







Languages
Pages
Legal


FATTY ACIDS AS ENERGY SOURCE
Fatty acid oxidation is the major source of energy for ATP
synthesis in the human.
This process generates acetyl CoA and energy in the form of
ATP. Oxidation of the acetyl CoA in TCA cycle produces
additional ATP.
Fatty acids are not a significant source of fuel for the brain and
other nervous tissue, and they cannot by utilized by red blood cells,
which have no mitochondria.
The liver obtains energy from β-oxidation of fatty acids, but
much of the acetyl CoA is converted within mitochondria to the
ketone bodies acetoacetate and β-hydroxybutyrate, which are
released into blood (KB serve as fuels also for muscle and kidney).
Tissue oxidize fatty acids whenever their concentration increases
in the blood. Fatty acids are predominant fuel during fasting when
they are released from adipose triacylglycerols.
The need for energy in the form of ATP controls the rate of
fatty acid oxidation.
Although β-oxidation is the major process for obtaining energy
from fatty acids, α-, ω- and peroxisomal oxidation also occur.
Peroxisomal fatty acid oxidationis similar to mitochondrial β-
oxidation except that it directly uses molecular oxygen and does not
produce ATP via electron transport chain.

FATTY ACID ACTIVATION
FA must be activated before they can be oxidized
R-COO-
fatty acid+ ATP
EPPi
thiokinase
O
R - C ~ O ~ P - O – adenosine
O– acyl adenylate intermediate
CoASH
O
R - C - S CoA + acyl CoA
E
AMP
Overall, activation process consumes two high energy phosphate bonds,
one in the formation of the fatty acyl-AMP and the second in the cleavage of
pyrophosphate.
Metabolism of fatty acids
Size class Number of
carbons
Site of catabolism
Membrane transport
Short chain 2-4 mitochondrion free diffusion
Medium chain
4-12 mitochondrion carnitine cycle
Long chain 12-20 mitochondrion carnitine cycle
Very long chain
> 20 peroxisome unknown

TRANSPORT OF LONG CHAIN FATTY ACIDS INTO MITOCHONDRIA
Carnitine is obtained from the diet or synthesized from amino acid lysineThe carnitine shuttle
R C O
O
(CH3)3 – N – CH2 – CH – CH2 – COO-+
acyl carnitine
Transfer of fatty acyl group from CoA to carnitine → carnitine : palmitoyltransferase I – CPTI (outer membrane)Fatty acyl carnitine cross the membrane → translocaseFatty acyl group is transferred back to CoA → CPT-IICarnitine returns to cytosolic site → translocaseLong chain fatty acyl CoA is substrate for β-oxidation
AMPATP
PPiCoA
fatty acid fatty acid CoACoA
E
thiokinase
E
CPT-I
cytosol
outer mitochondrial membrane
E
CPT-II
E
Carnitine-acyl carnitine
translocase
inner mitochondrial membrane
CoACoA
carnitine carnitine
fatty acid CoA
fatty acyl carnitine
carnitineCoA
carnitine
CoAfatty acyl
CoA
+ +
β-oxidation

Energy yield from glucose and palmitate
Substrate Molecular weight
Net ATP yield (mol/mol)
mol ATPg
Caloric value (Cal/g)
glucosepalmitate
180256
36-38129
0.20.5
49


Fatty acids that contain an odd number of carbon atoms undergo β-oxidation, producing acetyl CoA, until the last spiral, when five carbons remain in the fatty acyl CoA. In this case, cleavage by thiolase produces acetyl CoA and propionyl CoA. Carboxylation of propionyl CoA yields methylmalonyl CoA, which is ultimately converted to succinyl CoA.
The propionyl CoA-to-succinyl CoA pathway is a major anaplerotic route for the TCA cycle and is used in the degradation of valine, isoleucine, and a number of other compounds. In the liver, this route provides precursors of oxaloacetate, which is converted to glucose. Thus, this small proportion of the odd-carbon-number fatty acid chain can be converted to glucose. In contrast, the acetyl CoA formed from β-oxidation of even-chain-number fatty acids in the liver either enters the TCA cycle, where it is principally oxidized to CO2, or it is converted to ketone bodies.
ODD-CHAIN-LENGTH FATTY ACIDS

OXIDATION OF ODD CHAIN FATTY ACIDSβ-oxidation untill the last spiral, when 5 carbons remain in the fatty acyl CoA → THIOLASE → acetyl CoA + propionyl CoA
ω
O
C ~ SCoA
O
CH3 – CH2 – C ~ SCoA
O
CH3 – C ~ SCoA
Propionyl CoA Acetyl CoA
Carboxylation of propionyl CoA → methylmalonyl CoA → succinyl CoA → to TCA cycle
O
CH3 - CH2 - C - S - CoA propionil CoA
Propionyl CoA metabolism of odd-chain fatty acids
O O
-O - C - CH2 - CH2 - C - S - CoA succinyl CoA
C - S - CoA
H - C - CH3
COO-D-methylmalonyl CoA
O
C - S - CoA
H3C - C - CH
COO-
L-methylmalonyl CoA
O
E
E
E
CO2propionyl CoA carboxylaserequires biotin
methylmalonyl CoA racemase
methylmalonyl CoA mutase requires vitamin B 12, cobalamin

ESSENTIAL FATTY ACIDS
Two series of fatty acids – having double bonds
three carbons from the methyl end (ω-3 fatty acid)
and six carbons from the methyl end (ω-6 fatty
acid) – are required for the synthesis of eicosanoids.
Mainly, they are obtained from dietary plant oils:
these oils contain the ω-6 fatty acid, linoleic acid
and the ω-3 fatty acid, linolenic acid. Linoleic acid
is converted by to arachidonic acid, the precursor
for the synthesis of prostaglandin and other
eicosanoids in man.

OXIDATION OF UNSATURATED FATTY ACIDS
Approximately one half of the fatty acids in the human diet are unsaturated, containing cis double bonds, with oleate (C18:1, Δ9) and linoleate (18:2, Δ9,12) being the most common. Linoleate is obtained from the diet and cannot be synthesized in the human; thus, it is considered an essential fatty acid. Therefore, only that portion of linoleate that is not needed for other processes will be oxidized. For unsaturated fatty acids to undergo the β-oxidation spiral, their cis double bonds must be isomerized to trans double bonds that will end up between the second and third carbons during β-oxidation, of the double bond must be reduced. The process is illustrated for the polyunsaturated fatty acid linoleate.
1.Linoleate undergoes β-oxidation until one double bond is between carbons 3 and 4 near the carboxyl end and the other is between carbons 6 and 7. 2. An isomerase moves the double bond from the 3,4-position so that it is trans and in the 2,3-position, and β-oxidation continues. 3. When a conjugated pair of double bonds is formed (two double bonds separated by one single bond) at positions 2 and 4, an NADPH-dependent reductase reduces the pair to one trans double bond at position 3. 4. Then isomerization and β-oxidation resume.Fig. 1

Because the β-carbon of phytanic acids is at a branch point, it is not possible to oxidize this carbon to a ketone. The first and essential step in catabolism of phytanic acids is microsomal α-oxidation to pristanic acid, releasing the (α-)carbon-1 as carbon dioxide. Thereafter acetyl CoA and propionyl CoA are released alternately and in equal amounts.
Fatty acids that are not readily oxidized by the enzymes of β-oxidation enter alternative pathways of oxidation, including peroxisomal β- and α-oxidation and microsomal ω-oxidation. The function of these pathways is to convert as much as possible of the unusual fatty acids to compounds that can be used as fuels or biosynthetic precursors.

DEFICIENCIES IN CARNITINE METABOLISM
Occurs in infancy and is offen life threatening.
Characteristic features: hypoketotic hypoglycemia,
hyperammonemia, and altered plasma free carnitine concentration.
Hepatic damage, cardiomyopathy, and muscle weakness are
common.
Homozygous deficiencies in carnitine transport, CPTs-I and –II
and the translocase result in defects in long-chain fatty acid
oxidation.
Treatment: carnitine supplementation, frequent high-
carbohydrate feeding, avoidance of fasting.
LIPID ABNORMALITIES IN ALCOHOLISM
Higher serum concentration of TG and cholesterol →
infiltration of liver cells with fat.
Metabolism of alcohol produces increased hepatic NADH and
hydrogen ions. Increased NADH + H+/NAD+ rations inhibit the
oxidation of fatty acids.
Fatty acids reaching the liver are re-esterified with glycerol
to form TG.

FATTY ACID SYNTHESIS
The synthesis of fatty acids in mammalian systems can be considered as a two-stage process, both stages requiring acetyl Coenzyme A (acetyl CoA) and both stages using multifunctional proteins in multienzyme complexes:
• Stage 1: involves acetyl CoA carboxylase,• Stage 2: involves fatty acid synthetase.
In the first stage acetyl CoA is converted to malonyl CoA under the action of the enzyme, acetyl CoA carboxylase. This is a biotin-dependent enzyme with distinct enzyme functions and a carrier protein function: a biotin carboxylase, a transcarboxylase, and a biotin-carboxyl-carrier protein. 1.the carboxylation of biotin, involving adenosine triphosphate (ATP) 2.the transfer of this carboxyl group to acetyl CoA to produce the end-product, malonyl CoA, releasing the free enzyme-biotin complex.

Structure of fatty acyl synthase
FATTY ACID SYNTHETASE
The second stage of fatty acid biosynthesis is also accomplished by a
multienzyme complex, fatty acid synthetase. The protein contains seven
distinct enzyme activities and an acyl-carrier protein, the latter
replacing CoA. It consists of a dimer of identical polypeptides
arranged head to tail. Each chain contains all seven enzyme activities
and an acyl-carrier protein; however, the structure-function relationships
are shared between the two polypeptide chains.

1. Priming of the cysteine (-Cys-SH) group with acetyl CoA.2. Malonyl CoA then combines with the –SH of the pantetheine
group that is attached to the acyl-carrier protein of the other subunit.3. 3-ketoacyl synthase catalyzes the reaction between the
previously attached acetyl group and the malonyl residue. 4. The 3-ketoacyl group subsequently undergoes sequential
reduction, dehydration, and reduction to form a saturated acyl enzyme complex
5. The next molecule of malonyl CoA then displaces the acyl group from the pantotheine-SH group to the now free cysteine group, and the reaction sequence is repeated through six cycles, after which, thioesterase is activated and the 16-carbon fatty acid, palmitate, is released.

Acetyl CoA carboxylase requires the presence of citrate or isocitrate,
which allows the polymerization that produces the active form of the
enzyme. This process is also inhibited by palmitoyl CoA at the same
allosteric site. Under conditions of high citrate concentration, energy storage
is desirable, but decreased synthesis of fatty acids is appropriate when the
product (palmitoyl CoA) accumulates. An additional control mechanism
involving hormone-dependent protein phosphatase/kinase:
phosphorylation, which inhibits activity of the enzyme, and
dephosphorylation, which activates it, is promoted by glucagon.
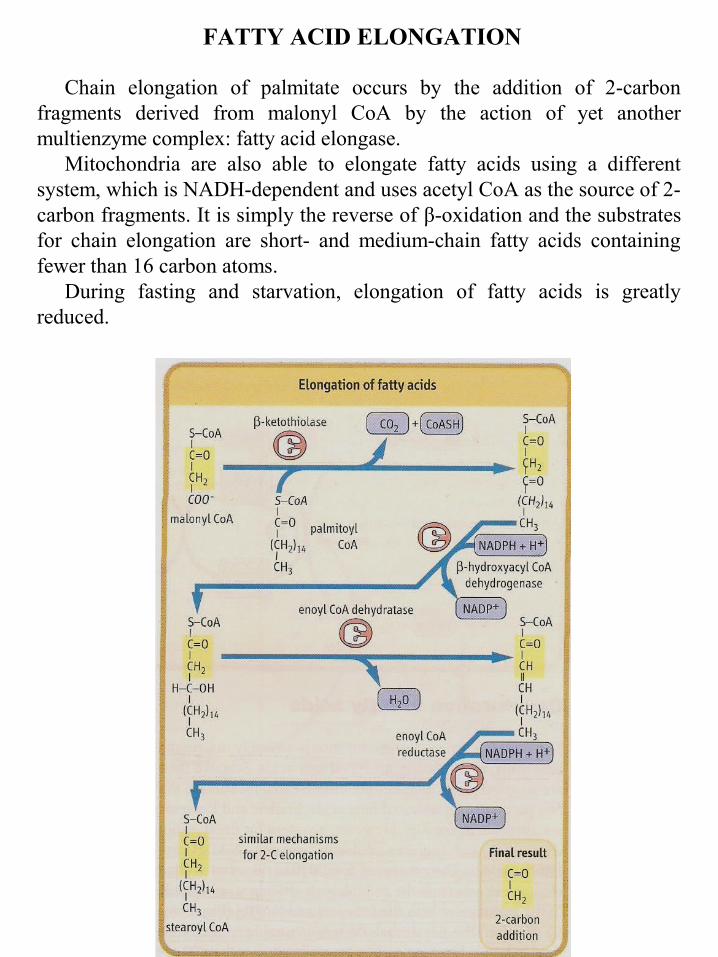
FATTY ACID ELONGATION
Chain elongation of palmitate occurs by the addition of 2-carbon fragments derived from malonyl CoA by the action of yet another multienzyme complex: fatty acid elongase.
Mitochondria are also able to elongate fatty acids using a different system, which is NADH-dependent and uses acetyl CoA as the source of 2-carbon fragments. It is simply the reverse of β-oxidation and the substrates for chain elongation are short- and medium-chain fatty acids containing fewer than 16 carbon atoms.
During fasting and starvation, elongation of fatty acids is greatly reduced.

DESATURATION OF FATTY ACIDS
The body has a requirement for mono- and polyunsaturated fatty
acids, in addition to saturated fatty acids. Some of these are supplied in
the diet. However, a system for desaturation does exist, and requires
molecular oxygen, NADH, and cytochrome b5. The process of
desaturation, like that of chain elongation, occurs on the endoplasmic
reticulum, and results in the oxidation of both the fatty acid and NADH.
In man, the desaturase system is unable to introduce double bonds
between carbon atoms beyond carbon 10 and the ω (terminal methyl)
carbon atom. The most common desaturation reaction to occur are
between carbon atoms 9 and 10.

KETOGENSIS FROM ACETYL COA
What does the liver do with the excess acetyl CoA that accumulates in fasting or starvation?
The problem of dealing with excess acetyl CoA is a critical one, because CoA is present only in catalytic amounts in tissues, and free CoA is required to initiate and continue the cycle of β-oxidation. To recycle the acetyl CoA, the liver uses a pathway known as ketogenesis, in which the CoA component is regenerated and the acetate group appears in blood in the form of three water-soluble lipid-derived products: acetoacetate, β-hydroxybutyrate, and acetone. The pathway of formation involves the synthesis and decomposition of hydroxymethylglutaryl (HMG) CoA in the mitochondrion. The liver is unique in its content of HMG CoA synthase and lyase, but is deficient in enzymes required for metabolism of ketone bodies, which explains their export from the liver.

SYNTHESIS OF KETONE BODIES(Acetoacetate, β-hydroxybutyrate, acetone)
KB synthesis when high level of FA in the blood (fasting,
starvation, high fat diet). Located in liver mitochondria.
1. High FA level → FA enter liver cells → β-oxidation generates
acetyl CoA, NADH, ATP → glucagon/insulin ratio is high →
synthesis of glucose (gluconeogenesis), conversion of pyruvate to
oxaloacetate → oxaloacetate + NADH = malate
2. Acetyl CoA + acetyl CoA → thiolase → acetoacetyl CoA +
acetyl CoA → HMG-CoA synthetase → 3-hydroxy-3-methylglutaryl
CoA (HMG-CoA) → HMG-CoA lyase → acetyl CoA + acetoacetate
a) directly enter the blood → to target tissue
b) NAD-dependent dehydrogenase → -hydroxybutyrate → to the
blood → to target tissue
c) spontaneous decarboxylation → acetone

OXIDATION OF KETONE BODIES
More energy is produced from β-hydroxybutyrate than from
acetoacetate
β-HB → β-HB denydrogenase → AA + NADH (ATP by OP)
AA → succinyl CoA : acetoacetate CoA transferase → acetoacetyl
CoA + succinate → β-ketothiolase → 2 acetyl CoA → TCA cycle
(23ATP)
FA from adipose TAG are the major fuel during fasting.
Some tissues are completely oxidized FA to CO2 and H2O.
Liver converts acetyl CoA to KB which are shipped to other tissues
by blood.
Advantages of ketone bodies
a) liver derives energy by partially oxidizing FA and forming KB
b) other tissues use KB as fuel
c) during starvation the brain can oxidize KB, reducing its need
for glucose.

Plasma concentrations of fatty acids and ketone bodies
Substrate Plasma concentration (mmol/L)
Normal Fasting Starvation
fatty acids
acetoacetate
β-hydroxybutyrate
0.6
0.1
0.1
1.0
0.2
1
1.5
1-2
5-10
Ketone bodies are taken up in extrahepatic tissues, including skeletal and heart muscle, where they are converted to CoA derivatives for metabolism. Ketone bodies are an efficient source of energy during fasting and starvation, and appear to be used in muscle in proportion to their plasma concentration. During starvation, the brain also converts to the use of ketone bodies for more than 50% of its energy metabolism, sparing glucose and reducing the demand on degradation of muscle protein for gluconeogenesis.

METABOLISM OF KETONE BODIES
E
CoA
OH O
hydroxybutyrate CH3 – CH – CH2 – C – O–
β-hydroxybutyratedehydrogenase
O O
acetoacetate CH3 – C – CH2 – C – O–
E succinyl CoA – acetoacetateCoA transferase
succinyl CoA
succinate
O O
acetoacetyl CoA CH3 – C – CH2 – C – S – CoA
E thiolase
2 x acetyl CoA
TCA cycle
Plasma concentrations of fatty acids and ketone bodies
Substrate Plasma concentration (mmol/L)
Normal Fasting Starvation
fatty acids 0.6 1.0 1.5
acetoacetate < 0.1 0.2 1-2
β-hydroxybutyrate
< 0.1 1 5-10

Top Related