







Languages
Pages
Legal


Vo1.153, No. 3,1988
June 30,1988
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Pages 1144-1151
CONPARZSON OF PATTERNS OF EXPRESSION OF TUNOUR NECROSZS
FACTOR. LYNPHOTOXINAND INTERLEUKZN-6 MRNA
MARTIN TURNER AND #ARC FELDMANN
Charing Cross Sunley Research Centre, Lurgan Avenue, Hammersmith, London W6 8LW
Received May 13, 1988
The expression of the mRNA encoding tumour necrosis factor, lymphotoxin and interleukin-6 by peripheral blood mononuclear cells was analysed. Unstimulated cells contained no detectable mRNA for these cytokines, however each mRNA was transiently expressed after stimulation with either the combination of phytohaemaglutinin and phorbol ester or the single stimulus of lipopolysaccharide. The dual stimulus yielded the stronger signal. The cytokine mRNA's had short half lives, but were stabilised following protein synthesis inhibition. Cyclosporin A completely blocked induction of lymphotoxin and partially inhibited induction of TNF and IL-6 mRNA. The features of regulation described in this paper suggest these genes belong within the "early" set of genes expressed following immune cell activation. ©1988AcademicPress, Inc.
Many of the inducible genes expressed by cells of the
immune system can be divided into two groups based on temporal
expression and sensitivity to various dru~s An "early" set of
genes includes those for IL-2, IFN~, c-fos, and c-myc. The
"late" set of genes include those encoding the transferrin
receptor, c-myb and N-ras. Induction of mRNA for the early set
of genes is not blocked by the protein synthesis inhibitor
cycloheximide, unlike mRNA accumulation of the late set (I-6).
The immunosuppressive fungal metabolite, Cyclosporin A has been
Abbreviations used: IL = interleukin; IFN = interferon; TNF = tumour necrosis factor; LT = lymphotoxin; PHA = phytohaemaglutinin; LPS = lipopolysaccharide; PMA = phorbol 12-myristate 13-acetate; CYA = cyclosporin A; CHX = cycloheximide; PBM = peripheral blood mononuclear cells.
0006-291X/88 $1.50 Copyright © 1988 by Academic Press, Inc. All rights of reproduction in any form reserved. 1144

Vol. 153, No. 3, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
found to block the accumulation of mRNA for a number of the
early genes; IL-2, IFN~, and c-myc (5,6).
In the case of IL-2 this has been shown to be due to a block at
the transcriptional level (I). We have previously characterised
the temporal pattern of expression of IL-2, IFN gamma and IL-2
receptor in peripheral blood mononuclear cells (7). It was
therefore of interest to extend this study to include other
lymphokines.
Tumor Necrosis Factor (TNF) and lymphotoxin (LT) are two
closely related lymphokines (8,9) with potent anti tumor
activities ~ and in vivo which are believed to be
important mediators in autoimmune and inflammatory diseases
(10). Interleukin-6, (IL-6) also called Interferon ~2 (11) or
B cell stimulating factor 2 (12) has also been implicated in
various autoimmune diseases, including rheumatoid arthritis
(13). In this paper we have analysed the effect of various
inducers on the induction of TNF, LT and IL-6 mRNA in
peripheral blood mononuclear cells, to determine whether they
are expressed in a manner similar to other lymphokines such as
IL-2 and IFN~.
MATERIALS AND METHODS
Reauents and cells Peripheral blood mononuclear cells (PBM) were obtained
from plateletpheresis residues by Ficoll hypaque density gradient centrifugation. Phorbol 12-Myristate 13-Acetate (PMA), cycloheximide (CHX), lipopolysaccharide from Salmonella typhimurium (LPS) and actinomycin D were purchased from Sigma chemical Co. (Poole, England), phytohaemaglutin (PHA) was from Difco laboratories, (Surrey, England). Cyclosporin A (Sandimmun, Sandoz Ltd.) was made lOOpg/ml in ethyl alcohol and stored at room temperature in the dark. Cells were maintained and stimulated at a density of 2 x106/ml in complete medium (RPMI 1640 supplemented with 10% fetal calf serum, 2mM Glutamine, 100 U/m1 Penicillin and 100 U/m1 Streptomycin) (Gibco, Paisley, Scotland).
RNA blottinu RNA was prepared by guanidinium isothiocyanate lysis
followed by caesium chloride gradient ultracentrifugation.
1145

Vol. 153, No. 3, 1 9 8 8 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Northern or slot blots, hybridisation, and densitometry were performed as described previously (7).
cDNA probe~ The TNF cDNA probe was a 800 bp Eco RI fragment (8), the
LT cDNA probe was a 950 bp Eco RI fragment (9), the IL-6 cDNA probe was a 440 bp Eco RI-Ban II fragment (12) and the 7B6 cDNA was a 708 bp Pst I-Dra II fragment, which contained the Pst I- Dra II region of pBR322. Inserts were labelled by random oligonucleotide priming (14).
Kinetics of ivmDhokine mRNA accumulation: PHA and PMA
are powerful mitogenic stimuli, since PHA induces calcium
flux in T cells (15), while PMA is a potent agonist of protein
kinase C (16). Following treatment with PHA and PMA, mRNA for
TNF, LT and IL-6 was rapidly induced (Figure I). Peak levels
were achieved after 8 hours of stimulation for all 3 mRNA's.
Each mRNA is expressed only transiently, and by 24 hours the
steady state levels of RNA were only about 20% of the maximal
levels seen at 8 hours. 7B6 mRNA was present in unstimulated
cells and its levels were not increased following stimulation,
which was expected since 7B6 has been shown to be cell cycle
independent in PBM (7,17,18).
We wished to use other stimuli to investigate the kinetics
of cYtokine mRNA production. LPS was chosen because unlike PHA
and PMA, LPS is not a primary T cell mitogen, but LPS is a
potent activator of macrophages which represent about 20% of
PBM. LPS transiently induced TNF mRNA with a peak of
expression at 4 hours (Figure 2). IL-6 and LT were also induced
by LPS however the duration of expression was shorter than that
seen with PHA and PMA. Comparison of the autoradiograms also
suggests the levels of mRNA induced were less when LPS was the
inducing stimulus.
mRNA for TNF. LT and IL-6 is unstable: Preliminary experiments
showed that the induction of lymphokine mRNA was not inhibited
1146

Vol. 153, No. 3, 1 9 8 8 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
• . .~ . .: .: .~;,~=:;::i~.~!,:~:':.;: . " .. ~ . : .
TNF a * 115-'tam'If b aiip TNF i••: ~ ~ ~iiii••i!i!-:•~i:
LT • • LT ~ ~ ; ~ ; ~ . . . . . . . . . . ~ ~ i ~ i ~
IL.-'6 i~:'~ . . . . . . ,,':'~ "-~;~:L~i" IL-6 i . ,.~i!...~:..W..~,,-, ~ ~ ,:~ ,,; "::,% . . . . . . . . ~ ~ ? ~ ~.: ~ :.:!:=._
~ ,~ 0 4 8 16 24 48 ~ , ) 0 4 8"16 24 48 v v
PBM were stimulated with PHA (lpg/ml) and PMA (50 ng/ml). RNA was extracted at the times indicated in hours and equal amounts of total RNA run on an agarose gel, transferred to nitrocellulose and hybridised sequentially with TNF, LT, 7B6 and IL-6 probes.
PBM were stimulated with LPS (10pg/ml) and total cellular RNA extracted at the indicated time points. Equal amounts of total RNA (20~g) were run on each track of a northern gel. RNA was transferred to a nitrocellulose filter and probed sequentially with TNF, LT, 7B6 and IL-6 cDNA.
by the presence of the protein synthesis inhibitor
cycloheximide (CHX) and that CHX alone was able to increase
steady state levels of lymphokine mRNA (data not shown).We
measured the half life of TNF, LT and IL-6 mRNA in PHA and PMA
stimulated cells in the presence and absence of CHX after
blocking RNA transcription with Actinomycin D. Total RNA was
collected at various intervals and the mRNA remaining
determined by filter hybridisation. The results show that after
addition of Actinomycin D, TNF, LT and IL-6 mRNA's are rapidly
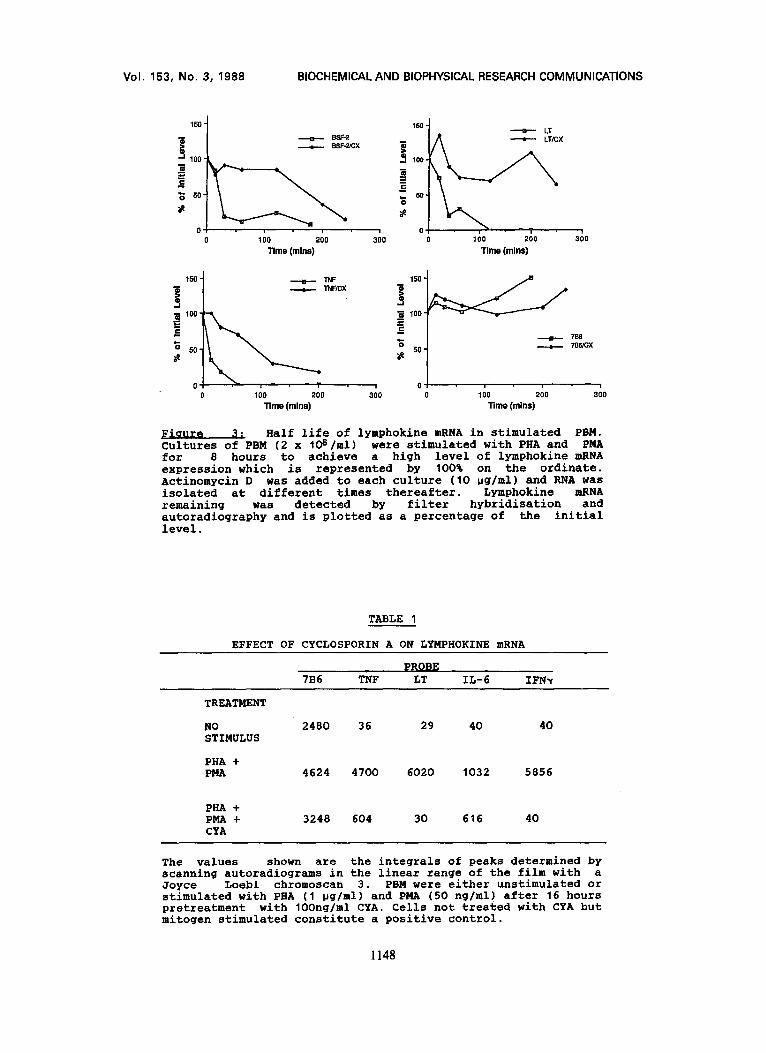
degraded while 7B6 mRNA is more stable (Figure 3). The presence
of CHX increases the half life of TNF, LT and IL-6 mRNA but has
no effect upon 7B6 mRNA demonstrating that the CHX effect is
specific for lymphokine mRNA. This experiment was also
performed on LPS stimulated PBM and gave identical results
(data not shown).
Effect of CyclosDorin A: Preincubation of PBM overnight with
100 ng/ml CYA resulted in the cells being unable to produce
detectable LT or IFN~ mRNA in response to PHA and PMA (Table
I). TNF and IL-6 mRNA induction was not completely abolished by
CYA, while 7B6 mRNA was only minimally affected.
1147
Voh 153, No. 3, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
150 -
>o 100'
m =
'~ 50.
• BSF-2/OX
Time (mine) 3;0
150,
~ 100'
0
LT
100 200
T i m e (ra ins)
3~o
/ 150 "1 - - - -e- - - 1NF
1 - TNF/GX ! !1oo
0 100 2OO
T l m e (ra ins)
150
100
=
50 ~e
• i
300
7B6
8 , 786 /0x
= i
100 200 300 Time (mlns)
Half life of lymphokine mRNA in stimulated PBM. Cultures of PBM (2 x I06/ml) were stimulated with PHA and PMA for 8 hours to achieve a high level of lymphokine mRNA expression which is represented by 100% on the ordinate. Actinomycin D was added to each culture (10 pg/ml) and RNA was isolated at different times thereafter. Lymphokine mRNA remaining was detected by filter hybridisation and autoradiograph¥ and is plotted as a percentage of the initial level.
TABLE I
EFFECT OF CYCLOSPORIN A ON LYMPHOKINE mRNA
p~OBE 7B6 TNF LT ZL-6 IFN~
TREATMENT
NO 2480 36 29 40 40 STIMULUS
PHA + PMA 4624 4700 6020 1032 5856
PHA + PMA + CYA
3248 604 30 616 40
The values shown are the integrals of peaks determined by scanning autoradiograms in the linear range of the film with a Joyce Loebl chromoscan 3. PBM were either unstimulated or stimulated with PHA (I pg/ml) and PMA (50 ng/ml) after 16 hours pretreatment with 10Ong/ml CYA. Cells not treated with CYA but mitogen stimulated constitute a positive control.
1148

Vol. 153, No. 3, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
DISCUSSION
There are striking similarities between our observations on
the modulation of TNF and IL-6 mRNA levels and previous reports
concerning other lymphokine and proto-oncogene expression in
various model systems (2-6). Human PBM can be induced to
transiently express the genes for TNF, LT and IL-6 following
treatment with PHA and PMA together and do so with kinetics
very similar to IFN~ and IL-2 mRNA (7). LPS, which acts
primarily on macrophages and B cells resulted in greater
amounts of TNF mRNA than LT or IL-6 mRNA. However, mRNA levels
peaked earlier and declined faster. Thus the single stimulus of
LPS is not as potent as the dual stimulus of PHA and PMA. We
have recently reported signal requirements for the induction
of TNF in purified T cells (18), optimal induction of TNF mRNA
displays the same two signal requirement in purified T cells
as that for IL-2 or IFN~.
The results with CHX suggest that this drug interferes with
posttranscriptional processing of TNF, LT and IL-6 mRNA by
stabilising the message. A "superinduction" phenomena has been
described for other lymphokine mRNA such as IL-2, IFN~ and
GM-CSF after protein synthesis inhibitors are added to cells
preactivated with mitogens (2, 4, 5). Superinduction of TNF, LT
and IL-6 mRNA can also be achieved if CHX is added after the
initial stimulus (ie: PHA and PMA) (unpublished data). However
culturing cells with CHX alone led to only small increases in
TNF, LT and IL-6 mRNA (unpublished data). Studies of IL-2 and
GM-CSF regulation suggest CHX stabilises preexisting lymphokine
mRNA and has little or no effect on gene transcription (3, 4,
19). Under the conditions of our assay for mRNA half life we
found that TNF LT and IL-6 mRNA's were short lived with a half
life of about 15-30 minutes, similar to that of GM-CSF (19) and
1149

Vol. 153, No. 3, 1 9 8 8 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
IL-la (unpublished data). CHX enhanced the half life of TNF, LT
and IL-6 mRNA but had no effect on the mRNA for 7B6. It has
been suggested (19), that inhibition of protein synthesis
prevents the synthesis of a labile ribonuclease which
recognises a target sequence present in the 3' non coding
region of these mRNAs. TNF, LT and IL-6 all contain variations
of this sequence (8, 9, 12), thus we have provided evidence
that this form of control operates for these mRNAs in activated
lymphoid cells.
The induction of LT and IFN~mRNA was completely blocked
by pretreatment of PBM with CYA. In contrast some TNF and IL-6
mRNA was produced, suggesting that cells (ie macrophages) that
produce IL-6 and TNF are not as sensitive to CYA inhibition as
those that produce only LT and IFN~ (T and B lymphocytes). This
data agrees with the findings of Espevik et.al. (20) which
demonstrated CYA reduced secretion of TNF, LT and IFN~ by
activated lymphoid cells, and demonstrates that this effect is
manifest at the level of mRNA.
The data presented here , together with previous results
(I-6), indicates that the regulation of inflammatory mediators
is complex, and occurs at a variety of levels; at the
transcriptional level, regulated by CYA and
posttranscriptionally, regulated by CHX. TNF, LT and IL-6
appear to be members of the early set of genes expressed during
the immume response.
ACKNOWLEDGMENTS We would like to thank Dr. Contreras (North London blood transfusion service) and her colleagues for platletpheresis residues, Dr. H. M. Shepard for TNF and LT cDNA, Dr. T. Kishimoto for IL-6 cDNA and Dr. U. Torelli for the 7B6 cDNA. This work was supported by the Nuffield foundation (0liver Bird Grant) and the Arthritis and Rheumatism Council.
1150

Vol. 153, No. 3, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
EEEZBEaCK~
I. Kronke, M. , Leonard, W. J. , Depper, J. M. , Arya, S. K. , Wong-Staal, F. , Gallo, R. C. , Waldmann, T.A. & Greene, W. C. (1984) Proc. Natl. Acad. Sci. U. S. A. 81, 5214-5218.
2. Reed, J. C. , Alpera, J. D. , Nowell, P. C. & Hoover, R. G. (1986) Proc. Natl. Acad. Sci. U. S. A. 83, 3982-3986.
3. Kronke, M. , Leonard, W. J. , Depper, J. M. & Greene, W. C. (1985) J. Exp. Med. 161, 1593-1598.
4. Shaw, J. , Meerovitch, K. , Elliot, J. F. , Bleackley, R. C. & Paetkau, V. (1987) Molecular immunology 24, 409-419.
5. Granelli-Piperno, A. , Andrus, L. & Steinman, R. M. (1986) J. Exp. Med. 163, 922-937.
6. Reed, J. C. , Nowell, P. C. & Hoover, R. G. (1985) Proc. Natl. Acad. Sci. U. S. A. 82, 4221-4224.
7. Buchan, G. , Barrett, K. , Fujita, T. , Taniguchi, T. 5, Maini, R. N. & Feldmann (1988) Clin. Exp. Immunol. 71, 29 - 301.
8. Pennica, D. , Nedwin, G. E. , Hayflickli J. S. , Seeburg, P. H. , Derynk, R. , Palladino, M. A. , Kohr, W. J.
24- Aggarwal, B. B. & Geoddel, D. V. (1984) Nature 312, 729.
9. Gray, P. W. , Aggarwal, B. B. , Benton, C. V. , Bringman, T. S. , Henzel, W.J. , Jarrett, J. A. , Leung, D. W. , Moffat, B. , Ng, P. , Svedersky, L. P. , Palladino, M. A. & Nedwin, G. E. (1984) Nature 312, 721-724.
10. Buchan, G. N: Barrett, K. , Turner, M. , Chantry, D. , Maini, R. & Feldmann, M. (1988) Clin. Exp. Immunol. in press.
11. Zilberstein, A. , Ruggieri, R. , Korn, J. & Revel, M. (1986) EMBO Journal 5, 2529-2537.
12. Hirano, T. , Yasukawa, K. , Hiranda, H. , Taga, T. , Watanabe, Y. , Matsuda, T. , Kashiwamura, S. Nakajima, K. , Koyama, K. , Iwamatsu, A. , Tsunasawa, S. , Sakiyama, F. , Matsui, H. , Takahara, Y. , Taniguchi, T. & Kishimoto, T. (1986) Nature 324, 73-76.
13. Hirano, T. , Matsuda, T. , Turner, M. , Miyasaka, N. , Buchan, G. , Tang, B. , Sato, K. , Shimizu, M. , Maini, R., Feldmann, M. & Kishimoto, T. (1988) (submitted).
14. Feinburg, A. P. & Vogelstein, B. (1984) Analyt. Biochem. 137, 266-277.
15. Hesketh, T. R. , Smith, G. A. , Houslay, M. D. , Warren, G. B. & Metcalfe, J. C. (1977) Nature 267, 490-494.
16. Nishizuka, Y. (1984) Nature 308, 693-698. 17. Kaczmarek, L. , Calabretta, B. & Baserga, R. (1985) Proc.
Natl. Acad. Sci. U. S. A. 82, 5375-5379. 18. Turner, M. Londei, M. & Feldmann, M. (1987) Eur. J.
Immunol. 17, 1807-1814. 19. Shaw, G. & Kamen, R. (1986) Cell 46, 659-667. 20. Espevik, T. , Figari, I. S. , Shalaby, M. R. , Lackides,
G. A. , Lewis, G. D. , Shepard, H. M. & Palladino, M. A. (1987) J. Exp. Med. 166, 571-576.
I151
Top Related