







Languages
Pages
Legal


Endogenous opioid-mediated antinociception in cholestatic
mice is peripherally, not centrally, mediated
Lisa Nelson1, Nathalie Vergnolle2, Charlotte D’Mello1, Kevin Chapman2,
Tai Le1, Mark G. Swain1,*
1Liver Unit, Gastrointestinal Research Group, Health Sciences Center, University of Calgary, 3330 Hospital Dr., NW, Calgary, AB, Canada T2N 4N12Mucosal Inflammation Research Group, University of Calgary, Calgary, AB, Canada T2N 4N1
Background/Aims: Cholestasis is associated with naloxone reversible antinociception and opiate receptor antagonists
are used clinically to treat pruritus. Pain and pruritus are closely interrelated and opioids modulate both sensations.
Therefore, we undertook a series of experiments to characterize opioid-mediated antinociception in cholestasis and
determine if it occurs inside or outside the CNS.
Methods: Antinociception scores to both thermal and mechanical stimuli were determined in mice with cholestasis
due to bile duct resection vs sham controls.
Results: Cholestatic mice demonstrated significant antinociception to both stimuli compared to controls, which
was reversible by the opiate receptor antagonist naloxone. The experiments were repeated with a naloxone
derivative, which does not cross the blood–brain-barrier (i.e. naloxone methiodide) with similar results, indicating
an opioid antinociceptive effect mediated outside of the CNS. Experiments with intraplantar injections of low dose
naloxone methiodide confirmed that cholestasis-associated antinociception occurs at the level of cutaneous nerve
endings. These findings were supported by findings of increased dermal met-enkephalin expression in cholestatic
mice.
Conclusions: Cholestasis in mice is associated with antinociception due to local effects of endogenous opioids (i.e. met-
enkephalin) at the level of sensory nerve endings. These findings may have direct implications in the management of
cholestasis associated pruritus.
q 2005 European Association for the Study of the Liver. Published by Elsevier B.V. All rights reserved.
Keywords: Opioids; Met-enkephalin; Analgesia; Opioid receptor; Naloxone; C-fibers
1. Introduction
Pruritus is commonly encountered in patients with
cholestatic liver diseases and can be a profoundly bother-
some and disabling symptom [1,2]. However, the patho-
genesis of pruritus in cholestasis remains poorly understood.
Traditionally, cholestasis associated itch has been postu-
lated as resulting from the interaction of free unmyelinated
cutaneous nerve endings with a substance(s) retained in the
circulation as part of the cholestatic syndrome. The likely
0168-8278/$32.00 q 2005 European Association for the Study of the Liver. Pub
doi:10.1016/j.jhep.2005.11.043
Received 22 June 2005; received in revised form 17 October 2005;
accepted 7 November 2005; available online 27 December 2005* Corresponding author. Tel.: C1 403 220 8457; fax: C1 403 270 0995.
E-mail address: [email protected] (M.G. Swain).
unmyelinated neuron involved in this process is the recently
defined C-fiber, which carries itch related nerve impulses
from the skin to the spinal dorsal horn [3,4]. Afferent spinal
projections then carry the itch signals in the contralateral
spinothalamic tract to the thalamus and ultimately to the
somatosensory cortex [5]. These central neural connections
involved in itch allow for the modulation of pruritus and
potentially for the de novo induction of pruritus within the
CNS [5,6].
Opioids have well documented roles in both antinocicep-
tion and pruritus [7]. In addition, although pruritus and pain
are distinct sensations, there exist complex interactions
between them. Specifically, itch can be inhibited by pain
and inhibition of the central processing of pain can induce
itch [8]. Therefore, increasing severity of pruritus appears to
Journal of Hepatology 44 (2006) 1141–1149
www.elsevier.com/locate/jhep
lished by Elsevier B.V. All rights reserved.

L. Nelson et al. / Journal of Hepatology 44 (2006) 1141–11491142
depend upon a fine balance between increased activity of
itch pathways and decreased activity in pain pathways [8,9].
Spinal neurons, which process pruritic signals are not
spontaneously active [4], whereas pain processing spinal
neurons are spontaneously active [8]. If spinal pain
processing neurons suppress itch processing neurons, then
inhibition of spinal pain neurons by opiates would be
expected to generate central itch [8,9].
In cholestasis there is clinical and experimental evidence
for both enhanced peripheral (i.e. outside the CNS) and
central opioidergic tone. Specifically, in cholestatic patients
and rodents there are increased levels of circulating
endogenous opioids [10,11]. In addition, there is evidence
of decreased opioid receptor numbers in brains of
cholestatic rats [12], and clinically, patients with cholestatic
itching can experience an opioid withdrawal-like syndrome
when given opioid receptor antagonists [13]. Moreover,
opioid receptor antagonists can ameliorate pruritus in
cholestatic patients [13,14]. Furthermore, cholestatic
rodents exhibit antinociception in response to a thermal
stimulus, which is reversed by the opioid receptor
antagonist naloxone [15]. No animal studies have reported
yet cholestasis-induced differences in nociceptive response
to mechanical stimuli.
Therefore, pruritus in cholestasis may be due to a
combination of the stimulation of cutaneous itch fibers by an
undefined pruritogen (possibly bile salts or histamine, levels
of both of, which are elevated in cholestasis [16,17], and the
action of endogenous opioids acting peripherally and/or
centrally to inhibit pain pathways. In support of this
hypothesis, opioid receptors have been identified on
cutaneous sensory nerve endings [18,19], and in patients,
opiate-mediated itch can be ameliorated by a opioid
receptor blocker, which does not penetrate the CNS [20].
Therefore, we undertook a series of experiments to, firstly,
further characterize cholestasis-associated antinociception
by examining different nociceptive stimuli (both thermal and
mechanical) and secondly, to determine whether cholestasis-
induced antinociception occurs as a result of opioid receptor
stimulation by opioids at peripheral (i.e. outside the CNS) or
central sites. To do this, we assessed anticociception in
cholestastic mice treated with opioid receptor blockers that
either act centrally and peripherally, or are excluded from the
CNS. Given the close association between pruritus and pain,
and the lack of suitable models with, which to study pruritus
directly in animal models of cholestasis, we used cholestasis
associated antinociception as a way to further understand
pruritus in cholestasis.
2. Materials and methods
2.1. Animals
Male C57BL/6 mice (6–8 week old) were obtained from Charles RiverLaboratories (Montreal, QC, Canada) and were housed in transparent
plastic cages in a room with a 12 h light/12 h dark cycle. Mice were fedstandard laboratory chow and tap water ad libitum. All experiments wereapproved by the Animal Care Committee of the University of Calgary andwere performed in accordance with the guidelines of the Canadian Councilon Animal Care.
2.2. Cholestasis induction
Under halothane anesthesia mice were either bile duct (cholestatic) orsham resected (control) as described previously [11,21]. Cholestasis wasconfirmed biochemically by determination of serum bilirubin and alanineaminotransferase levels using kits from Sigma.
2.3. Measurement of nociception
Nociceptive responses to thermal stimulation were evaluated bymeasuring paw withdrawal latency in response to a radiant heat stimulusapplied by a plantar test apparatus (Ugo Basile, Milan, Italy). Nociceptiveresponses to mechanical stimulation were assessed by examining theresponses to plantar stimulation with von Frey monofilaments with bendingforces of 0.166, 1.202 and 1.417 mN as previously described [22]. Briefly, ascore of 0 was given for no response, a score of 1 for withdrawal response,and a score of 2 for withdrawal response with licking or holding of the paw.All measurements were performed first under basal conditions (beforesurgery), and then at different time-points after surgery. Thermal analgesiawas defined as increased withdrawal latency compared to basal values.Mechanical analgesia was defined as decreased nociceptive score comparedto basal values.
2.4. Drug treatments
Systemic naloxone and naloxone methiodide (Sigma, St Louis, USA)treatments were given by daily sub-cutaneaous injections (dorso-scapularregion), at the dose of 1 mg/kg [15], the first injection startingimmediately after bile duct resection or sham surgery. Intraplantarinjection of naloxone methiodide was performed in a total volume of10 ml and at a dose of 15 mg/mouse [23]. The same volume of saline wasinjected into the paw of control mice. Intraplantar injections wereperformed 7-days after bile duct resection or sham surgery, andnociceptive responses were measured 30-min and 2-h after intraplantartreatments.
2.5. Paw Methionine(met)-enkephalin mRNA levels
and Immunohistochemistry
To determine whether the local production (i.e. intraplantar) ofendogenous opioids was increased in cholestatic mice, we removed pawtissues from BDR and sham mice 10 days post-surgery for determination ofpre-proenkephalin mRNA and met-enkephalin protein levels. We chose tospecifically examine met-enkephalin as previous reports have implicatedthis endogenous opioid in cholestasis associated itch [10]. Paw tissue waseither snap frozen on dry ice (for RNA extraction) or was placed in 10%formalin (for immunohistochemistry).
(i) Paw pre-proenkephalin mRNA expression: RNA was extracted fromindividual paw tissues removed from BDR and sham mice using TrizolReagent (Invitrogen, Burlington, ON) and steady-state pre-proenkephalinmRNA levels determined by RT-PCR using the ‘primer dropping’ method[24]. Pre-proenkephalin mRNA primer sequences were obtained fromMouse Genome Informatics (The Jackson Laboratories; MGI AccesssionNumber 1204569) which gave a product size of 141 bp. The internalhousekeeping gene control used was b actin with primers designed to give aproduct of 296 bp [25]. RT-PCR was performed using a ThermolyneAmplitron II cycler (Barnstead International, Dubuque, IO) with denaturing(30 s at 94 8C), annealing (30 s at 55 8) and extension (60 s at 72 8) with33 cycles for pre-proenkephalin and 24 cycles for b actin. RT-PCR productswere separated using a 2% agarose gel (Sigma), stained with ethidiumbromide, and band densitometry readings determined (Gel Doc 2000, BioRad Laboratories, Hercules, CA). Results are expressed as the ratio of banddensities of the target (pre-proenkephalin mRNA) to that of the

Table 1
Serum levels of total bilirubin and alanine aminotransferase (ALT) in bile duct (days 2, 4, 6, 8 and 10 post-surgery) and sham resected mice
Sham Day 2 Day 4 Day 6 Day 8 Day 10
ALT (U/L) 32.8G0.9 65.2G7.4 87.5G21.7* 97.6G11.5** 120.3G11.9*** 93.7G5.3**
Total Bilirubin (mg/dl) 0.25G0.01 1.74G0.38 3.34G0.57 5.42G1.60 24.12G1.25** 32.58G8.34***
Data represent the meanGSEM of results from four mice per group. *P!0.05; **P!0.01 and ***P!0.001 vs sham group.
L. Nelson et al. / Journal of Hepatology 44 (2006) 1141–1149 1143
housekeeping gene (b actin mRNA) and are reported as arbitrarydensitometric units.
(ii) Paw Met-enkephalin Immunohistochemistry: Paw tissues (nZ3mice/group) were removed from formalin, embedded in paraffin, andsectioned (6 m sections). Tissues were then processed for immunohisto-chemistry to detect tissue met-enkephalin expression using a specific met-enkephalin tissue immunohistochemistry staining kit following themanufacturers guidelines (Peninsula Laboratories, Inc., San Carlos, CA).Met-enkephalin was identified using AEC as chromagen (appears as a redcolor with light microscopy).
In additional immunohistochemistry experiments paw tissues fromBDR mice were stained with rat monoclonal antimouse F4/80 antibody(Serotec; stains cells of monocyte/macrophage lineage) followed by arabbit antirat biotinylated secondary antibody (Vector Labs) and DAB(Sigma) as chromagen (appears as reddish brown color under lightmicroscopy).
2.6. Statistical analysis
Data are presented as meanGSEM. For comparisons between twomeans a Student’s unpaired t-test was used. The withdrawal latency andnociceptive score data were analyzed using a one-way analysis of variancefollowed by a Dunnett’s test. With all statistical analyses, P!0.05 wasconsidered significant.
0 2 4 6 8 106
8
10
12
14A
**
Days after surgery
Days after surgery
With
draw
al la
tenc
y (s
)
0 2 4 6 8 100
5
10
15
20
25
30
35 Sham
Cholestatic Filament 4.08
* **
C
Noc
icep
tive
scor
e (%
)
Fig. 1. Effects of cholestasis on thermal (A), or mechanical (B-D) nociception
surgery. Withdrawal latency in response to a thermal stimulus (A) or nocicep
(3.22, 4.08 and 4.17 mN, panels B, C, and D, respectively) were recorded. Dat
surgery) values; P!0.05.
3. Results
3.1. Animal model of cholestasis
Bile duct resected mice developed biochemical evidence
of liver disease and cholestasis as demonstrated by an
increase in serum bilirubin and alanine aminotransferase
levels (Table 1).
3.2. Cholestasis caused increased nociceptice threshold in
response to both thermal and mechanical stimulation
in mice
While sham surgery did not significantly modify
nociceptive response to a thermal stimulus from day 6 to
day 8, the induction of cholestasis significantly increased
the withdrawal latency of mice when their paws were
exposed to a halogen lamp, as compared to basal (before
surgery, time 0) values (Fig. 1A). When a mechanical
Days after surgery
Days after surgery
0 2 4 6 8 100
1
2
3
4
5 Sham
Cholestatic Filament 3.22
B
Noc
icep
tive
scor
e (
%)
0 2 4 6 8 100
10
20
30
40
50 Sham
CholestaticFilament 4.17
D
Noc
icep
tive
scor
e (%
)
in mice at different days after bile duct resection (cholestatic) or sham
tive scores in response to von Frey filaments of different bending forces
a are meanGSEM; *, significantly different from basal (time 0, before

0 1 2 3 4 5 6 77.5
8.5
9.5
10.5
11.5
12.5 saline
nal.
nal. meth.
Sham
A
Days after sham surgery
With
draw
al la
tenc
y (s
)
With
draw
al la
tenc
y (s
)
0 1 2 3 4 5 6 77.5
8.5
9.5
10.5
11.5
12.5 saline
nal.
nal. meth.
Cholestatic
*
**
B
Days after ligation
0 1 2 3 4 5 6 70
5
10
15
20
25
30
35 salinenal.nal. meth.
Sham
Filament 3.84C
Days after sham surgery
Days after sham surgery
Noc
icep
tive
scor
e (%
)
0 1 2 3 4 5 6 70
10
20
30
40 saline
nal.
nal. meth.
Cholestatic
*
Filament 3.84
**
D
Days after ligation
Days after ligation
Noc
icep
tive
scor
e (%
)
0 1 2 3 4 5 6 715
20
25
30
35
40
45
saline
nal.
nal. meth. Sham
Filament 4.08E
Noc
icep
tive
scor
e (%
)
0 1 2 3 4 5 6 70
5
10
15
20
25
30
35
40
45 salinenal.nal. meth.
Cholestatic
Filament 4.08
* *
F
Noc
icep
tive
scor
e (%
)
Fig. 2. Effects of saline, naloxone (nal.) or naloxone methiodide (nal. meth.) treatments on thermal (A, B) and mechanical (C–F) nociception, in sham
control (A, C, E) and cholestatic (B, D, F) mice. Withdrawal latency in response to a thermal stimulus (A, B) or nociceptive scores in response to von
Frey filaments of different bending forces (3.84 mN panels C and D, and 4.08 mN, panels E and F) were recorded. Data are meanGSEM; *,
significantly different from basal (time 0, before surgery) values; P!0.05.
L. Nelson et al. / Journal of Hepatology 44 (2006) 1141–11491144
stimulus was applied to the mouse paw (von Frey filaments
of different sizes representing different bending forces) the
nociceptive score of sham animals was not different from
basal values. However, cholestatic mice exhibited decreased
nociceptive score for two filaments of small and intermedi-
ate sizes (Fig. 1B and C). In the case of the small filament
(Fig. 1B), cholestatic mice had no nociceptive reaction
(score of 0 from day 1 to day 10). For the large size filament,
cholestatic mice, like sham controls, did not show
nociceptive scores different from basal values (Fig. 1D).
Taken together, these results show that in mice induction of
cholestasis caused a decreased nociceptive threshold
characteristic of analgesia, in response to both a thermal
and light to moderate mechanical stimuli.
3.3. Involvement of opioid receptors in cholestasis-induced
increased nociceptice threshold
Further, we showed that systemic pre-treatment of mice
with the opioid receptor antagonist naloxone had no effect
Sham BDR0.0
0.5
1.0
1.5
2.0
2.5
3.0
*
A
B
PENK
Actin
sham BDR
Pre
-pro
en
kep
ha
linm
RN
A le
vels
(a
s a
ra
tioo
f β
Act
in m
RN
A le
vels
;A
rbitr
ary
De
nsi
tom
etr
icU
nits
)
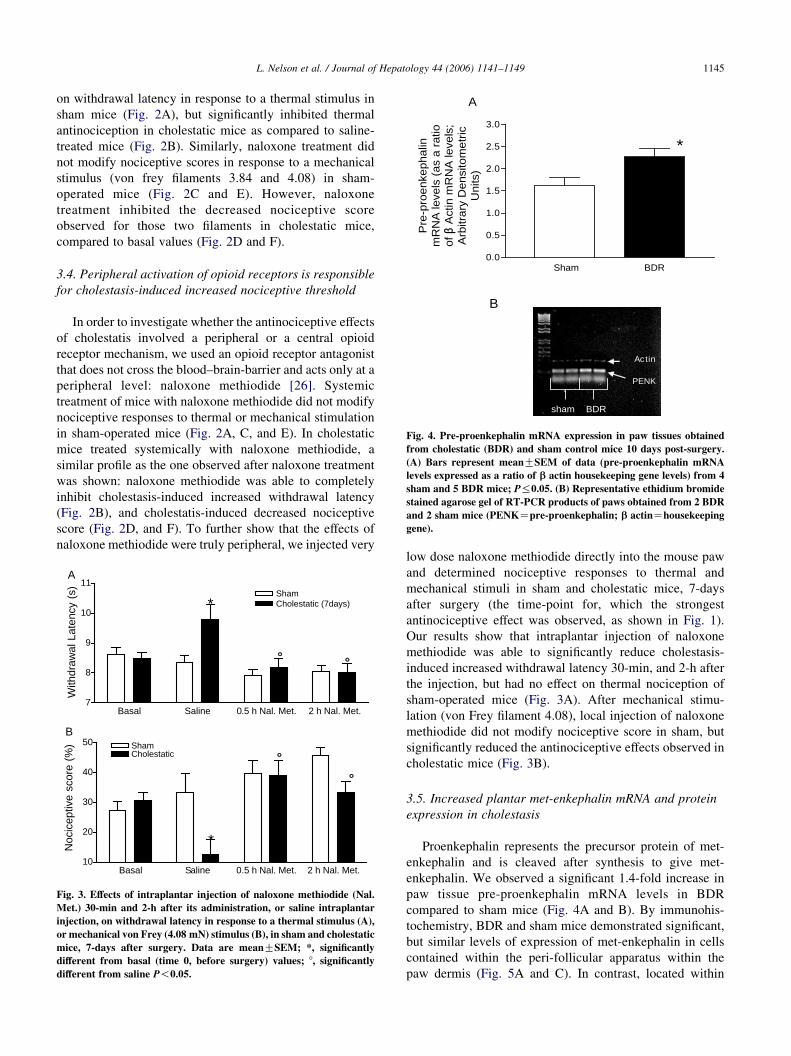
Fig. 4. Pre-proenkephalin mRNA expression in paw tissues obtained
from cholestatic (BDR) and sham control mice 10 days post-surgery.
(A) Bars represent meanGSEM of data (pre-proenkephalin mRNA
levels expressed as a ratio of b actin housekeeping gene levels) from 4
sham and 5 BDR mice; P%0.05. (B) Representative ethidium bromide
stained agarose gel of RT-PCR products of paws obtained from 2 BDR
and 2 sham mice (PENKZpre-proenkephalin; b actinZhousekeeping
gene).
L. Nelson et al. / Journal of Hepatology 44 (2006) 1141–1149 1145
on withdrawal latency in response to a thermal stimulus in
sham mice (Fig. 2A), but significantly inhibited thermal
antinociception in cholestatic mice as compared to saline-
treated mice (Fig. 2B). Similarly, naloxone treatment did
not modify nociceptive scores in response to a mechanical
stimulus (von frey filaments 3.84 and 4.08) in sham-
operated mice (Fig. 2C and E). However, naloxone
treatment inhibited the decreased nociceptive score
observed for those two filaments in cholestatic mice,
compared to basal values (Fig. 2D and F).
3.4. Peripheral activation of opioid receptors is responsible
for cholestasis-induced increased nociceptive threshold
In order to investigate whether the antinociceptive effects
of cholestatis involved a peripheral or a central opioid
receptor mechanism, we used an opioid receptor antagonist
that does not cross the blood–brain-barrier and acts only at a
peripheral level: naloxone methiodide [26]. Systemic
treatment of mice with naloxone methiodide did not modify
nociceptive responses to thermal or mechanical stimulation
in sham-operated mice (Fig. 2A, C, and E). In cholestatic
mice treated systemically with naloxone methiodide, a
similar profile as the one observed after naloxone treatment
was shown: naloxone methiodide was able to completely
inhibit cholestasis-induced increased withdrawal latency
(Fig. 2B), and cholestatis-induced decreased nociceptive
score (Fig. 2D, and F). To further show that the effects of
naloxone methiodide were truly peripheral, we injected very
Basal Saline 0.5 h Nal. Met. 2 h Nal. Met.7
8
9
10
11ShamCholestatic (7days)*
° °
A
With
draw
al L
aten
cy (
s)
Basal Saline 0.5 h Nal. Met. 2 h Nal. Met.10
20
30
40
50 ShamCholestatic °
°
*
B
Noc
icep
tive
scor
e (%
)
Fig. 3. Effects of intraplantar injection of naloxone methiodide (Nal.
Met.) 30-min and 2-h after its administration, or saline intraplantar
injection, on withdrawal latency in response to a thermal stimulus (A),
or mechanical von Frey (4.08 mN) stimulus (B), in sham and cholestatic
mice, 7-days after surgery. Data are meanGSEM; *, significantly
different from basal (time 0, before surgery) values; 8, significantly
different from saline P!0.05.
low dose naloxone methiodide directly into the mouse paw
and determined nociceptive responses to thermal and
mechanical stimuli in sham and cholestatic mice, 7-days
after surgery (the time-point for, which the strongest
antinociceptive effect was observed, as shown in Fig. 1).
Our results show that intraplantar injection of naloxone
methiodide was able to significantly reduce cholestasis-
induced increased withdrawal latency 30-min, and 2-h after
the injection, but had no effect on thermal nociception of
sham-operated mice (Fig. 3A). After mechanical stimu-
lation (von Frey filament 4.08), local injection of naloxone
methiodide did not modify nociceptive score in sham, but
significantly reduced the antinociceptive effects observed in
cholestatic mice (Fig. 3B).
3.5. Increased plantar met-enkephalin mRNA and protein
expression in cholestasis
Proenkephalin represents the precursor protein of met-
enkephalin and is cleaved after synthesis to give met-
enkephalin. We observed a significant 1.4-fold increase in
paw tissue pre-proenkephalin mRNA levels in BDR
compared to sham mice (Fig. 4A and B). By immunohis-
tochemistry, BDR and sham mice demonstrated significant,
but similar levels of expression of met-enkephalin in cells
contained within the peri-follicular apparatus within the
paw dermis (Fig. 5A and C). In contrast, located within

Fig. 5. Immunohistochemistry for met-enkephalin (stained bright red) in paw tissues obtained from sham (A and B) and BDR (C and D) mice 10 days
post-surgery. Filled black arrows point to met-enkephalin positively staining cells within the peri-follicular apparatus in the paws of BDR and sham
mice. Open arrows point to large macrophage-like cells in the dermis of BDR mice, which stain strongly for met-enkephalin. These large cells within
the dermis of BDR mice stain positively for F4/80, a marker for monocytes/macrophages (E; stained reddish-brown color). All pictures were taken at
40! power (For interpretation of the reference to colour in this legend, the reader is referred to the web version of this article.).
L. Nelson et al. / Journal of Hepatology 44 (2006) 1141–11491146
the dermis of only BDR mice were large cells, which stained
strongly for met-enkephalin expression (Fig. 5B and D) and,
which also stained positively for the monocyte/macrophage
marker F4/80 (Fig. 5E).
4. Discussion
The current study confirms previous experimental
observations indicating that cholestasis is associated with
naloxone reversible antinociception [15]; however our
current findings expand on previous observations and
demonstrate that cholestasis-associated antinociception is
elicited by using both mechanical and thermal nociceptive
stimuli. Peripherally administered naloxone readily crosses
the blood–brain-barrier and therefore blocks opioid
receptors on both peripheral nerves and within the CNS.
The blockade of cholestasis associated antinociception had
been attributed to a central effect of naloxone [12,15];
however, our results clearly demonstrate that cholestasis-
induced antinociception results from activation of opioid
receptors outside of the CNS. More specifically, our data
strongly suggests that endogenous opioids act at cutaneous
pain carrying nerve fiber endings in cholestasis to induce
antinociception. Furthermore, we have documented that
cholestasis is associated with the increased cutaneous
production of at least one endogenous opioid (i.e. met-
enkephalin). This is a novel finding with potential clinical
implications for both understanding the pathogenesis of
cholestasis associated pruritus as well as in its management.
Pruritus is a poorly understood sensation, which results
classically from a pruritogen activating specific itch-
dedicated unmyelinated cutaneous C-fiber nerve endings
[3,5,8,9]. The pruritogen(s) involved in cholestasis associ-
ated itch are poorly defined. However, both bile acids and
histamine, which accumulate in the circulation in choles-
tasis [16,17], are capable of activating cutaneous itch fibers
[3,9,27]. The bile salt binding resin cholestyramine has been

Fig. 6. Schematic drawing demonstrating the activation of cutaneous
itch fibers by an unknown pruritogen (e.g. bile acids) in cholestatic
mice, which increases activity in spinal itch pathways. In parallel,
activity in cutaneous pain fibers is suppressed by endogenous opioids
(e.g. met-enkephalin; M-enk) produced by macrophages within the
dermis, resulting in decreased activity in spinal pain pathways, which
leads to a reduction in the suppressive effect, which spinal pain
processing neurons normally exert upon spinal itch processing
neurons. The result is an augmentation in the activity of spinal itch
processing pathways and increased pruritus.
L. Nelson et al. / Journal of Hepatology 44 (2006) 1141–1149 1147
used clinically to effectively treat cholestasis associated
pruritus for decades consistent with a role for bile salts in the
generation of cholestatic pruritus [1]. The role for histamine
in cholestasis associated itch is more questionable as
antihistamines appear to be minimally effective in chole-
static pruritus [1].
Opiates are widely used clinically to relieve pain [7].
However, a common side effect of both peripheral and
centrally administered opiates (e.g. morphine) is pruritus
[7,28]. Opiate-induced pruritus is poorly understood but
may involve drug actions at a number of sites. Within the
CNS, an ‘itch center’ has been postulated as lying within the
medulla oblongata [29]. However, activation of this center
by central opiates results typically in pruritus confined to the
nose and upper part of the face [30]. This pattern of itching
is uncommonly observed in cholestatic patients.
Recently, opioid receptors have been identified on
cutaneous nerve endings [18,19]. Activation of these
receptors by opiates or endogenous opioids results in
antinociception [18,19,31], an effect blocked by local
peripheral application of opioid receptor antagonists [31].
Stimulation of cutaneous nerve pain fibers by opioids leads
to decreased activity within these nerves and ultimately
decreased pain processing activity within neurons in the
spinal dorsal horn [8,9,32] (Fig. 6). This effect could then
lead to an increase in pruritus through a decrease in
inhibitory activity of spinal itch processing neurons.
Elevated levels of circulating endogenous opioids have
been identified in patients and experimental animals with
cholestasis [10,11]. Moreover, rodents with experimental
cholestasis exhibit naloxone-reversible antinociception
[15], an observation confirmed and expanded upon in the
current study. These findings would be consistent with
enhanced opioid driven attenuation of pain processing
pathways in cholestatic rodents. However, the specific site
where endogenous opioids are producing this antinocicep-
tive effect (i.e. within or outside the CNS) is not known. Our
current results demonstrate that opioid-mediated antinoci-
ception in cholestasis is the result of endogenous opioid
effects outside of the CNS. However, opioids can inhibit
pain fiber nerve transmission at two main locations when
acting upon pain impulse carrying nerves outside of the
CNS; namely at the level of the cutaneous nociceptor or at
the level of the nerve cell body within the dorsal root
ganglion [33]. Our findings indicating a reversal of
cholestasis-associated antinociception by the intraplantar
injection of a very low dose of naloxone methiodide (i.e.
only blocking receptors on peripheral nerve endings)
strongly suggest that the effects of endogenous opioids in
cholestasis associated antinociception are occurring at the
level of these cutaneous nociceptors. Moreover, these
results may have direct implications for cholestasis
associated pruritus, and may also explain why scratching
does not typically alleviate cholestasis associated itch [1,2].
Interestingly, endogenous opioids can augment histamine-
induced cutaneous itch in normal volunteers [34],
suggesting a possible indirect effect of endogenous opioids
in the initiation of cholestasis associated itch via a
mechanism similar to that outlined in Fig. 6.
Our suggestion that cholestasis-associated antinocicep-
tion occurs at the level of cutaneous nerve endings is further
supported by our findings documenting an increase in the
local plantar (i.e. dermal) expression of the endogenous
opioid, met-enkephalin, in cholestatic compared to control
mice. We specifically examined the pattern of tissue met-
enkephalin expression in these mice as circulating met-
enkephalin levels are known to be elevated in cholestatic
patients and have been implicated in cholestasis associated
itch [10]. We have identified that BDR mice exhibit an
influx of macrophage-type cells (i.e. F4/80Cve) into the
paw dermis and these cells appear to be the cutaneous
source of met-enkephalin production in these cholestatic
mice. Moreover, monocytes/macrophages can be important
sources of met-enkephalin production in the skin [35,36].
Consistent with this observation, paw pre-proenkephalin
mRNA levels were significantly increased in BDR vs sham
mice. Our demonstration of increased pre-proenkephalin
mRNA levels in the paws of cholestatic mice does not
appear to be simply a non-specific effect of cholestasis as
pro-opiomelanocortin (POMC) mRNA levels are signifi-
cantly reduced in the paws of BDR compared to sham mice
(MG Swain, unpublished observation; POMC is the
precursor for another endogenous opioid, b endorphin).
Our current observations that antinociception in choles-
tasis is due to opioid effects outside of the CNS does not

L. Nelson et al. / Journal of Hepatology 44 (2006) 1141–11491148
preclude other actions of endogenous opioids within the
CNS during cholestasis. Opioid withdrawal-like symptoms
experienced by some pruritic cholestatic patients treated
with opioid receptor antagonists strongly implicate
enhanced central release of endogenous opioids within the
cortical regions of the CNS in these patients [13,14].
However, it is unclear whether the endogenous opioids
acting at these higher CNS centers mediate itch (at least in
part) or modulate the perception of itch.
In conclusion, we have identified that cholestasis-
associated antinociception is mediated by endogenous
opioids acting at the level of cutaneous pain fiber nerve
endings. These results may have direct relevance to
cholestasis associated pruritus and support clinical trials in
using opioid receptor blockers, which do not enter the CNS
to treat patients with cholestasis-induced itch, thereby
avoiding unwanted or clinically problematic CNS side
effects of opioid receptor blockade in cholestatic patients
[13,14,37].
Acknowledgements
This work was supported by operating grants from the
CIHR (MGS and NV). MGS is an AHFMR Senior Scholar
and a CIHR Investigator and NV is an AHFMR Scholar and
CIHR New Investigator.
References
[1] Mela M, Mancuso A, Burroughs AK. Review article: pruritus in
cholestatic and other liver diseases. Aliment Pharmacol Ther 2003;17:
857–870.
[2] Younossi ZM, Kiwi ML, Bopari N, Price LL, Guyatt G. Cholestatic
liver diseases and health related quality of life. Am J Gastroenterol
2000;95:497–502.
[3] Schmelz M, Schmidt R, Bickel A, Handwerker HO, Torebjork HE.
Specific C-receptors for itch in human skin. J Neurosci 1997;17:
8003–8008.
[4] Andrew D, Craig AD. Spinothalamic lamina 1 neurons selectively
sensitize to histamine: a central neural pathway for itch. Nat Neurosci
2001;4:72–77.
[5] Yosipovitch G, Greaves MW, Schmelz J. Itch. Lancet 2003;361:
690–694.
[6] Twycross R, Greaves MW, Handwerker H, Jones EA, Libretto SE,
Szepietowski JC, et al. Itch: scratching more than the surface. Q J Med
2003;96:7–26.
[7] Christo PJ. Opiod effectiveness and side effects in chronic pain.
Anesthesiol Clin North Am 2003;21:699–713.
[8] Ikoma A, Rukwied R, Stander S, Steinhoff M, Miyachi Y, Schmelz M.
Neurophysiology of itch: interaction of itch and pain. Arch Dermatol
2003;139:1475–1478.
[9] Schmelz M. Itch: mediators and mechanisms. J Dermatol Sci 2002;28:
91–96.
[10] Thornton JR, Lasowsky MS. Plasma methionine embaphalin
concentration and prognosis in primary biliary cirrhosis. Br Med J
1988;297:1241–1242.
[11] Swain MG, Rothman RB, Xu H, Vergalla J, Bergasa NW, Jones EA.
Endogenous opioids accumulate in plasma in a rat model of acute
cholestasis. Gastroenterology 1992;103:630–635.
[12] Bergasa NV, Rothman RB, Vergalla J, Xu H, Swain MG, Jones EA.
Central nu-opioid receptors are down-regulated in a rat model of acute
cholestasis. J Hepatol 1992;15:220–224.
[13] Bergasa NV, Yurdaydin C, Turner ML, Schmitt JM, Walker EC,
Jones EA. Effects of naloxone infusions in patients with pruritus of
cholestasis. A double-blind randomized, controlled trial. Ann Intern
Med 1995;123:161–167.
[14] Jones EA. Trials of opiate antagonists for the pruritus of cholestasis:
primary efficacy endpoints and opioid-withdrawl-like reactions.
J Hepatol 2002;37:863–865.
[15] Bergasa NV, Alling DW, Vergalla J, Jones EA. Cholestasis in the
male rat is associated with naloxone-reversible antinociception.
J Hepatol 1994;20:85–90.
[16] Schoenfield LJ, Sjovall J, Perman E. Bile acids on the skin of patients
with pruritic hepatobiliary disease. Nature 1967;213:93–94.
[17] Gittlen SC, Schulman ES, Maddrey WV. Raised histamine
concentration in chronic cholestatic liver disease. Gut 1990;31:96–99.
[18] Machelska H, Stein C. Pain control by immune-derived opioids. Clin
Exp Pharmacol Physiol 2000;27:533–536.
[19] Stander S, Gunzer M, Metze D, Luger T, Steinhoff M. Localization of
m-opioid receptor 1A on sensory nerve fibers in human skin. Regul
Pept 2002;110:75–83.
[20] Yan C-S, Foss JF, O’Connor M, Osinski J, Roizer MF, Moss J.
Efficacy of orally administered methylnaltrexone in decreasing
subjective effects after intravenous morphine. Drug Alcohol Depend
1998;52:161–165.
[21] Abe T, Arai T, Ogawa A, Hiromatsu T, Masuda A, Matsuguchi T,
et al. Kupffer cell-derived interleukin-10 is responsible for impaired
bacterial clearance in bile duct-ligated mice. Hepatology 2004;40:
414–423.
[22] Vergnolle N, Bunnett NW, Sharkey KA, Brussee V, Compton SJ,
Grady EF, et al. Proteinase-activated receptor-2 and hyperalgesia: a
novel pain pathway. Nat Med 2001;7:821–826.
[23] Stein C, Gramsch C, Herz A. Intrinsic mechanisms of antinociception
in inflammation: local opioid receptors and b-endorphin. J Neurosci
1990;10:1292–1298.
[24] Wong H, Anderson WD, Cheng T, Riabowol KT. Monitoring mRNA
expression by polymerase chain reaction: the ‘primer dropping’
method. Anal Biochem 1994;223:251–258.
[25] Pol O, Alameda F, Puig MM. Inflammation enhances opioid receptor
transcription and expression in mice intestine. Mol Pharmacol 2001;
60:894–899.
[26] Obara I, Przewlocki R, Przewlocki B. Local peripheral effects of
m-opioid receptor agonists in neuropathic pain in rats. Neurosci Lett
2004;360:85–89.
[27] Kirby J, Heaton KW, Button JL. Pruritic effect of bile salts. Br Med J
1974;4:693–695.
[28] Szarvas S, Harmon D, Murphy D. Neuraxial opioid-induced pruritus:
a review. J Clin Anesth 2003;15:234–239.
[29] Koenigstein H. Experimental study of itch stimuli in animals. Arch
Dermatol 1948;57:828–849.
[30] Scott PV, Fischer HB. Spinal opiate analgesia and facial pruritus: a
neural theory. Postgrad Med J 1982;58:531–555.
[31] Stein C, Comisel K, Haimerl E, Yassouridis A, Lehrberger K, Herz A,
et al. Analgesic effect of intraarticular morphine after arthroscopic
knee surgery. N Engl J Med 1991;325:1168–1169.
[32] Atanassoff PG, Brull SJ, Zhang J, Greenquist K, Silverman DG,
LaMotte RH. Enhancement of experimental pruritus and mechani-
cally evoked dysesthesia with local anesthesia. Somatosens Mot Res
1999;16:291–298.
[33] Stein C, Machelska H, Schafer M. Peripheral analgesic and anti-
inflammatory effects of opioids. Z Rheumatol 2001;60:416–424.

L. Nelson et al. / Journal of Hepatology 44 (2006) 1141–1149 1149
[34] Fjellner B, Hagermark O. Potentiation of histamine-induced itch and
flare responses in human skin by the enkephalin analogue FK-33-824,
beta endorphin and morphine. Arch Dermatol Res 1982;274:29–37.
[35] Przewlocki R, Hassan AH, Lason W, Epplen C, Herz A, Stein C. Gene
expression and localization of opioid peptides in immune cells of
inflamed tissue: functional role in antinociception. Neuroscience
1992;48:491–500.
[36] Stein C, Hassan AH, Przewlocki R, Gramsch C, Peter K, Herz A.
Opioids from immunocytes interact with receptors on sensory nerves
to inhibit nociception in inflammation. Proc Natl Acad Sci USA 1990;
87:5935–5939.
[37] McRae CA, Prince MI, Hudson M, Day CP, James OF, Jones DE. Pain
as a complication of use of opiate antagonists for symptom control in
cholestasis. Gastroenterology 2003;125:591–596.
Top Related