







Languages
Pages
Legal


.
Maximum Likelihood (ML) Parameter Estimation
with applications to inferring phylogenetic trees
Comput. Genomics, lecture 7a
Presentation partially taken from Dan Geiger, modified by Benny Chor.
Background reading: Durbin et al Chapter 8.

2
Our Probabilistic Model (Reminder)
Now we don’t know the states at internal node(s), nor
the edge parameters pe1, pe2, pe3
XXYXY YXYXX
YYYYX
pe1
pe2
pe3
A single edge is a fairly boring tree…
?????
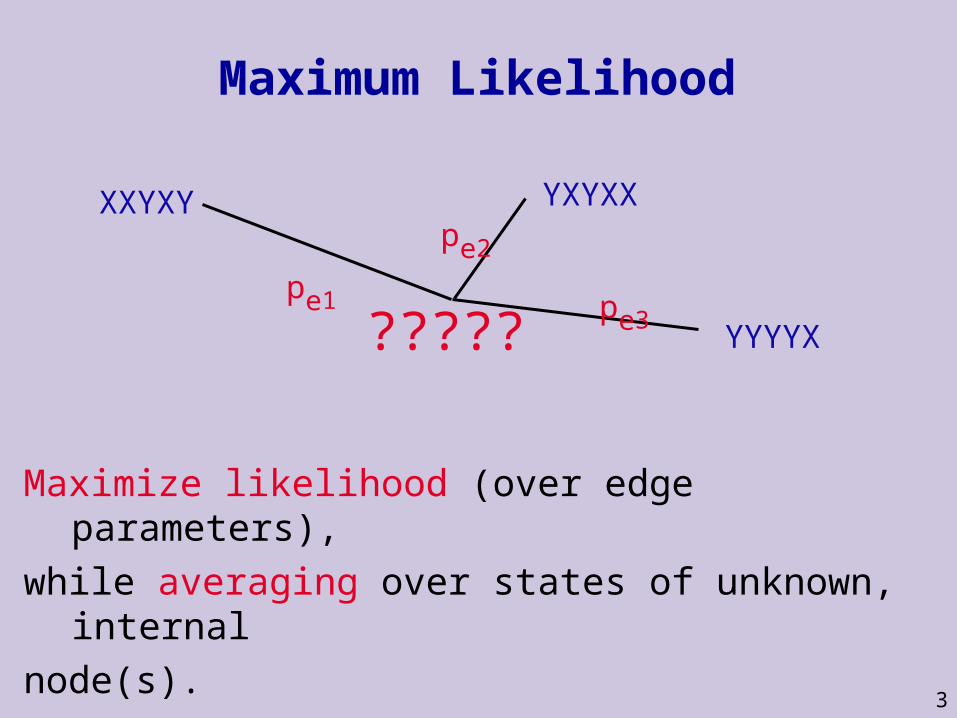
3
Maximum Likelihood
Maximize likelihood (over edge parameters),
while averaging over states of unknown, internal
node(s).
XXYXY YXYXX
YYYYX
pe1
pe2
pe3?????

4
Maximum Likelihood (2)Consider the phylogenetic tree to be a stochastic process.
XYXYYX
XXXXXY
XXX XYX
XXX
The probability of transition from character a to character b along edge e is given by parameters pe.
Given the complete tree, the likelihood of data is determined by the values of the pe‘s.
Observed
Unobserved

5
Maximum Likelihood (3)
We assume each site evolves independently of the others.
XY
XX
This allows us to decompose the likelihood of the data (sequences at leaves) to the product of each site, given the (same) tree and edge probabilities. This is the first key to an efficient DP algorithm for the tiny ML problem.(Felsenstein, 1981).
Will now show how Pr(D(i)|Tree, ) is efficiently computed.
YY
XX
XX
XY
Pr(D|Tree, )=i Pr(D(i)|Tree, )

6
X
p1 p2
tree1 tree2
Let T be a binary tree with subtrees T1 and T2.
Let Lx(D | T, ) be the
likelihood of T with X
at T’s root.
Define LY(D | T, )
similarly.
Computing the Likelihood
Y
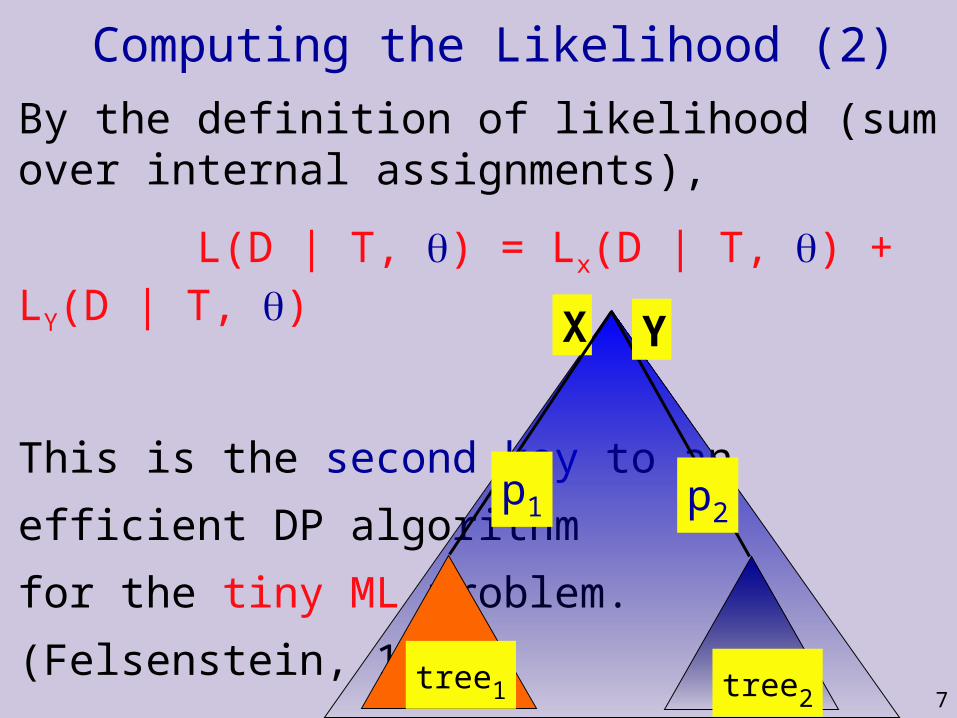
7
By the definition of likelihood (sum over internal assignments),
L(D | T, ) = Lx(D | T, ) + LY(D | T, )
This is the second key to an
efficient DP algorithm
for the tiny ML problem.
(Felsenstein, 1981)
Computing the Likelihood (2)
X
p1 p2
tree1 tree2
Y

8
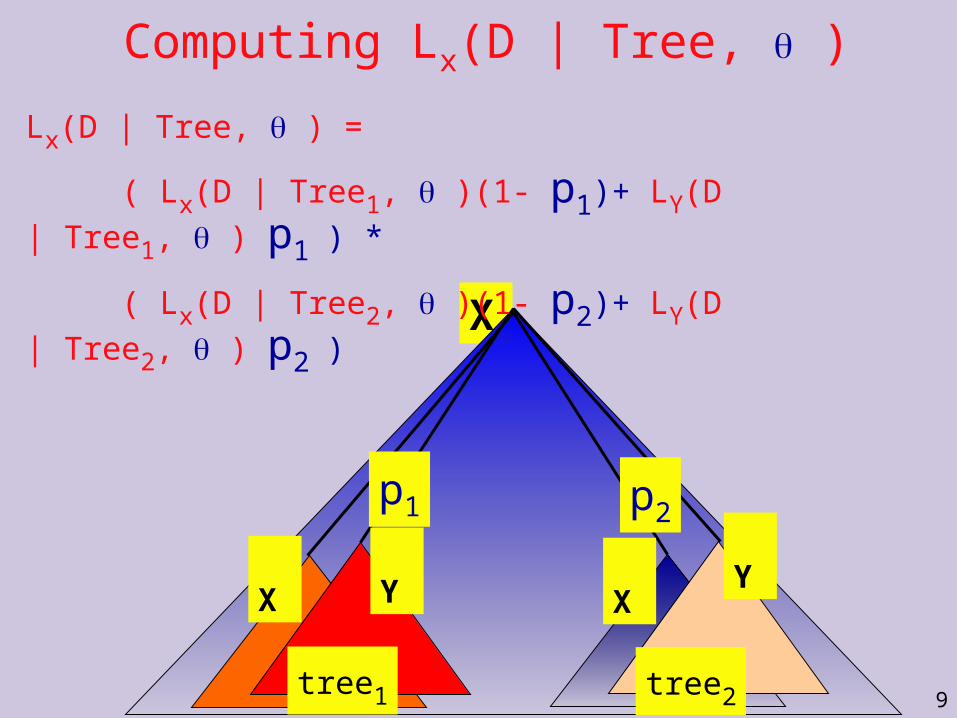
Computing Lx(D | Tree, ) X
p1 p2
X Y X Y
tree1 tree2
9
Computing Lx(D | Tree, )
X
p1 p2
X Y X Y
tree1 tree2
Lx(D | Tree, ) =
( Lx(D | Tree1, )(1- p1)+ LY(D | Tree1, ) p1 ) *
( Lx(D | Tree2, )(1- p2)+ LY(D | Tree2, ) p2 )

10
The Dynamic Programming Algorithm
X
p1 p2
X
Y
X
Y
tree1 tree2
The algorithm starts from the leaves and proceeds up towards the root. For each sub-tree visited, keep both Lx(D | sub-tree, ) and LY(D | sub-tree, ). This enables computing Lx and LY likelihoods w.r.t T using 5 multiplications and 2 additions.

11
The Dynamic Programming Algorithm
X
p1 p2
X
Y
X
Y
tree1 tree2
The algorithm thus takes O(1) floating point operations per internal node of the tree. If there are n leaves, the number ofinternal nodes is n-1, so overall complexity is O(n).

12
What About Initialization?
X
p1 p2
X Y XY
tree1 tree2
Well, this is easy. If T is a leaf that contains X, then Lx(D | T, ) = 1, and Lx(D | T, ) = 0.( the case where T is a leaf that contains Y is left as a bonusassignment )

13
A Few More Question Marks
X
p1 p2
X Y XY
tree1 tree2
• What if tree is not binary? Would it not effect complexity…• What if tree unrooted? Can show symmetry of substitution probabilities implies likelihood invariant under choice of roots.• Numerical questions (underflow, stability).• Non binary alphabet.
14
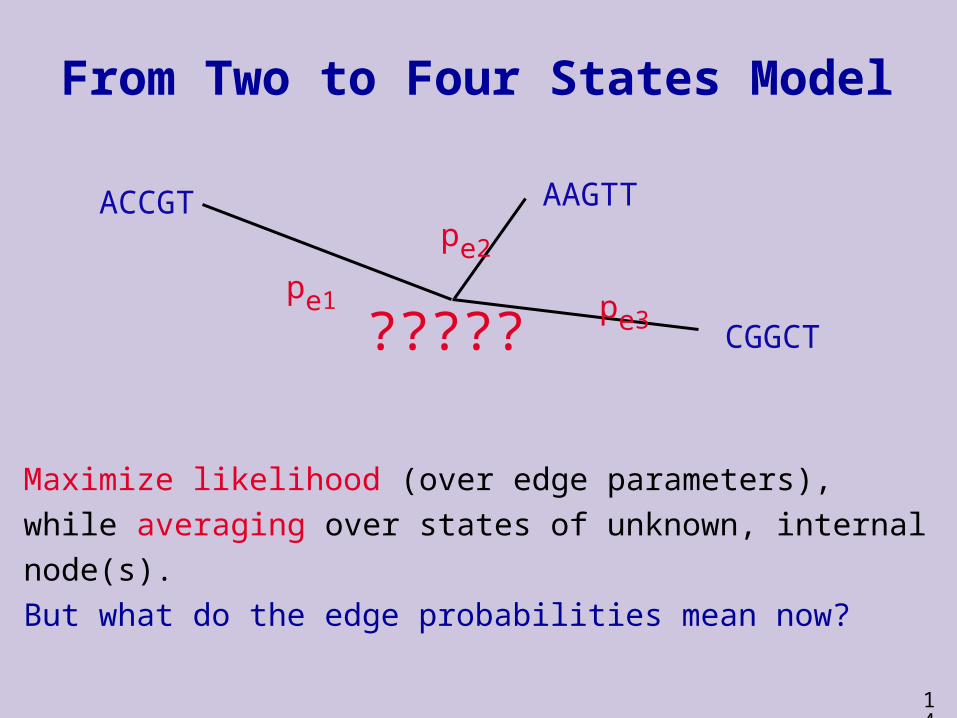
From Two to Four States Model
Maximize likelihood (over edge parameters),
while averaging over states of unknown, internal
node(s).
But what do the edge probabilities mean now?
ACCGT AAGTT
CGGCT
pe1
pe2
pe3?????

15
From Two to Four States Model (2)
So far, our models consisted of a “regular” tree, where in addition, edges are assigned substituion probabilities.
For simplicity, assumed our “DNA” has only two
states, say X and Y. If edge e is assigned probability pe , this means
that the probability of substitution (X Y)
across e is pe .
Now a single pe can no longer express all 16-4=12
possible substitution probabilities.

16
From Two to Four States Model Now a single pe can no longer express all 16-4=12
possible substitution probabilities. The most general model will indeed have 12
independent parameters per edge, e.g. pe (C->A),
pe (T->A), etc. It need not be symmetric.
Still, most popular models are symmetric, and use
far less parameters per edge. For example, the Jukes-Cantor substitution model
assumes equal substitution probability of any unequal pair of nucleotides (across each edge separately).

17
The Jukes-Cantor model (1969)
Jukes-Cantor assume equal prob. of change:
GA
TC
1-31 3
1 3Subst. Prob.
1 3
1 3
A C G T
A
C
G
T

18
Tiny ML on Four States : Like Before, Only More Cases
Can handle DNA subst. models, AA subst. models, ... Constant (per node) depends on alphabet size.
subtree)} ()({
subtree)} ()({
)(
rightPNGP
leftPNGP
subtreeP
NsNucleotideN
NsNucleotideN
G
A C G T
A C G TA C G T
P(GC) *PC(left subtree)
GleafG leafP
tionInitialisa
,)(

19
Kimura’s K2P model (1980)Jukes-Cantor model does not take into account that transitions rates (between purines) AG and (between pyrmidine) CT are different from transversions rates(AC, AT, CG, GT).
Kimura 2 parameter model uses a different substitution matrix:
1 2
1 2Subst. Prob.
1 2
1 2
A C G T
A
C
G
T

20
Kimura’s K2P model (Cont)
tttt
tttt
tttt
tttt
rsus
srsu
usrs
susr
tS )(
ttt
ttt
tt
usr
eeu
es
21
214
1
14
1
)(24
4
Leading using similar methods to:
Where:

21
Additional Models
There are yet more involved DNA substitution models, responding to phenomena occurring in DNA.
Some of the models (like Jukes-Cantor, Kimura 2 parameters, and others) exhibit a “group-like” structure that helps analysis.
The most general of these is a matrix where all rates of change are distinct (12 parameters).
For AA (proteins), models typically have less structure.
Further discussion is out of scope for this course. Pleaserefer to the Molecular Evolution course (life science).

22
Back to the 2 States Model
Showed efficient solution to the tiny ML problem.
Now want to efficiently solve the tiny AML problem.
XXYXY YXYXX
YYYYX
pe1
pe2
pe3?????

23
Two Ways to Go
In the second version (maximize over states of internal nodes)
we are looking for the “most likely” ancestral states. This is
called ancestral maximum likelihood (AML).
In some sense AML is “between” MP (having ancestral states)
and ML (because the goal is still to maximize likelihood).
XXYXY YXYXX
YYYYX
pe1
pe2
pe3?????

24
Two Ways to Go
In some sense AML is “between” MP (having ancestral states)
and ML (because the goal is still to maximize likelihood).
The tiny AML algorithm will be like Fitch small MP algorithm:
It goes up to the root, then back down to the leaves.
XXYXY YXYXX
YYYYX
pe1
pe2
pe3?????

25
Let T be a binary tree with subtrees T1 and T2.
Let LE(D | T, ) be the
ancestral likelihood of T
with E (X or Y) at the
node of T’s father.
Computing the Ancestral Likelihood
X
p1 p2
tree1 tree2
Y
Ep

26
By the definition of ancestral likelihood (maximizing over internal assignments),
LX(D| T, ) =
max((1-p)Lx(D | tree1, ) * Lx(D | tree2, ) ,
pLY(D | tree1, )* LY(D | tree2, ))
This is key to an efficient DP
algorithm for the tiny AML
problem (Pupko et. al, 2000)
Computing the Ancestral Likelihood (2)
X
p1 p2
tree1 tree2
Y
Xp

27
Boundary conditions: At leaves
LX(D| T, ) = 1-p if leaf label is X, p otherwise.
At root: We pick label E (X or Y) that maximizes LE(D | tree1, ) LE(D | tree2, ).
We now go down the tree. At each node we pick
the label that maximizes the likelihood,
given the (known) label of father.
Total run time is O(n).
Computing the Ancestral Likelihood (2)
X
p1 p2
tree1 tree2
Y
Xp
Top Related