







Languages
Pages
Legal


In the format provided by the authors and unedited.
Title
Transient, cell-specific EXO70A1 activity in CASP domain and Casparian strip
localisation
Authors
Lothar Kalmbach1,†, Kian Hématy1,2, Damien De Bellis1,3, Marie Barberon1, Satoshi
Fujita1, Robertas Ursache1, Jean Daraspe3, Niko Geldner1*
Affiliations 1 Department of Plant Molecular Biology, University of Lausanne, Biophore Building,
UNIL-Sorge, 1015 Lausanne, Switzerland
2 Institut Jean-Pierre Bourgin, INRA, AgroParisTech, CNRS, Université Paris-Saclay,
78000 Versailles, France
3 Electron Microscopy Facility, University of Lausanne, Biophore Building, UNIL-Sorge,
1015 Lausanne, Switzerland
† Present address: Sainsbury Laboratory, University of Cambridge, Bateman Street,
Cambridge CB2 1LR, United Kingdom
1
Transient cell-specific EXO70A1 activity in theCASP domain and Casparian strip localizationLothar Kalmbach1†, Kian Hématy1,2, Damien De Bellis1,3, Marie Barberon1, Satoshi Fujita1,Robertas Ursache1, Jean Daraspe3 and Niko Geldner1*
In a striking case of evolutionary convergence, polarized cell layers with ring-like diffusion barriers have evolved in bothplant and animal lineages independently. In plants, ring-like Casparian strips become localized by the CASPARIAN STRIPMEMBRANE DOMAIN PROTEINs (CASPs). The mechanism of this striking localization, however, has remained enigmatic.Here we present a genetic screen aimed at isolating determinants of CASP localization. One of the mutants, lotr2/exo70a1,displays dramatic de-localization of CASPs into randomly localized microdomains. EXO70A1 is a subunit of the exocystcomplex, a central component of secretion in eukaryotes. Irradiation of EXO70 subunit genes in plants has suggestedspecialization of this conserved complex. Intriguingly, lotr2/exo70a1 does neither affect secretion of the CASPs, nor that ofother membrane proteins in the endodermis, thus separating exocyst activity in localization from a general defect insecretion. Our results establish EXO70A1 as a central player in Casparian strip formation, generating a transient positionalinformation that will be translated into a precisely localized cell wall modification.
The endodermis is the innermost cortical cell layer in the root ofhigher plants and surrounds the central cylinder with thevascular tissues. Its physiological role is to allow controlled
and selective uptake of nutrients to enter the stele and to retainthem in favourable concentrations for the plant1–5 The Casparianstrip is a highly localized, ring-like lignin deposition around eachendodermal cell. It seals the apoplastic space between adjacentendodermal cells thus prevents extracellular diffusion, forcingpassage of nutrients across cellular membranes. Hence, it is amajor structural modification that allows the plant to control itsnutrient uptake. Therefore, the endodermis can be seen as aplant’s equivalent to intestinal epithelia in animals, with theCasparian strip being functionally analogous, yet molecularlydifferent, to tight/adherens junctions6,7.
Casparian strip formation is preceded by localization of theCASPARIAN STRIP MEMBRANE DOMAIN PROTEINs 1–5(CASP1-5), a family of endodermis-specific plasma membrane(PM) proteins, which initially localize ubiquitously at the PMbefore they rapidly fuse into a continuous, central ring. They serveas a platform to recruit lignin biosynthetic enzymes to ultimatelyset up the endodermal diffusion barrier8–11 Therefore, the ring-like CASP localization also represents a perfect system to studylocalized PM microdomain formation in an unbiased fashion.
Previously identified endodermal barrier mutants as for instancethe schengen (sgn) mutants or enhanced suberin 1 (esb1) displaydiscontinuities of the Casparian strips and the CASP domain inthe PM2,3,12. However, all described endodermis mutants to dateshowed only minor effects on CASP localization, mostly leadingto fragmented, slightly shifted, but otherwise normally localizedCASP domains2,3,12. The transcription factor mutant myb domainprotein 36 (myb36), which causes inactivity of most endodermalgenes, including the CASPs also cannot instruct us on howCASPs become localized into their central domain4,13. Thus, thefundamental mechanisms required for positioning of the CASPs
have remained obscure. Here, we now report the first completemis-localization phenotype of CASP1–GFP and loss of this centraldomain and identify loss-of-function of EXO70A1 as the causalmutation. We show that exo70a1 mutants specifically affect locali-zation, but not secretion of CASP1–GFP and also maintain normalsecretion and localization of other proteins tested, demonstratingthe surprising capacity of the plant exocyst for mediatingsubstrate-specific and spatially restricted protein localization.
ResultsA screen to identify spatial regulators of CASP positioning. Weisolated mutants through an epi-fluorescence-based, forwardgenetic screen scoring directly for mis-localization of CASP1–GFPin an ethylmethane sulphonate-mutagenized Arabidopsis thalianapopulation of 2,378 individual lines, (Fig. 1a). Since all mutations,in one way or another, affected the establishment of thesub-cellular CASP1–GFP ring, we named them ‘lord of the rings’(lotr) mutants, after the novel by J. R. R. Tolkien (for details seeSupplementary Information). This screen identified 48 mutants,falling into 15 complementation groups. Twenty-one of thesemutations were mapped to nine genes with previously unknownroles during endodermal differentiation or PM domainestablishment (LOTR1–LOTR9) and 27 are novel alleles of knownendodermis mutants (Supplementary Table 1). Intriguingly, mostmutants only showed discontinuous CASP1–GFP domains, whichremained centrally localized, implying that CASP localizationeither involves general trafficking regulators, whose mutants oftenare lethal, or that the required cellular factors are largely redundant.
LOTR2/EXO70A1 is required for median CASP targeting.Mutations in LOTR2, however, showed a dramatic CASP1–GFPmis-localization, suggesting that we had identified a centralregulator of CASP domain formation (Fig. 1b and SupplementaryFig. 1). The two lotr2 alleles showed excessive and ectopic
1Department of Plant Molecular Biology, University of Lausanne, Biophore Building, UNIL-Sorge, 1015 Lausanne, Switzerland. 2Institut Jean-Pierre Bourgin,INRA, AgroParisTech, CNRS, Université Paris-Saclay, 78000 Versailles, France. 3Electron Microscopy Facility, University of Lausanne, Biophore Building,UNIL-Sorge, 1015 Lausanne, Switzerland. †Present address: Sainsbury Laboratory, University of Cambridge, Bateman Street, Cambridge CB2 1LR, UK.*e-mail: [email protected]
ARTICLESPUBLISHED: XX XX 2017 | VOLUME: 3 | ARTICLE NUMBER: 17058
NATURE PLANTS 3, 17058 (2017) | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 1
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
SUPPLEMENTARY INFORMATIONVOLUME: 3 | ARTICLE NUMBER: 17058
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 1

Supplementary Information
Supplementary Figures 1-8
Supplementary Movie 1
Supplementary Material and Methods
Supplementary Table 1-3
Supplementary References
2
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 2
SUPPLEMENTARY INFORMATION

MedianSurface
exo70a1-4
(lotr2-1)
exo70a1-5
(lotr2-2)
exo70a1-1WT
48 dag
d
T3-8 T3-9 T3-14WT exo70a1-4 exo70a1-5a
exo70a1-1
CAS
P1-G
FP
b
Med
ian
Surfa
ce
WT exo70a1-4 exo70a1-5 exo70a1-1c
SGN3::EXO70A1 / exo70a1-4
exo70a1-4WTe
Supplementary Figure 1 | exo70a1 mutant phenotypes and complementation with transgenes. a, 3D maximum projections of CASP1-GFP in WT and three exo70a1 alleles with complementation of exo70a1-4 by three independent SGN3::EXO70A1 expressing lines to the right. b, Schematic representation as in Figure 1e. c, Surface and median confocal sections of WT and three exo70a1 alleles. d, Growth phenotypes of adult (48-days after germination) WT plants and exo70a1 mutants. e, Transmission electron micrographs of WT and exo70a1-4 depicting the endodermis-endodermis interface; sections were taken 2 mm from root tip. Image for exo70a1-1 in (a) and schematic representation in (b) are also shown in Figure 1b and 1e of main text, respectively. Scale bars represent 20 µm for 3D maximum projections in (a), 10 µm for surface and median confocal sections in (b), 10 cm in (d) and 500 nm in (e).
3
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 3
SUPPLEMENTARY INFORMATION

T3-8 T3-9 T3-14 T3-70 T3-73T3-39WT exo70a1-5 exo70a1-1exo70a1-4
010
4020
30C
ells
afte
r ons
et o
f elo
ngat
ion
a
bb
b
a a a a a a
SGN3::EXO70A1 / exo70a1-4 EXO70A1::EXO70A1-mVenus / exo70a1-1
c
Complemented transgenic lines
T3-8 T3-9 T3-14 T3-39 T3-70 T3-73a
15 c
ells
afte
r px
8 ce
lls a
fter p
xFu
chsi
n (li
gnin
)
WT exo70a1-2 exo70a1-5 exo70a1-1
T3-8 T3-9 T3-14 T3-70 T3-73T3-39WT exo70a1-5 exo70a1-1exo70a1-4
40-4
5 ce
lls20
-25
cells
Fluo
rol Y
ello
w (s
uber
in)
b
SGN3:EXO70A1 / exo70a1-4 EXO70A1::EXO70A1-mVenus / exo70a1-1
SGN3:EXO70A1 / exo70a1-4 EXO70A1::EXO70A1-mVenus / exo70a1-1
4
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 4
SUPPLEMENTARY INFORMATION

Supplementary Figure 2 | exo70a1 mutants display typical compensatory lignin and suberin deposition syndrome. a, 3D maximum projections of fuchsin-stained Casparian strips in WT, three exo70a1 alleles and transgenic complementations showing three independent T3 lines of homozygous exo70a1-4 mutants (T3-8, -9 and -14), as well as exo70a1-1 mutants (T3-39, -70 and -73) complemented withSGN3::EXO70A1 and EXO70A1::EXO70A1-mVenus, respectively. Upper panel depictsendodermal lignification in later stages (15 cells after onset of protoxylem lignification),lower panel in early stages (8 cells after onset of protoxylem lignification) ofendodermal development. b, Epifluorescence images of Fluorol Yellow-stained suberinlamellae of same genotypes as in (a); images depict suberization in late (40-45 cellsafter onset of elongation) and early (20-25 cells after the onset of elongation) parts ofthe root endodermis, demonstrating that the transgenic lines also complemented theprecocious suberization phenotype of lotr2/exo70a1. c, PI uptake assay of samegenotypes as in (a); PI uptake values for WT and exo70a1-1 are also shown inFigure 1c of main text. Scale bars: 10 µm in (a) and 100 µm in (b). Different lettersin (c) indicate statistically significant differences of means (p<0.005) by analysis ofvariance (ANOVA) and Tukey’s test as post-hoc analysis (n=10).
5© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 5
SUPPLEMENTARY INFORMATION

0
1
2
3
4
5
6
7
8
A1 B1 D1 D2 D3 E1 F1 H7 H8B2
Rel
ativ
e ex
pres
sion
Endodermis*Early endodermis**Differentiating endodermis**
-10 -10
-10 -10
-10
-10
co en ph xy
CASP15
0
-5
-10 ep co en ph xy
MYB365
0
-5
-10 epco en ph xy
EXO70H85
0
-5
-10 ep
co en ph xy
EXO70E15
0
-5
ep co en ph xy
EXO70F15
0
-5
ep
co en ph xy
EXO70D15
0
-5
ep co en ph xy
EXO70D25
0
-5
ep
co en ph xy
EXO70H75
0
-5
ep
co en ph xy
EXO70D35
0
-5
ep
co en ph xy
EXO70A15
0
-5
-10 ep co en ph xy
EXO70B15
0
-5
-10 ep co en ph xy
EXO70B25
0
-5
-10 ep
c exo70b1-1WT
10 ce
lls20
cells
exo70b1-2
Cel
ls a
fter o
nset
of l
igni
ficat
ion
a a a
1015
205
0
WT (Col-
0)
exo7
0b1-1
exo7
0b1-2
2530
d
Cel
ls a
fter o
nset
of l
igni
ficat
ion
a
b
6
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 6
SUPPLEMENTARY INFORMATION

Supplementary Figure 3 | EXO70A1 has a non-redundant function in the endodermis. a, Endodermal expression levels of the ten EXO70 subunits, which are expressed in the endodermis; data was normalised to the endodermal expression of EXO70A1; expression data extracted from endodermis-specific RNAseq (*)1 and from cell-type specific root transcriptome profiling of early and differentiating endodermal cells (**)2; threshold for significant expression in the endodermis was arbitrarily set to one third of the EXO70A1 expression in any sample. b, Relative expression of the ten EXO70 subunits shown in (a) across root tissues; data was extracted from cell-specific root transcriptome profiling as in (a)2 and is presented as log2 fold-change compared to each gene’s endodermal expression level; relative expression of two endodermis-specific genes (CASP1, MYB36) across root tissues is provided for comparison; epidermis (ep), cortex (co), endodermis (en), phloem (ph), xylem (xy). c, 3D maximum projections of fuchsin-stained Casparian strips in exo70b1 mutants in the early and later root do not shown any defects in endodermal lignification. d, PI uptake assay in exo70b1 mutants does not show any defects in apoplastic barrier establishment. Error bars in (a) represent standard deviations. Scale bar in (c) represents 25 µm. Identical letters in (d) indicate no statistically significant differences of means (p>0.5) by analysis of variance (ANOVA) and Tukey’s test as post-hoc analysis (n=10).
7© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 7
SUPPLEMENTARY INFORMATION

b
WT
Surfa
ceM
edia
nex
o70a
1-4
Surfa
ceM
edia
n
CASP1-GFP
RBOHF-mCherry Merged
aW
TSu
rface
Med
ian
exo7
0a1-
4Su
rface
Med
ian
CASP1-GFP
AtFH1-107-mCherry Merged
exo8
4b-4
-/-
3D m
ax. p
roje
ctio
nex
o84b
+/-
3D m
ax. p
roje
ctio
n
CASP1-GFP
c
Supplementary Figure 4 | exo70a1 has no effect on general secretion but only on the median CASP domain. a, b, Co-visualisation of CASP1-GFP with AtFH1-107-mCherry and RBOHF-mCherry, respectively, in WT and exo70a1-4; images shown in surface and median view. c, 3D maximum projections of CASP1-GFP in hetero- and homozygous exo84b-1; note that CASP1-GFP signal was very weak in homozygous exo84b-1 and non-endodermal autofluorescence was partially captured. Scale bars represent 5 µm in (a, b) and 20 µm in (c).
8© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 8
SUPPLEMENTARY INFORMATION

*
Inner PM
0
250 Root #1
RFU
*
0
250 Root #2
RFU
*
0
250 Root #3
RFU
*
0
250 Root #3
RFU
*
0
250 Root #2
RFU
*
0
250 Root #1
RFU
*
0
250 Root #2
RFU
*
0
250 Root #3
RFU
*
0
250 Root #1R
FU
EXO70A1-mVenusRoot #2 Root #3Root #1
GFP-SEC8Root #2 Root #3Root #1
SEC6-GFPRoot #2 Root #3Root #1
a
b
cOuter PM
Inner PMOuter PM
Inner PMOuter PM
Inner PMTubby-C-Citrine
Root #2 Root #3Root #1d
Outer PM
0
250 Root #1
RFU
0
250 Root #2
RFU
0
250 Root #3
RFU
*
*
*
0 255
Relative intensity
9
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 9
SUPPLEMENTARY INFORMATION

Supplementary Figure 5 | Exocyst and PI(4,5)P2 are ubiquitously present but accumulate at the site of the future Casparian strip. a-d, Confocal sections of the early root around the eighth endodermal cell after onset of elongation with quantifications of fluorescence intensities around the plasma membrane of the indicated endodermal cells for three roots per genotype. a, EXO70A1-mVenus. b, GFP-SEC8. c, SEC6-GFP. d, PI(4,5)P2 marker Tubby-C-Citrine. Transgenes in (a, b and c) were expressed for their respective native promoter; transgene in (d) was expressed from the UBQ10 promoter; fluorescence intensities were quantified along the outlined traces and in the indicated orientation and are presented in relative fluorescence units (RFU) for one endodermal cell of each root; quantifications include the outer endodermal plasma membrane domain, the central plasma membrane domain of the future Casparian strip and the inner plasma membrane domain; asterisks indicate the position of the central plasma membrane domain of the future Casparian strip. Scale bars indicate 50 µm.
10© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 10
SUPPLEMENTARY INFORMATION

***
Tubby-C-Citrine#39
#73GFP-SEC8SEC6-GFP
CASP1 expr.CASP1 accum.
EXO70A1-mVenus
PI block
Cells before PI block-2-4-6-8 +1-10
b
#70
c
b
encoep encoep encoep
0 255
Relative intensity
CAS
P1-m
Che
rry /
ind.
MYB
36
EXO
70A1
-mVe
nus
/ ind
. MYB
36
GFP
-SEC
8 / i
nd. M
YB36
DMSO
10 µM Estradiol
DMSO
10 µM Estradiol
DMSO
10 µM Estradiol
en co ep en co ep en co ep
en co ep en co ep en co ep
a
EXO
70A1
-mVe
nus
#70
CAS
P1-m
Che
rry
Mer
ged
EXO
70A1
+ C
ASP1
aaa
aaa
a
Supplementary Figure 6 | MYB36-dependent and Endodermis-specific accumulations of the exocyst and CASP1. a, CASP1-mCherry, EXO70A1-mVenus and GFP-SEC8 accumulation in late non-endodermal tissues in inducible MYB36 overexpressor upon induction with 10 µM estradiol and DMSO control. b, Quantification of endodermal accumulation events and start of CASP1::CASP1-GFP expression relative to the establishment of a functional apoplastic barrier (“PI block”) in each quantified transgenic line; different letters in indicate statistically significant differences of means (p<0.001) by analysis of variance (ANOVA) and Tukey’s test as post-hoc analysis (n=10); asterisks highlight statistically significant difference between CASP1 accumulation and all other events. c, EXO70A1-mVenus and CASP1-mCherry localisation in early root differentiation zone of a second, independent transgenic line; insets are magnifications of boxed areas at endodermal cell junctions. Scale bars represent 10 µm in (a) and 50 µm in (c).
11© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 11
SUPPLEMENTARY INFORMATION

0 255
Relative intensity
a
en co en co
Tubby-C-Citrine
Tubby-Cmut-Citrine
b
en co ep
en co ep
Tubb
y-C
-Citr
ine
/ ind
. MYB
36
DMSO
10 µM Estradiol
Supplementary Figure 7 | Endodermis-specific PI(4,5)P2 accumulation is MYB36 dependent. a, Overview of PI(4,5,)P2 marker Tubby-C-Citrine mutation and lack thereof for the mutated, non-PI(4,5)P2-binding Tubby-Cmut-Citrine in early root of WT; arrow heads indicate endodermal cell junctions. b, Tubby-C-Citrine accumulation in late non-endodermal cells of inducible MYB36 overexpressor upon induction with 10 µM estradiol and DMSO control; arrowheads indicate Tubby-C-Citrine accumulation at PM. Endodermis (en), cortex (co). Scale bars represent 20 µm in (a) and 10 µm in (b).
12© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 12
SUPPLEMENTARY INFORMATION

c
1 2 3 4 5 6 7 8 9 10
1.5
1.0
0.5
0
WTexo70a1-1
cells after CASP1 expression
ECL
inde
x
ctr.
lat.
ctr.
lat.
ctr.
lat.
ctr.
lat.
ctr.
lat.
ctr.
lat.
ctr.
lat.
ctr.
lat.
1st cell 3rd cell 5th cell 10th cell
WT
exo7
0a1-
1
ECL index =fluo (lat.)fluo (ctr.)
ECL index =2x fluo (lat.)
fluo (ctr.)
ECL index =fluo (lat.)fluo (ctr.)
cell #6 cell #7 cell #8 cell #9 cell #10 cell #11 cell #12
lat.
ctr.
lat.
ctr.
lat.
ctr.
lat.
ctr.
lat.
ctr.
lat.
ctr.
lat.
ctr.
d
EXO70A1-mVenus GFP-SEC8 SEC6-GFP EXO70A1-mVenus / myb36-2
root #1 root #2 root #3
5 6 7 8 9 10 11 120
0.20.40.60.81.01.2
root #1 root #2 root #3root #1 root #2 root #3root #1 root #2 root #3
00.20.40.60.81.01.2
5 6 7 8 9 10 11 125 6 7 8 9 10 11 12 130
0.20.40.60.81.01.2
0.20.40.60.81.01.2
5 7 9 11 13 15cells after onset of elongationcells after onset of elongation cells after onset of elongation cells after onset of elongation
a
b
****
Supplementary Figure 8 | EXO70A1 allows for endodermis-specific spatially-restricted CASP accumulation. a, CASP1-GFP transitions from ubiquitous to centrally localised in endodermis of WT but not in exo70a1; representative images of one root used for quantification; boxed areas were used in (b) for calculation of the endodermal central localisation (ECL) index. b, ECL in WT and exo70a1 along endodermal differentiation; (**) indicate statistically significant difference (p<0.05; n=4). c, Transient central accumulation of EXO70A1-mVenus; representative images of one root used for quantification; boxed areas were used in (d) for calculation of the ECL index. d, ECL of three roots per genotype along endodermal differentiation; quantification in all lines were performed in an identical fashion as for EXO70A1-mVenus depicted in (c). Scale bars represent 5 µm in (a).
13
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 13
SUPPLEMENTARY INFORMATION

Supplementary Movie 1 | CASP1-GFP gradually accumulates at ectopic patches in exo70a1. 4D movie of confocal stacks of CASP1-GFP taken every 15 minutes for 12 h at the early differentiating endodermis. The movie depicts a time lapse of the 12 h followed by a 360º turn of the endodermis.
14© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 14
SUPPLEMENTARY INFORMATION

Supplementary Material and Methods
Mutagenesis
A mutant population was generated from seeds of a pool of eight individuals containing a
homozygous, single-locus copy of a genomic fragment of CASP1, fused to GFP and expressed
under its native promoter 3. Seeds were mutagenized as previously described using 0.4 % ethyl
methanesulfonate (EMS) in 100 mM phosphate buffer, pH 7.5 for eight hours and then washed
20 times in 100 mM phosphate buffer, pH 7.5 4,5.
Forward genetic screen
2378 individual M2 lines were generated and ten to twelve seedlings per line were screened
for CASP1-GFP localization defects by epifluorescence microscopy, using a Leica DM5500
equipped with a standard GFP filter set. Isolated putative mutants were confirmed in the M3
generation and back-crossed to the parental line to test for recessivity and to generate an
isogenic mapping populations. Phenotypical groups were established and allelism tests
performed within these groups.
Mutant mapping
Mutants, which could not be identified through genetic complementation analysis were whole-
genome sequenced using an Illumina HiSeq 2500. The SNPtrack web-based pipeline 6 was
used to map raw reads against the Arabidopsis thaliana reference genome (TAIR10) and to
perform variant analysis. Subsequent Sanger-sequencing of the candidate locus in a second
allele confirmed the causative polymorphisms.
Allelism for lotr2/exo70a1 mutants was confirmed by non-complementation in the F1: lotr2-1
+/- X lotr2-2 -/-: 36/69 (52%) non-complementation; lotr2-1 +/- X exo70a1-1 +/-: 3/12 (25%)
non-complementation. Consequently, lotr2-1 and lotr2-2 were named exo70a1-4 and exo70a1-
5, respectively. exo70a1-4 and exo70a1-5 were back-crossed to the parental CASP1::CASP1-
GFP three and two times, respectively.
Generation of complementing transgenic lines
EXO70A1::EXO70A1-mVenus: PCR amplicons with approximately 20 bp overlaps were
generated for the pGreenII backbone 7, for a 6523 bp sequence covering the entire EXO70A1
upstream intergenic region (-2215 bp) and the EXO70A1 genomic fragment without stop
codon, for the mVenus coding sequence, and for the entire EXO70A1 downstream intergenic
region as terminator. PCR products were then assembled by circular polymerase extension
15
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 15
SUPPLEMENTARY INFORMATION

cloning 8. The generated plasmid was transformed into a segregating population of exo70a1-1.
Localisation was consistent for all 20 transformants. T1 individuals were genotyped for
exo70a1-1 and three independent complemented lines, homozygous for exo70a1-1, were used
for further characterisation.
For endodermal complementation EXO70A1-mCherry was driven by the SGN3 promoter
(referred to as SGN3::EXO70A1 since no mCherry fluorescence was detectable). The KpnI-
flanked upstream region of SGN3 (-2181 bp) was sub-cloned into a pGreenII plasmid,
containing the 3’ OCS terminator (pGreenII-SGN3). EXO70A1 genomic fragment was PCR
stitched to the 5’ end of mCherry and the entire fragment inserted in an EcoRV digested
pGreenII-SGN3 plasmid by Gibson assembly 9. The generated plasmid was transformed into a
segregating population of lotr2-1/exo70a1-4. All T1 individuals were WT-looking and were
genotyped for lotr2-1/exo70a1-4 and three independent lines, homozygous for lotr2-
1/exo70a1-4 were used for further characterisation.
Basic Fuchsin staining of cleared roots
Whole seedlings were cleared in 0.24 N HCl with 20% methanol for 15 min at 57° C. The
acidified methanol was replaced by 7% NaOH in 60% ethanol and incubated at room
temperature (RT). Seedlings were rinsed twice in 70% ethanol, stained 5 min in 0.01% (w/v)
basic fuchsin and briefly rinsed in 70% ethanol. Seedlings were rehydrated in 40%, 20% 10%
and 5% ethanol for 5 min. The samples were equilibrated in 5% ethanol with 25% glycerol for
at least 15 min at RT or overnight at 4° C. Seedlings were mounted in 50% glycerol, stored at
4° C and imaged within 2 weeks.
ClearSee for simultaneous staining of fluorescent proteins, lignin and cellulose
Five- to six-day-old seedlings were fixed in 4% paraformaldehyde in 1x PBS for 45-60 min
and washed twice in 1x PBS. Seedlings were then transferred into ClearSee solution [10%
(w/v) xylitol, 15% (w/v) sodium deoxycholate, 25% (w/v) urea, dH2O] and cleared for 2-3
days. Seedlings were stained for lignin using 0.2% (w/v) Basic Fuchsin in ClearSee for 16h,
rinsed twice for 30 min and then stained for cellulose in 0.1% (w/v) Calcofluor White in
ClearSee for 45-60 min and again rinsed twice for 30 min. Stained seedlings were kept in
ClearSee until microscopy for up to 3 weeks. All steps were carried out at RT.
Transmission electron microscopy of immuno-gold-labelled ultrathin sections
For immuno-gold transmission electron microscopy of CASP1-GFP, plants were fixed for 1 h
at RT in 0.1 M phosphate buffer pH 7.4 (Sigma, St. Louis, MO), containing 1% glutaraldehyde,
16
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 16
SUPPLEMENTARY INFORMATION

0.1% osmium tetroxide (both EMS, Hatfield, PA) and 1.5% potassium ferrocyanide (Sigma,
St. Louis, MO). Samples were then washed twice in dH2O water and dehydrated in 30%, 50%,
70%, 100% and 100% ethanol (Sigma, St Louis, MO, US) for 30 min each. Samples were
infiltrated in 33% and 66% (v/v ethanol) LR White resin (Sigma, St Louis, MO, US) for 4 h
each, followed by a final infiltration in 100% LR White for 2-8 h and subsequent
polymerization for 48 h at 60° C under nitrogen. 60 nm ultrathin transversal sections were
made at 2 mm from the root tip, using a Leica Ultracut (Leica Mikrosysteme GmbH, Vienna,
Austria). Sections were placed on a 2 x 1 mm copper slot grid (EMS, Hatfield, PA, US) and
coated with a polystyrene film (Sigma, St Louis, MO, US).
For immuno-gold labeling, Ab6556 rabbit polyclonal antibody against GFP (Abcam,
Cambridge, UK) was used. Ultrathin sections were incubated in 50 mM ammonium chloride
(NH4Cl) (Sigma, St Louis, MO, USA) solution for 10 min to block aldehydes. Then, sections
were treated with 1% of acetylated bovine serum albumine solution (BSAcTM) (Aurion,
Wageningen, The Netherlands) in 0.15 M potassium phosphate buffered saline (PBS), pH 7.4
(Sigma, St Louis, MO, USA) for 10 min to inhibit unspecific binding of antibodies. Sections
were incubated with the primary antibody, diluted 1/1000 in 0.1% BSAcTM in PBS for 1 h at
RT, washed six times in 0.1% BSAcTM in PBS, and then incubated with the secondary
antibody (Protein A gold 10nm, UMC, Utrecht), diluted 1/70 in 0.1% BSAcTM in PBS for 1
hour at RT. Sections were washed 4 times in 0.1% BSAcTM in PBS, followed by four washing
steps in PBS, fixed in 1% glutaraldehyde solution (EMS, Hatfield, PA, USA) in PBS for 5 min,
washed ten times in H2O and finally stained with 2% uranyl acetate (Sigma, St Louis, MO,
USA) in H2O for 10 min and then rinsed several times with H2O.
Endodermal central localisation (ECL) index
The ECL index was calculated as the mean fluorescence intensity at the lateral plasma
membrane between endodermis and cortex divided by the mean fluorescence intensity between
two endodermal cells. In an attempt to conservatively assess the local accumulation between
endodermal cells, raw values were obtained from rectangles of equal areas to ensure that the
local accumulation was rather under- than overestimated. Since CASP1-GFP is exclusively
present in the endodermis, its ECL index was calculated as twice the lateral fluorescence
intensity divided by the central fluorescence intensity, which takes into account two plasma
membranes with CASP1-GFP as opposed to one plasma membrane at the lateral endodermal
cell face. The binary comparisons were performed employing a Student t-test. In case data was
17
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 17
SUPPLEMENTARY INFORMATION

not following a linear model assumption a Wilcoxon-Mann-Whitney test was used to assess
statistically significant difference.
18
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 18
SUPPLEMENTARY INFORMATION

Supplementary Table 1: Mutants identified in the LOTR screen
Gene Alternative name
Phenotypical class Mutants Reference
LOTR1 Class II Class II Class II Class II Class II Class II Class II Class II
lotr1-1 lotr1-2 lotr1-3 lotr1-4 lotr1-5 lotr1-6 lotr1-7 lotr1-8
Li et al., Current Biology, 2017
LOTR2 EXO70A1 Class I Class I
lotr2-1 (exo70a1-4) lotr2-2 (exo70a1-5)
This study
LOTR3 Class III lotr3 L. Kalmbach, unpublished
LOTR4 Class IV Class IV Class IV
lotr4-1 lotr4-2 lotr4-3
Marhavy et al., unpublished
LOTR5 Class IV lotr5 Marhavy et al., unpublished
LOTR6 Class IV Class IV
lotr6-1 lotr6-2
Marhavy et al., unpublished
LOTR7 Class IV Class IV
lotr7-1 lotr7-2
Marhavy et al., unpublished
LOTR8 Class IV lotr8 L. Kalmbach, unpublished
LOTR9 Class I lotr9 L. Kalmbach, unpublished
SGN1 Class III Class III Class III
sgn1-3 sgn1-4 sgn1-5
10
SGN3 GSO1 Class III Class III Class III Class III Class III Class III Class III Class III Class III Class III Class III Class III Class III Class III
sgn3-5 sgn3-6 sgn3-7 sgn3-8 sgn3-9 sgn3-10 sgn3-11 sgn3-12 sgn3-13 sgn3-14 sgn3-15 sgn3-16 sgn3-17 sgn3-18
11
SGN5 SCR Class V Class II
sgn5-2 sgn5-3
12 L. Kalmbach, unpublished
ESB1 Class I Class I Class I Class I
esb1-3 esb1-4 esb1-5 esb1-6
Reyt et al., unpublished
MYB36 Class VI Class VI
myb36-3 myb36-4
13
SHR Class VI Class VI
shr-7 shr-8
L. Kalmbach, unpublished
19
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 19
SUPPLEMENTARY INFORMATION
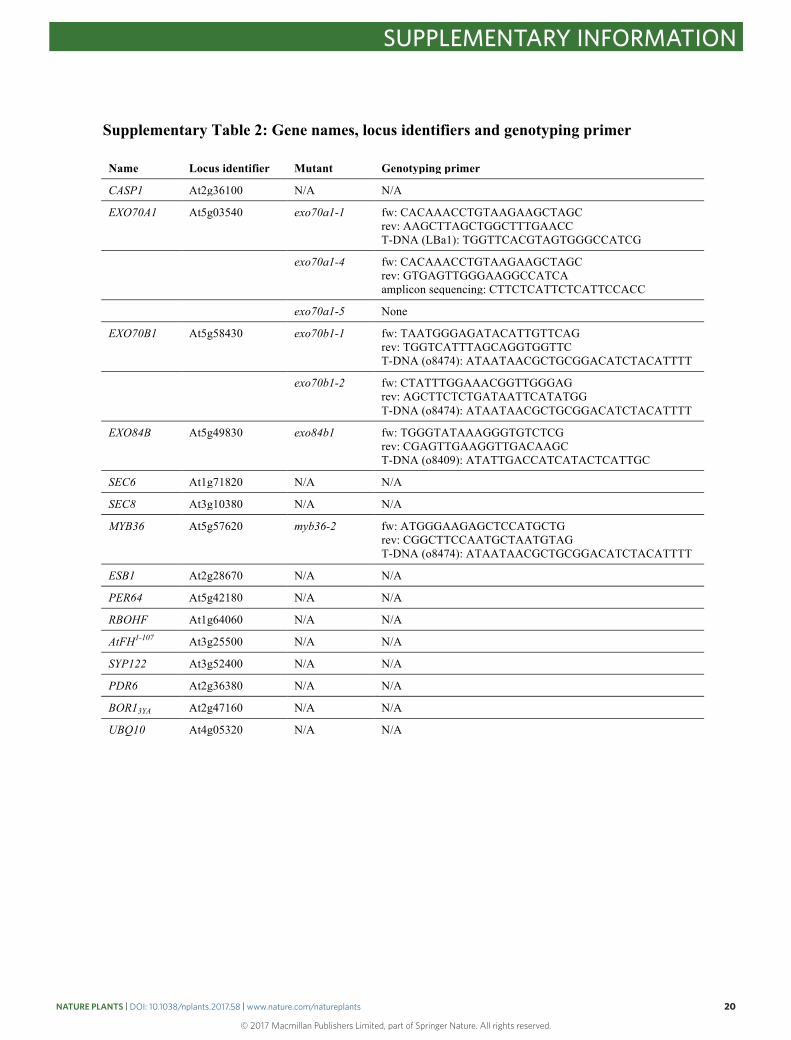
Supplementary Table 2: Gene names, locus identifiers and genotyping primer Name Locus identifier Mutant Genotyping primer
CASP1 At2g36100 N/A N/A
EXO70A1 At5g03540 exo70a1-1 fw: CACAAACCTGTAAGAAGCTAGC rev: AAGCTTAGCTGGCTTTGAACC T-DNA (LBa1): TGGTTCACGTAGTGGGCCATCG
exo70a1-4 fw: CACAAACCTGTAAGAAGCTAGC rev: GTGAGTTGGGAAGGCCATCA amplicon sequencing: CTTCTCATTCTCATTCCACC
exo70a1-5 None
EXO70B1 At5g58430 exo70b1-1 fw: TAATGGGAGATACATTGTTCAG rev: TGGTCATTTAGCAGGTGGTTC T-DNA (o8474): ATAATAACGCTGCGGACATCTACATTTT
exo70b1-2 fw: CTATTTGGAAACGGTTGGGAG rev: AGCTTCTCTGATAATTCATATGG T-DNA (o8474): ATAATAACGCTGCGGACATCTACATTTT
EXO84B At5g49830 exo84b1 fw: TGGGTATAAAGGGTGTCTCG rev: CGAGTTGAAGGTTGACAAGC T-DNA (o8409): ATATTGACCATCATACTCATTGC
SEC6 At1g71820 N/A N/A
SEC8 At3g10380 N/A N/A
MYB36 At5g57620 myb36-2 fw: ATGGGAAGAGCTCCATGCTG rev: CGGCTTCCAATGCTAATGTAG T-DNA (o8474): ATAATAACGCTGCGGACATCTACATTTT
ESB1 At2g28670 N/A N/A
PER64 At5g42180 N/A N/A
RBOHF At1g64060 N/A N/A
AtFH1-107 At3g25500 N/A N/A
SYP122 At3g52400 N/A N/A
PDR6 At2g36380 N/A N/A
BOR13YA At2g47160 N/A N/A
UBQ10 At4g05320 N/A N/A
20
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 20
SUPPLEMENTARY INFORMATION

Supplementary Table 3: Primer for Sanger sequencing of EXO70A1
Amplicon Primer for ~1000 bp amplicon Sequencing primer
1 fw: GGCTCGGCATTCCTCTGTC rev: GTGAGTTGGGAAGGCCATCA
seq: CTCTTTCTTTTTCTTCAACTTCC
2 fw: TTGGGAACTGGTCTTCGCTC rev: AGGAACGTGTGTCCCTGAAC
seq: CTATGATGGTCTTACTTCTTTAG
3 fw: TCCGCTGTTTGTGCTTACCT rev: TCTCTGTGTGCCATGCTGTT
seq: GTAATGCATTGGAGAAGCTTTG
4 fw: AGGATGCTGCAAACGTCTAGT rev: CCACTGCTGTTTCCTCCCTC
seq: TTGACACTGACACACATAACG
5 fw: AATTCGCAGCTAGCATCCGT rev: CCAAAGCTCGTCTCAAAGCA
seq: ACATGGTCAGATCTGTGCGC
21
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 21
SUPPLEMENTARY INFORMATION

Supplementary References
1. Liberman, L. M., Sparks, E. E., Moreno-Risueno, M. A., Petricka, J. J. & Benfey, P.
N. MYB36 regulates the transition from proliferation to differentiation in the
Arabidopsis root. Proc Natl Acad Sci USA 112, 12099–12104 (2015).
2. Li, S., Yamada, M., Han, X., Ohler, U. & Benfey, P. N. High-Resolution Expression
Map of the Arabidopsis Root Reveals Alternative Splicing and lincRNA Regulation.
Dev Cell 39, 508–522 (2016).
3. Roppolo, D. et al. A novel protein family mediates Casparian strip formation in the
endodermis. Nature 473, 380–383 (2011).
4. Kim, Y., Schumaker, K. S. & Zhu, J.-K. EMS mutagenesis of Arabidopsis. Methods
Mol. Biol. 323, 101–103 (2006).
5. Alassimone, J. et al. Polarly localized kinase SGN1 is required for Casparian strip
integrity and positioning. Nature Plants 2, 16113 (2016).
6. Leshchiner, I. et al. Mutation mapping and identification by whole-genome
sequencing. Genome Res. 22, 1541–1548 (2012).
7. Hellens, R. P., Edwards, E. A., Leyland, N. R., Bean, S. & Mullineaux, P. M. pGreen:
a versatile and flexible binary Ti vector for Agrobacterium-mediated plant
transformation. Plant Mol Biol 42, 819–832 (2000).
8. Quan, J. & Tian, J. Circular polymerase extension cloning of complex gene libraries
and pathways. PLoS ONE 4, e6441 (2009).
9. Gibson, D. G. et al. Enzymatic assembly of DNA molecules up to several hundred
kilobases. Nat Meth 6, 343–345 (2009).
10. Alassimone, J. et al. Polarly localized kinase SGN1 is required for Casparian strip
integrity and positioning. Nature Plants 2, 16113 (2016).
11. Pfister, A. et al. A receptor-like kinase mutant with absent endodermal diffusion
barrier displays selective nutrient homeostasis defects. Elife 3, (2014).
12. Alassimone, J. Molecular Dissection of Arabidopsis thaliana Endodermis
Development, Structure and Function. (PhD Thesis). (2012).
13. Kamiya, T. et al. The MYB36 transcription factor orchestrates Casparian strip
formation. Proc Natl Acad Sci USA 201507691–6 (2015).
doi:10.1073/pnas.1507691112
22
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.58 | www.nature.com/natureplants 22
SUPPLEMENTARY INFORMATION
Top Related