







Languages
Pages
Legal


International Journal of Pancreatology, 5 (1989) 229-238 $02.00 Copyright �9 1989 by The Humana Press Inc. All rights of any nature whatsoever reserved.
Effect of Alloxan Diabetes on Exocrine Pancreatic Secretion in the Anesthetized Rabbit
C a r m e n A l v a r e z * a n d M a r i a A . L @ e z
Departamento de Fisiologia y Farmacologia, Facultad de Biologia, Universidad de Salamanca, Pza~ ?r S/N, 37008 Salamanca, Spain
Received November 14, 1988; Revised February 16, 1989; Accepted February 20, 1989
Summary
The influence of the endocrine pancreas on the exocrine pancreatic secre- tion of both electrolytes and enzymes was studied in rabbits made diabetic by alloxan administration. No alterations were observed in the flow of pancreatic juice. Bicarbonate concentrations were considerably increased, whereas chloride concentrations were clearly reduced in the alloxan-diabetic rabbits compared with the control animals. Insulin treatment restored anion levels to normal. Similar, although less pronounced changes were seen in the output values of bicarbonate and chloride. There were no sig- nificant differences between the control and alloxan-treated animals in the output of sodium and potassium; however, potassium concentrations ex- hibited a significant rise both in untreated and insulin-treated diabetic rabbits compared with the controls. Total protein and amylase secretion decreased markedly in the diabetic animals. The secretion of amylase was not brought back to control values by additional administration of insulin. These results suggest that the endocrine pancreas plays an extremely im- portant role in exocrine pancreatic function.
Key Words: Insulin; pancreatic juice; electrolytes; amylase.
I N T R O D U C T I O N
T w o secre to ry c o m p o n e n t s can be d i f f e ren t i a t ed in the pancreas: an endo- crine par t and an exocr ine one. M o r p h o l o g i c a l studies have revealed the close con tac t be tween endocr ine and exocr ine cells (1,2). Addi t iona l ly , vascular connec t ions in the f o r m o f an insu lo-ac inar por ta l system have been d e m o n - s t ra ted be tween the islets and the ac inar tissue (3-5) . These m o r p h o l o g i c a l
*Author to whom all correspondence and reprint requests should be addressed.
International Journal of Pancreatotogy 229 Vohlme 5, 1989

230 Alvarez and L@ez
arrays suggest a possible direct influence of islet hormones on the function of the exocrine pancreas (6). The experimental basis for the existence of an islet- acinar axis is strongest for insulin (7).
In humans, diabetes mellitus may be associated with abnormal exocrine pancreatic responses to secretin and cholecystokinin (8-10). Pathologically, in recent-onset Type 1 diabetes the pancreas exhibits acinar cell atrophy sur- rounding insulin-deficient islets (11). The effects of diabetes mellitus have been studied in experimental animals using the B-cell toxins, alloxan, or strep- tozotocin. The acinar cells located close to the islets (peri-insular regions) of the rat pancreas were seen to contain more zymogen granules than distant cells (tele-insular regions) (12,13). This difference was abolished after injec- tion of alloxan and this abolition was not a direct effect of alloxan on the ex- ocrine pancreas (12). On the other hand, in streptozotocin-induced diabetic rats acinar cells located in the peri-insular regions of the pancreas appear totally devoid of amylase immunolabeling while positive and negative cells are found in the tele-insular regions (14). The presence of amylase in pan- creatic tissue (15-23) as well as the basal and stimulated amylase exocrine secretion (19,20,22-24) are severely reduced in diabetic rats. Likewise, in sheep, the volume of pancreatic juice and its protein content decrease mark- edly during alloxan-induced diabetes (25). Both in the rat and in the sheep, the effects of diabetes can be reversed by in vivo insulin administration (15- 21,25). However, Trimble et al. (26) and later Okabayashi et al. (23) found that protein and enzyme outputs in response to cerulein are considerably re- duced in insulin-treated diabetic rat pancreas despite restoration of the pan- creatic enzyme contents to control levels. The literature contains no references to the influence of experimentally-induced diabetes on the concentration of the major electrolytes (bicarbonate, chloride, sodium, and potassium) in pancreatic juice.
The aim of the present work was to study the function of the exocrine pan- creas in resting conditions in rabbits with alloxan-induced diabetes. We also investigated the effect of insulin treatment both on the content of electrolytes in pancreatic juice and enzyme secretion.
MATERIAL AND METHODS
Animals
Male New Zealand rabbits (1.5-2.5 kg b wt) were used in all experiments. The animals had free access to standard rabbit chow~ They were divided into three groups: A) Group I (untreated rabbits) served as nondiabetic controls. B) Group II rabbits comprised animals in which diabetes was induced by intra- venous injection of 75 mg/kg body weight of alloxan monohydrate (Sigma Chemical Co., St. Louis, MO). The diabetic state was confirmed by the posi- tive glucose reaction in urine. Urine samples were collected and analyzed periodically for the presence of glucose using reagent strips (Keto-Diabur Test 5000, Boehringer GmbH, Mannheim, FRG). The rabbits exhibited glyco- suria 3-6 d after alloxan injection. C) Group III, these were alloxan-diabetic
International Journal of Pancreatology Volume 5, 1989

Effect of A lloxan Diabetes on Exocrine Pancreatic Secretion 231
rabbits treated with a subcutaneous injection of 1-5 U of insulin (suspension of porcine protamine insulin, Nordisk, Gentofte, Denmark) given once daily. Treatment with insulin was begun when diabetes had been confirmed and was maintained until the day before the experiments when the rabbits were used to study pancreatic exocrine secretion. The insulin dose given to each animal was adjusted to the amount of urinary glucose excretion and judged to be suitable for avoiding glycosuria.
To examine the chronic effect of diabetes on pancreatic exocrine function, the rabbits were prepared surgically for collection of pancreatic juice 29 d after alloxan administration~ At the time of the experiments, blood samples were taken for glucose measurements from the ear vein before the rabbits were anesthetized.
Collection of Pancreatic Secretion
After a 14-15 h fasting period, but with free access to water, normal and diabetic rabbits were anesthetized by intravenous injection of t .0 g /kg b wt of urethane (Sigma Chemical Co., St. Louis, MO). The animals were then routinely tracheostomized. After performing a median laparotomy, the main pancreatic duct was exposed and cannulated near its entrance to the duod- enum following ligation of the pylorus and cannulation of the bile duct for deviation of bile to the exterior. Samples of pancreatic juice were taken after an equilibration period of 30 min. Pancreatic juice was collected over ice in preweighed plastic tubes in 40-rain batches. Throughout the experiments, the rectal temperature of the animals was kept at 38 _+ 1 ~ by a heating pad.
Assays
The volume of each sample of pancreatic juice was determined by weighing on a fully automatic electronic balance (Type H35, Mettler, Ztirich, Switzer- land) assuming the density of the juice to be 1.0. Bicarbonate concentrations were determined by measuring the total COz content with a Natelson micro- gasometer (Model 600, Scientific Industries, Inc., Bohemia, NY). Chloride concentrations were evaluated by a chloridometer (Model 106, Analytical Control, S.P.A. , Turin, Italy). Sodium and potassium were estimated with an atomic absorption/emission spectrophotometer (Model A-1800, Hitachi, Ltd., Tokyo, Japan). Total protein concentrations were measured by the method of Bradford (27). Amylase activity was determined according to the method of Noelting and Bernfeld (28) as modified by Hickson (29).
Blood glucose was measured with a commercially available kit (BM-Test- Glycemie 20-800R) using a reflectance photometer (Reflolux)o The glycemie test kit and photometer were supplied by Boehringer GmbH, Mannheim, FRG.
Statistical Analysis
Results are expressed as means +_ SEM. One-way analysis of variance fol- lowed by the Scheff~ test (30) were applied for comparisons between groups
International Journal o f Pancreatology Volume 5, 1989

232 A lvarez and L6pez
of animals. Differences with p values of less than 0.05 were considered statis- tically significant.
RESULTS
Rabbits given an alloxan injection characteristically developed polydipsia, polyuria, and gtycosuria, and during the first week showed a slight weight loss. The body weights of the animals on the day of alloxan injection were 1.95 _+0.04 kg in the controls (Group I; n = 20), 1.81 +0.09 kg in the diabetic animals (Group II; n = 7), and 2.00-+ 0.06 kg in the insulin-treated diabetic group (Group III; n = 14), and 29 d after the injection they were 2.35 +0.06 kg, 2.01 _+ 0.08 kg, and 2.32 +0.08 kg, respectively. Blood glucose concentra- tions in the insulin-treated diabetic rabbits at the time of the experiments (6.4-+0.4 mmol /L , n = 8) were similar to those found in the control group (6.1 _4_- 0.4 mmol /L , n = 6). The animals in Group II (alloxan-diabetic rabbits) showed glycosuria, such that it was not considered necessary to check state of hyperglycemia.
No differences were observed in the flow of pancreatic juice between the controls (5.58 _+ 0.53/~L/min, n = 20), treated with alloxan alone (4.95 _+ 0.59 tzL/min, n = 7) and the animals treated with alloxan plus insulin (5.28 +_ 0.56 /zL/min, n = 14). Bicarbonate concentrations were significantly higher in the untreated diabetic rabbits compared to both the controls (p<0 .01 ) and the insulin-treated diabetic animals (p < 0.05) (Fig. 1). The chloride concentra- tions in the rabbits treated with alloxan alone (Group II) were considerably lower than those found in the controls ( p < 0 . 0 t ) and additional treatment with insulin (Group III) led to chloride concentrations that were not signifi- cantly different to those in the control group (Fig~ 1). The changes in the out- put of pancreatic anions were only statistically significant in the case of chlo- ride secretion of the untreated diabetic group as compared with control values ( p < 0.05) (Fig. 1). Whereas sodium concentrations hardly showed any alter- ations, the concentrations of potassium exhibited clear rises in both alloxan- treated groups (Groups II and III) compared with the controls (p<0 .01) (Table 1). The outputs of both sodium and potassium were unaffected by alloxan treatment (Group II); neither were they affected by additional admin- istration of insulin (Group III) (Table 1).
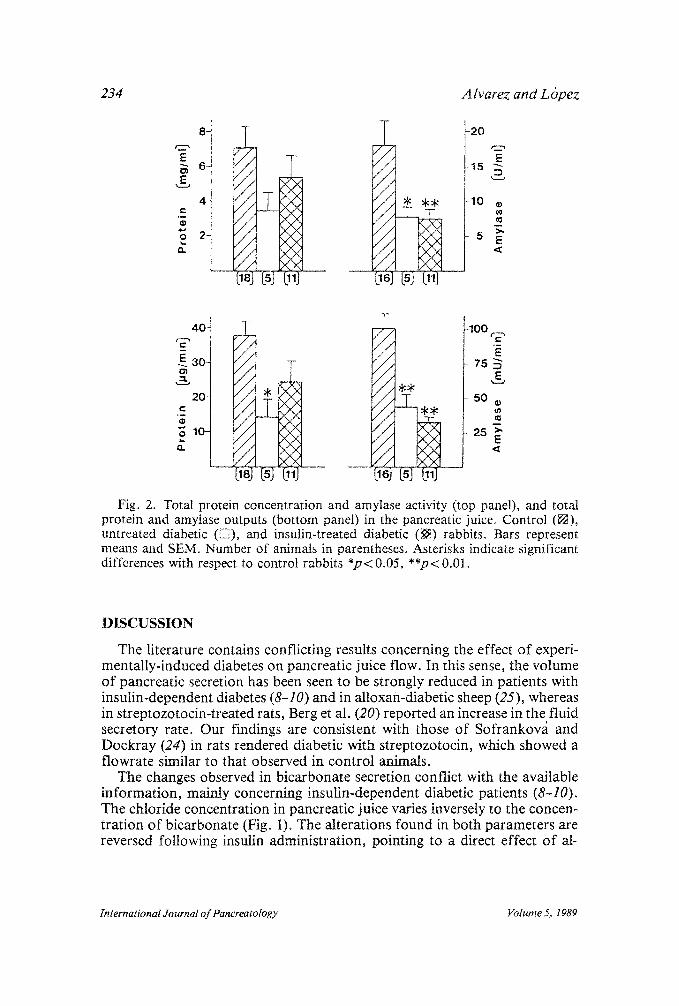
Protein concentrations were seen to decrease after alloxan treatment al- though the changes in this parameter were not significant (Fig. 2). The reduc- tion was more pronounced in output values; thus, the protein output of the untreated diabetic rabbits was significantly lower than that of the control group (p < 0.05) (Fig. 2). Amylase secretion was strongly reduced by alloxan administration (p < 0.01) and was not restored to control values by additional administration of insulin (p<0 .01) (Fig. 2). Furthermore, amylase activity was considerably decreased in both untreated diabetic (p < 0.05) and alloxan plus insulin-treated ( p < 0.01) groups (Fig. 2).
International Journal o f Pancreatotogy Volume 5, 1989

Effect o f A lloxan Diabetes on Exocrine Pancreatic Secretion 233
,__, 10o- .,,,...
E 75- E
SO- C o r~
~- 2 5 -
m
:r162 -7 I"
[18] [7] [137
/ / -V
/ /
/ / / / NN / / ~KN
/ / / / ",(N
[17] (7] Oa]
-120 m
"6 90 E
& 60
3 0 o e-
: 4 0 0 - E
E 3 0 0 - ,_%=
200- C
o too-
e m
/ /
/ /
/ /
/ /
/ / / /
bsl [7] 0a)
/ /
/ / / / / / / / / / / / / / / /
-800 , a
E
- 6 0 0
- 4 0 0
- 2 0 0
Fig. 1. Bicarbonate and chloride concentrations (top panel) and outputs (bottom panel) in the pancreatic juice in control ([]) , untreated diabetic (E~), and insulin- treated diabetic ([]) rabbits. Bars represent means and SEM. Number of animals in parentheses. Asterisks indicate significant differences between control and untreated diabetic rabbits *p<0.05, **p<0.01, and between untreated diabetic and insulin- treated diabetic rabbits ~ p < 0.05.
Table 1 Sodium and Potassium Concentrations and Outputs in the Pancreatic Juice
in Control, Untreated Diabetic, and Insulin-Treated Diabetic Rabbits a
Control Untreated diabetic Treated diabetic
Sodium concentration, mmol /L 157.6+ 4.8 (17) 162.6_+ 2.5 (7) 158.1 _+ 3,3 (14) output, nmol /min 989.7 + 98.3 (17) 816.3 + 111.5 (7) 817.0_+ 99.1 (14)
Potassium concentration, mmol /L 3.1 +_ 0.2 (16) 4.0_+ 0.2** (6) 4 .0+ 0.1"* (14) output, nmol /min 20.2 + t.7 (16) 21.8 +_ 3.3 (6) 20.6-+ 2.4 (14)
~Values are given as means + SEM with number of animals in parentheses. Asterisks indicate signifi- cant differences with respect to control rabbits **p < 0.01.
international Journal o f Pancreatology Volume 5, 1989
234 A lvarez and L @ez
E
&
, w
8-
6-
4-
2-
T
[la3 N i lIJ - -
/ / / / / / / / / /
-2O
E -15 '"
-10 ft./
5 E
,.~. 401 I" "A / / / / / /
20 ~ / / ~ / / . I
~.10 // / / / / / /
-100
75
50
25
&
1
<
Fig. 2. Total protein concentration and amylase activity (top panel), and total protein and amylase outputs (bottom panel) in the pancreatic juice. Control ([]), untreated diabetic ([]), and insulin-treated diabetic (~) rabbits. Bars represent means and SEM. Number of animals in parentheses. Asterisks indicate significant differences with respect to control rabbits *p< 0.05, **p< 0.01.
DISCUSSION
The literature contains conflicting results concerning the effect of experi- mentally-induced diabetes on pancreatic juice flow. In this sense, the volume of pancreatic secretion has been seen to be strongly reduced in patients with insulin-dependent diabetes (8-10) and in alloxan-diabetic sheep (25), whereas in streptozotocin-treated rats, Berg et at. (20) reported an increase in the fluid secretory rate. Our findings are consistent with those of Sofrankov~i and Dockray (24) in rats rendered diabetic with streptozotocin, which showed a flowrate similar to that observed in control animals.
The changes observed in bicarbonate secretion conflict with the available information, mainly concerning insulin-dependent diabetic patients (8-10). The chloride concentration in pancreatic juice varies inversely to the concen- tration of bicarbonate (Fig. 1). The alterations found in both parameters are reversed following insulin administration, pointing to a direct effect of al-
International Journal of Pancreatology Volume 5, 1989

Effect of A lloxan Diabetes on Exocrine Pancreatic Secretion 235
loxan-induced diabetes on the secretion of these anions. An explanation for the changes in anion levels would be the occurrence of an increase in pancre- atic proton flux to interstitial fluid, and, hence, in bicarbonate flux to pan- creatic juice, according to some models proposed for duct cell electrolyte secretion (31-33). Evidence has been advanced to suggest that the sodium/ proton exchange carrier is very efficient in the absence of insulin, at least in muscle cells (34). In untreated diabetic rabbits, this exchanger could be hyper- activated by a higher contribution of electrochemical energy that in animals with normal levels of insulin would be displaced toward other membrane car- riers that require this hormone to be able to function efficiently. According to Kuijpers et al. (35), chloride secretion is regulated secondarily to bicarbon- ate secretion and thus in the alloxan-diabetic rabbits a decrease might occur in chloride secretion to maintain the charge of the anion. Finally, it is also possible that in the secretory ducts of untreated diabetic animals the exchange of ductal bicarbonate with interstitial chloride, which has been shown to occur in rabbit pancreas (36,37), could be altered. Contrariwise, the rise observed in potassium concentrations in alloxan-diabetic rabbits is not reversed after insulin administration. It is clear that both groups treated with alloxan were subject to more stressful conditions, which probably induced an increase in aldosterone secretion, which in turn could account for the greater concentra- tion of this ion in pancreatic juice.
The changes observed in enzyme secretion in the alloxan-diabetic rabbits are consistent with the results found in previous studies in which the secretion of amylase was seen to be strongly decreased in rats with experimental dia- betes (19,20,22-24). However, there are discrepancies concerning the effects of diabetes on total protein secretion. Depending on the authors, in strepto- zotocin-treated rats the secretion of protein was similar to that observed in the control animals (24), or was enhanced (20), whereas in sheep (25), as in the case of our own animals, this parameter was clearly reduced during al- loxan diabetes.
Under our experimental conditions, insulin administration was unable to restore amylase activity and output to normal values. In most of the literature consulted, treatment with insulin has been reported to reverse the effects of chemically induced diabetes on pancreatic enzyme content (15-19,21,23,26) and both its in vitro (19) and in vivo (20) secretion. Thus, these effects of alloxan or streptozotocin on exocrine pancreas appeared to be owing to in- sulin deficiency. The doses of insulin given to normalize secretion were higher than those used in this study (19) or at least were higher than those required to avoid hyperglycemia (20). In in vitro assays with the pancreases from strep- tozotocin-diabetic rats treated with insulin, Trimble et al. (26) and Okabay- ashi et al. (23) found that although these organs had normal enzyme contents, the stimulated amylase and protein secretions were reduced. It is interesting to note that the protocols followed by these authors for the administration of insulin were the same as or similar to those used by us; thus, the insulin dose received by each animal depended either on the amount of urinary glucose excretion (23) or on the daily urine volume (26).
international Journal of Pancreatology Volume 5, 1989

236 A lvarez and L@ez
It is known that insulin has receptors on pancreatic acinar cells (38-41). Insulin regulates acinar protein synthesis, in particular, it promotes the syn- thesis of amylase (16-18,21,42) and it has been shown that insulin potentiates cholecystokinin- or acetylcholine-induced exocrine secretion (43-45). Owing to the existence of the insulo-acinar portal system, arterial blood supplying the pancreatic islets flows to the acini directly (3-5). As a result of this, under normal physiological conditions, the exocrine tissue around the islets is ex- posed to concentrations of insulin that are evidently higher than those found in the systemic circulation. Accordingly, in our assays, like those of Trimble et al. (26) and Okabayashi et al. (23), where insulin doses strictly necessary to normalize the general metabolism were employed, it is possible that the doses employed were insufficient to reverse the effects of diabetes on pancre- atic amylase secretion.
It seems feasible to conclude that the alloxan-induced diabetes affects both hydroetectrolytic and enzymatic exocrine pancreatic secretion, especially regarding the enzymatic component , where the insuto-acinar portal system might play a major role.
ACKNOWLEDGMENTS
This research was supported by a project from the Comisidn Asesora de Investigacidn Cientifica y T~cnica (PR 81-445).
REFERENCES
1 Bendayan M. Contacts between endocrine and exocrine cells in the pancreas. Cell Tiss. Res. 1982; 222: 227-230.
2 Leeson TS, Leeson R. Close association of centroacinar/ductular and insular cells in the rat pancreas. Histol. Histopath. t986; 1: 33-42.
3 Fujita T. Insulo-acinar portal system in the horse pancreas. Arch. Histol. Jpn. 1973; 35: 161-171.
4 Fraser PA, Henderson JR. The arrangement of endocrine and exocrine pancreatic micro- circulation observed in the living rabbit. Q. J. Exp. Physioto 1980; 65: 151-t58.
5 Lifson N, Kramlinger KG, Mayrand RR, Lender EJ. Blood flow to the rabbit pancreas with special reference to the islets of Langerhans. Gastroenterology 1980; 79: 466-473.
6 Henderson JR, Daniel PM, Fraser PA. The pancreas as a single organ: the influence of the endocrine upon the exocrine part of the gland. Gut 1981; 22: 158-167.
7 Williams JA, Goldfine ID. The insulin-pancreatic acinar axis. Diabetes 1985; 34: 980-986. 8 Domschke W, Tyrnpner F, Domschke S, Demling L. Exocrine pancreatic function in
juvenile diabetics. Am. J. Dig. Dis. 1975; 20: 309-312. 9 Frier BM, Saunders JHB, Wormsley KG, Bouchier IAD. Exocrine pancreatic function in
juvenile-onset diabetes mellitus. Gut 1976; 17: 685-691. 10 Sato M, Yamamoto K, Mayama H, Yamashiro Y. Exocrine pancreatic function in dia-
betic children. J. Pediatr. Gastroenterol. Nutr. 1984; 3: 415-420. 11 Foulis AK, Stewart JA. The pancreas in recent-onset Type 1 (insulin-dependent) diabetes
metlitus: insulin content of islets, insulitis and associated changes in the exocrine acinar tissue. Diabetologia 1984; 26: 456-461.
12 Kramer MF, Tan HT. The peri-insular acini of the pancreas of the rat. Z Zellforsch Mikrosk Anat (Berlin) 1968; 86: 163-170.
13 Bendayan M. Morphometfical and immunocytochemical characterization of peri-insular and tele-insular acinar cells in the rat pancreas. Eur. J. Cell Biol. 1985; 36: 263-268.
International Journal of Pancreatotogy Volume 5, 1989

Effect o f A lloxan Diabetes' on Exocrine Pancreatic Secretion 237
14 Bendayan M, Gr4goire S. Immunohisto- and cytochemical studies of pancreatic enzymes in peri-insular and tele-insular acinar cells of streptozotocin-induced diabetic rats. Pan- creas 1987; 2: 272-282.
15 Ben AbdeljliI A, Palta JC, Desnuelle P. Effect of insulin on pancreatic amylase and chymotrypsinogen. Biochem. Biophys. Res. Commun. 1965; 18: 71-75.
16 S61ing HD, Unger KO. The role of insulin in the regulation of c~-amylase synthesis in the rat pancreas. Eur. Jo Clin. Invest. 1972; 2: 199-212.
17 Adler G, Kern HF. Regulation of exocrine pancreatic secretory process by insulin in vivo. Horm. Metab. Res. 1975; 7: 290-296.
18 Korc M, Owerbach D, Quinto C, Rutter WJ. Pancreatic islet-acinar cell interaction: amy- lase messenger RNA levels are determined by insulin. Science 1981; 213: 351-353.
19 Otsuki M, Williams JA. Effect of diabetes mellitus on the regulation of enzyme secretion by isolated rat pancreatic acini. Jo Clin. Invest. 1982; 70: 148-156.
20 Berg T, Johansen L, Brekke IB. Insulin potentiates cholecystokinin (CCK)-induced secretion of pancreatic kallikrein. Acta Physiol. Scand. 1985; 123: 89-95.
21 Gregoire S, Bendayan M. Immunocytochemical studies of pancreatic acinar cells in nor- mal and streptozotocin-induced diabetic rats. Diabetologia 1986; 29: 655-660.
22 Kurahashi M, Inomata K. Amylase secretion by parotid glands and pancreas of diabetic rats during feeding. Am. J. Physiol. 1988; 254: G878-G882.
23 Okabayashi Y, Otsuki M, Ohki A, Suehiro I, Baba S. Effect of diabetes meItitus on pan~ creatic exocrine secretion from isolated perfused pancreas in rats. Dig. Dis. Sci. 1988; 33: 71t-717.
24 Sofrankova A, Dockray GJ. Cholecystokinin- and secretin- induced pancreatic secretion in normal and diabetic rats. Am. J. Physiol. 1983; 244: G370-G374.
25 Pierzynowski SG, Podgurniak P, Mikotajczyk M, Szczesny W. Insulin and the parasym- pathetic dependence of pancreatic juice secretion in healthy and alloxan diabetic sheep. Q. J. Exp. Physiol. 1986; 71: 401-407.
26 Trimble ER, Bruzzone R, Gjinovci A, Renold AE. Activity of the insulo-acinar axis in the isolated perfused rat pancreas. Endocrinology 1985; 117: 1246-1252o
27 Bradford MM. A rapid and sensitive method for the quantitation of microgram quanti- ties of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976; 72: 248-254.
28 Noetting G, Bernfeld P. Sur les enzymes amytolytiques. ItI. La 5-amylase: dosage d' activite et contrSle de 1' absence d' a-amylase. Helv. Chim. Acta 1948; 31: 286-290.
29 Hickson JCD. The secretion of pancreatic juice in response to stimulation of the vagus nerves in the pig. J. Physiol. (Loud.) 1970; 206: 275-297.
30 Scheff4 H. A method for judging all contrasts in the analysis of variance. Biometrika 1953; 40: 87-104.
3 t Swanson CH, Solomon AK. Micropuncture analysis of the cellular mechanism of electro- lyte secretion by the in vitro rabbit pancreas. J. Gen. Physiol. 1975; 65: 22-45.
32 Bonting SL, De Pont JJHHM, Jansen JWCM. The role of sodium ions in pancreatic fluid secretion in the rabbit. J. Physiol. (Lond.) 1980; 309: 533-546.
33 Kuijpers GAJ, Van Nooy IGP, De Pont JJHHM, Bonting SL. The mechanism of fluid secretion in the rabbit pancreas studied by means of various inhibitors. Biochim. Biophys. Acta 1984; 778: 324-331.
34 Clausen T. The significance of the effects of insulin on Na, K-transport in muscle cells. Regul. Peptides 1985; 4(Suppl): 59-67.
35 Kuijpers GAJ, Van Nooy IGP, De Pont JJHHM, Bonting SL. Anion secretion by the isolated rabbit pancreas. Biochim. Biophys. Acta 1984; 774: 269-276.
36 Schulz I, Yamagata A, Weske M. Micropuncture studies on the pancreas of the rabbit. Pfliigers Arch. 1969; 308: 277-290.
37 Mangos JA, McSherry NR. Micropuncture study of excretion of water and electrolytes by the pancreas. Am. J. Physiol. t971; 221: 496-503.
38 Korc M, Sankaran H, Wong KY, Williams JA, Gotdfine ID. Insulin receptors in isolated mouse pancreatic acini. Biochem. Biophys~ Res. Commun. 1978; 84: 293-299.
International Journal of Pancreatology Volume 5, 1989

238 A lvarez and L@ez
39 Sankaran H, Iwamoto Y, Korc M, Williams JA, Goldfine ID. Insulin action in pancreatic acini from streptozotocin-treated rats. IIo Binding of 125I-insulin to receptors. Am. J. Physiol. 1981; 240: G63-G68.
40 M6ssner J, Logsdon CD, Goldfine ID, Williams JA. Regulation of pancreatic acinar cell insulin receptors by insulin. Am. J. Physiol. 1984; 247: GI55-G160.
41 Sj6din L, Holmberg K, Lyden A. Insulin receptors on pancreatic acinar cells in guinea pigs. Endocrinology 1984; 115: 1102-1109.
42 Korc M. Iwamoto Y, Sankaran H, Williams JA, Gotdfine ID. Insulin action in pancreatic acini from streptozotocin-treated rats. I. Stimulation of protein synthesis. Am. J. Physiol. 1981; 240: G56-G62.
43 Kanno T, Saito A. The potentiating influences of insulin on pancreozymin-induced hyperpolarization and amylase release in the pancreatic acinar cell. J. Physiol. (Lond.) 1976; 261: 505-521.
44 Saito A, Williams JA, Kanno T. Potentiation of cholecystokinin-induced exocrine secre- tion by both exogenous and endogenous insulin in isolated and perfused rat pancreata. J. Clin. Invest. 1980; 65: 777-782.
45 Saito A, Williams JA, Kanno T. Potentiation by insulin of the acetylcholine-induced secretory response of the perfused rat pancreas. Biomed. Res. 1980; 1 : 101-103.
International Journal o f Pancreatology Volume 5, 1989
Top Related