







Languages
Pages
Legal


1
RESEARCH ARTICLE 1 2
Zipcode RNA-binding Proteins and Membrane Trafficking Proteins 3
Cooperate to Transport Glutelin mRNAs in Rice Endosperm 4
5 6
Li Tian1*, Kelly A. Doroshenk1, Laining Zhang1, Masako Fukuda1,2, Haruhiko Washida1,3, 7 Toshihiro Kumamaru2, Thomas Okita1* 8
9 1 Institute of Biological Chemistry, Washington State University, Pullman, WA 99164-6340, 10 USA 11 2 Faculty of Agriculture, Kyushu University, 744 Motooka Nishi-ku, Fukuoka 819-0395, Japan 12 3 Current address: Organic Nico Co., Ltd, Kyodai Katsura Venture Plaza, 1–36, Goryo-Ohara, 13 Nishikyo-ku, Kyoto 615–8245, Japan 14 * Correspondence should be addressed to [email protected] or [email protected]
16 Short Title: Glutelin mRNAs are transported on endosomes 17
18 One-sentence summary: Glutelin mRNAs are transported on endosomes through the direct 19 interactions of two RNA-binding proteins with two membrane trafficking factors. 20
21 The author responsible for distribution of materials integral to the findings presented in this 22 article in accordance with the policy described in the Instructions for Authors 23 (www.plantcell.org) is Thomas Okita ([email protected]) 24
25 26
ABSTRACT 27 28
In rice (Oryza sativa) endosperm cells, mRNAs encoding glutelin and prolamine are 29
translated on distinct cortical-endoplasmic reticulum (ER) subdomains (the cisternal-ER 30
and protein body (PB)-ER), a process that facilitates targeting of their proteins to 31
different endomembrane compartments. Although the cis- and trans-factors responsible 32
for mRNA localization have been defined over the years, how these mRNAs are 33
transported to the cortical ER has yet to be resolved. Here, we show that the two 34
interacting glutelin zipcode RNA-binding proteins (RBPs), RBP-P and RBP-L, form a 35
quaternary complex with the membrane fusion factors N-ethylmaleimide-sensitive factor 36
(NSF) and the small GTPase Rab5a, enabling mRNA transport on endosomes. Direct 37
interaction of RBP-L with Rab5a, between NSF and RBP-P, and between NSF and 38
Rab5a were established. Biochemical and microscopic analyses confirmed the co-39
localization of these RBPs with NSF on Rab5a-positive endosomes that carry glutelin 40
mRNAs. Analysis of a loss-of-function rab5a mutant showed that glutelin mRNA and 41
Plant Cell Advance Publication. Published on May 29, 2020, doi:10.1105/tpc.20.00111
©2020 American Society of Plant Biologists. All Rights Reserved

2
the quaternary complex were mis-targeted to the extracellular paramural body structure 42
formed by aborted endosomal trafficking, further confirming the involvement of 43
endosomal trafficking in glutelin mRNA transport. Overall, these findings demonstrate 44
that mRNA localization in plants co-opts membrane trafficking via the acquisition of new 45
functional binding properties between RBPs and two essential membrane trafficking 46
factors, thus defining an endosomal anchoring mechanism in mRNA localization. 47
48
INTRODUCTION 49 50
Localization of mRNAs is a universal mechanism to efficiently drive protein 51
targeting in eukaryotes and prokaryotes. The targeting of mRNAs facilitates the 52
accumulation of the locally translated proteins to specific cellular compartments and, 53
hence, is an essential mechanism in establishing cell polarity, patterning, and fate 54
determination as well as protein sorting (Herbert and Costa, 2019; Hughes and Simmonds, 55
2019; Tian et al., 2019b, 2020). 56
mRNA localization occurs as a multi-step process. After transcription, cis-acting 57
elements (RNA zipcodes) are recognized and bound by trans-acting factors, mainly 58
RNA-binding proteins (RBPs) to form a primary mRNA–nucleoprotein (mRNP) 59
complex. After export to the cytoplasm, the mRNP complex undergoes extensive 60
remodeling with recruitment of new factors and detachment of others enabling 61
cytoskeletal-based transport to the destination site (Blower, 2013; Weis et al., 2013; Tian 62
and Okita, 2014). 63
Although extensive knowledge on mRNA localization has been acquired by studies in 64
Drosophila, yeast and mammalian cells, only a few examples have emerged from higher 65
plants. The best defined model in plants is storage protein mRNA localization in 66
developing rice (Oryza sativa) endosperm cells, where mRNAs encoding glutelin and 67
prolamine are recognized by zipcode RBPs and transported to two distinct cortical 68
endoplasmic reticulum (ER) subdomains, the cisternal-ER, and protein body-ER (PB-69
ER), respectively (Chou et al., 2019; Tian et al., 2019b). Translation of prolamine 70
mRNAs on the PB-ER results in the assembly of prolamine intracisternal granules that 71
form an ER-derived protein body I (PB-I), while glutelin precursors are exported to the 72
Golgi and then transported to protein storage vacuoles (PSVs) for processing and storage 73

3
(Chou et al., 2019; Tian et al., 2019b). Although several cytoskeleton-associated RBPs 74
required for mRNA localization have been identified (Doroshenk et al., 2009, 2012), 75
information on how these mRNAs are transported to distinct ER subdomains remains 76
elusive. 77
Emerging evidence from fungal model systems reveals the intimate link of mRNA 78
transport with membrane trafficking (Schmid et al., 2006; Jansen et al., 2014; Haag et al., 79
2015; Niessing et al., 2018). Several mRNAs from Saccharomyces cerevisiae, Candida 80
albicans, and Ustilago maydis are co-transported with mobile ER or shuttling endosomes 81
(Schmid et al., 2006; Jansen et al., 2014; Haag et al., 2015; Pohlmann et al., 2015; 82
Niessing et al., 2018). ASH1 as well as other mRNAs are co-transported on tubular ER 83
that moves to the emerging bud or daughter cell in Saccharomyces cerevisiae. This 84
process is mediated by the RBPs She2p and She3p, with She2p having membrane 85
binding properties and She3p serving as an adaptor protein linking the mRNP-cER to 86
Myo4P protein (Schmid et al., 2006; Niessing et al., 2018). The cdc3 mRNA is 87
transported on shuttling endosomes in the smut fungus, Ustilago maydis, a process 88
requiring localization of the RBP Rrm4 on the endosomes and the interaction of a 89
membrane-associated linker protein Upa1 with Rrm 4 (Pohlmann et al., 2015; Niessing et 90
al., 2018). Specific adaptor proteins appear to be needed to hitch mRNPs on endosomes 91
for active transport over long distance. More recently, neuronal RNA granules have been 92
shown to hitchhike on moving lysosomes using annexin11 as a tether (Liao et al., 2019). 93
Although co-transport of mRNAs with membranous compartments was proposed to be a 94
common mechanism in higher eukaryotes (Jansen et al., 2014), whether the mechanism is 95
utilized by higher plants remains to be determined. 96
Previous studies suggested that endocytosis and membrane trafficking likely play a 97
role in mRNA localization in plants. For example the loss-of-function of the small 98
GTPase Rab5a and its cognate guanine nucleotide exchange factor (GEF) resulted in 99
defects in endocytosis and membrane trafficking and the mis-targeting of glutelin 100
proteins to the prolamine containing PB-I as well as to the extracellular paramural body 101
(PMB) in rice endosperm cells (Fukuda et al., 2011; Wen et al., 2015). As storage protein 102
targeting is regulated by their mRNA localization in rice endosperm cells, the mis-103
targeting of glutelin proteins in the mutant suggests a relationship between endosomal 104

4
transport and glutelin mRNA localization in rice. The extracellular distribution of glutelin 105
mRNAs within PMBs from a mutant expressing a defective GEF (Yang et al., 2018) 106
further supports the possible involvement of endosomal trafficking in glutelin mRNA 107
transport. However, direct evidence depicting the co-transport of glutelin mRNAs with 108
shuttling endosomes and how endosomal trafficking are engaged in glutelin mRNA 109
localization have yet to be established. Such mis-targeting of glutelin mRNAs in rice 110
lines expressing mutant Rab5a and GEF may simply be a consequence of pleiotropy. 111
Recent studies (Tian et al., 2018; Tian et al., 2019a) identified two RNA-binding 112
proteins, RBP-P and RBP-L, which contain two and three RNA recognition motif (RRM) 113
domains, respectively. These RBPs specifically bind to the glutelin zipcode mRNA 114
sequences and regulate glutelin mRNA localization. In this study, using these two 115
glutelin zipcode RBPs as entry points, we identified their interacting partners, N-116
ethylmaleimide-sensitive factor (NSF) and the small GTPase Rab5a, which participate in 117
endosomal membrane trafficking. The four proteins may form a quaternary complex 118
carrying glutelin mRNAs for active transport on endosomes to the cortical ER membrane. 119
The identification of these key linker proteins that enable endosome-mediated mRNA 120
transport in rice endosperm cells provides new insights on how mRNAs can be 121
distributed to specific locations in eukaryotes. 122
123
RESULTS 124
RBP-P interacts with membrane fusion factor NSF 125
Previous studies (Doroshenk et al., 2014; Tian et al., 2018; Tian et al., 2019a) 126
established that the glutelin zipcode RNA-binding proteins RBP-P and RBP-L, which 127
interact with each other, are essential for localization of glutelin mRNAs to the cisternal-128
ER, as mutations in these RBPs led to the mis-localization of glutelin mRNAs. To obtain 129
additional insight into how glutelin mRNAs are transported to the ER, we performed 130
immunoprecipitation-mass spectrometry (IP-MS) studies using affinity-purified anti-131
RBP-P antibody (Figure 1A). Tandem mass spectrometry of a major polypeptide band 132
observed in the IP generated with anti-RBP-P but not with control anti-GFP identified a 133
major interacting protein as N-ethylmaleimide sensitive fusion protein (NSF). 134

5
To determine whether RBP-P interacts directly with NSF, yeast two-hybrid studies 135
were carried out. In such two-hybrid screening, the two proteins of interest were fused to 136
activating domain (AD) and DNA-binding domain (BD) domain of yeast GAL4 137
transcription factor, respectively. Interaction of these proteins restore GAL4 that, in turn, 138
activate the transcription of histidine and adenine reporter genes. While no background 139
interaction of RBP-P and NSF with complementary empty vector was observed, yeast 140
cells carrying both RBP-P and NSF genes activated the reporter genes and survived in 141
selection medium lacking histidine and adenine (Figure 1B). These results indicate that 142
RBP-P and NSF interact under stringent binding conditions. 143
To further substantiate the interaction between RBP-P and NSF, bimolecular 144
fluorescence complementation (BiFC) analyses using tobacco (Nicotiana tabacum) BY-2 145
cells was performed. In addition to supporting an interaction between protein pairs, BiFC 146
also provides intracellular information on where this interaction occurs. RBP-P and NSF 147
were fused to two complementary non-fluorescent fragments of enhanced yellow 148
fluorescent protein (EYFP), nEYFP and cEYFP, respectively, and co-transformed into 149
live BY-2 cells. If the two proteins interact, the two EYFP fragments are brought in close 150
proximity to reform the native protein structure enabling emission of yellow fluorescence 151
and, in turn, direct visualization of the protein complex in live cells (Kerppola, 2006; 152
Miller et al., 2015). EYFP fragments lacking one of the putative protein partners were 153
used as negative control. As shown in Figures 2A-2C, while no interaction was detected 154
between RBP-P or NSF with the empty EYFP fragment controls, cells expressing both 155
RBP-P and NSF fused to EYFP fragments emitted bright yellow fluorescence, indicating 156
that RBP-P interacted with NSF. Closer examination reveals that RBP-P/NSF complexes 157
were distributed to the cytoplasm as intensely bright clusters together with more loosely 158
diffuse structures (Figure 2C). 159
NSF is a soluble hexameric ATPase commonly found in the cytoplasm of eukaryote 160
cells (Mastick and Falick, 1997; Zhao et al., 2007; Zhao et al., 2010; Guo et al., 2017), 161
which predominantly plays a major role as a chaperone in intracellular membrane fusion 162
events. Through its interaction with the adaptor protein, soluble NSF attachment protein 163
(SNAP), NSF binds to SNARE (soluble NSF activating protein receptor) complexes and 164
utilizes the energy of Mg2+
-dependent ATP hydrolysis to disassemble the SNARE 165

6
protein complex, and thus facilitating the recycling of SNARE proteins for further cycles 166
of membrane fusion (Zhao et al., 2007; Chang et al., 2012; Ryu et al., 2015). During this 167
process, SNAP serves as chaperone by stimulating the ATPase activity of NSF and 168
dissociating from NSF after ATP hydrolysis (Zhao et al., 2007). This transient interaction 169
between NSF and SNAP is only detected under conditions where a nonhydrolyzable ATP 170
is used as a substrate (Hanson et al., 1995; Barnard et al., 1997; Chang et al., 2012). 171
Alternatively, the addition of EDTA to chelate Mg2+
and thus inhibit ATPase activity has 172
been used to detect the transient interaction between NSF and SNAP (Hanson et al., 1995; 173
Barnard et al., 1997; Chang et al., 2012; Li et al., 2018). 174
Based on the established properties of NSF, we performed Co-IP experiments to 175
assess the formation of RBP-P/NSF complexes in rice endosperm cells. Rice seed lysates 176
supplemented with 1 mM ATP and 8 mM MgCl2 or EDTA were incubated with affinity 177
purified anti-RBP-P and NSF antibodies (Figure 2D) or anti-GFP antibody, the latter 178
used as a negative control. No proteins were captured by the control GFP antibody, 179
revealing the reliability of the IP experiments. Interestingly, RBP-P and NSF were co-180
precipitated in the presence of Mg2+
-ATP but not when EDTA was added to the seed 181
lysates (Figure 2D). This result indicates that formation of the RBP-P/NSF complex in 182
rice endosperm cells requires Mg2+
-ATP, a condition distinct from those complexes 183
involving the stable interaction between NSF and SNAP (Hanson et al., 1995; Barnard et 184
al., 1997; Chang et al., 2012; Li et al., 2018). The differences in binding properties, as 185
well as the absence of SNAP in RBP-P IPs (Figure 2D), indicate that the events of 186
membrane fusion requiring NSF-SNAP are not required for formation of the RBP-P/NSF 187
complex. On the other hand, the interaction between NSF with RBPs suggests that NSF 188
may function in mRNA metabolism by its interaction with RBP-P. 189
190
RBP-P indirectly interacts with Rab5a through NSF 191
The interaction of the glutelin zipcode trans-factor RBP-P with NSF supports a close 192
relationship between glutelin mRNA transport with membrane trafficking. Previous 193
studies demonstrated that Rab5a, an evolutionarily conserved key GTPase involved in the 194
biogenesis of early endosomes and membrane trafficking in the cytoplasm (Woodman, 195
2000; Saito and Ueda, 2009; Ito et al., 2018), is also required for glutelin mRNA 196

7
localization (Doroshenk et al., 2010) and vesicular membrane transport between the 197
Golgi and protein storage vacuole in rice endosperm cells (Wang et al., 2010; Fukuda et 198
al., 2011). Therefore, we investigated whether the abovementioned RBP-P/NSF complex 199
is associated with Rab5a to regulate the endosomal transport of mRNAs. 200
We first performed a transient expression study to confirm the association of rice 201
Rab5a (Figures 2E-2F, Supplemental Figure 1) with endosomes. FM4-64 is a lipophilic 202
dye that initially labels the plasma membrane and, subsequently, internalizes with 203
membrane vesicles transported along the endocytic pathway (Vida and Emr, 1995; Ueda 204
et al., 2001). In this experiment, BY-2 protoplasts expressing GFP-Rab5a were treated 205
with FM4-64 for 15 min before observation. As shown in Figure 2F, red fluorescence 206
derived from FM4-64 was evident on the plasma membrane and in internalized 207
endosomes. GFP-Rab5a displayed a similar distribution pattern and co-localized with the 208
internal FM4-64 labeled vesicles (Figure 2F), an observation confirming that Rab5a is 209
associated with endosomes. 210
We then studied the relationship of Rab5a with the abovementioned RBP-P/NSF 211
complex. Co-IP studies were carried out using affinity-purified antibodies to Rab5a, 212
RBP-P and NSF. All three proteins were found in IPs generated by anti-Rab5a, anti-RBP-213
P and anti-NSF (Figure 2D), suggesting that they form a multi-protein complex in rice 214
endosperm cells. Protein complex formation apparently required active ATPase 215
hydrolysis as all three protein interactions could only be simultaneously detected in the 216
presence of Mg2+
and ATP and not when EDTA and ATP were included (Figure 2D). 217
To further characterize this protein interactome and the formation of their complexes, 218
BiFC (Figures 2G-2J) studies were performed. Interaction between NSF and Rab5a was 219
observed in the tested cells, and their complexes existed in the cytoplasm. By contrast, 220
Rab5a does not interact with RBP-P (Figure 2J) and, therefore, the association of RBP-P 221
with Rab5a in the Co-IPs (Figure 2D) is mediated through the interaction between NSF 222
and Rab5a. 223
Rab GTPases are highly conserved small proteins and share two conserved regions 224
that are required for conformational switching between active and inactive states of GTP 225
hydrolysis (Figure 2E, Supplemental Figure 1). The switch regions are disordered in an 226
inactive conformation and adopt a well-defined conformation in the active stage 227

8
(Stenmark and Olkkonen, 2001). Amino acid substitutions within the switch regions 228
affects their conformational switching. While the G45D mutation in switch 1 inhibits the 229
conformation switching to an active GTPase stage and restricts Rab5a in a GDP-bound 230
form, a Q70L mutation in switch 2 locks it in a constitutively active GTP bound form 231
(Lee et al., 2009). Only the GTP-bound form of Rab5 are membrane-bound during 232
endosomal trafficking (Yuan and Song, 2020). 233
To investigate whether the NSF-Rab5a complex is associated with active endosomes, 234
the interaction of NSF with the two structural forms of Rab5a containing G45D or Q70L, 235
was analyzed by BiFC (Figures 2K-2L). Although NSF directly interacts with both GDP- 236
(Rab5aG45D
) and GTP- (Rab5aQ70L
) forms of Rab5a, their complexes are observed as two 237
distinct populations. When bound to the GTP-fixed Rab5a (Q70L) form (Figure 2L), the 238
NSF complex was distributed as endosomal punctate structures. By contrast, the NSF 239
complex constituted by GDP-fixed Rab5a (G45D) showed a diffuse distribution 240
throughout the cytoplasm (Figure 2K). These results are consistent with the view that the 241
GTP-fixed Rab5a (Q70L) form is membrane bound while the GDP-fixed Rab5a (G45D) 242
is mainly soluble. 243
The direct interaction between NSF and Rab5a provides an indirect link to RBP-P 244
and, in turn, RNA localization. To confirm the association of the RBP-P/NSF complex to 245
Rab5a-positive endosomes, we performed a three-way co-localization test consisting of 246
RBP-P and NSF as BiFC interacting partners in BY-2 cells expressing Rab5a tagged with 247
the red fluorescence protein (RFP) (Figures 2M-2N). Florescence analysis of BY-2 cells 248
showed that RFP-Rab5aWT
was distributed as diffuse signals throughout the cytoplasm 249
and as punctate structures (endosomes) near the plasma membrane (Figure 2M, middle 250
panel). By contrast, RFP-Rab5aQ70L
was present only as punctate structures (Figure 2N, 251
middle panel) indicating that the activated GTPase form of Rab5a is associated with 252
endosomes. The fluorescence distribution pattern seen for RFP-Rab5aWT
is consistent 253
with membrane-free Rab5a being distributed in the cytoplasm and active Rab5a 254
associated with endosomes (Yuan and Song, 2020). 255
As earlier seen in Fig. 2C, BiFC-linked RBP-P/NSF complexes are distributed as 256
fluorescent punctate structures in BY-2 cells (Figure 2M and N). These RBP-P/NSF 257
complexes co-localized with RFP-Rab5a-associated endosomes in the cytoplasm, 258

9
especially with membrane-associated GTP-fixed (Q70L) Rab5a (Figures 2M-2N). To 259
further confirm that the RBP-P/NSF complexes are associated with endosomes, we used 260
FM4-64, which specifically labels these small membrane compartments (Figure 2O). The 261
fluorescent signals from the RBP-P/NSF complexes co-localized with endosomal 262
compartments labeled by FM4-64, providing direct evidence that RBP-P/NSF complexes 263
co-localize with endosomes. Consistent with the abovementioned Co-IP analysis (Figure 264
2D) where RBP-P, NSF and Rab5a may co-assemble into a multi-protein complex in rice 265
endosperm cells, the BiFC results suggest that RBP-P is associated with endosomes 266
through a NSF-mediated interaction with Rab5a. 267
It should be noted that not all RBP-P/NSF complexes co-localized with Rab5a-linked 268
endosomes (Figures 2M-2N, open arrowheads). Similarly, the location of several Rab5a-269
active endosomes (Figures 2M-2N, open arrowheads) do not coincide with RBP-P/NSF 270
complexes. The lack of overlap in the distribution of a population of RBP-P/NSF 271
complexes and Rab5 endosomes suggests the multiple roles of these proteins in RNA 272
localization and membrane trafficking. 273
274
RBP-L is involved in the RBP-P/NSF/Rab5a complex 275
We had previously demonstrated that RBP-P co-assembles with RBP-L to form a 276
protein complex that is essential for storage protein mRNA localization (Doroshenk et al., 277
2014; Tian et al., 2018; Tian et al., 2019a). To determine whether RBP-L is also 278
involved in interacting with NSF or Rab5a, BiFC studies were conducted (Figures 3A-F). 279
Such analysis showed that RBP-L interacts directly with Rab5a but not with NSF 280
(Figures 3A-3C). The RBP-L/Rab5aWT
complex is observed as two distinct populations. 281
One population is distributed as endosomal punctate structures with the bulk located 282
close to the plasma membrane. The co-localization of RBP-L/ Rab5aWT
with endosomes 283
is supported by their close association with FM4-64 labeled endosomal compartments 284
(Figure 3D). A second population of RBP-L/ Rab5aWT
is viewed as a diffuse cloud 285
around the nucleus. Similar to the RBP-P/NSF/Rab5a complex, the interaction of RBP-L 286
with Rab5a was not dependent on the functional state of Rab5a as it interacts with both 287
the GTP-bound Rab5aQ70L
and GDP-bound Rab5aG45D
(Figures 3E and 3F), although the 288
distribution patterns are distinct. RBP-L/ Rab5aG45D
is distributed mainly as a diffuse 289

10
cloud around the nucleus and near the plasma membrane while RBP-L/ Rab5aQ70L
are 290
observed predominantly as discrete endosomal punctate structures. 291
To further determine whether RBP-L is associated with the RBP-P/NSF/Rab5a 292
complex, we performed Co-IP with paraformaldehyde-fixed seed extracts to optimize the 293
capture of any potential dynamic endosome-associated complexes formed in vivo. Such 294
results showed that, irrespective of the antibody used in the initial immunoprecipitation, 295
RBP-P, RBP-L, NSF and Rab5a were detected in IPs generated by all four antibodies 296
(Figure 3G). By contrast, SNAP is only present in IPs obtained with anti-NSF and anti-297
Rab5 but not in IPs generated by antibodies to RBP-P and RBP-L (Figure 3G). These 298
results are consistent with the Co-IP results depicted in Figure 2D where SNAP is present 299
in IPs generated with antibodies to NSF and Rab5a in the presence of EDTA but not in 300
the presence of MgCl2. Overall, these results are consistent with the view that NSF-301
Rab5a complexes exist as two separate populations. One NSF-Rab5a population together 302
with SNAP participates in Rab5-mediated endosomal fusion (Woodman, 2000; Zhao et 303
al., 2007), where NSF and SNAP disassemble the SNARE complex formed by Rab5-304
mediated membrane fusion. A second NSF-Rab5a complex contains RBP-P and RBP-L 305
and is independent of SNAP. 306
To further address whether RBP-P and RBP-L are attached to endosomes together, 307
sequential IPs were performed (Figure 3H). In this analysis, the RBP-P/NSF/Rab5a 308
complexes precipitated by anti-RBP-P antibodies were next subjected to an additional IP 309
using anti-RBP-L. All four proteins were detected in the second IP generated by anti-310
RBP-L, suggesting that RBP-P, RBP-L, NSF and Rab5 co-assemble to form a quaternary 311
protein complex. 312
313
The quaternary complex attaches to active endosomes carries glutelin mRNAs 314
To investigate whether the quaternary complex binds glutelin mRNA, we performed 315
RNA-IP analysis to detect the in vivo association of the complex with glutelin mRNAs 316
(Figure 3I). In this analysis, antibodies to RBP-P, RBP-L, NSF and Rab5a were utilized 317
to capture the associated RNA-protein complexes, and the RNA was subsequently 318
isolated from the IP fractions and subjected to RT-PCR using specific primers to amplify 319
glutelin transcripts. Compared with the negative empty-resin control and anti-GFP 320

11
antibody, glutelin mRNAs were highly enriched in IPs generated by all four antibodies 321
(Figure 3I). The mRNA amount associated with NSF and Rab5a was lower than that of 322
RBP-P and RBP-L, a result consistent with the roles of these proteins in membrane 323
fusion events. Overall, these results support the view that the quaternary complex 324
contains glutelin mRNAs. 325
Based on these results, a working model of cytosolic glutelin mRNA transport is 326
proposed in Figure 4. The mRNP complex containing glutelin mRNA, RBP-P and RBP-L 327
is bound to Rab5a-associated endosomes through a 4-way interactome, i.e. the direct 328
interaction of RBP-P with RBP-L, of RBP-L with Rab5a, of NSF with RBP-P, and of 329
Rab5a with NSF. The GTP-bound Rab5a is associated with endosomes (Yuan and Song, 330
2020) as suggested by distribution of this activated Rab5a form as endosomal punctate 331
structures (Figures 2N and 3F). 332
333
Loss-of-function of Rab5a results in mis-targeting of glutelin mRNAs 334
To provide evidence in support of this model, we analyzed a rab5a mutant EM960 335
(Fukuda et al., 2011) expressing a GDP-fixed (G45D) Rab5a (Figure 5A). Similar to the 336
phenotype shown in the EM956 mutant lacking Rab5a (Fukuda et al., 2011) or a mutant 337
line expressing a defective Rab5a effector GEF (Wen et al., 2015), normal endosomal 338
trafficking is disrupted in the endosperm cells of GDP-fixed rab5a mutant and leads to 339
the formation of PMBs (Figures 5B-5C), an aborted endosome complex containing mis-340
sorted endomembrane proteins. These extracellular PMBs, which contain numerous 341
electron-dense vesicles, are located in the space between the invaginating plasma 342
membrane and the cell wall in the mutant endosperm cells (Figures 5B-5C). 343
To investigate the co-localization of RBP-P, RBP-L, and NSF with Rab5a and the 344
subcellular localization of their complex in rice endosperm cells, we performed double 345
immuno-fluorescence labeling on thin sections of rice developing seeds using antibodies 346
raised against each of the four proteins. Although the bulk of these proteins were 347
evidently independent of Rab5, there was ample evidence for co-localization of RBP-P, 348
RBP-L, and NSF with Rab5a. The co-localization of these proteins with Rab5a was 349
apparent as punctate structures in the cytoplasm, particularly in the cortical region 350
underneath the plasma membrane (Figures 6A, 6C and 6E), an intracellular location 351

12
enriched in Rab5a-mediated endosome activity (Chavrier et al., 1990; Fischer von 352
Mollard et al., 1994). To directly assess the co-localization of these proteins, the 353
fluorescence intensity profiles of these proteins were quantified along a specific linear 354
distance (Figure 6, right panels). The fluorescence signals for the proteins examined 355
overlapped substantially indicating that RBP-P, RBP-L and NSF co-localized to Rab5a-356
labeled endosomal compartments in rice endosperm cells. The independent distribution 357
of RBP-P, RBP-L and NSF with Rab5a was also evident in the BiFC/RFP double 358
labeling (Figures 2M-N), which is indicative of their roles in other cellular processes. 359
This view is also supported by the Co-IP results (Figures 2F, 3H) where IPs by antibodies 360
to RBP-P, RBP-L and NSF contained only a small proportion of the total Rab5a amounts. 361
The RBPs retain their co-localization with Rab5a in the rab5a mutant (Figures 6B 362
and 6D), an expected observation as RBP-L as well as NSF interact with both the GDP- 363
and GTP-fixed Rab5a forms (Figures 2K-2L, Figures 3E-3F). These protein complexes 364
exist as punctate structures within the PMBs in the rab5a mutant, (Figures 6B, 6D and 365
6F). Hence, disruption of membrane trafficking in the rab5a mutant displaces not only 366
endosomal proteins to the extracellular PMBs but also RBP-mRNA complexes. 367
Transmission electron microscopy (TEM) analysis further confirmed the co-368
localization of RBP-P and RBP-L with Rab5a on endosomes in wildtype and aborted 369
endosome vesicles within PMBs in rab5a mutant (Figure 7). In wild type, Rab5a-370
mediated endosomes were observed as electron-dense vesicles with an irregular shape 371
likely due to endosomal fusion (Figures 7A-7E). Co-localization of RBP-P and RBP-L 372
with Rab5a was observed on those endosomes, further suggesting that mRNP complexes, 373
carrying glutelin mRNAs bound by RBP-P and RBP-L, are transported on endosomes. In 374
the rab5a mutant line, normal endosomal trafficking is disrupted resulting in the 375
displacement of mRNA-associated endosomes to the extracellular PMBs (Figures 7F-7J). 376
To further address whether glutelin mRNAs are associated with the Rab5a-mediated 377
endosomes, in situ reverse transcription (RT)-PCR on developing endosperm sections 378
was performed to locate glutelin mRNAs. In this experiment, Rhodamine B dye was used 379
to specifically stain the PB-ER (Muench et al., 2000) while glutelin mRNAs were labeled 380
with Alexa488-dUTP by in situ RT-PCR using glutelin-specific primers (Figure 8). In 381
wild type, glutelin mRNAs are localized on the cisternal-ER separate from PB-ER 382

13
(Figure 8). In rab5a mutant cells, however, they were mainly distributed to the PMBs and 383
with smaller amounts associated with the PB-ER. These results indicate that normal 384
glutelin mRNA localization is disrupted in the rab5a mutant. This result resonates with 385
the retention of the complexes formed by RBP-P, RBP-L, NSF and Rab5a in PMBs 386
(Figures 6 and 7), further supporting a Rab5a-dependent endosome transport of glutelin 387
mRNAs. 388
Taken all together, the results described here support the view that glutelin mRNAs 389
are transported to the cisternal-ER membrane via their hitchhiking on endosomes. In the 390
rab5a mutant, however, glutelin mRNA-containing mRNP complexes are not transported 391
to their normal location but are mis-targeted to the PB-ER and displaced to the 392
extracellular PMBs because of the disruption in normal endosomal trafficking. 393
394
395
DISCUSSION 396
The study of rice storage protein mRNA localization as a model has provided 397
considerable information on why and how plant mRNAs are localized to specific ER 398
compartments. Although zipcode cis-elements of glutelin and prolamine mRNAs and 399
several key RNA-binding proteins have been identified (Hamada et al., 2003; Washida et 400
al., 2009; Doroshenk et al., 2012; Washida et al., 2012; Tian et al., 2018; Tian et al., 401
2019a), the mechanism by which glutelin and prolamine mRNAs are transported to 402
distinct subdomains of the ER membrane has yet to be established. Here, we report that 403
rice endosperm cells employ Rab5a and NSF, proteins traditionally known for their 404
essential roles in endosomal trafficking and membrane fusion events, as adaptor proteins 405
linking mRNA-protein complexes to endosomes for active transport of glutelin mRNAs. 406
Active transport of mRNAs along cytoskeleton networks by hitchhiking on motile, 407
membrane-bound organelles has been reported in mammalian and fungi cells. In human 408
axons, the Ras GAP SH3 domain binding protein 1 (G3BP1)-labeled RNA granules were 409
found to co-localize and co-traffic with moving lysosomes along microtubules (Liao et al., 410
2019). The membranous amyotrophic lateral sclerosis (ALS)-associated 411
phosphoinositide-binding protein, annexin A11 (ANX11), functions as a molecular tether 412
between RNA granules and lysosomes (Liao et al., 2019). In addition to mRNAs, 413

14
neuronal precursor miRNAs are transported to the tip of growing axons by hitchhiking on 414
late endosomes/lysosomes (Corradi et al., 2020). 415
Endosomal transport of mRNAs has been defined in the filamentous fungus Ustilago 416
maydis, where mRNAs are transported throughout the growing hyphae via endosome 417
trafficking on microtubules (Baumann et al., 2012; Gohre et al., 2012; Vollmeister et al., 418
2012; Pohlmann et al., 2015; Niessing et al., 2018). mRNP complexes carrying cdc3 419
mRNAs are attached to endosomes via the adaptor protein Upa1, which associates with 420
the endosomal membrane through its FYVE domain and two RNA binding proteins, 421
Pab1 and Rrm4, through its PABP-associated motif 2 (PAM2) and PAM2-like (PAM2L) 422
domains, respectively (Pohlmann et al., 2015). 423
Unlike the abovementioned examples in mammalian and fungi cells, higher plants 424
apparently adapted the two membrane trafficking factors, NSF and Rab5a, to mediate 425
transport of storage protein mRNAs. The linking of glutelin mRNP complexes to 426
endosomes is mediated by the four-way interactions of NSF and Rab5a with the two 427
glutelin zipcode RNA-binding proteins, RBP-P and RBP-L. These interacting proteins 428
likely form a quaternary complex enabling endosomal transport of glutelin mRNAs. Such 429
protein to protein interactions are accomplished through a gain in binding properties by 430
Rab5a and NSF, and the glutelin zipcode mRNA binding proteins, RBP-P and RBP-L, 431
which allow for the heterotypic interaction of a RNA binding protein with a membrane 432
fusion factor (RBP-P/NSF and RBP-L/Rab5a). The interaction of an RBP with a 433
membrane trafficking factor, i.e. between RBP-P with NSF and between RBP-L and 434
Rab5a, as well as between NSF and Rab5a are unexpected findings as such interactions 435
have not been reported in other organisms. It remains unclear whether the interactions of 436
these membrane trafficking factors to RBPs and even between themselves is an inherent 437
property or whether they are unique to higher plants. While Upa1-mediated endosomal 438
mRNA transport is proposed as evolutionarily conserved in fungi (Muller et al., 2019), 439
further investigation is needed to assess whether endosomal mRNA transport by the NSF-440
Rab5a-RBP machinery is a widespread phenomenon in transporting mRNAs among 441
higher plants and other eukaryotic organisms. 442
It appears that the recognition of NSF and Rab5a with RBPs is highly selective. 443
These non-random interactions highlight the diverse binding capabilities of NSF, Rab5a, 444

15
as well as for RBP-P and RBP-L in rice endosperm cells. Both RBP-P and RBP-L 445
contain RNA recognition motifs (RRM) with the former having two RRM motifs and the 446
latter having three RRM motifs. Due to the conservation of the RRM motifs, the 447
recognition of NSF and Rab5a to RBP-P and RBP-L, respectively, is likely via the unique 448
N- and C-terminal regions that flank the RRM motifs. Indeed, our preliminary results 449
(Supplemental Figure 2) show that the N-terminal end of RBP-P is essential for its 450
interaction with NSF. Through the selective recognition of NSF and Rab5a to other 451
specific RBPs, these membrane trafficking factors could serve as the core components 452
enabling endosome-coupled mRNA transport with the RBPs specifying the mRNA 453
species. 454
NSF is homo-hexamer with three domains: the N-terminal domain (NSF-N) that is 455
required for SNAP-SNARE binding; the ATP-binding domain 1 (NSF-D1) responsible 456
for ATPase activity; and the ATP-binding domain 2 (NSF-D2) responsible for 457
hexamerization (Tagaya et al., 1993; Zhao et al., 2007). The NSF-N domain is the main 458
protein-protein interaction site for NSF binding to other proteins. NSF-N domain is likely 459
responsible for its interaction with the N-terminus of RBP-P in rice as well 460
(Supplemental Figure 2). Sequence alignment of NSF homologues from rice, Arabidopsis, 461
Drosophila, yeast and human show that the N-terminal region displays considerable 462
sequence diversity among these species (Supplemental Figure 3). Further structural 463
studies are required to investigate their binding mechanism and whether this kind of 464
interaction occurs in other species. 465
Rab5a interacts with a large number of proteins, including its regulators (activating 466
effectors and inhibitors), linkers to molecular motors, membranous factors, components 467
of membrane fusion complexes, protein kinases and phosphatases (Herve and 468
Bourmeyster, 2018; Pylypenko et al., 2018). Many of these interacting proteins function 469
as Rab5a effectors to activate endosomal transport and tightly control the specific 470
functions of Rab5a in membrane composition and modification, cytoskeleton regulation, 471
and intracellular trafficking (Herve and Bourmeyster, 2018; Pylypenko et al., 2018). 472
Although NSF has been reported to interact with several Rab5a effectors to drive 473
membrane fusion during endosomal docking (McBride et al., 1999; Grosshans et al., 474
2006), no direct relationship between Rab5a with NSF or with a RNA-binding protein 475

16
had been established until now. These findings will stimulate future research efforts on 476
identifying new alternative roles of Rab5a and NSF beyond membrane fusion. 477
Although the GTPase activity of Rab5a is not required for the apparent formation of 478
the quaternary protein complex as the four proteins still co-localized in the GDP-fixed 479
(G45D) Rab5a line (Figures 6-7), Rab5 GTPase activity is required for proper mRNA 480
transport. Endosome formation and membrane trafficking are dependent on Rab5 GTPase 481
activity (Woodman, 2000; Zeigerer et al., 2012). Null Rab5a activity results in aborted 482
endosomal transport in the mutant, which, in turn, disrupts glutelin mRNA transport on 483
endosomes and causes its mis-localization to the PB-ER as well as displacement to the 484
extracellular PMBs (Figure 8). Therefore, the active GTPase activity of Rab5a is essential 485
for the transport of glutelin mRNAs on endosomes. This also raises the question on 486
whether NSF and RBP-L act as Rab5a effectors to modulate the role of Rab5a in mRNA 487
transport and localization. While this hypothesis needs further examination, other 488
effectors, functionally equivalent to Rabaptin-5 and EEA1 that interact with both NSF 489
and Rab5a (McBride et al., 1999; Grosshans et al., 2006), may also be involved. These 490
effectors, including the abovementioned GEF, may allow further stabilization of the 491
linkage of mRNP complex to active endosomes and regulate endosomal mRNA transport 492
via the cytoskeleton (Figure 4). 493
The identification and study of rice lines expressing mutations in Rab5a were 494
instrumental in establishing its involvement in both RNA and membrane trafficking in 495
developing rice endosperm. While Rab5 is an essential growth factor, loss of Rab5a only 496
had a slight effect on rice growth and development as its activity is complemented by 497
other Rab5 activities. Rice expresses three other Rab5 genes, Rab5b, Rab5c, and Rab5d 498
(Supplemental Figure 4). Like Rab5a, Rab5c is a conventional type, while Rab5b and 499
Rab5d are plant-specific type homologous to the Arabidopsis plant-specific 500
Rab5F1/ARA6. During rice growth and development, the loss of Rab5a is offset by 501
Rab5c and possibly by Rab5b and Rab5d. Although the other Rab5 isoforms can 502
compensate for Rab5a, they are only able to partially fill this role as rab5a rice lines 503
grow slower and flower later than normal, likely due to their significant lower expression 504
compared to Rab5a (Supplemental Figure 5). At the grain filling stage where there is 505
massive protein transport from the ER via the Golgi to the storage vacuole, the reduced 506

17
expression of these other Rab5 isoforms fails to maintain normal membrane trafficking 507
resulting in the secretion of storage proteins, the formation of PMBs, and mis-localization 508
of glutelin mRNAs. 509
Efforts to identify rice lines expressing defective NSF have not been productive, 510
likely because of the importance of its ATPase activity in membrane fusion. Mutations in 511
NSF were reported to induce severe defect and cell lethality in several organism 512
(Boulianne and Trimble, 1995; Golby et al., 2001; Mohtashami et al., 2001; Horsnell et 513
al., 2002; Zhao et al., 2007). For example, Drosophila expresses two NSF isoforms, 514
dNSF1 and dNSF2. dNSF1 is dominant in the adult central nervous system while dNSF2 515
is broadly distributed at the larval/adult stages of development, respectively (Boulianne et 516
al., 1995). While dNSF1 null mutants perish as pharate adults, dNSF2 deletion mediates a 517
recessive lethal phenotype, which is not even rescued by the addition of a dNSF2 518
transgene (Golby et al., 2001; Mohtashami et al., 2001). In yeast, loss-of-function 519
mutation of the NSF gene, Sec18, resulted in a dominant lethal phenotype (Horsnell et al., 520
2002). These studies suggest that NSF mutations are pleiotropic and cause severe growth 521
problems. In Arabidopsis, even a subtle mutation of NSF caused severe abnormal Golgi 522
morphology (Tanabashi et al., 2018). Given that NSF is coded by a single gene copy in 523
the rice genome, mutations that affect NSF activity will likely confer a strong lethal 524
phenotype in rice. 525
Although beyond the scope of this study, NSF mutations that affect binding to RBP-P 526
would be a viable approach to obtain further insight on its role in mRNA trafficking. As 527
discussed earlier, the NSF-N domain is likely responsible for its interaction with the N-528
terminus of RBP-P. Selected residues in the NSF-N region can be replaced by amino 529
acids that alter charge or conformation and then tested for its protein-interactive 530
properties by yeast 2-hybrid analysis. Mutations in NSF that abolish its interaction with 531
RBP-P but not with Rab5a would be potential sites for genetic alteration by CRISPR 532
technology and, thereby, disrupting glutelin mRNA transport but maintaining normal 533
function in membrane vesicle transport. 534
Overall, this study provides evidence on how glutelin mRNPs are able to 535
hitchhike on trafficking endosomes in rice endosperm cells by exploiting the binding 536
properties of RBPs and membrane trafficking factors, NSF and Rab5a. These findings 537

18
will provide the basis for future research on membrane trafficking-mediated mRNA 538
transport and the unique functions of NSF and Rab5a in this cellular process. 539
540
MATERIALS AND METHODS 541 542 Plant materials and growth conditions. The rice wildtype (Oryza sativa japonica 543
variety TC65) and rab5a mutant line EM960 (Fukuda et al., 2011) were potted in 544
Sunshine Brand #1 soil (Sungro Horticulture) and grown in walk-in growth chambers 545
with a diurnal cycle of 12 h light/12 h dark at 27°C and a lighting intensity of 400 to 700 546
μmol m-2
s-1
using a combination of metal halide and high pressure sodium lamps . 547
Antibodies. Full length cDNAs encoding RBP-P, RBP-L, NSF, SNAP, BiP and GFP 548
were cloned into pET30a for His-tagged fusion protein expression. The His-tagged 549
proteins were then purified and used to immunize New Zealand White rabbits for 550
antibody production. Anti-Rab5a rabbit and mouse antibodies were obtained from an 551
earlier study (Fukuda et al., 2011). For immunofluorescence labeling, Alexa Fluor-488 552
labeled goat anti-mouse IgG antibody (Invitrogen, Cat # A32723) and Alexa Fluor-594 553
labeled goat anti-rabbit IgG antibody (Invitrogen, Cat # A32740) were used as secondary 554
antibodies. For immunogold labeling, EM-grade 10 nm-gold-conjugated goat anti-mouse 555
IgG (Electron Microscopy Science, Cat # 25128) and EM-grade 15 nm-gold-conjugated 556
goat anti-rabbit IgG (Electron Microscopy Science, Cat # 25112) were used as secondary 557
antibodies. 558
Immunoprecipitation (IP) analysis. Most IP experiments were conducted as previously 559
described (Doroshenk et al., 2014) except that incubations with antibody were conducted 560
in the presence/absence of 1mM ATP/GTP, 8 mM MgCl2 or EDTA. To maximize 561
capture of transient mRNP complexes on endosomes, developing seeds were treated with 562
1% paraformaldehyde (PFA). Detailed procedures for IP-MS, the identification of NSF 563
protein, and PFA fixation for the enhanced IP are described below. 564
IP-MS analyses. Affinity purified RBP-P or GFP antibodies were prepared using 565
immobilized metal affinity chromatography based on irreversibly oxidized Co(III)-IDA 566
resin as described previously (Crofts et al., 2010) and crosslinked to Protein A/G agarose 567
resin using the Pierce Crosslink Immunoprecipitation Kit according to the manufacturer’s 568
instructions (Thermo Fisher Scientific). Protein extraction and co-IP experiments were 569

19
performed at 4oC unless indicated. Two g of dehulled mid-developing wild type rice seed 570
harvested 12-14 days after flowering were frozen in liquid nitrogen and grounded to a 571
powder. Proteins were extracted in 6 ml of IP buffer (20 mM Tris-HCl pH 7.5, 0.15 M 572
NaCl, 1 mM EDTA, 0.5% v/v NP40) containing 1X protease inhibitor cocktail and 0.5X 573
phosphatase inhibitor (Sigma-Aldrich). The crude extract was clarified by twice 574
centrifuging at 12,000 g for 10 min. The resulting supernatant was gently rotated 575
overnight with agarose resin to eliminate non-specific interactions. 700 µL of the 576
unbound protein fraction were added to columns containing RBP-P or GFP antibodies 577
crosslinked to Protein A/G resin or naked resin and incubated approximately 7 hours with 578
rotation. The columns were washed 5 times with IP buffer and once with 1X conditioning 579
buffer, and bound proteins were eluted with 50 µL IgG elution buffer (Thermo Fisher 580
Scientific). The antibody conjugated Protein A/G resin columns were regenerated by 581
washing with 1X Coupling Buffer (Thermo Scientific) followed by IP buffer. Fresh, pre-582
cleared rice seed extract was added and incubated overnight. The columns were washed 583
and proteins eluted as above. 584
Eluted proteins from a total of five co-IP experiments using anti-RBP-P, anti-GFP 585
antibodies, or naked resin were pooled, precipitated with addition of 100% trichloroacetic 586
acid (TCA) to a 20% final concentration. The TCA precipitate was washed with acetone 587
and then resuspended in 30 µL SDS sample buffer containing 4 M urea and 5% v/v β-588
mercaptoethanol. Protein samples were resolved on 10% SDS-PAGE gels and stained 589
using a silver nitrate staining protocol (Chevallet et al., 2006). Because the protein profile 590
of the minus antibody control IP looked very similar to the GFP Co-IP, proteins from 591
only the GFP and RBP-P co-IPs were analyzed by mass spectrometry. Gel slices 592
corresponding to similar positions within each lane were excised, in-gel trypsin digested, 593
and subjected to liquid chromatography-tandem mass spectrometry as described 594
previously (Doroshenk et al., 2009). Proteins were identified by searching the Oryza 595
sativa NCBI non-redundant database (134548 sequences) using Mascot 596
(www.matrixscience.com) as previously described (Doroshenk et al., 2009). 597
IP with 1% PFA fixation. Antibodies raised against RBP-L, RBP-P, NSF, or Rab5a 598
were affinity purified as abovementioned. 20µL of resuspended protein A Magbeads 599
slurry (25% w/v slurry, GenScript) in a 1.5 mL tube was washed twice with 1X PBS and 600

20
then incubated overnight at 4 ºС under rotation with 30 µg affinity-purified antibody 601
(diluted to 500 µL by 1x PBS). Antibodies were cross-linked to protein A Magbeads 602
using the above-mentioned Pierce Crosslink Immunoprecipitation Kit. 603
Dehulled developing rice seeds were fixed with 1% paraformaldehyde (PFA) in 604
PBSM (1.76 mM KH2PO4, 10 mM Na2HPO4, 136 mM NaCl, 2.6 mM KCl, 5 mM 605
MgCl2, and 10% glycerol) under vacuum for 30 min. The reaction was stopped by 606
vacuuming in quenching buffer (0.333 M Tris and 10% glycerol) for 10 min. Ten to 607
twenty aleurone-layer peels collected from the fixed seeds were ground to a fine powder 608
using an ice cold mortar and pestle in 800 µL of lysis buffer (50 mM Tris-HCl pH 7.5, 609
150 mM NaCl, 10 mM EDTA, 0.5% NP-40, 0.1% Triton X-100, 1x proteinase inhibitor 610
cocktail, 100 µg/ml cycloheximide, 20 units/mL of RiboLock RNase Inhibitor (Thermo 611
Fisher Scientific) and then centrifuged at 1000 g for 5 min to remove starch followed by 612
centrifugation at 12,000 g for 10 min. The clear supernatant was added to the prepared 613
Magbeads for incubation overnight at 4ºС with gentle rotation. After washing twice with 614
lysis buffer, the bound fractions were eluted with IgG elution buffer (ThermoFisher 615
Scientific) and neutralized by addition of 1/10 volume of 1 M Tris-HCl (pH 8.8). The 616
neutralized elution samples were analyzed by immunoblotting. 617
In situ RT-PCR. In situ RT-PCR on developing rice seed sections was performed as 618
previously described (Washida et al., 2009). Specific primers Glutelin-F 5’-619
CCCTCAAGCATACAGGCGTG-3’ and Glutelin-R 5’-620
CGCTCTCTTGATTGCACTTGTCC-3’ were used in the PCR to amplify glutelin RNAs. 621
Construction of vectors. Gene sequences of RBP-P, NSF and their truncated forms were 622
cloned into pGAD T7 and pGBK T7 vectors and used as preys and baits, respectively, in 623
Y2H analyses for detection of protein-protein interaction. BiFC vectors of pSAT1-624
nEYFP-C1 and pSAT1-cEYFP-C1-B for N- and C-terminal EYFP fusion, respectively, 625
were obtained from the Arabidopsis Biological Resource Center 626
(https://www.arabidopsis.org). The cDNA sequences of RBP-P, RBP-L, NSF, Rab5a and 627
its mutant forms (Rab5aQ70L
and Rab5aG45D
) were cloned into pSAT1-nEYFP-C1 or 628
pSAT1-cEYFP-C1-B vectors for BiFC analysis. GFP or RFP-fusion vector driven by 629
double 35S promoters was constructed by replacing the N-terminal EYFP in pSAT1-630

21
nEYFP-C1 with GFP or RFP cDNA sequences. The cDNA sequences of NSF and Rab5a 631
were then cloned into GFP or RFP-fusion vector to obtain RFP protein fusions. 632
RNA-IP, Yeast two hybrid (Y2H), and BiFC assays. Experimental procedures for 633
these studies were performed as previously described (Doroshenk et al., 2014; Tian et al., 634
2018; Tian et al., 2019a). Briefly, for RNA-IP analysis, developing rice seeds, collected 635
10-14 days after flowering, were subjected to 1% PFA fixation and extracts prepared and 636
used for IP as mentioned above. The elution samples obtained from IP were incubated at 637
70 °C for 45 min to reverse the RNA-protein crosslinks followed by RNA extraction 638
using TRIzol (Invitrogen), cDNA synthesis using M-MLV reverse transcriptase 639
(Promega), and PCR with 20-25 cycles of amplification using glutelin and ACTIN 640
specific primers (Glutelin-F and Glutelin-R as mention in in situ RT-PCR, Actin-F 5’- 641
TCCATCTTGGCATCTCTCAG-3’, and Actin-R 5’- GTACCCGCATCAGGCATCT-642
3’). The antibodies used in the RNA-IP were affinity-purified as aforementioned. In Y2H 643
analysis, synthetic dropout (SD) growth media without leucine and tryptophan (SD/-644
Leu/-Trp) was used to screen positive transformants and selection media without leucine, 645
tryptophan, histidine, and adenine (SD/-Leu/- Trp/-His/-Adenine medium) with 646
supplement of 3 mM 3-amintriazole (3-AT) (SD/-Leu/-Trp/-His/-Adenine/+ 3-AT) or 40 647
mg/L X-α-Gal was used to verify protein-protein interaction. 648
BY-2 suspension cells were used for BiFC analyses. In brief, BY-2 cells were 649
treated with cell wall digestion buffer (1% cellulase (Onozuka RS, PhytoTechnology 650
Laboratories), 0.05% pectolyase (Seishin Pharmaceutical, Japan), 0.2% Driselase 651
(Sigma-Aldrich), 20 mM KCl, 10 mM CaCl2, 20 mM MES hydrate, and 0.5M sucrose, 652
pH 5.7) at room temperature for 3 h. After washing with W5 solution (154 mM NaCl, 653
125 mM CaCl2, 5 mM KCl, 5 mM glucose, pH 5.8-6.0), the BY-2 protoplasts were 654
subject to PEG-mediated transformation with the abovementioned vectors of pSAT1-655
nEYFP-C1 and pSAT1-cEYFP-C1-B as described previously (Tian et al., 2018). After 656
culture at 26oC for 16 hours, the BiFC fluorescence images were observed using a Leica 657
SP-8 confocal microscope. Negative controls using empty vectors were also examined to 658
check the reliability of the transformation procedure. The localization pattern of target 659
proteins or complexes was determined by examining at least 5 different protoplast cells. 660
To confirm the involvement of Rab5a and the corresponding complexes in endocytic 661

22
pathway, protoplast incubation was treated with the endocytic tracer FM4-64 (Invitrogen) 662
at a final concentration of 10 μM for 15-30 mins before observation. 663
Microscopy. Light microscopy was performed on 10 μm thick sections of developing 664
rice seed samples embedded in LR-white resin. The sections were positioned on Leica X-665
tra slides, stained by 1% Toluidine blue and observed using an Olympus BH-2 Light 666
microscope. Co-localization test of RBP-P, RBP-L, and NSF with Rab5a in rice 667
endosperm cells was performed through double-immunolabeling using the rabbit anti-668
RBP-P, RBP-L, NSF antibodies and mouse anti-Rab5a antibodies (see Section of 669
Antibodies) on 1 μm thick LR-white sections as described previously (Fukuda et al., 670
2011), and observed under a Leitz Epi-Fluorescent Microscope with Leica DFC425C 671
Camera. Fluorescence intensity of green and red signals was analyzed by plot profile tool 672
in FUJI (ImageJ) software. Transmission electron microscopy analysis was performed as 673
previously described (Tian et al., 2018). 674
675
ACCESSION NUMBERS 676
Sequence data from this article can be found in the GenBank/EMBL data libraries under 677
NCBI accession numbers shown in the legends of Supplemental Figures 3 and 4. 678
679
SUPPLEMENTAL DATA 680
Supplemental Figure 1. Sequence information of Rab5a in rice. 681
Supplemental Figure 2. Possible binding domains of RBP-P with NSF revealed by yeast 682
two hybrid (Y2H) analysis. 683
Supplemental Figure 3. Protein sequence alignment of NSF. 684
Supplemental Figure 4. Protein sequence alignment of Rab5 isoforms. 685
Supplemental Figure 5. Expression profile of Rab5 isoforms in rice plants. 686
687
ACKNOWLEDGEMENTS 688
This work was financially supported by grants from the National Science Foundation 689
(MCB-1444610 and IOS-1701061), from the USDA National Institute of Food and 690

23
Agriculture, Hatch umbrella project 899 1015621 and project WNP00119, and from the 691
Japan Society for the Promotion of Science (M.F. and T.K.). We thank Ai Nagamine for 692
her help to construction of BiFC plasmids and technical support provided by the 693
Franceschi Microscopy and Imaging Center at Washington State University. 694
695
AUTHOR CONTRIBUTIONS 696
L.T. designed the study; K.A.D. identified NSF as interacting partner of RBP-P 697
through IP-MS; L.T. discovered interaction of RBPs and NSF with Rab5a and conducted 698
BiFC, RNA-IP, yeast two hybrid, light microscopy and TEM analyses; L.T. and L.Z. 699
conducted co-IP analysis; M.F. conducted immunofluorescence microscopy; L.T. 700
constructed vectors; L.T. and H.W. conducted in situ RT-PCR; T.K. provided rab5a 701
mutant; T.W.O. supervised the project; L.T. and T.W.O. wrote the manuscript. 702
703

24
Reference 704 705
Barnard, R.J., Morgan, A., and Burgoyne, R.D. (1997). Stimulation of NSF ATPase 706 activity by alpha-SNAP is required for SNARE complex disassembly and 707 exocytosis. J Cell Biol 139, 875-883. 708
Baumann, S., Pohlmann, T., Jungbluth, M., Brachmann, A., and Feldbrugge, M. 709 (2012). Kinesin-3 and dynein mediate microtubule-dependent co-transport 710 of mRNPs and endosomes. J Cell Sci 125, 2740-2752. 711
Blower, M.D. (2013). Molecular insights into intracellular RNA localization. Int Rev 712 Cell Mol Biol 302, 1-39. 713
Boulianne, G.L., and Trimble, W.S. (1995). Identification of a second homolog of N-714 ethylmaleimide-sensitive fusion protein that is expressed in the nervous 715 system and secretory tissues of Drosophila. Proc Natl Acad Sci U S A 92, 716 7095-7099. 717
Chang, L.F., Chen, S., Liu, C.C., Pan, X., Jiang, J., Bai, X.C., Xie, X., Wang, H.W., and 718 Sui, S.F. (2012). Structural characterization of full-length NSF and 20S 719 particles. Nat Struct Mol Biol 19, 268-275. 720
Chavrier, P., Parton, R.G., Hauri, H.P., Simons, K., and Zerial, M. (1990). 721 Localization of low molecular weight GTP binding proteins to exocytic and 722 endocytic compartments. Cell 62, 317-329. 723
Chevallet, M., Luche, S., and Rabilloud, T. (2006). Silver staining of proteins in 724 polyacrylamide gels. Nat Protoc 1, 1852-1858. 725
Chou, H.L., Tian, L., Washida, H., Fukuda, M., Kumamaru, T., and Okita, T.W. 726 (2019). The rice storage protein mRNAs as a model system for RNA 727 localization in higher plants. Plant Sci 284, 203-211. 728
Corradi, E., Dalla Costa, I., Gavoci, A., Iyer, A., Roccuzzo, M., Otto, T.A., Oliani, E., 729 Bridi, S., Strohbuecker, S., Santos-Rodriguez, G., Valdembri, D., Serini, G., 730 Abreu-Goodger, C., and Baudet, M.-L. (2020). Axonal precursor miRNAs 731 hitchhike on endosomes and locally regulate the development of neural 732 circuits. The EMBO Journal 39, e102513. 733
Crofts, A.J., Crofts, N., Whitelegge, J.P., and Okita, T.W. (2010). Isolation and 734 identification of cytoskeleton-associated prolamine mRNA binding proteins 735 from developing rice seeds. Planta 231, 1261-1276. 736
Doroshenk, K.A., Crofts, A.J., Morris, R.T., Wyrick, J.J., and Okita, T.W. (2009). 737 Proteomic analysis of cytoskeleton-associated RNA binding proteins in 738 developing rice seed. J Proteome Res 8, 4641-4653. 739
Doroshenk, K.A., Crofts, A.J., Morris, R.T., Wyrick, J.J., and Okita, T.W. (2012). 740 RiceRBP: A Resource for Experimentally Identified RNA Binding Proteins in 741 Oryza sativa. Front Plant Sci 3, 90. 742
Doroshenk, K.A., Tian, L., Crofts, A.J., Kumamaru, T., and Okita, T.W. (2014). 743 Characterization of RNA binding protein RBP-P reveals a possible role in rice 744 glutelin gene expression and RNA localization. Plant Mol Biol 85, 381-394. 745
Doroshenk, K.A., Crofts, A.J., Washida, H., Satoh-Cruz, M., Crofts, N., Sugino, A., 746 Okita, T.W., Morris, R.T., Wyrick, J.J., Fukuda, M., Kumamaru, T., and 747 Satoh, H. (2010). Characterization of the rice glup4 mutant suggests a role 748

25
for the small GTPase Rab5 in the biosynthesis of carbon and nitrogen storage 749 reserves in developing endosperm. Breeding Science 60, 556-567. 750
Fischer von Mollard, G., Stahl, B., Walch-Solimena, C., Takei, K., Daniels, L., 751 Khoklatchev, A., De Camilli, P., Sudhof, T.C., and Jahn, R. (1994). 752 Localization of Rab5 to synaptic vesicles identifies endosomal intermediate 753 in synaptic vesicle recycling pathway. Eur J Cell Biol 65, 319-326. 754
Fukuda, M., Satoh-Cruz, M., Wen, L., Crofts, A.J., Sugino, A., Washida, H., Okita, 755 T.W., Ogawa, M., Kawagoe, Y., Maeshima, M., and Kumamaru, T. (2011).756 The small GTPase Rab5a is essential for intracellular transport of proglutelin757 from the Golgi apparatus to the protein storage vacuole and endosomal758 membrane organization in developing rice endosperm. Plant Physiol 157,759 632-644.760
Gohre, V., Vollmeister, E., Bolker, M., and Feldbrugge, M. (2012). Microtubule-761 dependent membrane dynamics in Ustilago maydis: Trafficking and function 762 of Rab5a-positive endosomes. Commun Integr Biol 5, 485-490. 763
Golby, J.A., Tolar, L.A., and Pallanck, L. (2001). Partitioning of N-ethylmaleimide-764 sensitive fusion (NSF) protein function in Drosophila melanogaster: dNSF1 is 765 required in the nervous system, and dNSF2 is required in mesoderm. 766 Genetics 158, 265-278. 767
Grosshans, B.L., Ortiz, D., and Novick, P. (2006). Rabs and their effectors: 768 achieving specificity in membrane traffic. Proc Natl Acad Sci U S A 103, 769 11821-11827. 770
Guo, Y., Yue, Q., Gao, J., Wang, Z., Chen, Y.R., Blissard, G.W., Liu, T.X., and Li, Z. 771 (2017). Roles of Cellular NSF Protein in Entry and Nuclear Egress of Budded 772 Virions of Autographa californica Multiple Nucleopolyhedrovirus. J Virol 91. 773
Haag, C., Steuten, B., and Feldbrugge, M. (2015). Membrane-Coupled mRNA 774 Trafficking in Fungi. Annu Rev Microbiol 69, 265-281. 775
Hamada, S., Ishiyama, K., Sakulsingharoj, C., Choi, S.B., Wu, Y., Wang, C., Singh, 776 S., Kawai, N., Messing, J., and Okita, T.W. (2003). Dual regulated RNA 777 transport pathways to the cortical region in developing rice endosperm. 778 Plant Cell 15, 2265-2272. 779
Hanson, P.I., Otto, H., Barton, N., and Jahn, R. (1995). The N-ethylmaleimide-780 sensitive fusion protein and alpha-SNAP induce a conformational change in 781 syntaxin. J Biol Chem 270, 16955-16961. 782
Herbert, S.P., and Costa, G. (2019). Sending messages in moving cells: mRNA 783 localization and the regulation of cell migration. Essays Biochem 63, 595-784 606. 785
Herve, J.C., and Bourmeyster, N. (2018). Rab GTPases, master controllers of 786 eukaryotic trafficking. Small GTPases 9, 1-4. 787
Horsnell, W.G., Steel, G.J., and Morgan, A. (2002). Analysis of NSF mutants reveals 788 residues involved in SNAP binding and ATPase stimulation. Biochemistry 41, 789 5230-5235. 790
Hughes, S.C., and Simmonds, A.J. (2019). Drosophila mRNA Localization During 791 Later Development: Past, Present, and Future. Front Genet 10, 135. 792

26
Ito, E., Ebine, K., Choi, S.W., Ichinose, S., Uemura, T., Nakano, A., and Ueda, T. 793 (2018). Integration of two RAB5 groups during endosomal transport in 794 plants. Elife 7. 795
Jansen, R.P., Niessing, D., Baumann, S., and Feldbrugge, M. (2014). mRNA 796 transport meets membrane traffic. Trends Genet 30, 408-417. 797
Kerppola, T.K. (2006). Design and implementation of bimolecular fluorescence 798 complementation (BiFC) assays for the visualization of protein interactions 799 in living cells. Nat Protoc 1, 1278-1286. 800
Lee, M.T., Mishra, A., and Lambright, D.G. (2009). Structural mechanisms for 801 regulation of membrane traffic by rab GTPases. Traffic 10, 1377-1389. 802
Li, Y., Wang, S., Li, T., Zhu, L., and Ma, C. (2018). Tomosyn guides SNARE complex 803 formation in coordination with Munc18 and Munc13. FEBS Lett 592, 1161-804 1172. 805
Liao, Y.C., Fernandopulle, M.S., Wang, G., Choi, H., Hao, L., Drerup, C.M., Patel, R., 806 Qamar, S., Nixon-Abell, J., Shen, Y., Meadows, W., Vendruscolo, M., 807 Knowles, T.P.J., Nelson, M., Czekalska, M.A., Musteikyte, G., 808 Gachechiladze, M.A., Stephens, C.A., Pasolli, H.A., Forrest, L.R., St George-809 Hyslop, P., Lippincott-Schwartz, J., and Ward, M.E. (2019). RNA Granules 810 Hitchhike on Lysosomes for Long-Distance Transport, Using Annexin A11 as 811 a Molecular Tether. Cell 179, 147-164 e120. 812
Mastick, C.C., and Falick, A.L. (1997). Association of N-ethylmaleimide sensitive 813 fusion (NSF) protein and soluble NSF attachment proteins-alpha and -gamma 814 with glucose transporter-4-containing vesicles in primary rat adipocytes. 815 Endocrinology 138, 2391-2397. 816
McBride, H.M., Rybin, V., Murphy, C., Giner, A., Teasdale, R., and Zerial, M. 817 (1999). Oligomeric complexes link Rab5 effectors with NSF and drive 818 membrane fusion via interactions between EEA1 and syntaxin 13. Cell 98, 819 377-386. 820
Miller, K.E., Kim, Y., Huh, W.K., and Park, H.O. (2015). Bimolecular Fluorescence 821 Complementation (BiFC) Analysis: Advances and Recent Applications for 822 Genome-Wide Interaction Studies. J Mol Biol 427, 2039-2055. 823
Mohtashami, M., Stewart, B.A., Boulianne, G.L., and Trimble, W.S. (2001). 824 Analysis of the mutant Drosophila N-ethylmaleimide sensitive fusion-1 825 protein in comatose reveals molecular correlates of the behavioural 826 paralysis. J Neurochem 77, 1407-1417. 827
Muench, D.G., Chuong, S.D., Franceschi, V.R., and Okita, T.W. (2000). Developing 828 prolamine protein bodies are associated with the cortical cytoskeleton in rice 829 endosperm cells. Planta 211, 227-238. 830
Muller, J., Pohlmann, T., and Feldbrugge, M. (2019). Core components of 831 endosomal mRNA transport are evolutionarily conserved in fungi. Fungal 832 Genet Biol 126, 12-16. 833
Niessing, D., Jansen, R.P., Pohlmann, T., and Feldbrugge, M. (2018). mRNA 834 transport in fungal top models. Wiley Interdiscip Rev RNA 9. 835
Pohlmann, T., Baumann, S., Haag, C., Albrecht, M., and Feldbrugge, M. (2015). A 836 FYVE zinc finger domain protein specifically links mRNA transport to 837 endosome trafficking. Elife 4. 838

27
Pylypenko, O., Hammich, H., Yu, I.M., and Houdusse, A. (2018). Rab GTPases and 839 their interacting protein partners: Structural insights into Rab functional 840 diversity. Small GTPases 9, 22-48. 841
Ryu, J.K., Min, D., Rah, S.H., Kim, S.J., Park, Y., Kim, H., Hyeon, C., Kim, H.M., Jahn, 842 R., and Yoon, T.Y. (2015). Spring-loaded unraveling of a single SNARE 843 complex by NSF in one round of ATP turnover. Science 347, 1485-1489. 844
Saito, C., and Ueda, T. (2009). Chapter 4: functions of RAB and SNARE proteins in 845 plant life. Int Rev Cell Mol Biol 274, 183-233. 846
Schmid, M., Jaedicke, A., Du, T.G., and Jansen, R.P. (2006). Coordination of 847 endoplasmic reticulum and mRNA localization to the yeast bud. Curr Biol 16, 848 1538-1543. 849
Stenmark, H., and Olkkonen, V.M. (2001). The Rab GTPase family. Genome Biol 2, 850 REVIEWS3007. 851
Tagaya, M., Wilson, D.W., Brunner, M., Arango, N., and Rothman, J.E. (1993). 852 Domain structure of an N-ethylmaleimide-sensitive fusion protein involved 853 in vesicular transport. J Biol Chem 268, 2662-2666. 854
Tanabashi, S., Shoda, K., Saito, C., Sakamoto, T., Kurata, T., Uemura, T., and 855 Nakano, A. (2018). A Missense Mutation in the NSF Gene Causes Abnormal 856 Golgi Morphology in Arabidopsis thaliana. Cell Struct Funct 43, 41-51. 857
Tian, L., and Okita, T.W. (2014). mRNA-based protein targeting to the endoplasmic 858 reticulum and chloroplasts in plant cells. Curr Opin Plant Biol 22, 77-85. 859
Tian, L., Chou, H.L., Zhang, L., and Okita, T.W. (2019a). Targeted Endoplasmic 860 Reticulum Localization of Storage Protein mRNAs Requires the RNA-Binding 861 Protein RBP-L. Plant Physiol 179, 1111-1131. 862
Tian, L., Chou, H.L., Fukuda, M., Kumamaru, T., and Okita, T.W. (2019b). mRNA 863 localization in plant cells. Plant Physiol. 864
Tian, L., Chou, H.L., Fukuda, M., Kumamaru, T., and Okita, T.W. (2020). mRNA 865 Localization in Plant Cells. Plant Physiol 182, 97-109. 866
Tian, L., Chou, H.L., Zhang, L., Hwang, S.K., Starkenburg, S.R., Doroshenk, K.A., 867 Kumamaru, T., and Okita, T.W. (2018). RNA-Binding Protein RBP-P Is 868 Required for Glutelin and Prolamine mRNA Localization in Rice Endosperm 869 Cells. Plant Cell 30, 2529-2552. 870
Ueda, T., Yamaguchi, M., Uchimiya, H., and Nakano, A. (2001). Ara6, a plant-871 unique novel type Rab GTPase, functions in the endocytic pathway of 872 Arabidopsis thaliana. EMBO J 20, 4730-4741. 873
Vida, T.A., and Emr, S.D. (1995). A new vital stain for visualizing vacuolar 874 membrane dynamics and endocytosis in yeast. J Cell Biol 128, 779-792. 875
Vollmeister, E., Schipper, K., and Feldbrugge, M. (2012). Microtubule-dependent 876 mRNA transport in the model microorganism Ustilago maydis. RNA Biol 9, 877 261-268.878
Wang, Y., Ren, Y., Liu, X., Jiang, L., Chen, L., Han, X., Jin, M., Liu, S., Liu, F., Lv, J., 879 Zhou, K., Su, N., Bao, Y., and Wan, J. (2010). OsRab5a regulates 880 endomembrane organization and storage protein trafficking in rice 881 endosperm cells. Plant J 64, 812-824. 882
Washida, H., Kaneko, S., Crofts, N., Sugino, A., Wang, C., and Okita, T.W. (2009). 883 Identification of cis-localization elements that target glutelin RNAs to a 884

28
specific subdomain of the cortical endoplasmic reticulum in rice endosperm 885 cells. Plant Cell Physiol 50, 1710-1714. 886
Washida, H., Sugino, A., Doroshenk, K.A., Satoh-Cruz, M., Nagamine, A., 887 Katsube-Tanaka, T., Ogawa, M., Kumamaru, T., Satoh, H., and Okita, T.W. 888 (2012). RNA targeting to a specific ER sub-domain is required for efficient 889 transport and packaging of alpha-globulins to the protein storage vacuole in 890 developing rice endosperm. Plant J 70, 471-479. 891
Weis, B.L., Schleiff, E., and Zerges, W. (2013). Protein targeting to subcellular 892 organelles via MRNA localization. Biochim Biophys Acta 1833, 260-273. 893
Wen, L., Fukuda, M., Sunada, M., Ishino, S., Ishino, Y., Okita, T.W., Ogawa, M., 894 Ueda, T., and Kumamaru, T. (2015). Guanine nucleotide exchange factor 2 895 for Rab5 proteins coordinated with GLUP6/GEF regulates the intracellular 896 transport of the proglutelin from the Golgi apparatus to the protein storage 897 vacuole in rice endosperm. J Exp Bot 66, 6137-6147. 898
Woodman, P.G. (2000). Biogenesis of the sorting endosome: the role of Rab5. 899 Traffic 1, 695-701. 900
Yang, Y., Chou, H.L., Crofts, A.J., Zhang, L., Tian, L., Washida, H., Fukuda, M., 901 Kumamaru, T., Oviedo, O.J., Starkenburg, S.R., and Okita, T.W. (2018). 902 Selective sets of mRNAs localize to extracellular paramural bodies in a rice 903 glup6 mutant. J Exp Bot 69, 5045-5058. 904
Yuan, W., and Song, C. (2020). The Emerging Role of Rab5 in Membrane Receptor 905 Trafficking and Signaling Pathways. Biochem Res Int 2020, 4186308. 906
Zeigerer, A., Gilleron, J., Bogorad, R.L., Marsico, G., Nonaka, H., Seifert, S., 907 Epstein-Barash, H., Kuchimanchi, S., Peng, C.G., Ruda, V.M., Del Conte-908 Zerial, P., Hengstler, J.G., Kalaidzidis, Y., Koteliansky, V., and Zerial, M. 909 (2012). Rab5 is necessary for the biogenesis of the endolysosomal system in 910 vivo. Nature 485, 465-470. 911
Zhao, C., Slevin, J.T., and Whiteheart, S.W. (2007). Cellular functions of NSF: not 912 just SNAPs and SNAREs. FEBS Lett 581, 2140-2149. 913
Zhao, C., Matveeva, E.A., Ren, Q., and Whiteheart, S.W. (2010). Dissecting the N-914 ethylmaleimide-sensitive factor: required elements of the N and D1 domains. 915 J Biol Chem 285, 761-772. 916
917 918 919
920

29
921 FIGURE LEGENDS 922 923 Figure 1. Identification of NSF as an interacting partner of RBP-P. (A) Precipitation 924 of NSF by RBP-P antibody as revealed by IP-MS. Left panel, immunoblot (IB) analysis 925 to test the IP reliability; right panel, silver stained SDS- polyacrylamide gel of eluted 926 samples from ()-GFP and RBP-P IPs. Input, starting material of rice lysate; Ub, 927 unbound fraction from IPs; B, bound fraction (eluted samples) from IPs. Blue asterisk (*) 928 indicates a modified form of RBP-P. The bands indicated by red and black arrows were 929 excised for MS analysis and NSF was identified as a specific protein precipitated by anti-930 RBP-P but not by anti-GFP. (B) Interaction between RBP-P and NSF revealed by yeast 931 two hybrid. Yeast colonies co-transfected with pGBK and pGAD constructs were labeled 932 1-4 as described in the upper Table. --, empty vector. Yeast cells carrying the 933 corresponding genes were grown on SD/-Leu/-Trp medium as growth control and SD/-934 Leu/-Trp/-His/-Ade/+ 3-AT selection medium to detect their interaction. Note that only 935 the yeast cells carrying both NSF and RBP-P survived on the strict selection medium 936 (lower panel), suggesting that RBP-P interacts with NSF. 937 938 Figure 2. RBP-P interacts indirectly with Rab5a through NSF, forming a complex 939 attached to endosomes. 940 (A-C) RBP-P interacts with NSF as revealed by BiFC studies. BF, bright field; Merge, 941 the merged images of BF and BiFC (middle panel); ---, empty vector as control. (D) 942 SNAP-independent formation of a protein complex consisting of RBP-P, NSF and Rab5a 943 as revealed by Co-IP. IP studies were conducted with ()-GFP, RBP-P, NSF, or Rab5a 944 antibodies in the presence of ATP and MgCl2 or with EDTA. GFP antibody was used as a 945 negative control. Input, the starting rice lysate. Note that the association of RBP-P with 946 NSF and Rab5a requires MgCl2 and ATP, unlike the interaction of SNAP with NSF and 947 Rab5a, which requires EDTA. (E) Schematic structure of Rab5a in rice. The two switch 948 regions are indicated as Switch 1 and 2. The two mutation sites (G45D and Q70L) to 949 generate GDP- and GTP-fixed forms of Rab5a, respectively, are indicated in red. (F) 950 GFP-Rab5a (green) co-localized with FM4-64 (magenta) labeled plasma membrane and 951 endosomal compartment. Co-localization of GFP-Rab5a with FM4-64 endosomes are 952 denoted by arrows. (G-J) Rab5a interacts with NSF (I), but not to RBP-P (J) as shown 953 by BiFC. (K-L) NSF interacts with both GDP- (panel J, Rab5a
G45D) and GTP-bound 954
(panel K, Rab5aQ70L
) Rab5a. (M-N) RBP-P/NSF complexes labeled by BiFC (yellow) 955 co-localize with RFP-Rab5a (magenta) in the cytoplasm. (M) The punctate RBP-P/NSF 956 structures align with the particle-like, endosome structures of RFP-Rab5a. (N) RBP-957 P/NSF complexes co-localize with the membrane-associated, GTP-fixed Rab5a
Q70L, 958
indicating RBP-P/NSF complexes are associated with active endosomes. In M-N, co-959 localized RBP-P/NSF complexes with endosomal Rab5a are indicated by white arrows. 960 Non-colocalization of RBP-P/NSF complexes and Rab5a-postive endosome are indicated 961 by empty arrowheads. (O) Co-localization of RBP-P/NSF complexes with FM4-64 962 labeled endosomes (magenta) are denoted by arrows. Scale bars, 20 μm. 963 964 Figure 3. RBP-L interacts directly with Rab5a, which collectively forms a multi-965 protein complexes with RBP-P and NSF to transport glutelin mRNAs on endosomes. 966

30
(A-C) RBP-L interacts directly with Rab5a (C) but not to NSF (B). Note that the RBP-967 L/Rab5a complex is detected in both the nucleus and cytoplasm. (D) RBP-L/Rab5a 968 complexes (yellow) co-localized with FM4-64 labeled endosomes (magenta) located 969 close to the plasma membrane. Co-localized signals are indicated by arrows. (E-F) in 970 vivo interactions of RBP-L with GDP- (F, Rab5a
G45D) or GTP-bound (G, Rab5a
Q70L)971
Rab5a. (G) Association of RBP-P, RBP-L with NSF and Rab5a as revealed by IP. The IP 972 experiment was conducted with developing seeds treated with 1% paraformaldehyde 973 (PFA) to preserve labile protein complexes. Input, the starting rice lysate. Control, GFP 974 antibody. IB, immunoblot. (H) RBP-P, RBP-L, NSF and Rab5a form a ternary complex 975 as revealed by sequential IPs. The first IP was conducted with ()-GFP (negative control) 976 or RBP-P antibodies. Proteins captured by RBP-P antibody were subjected to a 2
nd IP977
using RBP-L antibody. Subsequent immunoblot analysis showed the presence of NSF 978 and Rab5a together with RBP-P and RBP-L. Input, the starting rice lysate; FT, flow-979 through (unbound fraction); E, bound fraction; W1 and W2, washing fractions. (I) The 980 RBP-P/RBP-L/NSF/Rab5a complex contains glutelin mRNAs based on RNA-IP. RNAs 981 extracted from each IP generated by anti-RBP-P, anti-RBP-L, anti-GFP or empty resin (-982 CT) were subjected to RT-PCR using glutelin-specific primers and resolved by agarose 983 gel electrophoresis. GFP antibody (-GFP) and empty resin (-CT) were used as negative 984 controls. Input, PCR products using cDNA synthesized from total RNAs. Actin was used 985 as control genes to verify the specific binding of the complex to glutelin RNAs. Scale 986 bars, 20 μm. 987
988 Figure 4. A working model of glutelin mRNA transport via trafficking endosomes to 989 the cortical ER. 990 In rice endosperm cells, glutelin mRNA is bound by the scaffold complex of RBP-P and 991 RBP-L, forming a mRNP complex. Through direct or NSF-mediated indirect interaction 992 with membrane-associated Rab5a, RBP-L and RBP-P link the mRNP complex to 993 endosomes for active transport via the cytoskeleton. Rab5a effectors (e, light orange) may 994 be involved to stabilize the quaternary complex and modulate GTP-bound active Rab5a-995 drived endosomal trafficking on actin filaments in rice endosperm cells. Other unknown 996 RBPs or factors, showed in light grey shapes, may also be involved to constitute the 997 mRNP complex and define the linkage onto endosomes. 998
999 Figure 5. Rab5a mutation leads to abnormal trafficking of endosomes and 1000 formation of extracellular paramural bodies (PMBs). 1001 (A) Schematic representation of the Rab5a mutation site in the rab5a mutant. A G134A1002 base substitution within the Rab5a gene resulted in a G45D amino acid replacement. (B)1003 Formation of PMBs (white asterisks) was observed in endosperm cells of rab5a mutant1004 through light microscopy observations on seed sections stained with 1% Toluidine Blue.1005 Scale bar, 25 m. (C) Ultrastructure of PMBs formed in rab5a mutant due to aborted1006 endosomal trafficking in comparison to wildtype (WT) endosperm cells. Cell wall and1007 PMB boundaries are indicated by magenta and green dashed lines, respectively. SG,1008 starch granules; orange *, protein body I; blue *, protein storage vacuoles. Scale bar, 11009 m.1010
1011

31
Figure 6. Rab5a mutation leads to the transport of the RBP-P/RBP-L/NSF/Rab5a 1012 ternary complex to PMBs. 1013 (A-F) intracellular location of RBP-P (A-B, magenta), RBP-L (C-D, magenta), NSF (E-1014 F, magenta) and Rab5a (A-F, green) in wildtype (A, C, and E) and rab5a mutant (B, D, 1015 and F) rice endosperm cells as revealed by immunofluorescence labeling. Co-1016 localization patterns of RBP-P, RBP-L, and NSF with Rab5a are shown in the Merge 1017 panel, with rectangular areas enlarged in the adjacent fourth panels. Fluorescence 1018 intensity graphs on the very right show the relative strength of the magenta and green 1019 fluorescence signals as measured by scanning the region indicated by the white line in the 1020 fourth panel. X and y axes represent the fluorescence intensity and position of the signals 1021 (pixels), respectively, as evaluated by FUJI ImageJ. PMBs are indicated by asterisk (*) 1022 and co-localization signals are highlighted by white arrow heads. Scale bar, 20 m (the 1023 left three panels) and 10 m (the right panels). 1024 1025 Figure 7. Retention of RBP-P and RBP-L on the aborted endosomes in the PMBs of 1026 rab5a mutant as revealed by immunocytochemistry and transmission electron 1027 microcopy. 1028 (A-J) Localization of RBP-P (A-C, F-G) or RBP-L (D-E, H-J) on Rab5a-labeled 1029 endosomes in wildtype (A-E) and rab5a mutant (F-J) endosperm cells. RBPs (RBP-P 1030 and RBP-L) and Rab5a were labeled with 15 nm (blue arrowheads) and 10 nm (red 1031 arrows) gold particles, respectively. Panels B, C, E, G, I and J are the enlarged areas 1032 (rectangle) shown in A, D, F and H. Cell wall and PMB boundaries are indicated by red 1033 and yellow dashed lines, respectively. Scale bar, 2m (A, D, F and H) and 200 nm 1034 (B,C,E,G,I and J). Note that Rab5a-mediated endosomes are observed as electron-dense 1035 vesicles with an irregular-shape likely due to endosomal fusion (A to E) and as aborted 1036 endosome vesicles trapped within the PMBs in the rab5a mutant (F to J). 1037 1038 Figure 8. Rab5a mutation leads to mis-targeting of glutelin mRNAs to PB-ER and 1039 the PMBs as assessed by in situ RT-PCR. 1040 The PB-ER was stained by Rhodamine B dye (magenta), and glutelin mRNAs were 1041 labeled by in situ RT-PCR in the presence of Alexa-488-UTP (green). In wildtype (WT) 1042 endosperm cells (upper panel), glutelin mRNAs are localized on the cisternal-ER 1043 adjacent to the PB-ER. In rab5a mutant (lower panel), glutelin mRNAs are mis-targeted 1044 to the PMBs (asterisk) and PB-ER (arrows). Fluorescence intensity graphs on the very 1045 right show the relative position of magenta and green fluorescence signals in the regions 1046 indicated by rectangle in the Merge panels. X and y axes represent the fluorescence 1047 intensity and position of the signals (pixels), respectively, evaluated by FUJI ImageJ. 1048 Note the extensive overlap in the distributions between PB-ER and glutelin mRNAs in 1049 rab5a but not in wild type. Scale bar, 20m. 1050 1051

Figure 1. Identification of NSF as an interacting partner of RBP-P. (A) Precipitation of NSF by RBP-P antibody as revealed by IP-MS. Left panel, immunoblot (IB) analysis to test the IP reliability; right panel, silver stained SDS- polyacrylamide gel of eluted samples from (a)-GFP and RBP-P IPs. Input, starting material of rice lysate; Ub, unbound fraction from IPs; B, bound fraction (eluted samples) from IPs. Blue asterisk (*) indicates a modified form of RBP-P. The bands indicated by red and black arrows were excised for MS analysis and NSF was identified as a specific protein precipitated by anti-RBP-P but not by anti-GFP. (B) Interaction between RBP-P and NSF revealed by yeast two hybrid. Yeast colonies co-transfected with pGBK and pGAD constructs were labeled 1-4 as described in the upper Table. --, empty vector. Yeast cells carrying the corresponding genes were grown on SD/-Leu/-Trp medium as growth control and SD/-Leu/-Trp/-His/-Ade/+ 3-AT selection medium to detect their interaction. Note that only the yeast cells carrying both NSF and RBP-P survived on the strict selection medium (lower panel), suggesting that RBP-P interacts with NSF.
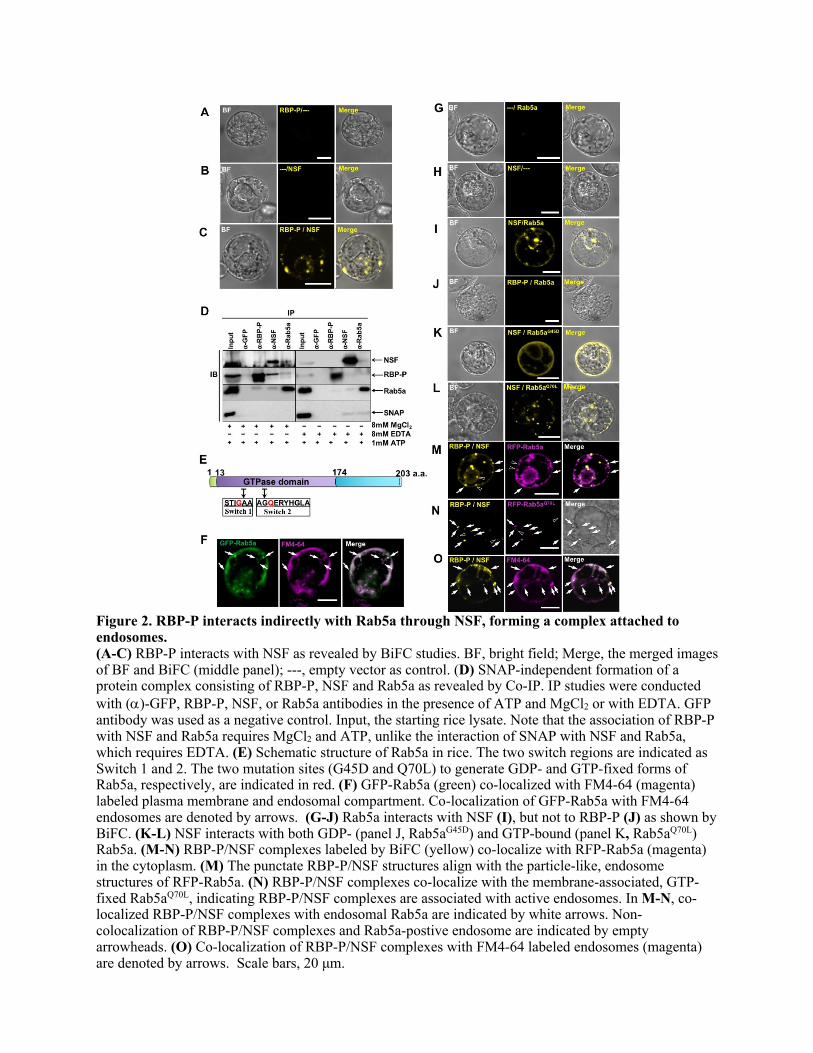
Figure 2. RBP-P interacts indirectly with Rab5a through NSF, forming a complex attached to endosomes. (A-C) RBP-P interacts with NSF as revealed by BiFC studies. BF, bright field; Merge, the merged images of BF and BiFC (middle panel); ---, empty vector as control. (D) SNAP-independent formation of a protein complex consisting of RBP-P, NSF and Rab5a as revealed by Co-IP. IP studies were conducted with (a)-GFP, RBP-P, NSF, or Rab5a antibodies in the presence of ATP and MgCl2 or with EDTA. GFP antibody was used as a negative control. Input, the starting rice lysate. Note that the association of RBP-P with NSF and Rab5a requires MgCl2 and ATP, unlike the interaction of SNAP with NSF and Rab5a, which requires EDTA. (E) Schematic structure of Rab5a in rice. The two switch regions are indicated as Switch 1 and 2. The two mutation sites (G45D and Q70L) to generate GDP- and GTP-fixed forms of Rab5a, respectively, are indicated in red. (F) GFP-Rab5a (green) co-localized with FM4-64 (magenta) labeled plasma membrane and endosomal compartment. Co-localization of GFP-Rab5a with FM4-64 endosomes are denoted by arrows. (G-J) Rab5a interacts with NSF (I), but not to RBP-P (J) as shown by BiFC. (K-L) NSF interacts with both GDP- (panel J, Rab5aG45D) and GTP-bound (panel K, Rab5aQ70L) Rab5a. (M-N) RBP-P/NSF complexes labeled by BiFC (yellow) co-localize with RFP-Rab5a (magenta) in the cytoplasm. (M) The punctate RBP-P/NSF structures align with the particle-like, endosome structures of RFP-Rab5a. (N) RBP-P/NSF complexes co-localize with the membrane-associated, GTP-fixed Rab5aQ70L, indicating RBP-P/NSF complexes are associated with active endosomes. In M-N, co-localized RBP-P/NSF complexes with endosomal Rab5a are indicated by white arrows. Non-colocalization of RBP-P/NSF complexes and Rab5a-postive endosome are indicated by empty arrowheads. (O) Co-localization of RBP-P/NSF complexes with FM4-64 labeled endosomes (magenta) are denoted by arrows. Scale bars, 20 μm.
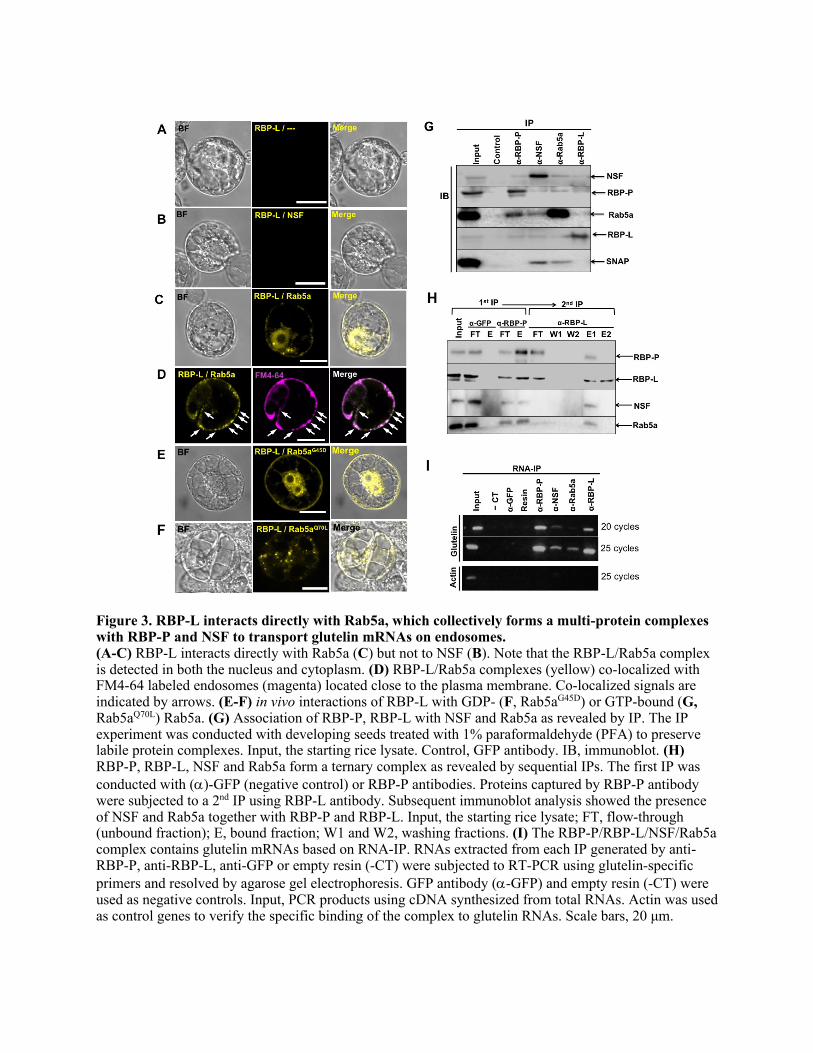
Figure 3. RBP-L interacts directly with Rab5a, which collectively forms a multi-protein complexes with RBP-P and NSF to transport glutelin mRNAs on endosomes. (A-C) RBP-L interacts directly with Rab5a (C) but not to NSF (B). Note that the RBP-L/Rab5a complex is detected in both the nucleus and cytoplasm. (D) RBP-L/Rab5a complexes (yellow) co-localized with FM4-64 labeled endosomes (magenta) located close to the plasma membrane. Co-localized signals are indicated by arrows. (E-F) in vivo interactions of RBP-L with GDP- (F, Rab5aG45D) or GTP-bound (G, Rab5aQ70L) Rab5a. (G) Association of RBP-P, RBP-L with NSF and Rab5a as revealed by IP. The IP experiment was conducted with developing seeds treated with 1% paraformaldehyde (PFA) to preserve labile protein complexes. Input, the starting rice lysate. Control, GFP antibody. IB, immunoblot. (H) RBP-P, RBP-L, NSF and Rab5a form a ternary complex as revealed by sequential IPs. The first IP was conducted with (a)-GFP (negative control) or RBP-P antibodies. Proteins captured by RBP-P antibody were subjected to a 2nd IP using RBP-L antibody. Subsequent immunoblot analysis showed the presence of NSF and Rab5a together with RBP-P and RBP-L. Input, the starting rice lysate; FT, flow-through (unbound fraction); E, bound fraction; W1 and W2, washing fractions. (I) The RBP-P/RBP-L/NSF/Rab5a complex contains glutelin mRNAs based on RNA-IP. RNAs extracted from each IP generated by anti-RBP-P, anti-RBP-L, anti-GFP or empty resin (-CT) were subjected to RT-PCR using glutelin-specific primers and resolved by agarose gel electrophoresis. GFP antibody (a-GFP) and empty resin (-CT) were used as negative controls. Input, PCR products using cDNA synthesized from total RNAs. Actin was used as control genes to verify the specific binding of the complex to glutelin RNAs. Scale bars, 20 μm.
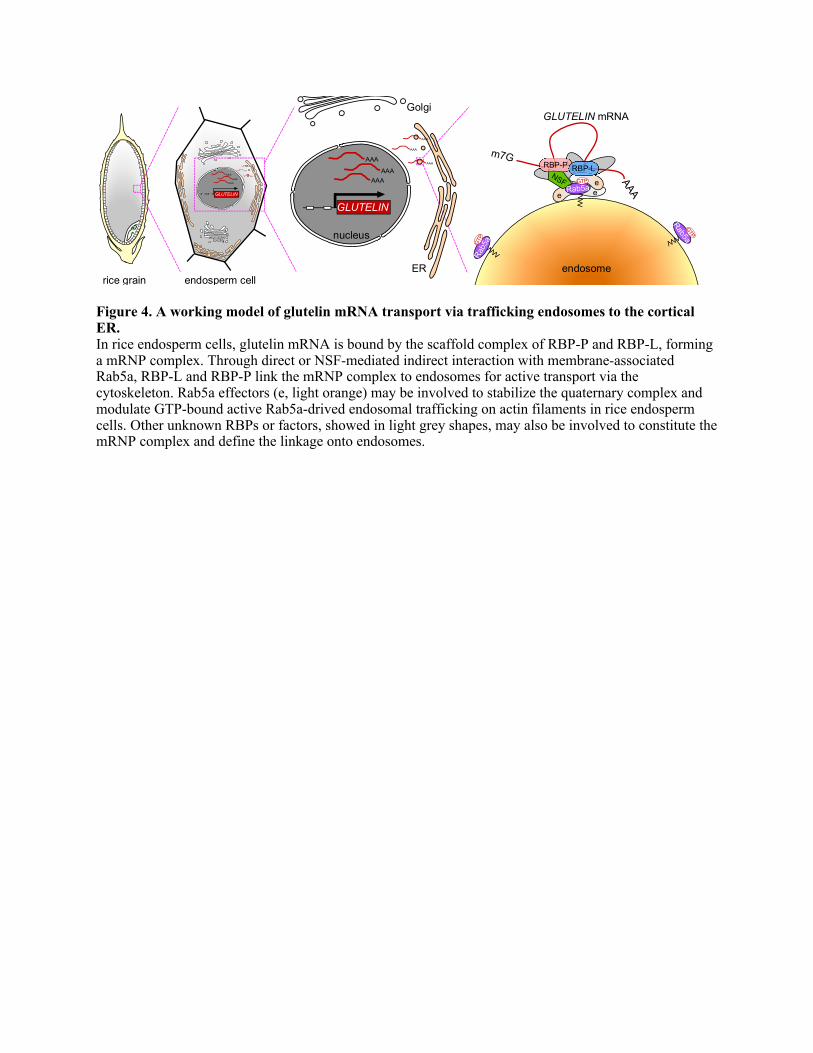
Figure 4. A working model of glutelin mRNA transport via trafficking endosomes to the cortical ER. In rice endosperm cells, glutelin mRNA is bound by the scaffold complex of RBP-P and RBP-L, forming a mRNP complex. Through direct or NSF-mediated indirect interaction with membrane-associated Rab5a, RBP-L and RBP-P link the mRNP complex to endosomes for active transport via the cytoskeleton. Rab5a effectors (e, light orange) may be involved to stabilize the quaternary complex and modulate GTP-bound active Rab5a-drived endosomal trafficking on actin filaments in rice endosperm cells. Other unknown RBPs or factors, showed in light grey shapes, may also be involved to constitute the mRNP complex and define the linkage onto endosomes.
e ee
RBP-P RBP-L
endosome
AAA
GLUTELIN
AAA
AAA
AAA
AAA
AAA
GLUTELIN mRNA
rice grain endosperm cell
AAAAAA
AAA
AAA
AAA
AAA
GLUTELIN
nucleus
ER
Golgi

Figure 5. Rab5a mutation leads to abnormal trafficking of endosomes and formation of extracellular paramural bodies (PMBs). (A) Schematic representation of the Rab5a mutation site in the rab5a mutant. A G134A base substitutionwithin the Rab5a gene resulted in a G45D amino acid replacement. (B) Formation of PMBs (whiteasterisks) was observed in endosperm cells of rab5a mutant through light microscopy observations onseed sections stained with 1% Toluidine Blue. Scale bar, 25 µm. (C) Ultrastructure of PMBs formed inrab5a mutant due to aborted endosomal trafficking in comparison to wildtype (WT) endosperm cells. Cellwall and PMB boundaries are indicated by magenta and green dashed lines, respectively. SG, starchgranules; orange *, protein body I; blue *, protein storage vacuoles. Scale bar, 1 µm.
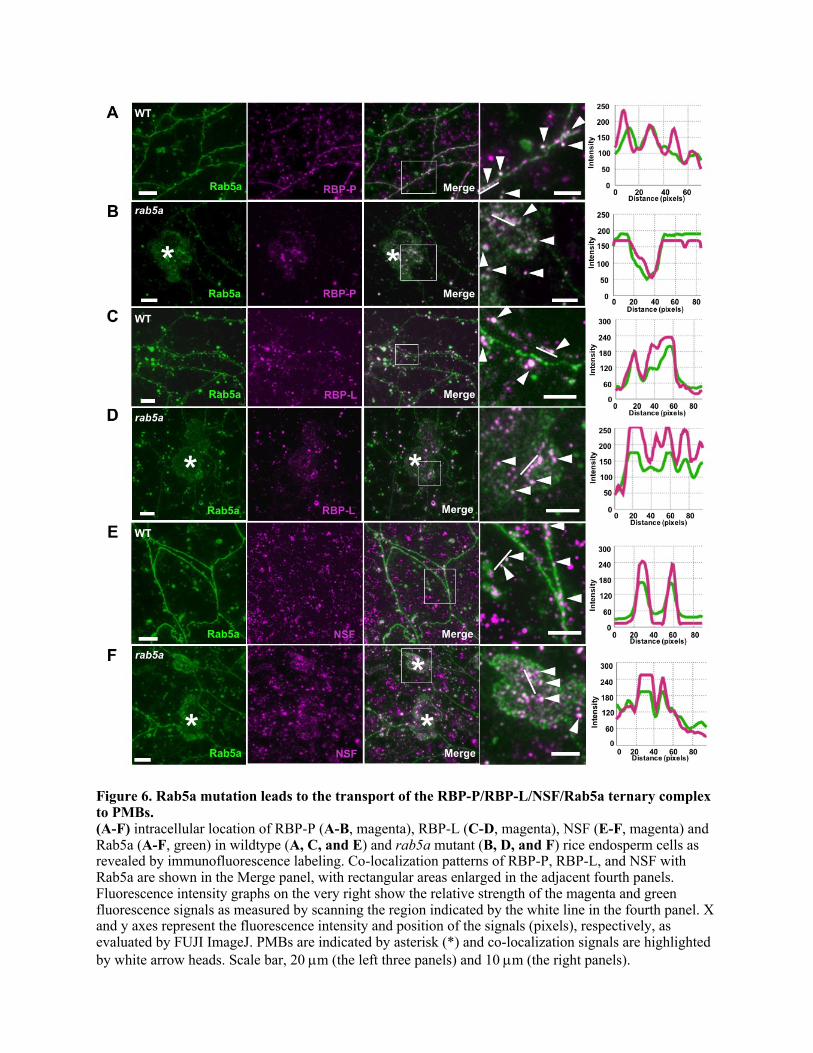
Figure 6. Rab5a mutation leads to the transport of the RBP-P/RBP-L/NSF/Rab5a ternary complex to PMBs. (A-F) intracellular location of RBP-P (A-B, magenta), RBP-L (C-D, magenta), NSF (E-F, magenta) and Rab5a (A-F, green) in wildtype (A, C, and E) and rab5a mutant (B, D, and F) rice endosperm cells as revealed by immunofluorescence labeling. Co-localization patterns of RBP-P, RBP-L, and NSF with Rab5a are shown in the Merge panel, with rectangular areas enlarged in the adjacent fourth panels. Fluorescence intensity graphs on the very right show the relative strength of the magenta and green fluorescence signals as measured by scanning the region indicated by the white line in the fourth panel. X and y axes represent the fluorescence intensity and position of the signals (pixels), respectively, as evaluated by FUJI ImageJ. PMBs are indicated by asterisk (*) and co-localization signals are highlighted by white arrow heads. Scale bar, 20 µm (the left three panels) and 10 µm (the right panels).

Figure 7. Retention of RBP-P and RBP-L on the aborted endosomes in the PMBs of rab5a mutant as revealed by immunocytochemistry and transmission electron microcopy. (A-J) Localization of RBP-P (A-C, F-G) or RBP-L (D-E, H-J) on Rab5a-labeled endosomes in wildtype (A-E) and rab5a mutant (F-J) endosperm cells. RBPs (RBP-P and RBP-L) and Rab5a were labeled with 15 nm (blue arrowheads) and 10 nm (red arrows) gold particles, respectively. Panels B, C, E, G, I and J are the enlarged areas (rectangle) shown in A, D, F and H. Cell wall and PMB boundaries are indicated by red and yellow dashed lines, respectively. Scale bar, 2µm (A, D, F and H) and 200 nm (B,C,E,G,I and J). Note that Rab5a-mediated endosomes are observed as electron-dense vesicles with an irregular-shape likely due to endosomal fusion (A to E) and as aborted endosome vesicles trapped within the PMBs in the rab5a mutant (F to J).
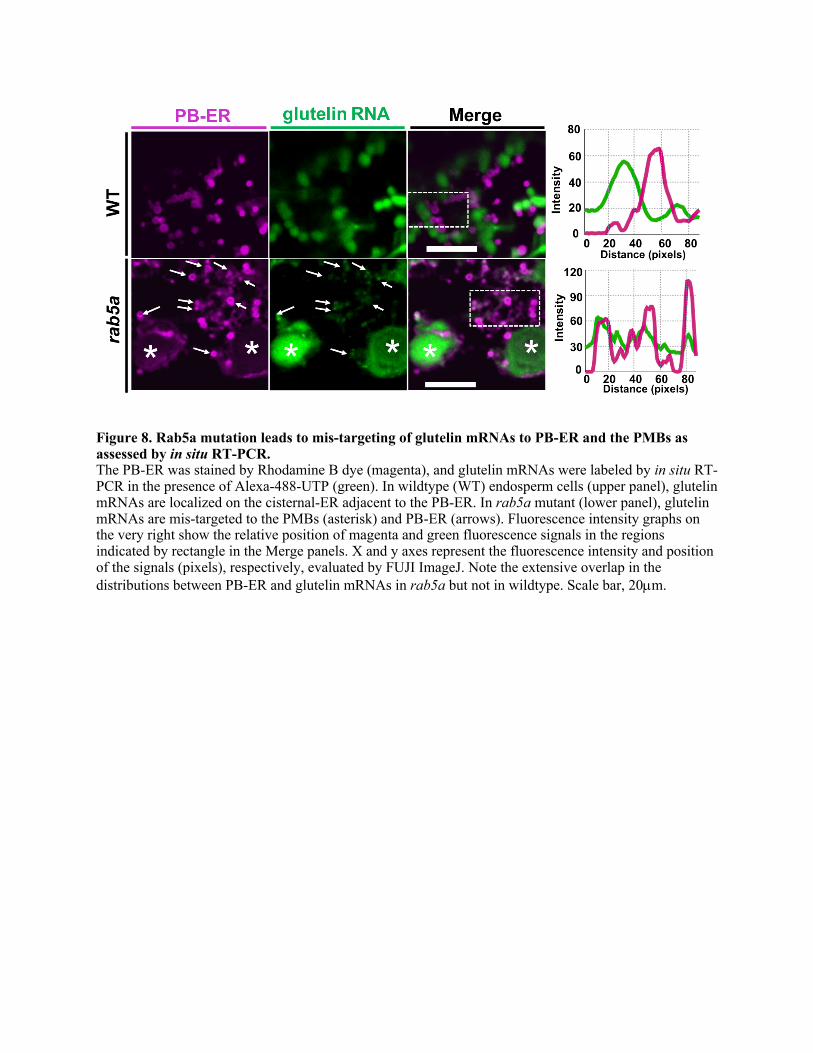
Figure 8. Rab5a mutation leads to mis-targeting of glutelin mRNAs to PB-ER and the PMBs as assessed by in situ RT-PCR. The PB-ER was stained by Rhodamine B dye (magenta), and glutelin mRNAs were labeled by in situ RT-PCR in the presence of Alexa-488-UTP (green). In wildtype (WT) endosperm cells (upper panel), glutelin mRNAs are localized on the cisternal-ER adjacent to the PB-ER. In rab5a mutant (lower panel), glutelin mRNAs are mis-targeted to the PMBs (asterisk) and PB-ER (arrows). Fluorescence intensity graphs on the very right show the relative position of magenta and green fluorescence signals in the regions indicated by rectangle in the Merge panels. X and y axes represent the fluorescence intensity and position of the signals (pixels), respectively, evaluated by FUJI ImageJ. Note the extensive overlap in the distributions between PB-ER and glutelin mRNAs in rab5a but not in wildtype. Scale bar, 20µm.

Parsed CitationsBarnard, R.J., Morgan, A., and Burgoyne, R.D. (1997). Stimulation of NSF ATPase activity by alpha-SNAP is required for SNARE complexdisassembly and exocytosis. J Cell Biol 139, 875-883.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Baumann, S., Pohlmann, T., Jungbluth, M., Brachmann, A., and Feldbrugge, M. (2012). Kinesin-3 and dynein mediate microtubule-dependent co-transport of mRNPs and endosomes. J Cell Sci 125, 2740-2752.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Blower, M.D. (2013). Molecular insights into intracellular RNA localization. Int Rev Cell Mol Biol 302, 1-39.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Boulianne, G.L., and Trimble, W.S. (1995). Identification of a second homolog of N-ethylmaleimide-sensitive fusion protein that isexpressed in the nervous system and secretory tissues of Drosophila. Proc Natl Acad Sci U S A 92, 7095-7099.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chang, L.F., Chen, S., Liu, C.C., Pan, X., Jiang, J., Bai, X.C., Xie, X., Wang, H.W., and Sui, S.F. (2012). Structural characterization of full-length NSF and 20S particles. Nat Struct Mol Biol 19, 268-275.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chavrier, P., Parton, R.G., Hauri, H.P., Simons, K., and Zerial, M. (1990). Localization of low molecular weight GTP binding proteins toexocytic and endocytic compartments. Cell 62, 317-329.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chevallet, M., Luche, S., and Rabilloud, T. (2006). Silver staining of proteins in polyacrylamide gels. Nat Protoc 1, 1852-1858.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chou, H.L., Tian, L., Washida, H., Fukuda, M., Kumamaru, T., and Okita, T.W. (2019). The rice storage protein mRNAs as a model systemfor RNA localization in higher plants. Plant Sci 284, 203-211.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Corradi, E., Dalla Costa, I., Gavoci, A., Iyer, A., Roccuzzo, M., Otto, T.A., Oliani, E., Bridi, S., Strohbuecker, S., Santos-Rodriguez, G.,Valdembri, D., Serini, G., Abreu-Goodger, C., and Baudet, M.-L. (2020). Axonal precursor miRNAs hitchhike on endosomes and locallyregulate the development of neural circuits. The EMBO Journal 39, e102513.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Crofts, A.J., Crofts, N., Whitelegge, J.P., and Okita, T.W. (2010). Isolation and identification of cytoskeleton-associated prolamine mRNAbinding proteins from developing rice seeds. Planta 231, 1261-1276.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Doroshenk, K.A., Crofts, A.J., Morris, R.T., Wyrick, J.J., and Okita, T.W. (2009). Proteomic analysis of cytoskeleton-associated RNAbinding proteins in developing rice seed. J Proteome Res 8, 4641-4653.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Doroshenk, K.A., Crofts, A.J., Morris, R.T., Wyrick, J.J., and Okita, T.W. (2012). RiceRBP: A Resource for Experimentally Identified RNABinding Proteins in Oryza sativa. Front Plant Sci 3, 90.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Doroshenk, K.A., Tian, L., Crofts, A.J., Kumamaru, T., and Okita, T.W. (2014). Characterization of RNA binding protein RBP-P reveals apossible role in rice glutelin gene expression and RNA localization. Plant Mol Biol 85, 381-394.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Doroshenk, K.A., Crofts, A.J., Washida, H., Satoh-Cruz, M., Crofts, N., Sugino, A., Okita, T.W., Morris, R.T., Wyrick, J.J., Fukuda, M.,Kumamaru, T., and Satoh, H. (2010). Characterization of the rice glup4 mutant suggests a role for the small GTPase Rab5 in thebiosynthesis of carbon and nitrogen storage reserves in developing endosperm. Breeding Science 60, 556-567.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fischer von Mollard, G., Stahl, B., Walch-Solimena, C., Takei, K., Daniels, L., Khoklatchev, A., De Camilli, P., Sudhof, T.C., and Jahn, R.(1994). Localization of Rab5 to synaptic vesicles identifies endosomal intermediate in synaptic vesicle recycling pathway. Eur J Cell

Biol 65, 319-326.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fukuda, M., Satoh-Cruz, M., Wen, L., Crofts, A.J., Sugino, A., Washida, H., Okita, T.W., Ogawa, M., Kawagoe, Y., Maeshima, M., andKumamaru, T. (2011). The small GTPase Rab5a is essential for intracellular transport of proglutelin from the Golgi apparatus to theprotein storage vacuole and endosomal membrane organization in developing rice endosperm. Plant Physiol 157, 632-644.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gohre, V., Vollmeister, E., Bolker, M., and Feldbrugge, M. (2012). Microtubule-dependent membrane dynamics in Ustilago maydis:Trafficking and function of Rab5a-positive endosomes. Commun Integr Biol 5, 485-490.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Golby, J.A., Tolar, L.A., and Pallanck, L. (2001). Partitioning of N-ethylmaleimide-sensitive fusion (NSF) protein function in Drosophilamelanogaster: dNSF1 is required in the nervous system, and dNSF2 is required in mesoderm. Genetics 158, 265-278.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Grosshans, B.L., Ortiz, D., and Novick, P. (2006). Rabs and their effectors: achieving specificity in membrane traffic. Proc Natl Acad SciU S A 103, 11821-11827.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Guo, Y., Yue, Q., Gao, J., Wang, Z., Chen, Y.R., Blissard, G.W., Liu, T.X., and Li, Z. (2017). Roles of Cellular NSF Protein in Entry andNuclear Egress of Budded Virions of Autographa californica Multiple Nucleopolyhedrovirus. J Virol 91.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Haag, C., Steuten, B., and Feldbrugge, M. (2015). Membrane-Coupled mRNA Trafficking in Fungi. Annu Rev Microbiol 69, 265-281.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hamada, S., Ishiyama, K., Sakulsingharoj, C., Choi, S.B., Wu, Y., Wang, C., Singh, S., Kawai, N., Messing, J., and Okita, T.W. (2003). Dualregulated RNA transport pathways to the cortical region in developing rice endosperm. Plant Cell 15, 2265-2272.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hanson, P.I., Otto, H., Barton, N., and Jahn, R. (1995). The N-ethylmaleimide-sensitive fusion protein and alpha-SNAP induce aconformational change in syntaxin. J Biol Chem 270, 16955-16961.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Herbert, S.P., and Costa, G. (2019). Sending messages in moving cells: mRNA localization and the regulation of cell migration. EssaysBiochem 63, 595-606.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Herve, J.C., and Bourmeyster, N. (2018). Rab GTPases, master controllers of eukaryotic trafficking. Small GTPases 9, 1-4.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Horsnell, W.G., Steel, G.J., and Morgan, A. (2002). Analysis of NSF mutants reveals residues involved in SNAP binding and ATPasestimulation. Biochemistry 41, 5230-5235.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hughes, S.C., and Simmonds, A.J. (2019). Drosophila mRNA Localization During Later Development: Past, Present, and Future. FrontGenet 10, 135.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ito, E., Ebine, K., Choi, S.W., Ichinose, S., Uemura, T., Nakano, A., and Ueda, T. (2018). Integration of two RAB5 groups duringendosomal transport in plants. Elife 7.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jansen, R.P., Niessing, D., Baumann, S., and Feldbrugge, M. (2014). mRNA transport meets membrane traffic. Trends Genet 30, 408-417.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kerppola, T.K. (2006). Design and implementation of bimolecular fluorescence complementation (BiFC) assays for the visualization ofprotein interactions in living cells. Nat Protoc 1, 1278-1286.

Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lee, M.T., Mishra, A., and Lambright, D.G. (2009). Structural mechanisms for regulation of membrane traffic by rab GTPases. Traffic 10,1377-1389.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li, Y., Wang, S., Li, T., Zhu, L., and Ma, C. (2018). Tomosyn guides SNARE complex formation in coordination with Munc18 and Munc13.FEBS Lett 592, 1161-1172.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liao, Y.C., Fernandopulle, M.S., Wang, G., Choi, H., Hao, L., Drerup, C.M., Patel, R., Qamar, S., Nixon-Abell, J., Shen, Y., Meadows, W.,Vendruscolo, M., Knowles, T.P.J., Nelson, M., Czekalska, M.A., Musteikyte, G., Gachechiladze, M.A., Stephens, C.A., Pasolli, H.A.,Forrest, L.R., St George-Hyslop, P., Lippincott-Schwartz, J., and Ward, M.E. (2019). RNA Granules Hitchhike on Lysosomes for Long-Distance Transport, Using Annexin A11 as a Molecular Tether. Cell 179, 147-164 e120.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mastick, C.C., and Falick, A.L. (1997). Association of N-ethylmaleimide sensitive fusion (NSF) protein and soluble NSF attachmentproteins-alpha and -gamma with glucose transporter-4-containing vesicles in primary rat adipocytes. Endocrinology 138, 2391-2397.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
McBride, H.M., Rybin, V., Murphy, C., Giner, A., Teasdale, R., and Zerial, M. (1999). Oligomeric complexes link Rab5 effectors with NSFand drive membrane fusion via interactions between EEA1 and syntaxin 13. Cell 98, 377-386.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Miller, K.E., Kim, Y., Huh, W.K., and Park, H.O. (2015). Bimolecular Fluorescence Complementation (BiFC) Analysis: Advances andRecent Applications for Genome-Wide Interaction Studies. J Mol Biol 427, 2039-2055.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mohtashami, M., Stewart, B.A., Boulianne, G.L., and Trimble, W.S. (2001). Analysis of the mutant Drosophila N-ethylmaleimide sensitivefusion-1 protein in comatose reveals molecular correlates of the behavioural paralysis. J Neurochem 77, 1407-1417.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Muench, D.G., Chuong, S.D., Franceschi, V.R., and Okita, T.W. (2000). Developing prolamine protein bodies are associated with thecortical cytoskeleton in rice endosperm cells. Planta 211, 227-238.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Muller, J., Pohlmann, T., and Feldbrugge, M. (2019). Core components of endosomal mRNA transport are evolutionarily conserved infungi. Fungal Genet Biol 126, 12-16.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Niessing, D., Jansen, R.P., Pohlmann, T., and Feldbrugge, M. (2018). mRNA transport in fungal top models. Wiley Interdiscip Rev RNA 9.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pohlmann, T., Baumann, S., Haag, C., Albrecht, M., and Feldbrugge, M. (2015). A FYVE zinc finger domain protein specifically linksmRNA transport to endosome trafficking. Elife 4.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pylypenko, O., Hammich, H., Yu, I.M., and Houdusse, A. (2018). Rab GTPases and their interacting protein partners: Structural insightsinto Rab functional diversity. Small GTPases 9, 22-48.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ryu, J.K., Min, D., Rah, S.H., Kim, S.J., Park, Y., Kim, H., Hyeon, C., Kim, H.M., Jahn, R., and Yoon, T.Y. (2015). Spring-loaded unravelingof a single SNARE complex by NSF in one round of ATP turnover. Science 347, 1485-1489.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Saito, C., and Ueda, T. (2009). Chapter 4: functions of RAB and SNARE proteins in plant life. Int Rev Cell Mol Biol 274, 183-233.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schmid, M., Jaedicke, A., Du, T.G., and Jansen, R.P. (2006). Coordination of endoplasmic reticulum and mRNA localization to the yeastbud. Curr Biol 16, 1538-1543.

Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Stenmark, H., and Olkkonen, V.M. (2001). The Rab GTPase family. Genome Biol 2, REVIEWS3007.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tagaya, M., Wilson, D.W., Brunner, M., Arango, N., and Rothman, J.E. (1993). Domain structure of an N-ethylmaleimide-sensitive fusionprotein involved in vesicular transport. J Biol Chem 268, 2662-2666.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tanabashi, S., Shoda, K., Saito, C., Sakamoto, T., Kurata, T., Uemura, T., and Nakano, A. (2018). A Missense Mutation in the NSF GeneCauses Abnormal Golgi Morphology in Arabidopsis thaliana. Cell Struct Funct 43, 41-51.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tian, L., and Okita, T.W. (2014). mRNA-based protein targeting to the endoplasmic reticulum and chloroplasts in plant cells. Curr OpinPlant Biol 22, 77-85.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tian, L., Chou, H.L., Zhang, L., and Okita, T.W. (2019a). Targeted Endoplasmic Reticulum Localization of Storage Protein mRNAsRequires the RNA-Binding Protein RBP-L. Plant Physiol 179, 1111-1131.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tian, L., Chou, H.L., Fukuda, M., Kumamaru, T., and Okita, T.W. (2019b). mRNA localization in plant cells. Plant Physiol.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tian, L., Chou, H.L., Fukuda, M., Kumamaru, T., and Okita, T.W. (2020). mRNA Localization in Plant Cells. Plant Physiol 182, 97-109.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tian, L., Chou, H.L., Zhang, L., Hwang, S.K., Starkenburg, S.R., Doroshenk, K.A., Kumamaru, T., and Okita, T.W. (2018). RNA-BindingProtein RBP-P Is Required for Glutelin and Prolamine mRNA Localization in Rice Endosperm Cells. Plant Cell 30, 2529-2552.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ueda, T., Yamaguchi, M., Uchimiya, H., and Nakano, A. (2001). Ara6, a plant-unique novel type Rab GTPase, functions in the endocyticpathway of Arabidopsis thaliana. EMBO J 20, 4730-4741.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Vida, T.A., and Emr, S.D. (1995). A new vital stain for visualizing vacuolar membrane dynamics and endocytosis in yeast. J Cell Biol 128,779-792.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Vollmeister, E., Schipper, K., and Feldbrugge, M. (2012). Microtubule-dependent mRNA transport in the model microorganism Ustilagomaydis. RNA Biol 9, 261-268.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang, Y., Ren, Y., Liu, X., Jiang, L., Chen, L., Han, X., Jin, M., Liu, S., Liu, F., Lv, J., Zhou, K., Su, N., Bao, Y., and Wan, J. (2010). OsRab5aregulates endomembrane organization and storage protein trafficking in rice endosperm cells. Plant J 64, 812-824.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Washida, H., Kaneko, S., Crofts, N., Sugino, A., Wang, C., and Okita, T.W. (2009). Identification of cis-localization elements that targetglutelin RNAs to a specific subdomain of the cortical endoplasmic reticulum in rice endosperm cells. Plant Cell Physiol 50, 1710-1714.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Washida, H., Sugino, A., Doroshenk, K.A., Satoh-Cruz, M., Nagamine, A., Katsube-Tanaka, T., Ogawa, M., Kumamaru, T., Satoh, H., andOkita, T.W. (2012). RNA targeting to a specific ER sub-domain is required for efficient transport and packaging of alpha-globulins to theprotein storage vacuole in developing rice endosperm. Plant J 70, 471-479.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Weis, B.L., Schleiff, E., and Zerges, W. (2013). Protein targeting to subcellular organelles via MRNA localization. Biochim Biophys Acta1833, 260-273.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

Wen, L., Fukuda, M., Sunada, M., Ishino, S., Ishino, Y., Okita, T.W., Ogawa, M., Ueda, T., and Kumamaru, T. (2015). Guanine nucleotideexchange factor 2 for Rab5 proteins coordinated with GLUP6/GEF regulates the intracellular transport of the proglutelin from theGolgi apparatus to the protein storage vacuole in rice endosperm. J Exp Bot 66, 6137-6147.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Woodman, P.G. (2000). Biogenesis of the sorting endosome: the role of Rab5. Traffic 1, 695-701.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yang, Y., Chou, H.L., Crofts, A.J., Zhang, L., Tian, L., Washida, H., Fukuda, M., Kumamaru, T., Oviedo, O.J., Starkenburg, S.R., and Okita,T.W. (2018). Selective sets of mRNAs localize to extracellular paramural bodies in a rice glup6 mutant. J Exp Bot 69, 5045-5058.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yuan, W., and Song, C. (2020). The Emerging Role of Rab5 in Membrane Receptor Trafficking and Signaling Pathways. Biochem Res Int2020, 4186308.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zeigerer, A., Gilleron, J., Bogorad, R.L., Marsico, G., Nonaka, H., Seifert, S., Epstein-Barash, H., Kuchimanchi, S., Peng, C.G., Ruda,V.M., Del Conte-Zerial, P., Hengstler, J.G., Kalaidzidis, Y., Koteliansky, V., and Zerial, M. (2012). Rab5 is necessary for the biogenesis ofthe endolysosomal system in vivo. Nature 485, 465-470.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhao, C., Slevin, J.T., and Whiteheart, S.W. (2007). Cellular functions of NSF: not just SNAPs and SNAREs. FEBS Lett 581, 2140-2149.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhao, C., Matveeva, E.A., Ren, Q., and Whiteheart, S.W. (2010). Dissecting the N-ethylmaleimide-sensitive factor: required elements ofthe N and D1 domains. J Biol Chem 285, 761-772.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

DOI 10.1105/tpc.20.00111; originally published online May 29, 2020;Plant Cell
and Thomas W OkitaLi Tian, Kelly A. Doroshenk, Laining Zhang, Masako Fukuda, Haruhiko Washida, Toshihiro Kumamaru
Glutelin mRNAs in Rice EndospermZipcode RNA-binding Proteins and Membrane Trafficking Proteins Cooperate to Transport
This information is current as of August 6, 2020
Supplemental Data /content/suppl/2020/06/09/tpc.20.00111.DC1.html
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists
Top Related