

![[XLS]12864_2006_834_MOESM1_ESM.xls - Springer Static ...10.1186/1471... · Web viewdataset b0080, ECs0084, transcriptional repressor of fru operon and others; fruR [BEZ] mwgecov2#0077](https://static.fdocuments.in/doc/165x107/5af8e7ae7f8b9a2d5d8c397a/xls128642006834moesm1esmxls-springer-static-1011861471web-viewdataset.jpg)





Languages
Pages
Legal


Gene regulation IIBiochemistry 302
February 27, 2006

Molecular basis of inhibition of RNAP by Lac repressor
CRP/DNA complex
−35 promoter site
−10 promoter site
Lewis, M. et al. (1996) Science 271:1247
−60
Lehninger Principles of Biochemistry, 4th ed., Ch 28
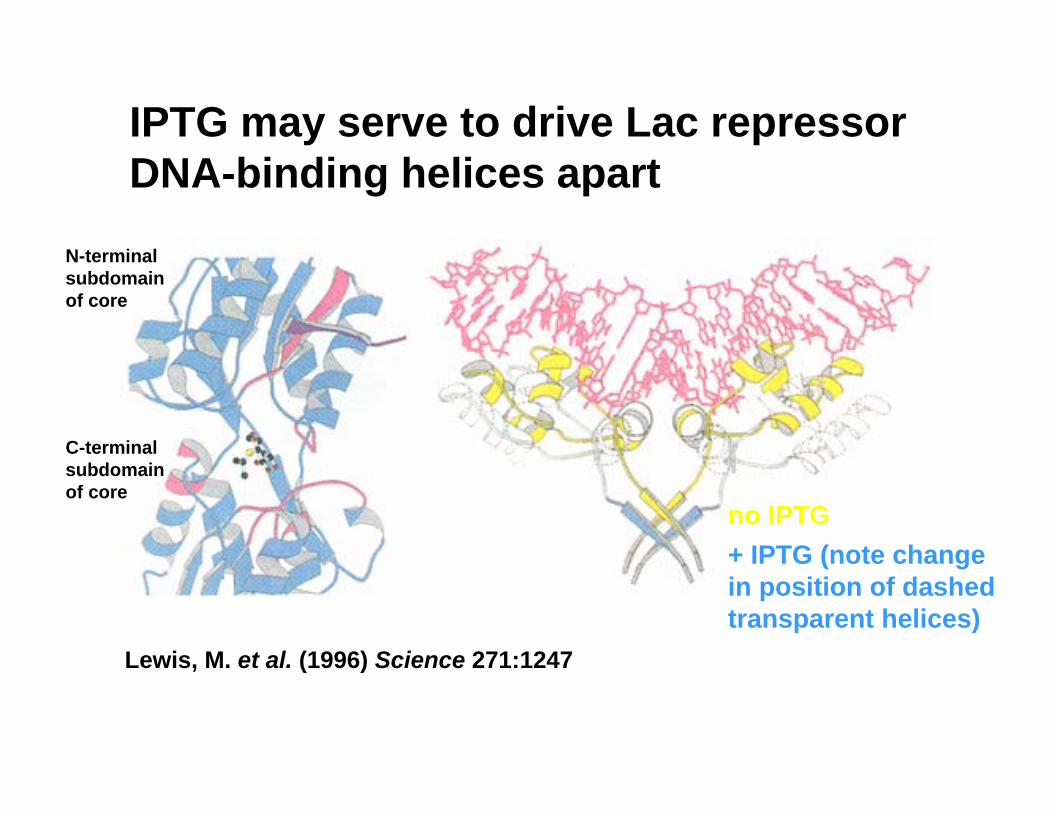
IPTG may serve to drive Lac repressor DNA-binding helices apart
Lewis, M. et al. (1996) Science 271:1247
no IPTG+ IPTG (note change in position of dashed transparent helices)
N-terminal subdomainof core
C-terminal subdomainof core

Hinge helix region: an allosteric switch
• no IPTG bound– Packed α-helical “hinge”– α-helices stabilized by DNA
(operator) and core contacts• w/ IPTG bound
– Rotation and translation of N-term subdomains
– Destabilization of the hinge helices
– Unpacking of hinge helix →headpiece “unfolding”
– Loss of specific DNA contacts: concerted folding and binding in reverse
– Repressor now has same low affinity for operator and non-operator sequences
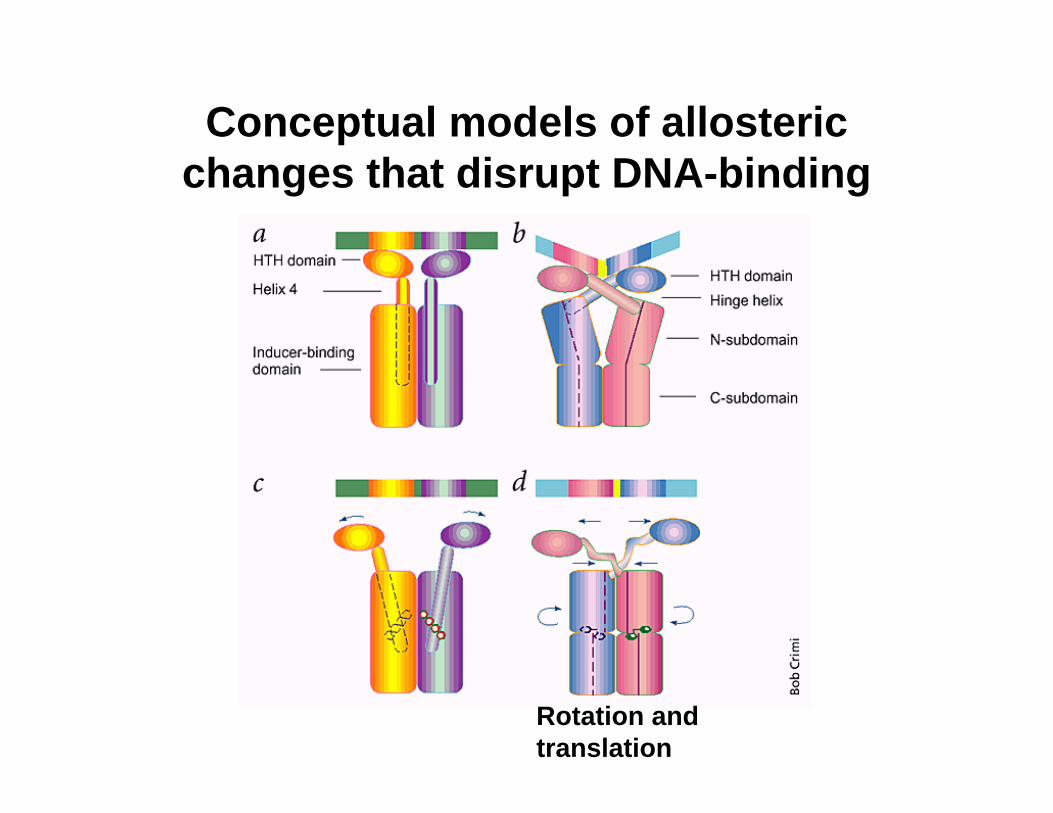
Conceptual models of allosteric changes that disrupt DNA-binding
Rotation and translation

Functional groups in DNA that facilitate protein (TF) binding
Groups used for base-pair recognition are shown in red.
Lehninger Principles of Biochemistry, 4th ed., Ch 28
3 2 3223
2 3

Amino acid-base pair interactions often observed in DNA-protein binding• Amino acid side chains
most often H-bonded to bases in DNA (Asn, Gln, Glu, Asp, Lys, Arg)
• No simple amino acid base interaction code….
• ….but some restricted interactions– Asn or Gln with N6 and N7
of adenine – Arg with N7 and O6 of
guanine• DNA binding sequences
cannot be inferred from protein structure.
Lehninger Principles of Biochemistry, 4th ed., Ch 28
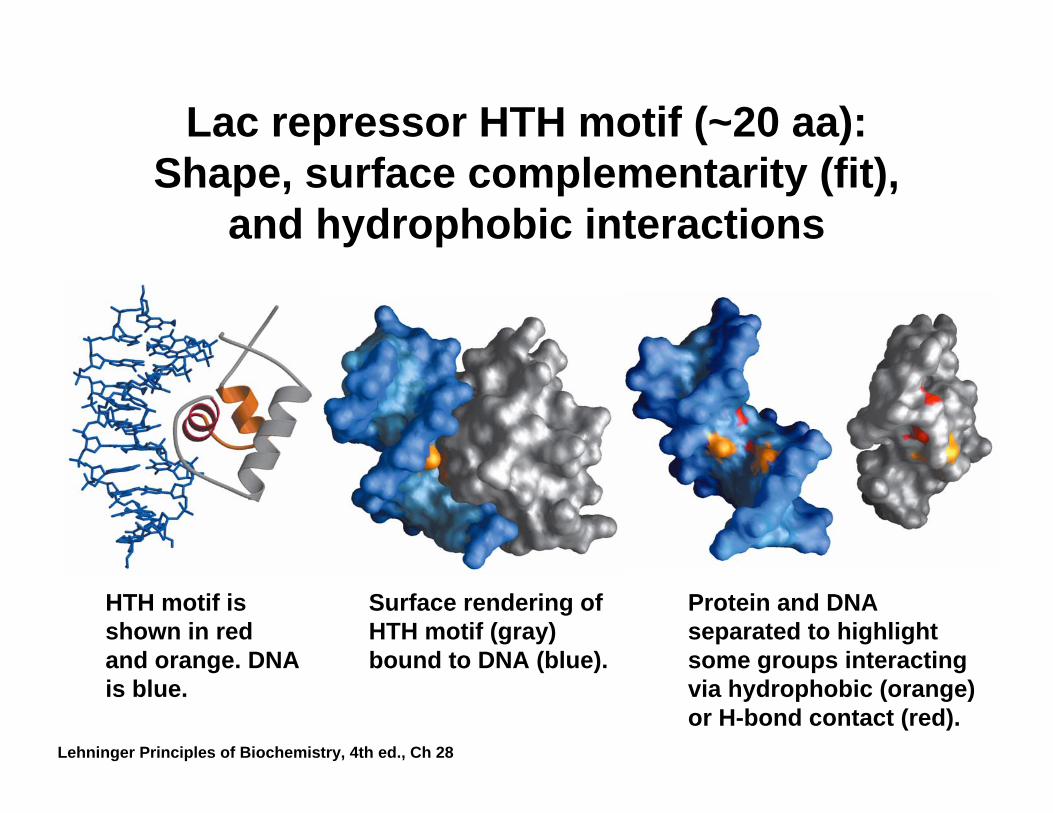
Lac repressor HTH motif (~20 aa): Shape, surface complementarity (fit),
and hydrophobic interactions
HTH motif is shown in red and orange. DNA is blue.
Surface rendering of HTH motif (gray) bound to DNA (blue).
Protein and DNA separated to highlight some groups interacting via hydrophobic (orange) or H-bond contact (red).
Lehninger Principles of Biochemistry, 4th ed., Ch 28

Catabolic operons: Regulation by multiple signals/ligands that target
different TFs
CRP homodimer (subunit Mr22,000) bound to DNA. cAMP“inducer” is in red. RNAP interaction domain is yellow.
Catabolite repression: Activity of lac operon is restricted when both glucose and lactose are present. E. coli would prefer to metabolize glucose directly (via glycolysis) rather than generating it from secondary sugars.
Lehninger Principles of Biochemistry, 4th ed., Ch 28

Other side of the coin: the biosynthetic trp operon• Amino acid biosynthesis consumes energy
– Advantageous to inhibit synthesis of biosynthetic enzymes when end product (amino acid) is available.
– Regulatory goal is to repress gene activity.• E. coli trp operon (in contrast to lac)
– Trp repressor is activated by ligand (Trp) binding.– Additional regulation by premature termination of
transcription (attenuation – regulatory dimmer switch involves ribosome positioning on 5′ mRNA)
• Discovered by Charles Yanofsky, common to many biosynthetic operons encoding enzymes needed for amino acid synthesis (including Trp, Leu, and His).
• Dictated by changes in RNA secondary structure• Extends the possible range of transcription rates
(moderate to high Trp levels)

Schematic of the E. coli trp operon (regulation by Trp-induced repression)
chorismic acid → Trp
Trp inducer
Dimeric HTH protein
aporepressor(when trp levels are low) Secondary mechanism of
repression: moderate to high Trp levelsFig. 26-33

Structure(s) of the trp operon mRNA leader (trpL) sequence (162 nt)
Does the 3:4 pair structure remind you of anything?
Lehninger Principles of Biochemistry, 4th ed., Ch 28
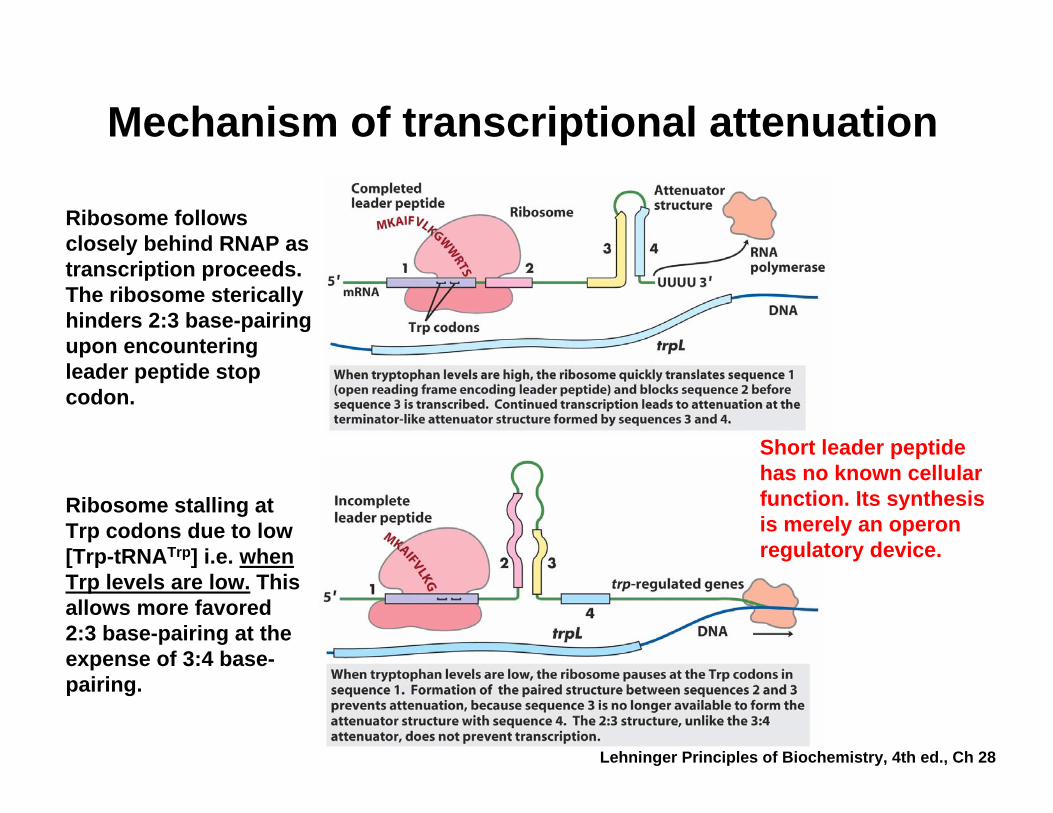
Mechanism of transcriptional attenuation
Ribosome stalling at Trp codons due to low [Trp-tRNATrp] i.e. when Trp levels are low. This allows more favored 2:3 base-pairing at the expense of 3:4 base-pairing.
Ribosome follows closely behind RNAP as transcription proceeds. The ribosome sterically hinders 2:3 base-pairing upon encountering leader peptide stop codon.
Short leader peptide has no known cellular function. Its synthesis is merely an operon regulatory device.
Lehninger Principles of Biochemistry, 4th ed., Ch 28
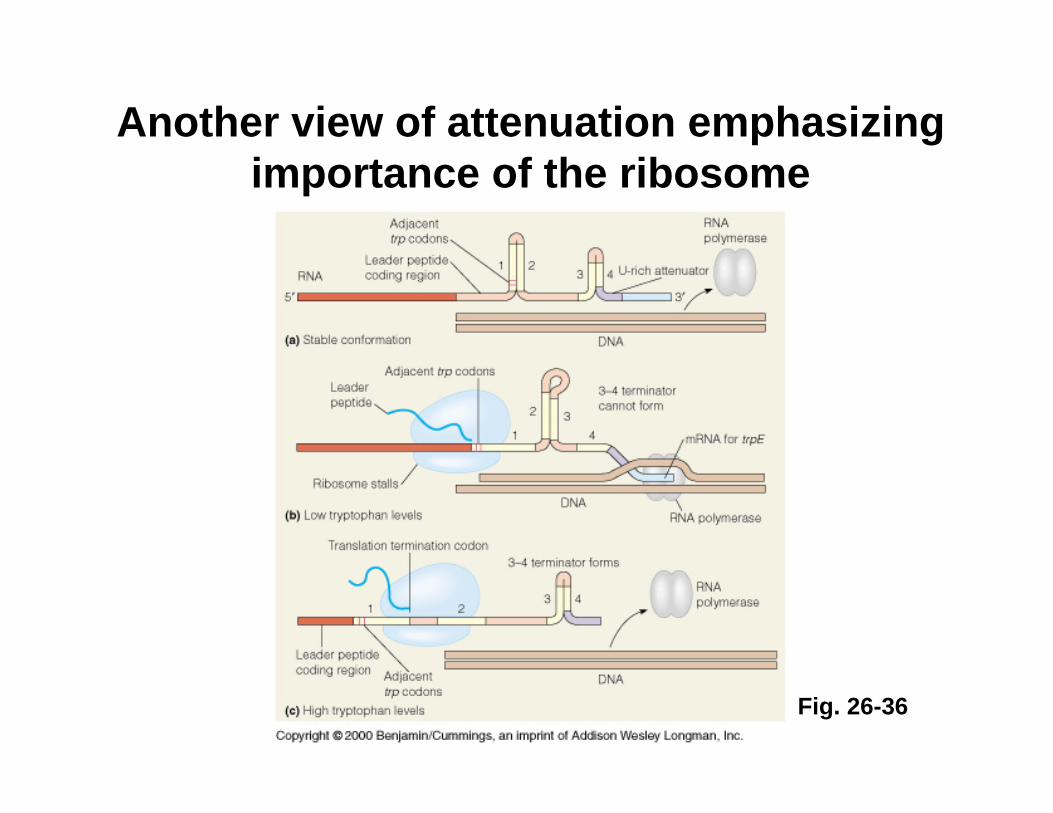
Another view of attenuation emphasizing importance of the ribosome
Fig. 26-36
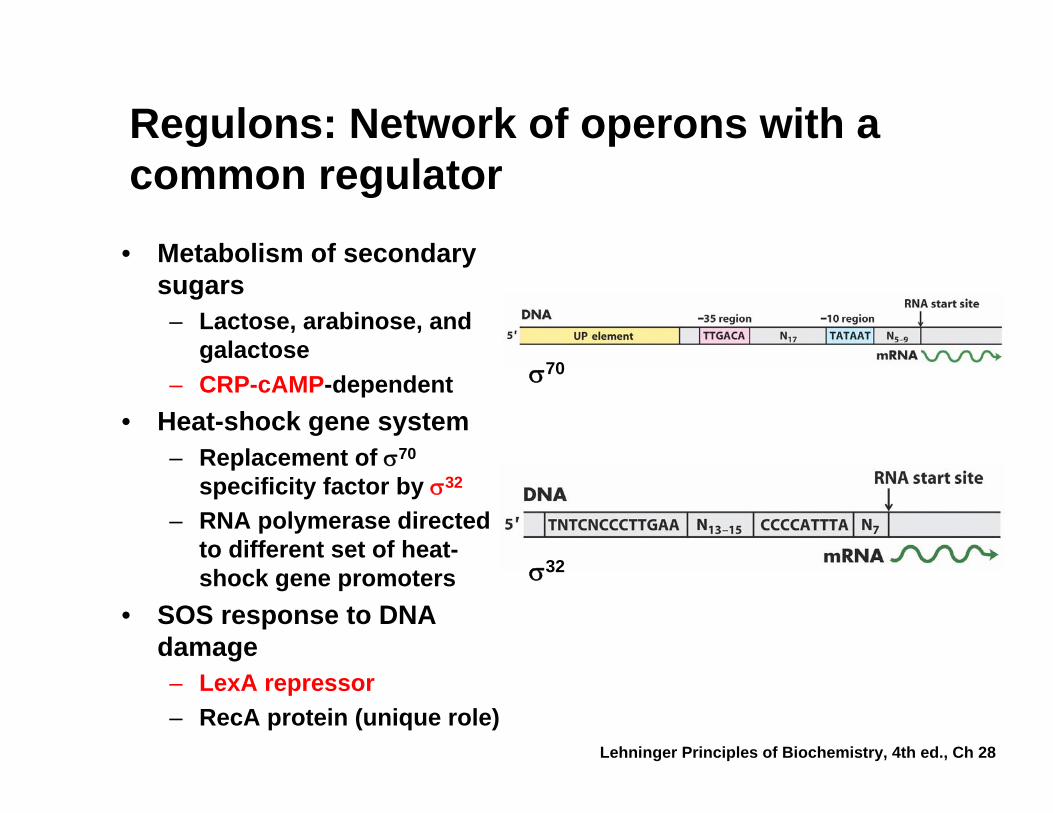
Regulons: Network of operons with a common regulator
• Metabolism of secondary sugars– Lactose, arabinose, and
galactose– CRP-cAMP-dependent
• Heat-shock gene system– Replacement of σ70
specificity factor by σ32
– RNA polymerase directed to different set of heat-shock gene promoters
• SOS response to DNA damage– LexA repressor– RecA protein (unique role)
σ70
σ32
Lehninger Principles of Biochemistry, 4th ed., Ch 28

Induction of SOS response in E. coli(LexA-dependent regulon)
• Cellular response to extensive DNA damage
• Induced genes mostly involved in DNA repair
• Mechanism: Proteolytic inactivation of LexArepressor– RecA/ssDNA-dependent– Interaction of ssDNA-bound
RecA stimulates intrinsic protease activity of LexA.
– LexA inactivates itself by catalyzing its own cleavage at a specific Arg-Gly bond in the middle of the protein.
Lehninger Principles of Biochemistry, 4th ed., Ch 28
Top Related