







Languages
Pages
Legal


Structural studies of small RNA phages
Lars Liljas, Uppsala UniversityStructure and Function of Large Molecular Assemblies
Erice, June 12, 2006
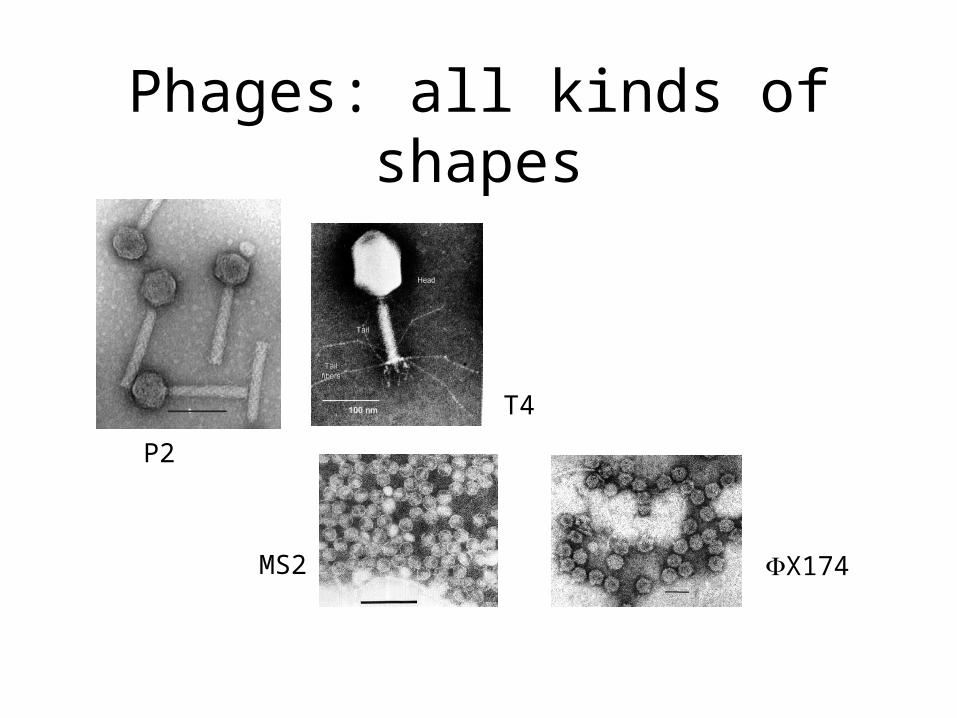
Phages: all kinds of shapes
P2
T4
MS2 X174

Phage MS2 infects E. coli
The virus particles attach to bacterial f-pili

Maturation Coat (0) Replicase (0)Lysis (+1)
Maturation-lysis CoatRead-through (A1)
Replicase
130 1308 1335 1724 1761 33951 3569
1678 1902
62 1305 1325 1743 2353 4119
4220
1
2331
MS2
Q •Coat protein binding site
RNA of leviviruses codes for 3-4 RNA of leviviruses codes for 3-4 proteins proteins
The genetic maps of The genetic maps of LevivirusLevivirus phage MS2 and phage MS2 and AllolevivirusAllolevivirus phage Q phage Q

MS2: function of components
• RNA: single-stranded positive sense• A-protein: attachment and entry• Coat protein: protein coat, translational repression
and RNA recognition• Replicase subunit: forms active RNA-dependent
RNA polymerase with host proteins• Lysis protein: lyses bacterial cell wall to release
new virions

MS2: components
• How is assembly controlled?
• What determines the specificity in the binding of coat protein to the translational operator/assembly initiation site in RNA?
• What is the position and conformation of the A protein?
• How is the replicase working?

The replicase
• The RNA polymerase is formed by – Replicase subunit (coded by viral RNA)– Elongation factors EF-Tu and EF-Ts– Ribosomal protein S1 (- strand synthesis)
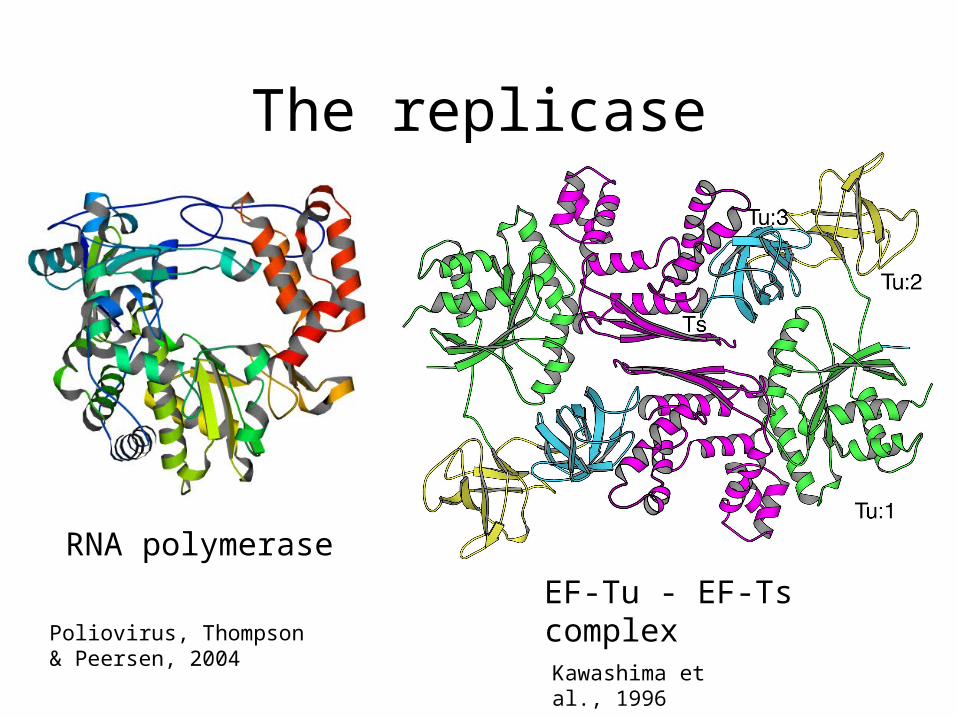
The replicase
RNA polymerase
EF-Tu - EF-Ts complexKawashima et al., 1996
Poliovirus, Thompson & Peersen, 2004

Levivirus capsid structures
Phage Resolution Particles
MS2 2.2 Å Phage and recombinant VLP
Q 3.5 Å Phage and recombinant VLP
fr 3.5 Å Phage
GA 3.4 Å Phage
PP7 3.7 Å Phage
PRR1 3.5 Å Recombinant VLP

Department of Cell and Molecular Biology, Uppsala UniversityKarin ValegårdKerstin Fridborg Roshan GolmohammadiSjoerd van den WormElin GrahnKaspars TarsCharlotte HelgstrandMagnus JohanssonPavel Plevka
Department of Molecular Biology, Latvian University, RigaMaija Bundule
School of Biology, University of LeedsNicola StonehousePeter StockleyWilf Horn
Small RNA phages

The virus particle is formed by • 90 coat protein dimers (180 subunits, T = 3) • 1 copy of A-protein• RNA molecule
MS2 particle

The coat protein dimer has a central sheet facing the interior. Helices and a hairpin form the outer surface
MS2 particle
B/A
CC

Role of A-proteinAttachment to pili is due to the A-protein
A fragment of the A-protein enters bacterium with viral RNA
The A-protein, 393 aa, has no sequence similarity to other proteins
Attempts to isolate (soluble) protein has failed

Role of A-protein
Where is the A-protein?How does the binding trigger release of the RNA?
Particle has holes at 5-fold and 3-fold axesPart of the A-protein might be exposed in one of these holes

Position of A-protein
Phage PRR1, Johansson et al., in prep. (Poster 26)
Fivefold and threefold axes surrounded by flexible loops
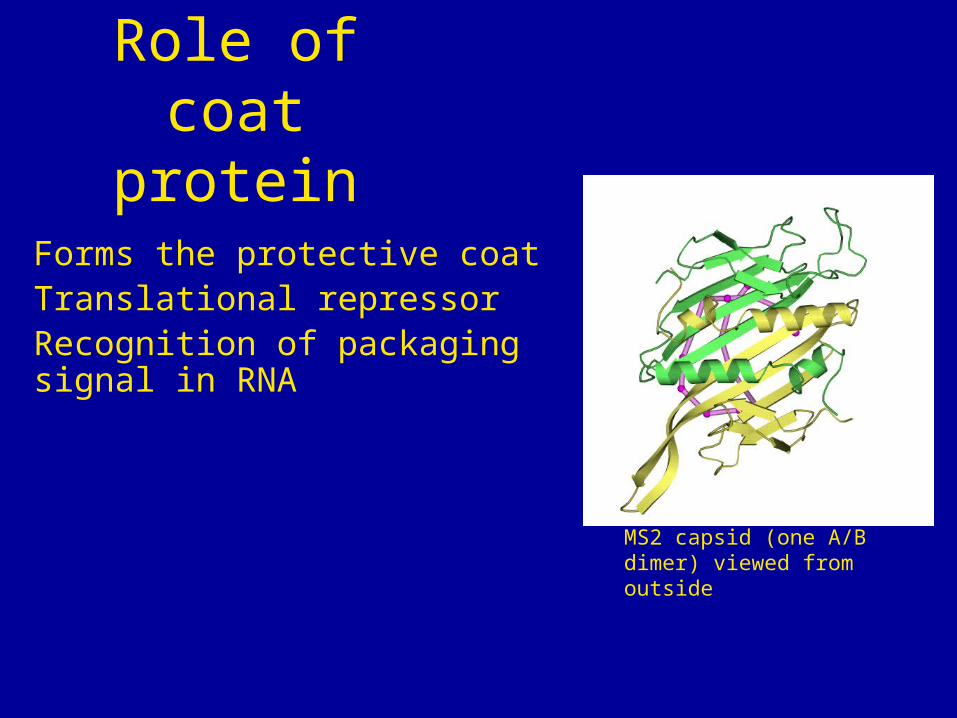
Forms the protective coatTranslational repressorRecognition of packaging signal in RNA
Role of coat protein
MS2 capsid (one A/B dimer) viewed from outside

Coat protein binding to RNA
A hairpin in the RNA is used both as a translational operator for the replicase and a packaging signal
-5 U U A A -4 G C G C -10 A G C -1 U A +1 A U C G -15 A U +4
A protein Coat Replicase
Lysis
Operator hairpin

Coat protein binding to RNA
-5 U U A A -4 G C G C -10 A G C -1 U A +1 A U C G -15 A U +4
N PyA A N N’ N N’Pu N N’ N N’ N N’ N N’ N N’
In MS2, four positions in single-stranded regions are recognized with specificity

RNA bindingCrystals of recombinant capsids can be soaked with RNA hairpins, which enter the particles and bind to the coat protein
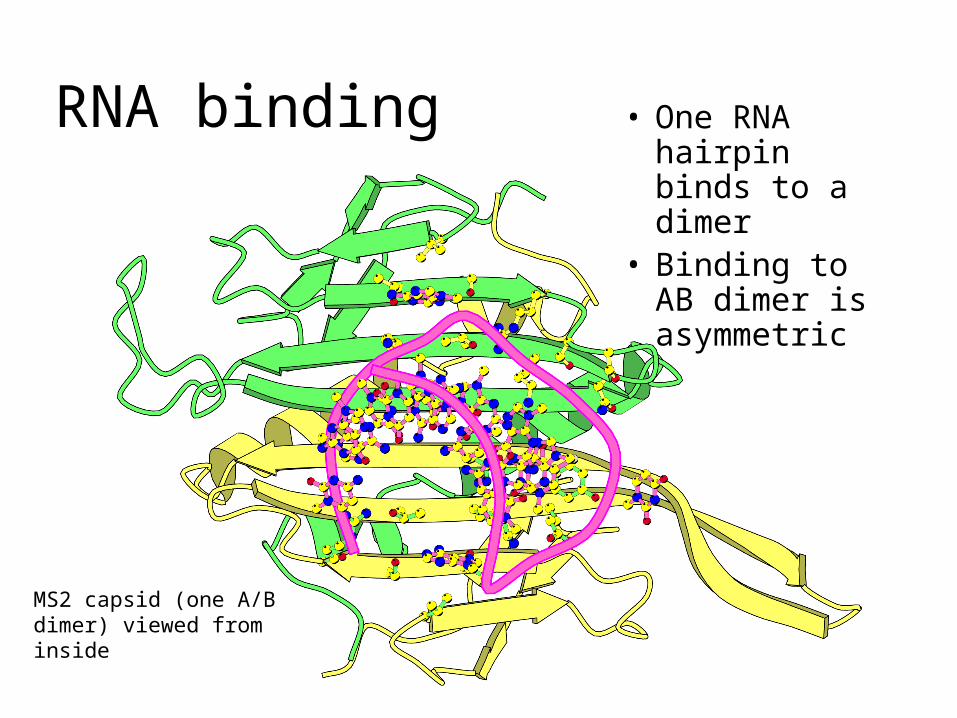
• One RNA hairpin binds to a dimer
• Binding to AB dimer is asymmetric
RNA binding
MS2 capsid (one A/B dimer) viewed from inside

Single-stranded regions of the RNA binds to three conserved sites in the protein
RNA recognition in MS2

What is determining the specificity?
How are differences in specificity between different phages achieved?
RNA recognition in MS2

-5 U U A A -4 G C G C-10 A G C -1 U A +1 A U C G -15 A U +4
A pocket with specificity for A
RNA recognition in
MS2: A-4
-5 U U A A -4 G C G C-10 A G C -1 U A +1 A U C G -15 A U +4
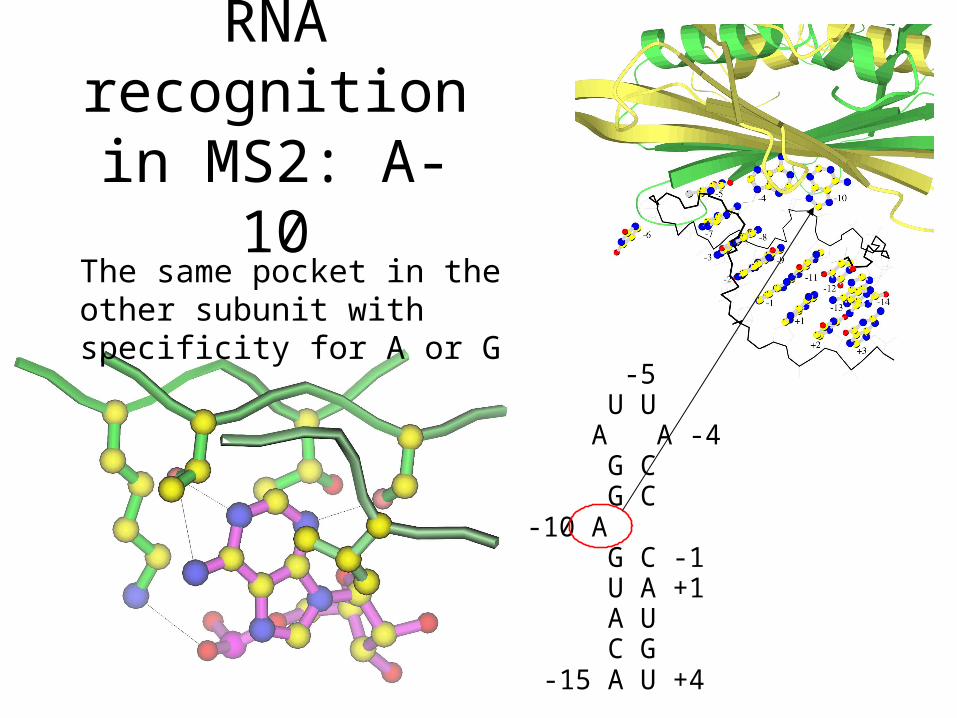
The same pocket in the other subunit with specificity for A or G
RNA recognition in
MS2: A-10

-5 U U A A -4 G C G C-10 A G C -1 U A +1 A U C G -15 A U +4
A-10
G-10
C-10
RNA recognition in
MS2: A-10

-5 U U A A -4 G C G C-10 A G C -1 U A +1 A U C G -15 A U +4
A binding site with specificity forU or C
RNA recognition in MS2: U-5

Of 9 natural and variant bases tested, 8 bind similar to wildtype U although the affinity differs by several orders of magnitude
U-5: 1A-5: 0.01
C-5: 10
RNA recognition in MS2: U-5
-5 U U A A -4 G C G C -10 A G C -1 U A +1 A U C G -15 A U +4
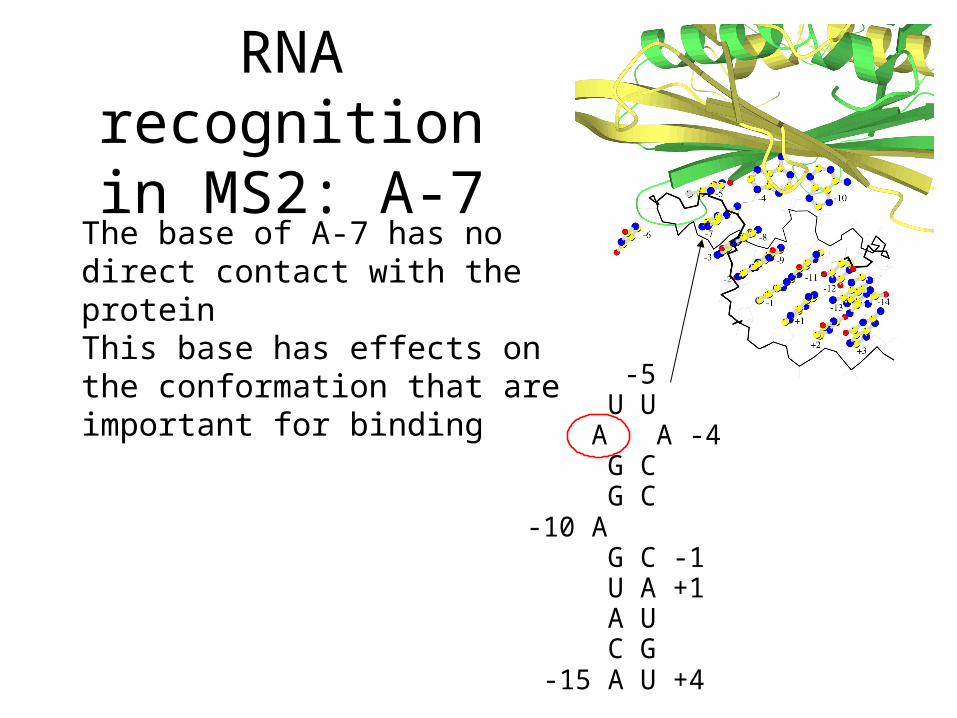
The base of A-7 has no direct contact with the proteinThis base has effects on the conformation that are important for binding
RNA recognition in MS2: A-7

RNA binding: specificity
-5 U U A A -4 G C G C-10 A G C -1 U A +1 A U C G -15 A U +4
The RNA-binding surface of the coat protein is very conserved, but Q coat protein binds to a hairpin with different structure
A U A +8 C G U A G C U A +1 A C G G C
MS2 Q
RNA binding
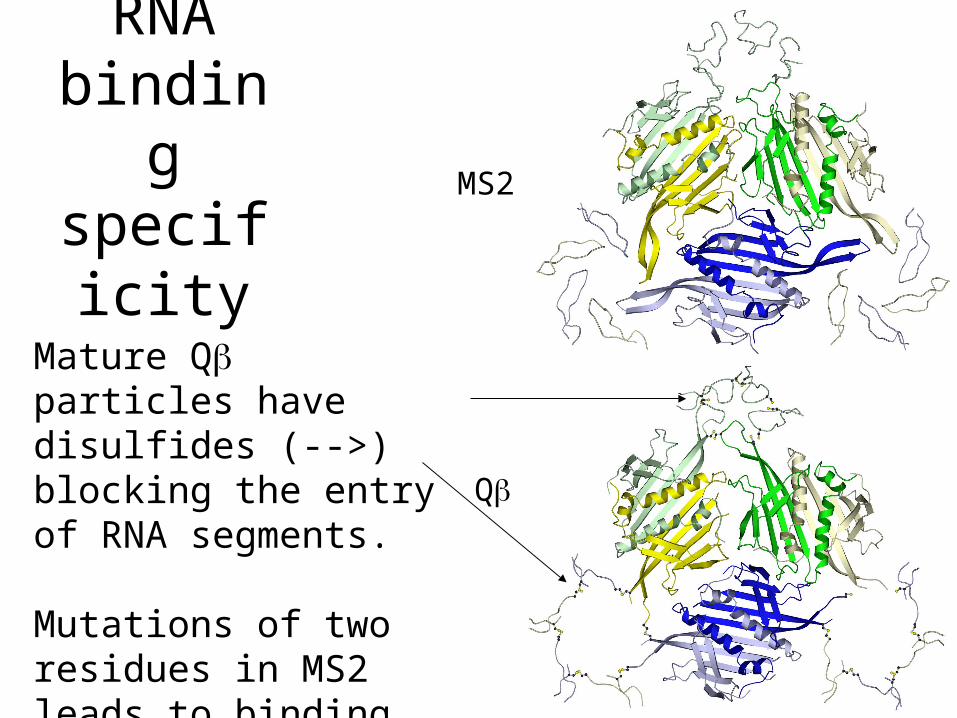
specificity
Mature Q particles have disulfides (-->) blocking the entry of RNA segments.
Mutations of two residues in MS2 leads to binding also of Q hairpin
MS2
Q

RNA binding: specificity
A U A G
U
C C :
TYR
Conformation of loop very similar and the protein-RNA contacts are conserved but the basepairs are shifted
A A G:C A:U
U TYR
MS2 hairpin Q hairpin

RNA binding: specificity
-5 U U A A -4 G C G C -10 A G C -1 U A +1 A U C G -15 A U +4
Phage PRR1 has still another variant of hairpin structure
A U A C G U A G C U A +1 A C G G C
MS2 Q PRR1
C G U A A G C G C A A U U A +1 A U C G C G

Assembly of MS2 particles
Dimers assemble into correctly formed T=3 particles

Two similar dimer conformations allow formation of quasi-equivalent contacts
Subunit packing in MS2
B/A
CC

In leviviruses, RNA stimulates assembly:•RNA binding to dimer - induces “assembly mode”•Strong binding to one site (“initiation site”) - controls encapsidation of correct RNA
Role of RNA in subunit packing

Control of quasiequivalence in other viruses
In many small viruses, a flexible part of the coat protein is used to control the packing of subunits – used as order-disorder switches

Control of quasi-equivalence
Leviviruses have no arms and order/disorder switches - assembly is controlled by the contact surfaces

Conclusions
The observed specificity of coat protein binding to the viral RNA is explained by the structure of the complex
The coat protein is able to form T=3 capsids without using an arm and order/disorder switches
Top Related