciliates. Tailed
microstomes (~ 90 µm) also consume bacteria, but unlike pyriform microstomes, are
narrow, elongate cells with the posterior drawn out into a stiff, tail-like structure (Kidder
et al. 1941, Williams 1961). New T. vorax populations (at low density and high resource
levels) initially consist entirely of pyriform microstomes. As population density
increases, some of the pyriform microstomes transform into macrostomes and tailed
microstomes. Each of the three morphs can transform into any of the others (Fig. 1).
Microstomes transform into macrostomes when exposed to known cues, the
nucleic acid catabolites uracil and hypoxanthine in the presence of ferrous iron (Smith-
Somerville et al. 2000). The cues are released by other ciliates that are competitors for
bacteria, such as Colpidium striatum Stokes (Ciliatea: Hymenostomatida) and also by
conspecifics. In the absence of predators, Colpidium can reduce bacterial densities to a
greater extent than T. vorax (Kaunzinger and Morin 1998, Price and Morin 2004), which
suggests that microstomes might be competitively excluded by Colpidium. The ability of
T. vorax to mount an inducible offense against its competitors may provide a potentially
significant fitness advantage (Thingstad et al. 1996). However, as with any inducible

10
response, there may be constraints and / or costs that prevent this response from being
constitutive (Relyea 2002, DeWitt and Scheiner 2004). If, for example, the induced
macrostomes cannot discriminate between conspecifics and heterospecifics, the potential
advantage of being able to eat interspecific competitors could be offset by the costs of
cannibalism (Polis 1981).
Cannibalism can be adaptive if it allows a lineage to survive and reproduce during
severe resource limitation (Crump 1983, Van den Bosch et al. 1988, Hoffman and
Pfennig 1999). However, the observed rarity of cannibalism under less severe conditions
and among taxa in general suggests that it exacts significant fitness costs (Pfennig and
Collins 1993, Pfennig and Frankino 1997, Kusch 1999, Pfennig 1999, Pfennig 2000,
Michimae and Wakahara 2002). Therefore, where inducible trophic polymorphisms
create the potential for cannibalism, there is likely to be strong selection for mechanisms
that minimize cannibalism on close relatives, or that divert attacks to non-kin (Gilbert
1973, Pfennig and Collins 1993, Pfennig and Frankino 1997, Kusch 1999, Pfennig 1999).
Grønlien et al. (2002) have demonstrated that macrostomes capture heterospecific
prey in preference to conspecific microstomes and will engulf available microstomes
only after exhausting the supply of heterospecific prey. These results suggest that T.
vorax may indeed use kairomones or other cues to discriminate between kin and non-kin,
but they also indicate that this ability does not actually prevent macrostomes from
ultimately consuming smaller conspecifics. Nevertheless, T. vorax may still minimize
the costs of cannibalism through reciprocal, coordinated, morphological changes among
macrostomes and microstomes (Agrawal 2001, Kopp and Tollrian 2003a,b). My
preliminary observations indicated that tailed microstomes may be less susceptible to

11
cannibalism by macrostomes because elongate tailed microstomes cannot be completely
engulfed. If so, tailed microstomes may constitute an inducible defense against
cannibalism that would allow for a highly adaptive mixed foraging strategy, assuming
they are consistently present when macrostomes are present.
I speculated that the polymorphism of T. vorax could be the result of size-
dependent differentiation. Microstomes vary somewhat in size because of recent feeding
history, position in the cell cycle, and chance events. The successful transformation from
a microstome into a macrostome could depend on cell size at the time of induction, where
larger microstomes transform into macrostomes upon induction, while smaller ones
transform into tailed microstomes. Presumably, the proportion of larger microstomes in
the population is determined by the overall abundance of energy and nutrients (in the
form of bacteria) within the system. If macrostome production is energetically costly, or
depends on higher levels of productivity, then decreased bacterial abundances should lead
to a decreased frequency of macrostomes within the population. If resource limitation is
not a significant constraint on macrostome production, however, intraguild predation
theory predicts that T. vorax should switch from feeding on bacteria, the shared resource,
when bacteria are abundant, to feeding on Colpidium, the intraguild prey, when
Colpidium is abundant relative to bacteria. Increasing bacterial abundance by increasing
productivity would, in this case, reduce the relative frequency of macrostomes in the T.
vorax population and decrease intraguild predation of Colpidium by T. vorax (Křivan
2000, van Baalen et al. 2001).
This paper describes a series of studies designed to elucidate the costs and
benefits associated with each morph of T. vorax. I explore three questions arising from

12
the inducible trophic polymorphism. (1) Do tailed microstomes serve as an inducible
defense against cannibalism by conspecific macrostomes? (2) Is size-dependent
differentiation responsible for the mix of macrostomes and microstomes seen in isogenic
T. vorax populations? (3) Is the proportion of macrostomes in T. vorax populations more
indicative of the energetic and physiological requirements of transformation or of the
relative advantage to being a predator?
Methods
I obtained T. vorax from the American Type Culture Collection (strain 30421).
Isogenic populations of T. vorax were cultured in 240-ml glass jars containing 100 ml of
complex organic medium. Medium consisted of 0.37 g of Carolina Biological Supply
protozoan pellets (Carolina Biological Supply Company, Burlington, NC 27215, USA)
per L of filtered well water. Autoclaved medium was inoculated after cooling with three
species of bacteria (prey for the ciliates – Serratia marcescens, Bacillus subtilis, and
Bacillus cereus). I autoclaved all culture vessels before use. Screw caps on the jars
permitted some air circulation, while restricting evaporation and limiting contamination.
The experimental units used in this study contained detritus-based food-chains,
consisting of organic matter, bacteria, and ciliates. The organic matter in the growth
medium provided a conveniently manipulated energy source. Differences in the
concentration of the medium create differences in bacterial densities, which simulate
changes in the productivity of the system (Morin 1999).
Prey Selection by Macrostomes
To determine the potential effectiveness of the tailed microstome as an inducible
defense against cannibalism, I evaluated rates of prey consumption by macrostomes

13
feeding on different densities of pyriform microstomes, tailed microstomes, and
Colpidium. Feeding took place in 15-mm Petri dishes in 0.2 ml of medium, to confine all
cells (macrostomes and their prey) within the field of view of the microscope.
Each trial initially contained 30 macrostomes exposed to one of nine
combinations of prey type (pyriform microstomes, tailed microstomes, Colpidium) and
prey density (low ~ 201 cells / mL, medium ~ 403 cells / mL, and high ~ 599 cells /
mL). Prey densities varied to ensure that any differences found among the capture rates
for different prey items would be due to prey morphology, rather than to density-
dependent encounter rates. The response was the number of successful captures observed
within a 15-minute interval. I removed macrostomes that engulfed prey items using a
micropipette, to keep track of captures by macrostomes. Individual macrostomes were,
therefore, limited to completing a single capture per the 15-minute observation period.
There were three replicates for each combination of prey type and prey density.
I compared the number of prey captured per unit time with an analysis of
covariance (ANCOVA), focusing on differences among the three prey types (Colpidium,
microstomes, and tailed microstomes) using prey density as a covariate. Differences in
the slope of the prey-captured-vs.-prey-density relation among prey types would indicate
differences in susceptibility to macrostomes.
Phenology of the Tailed Microstome
I grew isogenic populations of T. vorax with and without Colpidium across a
gradient of 5 different nutrient concentrations to explore whether the phenology of tailed
microstomes is consistent with the possibility of their serving as an inducible defense
against cannibalism by macrostomes. The nutrient concentrations used were 50% (0.19

14
g/L), 75% (0.28 g/L), 100% (standard, 0.37 g/L), 125% (0.46 g/L), and 150% (0.56 g/L).
I recorded the population density for each morph and for Colpidium on 6 days (days 1, 2,
3, 4, 5, and 7) over a 7 day interval, sampling approximately 0.3 ml from each jar without
replacement. I first combined these observations of the densities of tailed microstomes
and macrostomes in a single correlation analysis to evaluate whether the changing
population density (cells/ml) of tailed microstomes was consistently correlated with the
changing population density of macrostomes over time. One microcosm became
contaminated with flagellates on Day 4 of the experiment. The data from this replicate
was subsequently discarded for Days 4, 5, and 7, yielding a total of 177 data points for
the analysis. This analysis assumes that population dynamics are sufficiently rapid that
observations made on different days in the same microcosm are effectively independent.
Given that T. vorax has a generation time of about 6 hours, this is a reasonable
assumption. A more conservative analysis evaluated the correlation between densities of
tailed microstomes and macrostomes separately for each of the sampling days (30
replicates on days 1, 2, 3; 29 replicates on days 4, 5, 7), treating each microcosm as an
independent observation on a given day. A positive correlation between the densities of
tailed microstomes and macrostomes would be consistent with the hypothesis that tailed
microstomes are more abundant when macrostomes are present, though it would not
indicate whether they arise in response to cues released by macrostomes or as a size-
dependent response to a common cue released by prey. Inclusion of populations grown
in the absence of Colpidium in this experiment ensured that the presence or absence of
Colpidium had no bearing on whether or not tailed microstomes are more abundant when
macrostomes are more abundant.

15
Effect of Microstome Cell Size on Transformation
Effects of cell size at the time of induction on macrostome formation were
determined by sorting microstomes into two size classes, small (< 60 µm) and large (≥ 60
µm), and culturing them within15-mm Petri dishes containing 2 ml of medium (standard
concentration) with or without Colpidium. I then monitored microstomes every 12 hr,
over a 48-hr period, and recorded the number of each morph within experimental groups.
Each experimental group consisted initially of 5 pyriform microstomal cells, and there
were 3 replicates for each group. I first compared differences in macrostome frequency
using a pooled two-way analysis of variance (ANOVA), after confirming that the effects
of time were not significant. I then conducted a more conservative analysis comparing
differences in macrostome frequency for each sampling time individually.
Population Dynamics in the Presence and Absence of an Interspecific Competitor
across a Productivity Gradient
Using the data collected from the phenology experiment, I compared the
individual morph population densities of each treatment to determine the effect of
productivity on the frequency of macrostome formation. In addition, I measured the
population growth rates of T. vorax as a whole, calculated as the natural log of the mean
number of cells per ml over time, with and without Colpidium across the productivity
gradient, to measure fitness.
Results
Prey Selection by Macrostomes
Capture rates by macrostomes were influenced by both prey type (ANCOVA,
F2,21 = 6.84, p = 0.0052) and prey density (ANCOVA, F1,21 = 106.44, p < 0.0001). Below

16
approximately 200 prey individuals / ml, prey capture rate by macrostomes was near 0,
irrespective of prey type (Fig. 2). There was a significant interaction between prey type
and prey density (ANCOVA, F2,21 = 23.61, p < 0.0001), which indicates that the slope of
the relation between prey density and the number of prey captured differed among prey
types. Macrostomes captured Colpidium and pyriform microstomes at approximately the
same rate, while few tailed microstomes were captured even at the highest prey density.
Phenology of the Tailed Microstome
The correlation analysis for which I had combined population densities across
time revealed that the population density of tailed microstomes was significantly
correlated with that of macrostomes (r = 0.687, p < 0.0001, n = 177). The more
conservative analysis yielded a significant positive correlation between the densities on
Day 2 (r = 0.770, p < 0.0001, n = 30), Day 3 (r = 0.764, p < 0.0001, n = 30), Day 4 (r =
0.739, p < 0.0001, n = 29), and Day 7 (r = 0.502, p = 0.0055, n = 29). On Day 1, as one
might expect, given that the cells would not have had time to differentiate, the correlation
between the densities tailed microstomes and macrostomes was not significant (r = 0.054,
p = 0.7763, n = 30). The correlation was also not significant on Day 5 (r = 0.251, p =
0.189, n = 29). Since, even in the conservative analysis, the correlation is positive on
four of the six days, the pattern appears robust. At each simulated productivity, in both
the presence and absence of Colpidium, macrostomes and tailed microstomes appeared
simultaneously. At standard productivity, with Colpidium present, pyriform microstomes
disappeared by the second day of population growth and did not reappear until after
Colpidium disappeared (Fig. 3).
Effect of Microstome Cell Size on Transformation

17
ANOVA revealed a significant interaction between microstome cell size and
Colpidium presence / absence on the induction of macrostomes (F1,56 = 29.57, p <
0.0001). This interaction was still observed when the analysis was conducted on each
sampling time separately (F1,8 = 15.21, p = 0.0045 for Hour 12; F1,8 = 147.05, p < 0.0001
for Hour 24; F1,8 = 201.79, p < 0.0001 for Hour 36; and F1,8 = 387.68, p < 0.0001 for
Hour 48). Microstomes less than 60 m long did not transform into macrostomes in the
presence of Colpidium. Although some pyriform cells transformed into tailed
microstomes, the majority remained microstomes. The majority of microstomes greater
than 60m transformed into macrostomes when induced by the presence of Colpidium
(Fig. 4).
Effects of Productivity and Intraguild Prey on Population Dynamics
Although the population density of macrostomes was significantly correlated with
both productivity (r = 0.296, p < .0001, n = 177) and the total population density of T.
vorax (r = 0.795, p < .0001, n = 177), the proportion of macrostomes within the T. vorax
population was not significantly correlated with these variables (r = 0.314, p = 0.0748, n
= 177 for productivity and r = 0.048, p = 0.5229, n = 177 for total population density).
Pyriform microstomes, which, at low productivity and in the presence of Colpidium
achieved the lowest densities of all the morphs, increased and achieved a higher density
than macrostomes at high productivity, in the absence of Colpidium. Tailed microstomes
achieved the highest densities of all the morphs in each treatment (Fig. 5). The overall
population growth rate of T. vorax was significantly influenced by productivity and the
presence or absence of Colpidium (ANCOVA, F4,28 = 3.40, p = 0.0295). At higher levels

18
of productivity, Colpidium decreased the growth rate of T. vorax. At low productivity,
the pattern was reversed, with Colpidium increasing T. vorax growth rates (Fig. 6).
Discussion
The results of my investigation strongly support the hypothesis that tailed
microstomes constitute a defense by the smaller size fraction of microstomes to potential
cannibalism by macrostomes. I recognize that I cannot separate whether the appearance
of the tailed microstome is directly induced by the appearance of macrostomes, or
whether tailed microstomes and macrostomes constitute different size-dependent
developmental responses to the same cue released by high densities of prey or by
conspecifics. Tailed microstomes clearly exhibit greatly reduced susceptibility to
consumption by macrostomes, relative to pyriform microstomes. Macrostomes do not
appear to discriminate between consuming conspecific and heterospecfic cells, as shown
by the observation that the capture rates of pyriform microstomes and Colpidium by
macrostomes did not differ. Furthermore, tailed microstomes appeared simultaneously
with macrostomes, even in the absence of Colpidium. The very low consumption rates of
tailed microstomes by macrostomes, together with the phenology of tailed microstome
morphs, enables the tailed microstome to serve as a coordinated, induced defense against
cannibalism when macrostomes appear in the population. Synchronous inductions of
offensive and defensive phenotypes such as this are unlikely to have evolved under
conditions favoring cannibalism, as such a defense would negate any selective advantage
of cannibalism. It is instead conceivable that the simultaneous induction may be part of a
mixed strategy to reduce intraspecific competition and to capitalize on multiple types of
available resources (Fisher 1958, Ehlinger 1990, Dudgeon and Buss 1996). In the case of

19
T. vorax, as the macrostomes feed on available ciliates, the tailed microstomes continue
to graze on bacteria, while remaining immune to the predation pressure that their
interspecific competitors face from macrostomes.
Differences in cell size at the time of induction provide one explanation for how
the polymorphism allowing for this potential mixed strategy arises. The significant effect
of cell size on the proportion of macrostomes induced within T. vorax populations
suggests that a size threshold exists for the successful transformation from microstome to
macrostome. Given that well-fed protozoa are larger than starved individuals (Kidder et
al. 1941), high productivities, where bacteria are abundant, should yield larger
proportions of macrostomes than low productivities, where bacteria are scarce. However,
increasing nutrient concentration did not significantly increase the proportion of
macrostomes, but instead merely increased the overall population density of T. vorax.
The fact that cell division was favored over transformation in a situation where
Colpidium was present at high density, but was neither as available nor as abundant as
bacteria, indicates that macrostome induction is not as much determined by the energetic
costs of transformation as it is by the relative advantage to being a predator.
The dominance of tailed microstomes over the other morphs at both high and low
productivity and in both the presence and absence of Colpidium may indicate that tailed
microstomes have a greater conversion efficiency of prey than either macrostomes or
pyriform microstomes. This, in combination with the resistance of the tailed microstome
to predation by cannibalistic macrostomes (and perhaps other gape-limited predators),
begs the question of why T. vorax populations contain pyriform microstomes at all. The

20
fact that the tailed microstome does not appear to be constitutive implies the existence of
either an unknown trade-off or a physiological constraint.
The reduced growth rate of T. vorax in the presence of Colpidium at the higher
productivity levels can be attributed to the inefficiency of energy transfer between trophic
levels. All else being equal, T. vorax, has less energy available to it when feeding on
Colpidium than it does when feeding directly upon the basal resources (the bacteria).
However, at low productivity, T. vorax had a higher growth rate in the presence of
Colpidium than it did when feeding on bacteria alone. This could suggest that, at higher
productivities, Colpidium is more difficult to capture and, therefore, yields a lower
energy return. The cell size of Colpidium increases with productivity or nutrient
concentration (Balciunas and Lawler 1995), and it is possible that Colpidium becomes
too large for macrostomes to consume at higher productivities.
Alternatively, this might indicate that intraguild predation can be particularly
advantageous at low resource levels. I can infer that Colpidium, in the absence of
predation, is a superior competitor to T. vorax microstomes, knowing that, in
monoculture, Colpidium reduces bacterial densities to a greater extent than T. vorax
(Kaunzinger and Morin 1998, Price and Morin 2004). At low productivity, where
bacteria are relatively sparse, T. vorax may gain greater net benefits from feeding on
Colpidium than from feeding on the bacteria. In addition, cells that do not transform into
macrostomes may still benefit from the presence of macrostomes as a result of
macrostomes reducing the competitive ability of Colpidium (Tilman 1982, Thingstad
1997, Fox 2002). Measuring the impact of the presence of Colpidium on microstome
growth rates to test this hypothesis would be virtually impossible because Colpidium

21
induces transformation in microstomes. However, future studies might examine the
impact of Colpidium on closely related Tetrahymena species that do not exhibit inducible
trophic polymorphism, such as an amicronucleate strain of T. pyriformis (Nanney et al.
1998). By comparing the population dynamics of T. pyriformis with and without
Colpidium over a range of productivity, I could indirectly evaluate the potential costs of
interactions with Colpidium to T. vorax microstomes and directly test the predictions of
competitive outcomes based on resource use.
Inducible trophic polymorphisms as spectacular as those exhibited by T. vorax
may be unusual. Even among protozoa, where the rapid reversibility of transformations
that take place within a single generation is far more commonplace than in larger, more
complex organisms with developmentally stable exo- or endoskeletons, the changes in
size that allow for shifts in trophic position are very seldom accompanied by drastic
reconstructions of morphology (Kidder et al. 1941, Agrawal 2001). However,
functionally similar trophic polymorphisms do occur in a diverse array of organisms and
habitats, and not necessarily through changes in morphology. Behavior can be an
extremely plastic trait in many higher-level organisms (DeWitt and Scheiner 2004) and
can greatly influence the diet and diet breadth of those organisms (Schluter and Grant
1984, Packer and Ruttan 1988, Dean et al. 1990). Investigating the mechanisms of how
inducible trophic polymorphisms occur may shed new light on our understanding of how
phenomena such as diet breadth (Polis 1991, Morin 1999, Diehl and Feissel 2000, Jiang
and Morin 2005), cannibalism (Polis 1981), and intraguild predation (Polis et al. 1989)
impact community structure and dynamics, and even long-term evolutionary processes.
Acknowledgements

22
I would like to thank Timothy Casey, Lin Jiang, Ben Baiser, Andrea Kornbluh,
Jean Deo, Jack Siegrist, Maria Stanko, Holly Vuong, and Jennifer Adams Krumins for
their helpful comments and invaluable insights. Support for this research came from the
Rutgers School of Environmental and Biological Sciences and the New Jersey
Agricultural Experiment Station.
References
Abrahams, M. V., and L.D. Townsend. 1993. Bioluminescence in dinoflagellates: a test
of the burglar alarm hypothesis. Ecology 74(1): 258-260.
Afik, D., and W. H. Karasov. 1995. The trade-offs between digestion rate and efficiency
in warblers and their ecological implications. Ecology 76(7): 2247-2257.
Agrawal, A. A. 2001. Phenotypic plasticity in the interactions and evolution of species.
Science 294: 321-326.
Altwegg, R. 2002. Predator-induced life-history plasticity under time constraints in pool
frogs. Ecology 83(9): 2542-2551.
Baird, W. V., and J. L. Riopel. 1984. Experimental studies of haustorium initiation and
early development in Agalinis purpurea (L.) Raf. (Scrophulariaceae). American
Journal of Botany 71(6): 803-814.
Balciunas, D., and S. P. Lawler. 1995. Effects of basal resources, predation, and
alternative prey in microcosm food chains. Ecology 76(4): 1327-1336.
Barron, G. L. 1977. The nematode-destroying fungi. Canadian Biological Publications,
Guelph.
Barron, G. L. 1992. Lignolytic and cellulolytic fungi as parasites and predators. Pages
311-326 in G. C. Carroll and D. T. Wicklow, editors. The Fungal Community: Its
Organization and Role in the Ecosystem. Marcel Dekker, Inc., New York.
Berleman, J. E., and J. R. Kirby. 2007. Multicellular development in Myxococcus xanthus
is stimulated by predator-prey interactions. Journal Of Bacteriology 189(15):
5675-5682.
Crump, M. L. 1983. Opportunistic cannibalism by amphibian larvae in temporary aquatic
environments. The American Naturalist 121(2): 281-289.
Dean, W. R. J., W. R. Siegfried, and I. A. W. MacDonald. 1990. The fallacy, fact, and
fate of guiding behavior in the Greater Honeyguide. Conservation Biology 4(1):
99-101.
DeWitt, T. J., and S. M. Scheiner. 2004. Phenotypic variation from single genotypes: a
primer. Pages 1-9 in T. J. DeWitt and S. M. Scheiner, editors. Phenotypic
Plasticity: Functional and Conceptual Approaches. Oxford University Press,
Oxford.
Diehl, S., and M. Feissel. 2000. Effects of enrichment on three-level food chains with
omnivory. The American Naturalist 155(2): 200-218.

23
Dudgeon, S. R., and L. W. Buss. 1996. Growing with the flow: on the maintenance and
malleability of colony form in the hydroid Hydractinia. The American Naturalist
147(5): 667-691.
Ehlinger, T. J. 1990. Habitat choice and phenotype-limited feeding efficiency in bluegill:
individual differences and trophic polymorphism. Ecology 71(3): 886-896.
Fisher, R. A. 1958. Polymorphism and natural selection. The Journal of Ecology 46(2):
289-293.
Fox, J. W. 2002. Testing a simple rule for dominance in resource competition. The
American Naturalist 159(3): 305-319.
Fox, L. R. 1975. Cannibalism in natural populations. Annual Review of Ecology and
Systematics 6: 87-106.
Gilbert, J. J. 1973. Induction and ecological significance of gigantism in the rotifer
Asplanchna sieboldi. Science 181(4094): 63-66.
Grønlien, H. K., T. Berg, and A. M. Lovlie. 2002. In the polymorphic ciliate
Tetrahymena vorax, the non-selective phagocytosis seen in microstomes changes
to a highly selective process in macrostomes. The Journal of Experimental
Biology 205(14): 2089-2097.
Hoffman, E. A., and D. W. Pfennig. 1999. Proximate causes of cannibalistic polyphenism
in larval tiger salamanders. Ecology 80(3): 1076-1080.
Holt, R. D., and G. A. Polis. 1997. A theoretical framework for intraguild predation. The
American Naturalist 149(4): 745-764.
Jiang, L., and P. J. Morin. 2005. Predator diet breadth influences the relative importance
of bottom-up and top-down control of prey biomass and diversity. The American
Naturalist 165: 350–363.
Kaunzinger, C. M. K., and P. J. Morin. 1998. Productivity controls food chain properties
in microbial communities. Nature 395: 495-497.
Kerry, B. R., and B. A. Jaffee. 1997. Fungi as biological control agents for plant parasitic
nematodes. Pages 204-218 in D. T. Wicklow and B.E. Soderstrom, editors. The
Mycota: A Comprehensive Treatise on Fungi as Experimental Systems for Basic
Applied Research (Volume IV). Springer Verlag, Berlin Heidelberg.
Kidder, G. W., D. M. Lilly, and C. L. Claff. 1941. Growth studies on ciliates. IV. The
influence of food on the structure and growth of Glaucoma vorax sp. nov.
Biological Bulletin 80(1): 9-23.
Kopp, M., and R. Tollrian. 2003a. Reciprocal phenotypic plasticity in a predator-prey
system: inducible offences against inducible defences? Ecology Letters 6: 742-
748.
Kopp, M., and R. Tollrian. 2003b. Trophic size polyphenism in Lembadion bullinum:
costs and benefits of an inducible offense. Ecology 84(3): 641-651.
Křivan, V. 2000. Optimal intraguild foraging and population stability. Theoretical
Population Biology 58: 79-94.
Kusch, J. 1999. Self-recognition as the original function of an amoeban defense-inducing
kairomone. Ecology 80(2): 715-720.
Michimae, H., and M. Wakahara. 2002. A tadpole-induced polyphenism in the
salamander Hynobius retardatus." Evolution 56(10): 2029-2038.
Morin, P. J. 1999. Productivity, intraguild predation, and population dynamics in
experimental food webs. Ecology 80(3): 752-760.

24
Nanney, D. L., C. Park, R. Preparata, and E. M. Simon. 1998. Comparison of sequence
differences in a variable 23S rRNA domain among sets of cryptic species of
ciliated protozoa. The Journal of Eukaryotic Microbiology 45(1): 91-100.
Packer, C., and L. Ruttan. 1988. The evolution of cooperative hunting. The American
Naturalist 132(2): 159-192.
Padilla, D. K. 2001. Food and environmental cues trigger an inducible offense.
Evolutionary Ecology Research 3: 15-25.
Pfennig, D. W. 1992. Proximate and functional causes of polyphenism in an anuran
tadpole. Functional Ecology 6(2): 167-174.
Pfennig, D. W. 1999. Cannibalistic tadpoles that pose the greatest threat to kin are most
likely to discriminate kin. Proceedings of the Royal Society of London: Biological
Sciences 266(1414): 57-61.
Pfennig, D. W. 2000. Effect of predator-prey phylogenetic similarity on the fitness
consequences of predation: a trade-off between nutrition and disease? The
American Naturalist 155(3): 335-345.
Pfennig, D. W., and J. P. Collins. 1993. Kinship affects morphogenesis in cannibalistic
salamanders. Nature 362: 836-838.
Pfennig, D. W., and W. A. Frankino. 1997. Kin-mediated morphogenesis in facultatively
cannibalistic tadpoles. Evolution 51(6): 1993-1999.
Polis, G. A. 1981. The evolution and dynamics of intraspecific predation. Annual Review
of Ecology and Systematics 12: 225-251.
Polis, G. A. 1991. Complex trophic interactions in deserts: an empirical critique of food-
web theory. The American Naturalist 138: 123-155.
Polis, G. A., C. A. Myers, and R. D. Holt. 1989. The ecology and evolution of intraguild
predation: potential competitors that eat each other. Annual Review of Ecology
and Systematics 20: 297-330.
Porter, K. G., and J. W. Porter. 1979. Bioluminescence in marine plankton: a coevolved
antipredation system. The American Naturalist 114(3): 458-461.
Price, J. E., and P. J. Morin. 2004. Colonization history determines alternate community
states in a food web of intraguild predators. Ecology 85: 1017-1028.
Relyea, R. A. 2002. Costs of phenotypic plasticity. American Naturalist 159: 272-282.
Ryals, P. E., H. E. Smith-Somerville, and H. E. Buhse. 2002. Phenotype switching in
polymorphic Tetrahymena: a single-cell Jekyll and Hyde. International review of
cytology 212: 209-238.
Schluter, D., and P. R. Grant. 1984. Ecological correlates of morphological evolution in a
Darwin's Finch, Geospiza difficilis. Evolution 38(4): 856-869.
Smith-Somerville, H. E., J. K. Hardman, R. Timkovich, W. J. Ray, K. E. Rose, P. E.
Ryals, S. H. Gibbons, and H. E. Buhse Jr. 2000. A complex of iron and nucleic
acid catabolites is a signal that triggers differentiation in a freshwater protozoan.
Proceedings of the National Academy of Sciences 97(13): 7325-7330.
Thingstad, T. F. 1997. A theoretical approach to structuring mechanisms in the pelagic
food web. Hydrobiologia 363: 59-72.
Thingstad, T. F., H. Havskum, K. Garde, and B. Riemann. 1996. On the strategy of
"eating your competitor": a mathematical analysis of algal mixotrophy. Ecology
77(7): 2108-2118.

25
Tilman, D. 1982. Resource Competition and Community Structure. Princeton University
Press, Princeton.
Tollrian, R., and C. D. Harvell. 1999. The Ecology and Evolution of Inducible Defenses.
Princeton University Press, Princeton.
van Baalen, M., V. Krivan, P. C. J. van Rijn, and M. W. Sabelis. 2001. Alternative food,
switching predators, and the persistence of predator-prey systems. The American
Naturalist 157(5): 512-524.
Van den Bosch, F., A. M. De Roos, and W. Gabriel. 1988. Cannibalism as a life boat
mechanism. Journal of Mathematical Biology 26: 619-633.
Wainwright, P. C., C. W. Osenberg, and G. G. Mittelbach. 1991. Trophic polymorphism
in the Pumpkinseed Sunfish (Lepomis gibbosus Linnaeus): effects of environment
on ontogeny. Functional Ecology 5: 40-55.
Washburn, J. O., M. E. Gross, D. R. Mercer, and J. R. Anderson. 1988. Predator-induced
trophic shift of a free-living ciliate: parasitism of mosquito larvae by their prey.
Science 240: 1193-1195.
Williams, N. E. 1961. Polymorphism in Tetrahymena vorax. The Journal of Eukaryotic
Microbiology 8(4): 403-410.

26
low density,
low competition
high density,
high competition
Colpidium:
interspecific
competitor
Microstome:
inducible
bacterivore
Macrostome:
induced
predator
Tailed
Microstome:
induced
bacterivore
Colpidium:
intraguild
prey
Resource = bacteriaResource = bacteria
60 μm
90 μm
< 3 μm
75 μm
75 μm
< 3 μm
200 μm
Fig. 1. Diagram of the inducible trophic polymorphism of T. vorax. Gray arrows
represent developmental pathways. Black arrows represent trophic links. Arrow size
represents relative interaction strength. Transformation of microstomes into
macrostomes is triggered by chemical cues released by conspecifics and by interspecific
competitors for bacteria, such as Colpidium (Kidder et al. 1941; Smith-Somerville 2000;
Ryals et al. 2002).

27
Fig. 2. Comparison of the differences in susceptibility of microstomes, tailed
microstomes, and Colpidium to consumption by macrostomes. Black squares mark the
regression line for microstomes; black triangles mark that of tailed microstomes; and gray
circles mark that of Colpidium.
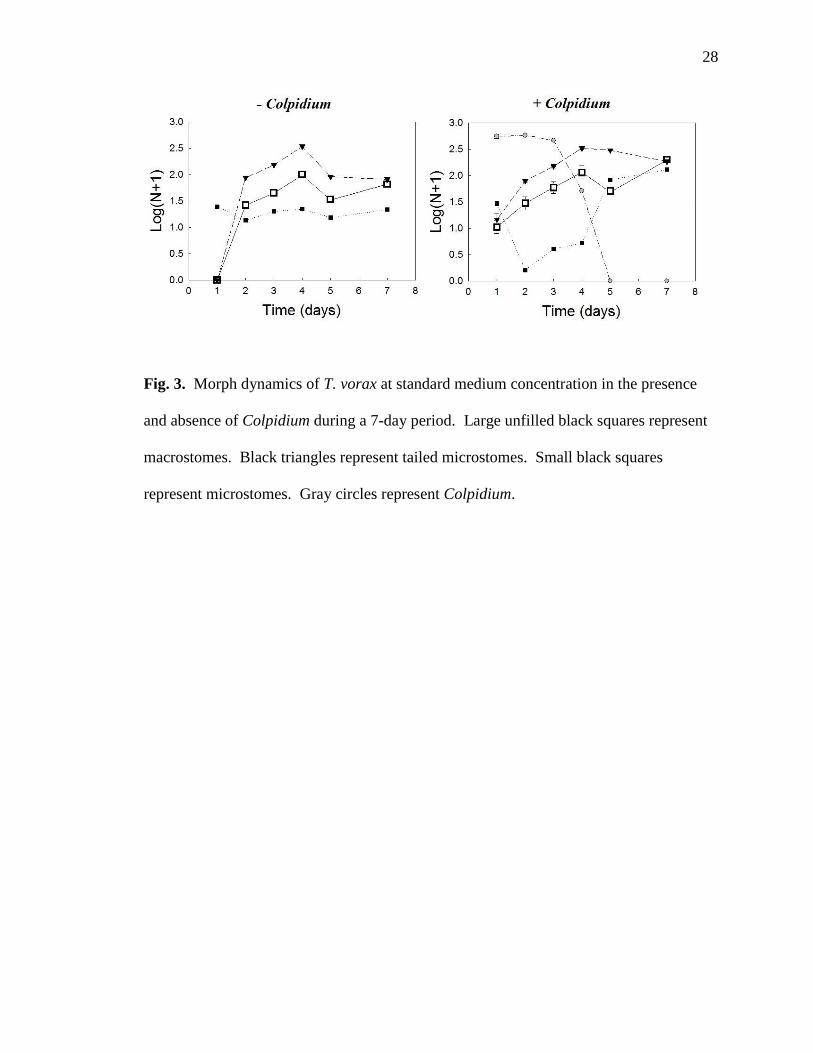
28
Fig. 3. Morph dynamics of T. vorax at standard medium concentration in the presence
and absence of Colpidium during a 7-day period. Large unfilled black squares represent
macrostomes. Black triangles represent tailed microstomes. Small black squares
represent microstomes. Gray circles represent Colpidium.

29
Fig. 4. Effect of microstome cell size (small < 60 µm; large ≥ 60 µm) on the outcome of
induction. Symbols are as in Fig. 3.

30
Fig. 5. Morph dynamics in the presence and absence of Colpidium at low (50% of
standard) and high (150% of standard) medium concentrations. Symbols are as in Fig. 3.

31
Fig. 6. Interaction between the effects of productivity and the presence or absence of
Colpidium on the rate of growth of T. vorax, calculated as the natural log of the mean cell
density of T. vorax over time. Black diamonds represent growth rates with Colpidium
present. White diamonds represent growth rates with Colpidium absent.

32
CHAPTER 2
Prey-Induced Gape Expansion Mediates Apparent Competition
Summary
1. Phenotypic plasticity allows certain predators to increase their gape by feeding
on increasingly large prey. Where predators increase their gape enough to
feed on initially inedible species, a novel, trait-mediated indirect interaction
may occur – one that is akin to apparent competition.
2. I tested the hypothesis that feeding on Colpidium kleini, a large intraguild
prey, enables macrostomes of Tetrahymena vorax to increase their gape
enough to engulf Paramecium aurelia, a species too large for macrostomes to
engulf prior to such feeding.
3. My results were consistent with the hypothesis. Macrostomes allowed to feed
on C. kleini showed a significant increase in their mean cell diameter over
time. When exposed to P. aurelia, these enlarged macrostomes were able to
engulf P. aurelia, whereas macrostomes that had been cultured in the absence
of C. kleini were unable to engulf P. aurelia. The population dynamics of T.
vorax, C. kleini, and P. aurelia during a 22-day period revealed that the
occurrence and outcome of expanded diet breadth resulting from increased
gape in T. vorax depended on the recent history of community assembly.
Key-words: Chasmatectasis, phenotypic plasticity, priority effect, inducible trophic
polymorphism (ITP), Tetrahymena vorax, trait-mediated indirect interaction (TMII)

33
Introduction
Phenotypic plasticity is the ability of a single genotype to produce two or more
alternative phenotypes in response to differing environmental conditions (DeWitt and
Scheiner 2004). One intriguing consequence of phenotypic plasticity is the generation of
inducible trophic polymorphisms, wherein morphologically or physiologically different
individuals within a population come to occupy different trophic positions (Kopp and
Tollrian 2003a, Banerji and Morin 2009). Inducible trophic polymorphisms can result in
the utilization of a broader range of available resources than would be used by
monomorphic populations. In addition, induced individuals may be capable of
consuming some of their natural enemies (Washburn et al. 1988, Padilla 2001, Price and
Morin 2004). Since a major determinant of whether or not a predator is able to capture
and consume its prey is the disparity in size between the two (Maret and Collins 1996,
Urban 2007), it is not surprising that, in inducible trophic polymorphisms, a shift from a
lower trophic position to a higher one often simply entails an increase in size – either of
the body in its entirety or of specific internal and external feeding structures (Polis 1981,
Ricci et al. 1989, Wainwright et al. 1991, Maerz et al. 2006, Olsson et al. 2007).
More surprising is that certain species with inducible predatory morphs can
continue to increase their gape after the initial induction of their predatory morphology
by ingesting increasingly large prey. As I have not been able to find a precise term for
this phenomenon anywhere in the ecological literature, I have borrowed the pseudo-
clinical term used jokingly by a handful of medical students with whom I am acquainted
to describe the preferred training method of professional gurgitators (Levine et al. 2007,
O’Connor 2007): chasmatectasis (from the Greek words khasma, “gape,” and ektasis,

34
“stretching”). Chasmatectasis is an inducible offense (Padilla 2001) that has the potential
to significantly alter the shape and outcome of ecological dynamics (Miner et al. 2005,
Jeschke 2006, Mougi et al. 2010). It may, for example, provide a counter-measure
against the inducible defenses of prey (Kopp and Tollrian 2003b, Kishida et al. 2006) or
enhance the ability of predators to survive and recover from periods of starvation (Hewett
1987). There is anecdotal evidence of the occurrence of chasmatectasis in rotifers
(Gilbert 1973), cladocerans (Abrusán 2003), amphibians (Collins and Cheek 1983,
Pfennig 1990, Walls et al. 1993), fish (Wainwright et al. 1991, Persson et al. 2004), and
reptiles (Queral-Regil and King 1998). Among protists, and particularly among ciliates,
definite examples of chasmatectasis have been observed relatively frequently (Giese and
Alden 1938, Rickards and Lynn 1985, Hewett 1987, Ricci et al. 1989, Kopp and Tollrian
2003a).
An ecological aspect of chasmatectasis that has not previously been explored is its
potential to give rise to novel indirect interactions (Wootton 1994). In particular,
chasmatectasis might enable an induced predator to increase its gape enough to consume
a species that would have been too large for it to consume at the time of its initial
induction. The resulting exchange of negative indirect effects between the smaller and
larger prey species via their now shared predator would constitute apparent competition
(Holt 1977, Bonsall and Hassell 1997, Chaneton and Bonsall 2000). However, the
effects would be mediated via the predator’s gape rather than its population size, making
this a “trait-mediated” indirect interaction, instead of a “density-mediated” indirect
interaction (Peacor and Werner 2001). To explore this possibility, I made use of the

35
inducible trophic polymorphism of the freshwater protozoan Tetrahymena vorax Kidder
(Ciliatea: Hymenostomatida).
Isogenic populations of T. vorax may contain both “microstomes” and
“macrostomes.” Microstomes are small, pyriform individuals that feed on bacteria.
Macrostomes are larger, ovoid individuals that consume other ciliates. Microstomes can
reversibly transform into macrostomes in the presence of intraguild prey, which may
include conspecifics (Kidder et al. 1941, Williams 1961, Banerji and Morin 2009). My
preliminary observations of macrostomes exposed to varying sizes of intraguild prey
indicate that macrostomes are capable of some degree of chasmatectasis. Here I describe
my experimental approach to quantifying these observations and testing the hypothesis
that feeding on accessible prey, such as Colpidium kleini Foissner (Ciliatea:
Hymenostomatida), enables macrostomes to attain sizes large enough to engulf species
such as Paramecium aurelia Müller (Ciliatea: Hymenostomatida), which, prior to such
feeding, would exceed the limit of their gape (Fig. 1).
Methods
Culturing of Ciliates
The distribution of T. vorax in nature is poorly known, a situation that is typical
for protists and other microbes (Fenchel et al. 1997, Finlay 2002, Foissner 2009).
Fortuitously, many of the protists known to occur with T. vorax have cosmopolitan
distributions (Finlay 2002). Among these are species of the genus Colpidium, which
Kidder et al. (1941) were able to identify as intraguild prey of T. vorax almost
immediately upon its discovery. Members of the widely distributed Paramecium aurelia
complex are also likely to be found where T. vorax is present (Sonneborn 1975, Finlay

36
2002). I chose C. kleini and P. aurelia for use in my investigation for the ease with
which they could be obtained and cultured.
I purchased T. vorax (strain 30421) and P. aurelia (Item # 131546) from the
American Type Culture Collection and the Carolina Biological Supply Company,
respectively, and isolated C. kleini from the Adelphia Plant Science Research and
Extension Center (Freehold, NJ 07728, USA). I grew separate, isogenic lines of each of
these three ciliates in 240-mL glass jars containing 100 mL of complex organic growth
medium. The medium I used consisted of 0.37 g of Carolina Biological Supply
protozoan pellets per L of filtered well water. I sterilized and cooled the medium to room
temperature and then inoculated it with three different species of bacteria – Serratia
marcescens, Bacillus subtilis, and Bacillus cereus – to serve as a basal resource for the
ciliates. Loosely tightened screw caps on the jars permitted air circulation, while limiting
evaporation and contamination.
Measuring Chasmatectasis in T. vorax
To verify that macrostomes are able to increase their size and consume larger
prey, I performed an induction experiment. I inoculated six 15-mL Petri dishes,
containing 7 mL of growth medium, with T. vorax. I allowed three of these T. vorax
cultures to feed solely on bacteria (control replicates), while immediately inoculating the
other three with C. kleini (treatment replicates). After C. kleini had been extirpated from
the treatment replicates and all six replicates had entered into logarithmic growth (~ 4.5
days), I inoculated all six cultures with P. aurelia. I sampled approximately 0.3 mL from
the cultures every 12 hours over the course of 10 days to record average macrostome cell
size and the frequency of predation.

37
I measured average macrostome cell size as the mean cell diameter, in
micrometers, of macrostomes that were present within the sample. I made individual cell
diameter measurements by taking digital photomicrographs of the macrostomes in my
samples at 400X magnification. I used a Nikon Eclipse 80i microscope equipped with a
Nikon DXM1200C camera and ACT-1C imaging software. I analyzed the photos using
the NIH program, ImageJ (see Rasband 2007). I measured frequency of predation on P.
aurelia as the total number of observed incidences of a macrostome having engulfed P.
aurelia during my sampling interval (~ 5 minutes per dish).
Determining the Effect of Community Assembly on the Occurrence and Outcome of
Chasmatectasis in T. vorax
I performed a community assembly experiment to address: (1) whether feeding on
C. kleini prior to being exposed to P. aurelia is required for chasmatectasis in T. vorax
and (2) how chasmatectasis in T. vorax affects the relative fitness of all three species. My
experimental design involved four treatments, with five replicates per treatment. Each
replicate consisted of 100 ml of growth medium in a covered 240-mL glass jar that was
initially inoculated with T. vorax. C. kleini and P. aurelia were later added to the
replicates in different temporal sequences (Table 1).
I monitored the populations of all of the species present in each jar approximately
every 24 hours, over the course of 22 days, sampling approximately 0.3 mL without
replacement from each jar. To ensure that effects of the treatments were not confounded
by the build-up of toxins and metabolic wastes in the system, I removed and discarded
approximately 10% of medium by volume from each jar and replaced it with an
equivalent volume of fresh medium on Day 16. No measurements were recorded on this

38
day. Adhering to the same procedure, I constructed and monitored a pair of controls (5
replicates per control), each containing both C. kleini and P. aurelia, but without T.
vorax. This provided a baseline to assess any indirect effects among C. kleini and P.
aurelia mediated by T. vorax (Table 1).
Results
Measurement of Chasmatectasis in T. vorax
Macrostomes fed C. kleini showed a significant increase in size (ANOVA, F1,277 =
87.41, p < 0.0001) over the 10-day period of study (Fig. 2, Fig. 3). On Days 6-8, Day 9,
and Day 10, I recorded no average cell diameter of macrostomes in the control replicates
because no macrostomes were present in the control replicates during this period.
Macrostomes in the control replicates that had been cultured in the absence of C. kleini
appeared to have reverted to microstomes in the presence of P. aurelia. While I observed
8 separate incidences of macrostomes engulfing P. aurelia in the treatment replicates, I
did not observe any incidence of macrostomes doing so in the control replicates.
Effect of Community Assembly on the Occurrence and Outcome of Chasmatectasis in
T. vorax
The order in which T. vorax was exposed to C. kleini and P. aurelia had a
significant effect on the mean population densities of both C. kleini (ANOVA, F1,8 =
407.57, p < 0.0001) and P. aurelia (ANOVA, F1,8 = 3162.48, p < 0.0001) across time. It
also had a significant effect on the mean densities of macrostomes (ANOVA, F1,8 =
144.2, p < 0.0001) and microstomes (ANOVA, F1,8 = 179.98, p < 0.0001) in the T. vorax
population. In replicates where T. vorax was exposed to C. kleini first and to P. aurelia
second, P. aurelia was ultimately extirpated. In replicates where T. vorax was exposed to

39
P. aurelia before being exposed to C. kleini, however, P. aurelia was able to persist
throughout the experiment. In the controls, where C. kleini and P. aurelia were paired in
the absence of T. vorax, both species persisted throughout the experiment (Fig. 4). C.
kleini and P. aurelia were both able to persist at relatively high population density when
paired alone with T. vorax (Fig. 5). T. vorax macrostomes, in the presence of both prey,
attained higher densities and persisted longer when exposed to C. kleini first. In the
presence of C. kleini alone, T. vorax macrostomes attained higher population densities
and persisted longer than in the presence of P. aurelia alone (Fig. 6).
Discussion
My results strongly support the hypothesis that chasmatectasis can enable a
predator to expand its diet breadth to include species that were previously inaccessible
due to their large size relative to the predator’s initial gape. By feeding on C. kleini, a
relatively large but still initially consumable prey, T. vorax was able to expand its gape
enough to consume P. aurelia, a prey that T. vorax could not consume upon initial
transformation. Furthermore, I have demonstrated that the occurrence and ecological
consequences of chasmatectasis can be influenced by the history of community assembly.
Where T. vorax was exposed to C. kleini before being exposed to P. aurelia, T. vorax
underwent chasmatectasis and became able to feed on P. aurelia. Yet, where T. vorax
had been exposed to P. aurelia first, T. vorax not only failed to undergo chasmatectasis,
but also maintained a lower population density of macrostomes throughout the
experiment. It is conceivable that competition with P. aurelia for bacterial resources
might have prevented T. vorax from crossing the minimum size threshold for
transformation into macrostomes (Banerji and Morin 2009) upon the arrival of C. kleini

40
and that maintenance of the macrostome morphology requires ingestion of prey. In the
replicates in which T. vorax was able to undergo chasmatectasis and feed on P. aurelia,
P. aurelia was extirpated. Given the low rate of consumption of P. aurelia by T. vorax
indicated by the results of the induction experiment, the extirpation of P. aurelia in these
replicates cannot be attributed to predation alone. However, given the persistence of P.
aurelia in the replicates in which T. vorax had been unable to undergo chasmatectasis, it
appears that predation must still have played a significant role.
Chasmatectasis is an intriguing manifestation of phenotypic plasticity that
warrants further investigation in other taxa. The underlying genetic and physiological
basis for chasmatectasis has yet to be revealed, and its environmental basis in natural
systems remains uncertain. In the case of my system, for example, it is unclear whether
the ability of C. kleini and other large, but accessible prey to trigger chasmatectasis in T.
vorax is a function of the size of the prey, in and of itself, or of the composition – i.e.,
nutritional or energetic content – of the prey. Still to be determined, also, is the upper
size limit of prey that can be consumed by T. vorax via chasmatectasis. Future studies
might readily address these questions by repeating my experimental design using
different prey species of varying size and composition.
Chasmatectasis is not limited to ciliates. Though they differ from the examples
seen in ciliates in that they are generally irreversible, examples of chasmatectasis, as
previously mentioned, can be found in fish, amphibians, reptiles, and a range of
invertebrates. Dynamics similar to those I have uncovered in this investigation may very
well occur in these other systems. Such dynamics could have repercussions for the study
of colonization and succession. For example, given that overall body size has been

41
shown to play a significant role in determining the invasion success of predators (Persson
et al. 2004, Schröder et al. 2009), diversity of body sizes among incumbent prey may
facilitate invasion by predators capable of undergoing chasmatectasis.
Dynamics resulting from chasmatectasis might also have implications for
management practice, as shown by my demonstration of chasmatectasis mediating
apparent competition. Apparent competition has been shown to be an important
determinant of community structure (Jeffries and Lawton 1984, Settle and Wilson 1990,
Bonsall and Hassell 1997, Juliano 1998, Orrock et al. 2008), and classical apparent
competition theory has been applied in the contexts of biological control (Karban et al.
1994, van Veen et al. 2006) and restoration (Norbury 2001, Hoddle 2004).
Understanding the workings of chasmatectasis could enable us to determine if apparent
competition that is mediated by chasmatectasis has the same significance in ecological
communities as classical apparent competition and enable us to avoid the potential
pitfalls (Howarth 1991, Paynter et al. 2010) associated with its use in biological control
and restoration.
Acknowledgements
I gratefully acknowledge Rebecca Jordan, Mike Pace, Kevin Aagaard, Laura
Chen, Jean Deo, Sean Hayes, Ruchi Patel, Jack Siegrist, Maria Stanko, Holly Vuong, and
Talia Young for their worthwhile comments and suggestions. Support for this research
came from the School of Environmental and Biological Sciences and the New Jersey
Agricultural Experiment Station.
References
Abrusán, G. 2003. Morphological variation of the predatory cladoceran Leptodora kindtii
in relation to prey characteristics. Oecologia 134(2): 278-283.

42
Banerji, A., and P. J. Morin. 2009. Phenotypic plasticity, intraguild predation and anti-
cannibal defences in an enigmatic polymorphic ciliate. Functional Ecology 23:
427-434.
Bonsall, M. B., and M. P. Hassell. 1997. Apparent competition structures ecological
assemblages. Nature 388: 371-373.
Chaneton, E. J., and M. B. Bonsall. 2000. Enemy-mediated apparent competition:
empirical patterns and evidence. Oikos 88(2): 380-394.
Collins, J. P., and J. E. Cheek. 1983. Effect of food and density on development of typical
and cannibalistic salamander larvae in Ambystoma tigrinum nebulosum. American
Zoologist 23: 77-84.
DeWitt, T. J., and S. M. Scheiner. 2004. Phenotypic variation from single genotypes: a
primer. Pages 1-9 in T. J. DeWitt and S. M. Scheiner, editors. Phenotypic
Plasticity: Functional and Conceptual Approaches. Oxford University Press,
Oxford.
Fenchel, T., G. F. Esteban, and B. J. Finlay. 1997. Local versus global diversity of
microorganisms: cryptic diversity of ciliated protozoa. Oikos 80: 220-225.
Finlay, B. J. 2002. Global dispersal of free-living microbial eukaryote species. Science
296: 1061-1063.
Foissner, W. 2009. Protist diversity and geographical distribution. Topics in Biodiversity
and Conservation 8: 1-8.
Giese, A. C., and R. H. Alden. 1938. Cannibalism and giant formation in Stylonychia.
Journal of Experimental Zoology 78(2): 117-134.
Gilbert, J. J. 1973. Induction and ecological significance of gigantism in the rotifer
Asplanchna sieboldi. Science 181(4094): 63-66.
Hewett, S. W. 1987. Prey size and survivorship in Didinium nasutum (Ciliophera:
Gymnostomatida). Transactions of the American Microscopical Society 106(2):
134-138.
Hoddle, M. S. 2004. Restoring balance: using exotic species to control invasive exotic
species. Conservation Biology 18: 38-49.
Holt, R. D. 1977. Predation, apparent competition, and the structure of prey communities.
Theoretical Population Biology 12: 197-229.
Howarth, F. G. 1991. Environmental impacts of classical biological control. Annual
Review of Entomology 36: 485-509.
Jeffries, M. J., and J. H. Lawton. 1984. Enemy-free space and the structure of ecological
communities. Biological Journal of the Linnean Society 23: 269-286.
Jeschke, J. M. 2006. Density-dependent effects of prey defenses and predator offenses.
Journal of Theoretical Biology 242(4): 900-907.
Juliano, S. A. 1998. Species introduction and replacement among mosquitoes:
interspecific resource competition or apparent competition? Ecology 79: 255-268.
Karban, R., D. Hougen-Eitzmann, and G. English-Loeb. 1994. Predator-mediated
apparent competition between two herbivores that feed on grapevines. Oecologia
97: 508-511.
Kopp, M., and R. Tollrian. 2003a. Trophic size polyphenism in Lembadion bullinum:
costs and benefits of an inducible offense. Ecology 84(3): 641-651.

43
Kopp, M., and R. Tollrian. 2003b. Reciprocal phenotypic plasticity in a predator-prey
system: inducible offences against inducible defences? Ecology Letters 6: 742-
748.
Kidder, G. W., D. M. Lilly, and C. L. Claff. 1941. Growth Studies on Ciliates. IV. The
influence of food on the structure and growth of Glaucoma vorax sp. nov.
Biological Bulletin 80: 9-23.
Kishida, O., Y. Mizuta, and K. Nishimura. 2006. Reciprocal phenotypic plasticity in a
predator-prey interaction between larval amphibians. Ecology 87(6): 1599-1604.
Levine, M. S., G. Spencer, A. Alavi, and D. C. Metz. 2007. Competitive speed-eating:
truth and consequences. American Journal of Roentgenology 189: 681-686.
Maerz, J. C., E. M. Myers, and D. C. Adams. 2006. Trophic polymorphism in a terrestrial
salamander. Evolutionary Ecology Research 8: 23-35.
Maret, T. J., and J. P. Collins. 1996. Effect of prey vulnerability on population size
structure of a gape-limited predator. Ecology 77(1): 320-324.
Miner, B. G., S. E. Sultan, S. G. Morgan, D. K. Padilla, and R. A. Relyea. 2005.
Ecological consequences of phenotypic plasticity. Trends in Ecology and
Evolution 20(12): 685-692.
Mougi, A., O. Kishida, and Y. Iwasa. 2010. Coevolution of phenotypic plasticity in
predator and prey: why are inducible offenses rarer than inducible defenses?
Evolution, no. doi: 10.1111/j.1558-5646.2010.01187.x
Norbury, G. 2001. Conserving dryland lizards by reducing predator-mediated apparent
competition and direct competition with introduced rabbits. Journal of Applied
Ecology 38(6): 1350-1361.
O'Connor, J. 2007. Feeding frenzy. Gastronomica: The Journal of Food and Culture 7(4):
92-94.
Olsson, J., M. Quevedo, C. Colson, and R. Svanbäck. 2007. Gut length plasticity in
perch: into the bowels of resource polymorphisms. Biological Journal of the
Linnean Society 90: 517-523.
Orrock, J. L., M. S. Witter, and O. J. Reichman. 2008. Apparent competition with an
exotic plant reduces native plant establishment. Ecology 89: 1168-1174.
Padilla, D. K. 2001. Food and environmental cues trigger an inducible offense.
Evolutionary Ecology Research 3: 15-25.
Paynter, Q., S. V. Fowler, A. H. Gourlay, R. Groenteman, P. G. Peterson, L. Smith, C. J.
Winks. 2010. Predicting parasitoid accumulation on biological control agents of
weeds. Journal of Applied Ecology 47(3): 575-582.
Peacor, S. D., and E. E. Werner. 2001. The contribution of trait-mediated indirect effects
to the net effects of a predator. Proceedings of the National Academy of Sciences
of the United States of America 98(7): 3904-3908.
Persson, L., D. Claessen, A. M. De Roos, P. Bystrom, S. Sjogren, R. Svanback, E.
Wahlstrom, and E. Westman. 2004. Cannibalism in a size-structured population:
energy extraction and control. Ecological Monographs 74(1): 135-157.
Pfennig, D. W. 1990. The adaptive significance of an environmentally-cued
developmental switch in an anuran tadpole. Oecologia 85: 101-107.
Price, J. E., and P. J. Morin. 2004. Colonization history determines alternate community
states in a food web of intraguild predators. Ecology 85(4): 1017-1028.

44
Queral-Regil, A., and R. B. King. 1998. Evidence for phenotypic plasticity in snake body
size and relative head dimensions in response to amount and size of prey. Copeia
1998(2): 423-429.
Rasband, W. 2007. ImageJ for microscopy: image processing and analysis in Java.
Retrieved November 24, 2008 from: http://rsbweb.nih.gov/ij/
Ricci, N., R. Banchetti, and R. Pelamatti. 1989. Reversible carnivory in Oxytricha
bifaria: a peculiar ecological adaptation. Hydrobiologia 182(2): 115-120.
Rickards, J. C., and D. H. Lynn. 1985. Can ciliates adjust their intermembranellar spacing
to prey size? Transactions of the American Microscopical Society 104(4): 333-
340.
Schröder, A., K. A. Nilsson, L. Persson, T. van Kooten, and B. Reichstein. 2009.
Invasion success depends on invader body size in a size-structured mixed
predation-competition community. Journal of Animal Ecology 78(6): 1152-1162.
Settle, W. H., and L. T. Wilson. 1990. Invasion by the variegated leafhopper and biotic
interactions: parasitism, competition and apparent competition. Ecology 71: 1461-
1470.
Sonneborn, T. M. 1975. The Paramecium aurelia complex of 14 sibling species.
Transactions of the American Microscopical Society 94(2): 155-178.
Urban, M. C. 2007. The growth-predation risk trade-off under a growing gape-limited
predation threat. Ecology 88: 2587-2597.
Van Veen, F. J. F., J. Memmott, and H. C. J. Godfray. 2006. Indirect effects, apparent
competition, and biological control. Pages 145-169 in J. Brodeur and G. Boivin,
editors. Trophic and Guild Interactions in Biological Control. Springer.
Wainwright, P. C., C. W. Osenberg, and G. G. Mittelbach. 1991. Trophic Polymorphism
in the pumpkinseed sunfish (Lepomis gibbosus Linnaeus): effects of the
environment on ontogeny. Functional Ecology 5(1): 40-55.
Walls, S. C., S. S. Belanger, and A. R. Blaustein. 1993. Morphological variation in a
larval salamander: dietary induction of plasticity in head shape. Oecologia 96(2):
162-168.
Washburn, J. O., M. E. Gross, D. R. Mercer, and J. R. Anderson. 1988. Predator-induced
trophic shift of a free-living ciliate: Parasitism of mosquito larvae by their prey.
Science 240(4856): 1193-1195
Williams, N. E. 1961. Polymorphism in Tetrahymena vorax. The Journal of Eukaryotic
Microbiology 8: 403-410
Wootton, J. T. 1994. The nature and consequences of indirect effects in ecological
communities. Annual Review of Ecology and Systematics 25: 443-466.

45
==========================================================
Species Added
on Day 0
Species Added
on Day 3
Species Added
on Day 6
Treatment
1 T. vorax C. kleini P. aurelia
2 T. vorax P. aurelia C. kleini
3 T. vorax C. kleini C. kleini
4 T. vorax P. aurelia P. aurelia
Control
1 C. kleini P. aurelia P. aurelia
2 P. aurelia C. kleini C. kleini
============================================================
Table 1. Inoculation sequence for community assembly experiment. Treatment
replicates were initially inoculated with T. vorax and later inoculated with C. kleini, P.
aurelia, or both in different orders. Control replicates consisted of C. kleini and P.
aurelia without T. vorax.

46
Fig. 1. Chasmatectasis in T. vorax may create negative trait-mediated indirect effects
between C. kleini and P. aurelia. Black arrows represent trophic links, while gray arrows
represent developmental pathways.

47
Fig. 2. Mean macrostome cell diameter over a 10-day period. White diamonds represent
diameters observed in the presence of C. kleini; black diamonds represent diameters in
the absence of C. kleini. On Days 6-8, Day 9, and Day 10, no macrostomes were
observed in the absence of C. kleini. On Day 8.5, only 3 macrostomes were observed in
the absence of C. kleini, one of which being a low-end outlier in terms of its cell
diameter.

48
Fig. 3. Representative micrographs (differential image contrast scans) of T. vorax
macrostomes observed on the last day of my induction experiment (Day 10). Shown on
the left is a macrostome from a control replicate (monoculture with bacteria as the only
available prey). Shown on the right is a macrostome from a treatment replicate (initially
containing C. kleini as well as bacteria).
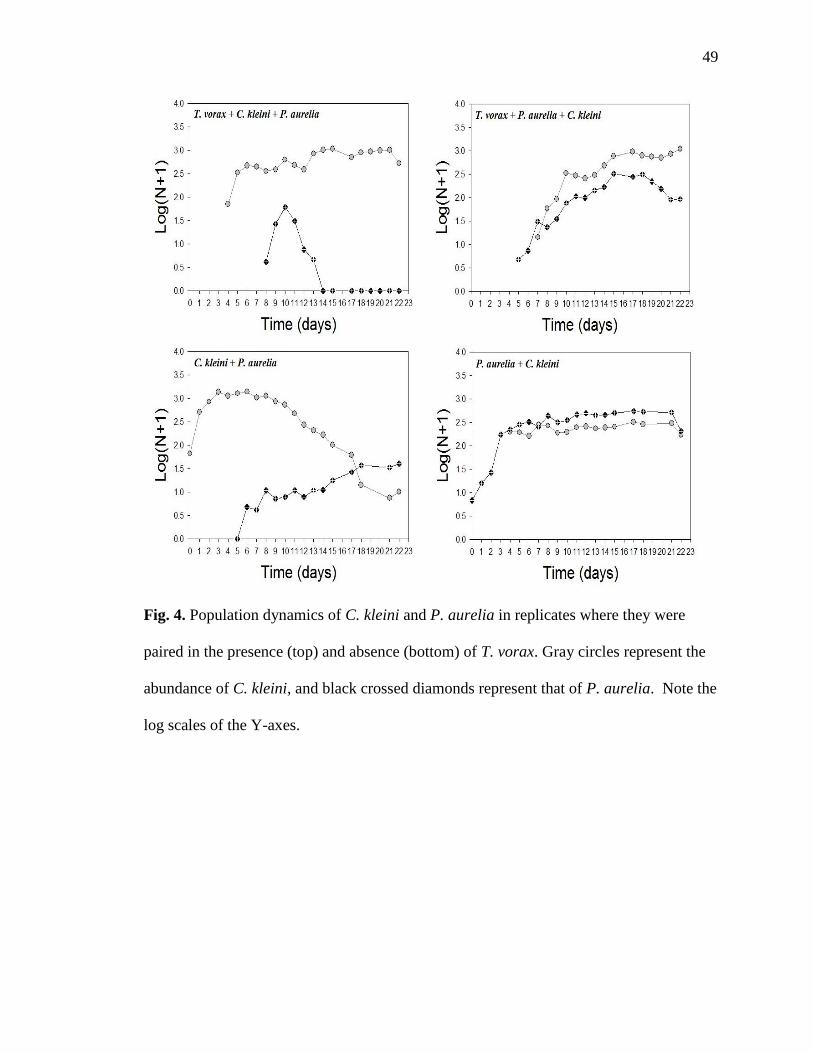
49
Fig. 4. Population dynamics of C. kleini and P. aurelia in replicates where they were
paired in the presence (top) and absence (bottom) of T. vorax. Gray circles represent the
abundance of C. kleini, and black crossed diamonds represent that of P. aurelia. Note the
log scales of the Y-axes.

50
Fig. 5. Population dynamics of C. kleini (left) and P. aurelia (right) alone with T. vorax.
Symbols are as in Fig. 4.

51
Fig. 6. Morph dynamics of T. vorax in the replicates shown at the top of Fig. 4 (top) and
shown in Fig. 5 (bottom). Large unfilled squares represent the abundance of
macrostomes. Small black squares represent that of microstomes. Note the log scales of
the Y-axes.

52
CHAPTER 3
Dynamics between Protists and Crustacean Zooplankton: Do Copepods
Show Preference for the Predatory Morph of Tetrahymena vorax?
Summary
1. Studies of protists in microcosms have made important contributions to
ecology, but some critics argue that the small spatial scale and trophic
simplicity of microcosms make them inappropriate as models of complex
natural systems. This concern is especially important to address in cases
where both the nature and strength of trophic links within the community are
density-dependent.
2. I used outdoor experimental mesocosms to assess whether increased trophic
complexity and variable abiotic conditions would alter the dynamics of a
simple freshwater protist community module that I had previously examined
in laboratory settings. This community contained the polymorphic ciliate
Tetrahymena vorax Kidder. I tested the following hypotheses: (1) that the
volume and ambient abiotic conditions of the mesocosm habitat would inhibit
the induction of predatory morphs of T. vorax, and (2) that predation by
crustacean zooplankton would result in the exclusion of T. vorax.
3. Differentiation did occur in the T. vorax population over the course of the
experiment. The proportion of T. vorax macrostomes was unaffected by the
presence or absence of crustacean zooplankton. T. vorax was not extirpated in
the presence of the zooplankton. However, the population densities of each of

53
the morphs of T. vorax, as well as that of Colpidium kleini (a competing
ciliate), were significantly reduced in the presence of zooplankton, relative to
what they were in the absence of zooplankton.
4. These results show that community patterns may change radically with the
scale and complexity of experimental systems, demonstrate the utility of
microcosms as a baseline for studies of larger systems, and offer insight into
possible reasons for the limited distribution of T. vorax in natural aquatic
systems.
Key-words: density-dependence, induced offenses, induced polymorphism, microcosm
experiments, predation, spatial scaling, Tetrahymena vorax, zooplankton

54
Introduction
The experimental tractability of protists in laboratory microcosms has made them
valuable tools for testing ecological and evolutionary principles and for uncovering the
mechanisms that underlie general patterns (Drake et al. 1996; Fraser and Keddy 1997;
Cadotte et al. 2005; Holyoak and Lawler 2005). The small size and self-contained
structure of microcosms allow for highly controlled experiments with adequate
replication (Jessup et al. 2004; Cadotte et al. 2005) – a feature that is sometimes lacking
and occasionally impossible in field experiments (Hurlbert 1984; Hargrove and Pickering
1992). These same convenient attributes can potentially make microcosms limited
models of larger, more complex systems (Carpenter 1996, 1999; Srivastava et al. 2004).
As noted by Schindler (1998) and Petersen and Hastings (2001), careful scaling
corrections must often be made even when the attempted extrapolation is simply from a
smaller habitat to a larger one of the same kind, located in the same geographic region,
and containing the same species. Seemingly contradictory results have been reported
even for investigations of community patterns thought to be scale-independent. For
instance, while productivity has been shown to be an important determinant of food chain
length in aquatic microbial communities (Kaunzinger and Morin 1998), it appears to be
unrelated to food chain length in lake systems (Post et al. 2000).
Recently, microcosm experiments with protists have been used to investigate the
phenomenon of inducible trophic polymorphisms, wherein phenotypic plasticity enables
organisms to alter their position in the food web and respond dynamically to changes in
resource availability and competitors (Kopp and Tolrian 2003; Price and Morin 2004;
Banerji and Morin 2009). In the case of the freshwater hymenostome ciliate

55
Tetrahymena vorax Kidder, exposure to chemical cues released by competing ciliates,
which include conspecifics, triggers a transformation from small (~ 60 µm), pyriform,
bacterivorous morphs, termed “microstomes,” to larger (~ 200 µm), ovoid, predatory
morphs, termed “macrostomes,” which can consume other ciliates (Kidder et al. 1941;
Smith-Somerville et al. 2000; Ryals et al. 2002). This ability to transform and
subsequently feed on competitors might not only enhance the fitness of T. vorax under a
variety of circumstances, but also give rise to trait-mediated indirect effects that would
influence the composition and stability of the community as a whole (Banerji and Morin
2009; Banerji and Morin in preparation). The fact that isogenic lines of T. vorax
differentiate into different morphs in response to high population densities of
conspecifics and/or heterospecifics indicates that transformation of microstomes into
macrostomes is a density-dependent response (Fig. 1).
The life-history of T. vorax in natural systems remains essentially unknown. All
lineages of T. vorax available in culture collections are descended from the collection
made by Kidder et al. (1940, see also Corliss 1952, 1953) from a small pond in the
vicinity of Providence, Rhode Island. T. vorax was also collected once from a stream in
Pennsylvania (Nanney et al. 1998). Other polymorphic Tetrahymena may enjoy less
restricted distributions (Ryals et al. 2002), but none appear to be as common or as widely
distributed as their smaller monomorphic congeners (Elliot 1973). Polymorphic
Tetrahymena species lack a drought-resistant resting stage and are consequently restricted
to permanent waters. Beyond this constraint, the reasons for the apparently limited
distribution of natural populations of T. vorax are unclear and raise fundamental
questions about the factors limiting the distribution of trophically plastic species.

56
Here I describe a study of T. vorax dynamics in outdoor experimental mesocosms
designed to uncover some of the possible causes of its limited distribution in nature. In
so doing, I also provide some insight as to whether or not the kinds of dynamics observed
in laboratory microcosms can be scaled up to larger, more complex, and more
realistically variable systems in the field.
Methods
Preparation of Protozoan and Bacterial Inoculants
I obtained T. vorax from the American Type Culture Collection (strain 30421)
and Colpidium kleini Foissner (Ciliatea: Hymenostomatida) from the Adelphia Plant
Science Research and Extension Center, Freehold, NJ 07728, USA. Species of the genus
Colpidium appear to have cosmopolitan distributions (Finlay 2002) and were found to be
naturally occurring prey / competitors of T. vorax in the original collections made by
Kidder et al. (1940). I cultured isogenic populations of T. vorax and C. kleini in 240-ml
glass jars containing 100 ml of complex organic medium. Loosely fastened screw caps
on the jars permitted some air circulation, while minimizing evaporation and
contamination. The complex organic culture medium used consisted of 0.37 g of
Carolina Biological Supply protozoan pellets (Carolina Biological Supply Company,
Burlington, NC 27215, USA) per L of filtered well water. After autoclaving and cooling,
I inoculated the medium with three species of bacteria (Serratia marcescens, Bacillus
subtilis, and Bacillus cereus) to standardize initial conditions and provide food for the
protozoa.
Construction of Outdoor Experimental Mesocosms

57
I modified twelve 19-L plastic buckets (Homer’s All-Purpose Bucket, The Home
Depot ®) to serve as my experimental units. I added 2 g of autoclaved dried alfalfa leaf,
14 L of well water, and 100 mL of bacterized alfalfa medium to each bucket. I covered
the buckets with 5-µm Nitex ® mesh glued to a removable plastic lid to prevent
contamination by larger aquatic organisms. The mesh size was small enough to exclude
the nauplii of cyclopoid copepods and enable me to maintain initial control over ciliate
species composition. The well water I used came from the same source used for the
medium of my laboratory cultures of T. vorax and C. kleini. The total volume in these
experimental units constituted a >140-fold increase from that of the jars used in my
microcosm experiments. The dilute alfalfa medium provided a nutrient source for the
bacteria, as well as some structural heterogeneity for the protozoa, that could be adjusted
to simulate the contributions of plant litter to natural pond systems. I placed the buckets
in a designated clearing located at the Hutcheson Memorial Forest of Rutgers University
(East Millstone, NJ, USA), arranging them with approximately 1 m of space between
them, with control and treatment replicates alternated to compensate for any potential
environmental gradients within the site (see Hurlbert 1984). Two days after filling and
positioning the buckets, I added a 2-day-old, 100-mL culture of T. vorax and a 2-day-old,
100-mL culture of C. kleini to each bucket.
Crustacean Zooplankton Treatment
There is an abundance of experimental and observational evidence implicating
cladocerans and copepods as important predators of the ciliates that persist in permanent
waters (Pace and Funke 1991; Burns and Gilbert 1993; Wickham 1995; Burns and
Schalenberg 1996; Adrian and Schneider-Olt 1999; Gaedke and Wickham 2004).

58
However, none of these studies address the very different problem of whether these
crustacean zooplankton actively exclude some ciliate taxa from otherwise favorable sites.
T. vorax in its macrostome state, for instance, could be more susceptible to grazing by
crustacean zooplankton due to its being more conspicuous than other, smaller co-
occurring ciliates. In addition, crustacean zooplankton could decrease the relative
abundance of macrostomes indirectly by reducing densities of all ciliates, and thereby
reducing the concentration of cues that induce macrostome formation in T. vorax.
After sampling to obtain baseline counts of the densities of T. vorax and C. kleini
in each bucket, I added 50 mL of unfiltered pond water from a local source to 6 of the 12
buckets (treatment replicates). The local source was a small permanent pond I had
chosen for the convenience of its proximity, containing cyclopoid copepods, cladocerans,
and other small aquatic organisms. To the remaining half of the buckets (controls), I
added pond water from the same source that had been filtered through 5-µm mesh to
remove the crustacean zooplankton, rotifers, and any large protozoa.
Sampling of Outdoor Experimental Mesocosms
I collected 7-mL samples from the surface and lower depths of each bucket (for a
combined total of 14 mL) in sterile Falcon® centrifuge tubes using a pair glass tubes that
I rinsed with hot water before and after each sampling interval. I counted copepods by
eye in the entire 14 mL sample, after confirming their identity via microscopy. I then
sub-sampled approximately 0.3 mL (measured by weight) from each of the 14 mL
samples using sterile glass pipettes to quantify the population densities of the protozoa. I
enumerated both macrostomes and microstomes of T. vorax and C. kleini. I also recorded

59
the presence (and, in some cases, density) of the other protozoa, algae, and macroscopic
zooplankton.
Results
Species Composition of Replicates after Initial Setup
The unfiltered pond water used for my treatment replicates introduced a number
of taxa to the treatment replicates. In addition, despite my use of the 5-µm mesh, all
replicates, including the control replicates, as of Day 26, were contaminated with large
densities of an alga of the genus Chlorella (mean average of 140.34 cells / mL) and a
species of microflagelate that I was unable to identify (mean average of 45.73 cells / mL).
As of Day 24, in treatment replicates Trt 1 and Trt 5 and in control replicate Con 4, I
observed hypotrichs of the genus Oxytricha (mean average of 10.84 cells / mL). During
the same period, in treatment replicate Trt 1 and control replicate Con 3, I observed
peritrichids of the genus Vorticella (mean average of 14.99 cells / mL).
Although I had observed species of Daphnia and other cladocerans in my source
pond, I did not observe any of these in my treatment replicates. The crustacean
zooplankton present in my treatment replicates consisted entirely of cyclopoid copepods,
the majority of which I could identify as being of the genus Acanthocyclops (either A.
vernalis or A. robustus), with two individuals (seen on Day 10 in Trt 6) possibly being of
the genus Diacyclops. One of the replicates with zooplankton added (Trt 1) also
contained eggs of the mosquito species Aedes albopictus (= Stegomyia albopicta Skuse),
which had presumably entered through my unfiltered pond water addition. These
emerged as larvae, developed into pupae, completed their metamorphosis into adults, and
then died between Days 17 and 26.

60
Effects of Zooplankton on Protist Community Dynamics
The population densities of T. vorax and C. kleini in treatment replicates were
significantly lower than in controls, which lacked large zooplankton (MANOVA, F3,8 =
10.83, p = 0.0034; Fig. 3). The appearance of the mosquitoes in Trt 1 did not appear to
change the overall patterns of abundance of T. vorax and C. kleini in that replicate, as
compared to the other treatment replicates. By Day 19, however, copepods were no
longer present in the replicate. In all of the treatment replicates, including Trt 1, protozoa
declined upon the addition of the unfiltered pond water containing macroscopic
zooplankton (Fig. 4). By contrast, without the zooplankton, T. vorax microstomes and C.
kleini maintained relatively constant population densities until the last three days of the
experiment (Fig. 5). Differences between the proportion of T. vorax macrostomes in the
presence and absence of crustacean zooplankton were not significant. In both cases, the
proportion of T. vorax macrostomes was significantly lower than what I had observed in
laboratory settings, across a range of productivities (ANOVA, F5,15 = 23.87, p < 0.0001;
see Banerji and Morin 2009). Follow-up contrast analysis of the means showed that
these differences were significant for each productivity-level compared to the outdoor
mesocosm settings.
Discussion
I am skeptical of there being a causal relationship between the presence of
mosquito larvae in Trt1 and the extirpation of copepods in that replicate. It is, for
example, highly unlikely that the copepods were consumed by the larvae. Apart from the
fact that I did not observe any such predation event in the course of my sampling,
copepods such as those I had observed prior to the emergence of the mosquito larvae

61
have been used as biological agents to kill mosquito larvae (Marten 1989, Marten and
Reid 2007). I can only speculate that other factors were responsible for the loss of
copepods in this replicate and that the timing was mainly coincidental. The steep decline
of the population densities of each of the protozoa in the control replicates during the last
three days of the experiment might be attributed to the fact that the water temperatures of
all of the replicates consistently exceeded 25 °C during this period.
Although T. vorax was not entirely extirpated from mesocosms in the presence of
crustacean zooplankton, the decline of T. vorax and C. kleini population densities
indicates that predation by crustacean zooplankton is still likely to be a factor in limiting
the distribution and abundance of T. vorax and other ciliates among natural aquatic
systems. This is not unexpected, as cladocerans and copepods have been shown to be
capable of affecting the abundance of protists in permanent ponds (Adrian and Schneider-
Olt 1999; Gaedke and Wickham 2004). Since the proportion of macrostomes in the T.
vorax population did not appear to differ between control and treatment replicates, it is
unlikely that crustacean zooplankton preferentially feed on macrostomes. It is uncertain
why the proportion of macrostomes in my outdoor mesocosms was smaller than what I
have observed in laboratory settings. The low density of ciliates and cues is a likely
cause, but I cannot rule out the possibility of the contents of the medium also having an
effect. It has been demonstrated, for example, that populations of T. vorax grown in
proteose peptone medium are unable to differentiate. Given that the dilute alfalfa
medium I used in this experiment resembled the environment of natural ponds more
closely than the protist pellet medium used for my laboratory cultures, it could be that the

62
expression of ITPs among protists raised in standard growth media is exaggerated,
relative to what it is in natural settings.
The results of my experiment show that community patterns may change
radically with the scale and complexity of experimental systems, but simultaneously
demonstrate the value of combining microcosm studies with field studies to uncover the
mechanisms underlying the dynamics of large, complex systems.
Acknowledgements
I thank Brian and Amy Clough for their aid in the setup of this experiment. I also
thank Kevin Aagaard, Jean Deo, Jack Siegrist, Maria Stanko, and Holly Vuong for their
helpful feedback. Support for this research came from the Rutgers School of
Environmental and Biological Sciences and the New Jersey Agricultural Experiment
Station, in the form of the Hutcheson Memorial Forest Summer Research Grant.

63
References
Adrian, R., and B. Schneider-Olt. 1999. Top-down effects of crustacean zooplankton on
pelagic microorganisms in a mesotrophic lake. Journal of Plankton Research
21(11): 2175-2190
Agrawal, A. A. 2001. Phenotypic plasticity in the interactions and evolution of species.
Science 294: 321-326
Altwegg, R. 2002. Predator-induced life-history plasticity under time constraints in pool
frogs. Ecology 83(9): 2542-2551
Banerji, A., and P. J. Morin. 2009. Phenotypic plasticity, intraguild predation and anti-
cannibal defences in an enigmatic polymorphic ciliate. Functional Ecology 23:
427-434
Burns, C. W., and J. J. Gilbert. 1993. Predation on ciliates by freshwater calanoid
copepods: rates of predation and relative vulnerabilities of prey. Freshwater
Biology 30: 377-393
Burns, C. W., and M. Schallenberg. 1996. Relative impacts of copepods, cladocerans and
nutrients on the microbial food web of a mesotrophic lake. Journal of Plankton
Research 18: 683-714
Cadotte, M. W., J. A. Drake, and T. Fukami. 2005. Constructing nature: laboratory
models as necessary tools for investigating complex ecological communities.
Advances in Ecological Research 37: 333-353
Carpenter, S. R. 1999. Microcosm experiments have limited relevance for community
and ecosystem ecology: Reply. Ecology 80: 1085-1088
Carpenter, S. R. 1996. Microcosm experiments have limited relevance for community
and ecosystem ecology. Ecology 77: 677-680
Corliss, J. O. 1952. Comparative studies on holotrichous ciliates in the Colpidium-
Glaucoma-Leucophrys-Tetrahymena group. I. General considerations and history
of strains in pure culture. Trans. American Microscopical Society 71: 159-184
Corliss, J. O. 1953. Comparative studies on holotrichous ciliates in the Colpidium-
Glaucoma-Leucophrys-Tetrahymena group. II. Morphology, life cycles and
systematic status of strains in pure culture. Parasitology 43:49-87
Drake, J. A., G. R. Huxel, C. L. Hewitt. 1996. Microcosms as models for generating and
testing community theory. Ecology 77(3): 670-677
Elliot, A. M. 1973. Life cycle and distribution of Tetrahymena. Pages 259-286 in A. M.
Elliot, editor. Biology of Tetrahymena. Dowden, Hutchinson, & Ross, Inc.,
Stroudsburg.
Finlay, B. J. 2002. Global dispersal of free-living microbial eukaryote species. Science
296: 1061-1063
Fraser, L. H., and P. Keddy. 1997. The role of experimental microcosms in ecological
research. Trends in Ecology and Evolution 12(12): 478-481
Gaedke, U., and S. A. Wickham. 2004. Ciliate dynamics in response to changing biotic
and abiotic conditions in a large, deep lake (L. Constance). Aquatic Microbial
Ecology 34: 247-261
Hargrove, W. W., and J. Pickering. 1992. Pseudoreplication: a sine qua non for regional
ecology. Landscape Ecology 6(4): 251-258

64
Holyoak, M., and S. P. Lawler. 2005. The contribution of laboratory experiments on
protists to understand population and metapopulation dynamics. Advances in
Ecological Research 37: 245-271
Hurlbert, S. H. 1984. Pseudoreplication and the design of ecological field experiments.
Ecological Monographs 54(2): 187-211
Jessup, C. M., R. Kassen, S. E. Forde, B. Kerr, A. Buckling, P. B. Rainey, B. J. M.
Bohannan. 2004. Big questions, small worlds: microbial model systems in
ecology. Trends in Ecology and Evolution 19(4): 189-197
Kaunzinger, C. M. K., and P. J. Morin. 1998. Productivity controls food-chain properties
in microbial communities. Nature 395: 495-497
Kidder, G. W., D. M. Lilly and C. L. Claff. 1941. Growth Studies on Ciliates. IV. The
influence of food on the structure and growth of Glaucoma vorax sp. nov.
Biological Bulletin 80(1): 9-23
Kopp, M., and R. Tollrian. 2003. Trophic size polyphenism in Lembadion bullinum: costs
and benefits of an inducible offense. Ecology 84: 641-651
Marten, G. G. 1989. A survey of cyclopoid copepods for control of Aedes albopictus
larvae. Bulletin of the Society for Vector Ecology 14(2): 232-236.
Marten, G. G., and J. W. Reid. 2007. Cyclopoid copepods. Reprinted from T. G. Floore,
editor, Biorational Control of Mosquitoes, American Mosquito Control
Association Bulletin No. 7.
Morin, P. J. 1999. Productivity, Intraguild Predation, and Population Dynamics in
Experimental Food Webs. Ecology 80(3): 752-760
Nanney, D. L., C. Park, R. Preparata, and E. M. Simon. 1998. Comparison of sequence
differences in a variable 23S rRNA domain among sets of cryptic species of
ciliated protozoa. Journal of Eukaryotic Microbiology 45: 91-100
Pace, M. L., and E. Funke. 1991. Regulation of planktonic microbial communities by
nutrients and herbivores. Ecology 72(3): 904-914
Petersen, J. E., and A. Hastings. 2001. Dimensional approaches to scaling experimental
ecosystems: designing mousetraps to catch elephants. Am. Nat. 157:324-333.
Price, J. E., and P. J. Morin. 2004. Colonization history determines alternate community
states in a food web of intraguild predators. Ecology 85: 1017-1028
Post, D. M., M. L. Pace, and N. G. Hairston Jr. 2000. Ecosystem size determines food
chain length in lakes. Nature 405: 1047-1049
Relyea, R. A. 2002. Costs of phenotypic plasticity. American Naturalist 159: 272-282
Ryals, P. E., H. E. Smith-Somerville, and H. E. Buhse. 2002. Phenotype switching in
polymorphic Tetrahymena: A single-cell Jekyll and Hyde. International review of
cytology 212: 209-238
Schindler, D. W. 1998. Whole-ecosystem experiments: replication versus realism: the
need for ecosystem-scale experiments. Ecosystems 1(4): 323-334
Smith-Somerville, H. E., J. K. Hardman, R. Timkovich, W. J. Ray, K. E. Rose, P. E.
Ryals, S. H. Gibbons, and H. E. Buhse Jr. 2000. A complex of iron and nucleic
acid catabolites is a signal that triggers differentiation in a freshwater protozoan.
PNAS 97(13): 7325-7330
Srivastava, D. S., J. Kolasa, J. Bengtsson, A. Gonzalez, S. P. Lawler, T. E. Miller, P.
Munguia, T. Romanuk, D. C. Schneider, and M. K. Trzcinski. 2004. Are natural

65
microcosms useful model systems for ecology? Trends in Ecology and Evolution
19(7): 379-384
Wickham, S. A. 1995. Trophic relations between cyclopoid copepods and ciliated
protists: complex interactions link the microbial and classic food webs.
Limnological Oceanography 40: 1173-1181
Williams, N. E. 1961. Polymorphism in Tetrahymena vorax. The Journal of Eukaryotic
Microbiology 8(4): 403-410

66
low density,
low competition
high density,
high competition
Colpidium:
interspecific
competitor
Microstome:
inducible
bacterivore
Macrostome:
induced
predator
Tailed
Microstome:
induced
bacterivore
Colpidium:
intraguild
prey
Resource = bacteriaResource = bacteria
60 μm
90 μm
< 3 μm
75 μm
75 μm
< 3 μm
200 μm
Fig. 1. Diagram of the inducible trophic polymorphism of T. vorax (reproduced from
Banerji and Morin 2009). Gray arrows represent developmental pathways. Black arrows
represent trophic links. Arrow size represents relative interaction strength.
Transformation of microstomes into macrostomes occurs in the presence of chemical
cues released by competitors for bacteria.

67
Fig. 2. Semi-quantitative measurements of the abundances of macroscopic zooplankton
in treatment replicates. Black hexagons represent the abundance of copepods
(Acanthocyclops spp.). Dark gray triangles represent that of mosquitoes (Aedes
albopictus). Note that mosquitoes were detected as larvae on Days 19 and 21, as pupae
on Day 24, and as (dead) adults on Day 26.

68
Copepods Present
no yes
Mean
Den
sit
y +
1 S
.E.M
. (n
o. / m
L)
0
20
40
60
80
100
T. vorax macrostomes
T. vorax microstomes
C. kleini
Fig. 3. Effects of copepods on protozoa. Black columns represent the mean population
density of macrostomes of T. vorax. Light gray columns represent the mean population
density of microstomes of T. vorax. Dark gray columns represent the mean population
density of C. kleini.

69
Fig. 4. Population dynamics of protozoa in treatment replicates (replicates containing
crustacean zooplankton). Large unfilled black squares represent abundances of
macrostomes of T. vorax. Small black squares represent abundances of microstomes of
T. vorax. Gray circles represent abundances of C. kleini.

70
Fig. 5. Population dynamics of protozoa in control replicates (replicates lacking
crustacean zooplankton). Symbols are as in Fig. 4.

71
CHAPTER 4
On the Strategy of Mounting an Indirect Offense: What If My Enemy Has
Allies, Too?
Summary
An indirect offense is the use of symbionts or recruited associates by consumers
to increase their rate of acquisition of prey or hosts. This versatile adaptive strategy is
employed by a wide array of consumers in various aquatic and terrestrial habitats. Here I
present a review of indirect offenses, the purpose of which being to draw attention to the
phenomenon and elucidate its significance within ecology and evolution. I explain the
utility of recognizing indirect offenses as a unique category of species interaction, present
examples to illustrate and clarify the modes and diversity of indirect offenses, and
highlight promising directions for future research.
Key-words: Functional response; indirect defense; indirect offense; parasite; parasitoid;
predator; symbiosis.

72
Introduction
In recent years, the phenomenon of species relying on symbionts or recruited
associates for an “indirect defense” against their natural enemies has been of growing
interest to ecologists (Poelman et al. 2008, Degenhardt 2009). Best-known are the cases
of terrestrial plants deterring herbivores using fungus-derived compounds (Huitu et al.
2008, Afkhami and Rudgers 2009, Hartley and Gange 2009) or higher-level consumers
(Dicke and Sabelis 1988, Turlings et al. 1995, Thaler 2002, Hountondji et al. 2006), but
indirect defenses are also employed by a variety of other taxa (McAtee 1944, Owens and
Goss-Custard 1976, Wheelwright et al.1997, Machado and Vital 2001, Oliver et al. 2003,
Hay et al. 2004, Damiani 2005, Fenton et al. 2011). Studies have shown that this use of
symbionts or recruited associates can drastically reduce the rate of acquisition of prey or
hosts by consumers (Damiani 2000, Kessler and Baldwin 2001, Sabelis et al. 2001,
Turlings and Wäckers 2004) and significantly influence the composition of ecological
communities (Poelman et al. 2008, Utsumi et al. 2010). What these studies have
generally failed to take into account, however, is that the use of symbionts and recruited
associates is not unique to species under threat. In the same way that a species might
make use of another species to protect itself from natural enemies, a species might also
make use of another species to enhance its ability to feed on prey or hosts. In other
words, not only are there indirect defenses in ecological communities, but also “indirect
offenses.”
The Utility of the Concept of an Indirect Offense
Indirect offenses occur when consumers make use of symbionts or recruited
associates to increase their rate of acquisition of prey or hosts. Many kinds of symbiosis

73
and recruited association can give rise to indirect offenses, including both resource-
exchange mutualisms (Hoeksema and Schwartz 2003, Holland et al. 2005) and parasite-
host interactions (Poulin 1994, Lafferty 1999). Since the term “indirect offenses” has
never before been used to categorize these interactions, it seems appropriate to briefly
address the question of how it would be meaningful to apply the term currently. Simply
stated, the term provides us with a conceptual framework. It implies, as it is meant to,
that these interactions are the logical counterpart to indirect defenses. Recognizing the
existence and common features of these interactions enables us to generalize across taxa
and make predictions regarding their consequences and dynamics. In turn, our ability to
generalize and make predictions allows for new avenues of academic and applied
research, such as the development of integrated pest management strategies that target the
symbionts and recruited associates of invasive species or that manipulate the symbionts
and recruited associates of biological control agents.
Examples of Indirect Offenses
Theory predicts that, while foraging, a consumer’s rate of acquisition of prey or
hosts – also known as its “functional response” – is constrained by two separate factors:
“search time” and “handling time” (Abrams 1982, Jeschke et al. 2002). Search time is
the amount of time that the consumer spends locating its prey or hosts. Handling time is
the amount of time that the consumer spends subduing and assimilating its prey or hosts.
In an indirect offense, symbionts or recruited associates may benefit the consumer by
reducing one or the other of these limiting factors. In this section, I present some of the
ways in which they go about this.
Indirect Offenses That Reduce Search Time

74
… By Revealing the Presence of Prey or Hosts
Many prey species rely on the cover of darkness to avoid detection by visual
predators (Nelson and Vance 1979, Rydell and Speakman 1995, Wcislo et al. 2004). In
response, many nocturnal and deep-sea visual predators have evolved specialized
adaptations for foraging under low-light conditions (Healy and Guilford 1990, Collin and
Partridge 1996). The loose-jawed dragonfish, Malacosteus niger, has evolved an
especially unique adaptation. Using the green-sulfur bacteria residing in its suborbital
photophores, the dragonfish is able to project far-red light. All other known animals of
the deep sea are blind to this wavelength, which enables the dragonfish to illuminate its
prey without giving its own presence away (Douglas et al. 2000, Herring and Cope
2005).
… By Locating Prey or Hosts
Typically, if two species rely on the same food source, one would expect their
interaction with each another to be mostly competitive. Interspecific cooperative
foraging associations that involve guiding behavior allow for a fascinating exception to
this rule (Morse 1977, Giraldeau 1984, Berner and Grubb 1985). One of the most
striking examples is the guiding behavior of the coral grouper, Plectropomus
pessuliferus, which often leads its fellow predator, the giant moray eel, Gymnothorax
javanicus, to the location of prey that have taken refuge in narrow crevices within a reef.
The eel, due to the shape of its body, is able to enter these crevices, but the grouper is not.
It is the grouper that recruits the eel in this association. The grouper signals the
eel to follow it by approaching the eel at the eel’s den and performing a ritualized set of
visual displays. This entails the grouper rapidly shaking its head directly in front of the

75
eel, a few centimeters away from the eel’s head. While doing so, the grouper keeps the
soft part of its dorsal fin erect and the bony part flat. Once the grouper has persuaded the
eel to leave the den and has led the grouper to the hiding place of the prey, the grouper
positions itself where it is best able to capture whatever prey the eel happens to flush out
without catching (Diamant and Shpigel 1985, Bshary et al. 2006).
… By Luring Prey or Hosts
For some consumers, the simplest way to reduce search time is to have their prey
or hosts come to them. The classic example of this kind of consumer is the bathypelagic
predatory female anglerfish, Melanocetus murrayi. The anglerfish uses the
bioluminescent, intracellular bacteria (Vibrionaceae) that reside in its light organ to lure
in its prey. It remains motionless with the exception of its light organ, which it wiggles
in order to mimic the movements of much smaller, bioluminescent organisms. Able to
discern only the wiggling light organ through the darkness, the anglerfish’s prey come
within range to investigate what they perceive as a potential meal. The moment they do,
the anglerfish is able to quickly swallow them whole (Hulet and Musil 1968).
Surprisingly, predators do not have to be larger than their prey to employ the
strategy of the anglerfish. In shallower waters, the squaloid shark, Isistius brasiliensis,
known also as the “cookiecutter shark,” uses its endosymbiotic bioluminescent bacteria to
camouflage all but a small portion of its body. Seeing it from below, larger pelagic
predators, such as tuna, mistake the shark for a very small, harmless prey. When these
larger predators attempt to consume the shark, the shark ambushes them and extracts
plugs of flesh from their bodies (Jones 1971, Widder 1998).

76
Another manifestation of the use of symbionts to lure prey or hosts is the
attraction of definitive hosts by parasites with complex life-cycles. This phenomenon has
been termed “parasite-increased trophic transmission” (Lafferty 1999) and involves
parasites modifying the behavior or physiology of their intermediate hosts in such a way
as to make them more vulnerable to capture and consumption. In many cases, it can be
readily distinguished from mere artifacts of nutrient and energy loss because of the nature
of the changes in the host (Poulin 2000, Brown et al. 2001). A wide range of parasites
make use of this strategy, including the tapeworm, Schistocephalus solidus. This
tapeworm manipulates its intermediate host, the threespine stickleback, Gasterosteus
aculeatus, so as to be able to infect its definitive host, the piscivorous common
kingfisher, Alcedo atthis. It dramatically reduces the melanin content of the stickleback’s
scales and darkens the stickleback’s eyes, while causing the stickleback to move slowly
and be less responsive to danger. The stickleback thereby becomes significantly more
noticeable from above to its visual predator and significantly easier to catch (Ness and
Foster 1999, Milinski 2006).
Indirect Offenses That Reduce Handling Time
… By Concealing the Presence of the Consumer
Just as prey species can use symbionts to avoid detection by their predators
(Gradstein and Equihua 1995), predators can use symbionts to avoid detection by their
prey. The Hawaiian bobtail squid, Euprymna scolopes, for example, uses the
bioluminescent bacteria in its mantle to create disruptive color patterns that break up its
silhouette from beneath and eliminate its shadow. This enables the squid to approach

77
prey such as red shrimp, Halocaridina rubra, without being seen (Travis 1996, Jones and
Nishiguchi 2004).
… By Enabling the Consumer to Metabolize Prey or Host Content
The presence of structural and chemical defenses can make feeding on prey or
hosts anything from unappealing to lethal for consumers (Simms and Fritz 1990, Jeschke
2006). Plants are especially well-equipped in terms of such defenses (Coley and Barone
1996). One of the ways in which herbivores and other consumers may respond to this is
by relying on specialized gut endosymbionts (Dyer 1989, Karban and Agrawal 2002).
The cigarette beetle, Lasioderma serricorne, for example, achieves this result using the
hydrolytic detoxifying enzymes of its symbiotic yeast (Dowd 1989, Dowd and Shen
1990, Dowd 1991).
It should be noted that a consumer’s inability to digest prey or host content is not
always a matter of the content being defensive in nature. In some cases, it is a matter of
the constraints set by the consumer’s own evolutionary history. The carnivorous plants,
Roridula gorgonias and Roridula dentata, for example, despite being fully capable of
trapping insects on their sticky leaves, lack the digestive enzymes necessary to
metabolize the insects. In lieu of the enzymes, these plants have developed a mutualistic
symbiosis with the predatory hemipteran, Pameridea roridulae. The hemipteran, which
is able to move freely across the plants’ sticky leaves, gains easy access to the already-
captured insects and enables the plants to assimilate nutrients from the insects via direct
absorption of its excrement (Anderson and Midgley 2003, Anderson 2005).
… By Poisoning Prey or Hosts

78
Many consumers use venom to immobilize and/or kill their prey or hosts
(Whittaker and Feeny 1971). Some, however, are able to use pathogens or symbiont-
derived toxins to envenomate their prey or hosts, in lieu of endogenously produced
toxins. Soil-dwelling, predatory nematodes of the genus Heterorhabditis, for example,
make use of Photorhabdus luminescens, a bioluminescent bacterial symbiont they carry
within their guts. The tiny nematodes burrow inside of their larval insect prey and release
P. luminescens into the insects’ circulatory system, causing a “raging septicemia” which
kills the larvae within 24 hours. As P. luminescens begins to decompose the dead larvae
and rapidly proliferate, the nematodes complete their life cycle by laying eggs. Newly
hatched nematodes of the next generation will later feed on a portion of the P.
luminescens colony for 10 to 14 days and then assimilate the remainder as endosymbionts
(Mlot 1997, Fenton et al. 2000).
Larger-scale predators can also rely on microbial pathogens for venom. On land,
the Komodo dragon, Varanus komodoensis, compounds the lethal effects of its
venomous saliva with the use of up to 57 different species of virulent bacteria which it
obtains by feeding on carrion and houses in its gingival tissue (Montgomery et al. 2002,
Fry et al. 2006). In rocky intertidal zones, the blue-ringed octopus, Octopus maculosus,
paralyzes and kills its prey using the neurotoxins it obtains from 16 of the 22 bacterial
strains that reside in its posterior salivary glands and intestines (Sheumack et al. 1978,
Hwang et al. 1989, White 1998, Bonnet 1999).
Certain parasitoids use a similar strategy to subdue their hosts. The wasp,
Campoletis sonorensis (Hymenoptera: Ichneumonidae), for example, makes use of its co-
evolved endosymbiotic virus (Polydnaviridae). The virus replicates in the ovary of the

79
wasp and is injected along with the wasp’s egg into a host caterpillar during oviposition.
There, it acts to suppress the caterpillar’s growth and encapsulating immune response to
the wasp’s egg, allowing the egg to successfully develop inside the caterpillar (Edson et
al. 1981, Theilmann and Summers 1988, Whitfield 1990, Shiga 2005).
… By Transporting the Consumers to Prey or Hosts
Phoresy is the symbiotic transportation of one organism by another (Binns 1982).
Consumers may make use of phoretic partners to acquire prey and hosts that are more
mobile than they are or are otherwise difficult to reach. One such consumer is the
mesostigmatid mite, Poecilochirus necrophori, which uses burying beetles of the genus
Necrophorus for transportation to the location of fly maggots and carrion on which to
feed. In return, the burying beetles may obtain the benefit of increased reproductive
success from the mite’s removal of their competitors (Springett 1968, Wilson 1983).
Pathogens commonly make use of phoretic partners, as well. In their case,
however, we refer to the partners as “vectors.” The mechanisms of recruitment of vectors
by pathogens are very similar to the attraction of primary hosts by parasites, in that they
often involve dramatic examples of host-manipulation. The fungus responsible for
Mummy-Berry Disease in blueberries and huckleberries, Monilinia vaccinii-corymbosi,
relies on pollinators for transportation from the leaves of its hosts to the flowers, where it
can sexually reproduce to make spores, which the pollinators then spread to other host
plants. To recruit the pollinators, the fungus causes the wilting leaves of its host to
secrete sugars, exude sweet odors, and reflect ultraviolet light in a manner similar to that
of nectar-rich flowers. While consuming the sugars, pollinators that have been attracted

80
to these imitation flowers take on some of the fungus and later deposit it into real flowers
as they make their normal feeding rounds (Ingvarsson and Lundberg 1993, Kaiser 2006).
Indirect Offenses versus Alternative Strategies for the Acquisition of Exogenous
Offensive Traits
Symbioses and recruited associations are favored where the benefits of
association outweigh the costs. Often included among the costs of association are
significant investments of nutrients, energy, and time (Dicke and Sabelis 1988, Thomas
et al. 1998, Siemens et al. 2002, van Rijn et al. 2002, Huntzinger et al. 2004, Wäckers
and Bonifay 2004). In some cases, the costs of association may also include conflicts
between partner-provided services and endogenous adaptations (Eisner et al. 1998) or
increases in the risk of attracting natural enemies and partners that are unsuitable
(Anderson and Midgley 2002, van Rijn et al. 2002, Horiuchi et al. 2003, Mouritsen and
Poulin 2003, Shenoy and Borges 2010). Given such costs, it would be reasonable to
assume that indirect offenses evolve when the alternative of developing and maintaining
endogenous offensive traits is still more costly. However, even in a situation where there
is strong selection for these traits and developing these traits endogenously is a
physiological impossibility, forming an indirect offense association is not the only
recourse. Besides indirect offenses, there are at least two other strategies for the
acquisition of exogenous offensive traits: opportunistic association and annexation. By
“opportunistic association,” I mean interspecific relationships such as scavengry (Tenney
1877, Farner and Kezer 1953), kleptoparasitism (Hamilton 2002), and aggressive
mimicry (Sazima 2002). By “annexation,” I mean the assimilation of exogenous traits

81
through collection (Levey et al. 2004) or consumption (Rothschild and Edgar 1978). In
this section, I expand on and clarify the distinctions between these alternative strategies.
Acquiring Exogenous Offensive Traits via Opportunistic Association
The ocellated antbird, Phaenostictus mcleannani, is an obligate associate of army
ants. It monitors the ants’ activity and follows them as they perform their raids, so that it
can capitalize on the swarms of hidden insect prey that the ants tend to flush out (Willis
and Oniki 1978, Swartz 2001, Chaves-Campos 2010). The antbird benefits from its
association with the ants in much the same way that other consumers benefit from
mounting an indirect offense. Yet, to obtain this benefit, the antbird neither has to recruit
the ants nor be recruited by the ants. It simply takes advantage of what the ants must do
irrespective of its presence or absence, scavenging off of them in a way that may actually
reduce their success rate in capturing prey (Wrege et al. 2005).
If being an opportunist is inherently less costly than initiating and maintaining an
indirect offense association, why, then, do species such as the coral grouper,
Plectropomus pessuliferus, invest in the latter (Diamant and Shpigel 1985, Bshary et al.
2006)? The likely scenario is that species such as the grouper, in reality, have more to
gain from recruiting and more to lose from scavenging. Unlike army ants, the moray eels
that the groupers rely on to flush out their hidden prey do not perform raids during the
grouper’s period of activity. Furthermore, if properly positioned, the eels are just as
capable as the grouper of chasing after the prey that flee vertically when flushed. As a
result, if the grouper were to simply follow the eel and scavenge off of it, rather than
recruit it, the grouper would be more likely to lose hidden prey to the eel when foraging

82
with the eel, as well as miss the opportunity to capture prey during its own optimal
foraging period.
Acquiring Exogenous Offensive Traits via Annexation
To infiltrate a colony of aphids and avoid detection by the ants protecting this
colony, antlions, the predatory larvae of the green lacewing, Chrysopa slossonae,
camouflage themselves using the waxy remains of aphids they have already captured
(Caltagirone 1999). Similarly, predatory nudibranchs, such as Aeolidia papillosa, can
assimilate the stinging cells of their sea anemone prey and use them not only as a defense
against their own predators, but also as a tool for capturing and killing additional prey
(Greenwood and Mariscal 1984, Östman 1997, Edmunds 2009). Considering the
previously mentioned potential costs of association, what incentive would a species
capable of annexation have for mounting an indirect offense? There are at least two that
are apparent: (1) reduction of the handling time necessary to obtain the traits of another
species; and (2) renewability of the traits upon assimilation. Species that acquire
exogenous traits via annexation may be unable to replenish their supply of the traits when
necessary (Williams 2010), due to the absence or inaccessibility of their suppliers, for
which they themselves may be responsible. By contrast, species that form indirect
offense associations have better assurance of the presence of their suppliers, due to their
symbiosis and/or recruiting investments. In addition, species that form indirect offense
associations can, in some cases, also manipulate the level of production of the traits they
obtain from their associates to suit their short-term needs.
Future Research Questions

83
The paucity of attention given to indirect offenses thus far leaves many openings
for potential exploration and research. There is still much to be learned, for instance,
with regard to how indirect offenses have evolved and might continue to evolve, what
consequences indirect offenses give rise to, and what undocumented forms of indirect
offenses may be present in ecological communities. In this section, I highlight a few key
examples of these promising future directions.
Determining How Indirect Offenses That Involve Symbiosis Differ in Terms of Their
Evolution from Indirect Offenses That Involve Recruited Association
Interspecific associations that do not involve one partner living within, on top of,
or beside the other can often be so tightly co-evolved that researchers might still deem it
appropriate to refer to them as symbioses (Sapp 2004, Cimino et al. 2010). This is
certainly true for many of the cases of recruited association that give rise to indirect
offenses. Yet, differences between these types of associations and ones in which the
participants function all together as a single biological unit (Dyer 1989) can have
meaningful effects on how the associations persist. The former may be more easily
disrupted, for example, by cascading trait-mediation (Liere and Larson 2010) and
fluctuating abiotic conditions (Peterson 1995, Kersch and Fonseca 2005) than the latter,
even when the latter is highly exploitative rather than reciprocally beneficial (Law and
Dieckmann 1998, Kiers and van der Heijden 2006). Several explanations have been put
forth regarding why certain taxa have evolved intricate – and perhaps even obligate –
symbioses, while other taxa – even when closely related – have not (Keeler 1981, Daida
et al. 1996, Stadler and Dixon 1999, Kramer et al. 2009, Schemske et al. 2009). How
well any of these explanations apply to indirect offense associations remains to be seen.

84
Exploring the Overlap between Indirect Offenses and Indirect Defenses
I refer earlier to indirect offenses as being the logical counterpart to indirect
defenses and depict them throughout this manuscript as essentially being analogous in
form, but diametrically opposed in purpose. This portrayal, though somewhat expressive
of how indirect offenses and defenses factor into consumer-resource dynamics, provides
little insight into the actual biological relatedness of these strategies. The relationship
between indirect offenses and defenses in general is often complex and can have
intriguing ecological and evolutionary significance. Indirect offenses can, for instance,
often be coupled with indirect defenses. Consider, for example, the case of the cassava
plant, Manihot esculenta. In response to herbivory by the green mite, Mononychellus
tanajoa, the plant releases a specific blend of volatile emissions that triggers infective
sporulation in the mite-pathogenic fungus, Neozygites tanajoae (Hountondji et al. 2006).
In this situation, the plant relies on the fungus for protection from the mite – meaning that
it is employing an indirect defense. Meanwhile, it simultaneously enhances the fungus’
ability to acquire new hosts – meaning that it is providing an indirect offense. Such
potential overlap between indirect offenses and indirect defenses leads to some
interesting questions. For example, how might we assess the relative importance of one
or the other in a situation like this?
Consider, also, the association between the red-billed oxpecker, Buphagus
erythrorhynchus, and the large mammals upon which it forages. This relationship is
obligate for the oxpecker and has, in classical literature, been cited as a mutualism – the
oxpecker reducing the mammals’ parasite loads by feeding. Tolerance of the plucking,
pecking, and scissoring behavior of the oxpecker foraging on their backs (Samish and

85
Rehacek 1999, Plantan et al. 2009), in other words, allows the mammals to obtain an
indirect defense against their ectoparasites. However, it has been demonstrated more
recently that the relationship between the oxpecker and the large mammals may be more
of a parasitism than a mutualism. Weeks (2000), for instance, reports that the exclusion
of oxpeckers from cattle in Zimbabwe has no significant effect on the adult tick load of
the cattle and that oxpeckers significantly prolong the healing time of wounds in cattle. If
open wounds in large mammals are attractive to other ectoparasites besides ticks, such as
flesh flies (Stevens et al. 2006), prolonging the healing time of wounds – and perhaps
even creating fresh wounds – could be an effective way for the oxpecker to acquire a
larger or steadier food supply. Employing this strategy would amount to exploiting the
traits of its mammalian hosts for an indirect offense. What factors might cause the
association to shift from being an indirect defense to being an indirect offense, or vice
versa?
Examining the Use of Symbionts and Recruited Associates in Interference Competition
Species in competition can make use of symbionts and recruited associates to
reduce the foraging ability, survival, or reproduction of their competitors. This
phenomenon constitutes a novel form of interference (Park 1962) and could also be
considered a “competitor” indirect offense. It has been observed in plants (Janzen 1966,
Janzen 1969, Schupp 1986, Agrawal 1998, Mattner and Parbery 2001, Mangla et al.
2008), insects (Yan and Stevens 1995, Yan et al. 1998, Currie et al. 2003, Little and
Currie 2007), free-living ciliates (Preer et al. 1953, Preer et al. 1974), and food-borne
microbial pathogens (Stecher et al. 2007, Brown et al. 2008). There is even some
evidence of its occurrence in mammals (Schmitz and Nudds 1994). What are the

86
consequences of this phenomenon in terms of population dynamics and community
structure? Do they fit the patterns we would expect to see with other trait-mediated
indirect interactions between competitors (Wootton 1994, Peacor and Werner 2001), such
as apparent competition (Holt 1977, Bonsall and Hassell 1997, Chaneton and Bonsall
2000)?
Conclusion
Indirect offenses are widespread, captivating, and diverse. Their occurrence
provides unique insights into the roles of symbiosis and solicited association in shaping
community structure and dynamics. There is still much to be learned with regard to how
indirect offenses have evolved and might continue to evolve, what consequences indirect
offenses give rise to, and what undiscovered forms of indirect offenses may occur in
natural systems. Although indirect offenses offer an enormous potential for promoting
interdisciplinary collaboration and fostering new ideas, their significance as a versatile
and general adaptive strategy has gone virtually unnoticed in the field of biology. It is
my hope that the contents of this review will draw attention to the phenomenon of
indirect offenses and stimulate further investigation into its mechanisms and potential
broader impacts in ecology and evolution.
Acknowledgements
I gratefully acknowledge Peter Morin, Jennifer Rudgers, Jeremy Fox, Nina
Fefferman, Michael Sukhdeo, Marilyn Scott, Robert Paine, Paul Craze, Mathew Leibold,
John Maron, Mark McPeek, Rebecca Jordan, Cesar Rodriguez-Saona, Michael Pace,
Timothy Casey, Peter Smouse, Jason Grabowsky, Deborah Goldberg, Robert Raguso,
James D. Thomson, John J. Wiens, Bradley Hillman, Claus Holzapfel, Michael May,

87
Zsofia Szendrei, Carolyn Norin, Ari Novy, Kristen Ross, my labmates (Kevin Aagaard,
Jean Deo, Jack Siegrist, Maria Stanko, and Holly Vuong), and a very long list of other
friends and colleagues for their helpful comments and invaluable insights. Support for
this review was provided by the Rutgers School of Environmental and Biological
Sciences and the New Jersey Agricultural Experiment Station.
References
Abrams, P. A. 1982. Functional responses of optimal foragers. The American Naturalist
120(3): 382-390.
Abrams, P. A., B. A. Menge, G. G. Mittelbach, D. A. Spiller, and P. Yodzis. 1996. The
role of indirect effects in food webs. Pages 371-395 in G. A. Polis and K. O.
Winemiller, editors. Food webs: integration of patterns and dynamics. Chapman
and Hall, New York.
Afkhami, M. A., and J. A. Rudgers. 2009. Endophyte-mediated resistance to herbivores
depends on herbivore identity in the wild grass, Festuca subverticillata.
Environmental Entomology 38: 1086-1095.
Alizon, S., and M. van Baalen. 2008. Transmission-virulence trade-offs in vector-borne
diseases. Theoretical Population Biology 74: 6-15.
Anderson, B. 2005. Adaptations to foliar absorption of faeces: a pathway in plant
carnivory. Annals of Botany 95: 757-761.
Anderson, B., and J. Midgley. 2002. It takes two to tango but three is a tangle: mutualists
and cheaters on the carnivorous plant Roridula. Oecologia 132(3): 369-373.
Anderson, B., and J. J. Midgley. 2003. Digestive mutualism, an alternate pathway in
plant carnivory. Oikos 102: 221-224.
Bascompte, J., P. Jordano, and J. M. Olesen. 2006. Asymmetric coevolutionary networks
facilitate biodiversity maintenance. Science 312: 431-433.
Belliure, B., A. Janssen, P. C. Maris, D. Peters, and M. W. Sabelis. 2005. Herbivore
arthropods benefit from vectoring plant viruses. Ecology 8: 70-79.
Berner, T. O., and T. C. Grubb, Jr. 1985. An experimental analysis of mixed-species
flocking in birds of deciduous woodland. Ecology 66(4): 1229-1236.
Bertness, M. D., and G. H. Leonard. 1997. The role of positive interactions in
communities: lessons from intertidal habitats. Ecology 78: 1976-1989.
Bonnet, M. S. 1999. The toxicology of Octopus maculosa: The blue-ringed octopus. The
British Homeopathic Journal 88(4): 166-171.
Bonsall, M. B., and M. P. Hassell. 1997. Apparent competition structures ecological
assemblages. Nature 388: 371-373.
Boucher, D. H., S. James, and K. H. Keeler. 1982. The ecology of mutualism. Annual
Review of Ecology and Systematics 13: 315-347.
Brasier, C. M. 1991. Ophiostoma novo-ulmi sp. nov., causative agent of current Dutch
elm disease pandemics. Mycopathologia 115: 151-161.

88
Bronstein, J. L. 1994. Our current understanding of mutualism. Quarterly Review of
Biology 69: 31-51.
Brooks, W. R., and R. N. Mariscal. 1985. Protection of the hermit crab Pagurus
pollicaris Say from predators by hydroid-colonized shells. Journal of
Experimental Marine Biology and Ecology 87: 111-118.
Brown, S. P., G. Loot, B. T. Grenfell, and J. F. Guégan. 2001. Host manipulation by
Ligula intestinalis: accident or adaptation? Parasitology 123: 519-529.
Brown, S. P., L. Le Chat, and F. Taddei. 2008. Evolution of virulence: triggering host
inflammation allows invading pathogens to exclude competitors. Ecology Letters
11: 44-51.
Bruno, J. F., J. J. Stachowicz, and M. D. Bertness. 2003. Inclusion of facilitation into
ecological theory. Trends in Ecology & Evolution 18: 119-125.
Bshary, R., A. Hohner, K. Ait-el-Djoudi, and H. Fricke. 2006. Interspecific
communicative and coordinated hunting between groupers and giant moray eels
in the Red Sea. PLoS Biology 4: e431.
Caltagirone, L. E. 1999. Adaptations of insects to modes of life: specialized adaptations
of predatory insects. Pages 211-230 in C. B. Huffaker and A. P. Gutierrez,
editors. Ecological Entomology. John Wiley & Sons, Inc., New York.
Chaneton, E. J., and M. B. Bonsall. 2000. Enemy-mediated apparent competition:
empirical patterns and evidence. Oikos 88(2): 380-394.
Chaves-Campos, J. 2010. Ant colony tracking in the obligate army ant-following antbird
Phaenostictus mcleannani. Journal of Ornithology 152(2): 497-504.
Cimino, G., S. De Stefano, S. De Rosa, G. Sodano, G. Villani. 2010. Novel metabolites
from some predator-prey pairs. Bulletin des Sociétés Chimiques Belges 89(12):
1069-1073.
Coley, P. D., and J. A. Barone. 1996. Herbivory and plant defenses in tropical forests.
Annual Review of Ecology and Systematics 27: 305-335.
Collin, S. P., and J. C. Partridge. 1996. Retinal specializations in the eyes of deep-sea
teleosts. Journal of Fish Biology 49: 157-174.
Currie, C. R., B. Wong, A. E. Stuart, T. R. Schultz, S. A. Rehner, U. G. Mueller, G-H.
Sung, J. W. Spatafora, and N. A. Straus. 2003. Ancient tripartite coevolution in
the attine ant-microbe symbiosis. Science 299(5605): 386-388.
Daida, J. M., C. S. Grasso, S. A. Stanhope, and S. J. Ross. 1996. Symbionticism and
Complex Adaptive Systems I: Implications of having symbiosis occur in nature.
Evolutionary Programming V: Proceedings of the Fifth Annual Conference on
Evolutionary Programming, Cambridge, MA.
Damiani, C. C. 2000. The relative importance of direct and indirect effects in a hermit
crab-hydroid symbiosis. Dissertation. Duke University, Durham, North Carolina,
USA.
Damiani, C. C. 2005. Integrating direct effects and trait-mediated indirect effects using a
projection matrix model. Ecology 86(8): 2068-2074.
Degenhardt, J. 2009. Indirect defense responses to herbivory in grasses. Plant Physiology
149: 96-102.
deMazancourt, C., M. Loreau, and U. Dieckmann. 2001. Can the evolution of plant
defense lead to plant-herbivore mutualism? American Naturalist 158: 109-123.

89
Diamant, A., and M. Shpigel. 1985. Interspecific feeding associations of groupers
(Teleostei: Serranidae) with octopuses and moray eels in the Gulf of Eilat
(Agaba). Environmental Biology of Fishes 13: 153-159.
Dick, J. T. A., I. Montgomery, and R. W. Elwood. 1993. Replacement of the indigenous
amphipod Gammarus duebeni celticus by the introduced G. pulex: differential
cannibalism and mutual predation. Journal of Animal Ecology 62: 79-88.
Dicke, M. 1999. Evolution of induced indirect defense of plants. Pages 62-88 in R.
Tollrian and C. D. Harvell, editors. The ecology and evolution of inducible
defenses. Princeton University Press.
Dicke, M., and M. W. Sabelis. 1988. How plants obtain predatory mites as bodyguards.
Netherlands Journal of Zoology 38: 148-165.
Douglas, R. H., C. W. Mullineaux, and J. C. Partridge. 2000. Long-wave sensitivity in
deep-sea stomiid dragonfish with far-red bioluminescence: evidence for a dietary
origin of the chlorophyll-derived retinal photosensitizer of Malacosteus niger.
Proceedings of the Royal Society of London. Philosophical Transactions:
Biological Sciences 355(1401): 1269-1272.
Dowd, P. F. 1989. In situ production of hydrolytic detoxifying enzymes by symbiotic
yeasts in the cigarette beetle (Coleoptera: Anobiidae). Journal of Economic
Entomology 82(2): 296-400.
Dowd, P. F., and S. K. Shen. 1990. The contribution of symbiotic yeast to toxin
resistance of the cigarette beetle (Lasioderma serricorne). Entomologia
Experimentalis et Applicata 56(3): 241-248.
Dowd, P. F. 1991. Symbiont-mediated detoxification in insect herbivores. Pages 411-440
in P. Barbosa, V. A. Krischik, and C. Jones, editors. Microbial Mediation of
Plant-Herbivore Interactions. Wiley & Sons, Inc., New York.
Dyer, B. D. 1989. Symbiosis and organismal boundaries. Integrative and Comparative
Biology 29(3): 1085-1093.
Edmunds, M. 2009. Do nematocysts sequestered by aeolid nudibranchs deter predators? -
a background to the debate. Journal of Molluscan Studies 75(2): 203-205.
Edson, K. M., S. B. Vinson, D. B. Stoltz, and M. D. Summers. 1981. Virus in a parasitoid
wasp: suppression of the cellular immune response in the parasitoid's host.
Science 211: 582-583.
Eigenbrode, S. D., H. Ding, P. Shiel, and P. H. Berger. 2002. Volatiles from potato plants
infected with potato leafroll virus attract and arrest the virus vector, Myzus
persicae (Homoptera: Aphididae). Proceedings of the Royal Society of London.
Series B, Biological Sciences 269(1490): 455-460.
Eisner, T., M. Eisner, and E. R. Hoebeke. 1998. When defense backfires: detrimental
effect of a plant's protective trichomes on an insect beneficial to the plant.
Proceedings of the National Academy of Sciences of the United States of America
95(8): 4410-4414.
Farner, D. S., and J. Kezer. 1953. Notes on the amphibians and reptiles of Crater Lake
National Park. American Midland Naturalist 50(2): 448-462.
Fenton, A., L. Magoolagan, Z. Kennedy, and K. A. Spencer. 2011. Parasite-induced
warning coloration: a novel form of host manipulation. Animal Behavior 81(2):
417-422.

90
Fry, B. G., N. Vidal, J. A. Norman, F. J. Vonk, H. Scheib, S. F. R. Ramjan, S. Kuruppu,
K. Fung, S. Blair Hedges, M. K. Richardson, W. C. Hodgson, V. Ignjatovic, R.
Summerhayes, and E. Kochva. 2006. Early evolution of the venom system in
lizards and snakes. Nature 439(7076): 584-588.
Giraldeau, L.-A. 1984. Group foraging: the skill pool effect and frequency-dependent
learning. The American Naturalist 124: 72-79.
Gradstein, S. R., and C. Equihua. 1995. An epizoic bryophyte and algae growing on the
lizard Corythophanes cristatus in Mexican Rain Forest. Biotropica 27(2): 265-
268.
Greenwood, P. G., and R. N. Mariscal. 1984. Immature nematocyst incorporation by the
aeolid nudibranch Spurilla neapolitana. Marine Biology 80: 35-38.
Hamilton, I. M. 2002. Kleptoparasitism and the distribution of unequal competitors.
Behavioral Ecology 13(2): 260-267.
Hartley, S. E., and A. C. Gange. 2009. Impacts of plant symbiotic fungi on insect
herbivores: mutualism in a multitrophic context. Annual Review of Entomology
54: 323-342.
Hartnett, D. C., B. A. D. Hetrick, G. W. T. Wilson, and D. J. Gibson. 1993. Mycorrhizal
influence on intra- and interspecific neighbour interactions among co-occurring
prairie grasses. Journal of Ecology 81: 787-795.
Hay, M. E., J. D. Parker, D. E. Burkepile, C. C. Caudill, A. E. Wilson, Z. P. Hallinan, and
A. D. Chequer. 2004. Mutualisms and aquatic community structure: the enemy of
my enemy is my friend. Annual Review of Ecology, Evolution and Systematics
35: 175-197.
Healy, S., and T. Guilford. 1990. Olfactory-bulb size and nocturnality in birds. Evolution
44(2): 339-346.
Herring, P., and C. Cope. 2005. Red bioluminescence in fishes: on the suborbital
photophores of Malacosteus, Pachystomias and Aristostomias. Marine Biology
148(2): 383-394.
Hoeksema, J. D., and M. W. Schwartz. 2003. Expanding comparative-advantage
biological market models: contingency of mutualism on partner's resource
requirements and acquisition trade-offs. Proceedings of the Royal Society of
London. Series B, Biological Sciences 270(1518): 913-919.
Holland, J. N., J. H. Ness, A. Boyle, and J. Bronstein. 2005. Mutualisms as consumer-
resource interactions. Pages 17-33 in P. Barbosa and I. Castellanos, editors.
Ecology of Predator-Prey Interactions. Oxford University Press, Inc.
Holt, R. D. 1977. Predation, apparent competition, and the structure of prey communities.
Theoretical Population Biology 12: 197-229.
Holt, R. D., and G. A. Polis. 1997. A theoretical framework for intraguild predation.
American Naturalist 149: 745-764.
Hountondji, F. C., R. Hanna, and M. W. Sabelis. 2006. Does methyl salicylate, a
component of herbivore-induced plant odour, promote sporulation of the mite-
pathogenic fungus Neozygites tanajoae? Experimental and Applied Acarology 39:
63-74.
Horiuchi, J-I, G-I Arimura, R. Ozawa, T. Shimoda, M. Dicke, J. Takabayashi, and T.
Nishioka. 2003. Lima beans exposed to herbivore-induced conspecific plant

91
volatiles attract herbivores in addition to carnivores. Applied Entomology and
Zoology 38(3): 365-368.
Huitu, O., M. Helander, P. Lehtonen, and K. Saikkonen. 2008. Consumption of grass
endophytes alters the ultraviolet spectrum of vole urine. Oecologia 156(2): 333-
340.
Hulet, W. H., and G. Musil. 1968. Intracellular bacteria in the light organ of the deep sea
angler fish, Melanocetus murrayi. Copeia 1968: 506-512.
Huntzinger, M., R. Karban, T. P. Young, and T. M. Palmer. 2004. Relaxation of induced
indirect defenses of acacias following exclusion of mammalian herbivores.
Ecology 85: 609-614.
Hwang, D. F., O. Arakawa, T. Saito, T. Noguchi, U. Simidu, K. Tsukamoto, Y. Shida,
and K. Hashimoto. 1989. Tetrodotoxin-producing bacteria from the blue-ringed
octopus Octopus maculosus. Marine Biology 100(3): 327-332.
Ingvarsson, P. K., and S. Lundberg. 1993. The effect of a vector-borne disease on the
dynamics of natural plant populations: a model for Ustilago violacea infection of
Lychnis viscaria. Journal of Ecology 81: 263-270.
Jeschke, J. M., M. Kopp, and R. Tollrian. 2002. Predator functional responses:
discriminating between handling and digesting prey. Ecological Monographs 72:
95-112.
Jeschke, J. M. 2006. Density-dependent effects of prey defenses and predator offenses.
Journal of Theoretical Biology 242(4): 900-907.
Jones, E. C. 1971. Isistius brasiliensis, a squaloid shark, the probable cause of crater
wounds on fishes and cetaceans. Fisheries Bulletin 69(4): 791-798.
Jones, B. W., and M. K. Nishiguchi. 2004. Counterillumination in the Hawaiian bobtail
squid, Euprymna scolopes Berry (Mollusca: Cephalopoda). Marine Biology 144:
1151-1155.
Kaiser, R. 2006. Flowers and Fungi Use Scents to Mimic Each Other. Science 311(5762):
806-807.
Karban, R., and A. A. Agrawal. 2002. Herbivore offense. Annual Review of Ecology and
Systematics 33: 641-664.
Karban, R., D. Hougen-Eitzmann, and G. English-Loeb. 1994. Predator-mediated
apparent competition between two herbivores that feed on grapevines. Oecologia
97: 508-511.
Kareiva, P. M., and M. D. Bertness. 1997. Re-examining the role of positive interactions
in communities. Ecology 78: 1945-1945.
Kaya, H. K. 1982. Parasites and predators as vectors of insect diseases. Pages 39-40 in
Proceedings of the IIIrd International Colloquium on Invertebrate Pathology and
XVth Annual Meeting of the Society for Invertebrate Pathology, Brighton, UK.
Keeler, K. H. 1981. A model of selection for facultative nonsymbiotic mutualism. The
American Naturalist 118(4): 488-498.
Kersch, M. F., and C. R. Fonseca. 2005. Abiotic factors and the conditional outcome of
an ant-plant mutualism. Ecology 86: 2117-2126.
Kessler, A., and I. T. Baldwin. 2001. Defensive function of herbivore-induced plant
volatile emissions in nature. Science 291(5511): 2141-2144.

92
Kiers, E. T., and M. G. A. van der Heijden. Mutualistic stability in the arbuscular
mycorrhizal symbiosis: exploring hypotheses of evolutionary cooperation.
Ecology 87: 1627-1636.
Kramer, A., J. L. Van Tassell, and R. A. Patzner. 2009. A comparative study of two goby
shrimp associations in the Caribbean Sea. Biomedical and Life Sciences 49(3):
137-141.
Lafferty, K. D. 1999. The evolution of trophic transmission. Parasitology Today 15(3):
111-115.
Law, R., and U. Dieckmann. 1998. Symbiosis through exploitation and the merger of
lineages in evolution. Proceedings of the Royal Society of London. Series B,
Biological Sciences 265(1402): 1245-1253.
Levey, D. J., R. S. Duncan, and C. F. Levins. 2004. Animal behavior: use of dung as a
tool by burrowing owls. Nature 431: 39-39.
Liere, H., and A. Larson. 2010. Cascading trait-mediation: disruption of a trait-mediated
mutualism by parasite-induced behavioral modification. Oikos 119(9): 1394-
1400.
Little, A. E. F., and C. R. Currie. 2007. Symbiotic complexity: discovery of a fifth
symbiont in the attine ant-microbe symbiosis. Biology Letters 3(5): 501-504.
Machado, G., and D. M. Vital. 2001. On the occurrence of epizoic cyanobacteria and
liverworts on a neotropical harvestman (Arachnida: Opiliones). Biotropica 33(3):
535-538.
Mattner, S. W., and D. G. Parbery. 2001. Rust-enhanced allelopathy of perennial ryegrass
against white clover. Agronomy Journal 93: 54-59.
Mauck, K. E., C. M. De Moraes, and M. C. Mescher. 2010. Deceptive chemical signals
induced by a plant virus attract insect vectors to inferior hosts. Proceedings of the
National Academy of Sciences of the United States of America 107(8): 3600-
3605.
McAtee, W. L. 1944. Birds pickaback. The Scientific Monthly 58(3): 221-226.
McLeod, G., R. Gries, S. von Reuß, J. Rahe, R. McIntosh, W. König, and G. Gries. 2005.
The pathogen causing Dutch elm disease makes host trees attract insect vectors.
Proceedings of the Royal Society of London. Series B, Biological Sciences 272:
2499-2503.
Menge, B. A. 1995. Indirect effects in marine rocky intertidal interaction webs: patterns
and importance. Ecological Monographs 65: 21-74.
Milinski, M. 2006. Fitness consequences of selfing and outcrossing in the cestode
Schistocephalus solidus. Integrative & Comparative Biology 46(4): 373-380.
Mlot, C. 1997. A soil story. Science News 152(4): 58-59.
Montgomery, J. M., D. Gillespie, P. Sastrawan, T. M. Fredeking, and G. L. Stewart.
2002. Aerobic salivary bacteria in wild and captive Komodo dragons. Journal of
Wildlife Diseases 38(3): 545–551.
Morse, D. H. 1977. Feeding behavior and predator avoidance in heterospecific groups.
BioScience 27(5): 332-339.
Mouritsen, K. N., and R. Poulin. 2003. Parasite-induced trophic facilitation exploited by
a non-host predator: a manipulator's nightmare. International Journal for
Parasitology 33(10): 1043-1050.

93
Nelson, B. V., and R. R. Vance. 1979. Diel foraging patterns of the sea urchin
Centrostephanus coronatus as a predator avoidance strategy. Marine Biology
51(3): 251-258.
Ness, J. H., and S. A. Foster. 1999. Parasite-associated phenotype modifications in
threespine stickleback. Oikos 85: 127-134.
Oliver, K. M., J. A. Russell, N. A. Moran, and M. S. Hunter. 2003. Facultative bacterial
symbionts in aphids confer resistance to parasitic wasps. Proceedings of the
National Academy of Sciences of the United States of America 100(4): 1803-
1807.
Östman, C. 1997. Abundance, feeding behaviour and nematocysts of scyphopolyps
(Cnidaria) and nematocysts in their predator, the nudibranch Coryphella
verrucosa (Mollusca). Hydrobiologia 355: 21-28.
Owens, N. W., and J.D. Goss-Custard. 1976. The adaptive significance of alarm calls
given by shorebirds on their winter feeding grounds. Evolution 30(2): 397-398.
Paine, R. T. 1966. Food web complexity and species diversity. American Naturalist 100:
65-75.
Park, T. 1962. Beetles, competition, and populations: an intricate ecological phenomenon
is brought into the laboratory and studied as an experimental model. Science 138:
1369-1375.
Peacor, S. D., and E. E. Werner. 2001. The contribution of trait-mediated indirect effects
to the net effects of a predator. Proceedings of the National Academy of Sciences
of the United States of America 98(7): 3904-3908.
Peckarsky, B. L., P. A. Abrams, D. I. Bolnick, L. M. Dill, J. H. Grabowski, B. Luttbeg, J.
L. Orrock, S. D. Peacor, E. L. Preisser, O. J. Schmitz, and G. C. Trussell. 2008.
Revisiting the classics: considering nonconsumptive effects in textbook examples
of predator-prey interactions. Ecology 89(9): 2416-2425.
Peterson, M. A. 1995. Unpredictability in the facultative association between larvae of
Euphilotes enoptes (Lepidoptera: Lycaenidae) and ants. Biological Journal of the
Linnean Society 55(3): 209-223.
Piersma, T., and J. Drent. 2003. Phenotypic flexibility and the evolution of organismal
design. Trends in Ecology and Evolution 18(5): 228-233.
Plantan, T. B., M. J. Howitt, A. le Roux, J. A. Heymans, A. Kotz, and M. S. Gaines.
2009. The capture of a large number of Red-billed Oxpeckers, Buphagus
erythrorhynchus, and their subsequent maintenance and behaviour. Ostrich -
Journal of African Ornithology 80(2): 103-105.
Poelman, E. H., J. J. A. van Loon, and M. Dicke. 2008. Consequences of variation in
plant defense for biodiversity at higher trophic levels. Trends in Plant Science
13(10): 534-541.
Polis, G. A., C. A. Myers, and R. D. Holt. 1989. The ecology and evolution of intraguild
predation: potential competitors that eat each other. Annual Review of Ecology
and Systematics 20: 297-330.
Poulin, R. 1994. Meta-analysis of parasite-induced behavioural changes. Animal
Behaviour 48: 137-146.
Poulin, R. 2000. Manipulation of host behaviour by parasites: a weakening paradigm?
Proceedings of the Royal Society of London. Series B, Biological Sciences
267(1445): 787-792.

94
Preer, J. R., L. B. Preer, and A. Jurand. 1974. Kappa and other endosymbionts in
Paramecium aurelia. Bateriological Reviews 38: 113-163.
Preer, J. R., R. W. Siegel, and P. S. Stark. 1953. The relationship between kappa and
paramecin in Paramecium aurelia. Proceedings of the National Academy of
Sciences of the United States of America 39: 1228-1233.
Robinson, J. V., and G. A. Wellborn. 1987. Mutual predation in assembled communities
of odonate species. Ecology 68: 921-927.
Rossignol, P. A., J. M. C. Ribeiro, M. Jungery, M. J. Turell, A. Spielman, and C. L.
Bailey. 1985. Enhanced mosquito blood-finding success on parasitemic hosts:
evidence for vector-parasite mutualism. Proceedings of the National Academy of
Sciences of the United States of America 82(22): 7725-7727.
Rothschild, M., and J. A. Edgar. 1978. Pyrrolizidine alkaloids from Senecio vulgaris
sequestered and stored by Danaus plexippus. Journal of Zoology 186(3): 347-349.
Rydell, J., and J. R. Speakman. 1995. Evolution of nocturnality in bats: potential
competitors and predators during their early history. 54(2): 183-191.
Sabelis, M. W., A. Janssen, and M. R. Kant. 2001. ECOLOGY: Enhanced: The enemy of
my enemy is my ally. Science 291(5511): 2104-2105.
Samish, M., and J. Rehacek. 1999. Pathogens and predators of ticks and their potential in
biological control. Annual Review of Entomology 44(1): 159-182.
Sapp, J. 2004. The dynamics of symbiosis: an historical overview. Canadian Journal of
Botany 82(8): 1046-1056.
Sazima, I. 2002. Juvenile snooks (Centropomidae) as mimics of mojarras (Gerreidae),
with a review of aggressive mimicry in fishes. Environmental Biology of Fishes
65(1): 37-45.
Schemske, D. W., G. G. Mittelbach, H. V. Cornell, J. M. Sobel, and K. Roy. 2009. Is
there a latitudinal gradient in the importance of biotic interactions? Annual
Review of Ecology, Evolution, and Systematics 40: 245-269.
Schmitz, O. J., and T. D. Nudds. 1994. Parasite-mediated competition in deer and moose:
how strong is the effect of meningeal worm on moose? Ecological Applications 4:
91-103.
Shenoy, M., and R. M. Borges. 2010. Geographical variation in an ant-plant interaction
correlates with domatia occupancy, local ant diversity, and interlopers. Biological
Journal of the Linnean Society 100(3): 538-551.
Sheumack, D. D., M. E. Howden, I. Spence, and R. J. Quinn. 1978. Maculotoxin: a
neurotoxin from the venom glands of the octopus Hapalochlaena maculosa
identified as tetrodotoxin. Science 199(4325): 188-189.
Shiga, D. 2005. Poisonous partnership. Science News 167: 136-137.
Siemens, D. H., S. H. Garner, T. Mitchell-Olds, and R. M. Callaway. 2002. Cost of
defense in the context of plant competition: Brassica rapa may grow and defend.
Ecology 83(2): 505-517.
Simms, E. L., and R. S. Fritz. 1990. The ecology and evolution of host-plant resistance to
insects. Trends in Ecology and Evolution 5(11): 356-360.
Smith, D. C., and A. E. Douglas. 1987. The biology of symbiosis. Cambridge University
Press.

95
Solla, A., J. A. Martín, P. Corral, and L. Gil. 2005. Seasonal changes in wood formation
of Ulmus pumila and U. minor and its relation with Dutch Elm Disease. New
Phytologist 166: 1025-1034.
Springett, B. P. 1968. Aspects of the relationship between burying beetles, Necrophorus
spp., and the mite, Poecilochirus necrophori Vitz. Journal of Animal Ecology
37(2): 417-424.
Stachowicz, J. J., and N. Lindquist. 2000. Hydroid defenses against predators: the
importance of secondary metabolites versus nematocysts. Oecologia 124: 280-
288.
Stadler, B., and A. F. G. Dixon. 1999. Ant attendance in aphids: why different degrees of
myrmecophily? Ecological Entomology 24(3): 363-369.
Stecher, B., R. Robbiani, A. W. Walker, A. M. Westendorf, M. Barthel, M. Kremer, S.
Chaffron, A. J. Macpherson, J. Buer, J. Parkhill, G. Dougan, C. von Mering, and
W-D. Hardt. 2007. Salmonella enterica serovar Typhimurium exploits
inflammation to compete with the intestinal microbiota. PLoS Biology 5: e244.
Stevens, J. R., J. F. Wallman, D. Otranto, R. Wall, and T. Pape. 2006. The evolution of
myiasis in humans and other animals in the Old and New Worlds (part II):
biological and life-history studies. Trends in Parasitology 22(4): 181-188.
Swartz, M. B. 2001. Bivouac checking, a novel behavior distinguishing obligate from
opportunistic species of army-ant-following birds. The Condor 103: 629-633.
Tenney, S. 1877. A few words about scavengers. The American Naturalist 11(3): 129-
135.
Thaler, J. S. 2002. Effect of jasmonate-induced plant responses on the natural enemies of
herbivores. The Journal of Animal Ecology 71: 141-150.
Theilmann, D. A., and M. D. Summers. 1988. Identification and comparison of
Campoletis sonorensis virus transcripts expressed from four genomic segments in
the insect hosts Campoletis sonorensis and Heliothis virescens. Virology 167(2):
329-341.
Thomas, F., F. Renaud, and R. Poulin. 1998. Exploitation of manipulators: 'hitch-hiking'
as a parasite transmission strategy. Animal Behavior 56: 199-206.
Travis, J. 1996. An illuminating partnership for squid. Science News 150: 167-167.
Turlings, T. C. J., J. H. Loughrin, P. J. McCall, U. S. R. Rose, W. J. Lewis, and J. H.
Tumlinson. 1995. How caterpillar-damaged plants protect themselves by
attracting parasitic wasps. Proceedings of the National Academy of Sciences of
the United States of America 92(10): 4169-4174.
Turlings, T. C. J., and F. L. Wäckers. 2004. Recruitment of predators and parasitoids by
herbivore-damaged plants. Pages 21-75 in R. T. Carde and J. G. Miller, editors.
Advances in Insect Chemical Ecology. Cambridge University Press, New York.
Utsumi, S., Y. Ando, and T. Miki. 2010. Linkages among trait-mediated indirect effects:
a new framework for the indirect interaction web. Population Ecology 52(4): 485-
497.
van Baalen, M., and V. A. A. Jansen. 2001. Dangerous liaisons: the ecology of private
interests and common good. Oikos 95: 211-224.
Van Rijn, P. C. J., Y. M. van Houten, and M. W. Sabelis. 2002. How plants benefit from
providing food to predators even when it is also edible to herbivores. Ecology
83(10): 2664-2679.

96
Vinson, S. B., and G. F. Iwantsch. 1980. Host regulation by insect parasitoids. Quarterly
Review of Biology 55: 143-165.
Wäckers, F. L., and C. Bonifay. 2004. How to be sweet? Extrafloral nectar allocation by
Gossypium hirsutum fits optimal defense theory predictions. Ecology 85(6): 1512-
1518.
Wcislo, W. T., L. Arneson, K. Roesch, V. Gonzalez, A. Smith, and H. Fernández. 2004.
The evolution of nocturnal behaviour in sweat bees, Megalopta genalis and M.
ecuadoria (Hymenoptera: Halictidae): an escape from competitors and enemies?
Biological Journal of the Linnean Society 83(3): 377-387.
Weeks, P. 2000. Red-billed oxpeckers: vampires or tickbirds? Behavioral Ecology,
11(2):154-160.
Werner, E. E., and S. D. Peacor. 2003. A review of trait-mediated indirect interactions in
ecological communities. Ecology 84: 1083-1100.
Wheelwright, N. T., J. J. Lawler, and J. H. Weinstein. 1997. Nest-site selection in
Savannah sparrows: using gulls as scarecrows? Animal Behaviour 53: 197-208.
White, J. 1998. Envenoming and antivenom use in Australia. Toxicon 36(11): 1483-1492.
Whitfield, J. B. 1990. Parasitoids, polydnaviruses and endosymbiosis. Parasitology
Today 6(12): 381-384.
Widder, E. A. 1998. A predatory use of counterillumination by the squaloid shark,
Isistius brasiliensis. Environmental Biology of Fishes 53(3): 267-273.
Wilson, D. S. 1983. The effect of population structure on the evolution of mutualism: a
field test involving burying beetles and their phoretic mites. The American
Naturalist 121(6): 851-870.
Williams, B. L. 2010. Behavioral and chemical ecology of marine organisms with respect
to tetrodotoxin. Marine Drugs 8(3): 381-398.
Willis, E. O., and Y. Oniki. 1978. Birds and army ants. Annual Review of Ecology and
Systematics 9: 243-263.
Wootton, J. T. 1994. The nature and consequences of indirect effects in ecological
communities. Annual Review of Ecology and Systematics 25: 443-466.
Wrege, P. H., M. Wikelski, J. T. Mandel, T. Rassweiler, and I. D. Couzin. 2005. Antbirds
parasitize foraging army ants. Ecology 86: 555-559.
Yan, G., and L. Stevens. 1995. Selection by parasites on components of fitness in
Tribolium beetles: the effect of intraspecific competition. The American
Naturalist 146(5): 795-813.
Yan, G., L. Stevens, C. J. Goodnight, and J. J. Schall. 1998. Effects of a tapeworm
parasite on the competition of Tribolium beetles. Ecology 79(3): 1093-1103.

97
CONCLUSION
The research presented in this dissertation has uncovered many of the ecological
and evolutionary facets of inducible trophic polymorphisms (ITPs), as well as a good deal
of the fascinating life history of Tetrahymena vorax. My investigation of the potential
costs and benefits of ITPs allowed me to document the significance of the “tailed
microstome” morph of T. vorax as a reciprocal induced anti-cannibal defense. I was able
to demonstrate that the co-occurrence of the tailed microstome and the predatory
“macrostome” morph of T. vorax allows for a coordinated adaptive response to the
presence of intraguild prey. In exploring the potential of ITPs to give rise to novel trait-
mediated indirect interactions, I was able to draw attention to “chasmatectasis,” the
phenomenon of induced predators increasing their gape via the ingestion of increasingly
large prey. I demonstrated that chasmatectasis can mediate a previously undescribed
form of apparent competition. In assessing the differences between the expression of
ITPs in artificial and natural settings, I was able to show how community patterns may
change with the scale and trophic complexity of experimental systems, demonstrate the
utility of microcosms as a baseline for studies of larger systems, and offer insight into
possible reasons for the limited distribution of T. vorax in natural aquatic systems.
Finally, I also introduced the concept of an “indirect offense,” the use of symbionts or
recruited associates to increase the rate of acquisition of prey or hosts.

98
Curriculum Vitae
Aabir Banerji
Education
2004-2011 Ph.D.
Ecology & Evolution
GS-NB, Rutgers University
2000-2004 B.S.
Natural Resource Management
Cook College, Rutgers University
Employment
2004-2011 Teaching Assistant
Rutgers University
Publications
2009 Banerji, A., and P.J. Morin. Phenotypic plasticity, intraguild
predation and anti-cannibal defences in an enigmatic
polymorphic ciliate. Functional Ecology 23: 427-434.


















