







Languages
Pages
Legal


Biochemistry 3300 Slide 1
III. MetabolismThe Citric Acid Cycle
Department of Chemistry and BiochemistryUniversity of Lethbridge
Biochemistry 3300
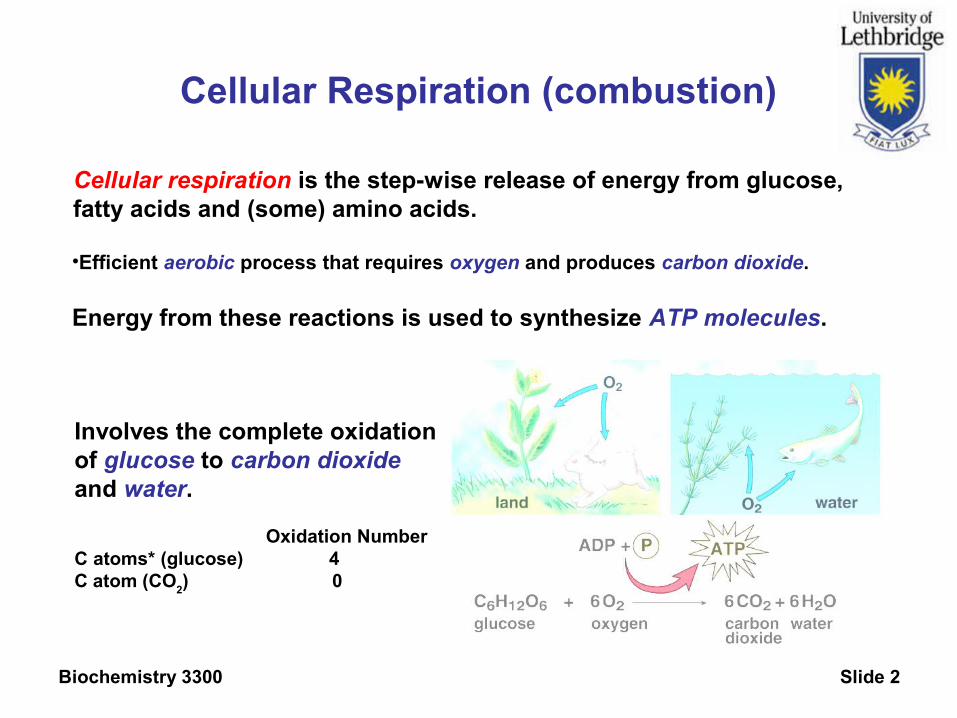
Biochemistry 3300 Slide 2
Cellular Respiration (combustion)
Cellular respiration is the step-wise release of energy from glucose,fatty acids and (some) amino acids.
•Efficient aerobic process that requires oxygen and produces carbon dioxide.
Energy from these reactions is used to synthesize ATP molecules.
Involves the complete oxidationof glucose to carbon dioxide and water.
Oxidation NumberC atoms* (glucose) 4C atom (CO
2) 0
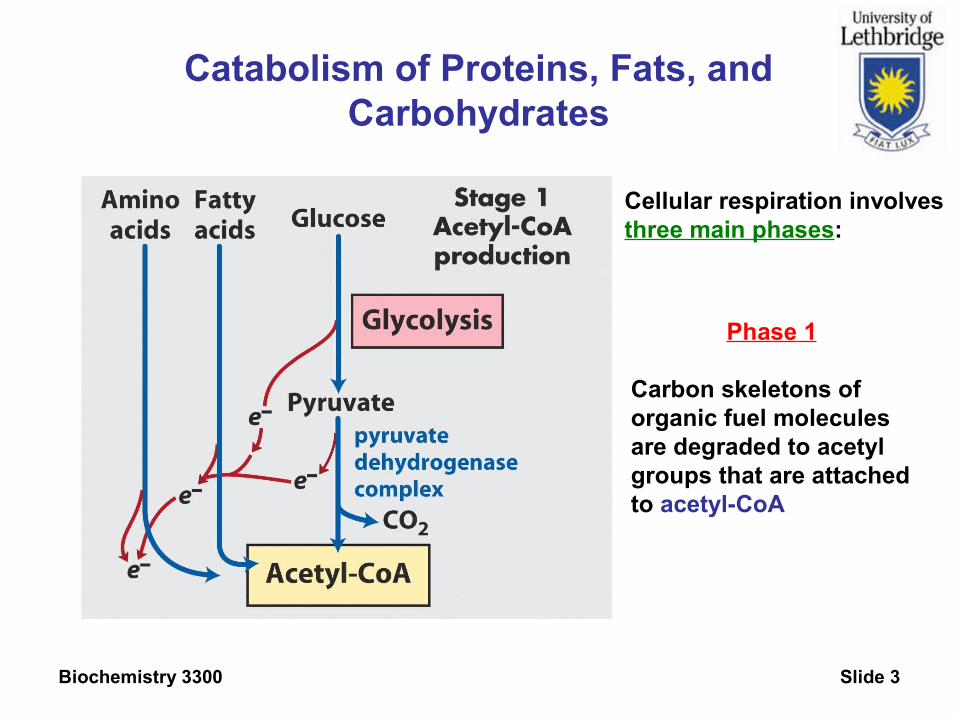
Biochemistry 3300 Slide 3
Catabolism of Proteins, Fats, and Carbohydrates
Phase 1
Carbon skeletons oforganic fuel moleculesare degraded to acetyl groups that are attachedto acetyl-CoA
Cellular respiration involvesthree main phases:
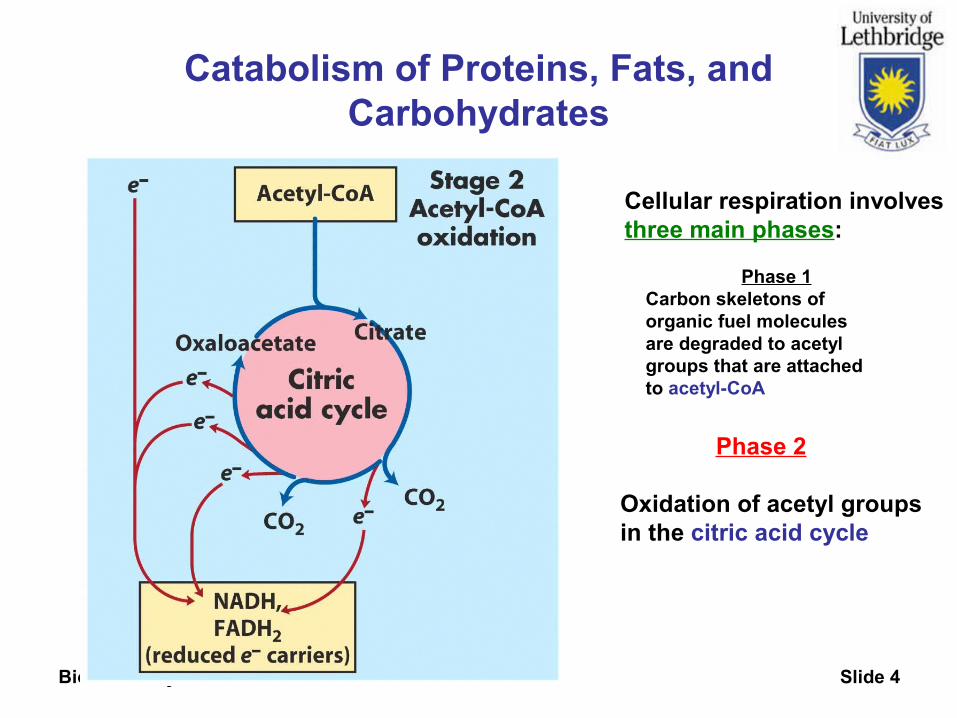
Biochemistry 3300 Slide 4
Catabolism of Proteins, Fats, and Carbohydrates
Phase 2
Oxidation of acetyl groupsin the citric acid cycle
Cellular respiration involvesthree main phases:
Phase 1Carbon skeletons oforganic fuel moleculesare degraded to acetyl groups that are attachedto acetyl-CoA
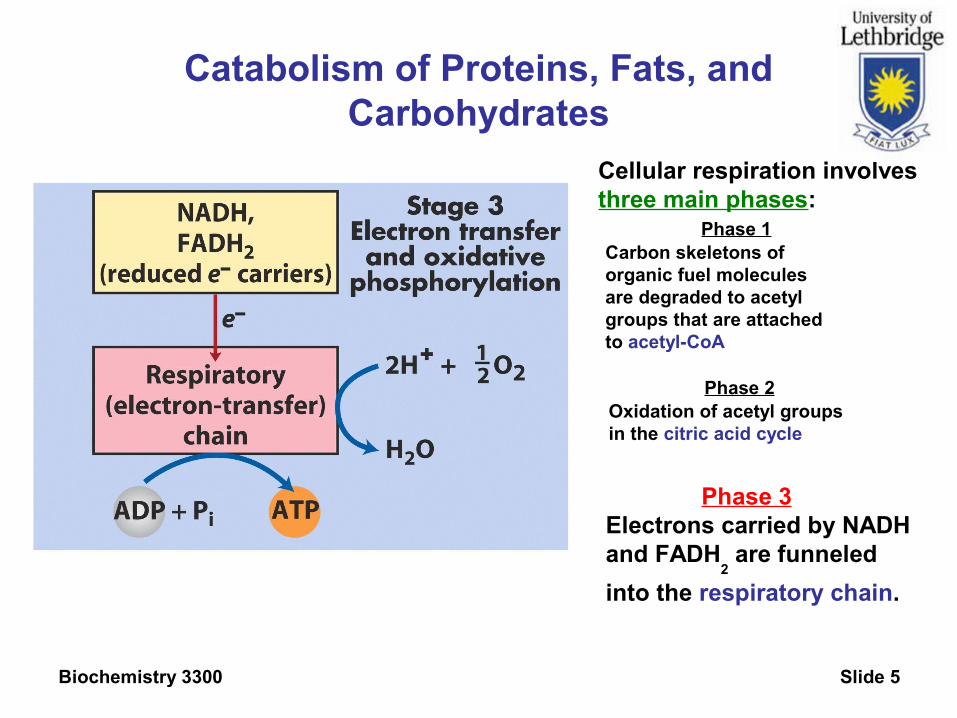
Biochemistry 3300 Slide 5
Catabolism of Proteins, Fats, and Carbohydrates
Phase 3Electrons carried by NADHand FADH
2 are funneled
into the respiratory chain.
Phase 2Oxidation of acetyl groupsin the citric acid cycle
Phase 1Carbon skeletons oforganic fuel moleculesare degraded to acetyl groups that are attachedto acetyl-CoA
Cellular respiration involvesthree main phases:
Biochemistry 3300 Slide 6
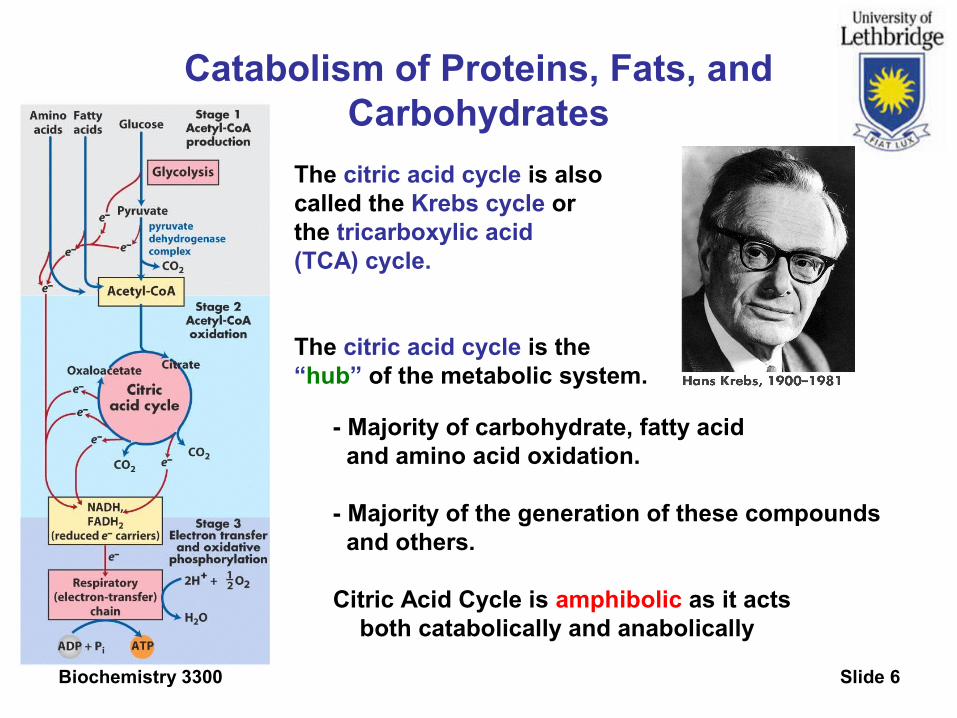
Catabolism of Proteins, Fats, and Carbohydrates
- Majority of carbohydrate, fatty acid and amino acid oxidation.
- Majority of the generation of these compounds and others.
Citric Acid Cycle is amphibolic as it acts both catabolically and anabolically
The citric acid cycle is alsocalled the Krebs cycle or the tricarboxylic acid (TCA) cycle.
The citric acid cycle is the“hub” of the metabolic system.

Biochemistry 3300 Slide 7
HistoryBy 1930, it was established that some compounds:
Carboxylic acids (acetate, lactate)Dicarboxylic acids (succinate, malate, -ketoglutarate) andTricarboxylic acids (citrate, isocitrate)
would stimulate O2 consumption and CO
2 production when added to
“minced” muscle
1935: Albert Szent-Gyorgyi Succinate → Fumarate → Malate → Oxaloacetate
Carl Martius & Franz KnoopCitrate [→ Cis-aconitate] → Isocitrate → -ketoglutarate →Succinate → Fumarate → Malate → Oxaloacetate

Biochemistry 3300 Slide 8
Subsequently, Martius & Knoop showed:
Pyruvate and Oxaloacetate can form citrate non-enzymatically (requires peroxide and basic conditions).
Odd: oxaloacetate is the product in their pathway!
And then Hans Krebs the showed:
Succinate is formed from fumarate, malate or oxaloacetate.
Odd: These appear to be the reverse reactions!
Citric Acid metabolic pathway is a CYCLE !!
History
Biochemistry 3300 Slide 9
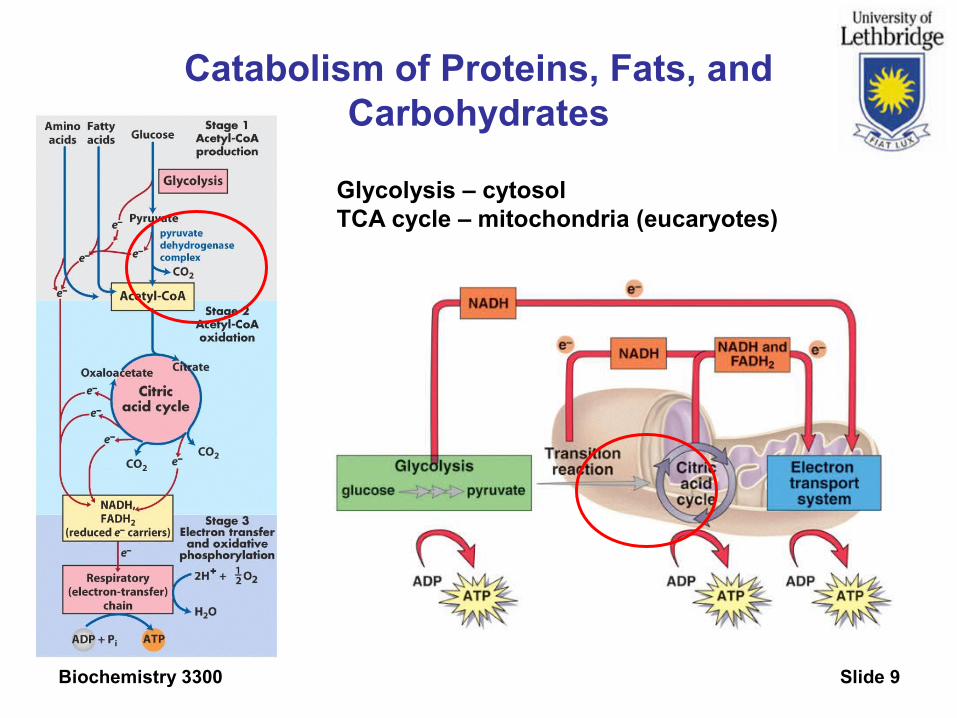
Catabolism of Proteins, Fats, and Carbohydrates
Glycolysis – cytosolTCA cycle – mitochondria (eucaryotes)

Biochemistry 3300 Slide 10
Citric Acid Cycle enzymes are in the mitochondrial matrix
Substrates must cross both the outer and inner mitochondrial membrane
Biochemistry 3300 Slide 11
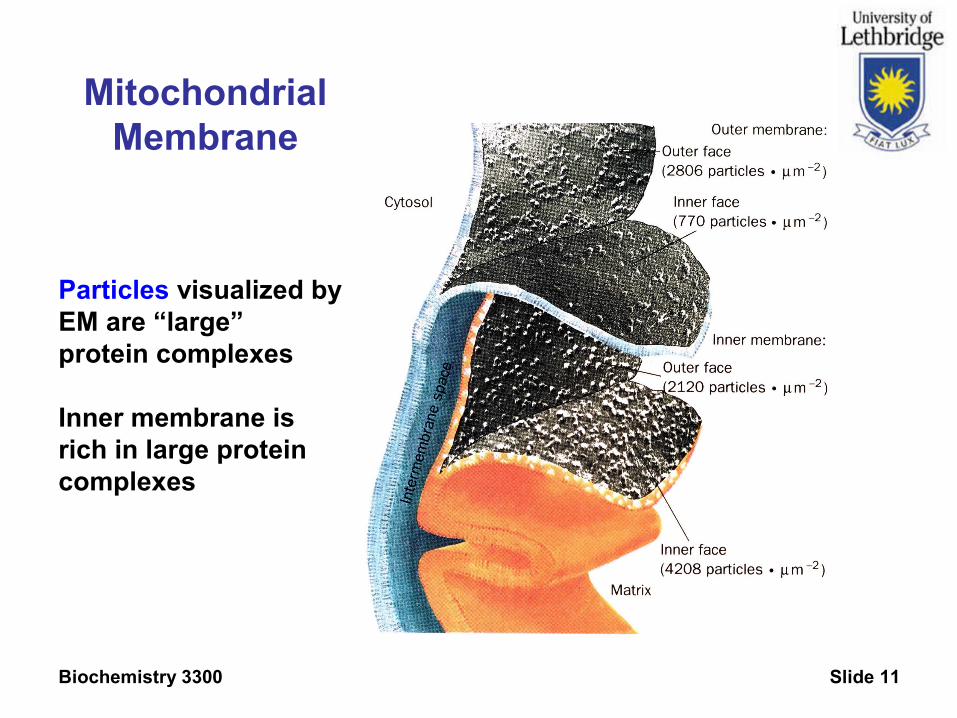
Mitochondrial Membrane
Particles visualized byEM are “large” protein complexes
Inner membrane isrich in large protein complexes
Biochemistry 3300 Slide 12
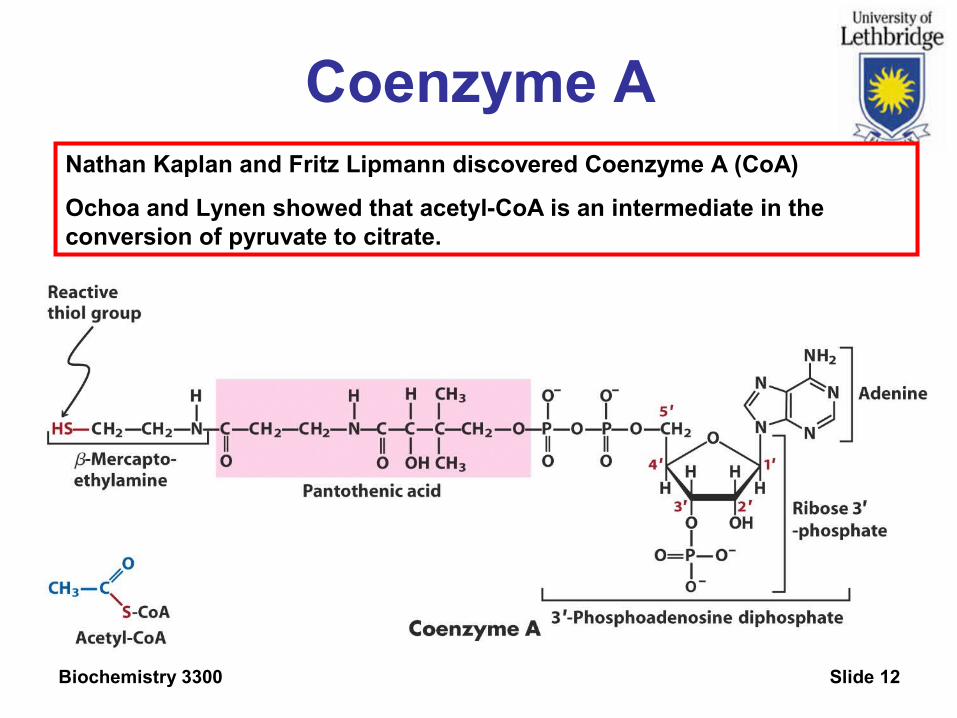
Nathan Kaplan and Fritz Lipmann discovered Coenzyme A (CoA)
Ochoa and Lynen showed that acetyl-CoA is an intermediate in the conversion of pyruvate to citrate.
Coenzyme A
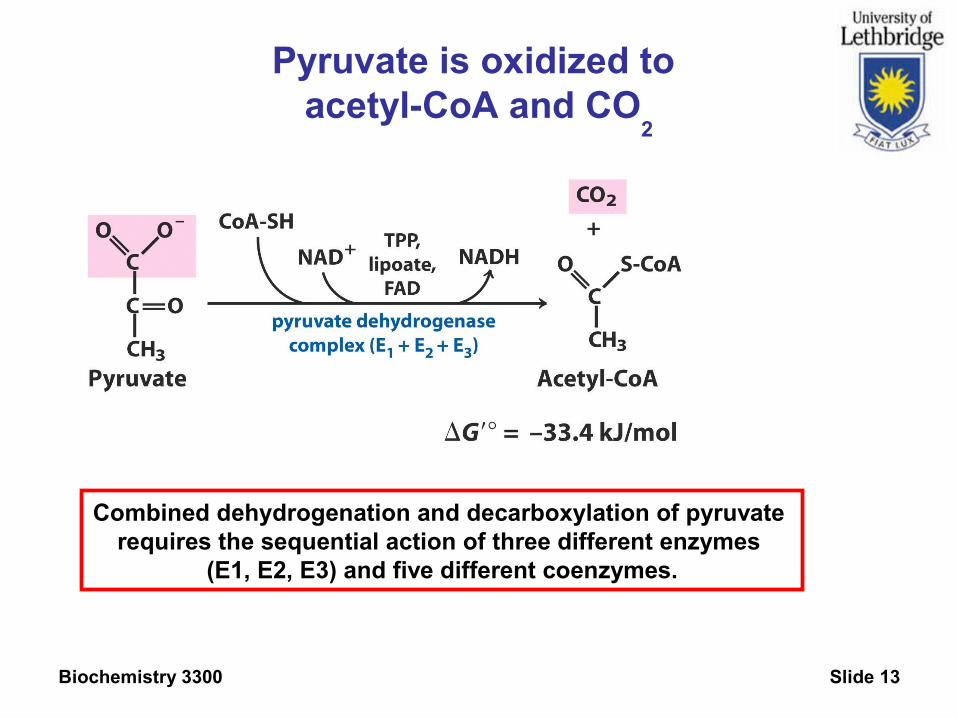
Biochemistry 3300 Slide 13
Pyruvate is oxidized to acetyl-CoA and CO
2
Combined dehydrogenation and decarboxylation of pyruvate requires the sequential action of three different enzymes
(E1, E2, E3) and five different coenzymes.

Biochemistry 3300 Slide 14
Pyruvate Dehydrogenase Complex (PDH)
E. coli yeast
Pyruvate dehydrogenase -- E1 24 60
Dihydrolipoyl transacetylase -- E2 24 60
Dihydrolipoyl dehydrogenase -- E3 12 12
Molecular weight of 4,600,000 Da ; 50 nm in diameter
PDH complex contains three subunits, present in multiplecopies. Number varies among species.
Lipoate is connected to E2
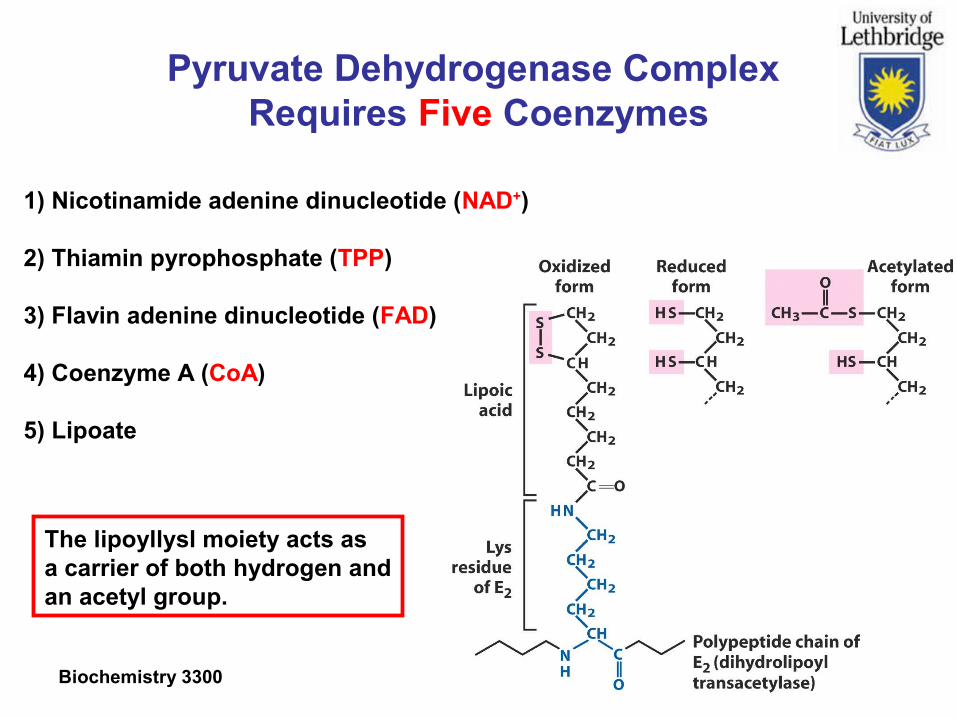
Biochemistry 3300 Slide 15
Pyruvate Dehydrogenase Complex Requires Five Coenzymes
1) Nicotinamide adenine dinucleotide (NAD+)
2) Thiamin pyrophosphate (TPP)
3) Flavin adenine dinucleotide (FAD)
4) Coenzyme A (CoA)
5) Lipoate
The lipoyllysl moiety acts as a carrier of both hydrogen andan acetyl group.
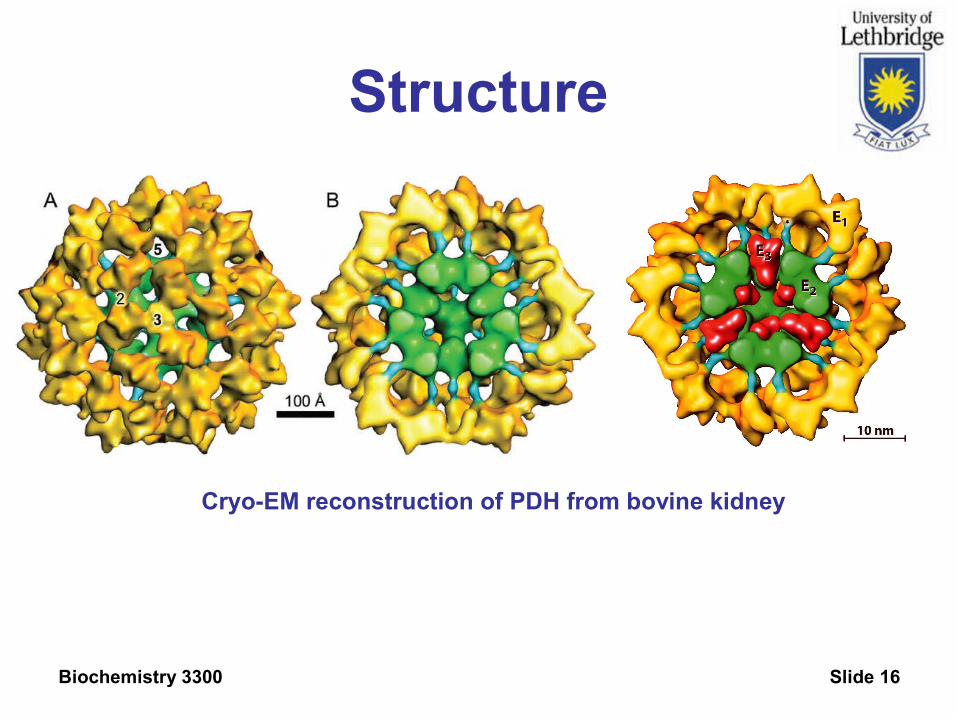
Biochemistry 3300 Slide 16
Structure
Cryo-EM reconstruction of PDH from bovine kidney
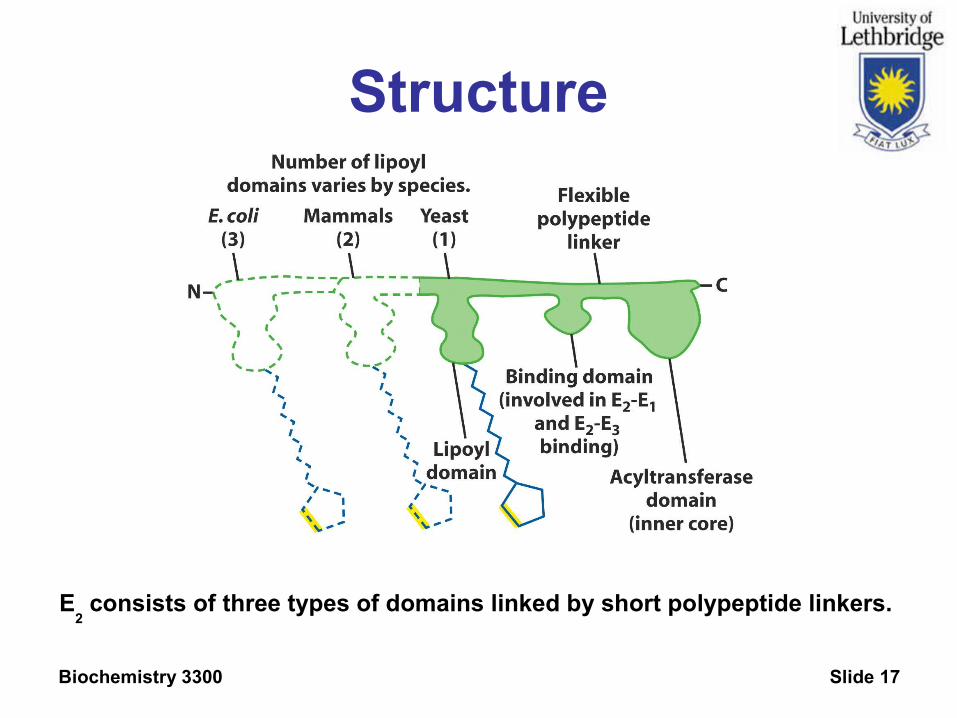
Biochemistry 3300 Slide 17
Structure
E2 consists of three types of domains linked by short polypeptide linkers.
Biochemistry 3300 Slide 18
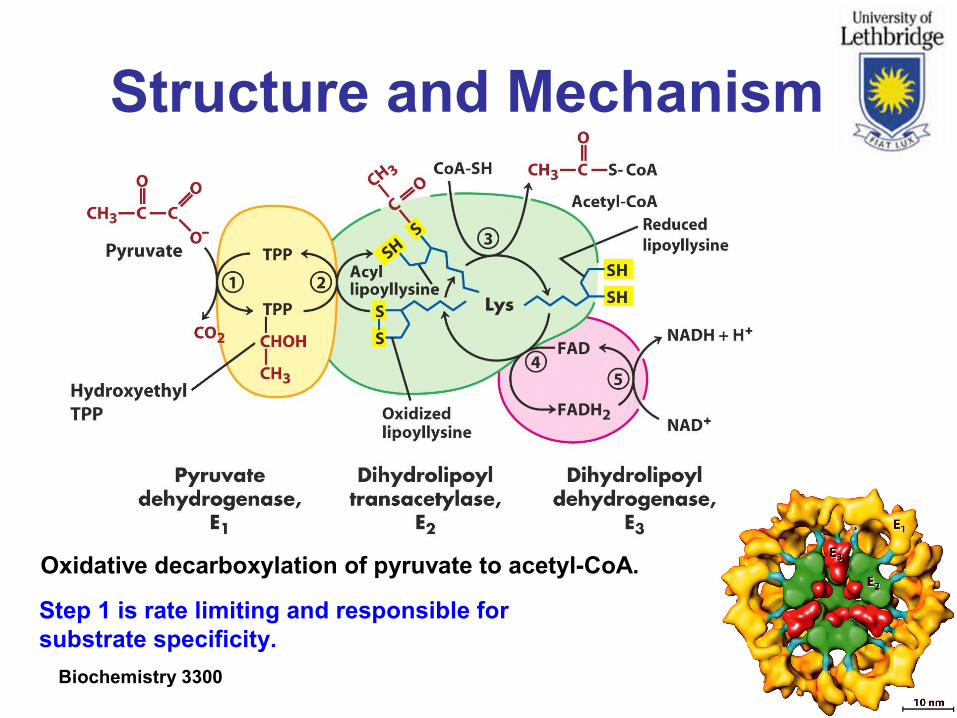
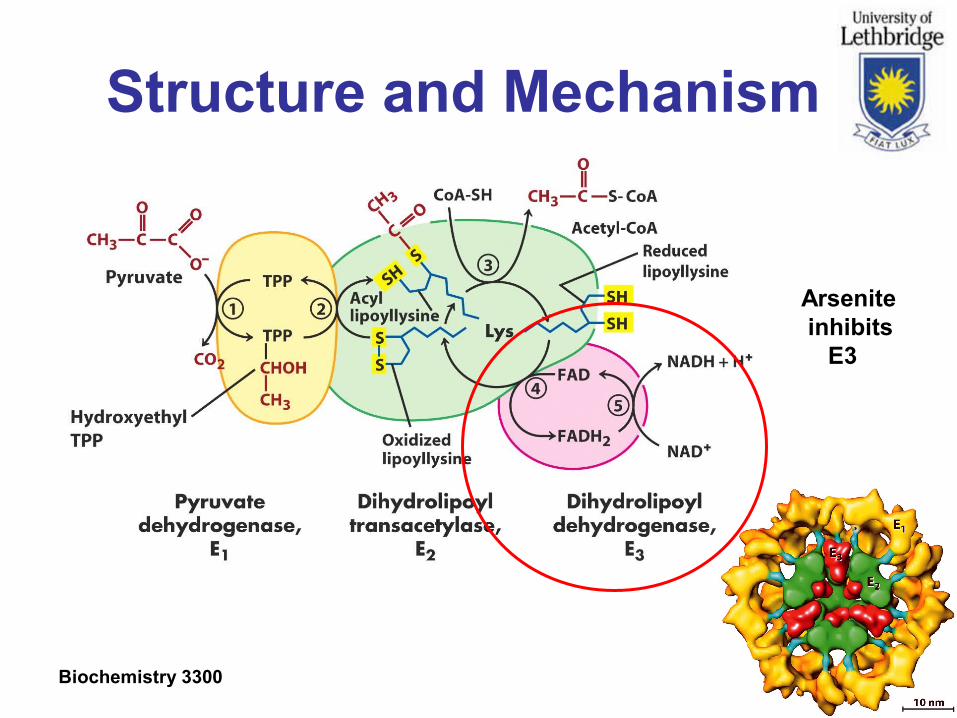
Structure and Mechanism
Oxidative decarboxylation of pyruvate to acetyl-CoA.
Step 1 is rate limiting and responsible for substrate specificity.
Biochemistry 3300 Slide 19
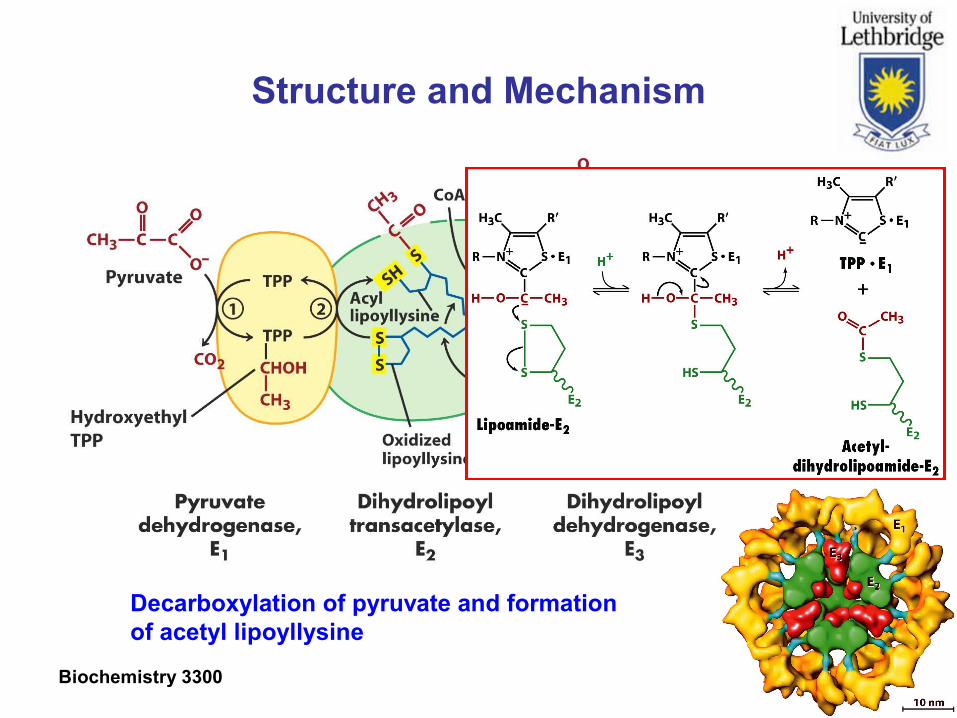
Structure and Mechanism
Decarboxylation of pyruvate and formationof acetyl lipoyllysine
Biochemistry 3300 Slide 20
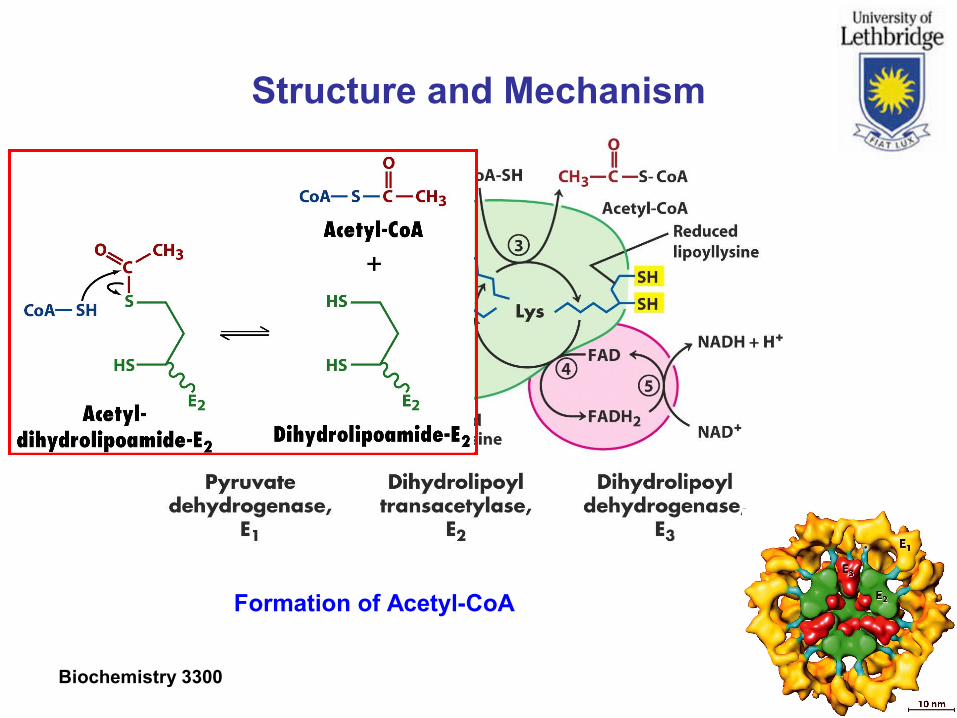
Structure and Mechanism
Formation of Acetyl-CoA

Biochemistry 3300 Slide 21
Why such a complex set of enzymes?
1. Enzymatic reaction rates are limited by diffusion, with shorter distance between subunits in an enzyme, the substrate can be directed from one subunit (catalytic site) to another.
2. Channeling metabolic intermediates between successive enzymes minimizes side reactions. (Substrate channeling).
3. Local substrate concentration is kept high.
4. The reactions of a multienzyme complex can be coordinately controlled / regulated.
Biochemistry 3300 Slide 22
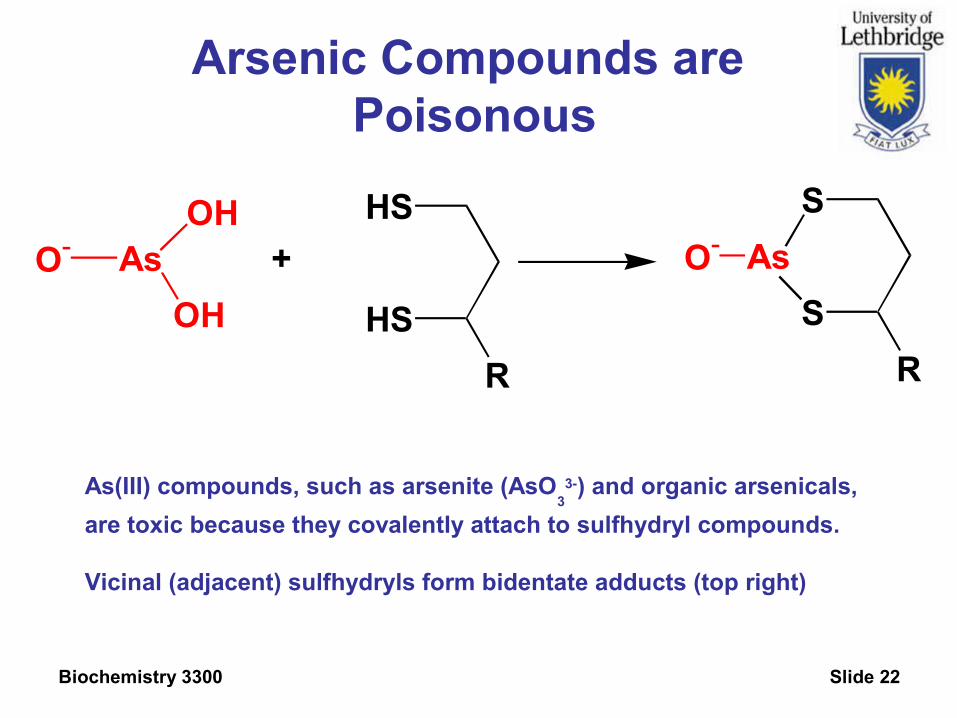
Arsenic Compounds are Poisonous
HS
HS
R
O- AsOH
OH
+
S
S
R
AsO-
As(III) compounds, such as arsenite (AsO3
3-) and organic arsenicals,
are toxic because they covalently attach to sulfhydryl compounds.
Vicinal (adjacent) sulfhydryls form bidentate adducts (top right)
Biochemistry 3300 Slide 23
Structure and Mechanism
Arsenite inhibits E3

Biochemistry 3300 Slide 24
Mechanism of Dihydrolipoyl Dehydrogenase.
More complicated than expected:
1. Spectra of oxidized dihydrolipoamide dehydrogenase (E3) isunaffected by arsenite.
2. NADH reaction with the oxidized enzyme in the presence ofarsenite → forms an enzymatically inactive species.
3. Spectrum of the arsenite-inactivated enzyme (2.) indicates that itsFAD prosthetic group is fully oxidized.
Recall: The oxidation state of the flavin in a flavoprotein is readily established from its characteristic UV-Vis Spectrum: FAD is intense yellow, whereas FADH
2 is pale yellow.
Explanation ?

Biochemistry 3300 Slide 25
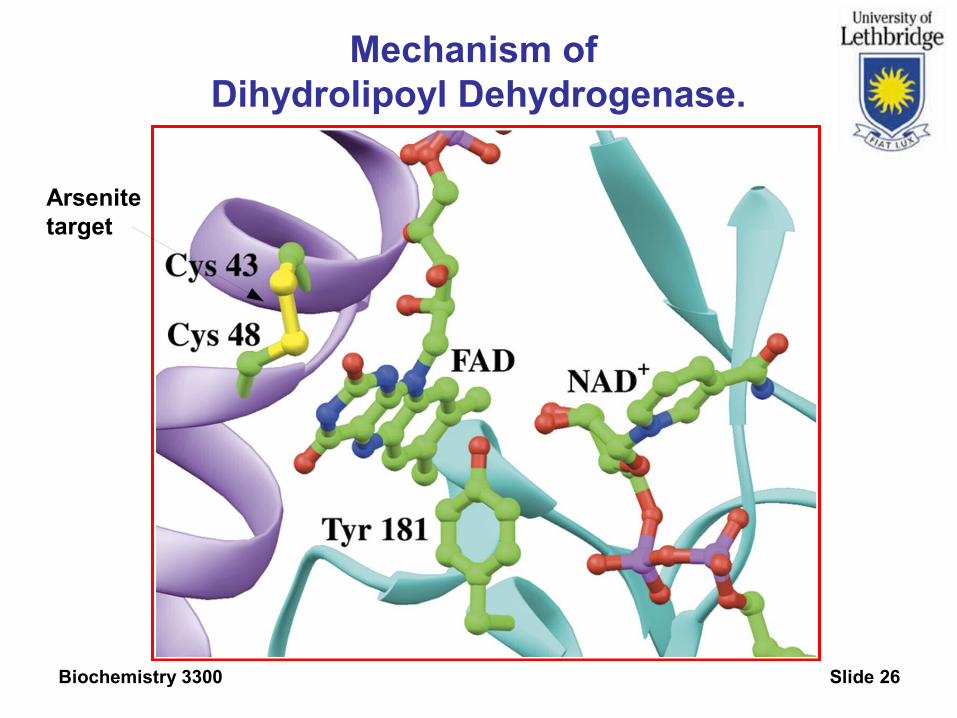
Mechanism of Dihydrolipoyl Dehydrogenase.
Oxidized dehydrolipoamide dehydrogenase has an additional electron acceptor.
Arsenite inhibition suggests a disulfide as acceptor.
See X-Ray structure of dehydrolipoamide DH from P. putida, PDBID 1LVL
Catalytic active residues: Cys 43 & 48 , Tyr 181
Biochemistry 3300 Slide 26
Mechanism of Dihydrolipoyl Dehydrogenase.
Arsenitetarget
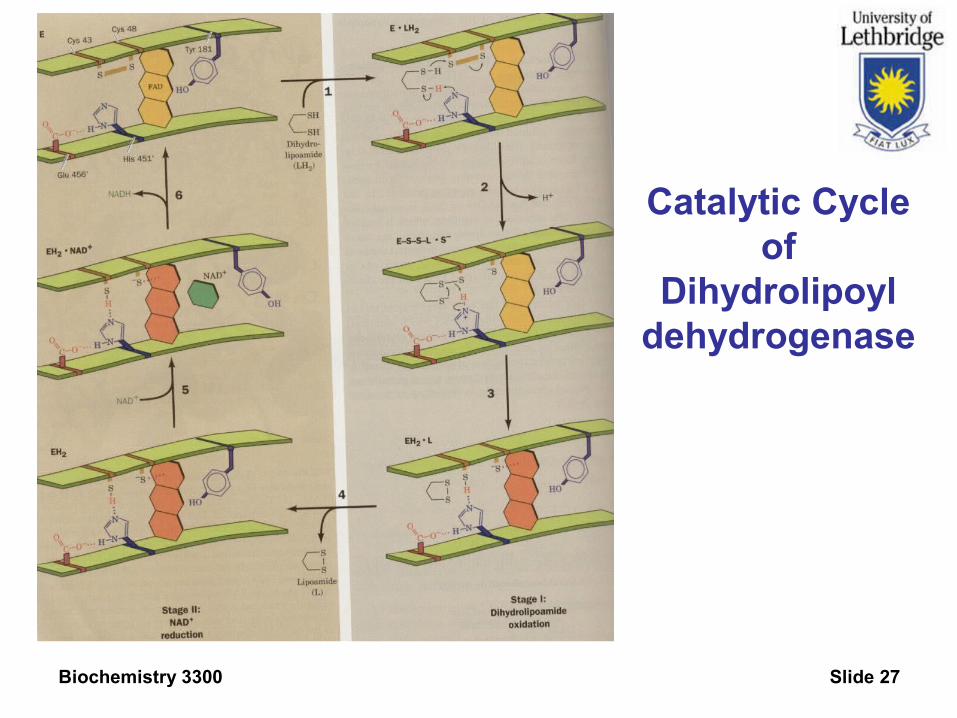
Biochemistry 3300 Slide 27
Catalytic Cycle of
Dihydrolipoyldehydrogenase
Biochemistry 3300 Slide 28
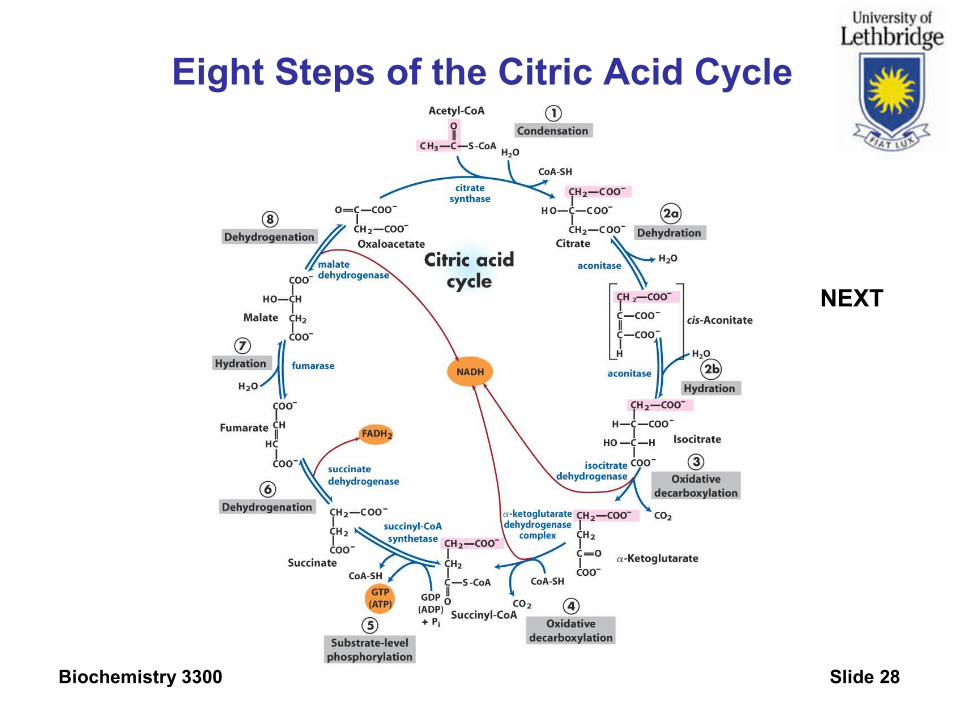
Eight Steps of the Citric Acid Cycle
NEXT
Top Related