







Languages
Pages
Legal


Algae Biomass for Bioenergy in Denmark Biological/Technical Challenges and Opportunities
Susse Wegeberg
Department of Biology, SCIENCE
Claus Felby
Forrest & Landscape, LIFE
University of Copenhagen, 2010

1
0. Contents
1. Dansk resumé ................................................................................................................................................ 4
2. Introduction ................................................................................................................................................... 7
3. Photosynthesis ‐ potential for biomass production ...................................................................................... 9
Can we improve the efficiency of photosynthesis? .................................................................................... 11
Recommended reading ............................................................................................................................... 12
4. The Chemistry of Algae ................................................................................................................................ 13
Algae cell walls ............................................................................................................................................. 13
Algae carbohydrates .................................................................................................................................... 15
Mannanes ................................................................................................................................................ 15
Cellulose .................................................................................................................................................. 16
Ulvan ........................................................................................................................................................ 16
Fucans ...................................................................................................................................................... 16
Xylanes ..................................................................................................................................................... 16
Alginic acid ............................................................................................................................................... 16
Sulfated carbohydrates ........................................................................................................................... 16
Intracellular carbohydrates ..................................................................................................................... 17
Algae oils ...................................................................................................................................................... 17
Inorganic elements ...................................................................................................................................... 17
5. Microalgal production for bioenergy .......................................................................................................... 18
Potential species .......................................................................................................................................... 19
Growth rate/lipid content ........................................................................................................................... 20
Mass cultivation ........................................................................................................................................... 23
Open ponds (Christi 2007, Lee 2001, Li et al. 2008; and references herein) .......................................... 23
Photobioreactors ..................................................................................................................................... 24
Alternative designs ‐ OMEGAs (Trent, WSA (2009)) ................................................................................ 26
Comparison of the cultivation technologies ........................................................................................... 27
Harvesting .................................................................................................................................................... 28
Centrifugation .......................................................................................................................................... 29
Chemical flocculation .............................................................................................................................. 29
Settling ponds .......................................................................................................................................... 29
Recommended reading ............................................................................................................................... 30

2
6. Possibilities for improvement of microalgal production systems ............................................................... 31
Yield optimization of photosynthetic microalgal production systems ........................................................ 31
Photosynthetic efficiency ........................................................................................................................ 31
Temperature ............................................................................................................................................ 33
Harvest timing ......................................................................................................................................... 34
Mixed cultures of algae and bacteria ...................................................................................................... 34
Heterotrophic microalgae ........................................................................................................................... 35
Mixotrophic algae ........................................................................................................................................ 37
7. Macroalgal production for bioenergy ......................................................................................................... 38
Species and growth potentials; which species are suitable for cultivation in DK? ..................................... 40
Growth rates ............................................................................................................................................ 41
Mass cultivation and biomass yield of seaweeds in tanks .......................................................................... 42
Sea‐based mass cultivation and biomass yield of seaweeds ...................................................................... 45
Designs and harvest of sea‐based cultivation systems ........................................................................... 45
Identification of optimal locations for sea‐based cultivation ................................................................. 48
Fouling ..................................................................................................................................................... 48
Comparison of sea‐based cultivation biomass yields .............................................................................. 49
Reductions in biomass yield .................................................................................................................... 49
Storage and down‐stream processing ......................................................................................................... 52
Recommended reading ............................................................................................................................... 53
8. Possibilities for improvement of macroalgal production systems .............................................................. 54
Species/strain screening .............................................................................................................................. 54
Sporophyte density ..................................................................................................................................... 55
Multiple harvests ......................................................................................................................................... 56
Manipulating spore formation ................................................................................................................ 56
Resting gametophyte cultures................................................................................................................. 56
9. Bioenergy conversion technologies for algae ............................................................................................. 58
Algae bio‐diesel ........................................................................................................................................... 58
Algae ethanol ............................................................................................................................................... 59
Combustion ................................................................................................................................................. 60
Biogas .......................................................................................................................................................... 60
Thermo‐chemical processing ....................................................................................................................... 61
Microalgae for H2 production ...................................................................................................................... 62

3
10. Sequestration of CO2, NOX and SOX ........................................................................................................... 63
Growth rates and CO2 uptake ...................................................................................................................... 63
Flue gas as CO2 source ................................................................................................................................. 63
NOx, SOx ....................................................................................................................................................... 64
11. Scenario for microalgal production in Denmark ....................................................................................... 66
Production and production systems ........................................................................................................... 66
Up‐scaling .................................................................................................................................................... 67
Costs ............................................................................................................................................................ 68
Fertilizers ................................................................................................................................................. 68
Seawater .................................................................................................................................................. 68
CO2 ........................................................................................................................................................... 68
Area needs ................................................................................................................................................... 69
Potential for energy production .................................................................................................................. 70
Prospects and limitations ............................................................................................................................ 70
12. Scenario of macroalgal production and conversion .................................................................................. 71
Area needs ................................................................................................................................................... 71
Production costs .......................................................................................................................................... 74
Reported production costs ...................................................................................................................... 75
Macroalgae potential for energy production .............................................................................................. 75
13. Recommendations ..................................................................................................................................... 76
Algae production ......................................................................................................................................... 76
Conversion technology development ......................................................................................................... 77
Fermentation to liquid energy carriers ................................................................................................... 78
Combustion and gasification ................................................................................................................... 78
Biogas ...................................................................................................................................................... 79
Algae oils .................................................................................................................................................. 79
14. Literature ................................................................................................................................................... 80
Appendix 1 ............................................................................................................................................... 88

4
1. Dansk resumé
Dette review er udført på foranledning af DONG Energy, Danmark, som også har støttet arbejdet
økonomisk.
Udnyttelse af biomasse til energiformål er centralt i forbindelse med en sikker og bæredygtig levering af
energi. For dog at opnå en succesfuld implementering af bioenergi må nye og bæredygtige
biomasseressourcer og tilsvarende konverteringsteknologier identificeres og udvikles.
En hidtil uudnyttet, men dog ikke helt uudforsket, biomasse er den såkaldte blå biomasse, den primære
produktion i vand ‐ alger.
Alger er fotosyntetiserende organismer, som, i kontrast til landplanter, ikke producerer blomster og frø,
men danner sporer til formering og som spredningsenhed. Enkelt‐cellede alger, mikroalger, findes især frit i
vandmassen og danner fytoplankton. De store alger makroalgerne, har ikke rødder, men blot
fasthæftningsorganer, og de har heller ikke specialiseret væv til transport af vand, sukkerstoffer og
næringssalte, idet de er omgivet af vand. Alger er en gammel og meget heterogen gruppe af organismer,
der spænder fra en størrelse på få nanometer til de store arter på op til 60 m længde.
Som alle andre planter, er alger primært opbygget af sukkerstoffer som dannes ved fotosyntesen.
Sammenlignet med landjordens planter indeholder alger kun en lille mængde cellulose og i enkelte tilfælde
meget små mængder lignin. Fraværet af lignin gør, at alger i stedet har udviklet svært nedbrydelige
sukkerstoffer, ligesom cellevæggen kan binde store mængder vand. Sidstnævnte egenskab udnyttes bl.a. i
flere geleringsmidler baseret på alger. Dertil kommer at alger ofte har svovl og nitrogen som strukturelle
komponenter i deres cellevægge og ydermere kan have et indhold af salte og uorganiske forbindelser på
over 30 %.
Alger kan danne meget produktive økosystemer. Sammenlignet med landjordens planter er det teoretisk
muligt at lave 3‐5 gange så meget biomasse på samme areal, og interessen for at udnytte alger som
biomasse til produktion af bioenergi har været kraftigt stigende de seneste år. De umiddelbare fordele ved
at udnytte algebiomasse til energi er flere, men her kan nævnes: Alger kan dyrkes i hav‐ eller brakvand
direkte i havet eller i anlæg på arealer, der ikke kan anvendes til landbrug. Produktionen af
mikroalgebiomasse kan kombineres med direkte fiksering af CO2 fra røggasser, og makroalgeproduktion i
havet kan muligvis få en positiv økologisk effekt ved at bidrage til fjernelse af næringssalte samt fungere
som skjulesteder for fiskeyngel
Som enhver anden biomasse kan alger omdannes til både faste, flydende og gasformige energibærere. Men
for alger gælder, at deres kemiske sammensætning med et højt indhold af svovl, nitrogen og salte gør dem
uegnede til afbrænding, ligesom deres indhold af forgærbare sukre er lavt og derfor ikke velegnede til f.eks
ethanolproduktion. I praksis er de umiddelbare muligheder for at konvertere alger til energibærere derfor
begrænset til:

5
Udnyttelse af mikroalgernes evne til at producere fedtstoffer, lipider, som kan omdannes til
biodiesel
Biogas dvs methanproduktion af mikro‐ og makroalgebiomasse og restprodukter fra alge‐biodiesel
Andre mulige konverteringsteknologier er pyrolyse og forgasning til efterfølgende fremstilling af dieselolier,
der vil dog formentlig være store problemer med uønskede sidereaktioner, tjære og belægninger grundet
algernes høje indhold af nitrogen, svovl og salte.
For algebiodiesel er der i laboratorieskala påvist et meget stort potentiale. Men der er problemer med den
efterfølgende processering til biodiesel. Algernes cellevægge er meget robuste og sammen med deres lille
størrelse er det omkostningstungt at ekstrahere olien fra algerne.
Den største udfordring i at kunne udnytte algebiomasse i stor skala til energi ligger ikke i konverteringen til
energibærere, men i dyrkning, høst og opbevaring af millioner af tons alger. Eksisterende danske naturlige
forekomster af alger vurderes ikke tilstrækkelige til at kunne understøtte en udnyttelse til energi.
For mikroalger arbejdes der både med bassiner (open ponds) og egentlige reaktorer (bioreactors). Begge
principper er afprøvet i pilot‐skala, men de opnåede udbytter er lave og der mangler stadig metoder og
systemer til at kunne håndtere nærings‐ og affaldsstoffer samt undgå infektioner.
For makroalger har der igennem århundreder været en egentlig kommerciel produktion til fødevarer og
tilsætningsstoffer. Der er dog ingen af disse systemer, der er i stand til at levere de udbytter og mængder
som vil være nødvendige for stor‐skala energiproduktion. Hertil kræves intensiv forskning både i de
forskellige algearter, marine dyrkningssystemer samt identifikation af optimale dyrkningslokaliteter.
I forhold til et dansk scenarie vurderes det umiddelbare energipotentiale dog til at være begrænset.
Indenfor en kort til mellemlang tidshorisont er den eneste tilgængelige konverteringsteknologi biogas.
Udbyttet fra 1 million tons makroalgebiomasse til biogas vil være i størrelsesordenen 10 GJ, hvilket
sammenlignet med andre biomasser er relativt lavt. Omkostningsniveauet herved vurderes også endnu at
være højt med en faktor 3‐10 i forhold til andre biomasser. Ifald det lykkes at etablere en teknologi til
produktion af algebiodiesel vil potentialet kunne være væsentligt højere, men der er endnu ikke et
teknologisk og biologisk grundlag herfor.
Set i et længere tidsperspektiv er det sandsynligt, at der vil kunne udvikles både biologi og teknologi som vil
kunne muliggøre et væsentligt bidrag fra alger til energiforsyningen. Men det forudsætter en betydelig og
målrettet forskning og udvikling. Fokus heraf bør være både på grundlæggende biologiske og biokemiske
problemstillinger samt udvikling af egentlige teknologier til stor‐skala produktion af algebiomasse med
tilstrækkeligt lave omkostninger. Dertil kommer, at der også forestår et stort arbejde med udredning af de
miljømæssige konsekvenser ved anvendelse af havet til algedyrkning i stor skala.
Der er iværksat en del projekter i udlandet, som søger at udvikle især mikroalger til produktion af biodiesel.
Projekterne er i sig selv ambitiøse og udfordrende, men må karakteriseres som værende risikofyldte. En del

6
af projekterne bygger på, at der foruden energi også kan produceres højværdi‐produkter såsom special‐
kemikaler, medicin m.m. Dette er absolut muligt, men i relation til en størrelsen af en energiproduktion vil
sådanne produkter kun bidrage marginalt til den overordnede økonomi.
Anvendelsen af alger til energi kan derfor karakteriseres ved både at have et meget stort potentiale, men
også en række teknologiske og biologiske udfordringer, som gør området risikofyldt ud fra et
investeringsmæssigt synspunkt. For energisektoren vil det derfor være fornuftigt at opretholde et
forsknings‐ og vidensmæssigt beredskab som vil muliggøre, at man kan udnytte eventuelle gennembrud
indenfor algeproduktion. Især vil det være vigtigt at identificere de anvendelser, hvor alger vil kunne have
synergi med den nuværende udnyttelse af biomasse til energi.
I praksis bør man holde sig grundigt orienteret om resultaterne af den igangværende udvikling for at kunne
identificere potentielle muligheder der måtte opstå. Derudover er der også behov for at afklare potentialet
for flere forskellige algeproduktioner under danske forhold f.eks ved gennemførelse af pilotforsøg. Dette vil
muliggøre, såfremt der opnås et eller flere gennembrud, nationalt så vel som internationalt, at der kan
åbnes nye muligheder for udnyttelse af alger til energi, og man vil kunne handle hurtigt derefter.

7
2. Introduction
This review is developed on behalf of DONG Energy, Denmark, whom also funded the work.
The utilization of biomass for energy is one of the key elements in a sustainable and secure energy supply.
However, the successful implementation of bioenergy requires the development of new and sustainable
sources of biomass and associated conversion technologies. Biomass production must be adapted and
developed to secure and maintain a proper balance between food, feed and energy production in an
ecologically and economically responsible way. The development of high yield sustainable production
systems is a major challenge for the industrial and scientific communities if the very high potential of
biomass for energy is to be realized.
The focus has increasingly been directed to algae biomass as a resource for bioenergy. The advantages of
algal biomass to terrestrial plants as a source of transportations fuels seem to be numerous according to
Rodolfi et al. 2009:
Oil yield per area of microalgae could greatly exceed the yield of the best oilseed crops
Microalgae grow in an aquatic medium, but need less water than terrestrial crops
Microalgae can be cultivated in seawater or brackish water on non‐arable land, and do not
compete for resources with conventional agriculture
Microalgae biomass production may be combined with direct bio‐fixation of waste CO2
Fertilizers for microalgae cultivation can be obtained from wastewaters
Algae cultivation does not need herbicides or pesticides
The residual biomass after oil‐extraction may be used as feed or fertilizer, or fermented to produce
ethanol or methane
The biochemical composition of the algal biomass can be modulated by varying growth conditions
the oil content can be highly enhanced
Furthermore, solely considering sea‐based cultivation of macroalgae:
Seaweed farms will not involve land areas at all
Seaweed farms may have a positive ecological impact by nutrient stripping and providing nursery
grounds for fish fry
As indicated above there are several ways of achieving energy from algal biomass (Fig. X):
Oil from lipid containing especially microalgae
Ethanol from fermentation of sugars in especially macroalgae
Methane from fermentation of algal biomass in general
Heat and power by burning of algal biomass in general

8
However, in the comprehensive work of the Aquatic Species Program by the North American National
Renewable Energy Laboratory and their subcontractors, which was conducted in the 1980’s and 90’s
(Sheehan et al. 1998), four major difficulties associated with microalgal mass culture for fuel production
were identified connected to open pond systems:
Photosynthesis efficiency for light energy and high lipid production
Fundamentals of species selection and control in open pond systems
Mechanisms (and control) of algal bio‐flocculation
Effects of non‐steady‐state operating conditions on algal metabolism
as only a very low‐cost system, based on open ponds mixed on low velocities and using very simple
harvesting process, could be considered in such a production.
They also conclude that even with these rather favourable, though plausible, assumptions, costs would still
be well above those for current, or projected, oil prices.
The program was closed by the Clinton‐government in the 1990’s, but here 10 years after algal biomass
derived bioenergy is intensely hyped. What is the actual present status of research and commercialization
of algal biomass based bio‐fuels? This project seeks to identify and investigate the challenges in producing
algal biomass for bioenergy production as well as evaluate the potential at present.

9
3. Photosynthesis ‐ potential for biomass production
How much biomass and thus bioenergy can we harvest from plants regardless if they are grown in the sea
or on land? As any other living organisms the growth of plants are dependent upon the rate by which they
can feed their metabolism with energy and nutrients. In practice this means that the levels of water, CO2,
nutrients and light that is available, limit the efficiency by which plants convert solar energy to biomass.
Data on biomass production from algae are often reported as the production in kg m− 2 d− 1. This unit is
valuable from a laboratory scale point of view, but when looking at the millions of tones biomass required
for the energy sector as well as comparing to current use of land based biomass for energy, it is more
relevant to look at the annual production of biomass from 1 ha.
Current biomass potentials from agricultural crops range from 7 to more than 120 ton ha‐1 with the highest
levels found in the tropical regions (Table 3.1). Translating reported algae potentials to t ha‐1 (Table 3.2) it
can be seen that they are potentially capable of producing more biomass pr. area unit than current
agricultural crops.
Crop Record yield t/ha Average yield t/ha
Corn 19.3 4.6
Wheat 14.5 1.9
Soybeans 7.4 1.6
Sorghum 20.1 2.8
Oats 10.6 1.7
Barley 11.4 2.1
Potatoes 94.1 28.2
Sugar beets 121.0 42.6
Table 3.1. Comparison of record and average yields for agricultural crops (Chrispeels & Savada 2003).
kg m− 2 d− 1 t /ha
Photobioreactor I 0,072 262
Photobioreactor II 0,048 175
Raceway ponds 0,035 127
Table 3.2. Potential yields from microalgae production (Christi 2007). Note that data have not been obtained in large‐scale.
The major limiting factor for biomass production in land‐based plants is the amount of water available. For
algae there is no limitation of water just as the nutrients are more mobile in an aqueous environment and
can be maintained at optimum levels. Furthermore we may have the opportunity of raising the CO2 level by
dissolving flue gas in the aqueous environment of the algae. Therefore it is possible to provide conditions
where the only limiting factor is the level of light available to the algae, and we may calculate the
theoretical maximum for biomass production at a given amount of solar radiation.

10
The basic process in photosynthesis is about transferring the energy from the incoming photons to
electrons in the photosynthetic pigments. This transfer of light energy to electrons is subsequently
transformed into chemical energy that is used to fixate CO2 into carbohydrates. An outline of
photosynthesis can be seen in Fig. 3.1, note the division between light reactions capturing the solar
radiation and the Calvin cycle responsible for the fixation of CO2 into carbohydrates.
Fig. 3.1. Outline of the reactions taking place during photosynthesis. The light reactions capture the energy in the light and the fixation of CO2 into sugars (production of carbohydrates takes place in the Calvin cycle).
The rate limiting step in the process is not the rate by which plants are able to convert photons into excited
electrons, but the rate by which the carbohydrates are synthesized in the Calvin cycle.
The electrons in the photosynthetic pigments absorb the solar energy by moving between orbitals i.e. the
electron is excited into an orbital with a higher energy level. Subsequently as the electrons move to lower
orbitals, the energy is released and used to split water and create energy in chemical bonds driving the
process of CO2 fixation.
Looking at the overall equation (1) for the photosynthesis it can be seen that the levels of CO2, water and
light are interdependent. The level of solar radiation is specific for a given area at a given latitude, thus if
we know the latitude we can determine the upper level of productivity by algae or any other plant for a
given location.
(1) 6CO2 + 12H2O + light (photons) ‐> C6H12O6 + 6O2 + 6H2
But how do we convert the level of solar energy into the production of biomass? When looking at the
photosynthesis reaction to produce one carbohydrate (CH2O) we need 8 photons, one molecule of CO2 and
two molecules of H2O.
Not all wavelengths of light can be absorbed by the plant pigments, the reason being that the energy level
of the excited electrons has to match the energies of the reactions creating the chemical energy driving the
Calvin cycle. Thus the level of energy in the photosynthetically available region (PAR) of light is roughly 217
kJ mol‐1 of photons, accounting for 43% of the incident sunlight. The efficiency by which the energy from

11
the protons is transferred to electrons within the plant photosynthesis is very high and for simplicity we can
assume it to be 100%. With the single carbohydrate having 1/6 of the glucose energy content i.e. 467 kJ
mol‐1, the quantum limit of the photosynthetic efficiency is 11.6 %.
This value is the maximum theoretical level of efficiency by which plants are able to convert solar radiation
into biomass. In practice such a high level can never be realized as the plants need to divide part of the
energy to build and maintain their metabolic system, including the molecules in the photosynthetic centres
as well as many plants are dormant in the winter season.
However, if we assume that there is a 100% conversion of solar radiation into biomass, what is the
maximum level of biomass production in Denmark with an average solar radiation of 1000 kWh m‐2 on a
yearly basis? The answer can be seen in Table 3.3. These figures are in fact valid for any type of plant
regardless if it is a C3 or C4 type of photosynthesis, and cannot be exceeded no matter how much CO2 or
nutrients we apply.
Solar radiation PAR (GJ) Biomass (t/ha)
1000 kWh/m2 (Denmark) 15500 220
1500 kWh/m2 (Spain) 23200 330
2000 kWh/m2 (Sahara) 31000 439
Table 3.3. The levels of photosynthetic available light (PAR), and the corresponding theoretical maximum levels of biomass production assuming that 100% of the light is converted into biomass.
Again it should be stressed that the numbers here are the theoretical maximum assuming that all solar
energy is converted to biomass, which is of course not the case. In the literature figures above these
theoretical limits can sometimes be seen. Such figures should indeed be viewed with outmost caution as
they need to include excessive amounts of artificial lightning. Also production figures approaching the
theoretical possible maximum levels should be viewed with caution, as plants needs to devote a substantial
part of their energy to maintain respiration, synthesis of functional cell wall components as well as
proteins. An estimate of what level of practical biomass production that can possible be obtained in highly
efficient crops and algae systems under natural light conditions would be within 30‐50% of the theoretical
maximum, which is indeed still a very high productivity.
Can we improve the efficiency of photosynthesis?
Given the structure of the light capturing complexes in the photosynthesis and the matched coupling to the
reactions in the Calvin cycle, it will not be possible to increase the theoretical efficiency of photosynthesis
by which light energy can be converted to biomass above the theoretical maximum of 11,6%.
What will be possible is to increase the rate by which the Calvin cycle are capable of fixating CO2, as well as
the recovery rate due to the photoprotection of the reaction centres. Photoprotection is necessary as the
reaction centres of photosynthesis are continuously exposed to high energy radiation, and are on average
replaced every 20 min. Pulsating light sources can be used to reduce the need for photoprotection, but for
practical large scale applications this not likely to become economically feasible.

12
Improvement of photosynthesis will depend upon molecular engineering of the reaction centres as well as
breeding of crops or algae. It will require in the order of 10‐20 years before significant results can be
expected, but looking at the current potentials we can come a long way using the crops and algae that are
already available to us.
Recommended reading
Chrispeels MJ, Sadava DE 2003. Plants, genes and crop biotechnology. Jones and Bartlett Publishers,
Boston, MA.

13
4. The Chemistry of Algae
By looking at algae it is clear that, though they are photosynthetic organisms and thus by definition plants,
they are very different from land based plants. Not only by their physiology and morphology but also by
their chemical and physical composition ranging from molecular‐ to macro‐scale, it is evident that they
have followed a very different evolutionary path.
As with all other plants a main component in algae is carbohydrates (sugars). But on an overall basis the
chemical composition and structure of algae are considerably more heterogeneous than what is found
among land plants. The evolution path of algae has allowed for a far greater diversity (Domozych et al.
1980), and a large number of carbohydrates of different composition and organisation can be found.
Basically it appears that any carbohydrate in any conceivable combination can be found in algae cell walls,
and in many cases each species has a unique carbohydrate composition. This is indeed impressive, and
tough it adds to the complexity of utilizing algae for energy, it also underlines the potential for using algae
as chemical factories producing a number of organic chemicals or energy carriers.
Due to the large chemical diversity within algae a complete picture of the algae chemistry has not yet been
established. However, the diversity by itself is an important characteristic and also there some compounds
more common than others.
The bulk amount of biomass exploited for energy is based on the plant cell walls, but also storage
compounds such as starch and oils are of relevance for e.g. fermentation or extraction of the oils. The
following overall description is divided into the chemistry of algae cell walls storage compounds.
Algae cell walls
All plants have a need for structural rigidity of their cell wall in order to maintain their physical structure,
but depending on whether the plant has to function in an aqueous or atmospheric environment quite
different cell wall structures have evolved. Land based plants have as their adaptation to gravity developed
a sturdy cell wall, based on a combination of the polysaccharide pectin (herbs and young plants) and the
aromatic compound lignin (woody plants). Grasses as well as other plants use a combination of pectin and
lignin during their life cycle, where pectin is used in the young plants and lignin is build into the cell walls of
the mature plants in order to support the heavy kernels.
For algae, with the exception of diatoms which build their cell wall from silicic acid, the cell wall is based on
a wide range of carbohydrates and organic acids build into a polymeric matrix capable of forming aqueous
gels through ionic interactions and thereby providing the necessary structural rigidity.
Algae cell walls are thin and do not have a division in primary and secondary cell walls. Different cell wall
layers with different chemical composition can be seen in a number of algae, but there is no overall generic
structure as seen for land plants. Fig. 4.1 shows the chemical structure of sea lettuce (Ulva spp.), note the
organisation at the organelle level for the thallus as well as the cell wall. The ionic interaction between

14
acids and cations indicated in B at Fig. 4.1 is a common mechanism in algae cell walls for providing strength
and rigidity. The structure in Fig. 4.1 is only an example for sea lettuce, different structures can be found in
other algae.
Fig. 4.1. Distribution of the different Ulva sp. cell wall polysaccharides in a schematic cross section of a thallus (A) and proposed associations between the different cell wall polysaccharides (B). After Lahaye & Robic (2007).
Another distinct feature of algae cell walls is that they contain a high amount of protein compared to land
plants. The proteins in the algae cell walls are structural glycoproteins i.e. proteins with carbohydrate
oligomers linked to their polypeptide side chains. The glycoproteins are a structural part of the algae call
wall and may also serve as a cross linking matrix between cells in macroalgae. In some cases they even
constitute a larger part of the cell wall matrix than the carbohydrates, see Table 4.1.
Another common group of proteins in algae are the proteins in the photosynthesis complexes. Compared
to other plants they constitute a higher amount of the total biomass, and levels of more than 5% of the
total dry weight can be found.
The high protein content of algae could provide opportunities for preparing algae based protein feed
products, and such feed products are manufactured and sold on a small scale. But in general structural
proteins such as glycoproteins have a lower feed value and less favourable amino acid composition
compared to other plant proteins, and they may therefore be less suitable for animal feed.

15
Strain Protein Carbohydrates Lipids Nucleic acid
Scenedesmus obliquus 50‐56 10‐17 12‐14 3‐6
Scenedesmus quadricauda 47 ‐ 1.9 ‐
Scenedesmus dimorphus 8‐18 21‐52 16‐40 ‐
Chlamydomonas rheinhardii 48 17 21 ‐
Chlorella vulgaris 51‐58 12‐17 14‐22 4‐5
Chlorella pyrenoidosa 57 26 2 ‐
Spirogyra sp. 6‐20 33‐64 11‐21 ‐
Dunaliella bioculata 49 4 8 ‐
Dunaliella salina 57 32 6 ‐
Euglena gracilis 39‐61 14‐18 14‐20 ‐
Prymnesium parvum 28‐45 25‐33 22‐38 1‐2
Tetraselmis maculata 52 15 3 ‐
Porphyridium cruentum 28‐39 40‐57 9‐14 ‐
Spirulina platensis 46‐63 8‐14 4‐‐9 2‐5
Spirulina maxima 60‐71 13‐16 6‐7 3‐4.5
Synechoccus sp. 63 15 11 5
Anabaena cylindrica 43‐56 25‐30 4‐7 ‐
Table 4.1 ‐ Chemical Composition of some microalgae. After Becker (1994).
Algae carbohydrates
As in other plants, carbohydrates are the most common single component found in algae, tough the ratio
of carbohydrates is lower compared to land plants. But unlike most land plants marine algae with a few
exceptions do not contain simple carbohydrates or easily hydrolysable polysaccharides. Starch and cellulose
are only present in minor quantities.
Algae carbohydrates may be linear, but are often derivatized with acids or sulphate groups. These
structural characteristics of algae carbohydrates means that biochemical conversion of algae is not a
straightforward task, as no known industrial yeast strains are capable of fermenting the major algae
carbohydrates. Below is a description of some of the most common carbohydrates found in algae, but it
must be emphasized that they only represent a small number of the carbohydrates that can be found in
algae cell walls.
Mannanes
In a number of marine green algae as well as in the walls of some red algae the C6 mannanes are main
structural elements. They can be both branched and linear and the mannosyl residues are 1 > 4
glycosidically linked. The supramolecular structure of mannanes often form fibrils in a partly crystaline
organisation similar to cellulose. The mannanes may be tightly associated with the cell wall glycoprotein.

16
Cellulose
The glucose based C6 polymer of cellulose is present in a number of both green brown and red algae. The
amount is typically below 10%. The low amount of cellulose compared to land plants may be caused by the
fact that algae are exposed to a lower level of tensile stress. The fibrillar structure is important for the
physical reinforcement of the cell walls.
Ulvan
A typical representative of the complex carbohydrates found in algae is ulvan grom the genus Ulva also
known as Sea lettuce. The carbohydrate polymer constitute up to 30% of the cell wall in Ulva and is made
from a number of C5 and C6 carbohydrates including; rhamnose, xylose, glucose, uronic acid and glucoronic
acid, where rhamnose and glucoronic acid forms the complex aldobiouronic acid. Often the rhamnose has a
sulphate group and the linkage between the individual carbohydrate monomers vary. Thus the complexity
of ulvan is impressive, but it also shows the challenge in utilizing algae cell wall components for large scale.
Fucans
In brown algae the sulphated fucan polysaccharides are common. A number of structural variations can be
found between the species. The structure is based on the C6 carbohydrate fucose which is sulphonated and
forms the polymeric backbone. Different branches of galactose and acetate side groups can be found as
part of the fucan structure. Fucans are known to have therapeutic properties for blood coagulation or
inflammation.
Xylanes
Xylanes are polymers made from C5 sugars where the xylosyl residues are linked via 1 > 3 and 1 > 4
glycosidic bonds. Xylans are partly branched. As a rule of thumb layered cell walls can found in xylan
containing algae.
Alginic acid
Alginic acid and its salts, the alginates are components of the walls of brown algae. They consist of uronic
acids: mannuronic acid and glucuronic acid in varying ratios. The alginates of brown algae exist both within
the cell wall and in the intercellular substance. Their part in the cell wall may be as high as 40 per cent of
the dry matter. They have a high affinity for divalent cations (calcium, strontium, barium, magnesium)
which are important for their tendency to gel.
Sulfated carbohydrates
A distinct feature of algae is that they often contain sulfated carbohydrates. That is carbohydrates with –
SO3 groups which have been found in nearly all marine algae. They occur partially in the cell wall itself and
partially in the intercellular substance. Sulfonated galactanes are typical for many red algae, depending on
their origin they are named agarose, porphyran, furcelleran and funoran.

17
Intracellular carbohydrates
In both green and red algae starch is a common metabolic energy storage compound. In a few cases such as
Chlorella vulgaris a starch content of up to 37% has been reported, but normally lower contents are found.
Mannitol and laminarin are storage carbohydrates found in brown algae. During the summer season they
may constitute up to 40% of the dry matter.
Algae oils
Like other vegetable oils or lipids, the algae oils are composed of triglycerides i.e. glycerol esterified with
fatty acids. When processed to biodiesel the glycerine is removed by de‐esterification.
Just as for land plants, triglycerides in algae are energy storage compounds. But there is an important
difference in the metabolism of algae as they produce the highest content of lipids when exposed to
nutrient stress. Unfortunately this also means that high content of lipids correspond to low growth rates,
which is unfavourable for large scale production. Algae oils are high in unsaturated fatty acids. The most
commonly found oils in algae are:
Arachidonic acid
Eicospentaenoic acid
Docasahexaenoic acid
Gamma‐linolenic acid
Linoleic acid
Inorganic elements
With the exception of grasses algae has a high level of inorganic elements compared to land based plants.
The most common elements found in algae are; calcium, potassium, sodium, phosphorous, sulphur,
chlorine and magnesium. The most elements are calcium and sodium which are typically found at levels of
1‐3%.

18
5. Microalgal production for bioenergy
From an application point of view microalgae can be described as a single cell photochemical factory. They
can be used to produce not only biomass for energy but also, pharmaceuticals, nutraceuticals, pigments
(e.g. Dunaiella, Fig. 5.1) etc.
Fig. 5.1. From left to right: Dunaliella salina, a natural bloom of D. salina and cultivation of D. salina (for natural beta carotene) in Western Australia.
The overwhelming interest in microalgae is due to the high lipid content of some species, and that the lipid
synthesis, especially of the non‐polar triacylglycerols (TAGs), which are the best substrate to produce
biodiesel, can be modulated by varying growth conditions. The total content of lipids in microalgae vary
from about 1‐85% of the dry weight (Rodolfi et al. 2009, and references herein) with values higher than
40% typically achieved under nutrient limitation.
The potential has been heavily investigated during the 1980 and 1990’s dominated by the North American
Aquatic Species Program headed by the National Renewable Energy Laboratory (NREL) (Sheehan et al.
1998), which was closed by the Clinton government). Now, ten years later, the interest in microalgae for
bio‐diesel is peaking and research institutes as well as private companies are allocating resources to this
area as the benefits of this renewable energy seems obvious, as according to Rodolfi et al. (2009, and
references herein):
Oil yield per area of microalgae cultures could greatly exceed the yield of the best oilseed crops
Microalgae grow in an aquatic medium, but need less water than terrestrial crops
Microalgae can be cultivated in seawater or brackish water on non‐arable land, and do not
compete for resources with conventional agriculture
Microalgae biomass production may be combined with direct bio‐fixation of waste CO2
Fertilizers for microalgal cultivation can be obtained from wastewaters
Algae cultivation does not need herbicides or pesticides
The residual microalgal biomass after oil extraction may be used as feed or fertilizers, or fermented
to produce ethanol or methane
The biochemical composition of the microalgal biomass can be modulated by varying growth
conditions and the oil contents can be highly enhanced
Regardless the technological and biological advantages the question is if at present is possible to produce
microalgal biodiesel at costs competitive with petroleum derived diesel?

19
Christi (2007, 2008) claims that it is possible to produce feasible algae source bio‐diesel. He finds a gap in
prices of petroleum and algae source diesel of a factor 9 and 7, respectively, according to crude oil prices,
but still consider it attainable due to optimization of yield and production chain. However, Borowitzka
(2008) presents a gap of a factor 5, but considers it to be impossible to close this gap as production price is
interconnected with energy price.
The interest for using microalgae for bio‐diesel production is large. There are several companies whom
offer algae growing systems (AlgaeLink, GreenFuel); Carbon Trust in UK launched late 2008 the by then
largest algal biofuel project (http://www.carbontrust.co.uk/technology/directedresearch/algae.htm);
Seambiotic claims to be the first company that is utilizing flue gas from coal burning power stations for
algae cultivation. Seambiotic (http://www.seambiotic.com/) was founded in 2003 with the objectives to
grow and process marine microalgae for profitable exploitation of microalgae in two areas, bio‐fuels and
food additives; Bill Gates invests in algal‐based biofuels (http://www.sapphireenergy.com/)!
To evaluate the potential of a feasible production of microalgae for bio‐diesel in Denmark we need to look
through both hype as well as technology pessimism:
1. Which species or strains of species of microalgae will be suitable for Danish conditions, considering
growth rates and lipid yields and thereby the actual lipid production
2. Can the production be optimized to meet the production cost of petroleum source fuels
3. Can microalgal mass cultures serve as flue gas bioremediation and reduce the microalgal biomass
production costs
Potential species
Few works have focused on identifying the most suitable microalgal species or strains for oil‐production.
The most comprehensive work being that of the North American Aquatic Species Program (Sheehan et al.
1998), but more recently a nice work of Rodolfi et al. (2009) was published on species and strains tested in
Italy.
It is well known that strains of a species might perform very different according to growth rate and lipid
contents under different conditions (Hu et al. 2008, Rodolfi et al. 2009, Sheehan et al. 1998). The list of
promising species and their strains are available in Sheehan et al. (1998). More than 3000 strains were
collected whereas 300 strains remain in the collection. From this and other works (Christi 2007, and
references herein, Li et al. 2008, and references herein, Rodolfi et al 2009) the species listed in Table 5.1
seem to be the most promising candidates for mass cultivation and lipid production. In the original 1984‐
1985 Microalgae Culture Collection Catalogue (SERI 1984) the 10 highest lipid yielding species were listed,
of which 7 are on the list of microalgae accumulating lipids by Christi (2007). In the same work Christi lists
further 7 species with high oil contents. All species have been evaluated for lipid production and a few of
them are commented on below:

20
Cryptothecodinium cohnii (Fig. 5.2) is a colourless marine dinoflagellate, which means that it is a
heterotrophic organism. It will be treated below in connection with microalgae not obligate autotrophic.
In Christi (2007) Schizochytrium sp. (Fig. 5.2) is listed as an oil containing microalgae, but this organism is a
thraustochytrid, a microbe which is a heterotrophic marine fungi. It is sometimes referred to as a marine
alga, perhaps because of reproduction by zoospores? As Schizochytrium sp. is obligate heterotrophic as well
as not being an alga it is not included in this evaluation.
Fig. 5.2. From left to right; Cryptothecodinium cohnii, Schizochytrium sp.
Of the species listed in Table 5.1, 9 are registered in Denmark (Thomsen 1992, Hällfors 2004) and therefore
are potential species for cultivation under Danish environmental conditions.
In Table 5.1, when Christi (2007) is used as reference, the numbers are homologous to numbers
corresponding to Sheehan et al. (1998).
Growth rate/lipid content
Under optimal conditions of growth, algae synthesize fatty acids principally for etherification into glycerol‐
based membrane lipids, while under suboptimal or stress conditions for growth many algae alter their lipid
biosynthetic pathways towards the formation and accumulation of neutral lipids mainly in the form of TAGs
(Hu et al. 2008). TAGs composed primarily of saturated and monounsaturated fatty acids can be efficiently
packed into the cell and generate more energy than carbohydrates upon oxidation, thus constituting the
best reserve for rebuilding after the stress (Rodolfi et al. 2009, and references herein).
Therefore, as very high lipid production usually is correlated with stress conditions (nutrient deprivation)
(Chiu et al. 2009, Rodolfi et al. 2009, Sheehan et al. 1998), this results in decreased photosynthetic
efficiency and decreased growth. Thus the two conditions of high lipid content and high biomass
productivity are mutually exclusive. In general, productivity and lipid content is inversely related, a fact that
further has its rationale in the high metabolic cost of lipid synthesis (Rodolfi et al. 2009).
The best lipid producers are the strains showing the best combination of biomass productivity and lipid
content (Rodolfi et al. 2009). However, often only data of the lipid contents are published as summed in
Table 5.1. It is quite difficult to obtain published data on growth rates obtained in the same study as lipid
contents, which would give the true picture of the lipid productivity potential of a species or strain.
Apparently more data of this kind is in press, for example on Nannochloris oleoabundans, as presented on
the XIth International Conference on Applied Phycology in Galway, 2008 (Fouchard et al. 2008).

21
Species Phylum/Class Lipid yield(% DW)
References F M
DK
Botryococcus braunii Chlorophyta 44‐54 Banerjee et al. 2002, Ben‐Amotz et al. 1985, Christi 2007, Sheehan et al. 1998
F •
Chlorella spp. Chlorophyta 22‐63 Christi 2007, Illman et al. 2000, Liu et al. 2008
F •
Cylindrotheca sp. Bacillariophyceae 16‐27 (37*) Ying et al. 2002, *Christi 2007
M
Dunaliella primolecta Bacillariophyceae 9‐25 Ben‐Amotz et al. 1985, Christi 2007
M •
Isochrysis sp. Prymnesiophyta 7‐33 Christi 2007, Ben‐Amotz et al. 1985, Sukenik & Wahnon 1991
M •
Monallanthus salina
Chlorophyta 21‐22 (41‐72)
Sheehan et al. 1998, Christi 2007, (Shifrin et al. 1981)
M
Nannochloris sp. Chlorophyta 20‐36 Ben‐Amotz et al. 1985, Christi 2007
M •
Nannochloropsis sp. Eustigmatophyta 31‐60 (68*) Chiu et al. 2009, *Christi 2007, Rodolfi et al. 2008
M •
Neochloris oleoabundans
Chlorophyta 35‐54 Christi 2007 F?
Nitzschia sp. Bacillariophyceae 22‐47 Ben‐Amotz et al. 1985, Christi 2007
M •
Phaeodactylum tricornutum
Bacillariophyceae 20‐30 Christi 2007 M •
Tetraselmis suecica Prasinophyta 9‐15 (23*) Rodolfi et al. 2009, *Christi 2007
M •
Table 5.1. List of selected species for cultivation due to relatively high lipid contents. *It has not been possible to verify
the lipid yield stated by Christi (2007, table 2). F: freshwater, M: marine.
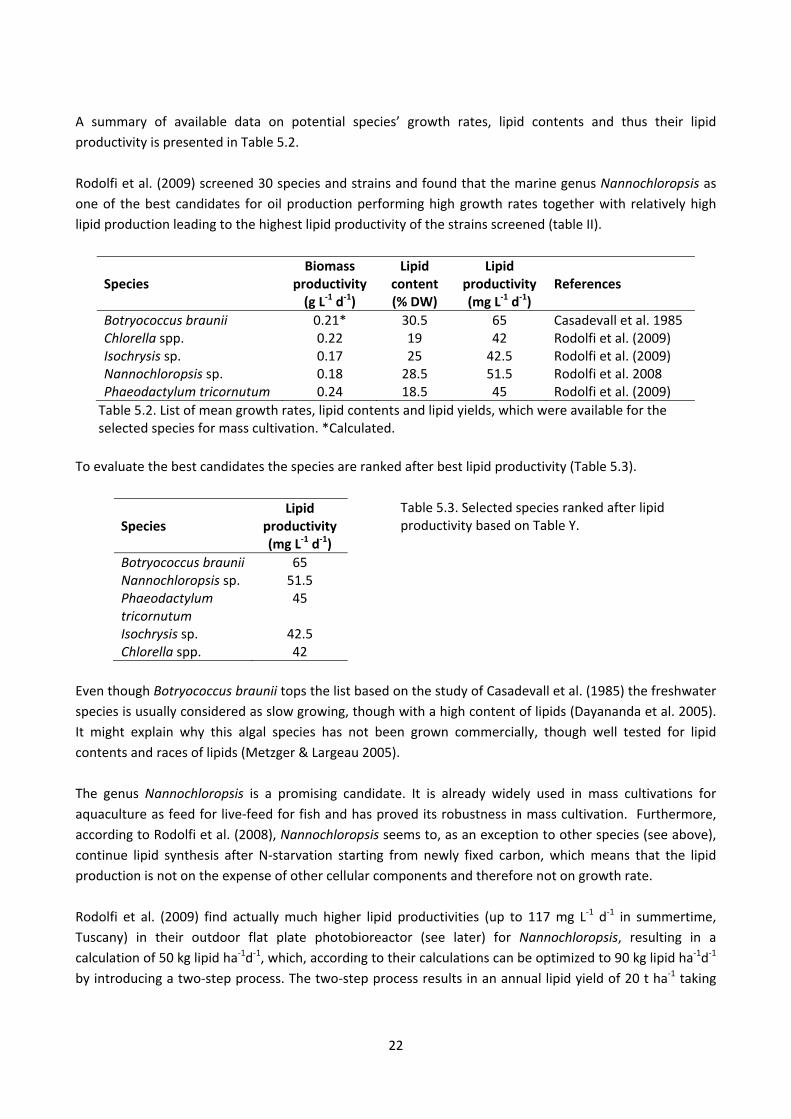
22
A summary of available data on potential species’ growth rates, lipid contents and thus their lipid
productivity is presented in Table 5.2.
Rodolfi et al. (2009) screened 30 species and strains and found that the marine genus Nannochloropsis as
one of the best candidates for oil production performing high growth rates together with relatively high
lipid production leading to the highest lipid productivity of the strains screened (table II).
Species Biomass
productivity (g L‐1 d‐1)
Lipid content (% DW)
Lipid productivity (mg L‐1 d‐1)
References
Botryococcus braunii 0.21* 30.5 65 Casadevall et al. 1985 Chlorella spp. 0.22 19 42 Rodolfi et al. (2009) Isochrysis sp. 0.17 25 42.5 Rodolfi et al. (2009) Nannochloropsis sp. 0.18 28.5 51.5 Rodolfi et al. 2008 Phaeodactylum tricornutum 0.24 18.5 45 Rodolfi et al. (2009)
Table 5.2. List of mean growth rates, lipid contents and lipid yields, which were available for the selected species for mass cultivation. *Calculated.
To evaluate the best candidates the species are ranked after best lipid productivity (Table 5.3).
Species Lipid
productivity (mg L‐1 d‐1)
Botryococcus braunii 65 Nannochloropsis sp. 51.5 Phaeodactylum tricornutum
45
Isochrysis sp. 42.5 Chlorella spp. 42
Table 5.3. Selected species ranked after lipid productivity based on Table Y.
Even though Botryococcus braunii tops the list based on the study of Casadevall et al. (1985) the freshwater
species is usually considered as slow growing, though with a high content of lipids (Dayananda et al. 2005).
It might explain why this algal species has not been grown commercially, though well tested for lipid
contents and races of lipids (Metzger & Largeau 2005).
The genus Nannochloropsis is a promising candidate. It is already widely used in mass cultivations for
aquaculture as feed for live‐feed for fish and has proved its robustness in mass cultivation. Furthermore,
according to Rodolfi et al. (2008), Nannochloropsis seems to, as an exception to other species (see above),
continue lipid synthesis after N‐starvation starting from newly fixed carbon, which means that the lipid
production is not on the expense of other cellular components and therefore not on growth rate.
Rodolfi et al. (2009) find actually much higher lipid productivities (up to 117 mg L‐1 d‐1 in summertime,
Tuscany) in their outdoor flat plate photobioreactor (see later) for Nannochloropsis, resulting in a
calculation of 50 kg lipid ha‐1d‐1, which, according to their calculations can be optimized to 90 kg lipid ha‐1d‐1
by introducing a two‐step process. The two‐step process results in an annual lipid yield of 20 t ha‐1 taking

23
low/no production periods due to the varying solar irradiance in the Tuscany area into account. They
estimate a potential annual lipid production of 30 t ha‐1 for the tropics (for detailed information, see Rodolfi
et al. 2009, p. 110).
Nannochloropsis is already available in culture in Denmark. See Scenario for Microalgae Production for an
estimation of lipid yield under Danish conditions.
Mass cultivation
Ugwu et al. (2008) present a review on photobioreactors for mass cultivation of microalgae. They include
following photobioreactor systems:
Open ponds
Tubular photobioreactors
Vertical‐column photobioreactors
Flat‐plate photobioreactors
Raceway ponds are widely used as an open pond system since it is easy to operate. Production of
microalgae biomass has been extensively evaluated in raceway ponds by United States Department of
Energy (Sheehan et al. 1998).
Closed photobioreactors were also recommended for scaling up production of autotrophic microalgae since
this kind of bioreactor could save water, energy and chemicals compared to some other open cultivation
systems (Peer et al. 2008). The most widely used closed photobioreactors are designed as tubular reactors
or plate reactors as will be presented below.
A new “technology” has recently been presented at the International Workshop on Offshore Algae for
Biofuels and Beyond, April 2009, Lolland, Denmark, which is sea‐based cultivation of marine species in sacs
anchored to wind‐mill foundations. The technology has not been tested but theoretical pros and cons of
this land‐saving technology will be discussed together with the more established and tested cultivation
technologies.
Open ponds (Christi 2007, Lee 2001, Li et al. 2008; and references herein)
Raceway‐shape culture ponds are used in Israel, USA, China and other countries. Generally, the structure of
the ponds is almost the same as the sewage treatment system (Fig. 5.3). Fertilizer is used and the culture is
agitated by paddle wheel (Fig. 5.3).
The advantages of open ponds are low costs and that they are easy to operate. However, they are sensitive
to contamination leading to introduction of unwanted fast growing organisms in the ponds. Especially
heterotrophic organisms will graze on the autotrophic biomass and lead to loss of productivity. Therefore
the present commercial production of microalgae in open culture systems is restricted to only those
organisms that can grow under extreme conditions i.e. high pH or salinity. Thus a limited range of
microalgae can be maintained as monoculture in open ponds in long‐term operation. Presently, only

24
Dunaliella in high salinity, Spirulina in high alkalinity and Chlorella in high levels of nutrients have been
successfully mass cultured and marketed commercially. To overcome the problem of infections in the open
pond systems the solution may be to mass cultivate mixotrophic organisms that can “clean up” themselves
(se Mixotrophic organisms).
Another major barrier is harvesting of the microalgae biomass due to its dilute form in open ponds (see
Harvest).
Fig. 5.3. At left; outline of a raceway open‐pond (after Li et al. 2008), and above and right, examples of plants.
Photobioreactors
According to the consensus on which photobioreactors systems to be most widely investigated (IVXth
Congress on Applied Phycology, Galway, April 2008) and most recent publications, the applicable
photobioreactor systems are, the tubular and flat plate designs.
Tubular photobioreactor system (Christi 2007, Li et al. 2008; and references herein)
A tubular photobioreactor is usually equipped with fencelike solar collectors. Microalgal broth is
continuously pumped through the solar array, where sunlight is absorbed. Fresh culture medium is fed
continuously to the degassing column during daylight, and an equal quantity of broth is harvested at the
same time. The degassing column is continuously aerated to remove the oxygen accumulated during
photosynthesis and the oxygen‐rich exhaust gas is expelled from the degassing column.
Photosynthesis generates oxygen. A high concentration of dissolved oxygen in combination with intense
sunlight produces photooxidative damage to algal cells. To prevent inhibition and damage, the oxygen
needs to be removed through a degassing column as the oxygen cannot be removed within the
photobioreactor tube.
As the broth moves along the photobioreactor tube, pH increases because of consumption of CO2. Feeding
of CO2 to the reactor is done in the degassing zone in response to a pH controller. Additional CO2 injection

25
points may be necessary at intervals along the tubes, to prevent carbon limitation and an excessive
increase in pH. A typical tubular photobioreactor is designed as in Fig. 5.4.
Fig. 5.4. Typical outline of tubular photobioreactors, right. After Li et al. 2008. For explanation, see text.
Flat plate technology
The flat plate photobioreactor is a vertical reactor made of transparent plates glued together to form a
500‐1000 L unit (Cheng‐Wu et al. 2001), and elaborated by Rodolfi et al. (2008) to the patented “green wall
panel” (GWP)) (Fig. 5.5).
The GWP comprises a culture chamber made of 0.3‐mm thick flexible LDP film enclosed in a rectangular
metal frame. The modules are 1 m high, 2.5 m long and ca 4.5 cm thick resulting in a culture volume of 110
L. For mixing, compressed air was bubbled at the bottom of the reactor through a perforated plastic tube.
CO2 was injected into the culture through a gas diffuser placed in an un‐aerated zone, as carbon source and
for pH regulation. A control unit provided temperature regulation of the cultures by automatically
activating water spraying on the reactor surface, when temperature exceeded the pre‐set value.
Fig. 5.5. Flat plate (green wall panel (GWP)). After Rodolfi et al. 2008. For explanation, see text.
In general, for the above described photobioreactors, the advantages are no contamination and controlled
production parameters but the design is expensive and will also require cooling.

26
Alternative designs ‐ OMEGAs (Trent, WSA (2009))
This technology is ocean mega enclosures of semi‐permeable membranes for growing algae (OMEGAs) in
freshwater (Fig. 5.6). The idea is to fill the enclosures with nutrient rich freshwater (sewage?) and grow
microalgae species which are not supposed to survive if leaked to the surrounding marine environment.
The idea is also that the osmotic force by placing the OMEGA in sea with freshwater content can be used
for concentrating the biomass.
It is a land‐saving technology for microalgal cultivation, and is also bioremediative in case of using sewage
as growing medium, as suggested.
However, risk of leakages is the major concern for this technology, regarding both introductions of invasive
species as well as sewage to the environment. The technology is as yet not sufficient developed concerning
robustness and especially mixing of the internal content for nutrient and light distribution. As the OMEGA is
an alien to the environment, prototypes have been attacked by sea gulls as well as marine animals
(dolphins, sharks) (McGillivary, WSA 2009).
Fig. 5.6. Principles of the OMEGA system. An example of an offshore membrane enclosure as a fresh water reservoir for cultivating freshwater algal species, which will take up nutrients from treated wastewater and be harvest by concentrating the algal biomass by osmosis.

27
Comparison of the cultivation technologies
Cultivation of microalgae using open‐ponds is technologically simple, but not necessarily cheap due to the
high downstream processing costs. The need to achieve higher productivity and to maintain monoculture
of algae led to the development of enclosed tubular or flat plate photobioreactors. Despite higher biomass
concentration and better control of culture parameters, data accumulated in the past 30 years have shown
that the illuminated area, volumetric productivity and cost of production in these enclosed
photobioreactors are not better than those achievable in open‐pond cultures (Lee 2001).
Christi (2007, 2008) provides calculations for comparison of photobioreactor and raceway production
methods (Table 5.4). His main conclusions from these calculations are that a larger area is needed with the
open pond system (7828 vs. 5681 m2, 38%) and the oil yield is correspondingly lower per area (99.4 vs.
136.9 m3 ha‐1). He does not, though, credit the data or indicate which species are considered.
Table 5.4. Comparison of photobioreactor and raceway production methods. Based on afacility area, bactual pond area, cprojected area of photobioreactor tubes, d70% by wt oil in biomass, e30% by wt oil in biomass. After Christi (2007).
Rodolfi et al. (2009) achieved very good results on cultivation of the genus Nannochloropsis in flat plate
reactors, but they did not perform either a practical (harvest, cleaning) or an economical evaluation on this
production system.
However, Rodolfi et al. (2008) presented a comparison of the GWP and open pond systems. They found
that the GWP technology was severely hampered by a low net energy ratio (NER). The high‐embodied

28
energy of the reactor and the high‐energy expenditures for mixing, algae harvesting and dewatering
appeared difficult to reduce in a significant way. The presented preliminary life cycle analysis indicated that
raceway ponds, requiring no energy for cooling and a much lower energy input per unit area for mixing,
have much more favourable NER. However, they consider algae ponds to be very unstable ecosystems,
which suffer from the impossibility to maintain the desired organism in culture long enough. Therefore,
they proposed a strategy combining reactors and ponds with the aim to achieve both positive NER and the
process reliability necessary for cost‐effective production.
In Table 5.5 a summary of prospects and limitations of various culture systems for microalgae is presented
as considered by Ugwu et al (2008). Ugwu et al. (2008) also include the main issues of the photobioreactor
systems, the need for cooling.
Table 5.5. Prospects and limitations of various culture systems for microalgae. After Ugwu et al. (2008).
The temperature problem is logically interconnected with the need for maximal solar irradiance, but this
will increase the cooling requirements. At present there is no cooling technology available that will make
possible to achieve a high NER.
Harvesting
Efficient harvesting of microalgae is one of the major factors to be overcome in order for them to be used
as a fuel source. According to Benemann & Oswald (1996) the problem is that microalgae mass cultures are
dilute, typically less than 500 mg L‐1 on a dry weight organic basis, and the cells are very small, like the
genus Nannochloropsis with cell sizes of2‐5 um. In order to extract and process the lipids into bio‐diesel the
algae must be available at 15% solids, and in raceway ponds, for example, there is a 1% solids level
(Benemann & Oswald 1996). Therefore the culture must go through a process, which will result in a
sufficient concentration of the algae biomass.
Many different algae harvesting processes have been studied. In Table 5.6 technologies is compared.

29
Table 5.6. Comparative evaluation of harvesting technologies. After Benemann & Oswald 1996.
Centrifugation
The algae pond solution is pumped into a large centrifuge, which rotates at several thousand RPM and has
a filter of few microns. This is a proven method but has high cost (Table 5.6).
Chemical flocculation
Chemicals like lime, alum or chitosan can be added to the algae pond solution flocculating the algae.
However, this technology also has relatively high costs associated with the large amounts of chemicals
required.
Settling ponds
A raceway open ponds system may include settling ponds. The settling ponds may serve as the initial
harvesting method, where settling of the microalgae will bring the solution to 3% algae biomass. After the
settling ponds one of the above technologies then will need to be applied, but use of the settling ponds will
reduce costs accordingly.

30
Recommended reading
Hu Q, Sommerfeld M, Jarvis E, Ghirardi M, Posewitz M, Seibert M, Darzins A. 2008. Microalgal triacyl‐
glycerols as feedstocks for biofuel production: perspectives and advances. The Plant Journal 54: 621‐639.
Rodolfi L, Zittelli GC, Bassi N, Padovani G, Biondi N, Bonini G, Tredici MR. 2009. Microalgae for oil: strain
selection, induction of lipid synthesis and outdoors mass cultivation in a low‐cost photobioreactor.
Biotechnology and Bioengineering 102: 100‐112.

31
6. Possibilities for improvement of microalgal production systems
Yield optimization of photosynthetic microalgal production systems
Improving the yield of a given photosynthetic algae species provided equal input of energy and nutrients
will improve the economy of microalgae biodiesel accordingly. According to the rapport on the Aquatic
Species Program (Sheehan et al. 1998) a technical evaluation identified two major requirements for
economic feasibility of algal biodiesel production:
1. Photosynthetic efficiency, i.e. the percentage of incident radiation converted into biomass, needs
to be 18%, which is above the theoretical maximum for photosynthesis efficiency of approximately
12%.
2. Algal biomass needs to consist of 60% lipid
At present we cannot meet these requirements. Suggestions to improve performance of microalgae for
biomass and lipid production is presented and discussed below.
Photosynthetic efficiency
Flashing light
Studies have shown that intermittent light increased the rate and efficiency of photosynthesis compared to
the energy utilization efficiency under constant illumination (Grobbelaar 2008, Grobbelaar et al. 1996,
Nedbal et al. 1996, Terry 1986). The shift between light and dark speeds up the rate of photosynthesis as
the enzymatic process of CO2 reduction to photosynthetic intermediate products is accomplished in the
dark, and the chlorophyll is set free to take part again in the photochemical reaction (Emerson & Arnold
1932a, b).
This flashing light effect can be mimicked in vertically mixed algal ponds, as cells circulate to the surface
and back down to the lower levels in the pond thereby creating a transition between the photic and
aphotic layers. Thus, the photosynthetic efficiency of algal grown in ponds and photobioreactor designs
may be increased in high light by using mixing strategies that optimize the effects of an intermittent light
regime (Grobbelar 1996, Nedbal et al. 1996, Sheehan et al. 1998).
Light adaptation
Can adapting microalgal cells to lower light regimes optimize photosynthetic yield? The historical
background on adaptation of algae to various light intensities is that it is argued that there is little evidence
to suggest that previous growth at low light intensities enhances the ability of an alga to utilize these low
light levels (Beardall & Morris 1976, and references herein) and may even result in lowered maximum levels
of photosynthesis at light saturation (Pmax). However, both Beardall & Morris (1976) and Neale & Melis
(1986) do find an enhanced photosynthetic efficiency in Phaeodactylum tricornutum and Chlamydomonas
reinhardtii, respectively, adapted to low light regimes. According to Neale & Melis (1986, and references

32
herein) most species of algae increase the chlorophyll cell‐1 under low light conditions. In their study of
Chlamydomonas reinhardtii they found that the photosynthetic performance of cells grown in low light (LL
cells) was higher in low light regimes than cells grown in high light (HL cells) indicating the benefit, which
the algae derive from the adaptation of their photosynthetic apparatus. Overall they concluded that LL cells
had a greater ability for photosynthesis at low light, but maximum photosynthetic capacity per cell was not
lower.
In connection with mass cultivation of microalgae it was suggested that this light adaptation ability was
utilized to optimize photosynthetic production in connection with photobioreactor designs (Weismann
2008), as well as the photosynthetic production may be higher than expected when culturing algae in areas
with lower solar radiation (Tredici 2008).
Cell engineering
According to Rodolfi et al. (2009), considering genetic and metabolic engineering of microalgae, it would be
prudent to limit projections to what can be achieved with wild‐type strains for the foreseeable future as
they state that: “It seems likely that among the thousands of microalgal species and billions of strains that
colonize almost every photic niche of the earth, many organisms suitable for outdoor biomass culture and
biofuel production might be found. Thus there is no apparent need to genetically modify microalgae to
achieve the immediate objectives of such a process, [which] is stable mass cultures exhibiting relatively
high oil contents and productivities.”
Eliminating photosaturation and photoinhibition
Christi (2007) finds cellular engineering useful in many respects, including enhancing biomass growth rate,
oil contents in biomass, improving temperature tolerance, but focuses on eliminating photosaturation and
photoinhibition, i.e. rising Pmax by a delayed onset of photosaturation and increasing the “window” of Pmax
by a higher tolerance to high light intensities, delaying the onset of photoinhibition (see also Chapter 2,
Photosynthesis).
Cell engineering focusing on eliminating photosaturation and photoinhibition by cell engineering, Rodolfi et
al. (2008) also expect can significantly increase productivity of outdoor cultures and greatly improve the
economics of microalgal oil production. But they also take into account that it will require long‐term
research and funding, and, in many countries, overcoming current strictures against the release of
genetically modified organism. And, it must be recognized, that improving the photosynthetic efficiency by
eliminating photosaturation and photoinhibition will require a major redesign of the very advanced
photosynthetic molecular complex, which we even today only partially understand.
Reducing antenna size
Another approach to optimizing photosynthesis is reducing antenna size. The actual photochemistry in the
reaction centres of the two photosystems involves only two or three chlorophylls but the membranes
contain many more. These additional chlorophylls together with other accessory pigments (phycobilisomes,
carotenoids) are known as antenna pigments because they substantially increase the likelihood that
sunlight falling on the organism will be captured and transferred to the photosynthetic apparatus (Prince &
Kheshgi 2005, and references herein). These very large antenna systems collect far more photons than can

33
be used by the reaction centres. Therefore efforts are put into manipulating the system to decrease the
antenna size to achieve higher yields and photosynthetic efficiencies by a smaller energy budget (Prince &
Kheshgi 2005, Grobbelaar 2008). Melis et al. (1999) showed that the green alga, Dunaliella salina, with
manipulated small antenna sizes exhibited higher photosynthetic productivities and photon use efficiencies
than normally pigmented cells.
In contrast to Rodolfi et al (2008), Prince & Khesghi (2005) philosopher about evolution concerning
photosynthetic efficiency and find that natural selection “unfortunately” not seems to have selected for
maximal energy usage or efficiency! They write:” We do not understand why natural selection has resulted
in a system that is optimized for maximal photon use rather than maximal energetic efficiency.” One reason
could be that within large algae populations, the algae, which are not directly light exposed at the surface,
are limited by suboptimal light levels, thus favouring apparently oversized photon capturing systems.
It should be noted that none of the methods and ideas described above would overcome the limitation in
photosynthetic efficiency caused by the level of energy present in energy that can be transferred from the
photons to the photosynthetic system. Therefore the maximum efficiency of the photosynthesis cannot be
higher than 11.8% of the incident solar radiation at a given surface (see Chapter 2 on photosynthesis).
Temperature
As temperature regulation, in the form of cooling or heating will add significant costs to the production of
microalgal biomass it has been argued that the species/strain selected for production will need to be
temperature tolerant in a wide range relevant for the actual climate (Christi 2007).
The typical response is for photosynthesis to increase progressively with increasing temperature, with a Q10
(temperature coefficient for ratio of rate at T°C to that of T + 10°C) of approximately 2 (Davison 1991).
However, if the temperature exceeds the temperature range tolerated by the organism the growth rate will
decrease with increasing respiration/photorespiration.
Therefore the local temperature and the range of optimal/tolerated temperatures of the selected
species/strain must walk and in hand. The solar irradiance crucial for photosynthesis may also elevate the
temperature into non‐tolerated temperature regimes for the cultivated algae; especially this may be a
problem in photobioreactors. Species and strains with wide temperature tolerance ranges are thus
desirable for mass cultivation (Christi 2007).
Mass culturing microalgae in temperate climates may diminish this problem but will have to compensate
for less solar irradiance. One should also bear in mind that species from temperate climates are adapted to
lower temperatures and the elevated temperatures in ponds with low water levels as well as
photobioreactors may also well be outside the range of their temperature tolerances.
On the other hand low temperatures will make processes go to slow resulting in low growth rates.

34
Harvest timing
To overcome the inverse relation between growth rate and lipid yield an experiment was conducted as part
of the Aquatic Species Program (Sheehan et al. 1998) to help understand the effects of nitrogen deprivation
and lipid accumulation on photosynthetic efficiency. The results showed that N starvation triggered the
lipid accumulation after the culture was grown to a certain density and reaching the maximum of
photosynthetic efficiency. Therefore, according to Sheehan et al. (1998) summing up the results of Lien &
Roessler (1986), to obtain maximal lipid yield the best time for harvesting occurs just after the N is depleted
from the cultures.
Mixed cultures of algae and bacteria
Cole (1982) reviewed the interaction between bacteria and algae in aquatic systems and concluded that
laboratory evidence strongly suggested that a zone existed within which bacteria and algae interact in both
stimulatory and inhibitory ways. Banerjee et al. (2002) also include this matter in their review of
Botryoccocus braunii as a renewable source of hydrocarbons. They list a number of impacts of bacteria
presence on algal cultures; stimulation of algal growth by releasing substances (vitamins, organic chelating
agents), remineralisation of inorganic nutrients as well as influence on pH and redox potential by CO2
production. They conclude that: “In many cases, stable association [of algae and bacteria, eds] will provide
a large increase in algal biomass and hydrocarbon production, a reduced contamination by other
microorganisms, and an improved supply of CO2.”
However, the choice of bacteria culture will be important as some bacteria cultures may provide inhibitory
effects (Banerjee et al. 2002, Cole 1982, and references herein), i.e. Pseudomonas oleovorans caused a
marked reduction in algal biomass and hydrocarbon production in Botryococcus braunii (Jones 1972).
The inhibition of bacteria on algal growth, according to Banerjee et al. (20022) and Cole (1982) can be:
secretion of algaecides
degradation of algal polysaccharindes
decomposition of algal cell walls causing algae cells to lyse (rupturing/puncturing of the cell wall) –
which may be interesting as an oil extraction process?
competition for nutrients
Banerjee et al. (2002) describes that the effects of algae and bacteria interactions depend on culture
conditions and the microbial species involved. This is based on several examples of tests performed to
elucidate the impact of different bacteria cultures on B. braunii growth and hydrocarbon content (for
references, see Banerjee et al. 2002).
The bacteria culture, which has been demonstrated to have the best and most profound effect on both the
biomass and hydrocarbon yield in B. Braunii is Flovobacterium aquatile (Jones 1972). But Grossart (1999)
tested the interactions between marine bacteria and three diatom species and also found a significant
stimulating effect of especially one bacteria culture (Fl17) on the growth rates of all three diatom species
tested.

35
However, in general conclusions are somewhat ambiguous probably due to:
a range of environmental parameters affecting the interaction
different impacts of different bacteria cultures
that even though stimulatory effects have been observed, the stimulatory substances remains
unknown
the complex nature of the stimulatory effects
These issues are also recognized by Cole (1982), and primarily, the interaction between algae and bacteria
is not treated when concerning oil‐rich algal biomass production.
Can bacteria that cause algal cells to lyse be used in extracting oil from an algal biomass?
According to the review of Cole (1982) the bacterium induced lysis of an algal cells is caused by the
bacterial release of a substance (lysozyme) which causes breakdown of the algal cell wall and release of the
cell contents. Probably owing to the need for attachment and chemotaxis, lysis occurs most rapidly in static
liquid cultures.
Heterotrophic microalgae
To solve problems on light requirements, eg., in countries on higher latitudes, some microalgae can be
cultured heterotrophically on sugar or organic acids (Table 6.1). However, the maximum specific growth
rate of algae cultures heterotrophically on simple sugars and organic acids are usually slower than those
cultured photosynthetically, sometimes up to a third (Table 6.2), which implies that the cell concentration
of heterotrophic cultures needs to be, in that case, three times higher than that of a photosynthetic culture
to achieve the same product output rate (Lee 2001).
This is, though, also one of the main advantages of producing heterotrophic microalgae as they do not
require light, it is possible to grow them in much higher cell densities. Cultures with high cell densities have
several advantages in industrial applications, as they occupy less space and reduce the expense of
extraction and other downstream operations (Chen 2008).
Xu et al. (2006) cultured Chlorella protothecoides heterotrophically on corn powder and reached a maximal
cell concentration of 15.5 g L‐1. Compared to autotrophically cultured species this is about 10 fold higher!
For example, the maximal cell concentration found in Chlorella sp. and Nannochlopsis oculata was 1.4 and
1.2 g L‐1 (Chiu et al. 2008, 2009), respectively.
However, the need for a carbon supply implies a major barrier for a bioenergy production unless it is
possible to find a waste source.
Thus heterotrophic microalgae are not considered suitable for mass cultivation for high biomass yields at
present. However, heterotrophic algae may serve as production apparatus for high quality products
(nutraceuticals, pharmaceuticals, pigments) and thereby high‐price products.

36
Table 6.1. Commercially important microalgal species that could be cultured heterotrophically. After Lee (2001).
Table 6.2. Comparison of the maximum specific growth rate of some commercially important microalgae cultured photosynthetically, heterotrophically and mixotrophically. After Lee (2001).

37
Mixotrophic algae
Mixotrophic cultivation of microalgae takes advantages of their ability to simultaneously feed on organic
energy and carbon substrates while performing photosynthesis concurrently. Some find a need for
sterilizable bioreactors for mixotrophic cultivation of microalgae a challenge (Lee 2001), while others find
the mixotrophic behaviour as a prerequisite for mass cultivation in open ponds as the mixotrophic
organism being able to “clean up” unwanted organisms (PJ Hansen, pers. comm.).
According to Yu et al. (2009, and references herein), for some kinds of algal strains, photosynthesis and
oxidation of organic substrates proceed independently in an additive manner under mixotrophic condition
so that the growth rate of cells in mixotrophic condition is equal to the sum of those in phototrophic and
heterotrophic cultures. This phenomenon has been reported from several strains of Chlorella vulgaris and
some cyanobacteria, although there are several examples which diverge from this pattern, where the
addition of glucose or like‐wise may slow down photosynthetic rate and enhance the dark respiration
(Yu et al. 2009, and references herein).
Table 6.3 lists some microalgae capable of mixotrophic cultivation and suitable substrates.
Table 6.3. Commercially important microalgae that could be cultured mixotrophically. After Lee (2001).
The benefits of culturing mixotrophic microalgae definitely require further investigations to find suitable
species or strains and optimal culturing design.

38
7. Macroalgal production for bioenergy
Seaweeds are a little exploited source of biomass for energy production. Since seaweeds are of low
importance for human nutrition worldwide they can potentially provide a basis for non‐food bioenergy. Oil
in macroalgae is not considered of any importance although some authors argue for relative high contents
of lipids in macroalgae (Hossain & Salleh 2008). Ethanol has been made from kelp‐species but mainly as
conversion of free sugars (e.g. Coastal Research Management, Kiel, Horn et al. 2000, Adams et al. 2008).
The following will only focus on cultivation of seaweed for industrial uses, as harvest of natural stocks, as in
Iceland, Ireland and Norway, is not of relevance for Denmark. Due to lack of suitable substratum in
Denmark (Wegeberg & Felby 2009) the natural stocks of seaweeds are relatively small, and harvest of these
occurrences will be neither sustainable nor economically feasible. For information on worldwide harvest of
natural seaweed stocks, see for example Critchley et al. (2006) or Steen (2005) on data from Norway.
Traditionally species of seaweeds are cultivated on industrial scale worldwide and algal polysaccharides
such as alginates and carrageenans are widely used as texturisers, gelling agents, emulsifiers and stabilisers.
There are also specialized niche markets for algal polysaccharides. Alginate is used in various
pharmaceutical preparations such as Gaviscon®, Bisodol®, Asilone® and is also used extensively as a mold‐
making material for dentistry and prosthetics. Calcium alginate is used in different types of medical
products, including burn dressings that promote healing.
The market for seaweed compounds and conversion technologies is relatively immature and, unsurprisingly
given the potential gains, highly competitive.
From the onset of the Aquatic Species Program by NREL it also included pilot‐scale cultivation of
macroalgae subcontracted by the Solar Energy Research Institute (Macintosh 1984). The original concept
was an energy farm based on macroscopic algae in open‐sea; a suspended framework structure, buoyed
and moored at great depths (> 700m) to which macroalgae as Macrocystis pyrifera (giant kelp) (Fig. 6.1)
would be attached. Very early doubts about its economic feasibility and cost‐effectiveness was raised as
the great depths required pumping of nutrient rich bottom water to the surface to nourish the growth of
the surface cultivated macroalgae biomass. The trial was beset with technical problems probably mainly
due to the open‐sea location and the test module sank to the bottom before any useful data was obtained
(Macintosh 1984)! The story does not tell the reason to this disaster but when buoyancy is calculated one
has to take into account that the growing algae eventually get heavier.
An alternative concept by the Aquatic Species Program was a land‐based energy production system of
burial vaults. 42 macroalgal species (16 green algae, 2 brown algae and 18 red algae) were screened and
two species (Gracelaria sp. and Ulva sp.) were the most successful and suitable species for cultivation; they
demonstrated the highest carbohydrate content considered to be anaerobically digested for methane gas
(Macintosh 1984).

39
Fig. 7.1. Macrocystis pyrifera. Left; creating underwater forest of up to 60m high plants. Above; bladders at base of each blade leading to buoyancy.
They also tested the lipid contents of macroalgae but initial results indicated that macroalgae produce very
low quantities of lipids ranging from 1‐5% of the total organic weight; the majority are polar, membrane‐
bound lipids, which are less valuable as fuels (Macintosh 1984).
After this 1984‐report the emphasis of the Aquatic Species Program has been on the production of
microalgae (see Microalgae Production for Bioenergy).
However, other countries have well‐established sea‐based mass production of macroalgal species. In Asian
countries as China, Japan and Korea kelp species (mainly Saccharina japonica, Porphyra yezoensis, Undaria
pennitifida) are cultivated for commercial purposes.
Japan has developed a comprehensive program (the Ocean Sunrise
Program) on large‐scale production of bioethanol made from cultivated
seaweed (Sargassum fulvellum, Fig. 7.2) for bio‐fuels (Aizawa et al. 2007).
The mid‐ to long‐term goal of the Ocean Sunrise Project is to produce 5
million kL of bio‐ethanol by farming 150 million tons of S. fulvellum.
In Europe Ireland has long tradition on utilization of especially natural
harvest of seaweed species as Ascophyllum nodosum. Irish Seaweed Center
(ISC) has been up front in developing protocols for cultivation and testing
sea‐based cultivation systems of various macroalgal species, like Alaria
esculenta (Kraan & Gury 1998, Kraan et al. 2000, Morrissey 2002),
Laminaria digitata and Palmaria palmata (Morrissey 2002, Edwards 2008)
(Fig. 7.3), all of which are commercial important species.
Fig. 7.2. Sargassum fulvellum
Also Scotland has initiated a program on sustainable fuels from marine biomass, BioMara. The Scottish
Association for Marine Sciences (SAMS) is the lead partner and the project involves research on both
macro‐ and microalgae (Gunn 2007). MHT seaweed biomass they focus on sea‐based cultivation and
conversion to methane gas and bio‐ethanol/butanol (see Mass Cultivation).

40
Fig. 7.3. Macroalgal species. From left to right: Alaria esculenta, Laminaria digitata, Palmaria palmata.
Neighbouring regions seems to stake on industrial production of seaweeds. Thus it may be possible by
acquiring the appropriate basic knowledge on optimizing cultivation processes, to develop a large‐scale
cultivation of seaweed species in Denmark for bioenergy and other biomass components? In the following
we will evaluate the potential and need for optimizations as well as knowledge of a feasible production of a
macroalgal sourced bioenergy in Denmark.
Species and growth potentials; which species are suitable for cultivation in DK?
Irradiance, temperature, nutrients, grazing and stand density are the most important parameters affecting
biomass production in the sea. Parameters, which affect photosynthesis, affect productivity. Macroalgae
show variations over day and season in photosynthetic rate, as well as variation in pigmentation related to
light conditions, such as shading or with increased depth (Dring 1986). Photosynthetic rates also depend on
interconnected external parameters as nutrients, CO2 availability and water motion, and biological
parameters such as age and species (strain). The photosynthetic rate can be enhanced up to 4 times that in
still water by increased water motion. Rapid water motion facilitates the availability and uptake of CO2 and
nutrients by the macroalgae enhancing the photosynthetic rate together with the flashing light
phenomenon (Leigh et al. 1987, Gao & McKinley 1994, and references herein, Grobbelaar 2008) (see also
Yield Optimization for Microalgae Production Systems).
Countries in temperate to arctic climates such as Canada, Faroe Islands, France, Greenland, Iceland, Ireland
and Norway and North America, which have oceanic waters as well as rocky coasts, have a high occurrence
of kelp species and kelp forests (Table 7.1). Despite of a relatively low level of irradiance, due to adaptation
and acclimation these macroalgae utilizes light so efficiently that they build up large biomasses.
Kelp species are well‐adapted to short light seasons by an ecological strategy of these perennial kelps,
which synthesize and store reserve products in summer and remobilize them for growth in winter and
spring making them capable of initiating growth under suboptimal light conditions (Bartsch et al. 2008).
Country Species Biomass (kg m‐2) References
Canada Saccharina longicruris 8 Sharp et al. (2008) Greenland (SW) Alaria esculenta +
Laminaria nigripes 8 Wegeberg (2007)
Ireland Laminaria hyperborea 19 Werner & Kraan (2004) Norway Laminaria hyperborea 20‐30 Steen (2005)
Table 7.1. Maximal biomasses of kelp species in the North Atlantic.

41
A specimen collected at the North coast of
Zealand, Denmark, in April (Fig. 7.4) was 90
cm long consisting of the old blade at the top
and the new blade from present year’s
growth at basis, already 45 cm long.
Fig. 7.4. Saccharina latissima collected at Tisvilde, Denmark, 21st March 2009. Bottom‐right; new blade, top‐left; old blade.
Therefore the relative number of red, brown and green algae species changes with latitudes. From tropical
to cold temperate regions the diversities of red and green algae are drastically reduced and the brown
algae begin to dominate also expressed in an index showing the relationship of red (R), brown (P) and green
algae (G) (R:P:C), which is 1.0:1.6:1.0 in Greenland and 1.4:1.2:1.0 in the Baltic Sea Area (based on Lüning
1990, Nielsen et al. 1995, Pedersen 1976).
The acclimation potential of some species, such as Saccharina latissima, to different irradiances is very
high. Photoprotection enables photosynthesis to acclimate both to high and very low irradiances (e.g. 1100
‐ 5 μmol m‐2 s‐1, respectively). Furthermore, respiration rates decrease under low light conditions, lowering
the light compensation point to about 2 μmol m‐2 s‐1 so that photosynthesis is balanced even during periods
of, for example, ice cover (Bartsch et al. 2008, and references herein).
Growth rates
Growth rates of kelp species can be very high in the light season with a length increase of 2 cm d‐1 in the
exponential growth phase (Mortensen & Wegeberg 2007).
In a pilot‐cultivation study of Alaria esculenta in the Faroe Islands the mean length reached approximately
1.5 m after 3.5 months of growth. The maximal individual length reached 270 cm within the investigation
period (Mortensen & Wegeberg 2007) (Fig. 7.5). In the North Sea, testing design for cultivation of
Saccharina latissima, the cultivated plants reached a mean length of 1.5‐2.0 m within one growth season
(February to June) (Buch & Buchholz 2004) (Fig. 7.5).
Estimating growth rates of the pacific kelp species, Eisenia arborea, showed that, comparing growth rates
expressed as length increase and weight increase, the weight increase was up to 3 fold the length increase
(Zertuche‐González et al. 2008). It was therefore concluded that previous estimates of the growth of E.
arborea in Mexico based on total length might have been underestimated.
Thus an obvious choice when producing large‐scale macroalgal biomasses in the North Atlantic will
therefore be cultivation of kelp species, such as especially Alaria esculenta and Saccharina latissima, but
also Laminaria digitata and Sacchorhiza polyschides, being screened by ISC and SAMS, respectively, may
also prove to be potential candidates.

42
Fig. 7.5. Kelp species biomass yield within on growth season of 1) Alaria esculenta in the Faroe Islands, and 2) Saccharina latissima in the North Sea (after Mortensen & Wegeberg (2007) and Buch & Buchholz (2004), respectively).
Mass cultivation and biomass yield of seaweeds in tanks
When cultivation of seaweeds is conducted in tanks, it usually is for high quality products as Porphyra (Levy
2008) or for bioremediation of effluent water from aquacultures of, e.g., oysters (Lüning 2008), abalone
(Robertson‐Andersson et al. 2008) and as animal (abalone) feed.
For the bioremedative effect of the macroalgae, benefitting the recycling of water, tanks may be
interconnected with water pipes and aerated individually. The residence time of the water is regulated
according to the turbulence of the aeration. The air diffusers make the algae culture rotates for enhanced
nutrient uptake and flashing light effect (Pang & Lüning 2004) (Fig. 7.6).
Sylter Algenfarm, Germany, produces four macroalgal species, Saccharina latissima, Palmaria palmate,
Soliera chordalis and Gracelaria vermiculaphylla, integrated in oyster culture on test‐scale (Lüning 2008) for
human nutrition or as raw material for the cosmetics industry (Fig. 7.7). The tanks are placed in
greenhouses with re‐circulated water.
Fig. 7.6. Tumble culture of macroalgae using specialized tanks. A) Longitudinal section with a pipe as air diffuser at bottom. B) Transverse section of tank. C) Tumble culture of Palmaria palmata, Irish Seaweed Centre. Photo: K Sjøtun.
Figure continues next page
A

43
Fig. 7.6, continued.
Fig. 7.7. Sylter Algenfarm producing four species; one brown kelp, Saccharina latissima, and three red algae, Gracelaria vermiculaphylla, Palmaria palmata and Solieria chordalis. After www.algenfarm.de.
A production of Pacific dulse (Palmaria mollis) on Big Island, Hawaii, is co‐cultured with abalone in a land‐
based system (Demetropoulos & Langdon 2004) for bioremediation and feed supply. The production of P.
mollis averages 70 t month‐1.
B C

44
In Israel NoriTechTM focuses on the production of Porphyra, the most valuable commercial seaweed,
derived from land‐based, scalable, artificial ponds, using proprietary, cost effective technology
(www.noritech.co.il), and, according to Levi & Friedländer (2008) the production can prize‐wise compete
with the sea‐based cultivation of Porphyra performed in, e.g., Japan. The production system is patented
(WO/2008/013667), and the invention claims to provide unique technology, systems and methods of
cultivating different types of seaweeds.
In Ireland intensive tank cultivation of Porphyra spp. is under trial. The project examines the different
factors affecting the vegetative growth of P. dioica in indoor tank cultures under controlled conditions, and
a small‐scale demonstration unit is developed to evaluate the production in view of economic feasibility
under Irish conditions (Hanniffy & Kraan 2008). Also in USA tank cultivating of Porphyra umbilicalis as
bioremediation for Atlantic cod production is under trial (Neefus et al. 2008).
The green algae Ulva sp. is cultivated in tanks in Denmark (Fig. 7.8) for the purpose of bioenergy production
(Rasmussen et al. 2009). According to Rasmussen et al. (2009), the production of Ulva is estimated to an
average of 45 t dry weight/ha on annual basis (200 days of sufficient light conditions).
Fig. 7.8. Cultivation of Ulva sp. in outdoor tanks at Danish Shellfish Centre, Nykøbing Mors, Denmark. Photo
from presentation by PD Jensen, Bioenergidag, 24.11.08.
Tank cultivation of Ulva spp. is critical because the frequently occurring sporulation events result in loss of
algal tissue (Werner et al. 2003, and references herein) (see Biomass Reduction). This phenomenon of
sporulation and the resulting reductions in biomass was also observed by Rasmussen et al. (2009).

45
In Mexico cultivation of Ulva spp. (former Enteromorpha spp.) was performed in ponds and in open water
in the Bay of La Paz for evaluation of biomass production in commercial scale to food and feed industries.
According to Sánchez et al. (2008) preliminary results showed that cultivation of Ulva spp. in their studies
was feasible. However, data are yet not released as well as the cultivation methodology in open water.
Sea-based mass cultivation and biomass yield of seaweeds
Most bulk biomasses of seaweed are obtained by sea‐based cultivation on sticks or ropes as known from,
e.g., China, Japan, Philippines, and in the light of globally rising demand for agricultural land focus has been
on utilizing resources from sea. Sea‐based cultivation processes and development of large‐scale seaweed
farms may deliver the required supply of biomass for an energy production. Such seaweed farms may have
a positive ecological impact by nutrient stripping and provide nursery grounds for fish fry, and will not
involve areas suitable for cereals.
The protocols of sea‐based cultivation of kelp species are known and documented by several authors
(Blouin et al 2007, Buch & Buchholz 2004, Lüning & Müller 1978, Lüning & Pang 2003, Kraan & Guiry 1998,
Mortensen & Wegeberg 2007, Shea & Chopin 2007) and goes through following steps (Fig. 7.9):
1. Collection of fertile specimens in winter months
2. Sporulation and inoculation; release of spores which are added to enriched seawater containing
items for settling (ex. lines)
3. Establishment of sporelings; life cycle accomplishment under specified laboratory condition
4. On‐grow of sporophytes in sea from February/March until harvest in summer
5. Harvest
Fig. 7.9. Production cycle of kelp species. For a detailed description of cultivation of Laminaria digitata and Saccharina latissima in Denmark, see Wegeberg (2010).
Designs and harvest of sea‐based cultivation systems
Several designs of sea‐based cultivation methods have been developed and tested. The most simple, but
also the most applicable and most commonly used, is the long‐line (Figs 7.10‐13). In Germany and in Ireland
the grid has been tested (Buch & Buchholz 2004, Morrissey 2003) (Fig. 7.14), and in Germany a cultivation
ring was developed for cultivation in high‐turbid waters (North Sea) (Buch & Buchholz 2004) (Fig. 7.15).

46
The long‐line and grid are both systems build up by ropes of 16‐18 mm thickness anchored by concrete
blocks or, for example, in between wind mills (Figs 7.11, 12). For details on mooring gear and establishment
of long‐lines and grid, see Morrissey (2003) and Buch & Buchholz (2004), respectively.
To harvest from long‐lines and grids small boats can be employed. Either the kelp biomass is cut off the
lines into the boats or the lines are pulled into the boat. So far no development has been performed on
optimizing the process of harvesting from long‐lines and grids.
The off‐shore cultivation ring developed by Buch & Buchholz (2004) (Fig. 7.15) has proved resistant to
rough weather conditions and water velocities and also permits easy handling. The ring constructed by
Buch & Buchholz (2004) carried 84 m of seed lines leading to a harvest of approximately 300 kg kelp
biomass in wet weight. To harvest the ring construction the rings can be towed to the shore and lifted by
cranes, or harvest at sea can be performed by boat‐based cranes. However, the costs of a fully mounted
ring are quite high, € 1000, as suggested by Buch & Buchholz (2004) (see Scenario for Macroalgae).
Fig. 7.10. Model of a floating raft consisting of seed line (red) with cultivated kelp on long line anchored by concrete blocks and kept floating by buoys. After presentation by Kraan & Guiry (ISS 1994).
Fig. 7.11. Line with cultivated kelp species anchored between wind turbines and kept buoyant by buoys.

47
Fig. 7.12. Long line with a twisted seed line. Saccharina latissima, Denmark.
Fig. 7.13. Long lines with mature kelp. Saccharina japonica, China.
Fig. 7.14. Example of grid‐model floating rafts system with dimension suggestions. Marker buoys (arrow). After Buch & Buchholz (2004, ladder‐model).
Fig. 7.15. Ring‐construction for seaweed cultivation. For details, see Buch & Buchholz (2004).

48
Identification of optimal locations for sea‐based cultivation
Identification of specific locations with ideal environmental conditions for optimal growth and yield,
minimal risks of impact on natural stocks and overgrowth of unwanted organisms is essential for sea‐based
mass cultivation of macroalgal species.
Following parameters have to be taken into account:
Level of exposure/wave action is important to consider as a cross point of ensuring good water
exchange and thereby continuous nutrient supply and level of mechanical stress on the cultivated
species as well on the mooring system.
Water depth is essential in several ways; too shallow sites with soft bottom may cause high siltation
and suspended solid load causing lower light conditions as well as sedimentation of particles on the
kelp biomass; shallow sites with hard bottom may cause shading of natural stands of macroalgae by the
cultivation system; very deep sites will require more costly mooring systems as well as may lead to
nutrient deficiency as cultivated seaweeds in the surface on deep water may lead to nutrient deficiency
as the re‐mineralized nutrients from the bottom only will reach the surface in areas of natural
upwelling. The optimal water depths are estimated to range between 5 – 20 m (Morrissey 2003).
Salinity, especially in the inner Danish waters (Wegeberg & Felby 2010) is a major factor influencing
growth potential of macroalgae. Therefore it has to be considered carefully if the salinity of a site in
question is optimal for the projected species to be cultivated. In general, the best results will be
reached in full strength ≈ 30‐35 psu (Wegeberg 2008, Morrissey 2003)
Fouling of unwanted organisms such as other macroalgae or mussels, sea‐squirts, hydroids, barnacles,
which will decrease growth of kelp as well as constituting a problem for the quality of the mature
biomass (see Fouling).
Bottom type is essential to the anchoring potential, as very soft bottom, for example, will constitute a
problem for the stability of the mooring system.
Congested sea areas with many other stakeholders (recreational, fishing) might need to be avoided.
According to B. Buch (pers. comm.) optimal locations are very local, even in the North Sea, and therefore
efforts have to be invested in identifying these optimal sea areas for kelp cultivation.
Fouling
Unwanted organisms may constitute a major problem on cultivation ropes as well as on the cultivated
macroalgal biomass. Both animals as well as other macroalgal species are known to foul on the available
substratum, which the cultivation system provides (for actual infestations and diseases, see Biomass Yield).
Macroalgae species, most often of the filamentous types (species of Ectocarpus, Pylaiella, Sagassum,
Ceramium, Polysiphonia), constitutes competitors for light and nutrients leading to decrease in growth
rates and final biomass yield. The epiphytic macroalgae may not threat the quality of the mature biomass
unless they appear in dominating masses.

49
As macroalgae often are cultivated in proximity to mussel farms mussel spat settlement on the macroalgal
mooring gear may constitute a problem on the weight on the mooring system (Morrissey 2003) as well as
on the quality of the mature algal biomass. The same problem is constituted by other fouling organisms as
tunicates and hydroids, which also was experience in connection with a pilot project on cultivating
Laminaria digitata and Saccharina latissima at a mussel farm in the Limfjord (2010); the cultivation lines
with relatively well‐developed kelp was in late spring/early summer completely fouled by sea squirts and
hydroids.
Also Buch & Buchholz (2004) observed heavy fouling in testing cultivation systems in a harbour. Actually
the fouling of especially sea squirt (Ciona intestinalis) was so heavy that, in the end, many algae were
basically reduced to the meristematic area leading to a negative growth rate.
Localities with high water velocities as well as lower nutrient load, e.g. the northern part of Kattegat and
the North Sea, will probably not experience the same degree of animal fouling, as wave action may knock
away the enemies of the seaweeds and also prevent grazers to feed on them (Leigh et al. 1987).
Comparison of sea‐based cultivation biomass yields
As sea‐based cultivation seaweeds on a commercial scale is premature in our part of the world not many
data are available on the actual yield obtained on sea‐based cultivation systems.
In Table 7.2 comparative available cultivation data on the kelp species, Saccharina latissima, from the
temperate Atlantic are presented.
Country kg m‐1 rope kg m‐2 t ha‐1 References
Denmark (Great Belt) 5 12 1202 Birkeland (2009) Canada 12 351 3502 Chopin et al. (2004) Germany (North Sea) 4 121 1202 Buch & Buchholz (2004) Ireland 9 Kraan & Guiry (2004) “Temperate Atlantic” 28 Ratcliff & Kraan (2008)
Table 7.2. Biomass yields of cultivated Saccharina latissima from relevant countries. 1Computed from published data. 2Extrapolated.
The extreme variation in biomass yields may be due to different inoculation conditions, for example, spore
density, spore viability, size of sporophytes when out‐planted, just to mention a few possibilities.
Furthermore, the location suitability may be reflected in the yield as well as improper extrapolations may
lead to numbers out of proportion (see Recommendations).
Reductions in biomass yield
High‐density commercial farming of macroalgal monocultures may be plagued with diseases and
infestations of algal epiphytes leading to reduction in biomass production. Several cases have been
reported for green, red and brown macroalgae.

50
Red rot disease
Red rot disease (in Japanese: Akagusare) of Pophyra caused by Pythium porphyrae is one of the most
destructive fungal diseases affecting Porphyra cultivation farms in Japan and Korea. The causative organism
of this disease is spread by fungal zoospores released into seawater. When the zoospores attach to
Porphyra thalli, they grow hyphae and kill the algae within a few days along with release of a large number
of new zoospores (Park et al. 2001, and references herein).
Therefore, in order to reduce the incidence of red rot disease in cultivation farms, it is important to detect a
small amount of zoospores in seawater before onset of infection. Park et al. (2001) tested specific primers
in detection of the red rot disease fungi Pythium spp. by polymerase chain reaction (PCR). Their results
supported that using the primers, PP‐1 and PP‐2, against P. porphyrae and P. marinum parasitic to Porphyra
is effective and sensitive for detection of hyphae and germ‐tubes of zoospores in the early infection as well
as non‐germinated zoospores in seawater before infection.
To prevent red dot disease in Pophyra cultivation farms an acidic fungicide treatment is applied (Park et al.
2008).
The red rot disease is also known in terrestrial crops but caused by other species of Phytium.
Pinhole disease
Pinhole disease is known from cultivated Undaria pinnatifida, a brown macroalga, in Korea (Park et al.
2008). The disease is characterized by numerous small holes in the thallus of the alga, and infected algae
are not considered to be suitable for human consumption. It is called a disease although the pinholes are a
result of grazing by species of phytal copepods, but the macroalga react by the cells on the periphery of the
excavated pinholes contained more pigment than normal cells in un‐infested parts of the plants (Fig. 7.16).
Fig. 7.16. Undaria pinnatifida with pinhole disease caused by phytal copepods. The area encircling the pinholes is more heavily pigmented than normal thallus. After Park et al. (2008).
Epiphytes
Floral and faunal epiphytes are common in natural macroalgae beds, but may constitute a problem in
connection with cultivated macroalgae if in amounts causing an actual infestation.
Algal epiphytes
Farms of a red alga, Kappaphycus alvarezii, for carrageen in the Philippines regularly experience epiphyte
outbreak of species of Neosiphonia (Fig. 7.17) (Vairappan et al. 2008). These infections may lead to reduced
production and in some cases to collapse of crop.

51
In 2005 a bad infection led to a desperate measure to safe the industry by importing new seedlings.
However due to insufficient monitoring as well as quarantine these efforts resulted in an introduction of
yet another species of Neosiphonia, N. apiculata. This new epiphyte has caused severe outbreaks of with
dense epiphyte coverage, secondary infections by microbes (“ice‐ice” infection) and breaking‐off of the
seaweed thallus of K. alvarezii.
Fig. 7.17. Kappaphycus alvarezii infested with the epiphyte Neosiphonia. After Vairappan et al. 2008.
Cultivating the green macroalga Ulva in culture tanks integrated in an abalone farm in South Africa
infestation of a brown algal epiphyte Myrionema stangulans has been reported (Bolton et al. 2008,
Robertson‐Andersson et al. 2008).
M. strangulans occurs naturally in wild populations of Ulva spp. It persists as spots, numbering between 5
and 10 on the holdfast section. The spots are brown regular discs, 1 ‐ 3 mm in diameter.
In Fig. 7.18 the effects of a M. strangulans infestion are presented. Typical effects on Ulva thalli in culture
are that the M. strangulans spots increase in number and move over the entire thallus surface. The Ulva
thallus becomes thinner and yellow in colour. Then, due to some unknown process (as M. strangulans is an
epiphyte with no endophytic filaments), the host thallus breaks up into pieces of 1‐3 cm in diameters. Once
this has happened, the culture populations never recover.
Fig. 7.18. Ulva without and with infestation of Myrionema strangulans. After D. Robertson‐Andersson (www.botany.uwc.ac.za)

52
According to a net‐based article by D. Robertson‐Andersson (www.botany.uwc.ac.za) M. strangulans is
called The Brown Strangler!
Infestation is particularly prevalent in culture tanks that are carbon limited due to low water exchange rates
or in tanks that have a high pH. Pulse fertilization and improved water exchange rates (12 ‐ 20 volume
exchanges per day) appear to control or minimize the infections.
It is listed as being widespread in temperate seas. Its infection and subsequent decimation of Ulva in
culture means that cultivating Ulva one needs to be able to recognize and treat infestations rapidly before
they become too severe.
Faunal epiphytes
Tamigneaux et al. (2008) experienced colonization of the invasive bryozoan Membranipora membranacea
on kelp blade and stipe in Quebec, Canada. The cover of the thin incrusting bryozoan led to loss of the
blades in 68% of plants. They therefore recommend setting up cultivation and harvesting before settlement
of bryozoans larvae in, in this case, July.
Sporulation
As already mentioned sporulation may result in loss of biomass, and is a phenomenon known for especially
green parenchymatic macroalgae, like Ulva spp. and Monostroma spp. In these species reproduction
involves transformation of somatic cells to sporangia (Wegeberg & Felby 2010).
To overcome the problem of repeated sporulations illumination by low light throughout dark‐phase can be
installed (Werner et al. 2003, and references herein). Ulva gets reproductive about every 2 weeks near full
or new moon in a semi‐lunar rhythmicity depending on season (Lüning et al. 2008). By applying artificial
light during dark‐phase the endogenous clock is disturbed. Sporulation may still occur but is restricted to
small portions of the blades. See Lüning et al. (2008) for more details.
A more elegant and cheaper solution to the problem would be to produce non‐sporulating varities through
selection of natural non‐fertile ecotypes and breeding. But compared to terrestrial crops and plants the
breeding programmes are still in their infancy.
Storage and down-stream processing
After harvest before conversion to energy the biomass may need to be stored. Not much knowledge is
provided on this issue. However, Liot et al. (1993) studied the microbiology and storing life of fresh edible
seaweeds. They concluded that the seaweeds could be stored in the cold (4°C) up to 14 days and these
compare favourably with stored fresh vegetable such as lettuce. The use of tap water to wash seaweeds
quickly altered their quality, whereas seawater washing resulted in low microbial densities during storage.
Arramara Teoranta is an example of how seaweed biomass is down‐stream processed (Fig. 7.19). The old,
low‐tech factory is placed in Connemara in Ireland, and processes natural harvested Ascophyllum nodosum.
On an annual basis they process 20.000 tons of Ascophyllum‐biomass. They dry and mill the biomass for

53
feed supplements for domestic animals and aquaculture as well as fertilizers. The products are exported to
12 different counties. When the seaweed arrives in large lorries the seaweed biomass is stored outside only
for a few days before processing is needed.
Fig. 7.19. Seaweed processing at the seaweed factory Arramara Teoranta in Connemara, Ireland.
Desiccated seaweed should be stored only for few days or in the cold. An alternative could be to leave the
harvested seaweed biomass in the sea in great nets until use or to harvest when needed. However,
weather and/or fouling may constitute a problem in both cases and trial will be required to confirm their
appropriateness.
Recommended reading
Buch BH, Buchholz CM. 2004. The offshore‐ring: A new system for the open ocean aquaculture of
macroalgae. Journal of Applied Phycology 16: 355‐368.
Werner A, Clarke D, Kraan S. 2003. Strategic Review of the Feasibility of Seaweed Aquaculture in Ireland.
Marine Institute, Technology Park, Parkmore, Galway, Ireland (www.marine.ie/marinertdi). Report, 120 pp.

54
8. Possibilities for improvement of macroalgal production systems
When the optimal species and optimal localities for cultivation in Denmark are identified, what can then
optimize the macroalgal biomass production?
1. Strain selection/breeding?
2. More efficient production and harvest technologies?
Species/strain screening
Characterization and screening of species and genotypes for biomass production regarding productivity and
yield of biomass components including bioenergy performance has only been conducted in limited extent.
In China molecular studies, amplified fragment length polymorphism (AFLP) analyses, have shown that
natural populations of Saccharina japonica, now under cultivation in China, are genetically mixed in nature
and have a high level of hybrididzation (Li et al. 2008). Some varieties have undergone several generations
of inbreeding (5‐15), selection and X‐ray treatments, and are characterized by higher growth rate (frond
elongation) at higher temperatures, higher iodine content and lower water content resulting in higher
productions than natural populations. Large‐scale cultivation showed that the production of these breed
varieties was 8‐40% higher than that of control (Wu & Pang 2006).
Also methods for strain selection of Alaria esculenta has been developed (Kraan & Guiry 1998), and it has
been shown for A. esculenta in Ireland that genotypic variation caused differences in relative growth rates
(Kraan et al. 2000).
The significant salinity gradient in the Danish inland waters provides unique environmental conditions for
the possibilities of seaweed breeding. Several brown seaweed species adapt to temperatures and salinities
and thereby establish ecotypes with genotypic variation (e.g. Kristiansen et al. 1994). These ecotypes may
provide a natural resource for breeding according to optimizing growth rates and contents of attractive cell
components.
AFLP analyses, did not, however, reveal any differences between different morphotypes of Laminaria
digitata and Saccharina latissima from Danish coastal waters (Erting et al. 2004). Differences in
morphology, established as morphotypes, may be expected, though, to be a result of the level of water
velocity and not a result of genotypic variation. However, comparison of morphotypes as a function of
water velocities was not included in the investigation.

55
Sporophyte density
Biomass yield can vary with stand density. When the density is high, light availability, nutrients and
inorganic carbon sources become limited. Optimal density for maximal yield of, e.g., a natural Macrocystis
pyrifera population in central California has been estimated at 5‐6 kg m‐2 in wet weight (Gao & McKinley
1994, and references herein.
However, when it comes to sea‐based production of kelp species the ideal density of sporephytes has not
been investigated. It should leave sufficient space for the individual macroalga to grow and develop
optimally according to light and nutrient conditions but also deny space for smothering of unwanted
organisms that compete for substrate.
Buch & Buchholz (2004) did not, either, estimate the optimal spore density in their tests of different
cultivation systems and localities. They conclude that: “Optimum density of sporophytes on the culture
lines is still unknown and requires further well‐designed investigations. “
Such investigations can be studies of different spore concentrations in the line‐seeding process at different
conditions.
Reed (1990) studied the effects of spore density on kelp recruitment in a natural habitat, and obtained
results that indicated that a settlement of at least 1 spore mm‐2 is necessary for successful recruitment of
two kelp species from the Pacific Ocean (Macrocystis pyrifera and Pterygophora californica) to macroscopic
sizes. This minimum density may be determined in part by the maximum distance separating male and
female gametophytes at which fertilization is still possible. Fertilization in kelps is enhanced by a
pheromone emitted from the egg that triggers both the release and attraction of sperm from the male
gametophyte (Wegeberg & Felby 2010). Laboratory studies indicate that the range of attraction of an egg
emitting the pheromone does not exceed 1 mm (Reed 1990, and references herein). Reed (1990) also
found that although the proportion of female gametophytes producing sporophytes was negatively
correlated with spore density in all experiments, higher densities of settled spores often resulted in greater
numbers of sporophyte recruits (Table 8.1).
Table 8.1. Settlement densities of pores on glass and rock in laboratory using three different concentrations of spore suspensions for two pacific kelp species. The differences in settlement densities among the three seeding concentrations are significant. After Reed (1990).
Although the studies of Reed (1990) do not provide any idea of the range of optimal spore density for kelp
cultivation, his results are worth bearing in mind when designing investigation on spore density
optimization as fertilization of the female gametophyte’s eggs is crucial for development of sporophytes for
on‐grow in sea.

56
Multiple harvests
To achieve higher biomass yield it may be possible to perform more than one production cycle per year. As
most kelp species are fertile during winter the seeded lines are ready for on‐grow in sea in late winter/early
spring (Wegeberg 2010). Breaking this seasonal limitation can be performed in several ways; by induction
of sporogenesis (Pang & Lüning 2004, Lüning 2008) or by resting gametophytes (Lüning & Dring 1978, Kraan
& Guiry 1998). By manipulating the timing of reproduction it is possible to achieve seed lines ready for on‐
grow in sea either earlier or later than those of the natural seasonal cycle opening up for more than just
one annual harvest.
Manipulating spore formation
Breaking seasonal limitation for year‐round zoospore formation (sporogenesis) in kelp species isolation of
blade parts of kelp species together with certain laboratory conditions is required. By mechanically blocking
the transport of the putative sporulation inhibitors produced by the blade meristem and culturing the
plants in constant short day (8 h light d‐1) sporogenesis was obtained in Saccharina latissima (Pang & Lüning
2004, Lüning 2008). Sporogenesis was successfully induced by removal of the blade meristem, either by
cultivating distal blade fragments or by performing a suitably long transverse cut in the frond (Fig. 8.1).
Fig. 8.1. Saccharina latissima. Sorus induction induced by culturing fragments in constant short days. A) Fragmentation scheme showing the middle blade portion used for sorus induction. B) Middle blade proportion with transverse cut, being too short for sorus induction (left) or suitablyy long for sorus induction (right). After Pang & Lüning 2004.
Hwang et al. (2008) also induced sori formation on isolated discs from distal parts of the blades of Ecklonia
stolonifera, an important kelp species for food and feed in Korea, in laboratory.
Resting gametophyte cultures
When gametophytes have been obtained from zoospores (Wegeberg 2010), the gametophytes can be
maintained in the vegetative state in red fluorescent light at an irradiance of 10 μmol photons m‐2s‐1 in glass
dishes containing sterile enriched seawater (Kraan & Guiry 1998) (Fig. 8.2). Gametophyte cultures grown in
red light will not become fertile as blue light induces fertility (Lüning & Dring 1978).
The resting gametophytes can be kept stored in red light, which can be obtained just by covering with red
cellophane, and change of media every month.
A B

57
For obtaining sporophytes the filamentous gametophytes are gently ground with mortar and pestle. The
few‐celled gametophytes fragments can be innoculated into tanks with seeding lines and the sticky
gametophytes fragments will settle on the lines within short time (Lüning 1979). In white flourescent light
the gametophytes mature and accomplish life history to obtain sporophytes (Wegeberg 2010).
Fig. 8.2. Resting gametophytes of Alaria esculenta in glass
dishes (After Mortensen & Wegeberg 2007).

58
9. Bioenergy conversion technologies for algae
From an energy point of view conversion of algae biomass to energy carriers the overall is identical to other
types of biomass. Thus biomass from micro‐ and macroalgae can be converted into solid, liquid or gaseous
energy carriers. But there are some major differences in the chemical and physical chemical composition of
algae as well as their physical composition which make them more or less suitable for a given conversion
technology.
In general the most important issues to remember when evaluating bioenergy conversion technologies for
algae are their high content of inorganics (more than 30% can be found), a low level of fermentable sugars,
high protein and sulphur content and a potentially very high content of oils. The glycerine has many uses,
such as in the manufacture of food, in the production of pharmaceuticals as well as a feedstock for biogas.
Algae bio-diesel
The term bio‐diesel covers a diesel type oil made from vegetable oils. In plants oils are found as tri‐
glycerides, where three fatty acids of different chain lengths are coupled to a glycerol molecule by ester
bonds (Fig. 9.1). By splitting the triglycerides into their components via a transesterification reaction the
single fatty acids can be separated for used as fuel in diesel engines. In principle the plant oils can be used
directly as diesel fuels, but triglycerides have a high melting point and can be solid at low temperatures.
The amount of glycerol relative to fatty acids depends on the chain length of the acids, but as a rule of
thumb the ratio of glycerine to fatty acids is 1:10. The glycerine has many uses, such as in the manufacture
of food, in the production of pharmaceuticals as well as a feedstock for biogas.
Fig. 9.1. Triglyceride.
The processing of plant oils to biodiesel is technically simple, the transesterification is typically done by
applying a catalyst e.g. methanol at alkaline pH, whereupon the glycerol and fatty acids can be separated.
Algae biodiesel are derived from microalgae only, as macroalgae does not have sufficient high oil content.
The production of bio‐diesel from algae oil is not any different than for other vegetable oils, but the
composition of fatty acids in algae is not the same as for other plant oils. Algae generally have a higher
content of polyunsaturated fatty acids with four or more double bonds. These acids are susceptible to
oxidation during storage and are not accepted as motor grade diesel. The problem can be overcome by a
simple hydrogenation of the oils, similar to the production of margarine.

59
From a processing point of view the main difference between algae bio‐diesel and oil crop bio‐diesel is the
different properties with regard to the applicable methods for extraction of oil from the biomass. More
than 90% of the oil from seeds can be extracted using a simple mechanical press; this is, however, not the
case for microalgae. Microalgae cell walls are very sturdy and do not break upon mechanical pressing.
Furthermore the small size of the algae makes it difficult to do mechanical extraction, as the press cake will
block the oil. Therefore, there are at present no feasible methods for mechanical extraction available.
Several methods can be applied for extraction of the algae oil. The most commonly used method is
extraction using organic solvents, typically hexane. The use of hexane for extraction of oils is already done
at a large scale in the food industry, but it is a more costly process requiring energy and chemicals
compared to mechanical processing. Another option is the use of supercritical CO2 for extraction. The
process is efficient but supercritical processes have not yet been implemented at such a scale that would be
required for bio‐diesel oil extraction. Supercritical extraction will require some input of energy (electricity),
which yet has to be quantified. Extraction using different solvents may be assisted by the use of ultrasound
to break down the algae cell wall.
From a commercial point of view algae biodiesel has a large potential provided that algae with high oil
levels are available. But it does not appear that a feasible process for large‐scale production of algae oil to
bio‐diesel is available at present. Further development of the technology will be required.
Algae ethanol
The production of ethanol by fermentation of sugars is technically well developed. On commercial scale the
sugar glucose is fermented by ordinary yeast to produce ethanol. Glucose is a so called C6 sugar with 6
carbon atoms and can be found in a number of different plant sugars of which cellulose, starch and sucrose
are the most common. New microorganisms capable of fermenting C5 sugars are under development, but
have not yet reached a commercial scale and applicability.
An important parameter in ethanol production is an ethanol level following fermentation of at least 8 %. At
lower levels the energy requirement for separation (distillation) of the ethanol rises exponentially.
The potential for production of ethanol from any biomass is dependent upon the presence of fermentable
sugars. Algae only have a low level of fermentable C6 and C5 sugars, typically below 10%. Therefore it is not
possible to obtain sufficient yields for efficient ethanol production, just as the high salt content requires
extensive washing. It may be possible to breed new algae strains capable of higher production of
fermentable sugars, but it does not seem likely that the carbohydrate content can be increased to levels
that will provide sufficient ethanol for efficient distillation.
Algae ethanol is therefore neither technically nor economically competitive with current use.
A radically different approach is to develop cyanobacteria capable of directly producing ethanol from
photosynthesis. The company Algenol is currently building a large‐scale unit in Mexico and claim to go on‐
line by 2009. Very few details about the process and the biochemistry involved are available.

60
Combustion
Efficient combustion of biomass requires a low content of inorganic components as well as of nitrogen and
sulphur compounds.
The chemical composition of algae biomass (see Chapter 4) shows that it is not a suitable feedstock for
combustion. A high content of sulphur, nitrogen, alkali and inorganic compounds requires extensive
cleaning of the flue gas just as fouling of the boilers will be a major problem. Raw algae biomass is thus not
suitable for heat and electricity production by combustion. Application of algae for combustion will require
extensive washing and separation of inorganics, N‐ and sulphur compounds from the carbohydrates and
oils. No reports on combustion of algae biomass have been found.
Biogas
Biogas is produced by methanogenic bacterial breakdown of organic matter under anaerobic conditions. It
is also known as anaerobic digestion or mesophilic fermentation, and consists of at least three bacterial
processes yielding the main final products methane (CH4) and CO2. A wide range of organic matter can be
converted in a biogas reactor, with lignin and cellulose being the main exceptions. The processing of
biomass in a biogas reactor typically takes weeks, but the process as such is robust. The composition of
biogas varies depending upon the feedstock. Biogas will contain CH4, CO2, H2, N2, O2, H2S and water vapour.
Typically the methane content of biogas is 55‐75%CH4, 20‐40% CO2 and smaller amounts of the other gases.
Purification of the gas is therefore needed and several large‐scale methods are available. Following the
conversion to biogas there will be residual liquid and fibre fractions. The liquid fraction typically has a high
content of salts and nitrogen, whereas the fibre fraction contains the biomass that could not be digested by
the bacteria.
Biogas production from of algae by methanogenic bacteria is probably the best suited technology for
converting both macro‐ and microalgae biomass into a practical energy carrier.
For algae biomass the high content of sulphur is a challenge as H2S is corrosive and will require special
considerations in design of the biogas plant. Also the high content of silicium in algae may produce
siloxanes that need to be removed.
The energy content of biogas varies from 16‐30 MJ m3 depending on the feedstock composition and thus
the final concentration of methane. For algae the yields has been reported to be between 0.15 to 0.65 m3
kg‐1 of dry biomass. However, the published most of the published reports show somewhat lower yields e.g.
0.18 m3 kg‐1 using different species (Fernandes et al. 2007) and 0.20 m3 kg‐1 using Ulva (sea lettuce) (Brian &
Moran 1997). A reasonable average yield from algae biomass is 10 GJ of energy ton‐1 of biomass; this is the
net yield including purification. This puts the biogas yield higher than manure but lower than e.g. clover
grass or whole crop corn, therefore algae biomass is not suitable to boost the conversion of manure to
biogas.
The biggest practical challenges with regard to biogas from algae are the residual fractions. For the fibre
fraction the high sulphur and nitrogen content makes it less suitable for combustion. The liquid fraction will

61
have a high content of nitrogen which making it well suited as a fertilizer, but in the case of marine algae
the residual salt could make application in the field difficult.
No technical development has addressed these problems at present, but they need to be solved in order to
make algae biogas commercially viable.
In general biogas is the currently best suited technology for converting algae biomass into practical energy
carriers. Some design considerations are needed for the biogas plant but for the digester itself there are no
major technical constraints. However, more development is needed with regard to the residual fibre and
liquid fraction. It has not been tested if the liquid fraction is suitable as fertilizer or if the fibre fraction can
be combusted, therefore more knowledge is needed in order to evaluate the full potential of algae biogas.
Thermo-chemical processing
Algae biomass can be processed using thermochemical processes, no different from those applied for
conversion of biomass residues. This may include pyrolytic cracking or gasification and subsequent
reforming of the syngas. The products from these processes are a crude bio oil or synthetic diesel oils, he
latter also known as biomass to liquid (BtL) diesel.
Patil et al. (2008) describe hydrothermal liquefaction as an efficient option for converting wet biomass to
oil. Hydrothermal technologies are broadly defined as pyrolysis at 200‐600 C and pressures of 5‐40 MPa in
liquid or supercritical water (Peterson et al. 2008). The bio‐oil produced by such a process will have a high
content of organic acids and the shelf life will be short, as the unrefined oil will undergo further oxidation.
Therefore it will require further processing by purification and hydrogenation in order to produce a suitable
fuel.
Patil et al. (2008) suggest that the moist biomass can be heated by microwave power as performed by
Bohlmann et al. (1999) on sewage sludge, but from a processing point of view the large amounts of electric
power required will probably make this too expensive.
Minowa et al. (1995) liquified algal cells of Dunaliella tertiolecta with a moisture content of 78.4% by
termochemical liquefaction. At 300°C and 10 MPa the oil yield was about 37%, and the oil calorific value
was comparable to that of fuel oil.
In Switzerland Haiduc et al (2008) are developing: “an innovative process (SunChem) for the production of
methane via hydrothermal liquefaction and gassification of microalgae.” The process consists of five steps:
1. Photosynthetic production of biomass in photobioreactor
2. Mechanical dewatering (to 15‐20% dry weight)
3. Hydrothermal liquefaction of biomass sludge and salt separation for reuse of nutrients
4. Catalytic hydrothermal production of CH4
5. CO2 separation from the product gas and recycling to the photobioreactor

62
The Swiss group is currently working towards demonstrating the technical and economical feasibility of the
SunChem process for sustainable production of CH4 (Haiduc et al. 2008).
The process may be an economic alternative to other harvest and conversion technologies depending on
the extent to which otherwise necessary costs for harvest, drying and conversion can be dispensed with.
Microalgae for H2 production
A different technology for producing energy caariers from algae is the photobiological production of
hydrogen. Prince & Khesghi (2005) present a review of photobiological production of hydrogen.
Prince & Khesghi (2005) describes how cyanobacteria and microalgae emits hydrogen only in small
amounts under normal photosynthetic conditions. But many microalgae and cyanobacteria can synthesize
enzymes (nitrogenase, hydrogenases) that can reduce protons to gaseous hydrogen. These enzymes
provide proton‐reducing activity and actively evolve H2 if provided with reductant (e.g. N2). Thus the
presence of hydrogenase or nitrogenase will divert electrons to these enzymes and produce H2. From a
biochemical point of view the advantage is that the formation of H2 it is not limited by the slow CO2 fixation
rates but build upon the much faster light harvesting complexes. For more details see Prince & Khesghi
(2005).
Production of H2 involves some of the same barriers as those for lipid production; the photosynthetic
efficiency and mass cultivation. Furthermore, production of H2 will imply sterile photobioreactors to
minimize the biological consumption of hydrogen.
Some efforts have been focused on H2 production using the green microalga Clamydomonas reinhardtii.
Lehr et al. (2008) and Torzillo et al. (2008) investigated the possibilities of optimizing H2 production as
hydrogen photoproduction with C. reinhardtii is sustained by depriving the cultures of sulphate. They
tested switch from growth to H2 production phase with controlled sulphur feeding and aeration as well as
flashing light effects.
If photobiological hydrogen production is to achieve commercial success, it must use solar photons
efficiently. According to Prince & Khesghi (2005) the highest photon efficiency of the energetic systems of
hydrogen evolution is approximately 40% and they suggest several prospects of cell engineering to bring up
the photosynthetic efficiency by bringing down energetic costs. See Prince & Khesghi (2005) for more
details.

63
10. Sequestration of CO2, NOX and SOX
As any other photosynthetic organisms algae are capable of fixating CO2 from the atmosphere. Therefore
they present an opportunity to serve as carbon sinks and an approach to CO2 sequestration is to use plant
biomass, with a relatively long residence time, e.g. wood to fix CO2. It has also been suggested to use large,
perennial kelp species in the sea to increase living plant biomass in the sea (Fei et al. 2007). In order to
meet the CO2 sink purpose some kinds of artificial large facilities should be constructed and cultivation of
long‐living kelp species without blade‐change as, for example suggested, Sargassum spp., has to be
developed. However, the scale of such systems would be very large, and the use of algae biomass as CO2
sink should only be seen as a side effect of biomass production for energy.
Growth rates and CO2 uptake
To estimate the sequestering effect of algal biomass on CO2, one has to take into account that the uptake
rates as well as the ability to take up dissolved inorganic carbon (DIC) as eitherCO2, HCO3‐ or CO32‐, may be
specific or even strain‐specific (Gao & McKinley 1994, Douskova et al. 2009), in the case of macroalgae,
depending on the presence or absence of extracellular carbonic anhydrase (CA) (Gao & McKinley 1994), for
more details, see Gao & McKinley (1994).
Chiu et al. (2008, 2009) tested the growth rate and lipid accumulation of the microalgal species, Chlorella
sp. and Nannochloropsis oculata, respectively, cultivated at different CO2 concentrations. They found that
at a 2% CO2 concentration the optimal CO2 removal efficiency was reached for Chlorella sp. (Chiu et al.
2008) as well as the maximal biomass and lipid productivity was reached with 2% CO2 aeration for N.
oculata (Chiu et al. 2009).
For macroalgae studies have shown that to CO2 saturate photosynthesis concentrations of up to 5 times
that of the ambient for ordinary air were needed for a number of macroalgal species (e.g., Cladophora
rupestris, Macrocystis pyrifera, Gracelaria sp., Ulva lactuca) (Gao & McKinley 1994, and references herein).
A better understanding of the use of inorganic C by algae under elevated CO2 concentrations is important
for predicting the effects of CO2 on biomass production, and thereby the CO2 bioremediation effect.
Flue gas as CO2 source
Douskova et al. (2009) tested flue gas originating from a municipal waste incinerator (MWI) as a source of
CO2 for the cultivation of a microalga, Chlorella vulgaris, in order to decrease the biomass production costs
and to bioremediate CO2 simultaneously. Flue gas emission limits from MVIs are very strict, in most
parameters by over an order of magnitude lower than those valid for the coal‐fired power plants.
For most microalgae cultivation the optimum concentration of CO2 in the gas mixture is 2%, as also shown
by Chiu et al. (2008, 2009) (see above). However, the strain of Chlorella vulgaris used by Douskova et al.

64
(2009) allowed its cultivation directly on the cooled flue gas containing 10‐13% of CO2, and showed a higher
growth rate than the growth of the control culture grown under conditions usually regarded as optimal.
Acién and co‐workers (Acién et al. 2008, González et al. 2008, Sierra et al. 2008) conducted investigations
to elucidate the CO2 fixation of microalgae. They tested a number of cyanobacteria species for their ability
to fix CO2, and developed a model for predictive control of bioreactor operation for CO2 removal from flue
gases by microalgal systems. They found that the best results were obtained from Anabaena sp., which,
when cultivated on flue gas containing 10% CO2, reached a CO2 fixation rate of 15‐90 g CO2 m‐2 d‐1 or 1.5 g
CO2 L‐1 d‐1.
According to Douskova et al. (2009) the toxicological analysis of the biomass produced using untreated flue
gas showed only a slight excess of mercury while all the other compounds; other heavy metals, polycyclic
aromatic hydrocarbons, polychlorinated dibenzodioxins and debenzofurans, and polychlorinated biphenyls,
were below the limits required by the European Union foodstuff legislation. Extending the flue gas
treatment prior to cultivation by active carbon led to an efficient absorption of gaseous mercury and to the
algal biomass composition compliant with all the foodstuff legislation requirements.
Douskova et al (2009) explain that at the assumed 47% of carbon in algal dry biomass, about 1.72 kg CO2 kg‐
1 biomass is required, which is in accordance with Hall et al. 1991, Sánchez Mirón et al. 2003, Christi 2007,
Patil et al. 2008, Rodolfi et al 2009, all considering 1 kg of dry algal biomass requiring about 1.6‐1.8 kg of
CO2. However, Douskova et al. (2009) computed by a mathematically model, which permitted the
calculation of optimum flue gas injection rate into the photobioreactor containing Chlorella sp. From this
theoretical model on the mass balance of CO2 obtained from the flue gas stream and for the algal
suspension it was estimated that about 50% of flue gas decarbonization can be attained in the
photobioreactor and that 4.4 kg of CO2 is needed for production of 1 kg (dry weight) algal biomass. That is
about 2.5 fold higher than generally stated (1.6‐1.8 kg CO2 kg‐1 biomass) and definitely needs verification by
similar studies.
NOx, SOx
It has been reported that sulphur and nitrogen oxides (SOx, NOx), which were present in flue gases from
power plants boilers, inhibited algal growth, as well as the presence of these gases (at low levels) had
barely any influence on algal growth (Doucha et al. 2005, and references herein).
Doucha et al. (2005) tested the effect of NOx present in flue gas on the microalgae Chlorella sp. They found
that the presence of NOx did not inhibit the growth of microalgae as well as the degree of denitrification of
the flue gas, estimated from NOx absorption in the algal suspension was about 10% of the NOx content in
the inlet gas stream.

65
The studies of Doucha et al. (2005) and Douskova et al. (2009), although some of their results need support
by similar studies, are excellent examples of commercial useful research, which actually evaluates some of
the obvious potential benefits of producing biomass for bioenergy. More studies of this kind are definitely
required for evaluating this flue gas bioremediation effect, but also others of the non‐evaluated suggested
potential benefits of using algal biomass for bioenergy.

66
11. Scenario for microalgal production in Denmark
In the comprehensive work of the Aquatic Species Program (Sheehan et al. 1998) four major difficulties
associated with microalgal mass culture for fuel production were identified connected to open pond
systems:
Photosynthesis efficiency for light energy and high lipid production
Fundamentals of species selection and control in open pond systems
Mechanisms (and control) of algal bio‐flocculation
Effects of non‐steady‐state operating conditions on algal metabolism
as only a very low‐cost system, based on open ponds mixed on low velocities and using very simple
harvesting process, could be considered in such a production.
They also conclude that even with these rather favourable, though plausible, assumptions, costs would still
be well above those for current, or projected, oil prices. These considerations seem still to be valid, and
given the current state‐of‐the‐art oil prices above 200 USD/barrel would be needed in order to make
microalgae based bio‐fuels competitive.
As no research has been performed in Denmark using open‐pond or photobioreactor systems in open air or
on best suited microalgal species or strains for optimal biomass production and lipid yield, the scenario will
present calculations on realistic data from productions in Eilat, Israel, which actually are running. The
production of Dunaliella is for production of carotenoid‐rich algae powder for the health product market in
Asian countries, but the recently initiated production of Nannochloropsis will be evaluated for its biofuel
potential (Ben‐Amotz, WSA 2009). At present the total scale of the Dunaliella sale is approximately 100 mill
$ year‐1.
Prospects and limitations comparing the geographical parameters (temperature, hours of sunlight, non‐
arable land) of Israel and Denmark will be discussed.
Production and production systems
Open‐ponds seem at the moment to be the best solution for microalgal cultivation, which also was
supported by Dr Ami Ben‐Amotz at the International Workshop on Offshore Algae Cultivation for Biofuels
and Beyond (WSA), Lolland, April 2009. Dr Ben‐Amotz did not want to discuss open‐ponds vs.
photobioreactor systems, but stated that all commercial productions at present were open‐ponds.
The photosynthetic limitation of long‐term algal productivity leads to a maximum theoretical microalgal
productivity of 25 g m‐2 d‐1 (Table 11.1) (Ben‐Amotz, WSA 2009). According to Ben‐Amotz (WSA, 2009) the
actual productivity of Dunaliella in Eilat, Israel, is 2‐4 g m‐2 d‐1.

67
Environmental Factor Reduction (%)
Solar light ‐ 100
Scattering and eflecting properties of surface 10% 90
Absorption spectrum (depth of culture) 50% 45
Photosynthetic efficiency (25%) 75% 11.3
Light saturation (7‐95%) 60% 4.5
Respiration, photo‐respiration, excretion 5% 4.3
Photo‐inhibition 10% 3.8
Temperature 20% 3.1
Productivity
Mean daily solar intensity 4.000 kcal m‐2 day‐1
Energy production at 3% efficiency 120 kcal m‐2 day‐1
Algal biomass productivity (5 kcal g‐1) 25 g m‐2 day‐1
Table 11.1. The theoretical maximum production of microalgae following Ben‐Amotz (WSA, 2009).
Up-scaling
Up‐scaling is one of the major bottlenecks in cultivation of microalgae. A successful up‐scaling methodology
has been developed in Israel by NBT Ltd and Seambiotic in Eilat. They followed a step‐wise up‐scaling
approach (see below). This method has the advantages that the technologies and economics can follow
each other tightly. The up‐scaling is thus based on real biological data at the actual location as well as
present market prices.
The production starts with flask cultures in‐door and then cultivation trials in small out‐door containers
followed by up‐scaling to open‐pond systems; race‐ways with paddle wheel mixing (Fig. 11.1). The race‐way
systems also are scaled up from micro bioreactor (200‐300 L), mini bioreactor (3.000‐5.000 L), inoculation
bioreactor (30.000‐70.000 L) to process bioreactor (200.000‐1.2 mill L). The up‐scaled bioreactors are fully
technological controlled resulting in low manpower requirements.
Fig. 11.1. Up‐scaling of cultivation of Nannochlopsis. From left to right: In‐door cultivation of stock‐cultures; Out‐door cultivation of stock‐cultures; Up‐scaling cultivation.

68
The intensive plant of Dunaliella production (NBT Ltd) is covering 100.000 m‐2, and has been producing
since 1990.
Costs
According to the calculations by Ben‐Amotz (WSA 2009) (Table 11.2) the heaviest costs in cultivating land‐
based microalgae, besides electricity, are fertilizers, seawater and CO2.
Fertilizers
Producing fertilizers has energy costs resulting in, e.g., ammonium costs track with oil/gas prices, and
trending upward (Fig. 11.2). Furthermore, fertilizers are not an infinite resource and therefore the fertilizers
will need to be recycled in a sustainable way (Pate, WSA 2009).
Seawater
Seawater is not of the same value as freshwater, but for land‐based plants with large up‐scaled open
bioreactors the supply of seawater is a major production cost. The filtration and chlorination of seawater is
estimated to $ 0.25 m‐3 (Ben‐Amotz, WSA 2009).
CO2
The supply of liquid CO2 for optimal microalgal production is another major cost on the budget (Table 11.2)
as there is a significant loss in the open ponds (Ben‐Amotz, WSA 2009).
Dunaliella
NBT Ltd., Eilat
Seambiotic/IEC Plant
(estimated)
Cost in US$ year‐1
Manpower
Electricity ($ 0.125 KW‐1) and residual energy 180.000 30.000
Fertilizers (N, P, K, Fe) and other chemicals 36.000 36.000
Domestic land taxes 50.000 10.000
CO2 150.000 5.000
Seawater 200.000 5.000
Freshwater 20.000 10.000
Other supplies and miscellaneous 30.000 20.000
Total 1.166.000 236.000
Yearly production of dry algae biomass 70 tons
(2 g m‐2 day‐1)
700 tons
(20 g m‐2 day‐1)
Cost per 1 kg dry microalgae $ 17.00 $ 0.34
Market price β‐carotene health food
$ 4.000
For bio‐fuel costs should be
< $ 0.5 per kg algal dw
Table 11.2. Microalgae production costs. Slide from the presentation of Ben‐Amotz (WSA, 2009).
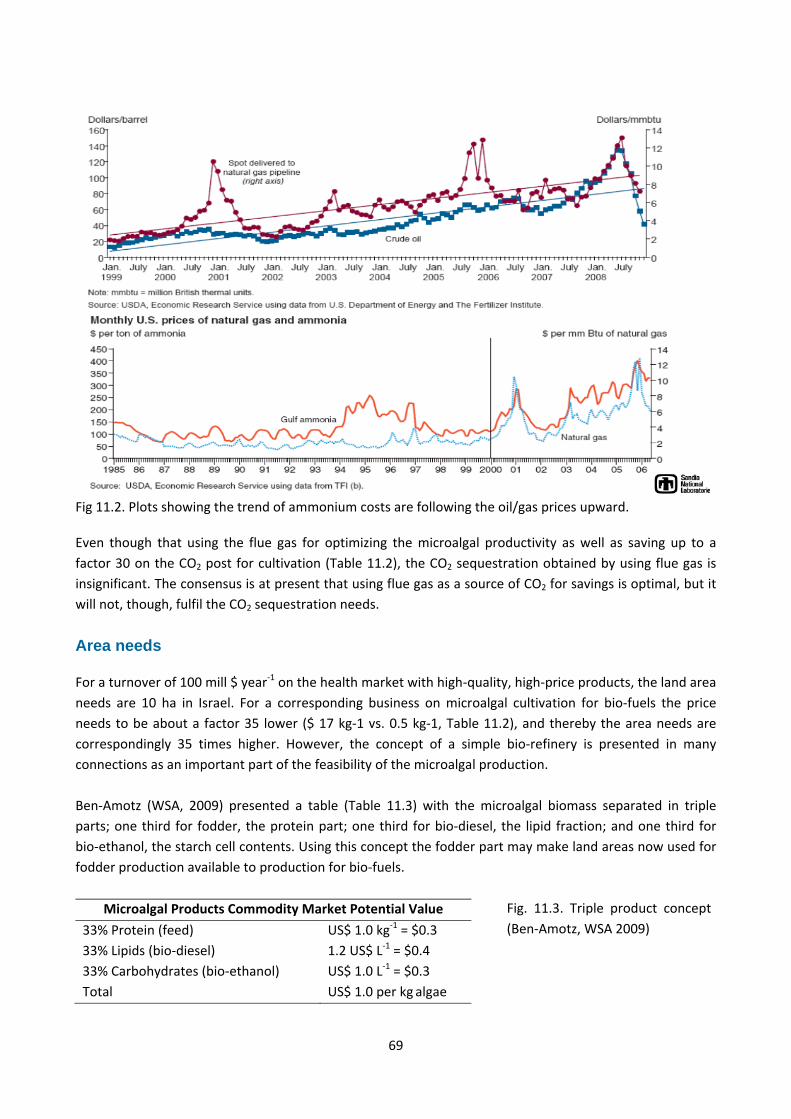
69
Fig 11.2. Plots showing the trend of ammonium costs are following the oil/gas prices upward. Even though that using the flue gas for optimizing the microalgal productivity as well as saving up to a
factor 30 on the CO2 post for cultivation (Table 11.2), the CO2 sequestration obtained by using flue gas is
insignificant. The consensus is at present that using flue gas as a source of CO2 for savings is optimal, but it
will not, though, fulfil the CO2 sequestration needs.
Area needs
For a turnover of 100 mill $ year‐1 on the health market with high‐quality, high‐price products, the land area
needs are 10 ha in Israel. For a corresponding business on microalgal cultivation for bio‐fuels the price
needs to be about a factor 35 lower ($ 17 kg‐1 vs. 0.5 kg‐1, Table 11.2), and thereby the area needs are
correspondingly 35 times higher. However, the concept of a simple bio‐refinery is presented in many
connections as an important part of the feasibility of the microalgal production.
Ben‐Amotz (WSA, 2009) presented a table (Table 11.3) with the microalgal biomass separated in triple
parts; one third for fodder, the protein part; one third for bio‐diesel, the lipid fraction; and one third for
bio‐ethanol, the starch cell contents. Using this concept the fodder part may make land areas now used for
fodder production available to production for bio‐fuels.
Microalgal Products Commodity Market Potential Value
33% Protein (feed) US$ 1.0 kg‐1 = $0.3
33% Lipids (bio‐diesel) 1.2 US$ L‐1 = $0.4
33% Carbohydrates (bio‐ethanol) US$ 1.0 L‐1 = $0.3
Total US$ 1.0 per kgalgae
Fig. 11.3. Triple product concept
(Ben‐Amotz, WSA 2009)

70
Potential for energy production
In the short to medium term the only relevant technology for converting microalgae to bioenergy is biogas.
Under the assumption that we under Danish conditions can achieve a microalgae productivity of 25 g m‐2 d‐
1 we will achieve a yield on a yearly basis corresponding to 91 t/ha. Converting this figure to biogas (see
Chapter 9 on conversion) the energy yield will be 910 GJ/ha. This is a 2.5 times higher energy production pr.
area unit compared to biogas from whole crop corn. However, looking at the estimated cost for producing
microalgae we will need a substantial cost reduction in order to make energy production from microalgae
economically viable. Further technological development of algae bio‐diesel or more efficient photo
bioreactors may change this, but at present these technologies are not available
Prospects and limitations
The above example is primary based on an existing production of microalgae biomass in Israel. Therefore,
to complete the scenario a comparison of geographical factors of the two countries is needed to evaluate
the potential for economical and ecological feasible microalgae production in Denmark (Table 11.4).
The large amount of sunlight and the high salinity at Eilat, Israel, makes this area suited for especially
cultivation of the halophile and sunlight tolerant Dunaliella. Furthermore, mass‐cultivation of microalgae
utilizes the non‐arable land in southern Israel for biomass production. However, the overly high summer
temperatures constitute a problem to cultivation (Ben‐Amotz, WSA 2009).
On the contrary, in Denmark the limited hours of sunlight result in a limited numbers of days for growth.
The relatively low temperatures may, however, have a positive impact on the production costs in, e.g.,
photobioreactors, which main problem is over‐heating.
Conclusively, the potential and size of microalgal biomass production in Denmark in either open‐ponds or in
photobioreactors may depend on trial.
Israel, Eilat Denmark
Sunlight ≈ 3000 h year‐1 ≈ 1500 h year‐1 Temperature > 30 °C in summer ≈ 20°C Non‐arable land
Israel is about half the size of Denmark but have the same number of inhabitants. In south the dessert is unsuitable for crops but suitable for algal cultivation. Only a minor part of this dessert is connected to sea at Eilat.
60% of the area is arable land. Therefore non‐arable land is limited.
Table 11.4. Comparison of geographical factors for Israel and Denmark.

71
12. Scenario of macroalgal production and conversion
In this scenario emphasis will be put on sea‐based cultivation as cultivation of, for instance, Ulva lactuca in
ponds can be obtained from the research group on this subject in Denmark (contact: Michael B.
Rasmussen, Aarhus University).
Also for the land and water saving issues the emphasis is put on sea‐based cultivation. Furthermore,
controlled sea‐based cultivation of macroalgal biomass may have a positive impact on the Danish sea
nutrient levels, which at present lead to phytoplankton blooms which eventually lead to dead zones; sea
areas with oxygen depletion at the bottom. By cultivating macroalgae at sea and harvest the biomass,
nutrients are actively removed from this ecological system.
To evaluate the potential of cultivated macroalgal biomass production at sea the species with the highest
production needs to be selected as well as data on production rates per mooring unit (ex. m rope‐1) is
needed.
From the preliminary results of cultivating kelp species in the Limfjord the best candidate for cultivation of
the largest biomasses is Saccharina latissima (Fig. 12.1), the species for which growth rates are provided in
literature. Therefore, the following scenario is based on production of this species.
Fig. 12.1. Preliminary results on growth rates of Saccharina latissima and Laminaria digitata in the Limfjord, January to April 2009.
Area needs
As the cultivation of Saccharina latissima in the Limfjord will continue to June 2009, the data presented
here are obtained from the literature: Birkeland (2008), Buch & Buchholz (2004), and data stated by Buch
(WSA, 2009) (Tables 12.1‐4).

72
The results obtained on cultivated biomass production of S. latissima in the closest proximity of Denmark
are in the Danish inland waters in the Great Belt in connection with a salmon farm (Birkeland 2008). This
work was part of a master thesis and tested the bioremediation impact of S. latissima on the salmon farm,
Musholm Lax, in the concept of Integrated Multi‐Trophic Aquaculture (IMTA). The results on production are
presented in Table 12.1.
Author Months of growth
Kg m cord‐1 m cord m‐2 Kg m‐2 Kg ha‐1 t ha‐1 ha mill t‐1
Birkeland (2008)
6 5,3(1) 2,2/(0,4) 11,66 116.600 116,6(21) 8.576(47.650)
Table 12.1. Biomass (wet (dry) weight) and area needs for 1 t of biomass of Saccharina latissima as cultivated as part of IMTA in Denmark. Data on wet biomass (kg) m‐1 cord and m cord m‐2 from Birkeland (2008).
The production results on S. latissima obtained by Buch & Buchholz (2004) was part of testing of different
mooring systems in the North Sea and the attachment strength of the kelp (Buch & Buchholz 2005). The
most exposure withstanding and most easy‐handling system was a ring construction (see Macroalgae
production, Cultivation), and the only offshore construction from which they obtained biomass production
results. The results are presented in Table 12.2.
Author Months of growth Ring area (m2) Kg ring‐1 Kg ha‐1 t ha‐1 ha mill t‐1
Buch & Buchholz (2004)
5 25 300(54) 120.000 120(22) 8.333(46.300)
Table 12.2. Biomass (wet (dry) weight) and area needs for 1 t (wet (dry) weight) of Saccharina latissima when cultivated in the North Sea. Ring diameter and biomass (wet weight) ring‐1 are obtained from Buch & Buchholz (2004).
In connection with a presentation on the WSA, 2009, Dr. Bela H. Buch was asked about the amount of
biomass production on one ring, which he said was 800‐1000 kg of wet biomass (≈ 150‐180 kg dry weight,
where dry weight constitutes 18 % of wet weight) per 80 m’s of seed line per ring. This number is 2 to 3
times higher than presented in the literature (Buch & Buchholz 2004), which, however, also seemed
somewhat low. The results of Buch & Buchholz (2004) from the North Sea equalize the results of Birkeland
(2008) from the Danish inland waters, but due to environmental factors, such as salinity and exposure, the
productivity is expected to be higher in the North Sea. Specimens of S. latissima casted onshore on the
West coast of Denmark are usually much larger than those to be collected in Kattegat according to Dr Aase
Kristiansen (pers. comm.). Therefore these newly introduced data are presented in Table 12.3.
Author Kg ring‐1 Kg ha‐1 t ha‐1 ha mill t‐1
Buch (WSA, 2009)
800‐1000 (140‐180)
320.000‐400.000 320‐400 (60‐70)
3.125/2.500 (17.500/14.000)
Table 12.3. Biomass (wet (dry) weight) and area needs for 1 t of biomass (wet (dry) weight) of Saccharina latissima as cultivated in The North Sea off Germany. Ring diameter and biomass (kg) ring‐1 are obtained from Buch & Buchholz (2004) and Buch (WSA, 2009).

73
Areas needed for production as calculated in Tables 12.1‐3 have also been calculated for 5 t, which is
considered as the most wanted scenario for a secure supply of biomass for energy production. The
calculations of needed areas for 1 and 5 t wet weight kelp biomass, respectively, are presented in Table
12.4.
Table 12.4. Calculations of the area needed for 1 mill and 5 mill t wet weight of Saccharina latissima.
To illustrate the area needs according to the different biomass data obtained by the different authors,
squares of 3.000, 10.000, 15.000 and 40.000 ha are presented in Fig. 12.2, only for wet weight, though.
5,5 km
5,5 km a. 1 t
3.000 ha
10 km
c. 1 t 10.000 ha
10 km
12 km
b. 5 t 15.000 ha
12 km
20 km
d. 5 t 40.000 ha
20 km
Fig. 12.2. The area required for cultivation of 1 or 5 mill t kelp biomass (wet weight) according to the different data available summed up in Table 12.4; a. 1 t and b. 5 t (Buch WSA, 2009); c. 1 t and d. 5 t (Birkeland 2008, Buch & Buchholz 2004).
1 mill t wet weight kelp biomass (ha‐1)
5 mill t wet weight kelp biomass (ha‐1)
Birkeland (2008) 8.576 42.880 Buch & Buchholz (2004) 8.333 41.665 Buch (WSA, 2009) 3.125/2500 15.625/12.500

74
Production costs
The cost will vary according to cultivation methodology and location. Local parameters as exposure (wind,
currents, tides) decide the amount of anchoring required as well as number of buoys and marking.
It is most wanted to cultivate kelp biomass in the North Sea due to many conflicting interests in the area of
Kattegat as well as the suitable salinity in the North Sea. For cultivation in the North Sea the ring
construction presented by Buch & Buchholz (2004) will be suitable due to its robustness to wave exposure,
and easy handling both off‐ and on‐shore. However, according to Dr Bela H. Buch (pers. comm.) the ring
construction is much too expensive and he recommended long lines.
Therefore, a calculation of estimations of the costs of a long line/grid plant has been performed (Table
12.5), as developed for cultivation of kelp species in Greenland (Wegeberg 2008). The original plant design
was presented by Kraan & Guiry (ISS 1994) (Fig. 12.3). The plant consists of main lines and cords, which is
seeded with kelp species. The line system is anchored by wires in concrete blocks and held buoyant by large
corner buoys (1000L), smaller middle buoys (200L) and cord buoys (25 L). The actual investment is DKK
60.000 and includes; laboratory equipment (vessels, projectors, pumps, weight, etc.), wires, lines, cords and
concrete blocks. The running costs are estimated to DKK 15.000 and include; seed lines, nutrients, sterile
filtered seawater, etc. For more details, see Appendix 1.
Year 1 Year 2
Seed lines 45.000 15.000 Sea plant 30.000 Total 75.000 15.000 Investments 60.000
Table 12.5. The costs (DKK) of the seeding lines in the laboratory and the actual plant in the sea for on‐grow.
Fig. 12.3. Sea‐based plant design presented by Kraan & Guiry (ISS 1994). For detailed description, see text.
The variable costs of production of 1 t wet weight of kelp according to literature follow in Table 12.6.
Table 12.6. Estimates of the tons‐wise costs (DKK) of kelp biomass cultivated in a line system anchored by concrete blocks. *The conversion factor from wet weight to dry weight is conservatively 5.55 (17‐19 % dry weight in kelp species (Wegeberg et al. 2007)).
It must be emphasized that the above biomass costs are based on a very simple cost system, and that no
land‐based or boat facilities as well as labour costs are included.
Kelp biomass (t ha‐1) DKK (t wet weight‐1) DKK (t dry weight‐1)*
Birkeland (2008) 116,6 130 700 Buch & Buchholz (2004) 120 125 690 Buch (WSA, 2009) 320‐400 40‐50 210‐260

75
Reported production costs
The above calculated costs are in the range of other known or calculated prices.
A cost of DKK 500 t‐1 has been introduced in Denmark by Thomas Bjerregård, Hov, who cultivates
Saccharina latissima at mussel lines for Sylter Algenfarm.
In connection with a presentation of Dr Andrew Ross, Leeds University, on Marine Biomass Potential and
Limitation following prices were calculated and presented for the macroalgae: US$ 0.045–0.31 kg‐1 (DKK
225‐1550 t‐1) based on Chynoweth (2002). Compared to these costs the production costs of wheat straw
were presented to be US$ 0.04 (DKK 200 t‐1).
To improve costs the productivity and biomass yield might be enhanced by:
breeding on selected strains from Denmark to improve productivity and biomass yield within one
harvest
improving the technology on seeding the lines by manipulation of fertility leading to more than one
harvest per growth season
Macroalgae potential for energy production
For macroalgae the only relevant technology also in the short to medium term seems to be biogas. The
composition of the algae does not allow for either bio‐diesel or fermentation to e.g. ethanol.
Thermochemical processing may be an option but given the high content of salts it will be technically
difficult to achieve an economic process.
Applying the figures from above producing 1 and 5 mill t of wet algae biomass, the energy yield in a
macroalgae based biogas process would be in the order of 1,8 to 9 PJ. Comparing to e.g. a potential of 40 PJ
based on manure from Danish livestock production the energy potential from macroalgae is low in the
current scenario.
Higher energy production from a given area, require significant breakthroughs in the yield by which we can
grow macroalgae. The current yield from macro algae biomass is limited by both the lack of well developed
systems for marine farming as well as the fact that there has been no breeding of macroalgae varities as for
agricultural crops. Most likely it will be possible to increase the energy production from a given area but it
will need substantial investments not only in marine farming but also in basic science and knowledge within
the field of algae.

76
13. Recommendations
The use of algae as an alternative source for bioenergy has a large potential. Looking at algae as a
photosynthetic unit for production of biomass it could provide billions tons of biomass without
compromising food supply or take up more agricultural land.
But as can be seen from this report, it is clear that the use of algae biomass for energy is still in its infancy.
Neither the technology nor the biology for a large scale production of algae based bioenergy is ready at this
period in time.
In the medium to long term algae biomass can be developed to a scale where it may provide cost effective
production of energy. However, large efforts within basic research on algae biology, marine farming and
conversion technologies will be needed. Also it will be necessary to apply the very latest techniques within
molecular biology as well as breeding in order to overcome some of the constraints facing large scale
production of algae biomass.
A possible commercial approach without investing in full‐scale programs for algae development would be
to monitor the development within algae technology in order to spot new opportunities. Special attention
should be given to potential synergies with other large scale applications e.g. animal feed protein derived
from algae.
Specifically within the areas of algae production and conversion technologies, the following comments and
recommendations are made.
Algae production
For micro‐ and macro‐algae cultivation both on land and off shore several questions need to be answered:
Which species?
Which cultivation techniques?
Harvesting technique?
Where to do it? Land‐ or sea‐based? Where to find the optimal localities?
To identify ideal cultivation species cross points (or compromises?) for maximal yields may need to be
mapped, for instance:
‐ A high growth rate with high lipid production is desirable, but when the growth rates rise with
increasing available nitrogen, the lipid production is positively correlated to nitrogen starving.
‐ An algae species which is easy to cultivate and easy to extract/convert energy from could be
hypothetic as these requirements might be represented by opposing properties, e.g. robust cell

77
walls may increase the cultivation robustness but may at the same time increase the difficulties in
extracting oil.
‐ The extremely high variety of carbohydrates in e.g. macroalgae emphasises the potential of
utilisation possibilities which have not yet been mapped. However, this variety and the seasonality
in the carbohydrate content also make it difficult to identify which species and compound to focus
on.
Thus, what does an ideal species look like and is it already present in nature?
Several cultivation techniques are developed and tested to some extent (with a limited number of species),
and they may all have their pros and cons, for instance:
‐ Open ponds are relatively cheap and the efficiency may be relasonably highs as seen in Israel (see
Chapter 11). However, the demands on land area as well as cultivation robustness of species due to
high risk of contamination may be high, which limits the suitability of this cultivation technique in
countries with a limited area of non‐arable land such as Denmark as well as the number of algal
species candidates.
‐ Photobioreactors (flat plates, tubes) are effective systems which provide controlled environments
and low risk of contamination. However, these expensive cultivation systems struggle with
unwanted high levels of O2 and low CO2 as well as temperature. Furthermore, cleaning and
sterilisation of the cultivation containers may provide sensitive and costly steps in the production.
‐ Sea‐based cultivation of macroalgae has many advantages such as, eg, no use of arable land and
utilisation of the surplus of nutrients in the coastal waters. However, the production is extensive
and also in these open cultivation systems the risk of contamination (fouling) is high.
Thus, the available production systems still seem to have childhood diseases according to productivity and
costs.
At present the ideal species or techniques seem not be identified, but the research over the last 20‐30
years may provide a base upon which qualified research suggestions/proposals can be developed and from
which the continued research should rise.
According to state‐of‐the‐art, identification/description/development of the ideal species needs to go hand
in hand with development of cultivation and conversion technologies and during this process the ideal
species and technologies might change accordingly.
Conversion technology development
The photosynthetic potential of algae allows for bulk production of biomass. Given that production
methods and technologies are developed to a level providing economically efficient production of algae
biomass at the million tons scale algae are capable of providing large amounts of bulk biomass for
bioenergy. However, this is by itself not sufficient as the current conversion technologies for biomass to
energy carriers cannot provide efficient conversion of algae biomass. This does not imply that these issues

78
cannot be overcome and new conversion technologies developed, but currently they are bottlenecks
preventing an efficient large scale use of algae biomass.
Fermentation to liquid energy carriers
Conversion of biomass to liquid fuels such as ethanol and butanol requires fermentable carbohydrates. And
though the content of carbohydrates in algae appears high, the levels of carbohydrate polymers such as
mannans, starch and cellulose that can be hydrolyzed into fermentable monomers are low, typically less
than 10%. Compared to land based plants the structural carbohydrates in algae are heterogeneous, highly
sulfonated and with organic acid side groups. There are no known microorganisms capable of fermenting
these carbohydrates to alcohols, nor do we have enzymes available capable of deconstructing the algae cell
wall carbohydrates to monomers.
Thus one is faced with two options for developing conversion technologies for fermentation of algae
biomass; either we develop a new range of biotechnological tools in the form of industrial microorganisms
and enzymes or we develop and breed new strains of algae capable of producing fermentable sugars. A
new set of biotechnological tools more or less requires that one start from scratch, whereas breeding of
algae high in fermentable sugars can be based on already established knowledge and technologies.
Therefore the initiation of dedicated and large breeding programmes for algae are suggested to be the
most sensible and efficient approach enabling conversion of algae biomass to liquid energy carriers. A
continuous research effort through 10‐25 years will be necessary in order to breed commercially viable
micro‐ or macroalgae strains. That being said, technical issues still remains with regard to handling and
storage as well as the compatibility of enzymes and microorganisms to the inevitable residual salts in
especially the marine algae biomass.
Combustion and gasification
Preferably biomass for combustion is composed only of carbon, oxygen and hydrogen with a content of
salts less than 1%. For algae the content of salt (especially for the marine algae), protein and sulphur makes
them a difficult feedstock for combustion processes. The high level of structural proteins and thus nitrogen
and the sulfonated carbohydrates in a number of micro‐ and macroalgae are basic bottlenecks for obtaining
a technically good feedstock. Furthermore the high salt content from both seawater and the cationic
minerals in the cell walls are problematic. The salt levels can be reduced in marine algae by extensive
washing, but a significant amount of residual salts will remain.
For gasification of algae biomass the same technical problems as for combustion will be encountered, and
there are no readily available solutions.
Identical to the liquid biofuels, breeding of new algae strains may improve the properties for combustion
and gasification. However, algae are a very difficult feedstock for this type of conversion, and it is not likely
they will become a significant source. This also means that in the case of residual algae biomass from
biogas, biodiesel or fermentation other uses such as animal feed or fertilizers should be pursued rather
than combustion.

79
Biogas
The most readily accessible conversion technology for algae is the use of methanogenic bacteria for
production of methane. A number of the carbohydrates in algae cell walls can be converted by the bacteria.
The yield is higher than for manure but less than what is found for e.g. whole crop corns. It will be possible
to increase the biogas yield from algae through process optimization, but lower yields than for e.g. whole
crop corns are to be expected. Some technical issues remain for algae biomass; the high level of residual
salts will increase the time in the biogas digester and the sulfonated carbohydrates will increase the
amount of H2S in the biogas requiring more intensive purification.
Biogas from algae seems currently to be the most economical and technically optimal conversion of algae
to a gaseous energy carrier. It is recommended to focus on process development of algae biogas. This can
be done using already existing technologies and methods, why it will be a relatively limited effort to
develop an optimized algae biogas process.
Algae oils
The production of biodiesel from microalgae oils is technically straightforward. The potential is high but
also the technical and biological barriers to be overcome are quite substantial. A number of large public and
private research programmes are addressing these issues, and significant progress is to be expected within
the next decade.
The biological challenge is the fact that, the rate by which algae produce oils is inversely related to the
overall growth, meaning that oil production is induced by nitrogen starvation. It is therefore not in present
time possible to obtain a high overall growth rate and a simultaneous high oil production. A number of
projects have addressed this issue by metabolic engineering of existing strains or by searching for new
strains capable of both having a high biomass growth rate as well as high oil productivity. Recently some
advances have been reported, but still major developments will be necessary.
The technical challenge of algae biodiesel is the extraction of the oils. The small size of the microalgae
combined with their cell wall structures makes simple mechanical pressing as done for e.g. rape seed
biodiesel not possible. The techniques in use today are all based on extraction with organic solvents. The
problem of using organic solvents for extraction is not only the cost, but also the scale of the production.
Other possibilities may be the use of supercritical CO2 for extraction, but they are still under development
and quite costly.
Though the challenges and barriers for algae biodiesel are substantial, the R&D investments are at such a
high level that they may become a commercial reality within 10‐20 years. But what has been somewhat
overlooked is the fact that if this is to happen also the residual biomass form the biodiesel production will
need to be processed into biogas, feed and fertilizers. Therefore equal attention is required into the
processing of residual algae biomass.

80
14. Literature
Acién FG, González CV, Fernández JM, García‐González M, Moreno J, Sierra E, Guerro MG, Molina E. 2008.
Removal of CO2 from flue gases coupled to the photosynthetic generation of biomass and
exopolysaccharides by cyanobacteria. 11th International Conference on Applied Phycology, Galway, June
2008. Abstracts: 138
Adams J, Gallagher J, Donnison I. 2008. Fermentation studies on selected macroalgae species for bioethanol
production. 11th International Conference on Applied Phycology, Galway, June 2008. Abstracts: 95
Aizawa M, asaoka, K, Atsumi M, Sakou T. 2007. Seaweed Bioethanol Production in Japan – The Ocean
Sunrise Project. Report, 5pp
Banerjee A, Sharma R, Christ Y, Banerjee UC. 2002. Botryococcus braunii: a renewable source of
hydrocarbons and other chemicals. Critical Reviews in Biotechnology 22: 245‐279.
Bartsch I, Wiencke C, Bischof K, Buchholz CM, Buck BH, Eggert A, Feuerpfeil P, Hanelt D, Jacobsen S, Karez
R, Karsten U, Molis M, Roleda MY, Schubert H, Schumann R, Valentin K, Weinberger F, Wiese J. 2008. The
genus Laminaria sensu lato: recent insights and developments. European Journal of Phycology 43: 1‐86
Beardall J, Morris I. 1976. The concept of light intensity adaptation in marine phytoplankton: some
experiments with Phaeopdactylum triconutum. Mar Biol 37: 377‐388
Becker EW. Microalgae: Biotechnology and Microbiology. 1994. Cambridge University Press
Ben‐Amotz A. 2009. Presentation at WSA; International Workshop on Offshore Algae for Biofuels & Beyond,
Maribo; Denmark, in April.
Ben‐Amotz A, Tornabene TG,Thomas WH. 1985. Chemical profile of selected species of microalgae with
emphasis on lipids. Journal of Phycology 21: 72–81
Benemann JR, Oswald WJ. 1996. Systems and economic analysis of microalgae ponds for conversion of CO2
to biomass. Final report to the US Department of Energy, Pittsburgh Energy Technology Center
Blouin N, Xiugeng F, Peng P, Yarish C, Brawley SH. 2007. Seeding nets with neutral spores of the red alga
Porphyra umbilicalis (L.) Kützing for use in integrated multi‐trophic aquaculture (IMTA). Aquaculture 270:
77–91
Birkeland M. 2009. Biofiltering efficiency and primary production by Saccharina latissima, estimated from
mathematical modelling and experimental cultivation near a sea cage farm. Department of Biology,
University of Copenhagen. MSc Thesis, 28 pp
Bohlmann JT, Lorth CM, Drews A, Buchholz R. 1999. Microwave high pressure thermochemical conversion
of sewage sludge as an alternative to incineration. Chem Eng Technol 21: 404‐409.
Bolton JJ, Robertson‐Andersson D, Shuuluka D, Kandjengo L. 2008. Growing Ulva as a commercial crop for
Abelone feed in South Africa. 11th International Conference on Applied Phycology, Galway, June 2008.
Abstracts: 153

81
Borowitzka MA. 2008. Algae and biofuels: quo vadis? 11th International Conference on Applied Phycology,
Galway, June 2008. Abstracts: 1
Briand X, Morand P. 1997. Anaerobic digestion of Ulva sp. 1. Relationship between Ulva composition and methanisation. Journal of Applied Phycology 9: 511-524 Buch BH. 2009. Presentation at WSA; International Workshop on Offshore Algae for Biofuels & Beyond,
Maribo; Denmark, in April.
Buch BH, Buchholz CM. 2004. The offshore‐ring: A new system for the open ocean aquaculture of
macroalgae. Journal of Applied Phycology 16: 355‐368.
Casadevall E, Dif D, Largeau C, Gudin D, Chaumont D, Desanti O. 1985. Studies on batch and continuous
cultures ofBotryococcus braunii: Hydrocarbon production in relation to physiological state, cell
ultrastructure and phosphate nutrition. Biotechnol Bioeng 27:286–295
Chen F. 2008. High‐cell density heterotrophic culture of microalgae for mass production of long‐chain
polyunsaturated fatty acids. 11th International Conference on Applied Phycology, Galway, June 2008.
Abstracts: 11
Cheng‐Wu Z, Zmora O, Kopel R, Richmond A. 2001. An industrial‐size flat plate glass reactor for mass
production of Nannochloropsis sp. (Eustigmatophyceae). Aquaculture 195: 35‐49.
Chiu S‐Y, Kao C‐Y, Chen C‐H, Kuan T‐C, Ong S‐C, Lin C‐S. 2008. Reduction of CO2 by a high‐density culture of
Chlorella sp. In a semicontinous photobioreactor. Bioresource Technology 99: 3389‐3396.
Chiu S‐Y, Kao C‐Y, Tsai M‐T, Ong S‐C, Chen C‐H, Lin C‐S. 2009. Lipid accumulation and CO2 utilization of
Nannochloropsis oculata in response to CO2 aeration. Bioresource Technology 100: 833‐838.
Chopin T, Robinson S, Sawhney M, Bastarche S, Belyea E, Shea R, Armstrong W, Stewart I, Fitzgerald P.
2004. The AquaNet integrated multi‐trophic aquaculture project: rationale of the project and development
of kelp cultivation as the inorganic extractive component of the system. Proceedings of the Integrated
Multi‐TRophic Aquaculture Workshop, Saint John, NB March 2004: 11‐19
Christi Y. 2007. Biodiesel from microalgae. Biotechnology Advances 25:294‐306.
Christi Y. 2008. Biodiesel from microalgae beats bioethanol. Trends in Biotechnology 26: 126‐131
Chrispeels MJ, Sadava DE 2003. Plants, genes and crop biotechnology. Jones and Bartlett Publishers,
Boston, MA.
Chynoweth DP. 2002. Review of Biomethane from Marine Biomass. Draft report. University of Florida. 207
pp
Cole JJ. 1982. Interactions between bacteria and algae in aquatic ecosystems. Ann Rev Ecol Syst 13: 291‐314
Critchley AT, Ohno M, Largo DB (eds). 2006. World Seaweed Resources. An authorative refrence system.
DVD‐ROM, ETI.
Davison IR. 1991. Environmental effects on algal photosynthesis: temperature. Journal of Phycology 27: 2‐8
Dayananda C, Sarada R, Bhattacharya S, Ravishankar GA. 2005. Effect of media and culture conditions on
growth and hydrocarbon production by Botryococcus braunii. Process Biochemistry 40: 3125‐3131.

82
Demetropoulos CL, Langdon CJ. 2004. Enhanced production of Pacific dulse (Palmaria mollis) for co‐culture
with abalone in a land‐based system: effects of seawater exchange, pH, and inorganic carbon
concentration. Aquaculture 235: 457‐470
Domozych DS, Stewart KD, Mattox KR. 1980. The comparative aspects of cell wall chemistry in the green
algae (Chlorophyta). Journal of molecular evolution 15 1‐12
Doucha J, Straka F, Lívanský K. 2005. Utilization of flue gas for cultivation of microalgae (Chlorella sp.) in an
outdoor open thin‐layer photobioreactor. Journal of Applied Phycology 17: 403‐412
Douskova I, Doucha J, Livansky K, Machat J, Novak P, UmysovaD, Zachleder V, Vitova M. 2009.
Simultaneous flue gas bioremediation and reduction of microalgal biomass production costs. Applied
Microbiology and Biotechnology 82: 179‐185
Dring MJ. 1986. Stimulation of light‐saturated photosynthesis in Laminaria (Phaeophyta) by blue light.
Journal of Phycology 25: 254‐258
Edwards M. 2008. Furhter advances in the cultivation techniques of Palmaria palmate, Laminaria digitata
and Porphyra sp. in Ireland. 11th International Conference on Applied Phycology, Galway, June 2008.
Abstracts: 152
Emerson W, Arnold J. 1932a. A separation of the reactions in photosynthesis by means of intermittent light.
The Journal of General Physiology 15: 391-420
Emerson W, Arnold J. 1932b. The photochemical reaction in photosynthesis. The Journal of General
Physiology 16: 191-205
Erting L, Daugbjerg N, Pedersen PM. 2004. Nucleotide diversity within and between four species of
Laminaria (Phaeophyceae) analysed using partial LSU and ITS rDNA sequences and AFLP. European Journal
of Phycology 39: 243‐256
Fei XG, Pang SJ, Bao Y, Li DPl. 2007. How to use seaweeds as a CO2 sink in the sea. XIXth Internation Seaweed Symposium, Kobe Japan March 2007. Abstracts: 24 Fernandes AV, Vargas G, Alarcon N, Velasco A. 2007. Evaluation of marine algae as a source of biogas in a two-stage anaerobic reactor system. Biomass and Bioenergy 32: 338-344 Fouchard S, Pruvost J, van Vooren G, Legrand J. 2008. Evaluation of Neochloris oleoabundis lipids (TAGs)
productivities in photobioreator for biodiesel application. 11th International Conference on Applied
Phycology, Galway, June 2008. Abstracts: 92
Gao K, McKinley KR. 1994. Use of macroalgae for marine biomass production and CO2 remediation: a
review. Journal of Applied Phycology 6: 45‐60
González C, Acién FG, Fernández JM, Cerón MC, Sierra E, Sánchez JF, Molina E. 2008. Comparative analysis
of the carbon fixation rate of several microalgae and cyanobacteria. 11th International Conference on
Applied Phycology, Galway, June 2008. Abstracts: 233
Grobbelaar JU. 2008. Upper limit of photosynthetic productivity and problems of scaling. 11th International
Conference on Applied Phycology, Galway, June 2008. Abstracts: 26

83
Grobbelaar JU, Nedbal L, Tichý V. 1996. Influence og high frequency light/dark fluctuations on
photosynthetic characteristics of microalgae photoacclimated to different light intensities and implications
for mass algal cultivation. Journal of Applied Phycology 8: 335‐343
Grossart H‐P. 1999. Interactions between marine bacteria and axenic diatoms (Cylindrotheca fusiforme,
Nitzschia laevis and Thalassiosira weissflogi) incubated under various conditions in the lab. Aquatic
Microbial Ecology 19: 1‐11
Gunn D.2007. The Scottish Association for Marine Science. PPT presentation. 24 slides
Haiduc AG, Brandenberger M, Suquet S, Ludwig C, Vogel F, Bernier‐Latmani R, Stucki S. 2008. Hydrothermal
methane from microalgae. 11th International Conference on Applied Phycology, Galway, June 2008.
Abstracts: 141
Hall DO, Mynick HE, Williams RH. 1991. Cooling the greenhouse with bioenergy. Nature 353: 11‐12
Hanniffy D, Kraan S. Intensive tank cultivation of Porphyra spp. 2008. 11th International Conference on
Applied Phycology, Galway, June 2008. Abstracts: 189
Horn SJ, Aasen, IM, Østgaard K. 2000. Production of ethanol from mannitol by Zymobacter palmae. Journal
of Industrial Microbiology & Biotechnology 24: 51‐57.
Hossain ABMS, Salleh A, Boyce AN, Chowdhury P, Naqiuddin M. 2008. Biodiesel fuel production from algae
as renewable energy. American Journal of Biochemistry and Biotechnology 4: 250‐254
Hu Q, Sommerfeld M, Jarvis E, Ghirardi M, Posewitz M, Seibert M, Darzins A. 2008. Microalgal triacy‐
lglycerols as feedstocks for biofuel production: perspectives and advances. The Plant Journal 54: 621‐639.
Hwang EK, Baek JM, Park CS. 2008. Potential and utilization of the mass cultivation of Ecklonia stolonifera
Okamura as a summer feed for Abalone industry in Korea. 11th International Conference on Applied
Phycology, Galway, June 2008. Abstracts: 290
Hällfors G.2004. Checklist of Baltic Sea phytoplankton species (including some heterotrophic protistan
groups). Baltic Sea Environment Proceedings 95. 210 pp
Illman AM, Scragg AH, Shales SW. 2000. Increase in Chlorella strains calorific values when grown in low
nitrogen medium. Enzyme Microb 27: 631–635
Jones JG. 1972. Studies on freshwater micro‐organisms: phosphatase activity in Lakes of Differing Degrees
of Eutrophication. Journal of Ecology 60: 777‐791
Kraan S, Guiry M. 2004. Presentation at ISS; XVIII International Seaweed Symposium, Bergen, Norway, in
June.
Kraan S & Guiry M. 1998. Strain selection in the edible brown seaweed Alaria esculenta: Genetic
fingerprinting and hybridization studies under laboratory conditions. Marine Resource Series. 30 pp.
Kraan et al. 2000
Kristiansen A, Pedersen PM, Moseholm L.1994. Salinity–temperature effects on growth and reproduction of
Scytosiphon lomentaria (Fucophyceae) along the salinity gradient in Danish waters. Phycologia 33: 444–454
Lahaye M, Robic A. 2007. Structure and functional properties of Ulvan, a polysaccharide from green
seaweeds. Biomacromolecules 8, 1765‐1774

84
Lee Y‐K. 2001. Microalgal mass culture systems and methods: their limitations and potential. Journal of
Applied Phycology 13: 307‐315
Lehr F, Ivanova D, Hankamer B, Kruse O, Posten C. 2008. Kinetic investigations for hydrogen production
with Chlamydomonas reinhardtii. 11th International Conference on Applied Phycology, Galway, June 2008.
Abstracts: 31
Leigh EO (jr), Paine RT, Quinn JF, Suchanek TH. 1987. Wave energy and intertidal productivity. Proc Natl
Acad Sci USA 84: 1314‐1318
Levy I, Israel A, Friedländer M. 2008. Noritech’s Porphyra cultivation technology. 11th International
Conference on Applied Phycology, Galway, June 2008. Abstracts: 156
Levy B, Friedlander M. 2008. Identification of two putative adhesive polypeptides in Caulerpa prolifera
rhizoids using an adhesion model system. Journal of Applied Phycology 16: 1‐9
Li Q, Du W, Liu D. 2008. Perspectives of microbial oils for biodiesel production. App Microbiol Biotechnol
80: 749-756
Lien S, Roessler P. 1986. The energetics of biomass and lipid production by lipogenic microalgae under
nitrogen deprivation. Aquatic Species Program Review: Proceedings.
Liu Z‐Y, Wang G‐C, Zhou B‐C. 2008. Effect of iron on growth and lipid accumulation in Chlorella vulgaris.
Bioresource Technology 99: 4717‐4722
Liot F, Colin A, Mabeau, S. 1993. Microbiology and storage life of fresh edible seaweeds. Journal of Applied
Phycology 5: 243–247.
Lüning K. 2008. Integrated macroalgae‐oyster aquaculture on a North Sea Island: seasonal productivity of
the brown alga Laminaria saccharina and the red algae Palmaria palmate; Soliera chordalis; Gracelaria
vermiculophylla, and the use of these seaweeds in human nutritions or as raw material for the cosmetic
industry. 11th International Conference on Applied Phycology, Galway, June 2008. Abstracts: 151
Lünnig K. 1990. Seaweeds. Their environment, biogeography, and ecophysiology. John Wiley & Sons, Inc.
New York. 527 pp.
Lüning K, Dring MJ. 1978. Reproduction induced by blue light in female gametophytes of Laminaria
saccharina. Planta 104: 252‐256
Lüning K, Müller DC. 1978. Chemical interaction in sexual reproduction of several laminariales
(Phaeophyceae): release and attraction of spermatozoids. Aquatic ecology: plant genetics and breeding 89:
333‐341
Lüning K, Kadel P, Pang SJ. 2008. Control of reproduction rythmicity by environmental and endogenous
signals in Ulva pseudocurvata (Chlorophyta). Journal of Phycology 44: 866‐873
Neale PJ, Melis A. 1986. Algal photosynthetic membrane complexes and the photosynthesis‐irradiance
curve: a comparison of light‐adaptation response in Chlamydomonas reinhardtii (Chlorophyta). Journal of
Phycology 22: 531‐538
Nedbal L, Tichý V, Xiong F, Grobbelaar JU. 1996. Microscopic green algae and cyanobacteria in high‐
frequency intermittent light. Journal of Applied Phycology 8: 325‐333

85
Neefus CD, Day JP, Walker AB, Berlinsky DL, Yarish C. 2008. Optimization of an integrated cod/nori
recirculating aquaculture system. 11th International Conference on Applied Phycology, Galway, June 2008.
Abstracts: 54
Nielsen R, Kristiansen Aa, Mathiesen L & Mathiesen H. 1995. Distributional index of the benthic macroalgae
of the Baltic Sea area. Acta Bot Fen 155: 1‐51.
McGillivary P. 2009. Presentation at WSA; International Workshop on Offshore Algae for Biofuels & Beyond,
Maribo; Denmark, in April.
Metzger P, Largeau C. 2005. Botryococcus braunii: a rich source for hydrocarbons and related ether lipids.
Appl Microbiol Biotechnol 66: 486‐496.
Minowa T, Yokoyama S, Kishimoto M, Okakurat T. 1995. Oil production from algal cells of Dunaliella
tertiolecta by direct thermochemical liquefaction. Fuel 74: 1735‐1738
Morrissey J. 2002. Roaringwater Bay Pilot Seaweed Cultivation Project Phase I, 2001‐2002. Irish Seaweed
Centre,full report, 90 pp
Mortensen AM, Wegeberg S. 2007. Forsøgsdyrkning af Alaria esculenta på Færøerne. Rapport. 12 pp.
Olrik K. 2009. Presentation at WSA; International Workshop on Offshore Algae for Biofuels & Beyond,
Maribo; Denmark, in April.
Pang SJ, Lüning K. 2004. Breaking seasonal limitation: year‐round sporogenesis in the brown alga Laminaria
saccharina by blocking the transport of putative sporulation inhibitors. Aquaculture 240: 531‐541
Park CS, Kakinuma M, Amano H. 2001. Detection of red rot disease fungi Phytium spp. by polymerase chain
reaction. Fisheries Science 67: 197‐199
Pate RC. 2009. Presentation at WSA; International Workshop on Offshore Algae for Biofuels & Beyond,
Maribo; Denmark, in April.
Patil V, Tran K‐H, Giselrød HR. 2008. Towards sustainable production of biofuels from microalgae.
International Journal of Molecular Sciences 9: 1188‐1195
Pedersen P M. 1976. Marine, benthic algae from southernmost Greenland. Meddel. Grønland 199: 1–80
Peer M, Skye R, Thomas H, Evan S, Ute C, Jan H, Clemens P, Olaf K, Ben H. 2008. Second generation
biofuels: high‐efficiency microalgae for biodiesel production. Bioenergy Research 1:20–43
Peterson AA, Vogel F, Lachance RP, Fröling M, Antal MJ (jr), Tester JW. 2008. Thermochemical biofuel
production in hydrothermal media: a review of sub‐ and supercritical water technologies. Energy &
Environmental Science 1: 32‐54
Prince RC, Kheshgi HS 2005. The photobiological productivity of hydrogen: potential efficiency and
effectiveness as a renewable fuel. Critical Reviews in Microbiology 31: 19‐31.
Rasmussen MB, Fossing H, Markager S, Bruhn A. 2009. Anvendelse af marine algaer til fremstilling af
biobrændstof. 15. Danske Havforskermøde, Helsingør, January 2009. Abstracts p.67
Ratcliff J, Kraan S. 2008. Supergen II: bioenergy marine biomassproject. 11th International Conference on
Applied Phycology, Galway, June 2008. Abstracts: 177

86
Reed DC. 1990. The effects of variable settlement and early competition on patterns of kelp recruitment.
Ecology 71: 776‐787
Robertson‐Andersson DV, Potgieter M, Hansen J, Bolton JJ, Troell M, Anderson RJ, Halling C, Probyn T.
2008. A pilot‐scale integrated seaweed (Ulva lactuca) ans Abalone (Haliotis midae) recirculation system.
11th International Conference on Applied Phycology, Galway, June 2008. Abstracts: 57
Robertson‐Andersson DV, Potgieter M, Hansen J, Bolton JJ, Troell M, Anderson RJ, Halling C, Probyn T.
2008. Integrated seaweed cultivation on an abalone farm in South Africa. Journal of Applied Phycology 20:
129‐145
Rodolfi L, Zittelli GC, Bassi N, Padovani G, Biondi N, Bonini G, Tredici MR. 2009. Microalgae for oil: strain
selection, induction of lipid synthesis and outdoors mass cultivation in a low‐cost photobioreactor.
Biotechnology and Bioengineering 102: 100‐112.
Sánchez et al. 2008. Seaweed culture of Ulva spp. (Enteromorpha): experimental study in laboratory, open‐
air pond and open‐water in The Bay. 11th International Conference on Applied Phycology, Galway, June
2008. Abstracts: 186
Sánchez Mirón A, Garcı́a MCC, Gómez AC, Camacho FG, Grima EM, Christi Y. 2003. Shear stress
tolerance and biochemical characterization of Phaeodactylum tricornutum in quasi steady‐state continous
culture in outdoor photobioreactors. Biochemical Engineering Journal 16: 287‐297
SERI. 1984. Microalgae Culture Collection 1984‐1985. Solar Energy Research Institute, Golden, Colorado,
Publication SERI/SP‐231‐2486, 59 pp.
Sharp G, Allard M, Lewis A, Semple R, Rochefort G. 2008. The potential for seaweed resource development
in subarctic Canada; Nunavik, Ungava Bay. Journal of Applied Phycology 20: 41‐48
Shea R, Chopin T. 2007. Effects of germanium dioxide, an inhibitor of diatom growth, on the microscopic
laboratory cultivation stage of the kelp, Laminaria saccharina. Journal of Applied Phycology 19: 27‐32.
Sheehan J, Dunnahay T, Benemann J, Roessler P. 1998. A look back at the US Department of Energy’s
Aquatic Species Program – Biodiesel from Algae. National Renewable Energy Laboratory, report NREL/TP‐
of La Paz, Baja California, Mexico.
Shifrin NS, Chisholm SW. 1981. Phytoplankton lipids: interspecific differences and effects of nitrate, silicate
and light‐dark cyckles. Journal of Phycology 17: 374‐384
Sierra E, Acién FG, Fernández JM, García JL, González C, Molina E. 2008. Characterization of a flat plate
photobioreactor for the production of microalgae. Chemical Engineering Journal 138: 136‐147
Steen H. 2005. Stortare. Kyst og Havbruk 2; Kystressurser: 111‐114
Sukenik A, Wahnon R. 1991. Biochemical quality of marine unicellular algae with special emphasis on lipid
composition. I. Isochrysis galbana. Aqauculture 97: 61‐72
Tamigneaux E, Gendron L. 2008. Cultivation of kelp in Chaleurs Bay (Quebec, Canada): influence if invading
epibionts (Membranipora membrancea) on the growth of Saccharina longicruris (Bachelot de la Pylaie)
Küntze. 11th International Conference on Applied Phycology, Galway, June 2008. Abstracts: 255

87
Terry KL. 1986. Photosynthesis in modulated light: quantitative dependence of photosynthetic
enhancement on flashing rate. Biotechnology and Bioengineering 28: 988‐995
Thomsen HA. 1992. Plankton i indre danske farvande. En analyse af forekomsten af alger og heterotrofe
protister (ekskl. ciliater) i Kattegat. Havforskning fra Miljøstyrelsen 11. Miljøstyrelsen, Miljøministeriet, 340
pp
Torzillo G, Giannelli L, Scoma A. 2008. Optimization of H2 production with C. reinhardtii in laboratory
photobioreactors. 11th International Conference on Applied Phycology, Galway, June 2008. Abstracts: 140
Tredici MR. 2008. Maximum and actual productivities of out‐door algae. 11th International Conference on
Applied Phycology, Galway, June 2008. Abstracts: 14
Trent J. 2009. Presentation at WSA; International Workshop on Offshore Algae for Biofuels & Beyond,
Maribo; Denmark, in April.
Ugwu CU, Aoyagi H, Uchiyama H. 2008. Photobioreactors for mass cultivation of algae. Bioresource
Technology 99: 4021‐4028
Vairappan CS, Chung CS, Hurtado AQ, Soya RE, Lhonneur GB, Critchley A. 2008. Distribution and symptons
of epiphyte infection in major carrageenophyte‐producing farms. Journal of Applied Phycology 20: 27‐33
Wegeberg S. 2010. Cultivation of kelp species in the Limfjord, Denmark. Department of Biology, University
of Copenhagen. 12 pp
Wegeberg S.2007. Er tang en ny marin ressource i Grønland? Vand & Jord 3: 117‐120
Wegeberg S, Felby C. 2010. Introduktion til Alger. University of Copenhagen. 32 pp
Weismann JC. 2008. Factors limiting photosynthetic efficiency in outdoor mass cultures of microalgae. 11th
International Conference on Applied Phycology, Galway, June 2008. Abstracts: 15
Werner A, Kraan S. 2004. Review of the potential mechanization of kelp harvesting in Ireland. Marine
Environment and Health Series 17. 52 pp
Werner A, Clarke D, Kraan S. 2003. Strategic Review of the Feasibility of Seaweed Aquaculture in Ireland.
Marine Institute, Technology Park, Parkmore, Galway, Ireland (www.marine.ie/marinertdi). Report, 120 pp
Wu CY, Pang SJ. 2006. The Seaweed Resources of China
Xu H, Miao X, Wu Q. 2006.High quality biodiesel from a microalga Chlorella protothecoides by
heterotrophic growth in fermenters. Journal of Biotechnology 126: 499‐507
Ying L, Kang‐sen M, Shi chun S. 2002. Effect of harvest stage on the total lipid and fatty acid composition of
four Cylindrotheca strains. Chinese Journal of Oceanology and Limnology 20: 157‐161
Yu H, Jia S, Dai Y. 2009. Growth characteristics of the cyanobacterium Nostoc flagelliforme in
photoautotrophic, mixotrophic and heterotrophic cultivation. Journal of Applied Phycology 21: 127‐133
Zertuche‐González JA, García‐Lepe G, Ballesteros‐Grijalva G, Guzmán‐Calderón JM. 2008. In situ growth rate
of the commercial kelp Eisenia arborea Areschoug within a coastal lagoon in Baja California, Mexico. 11th
International Conference on Applied Phycology, Galway, June 2008. Abstracts: 257

88
Appendix 1
Omkostninger for tanganlæg. Havnanlægsdesign følger forlæg fra Irish Seaweed Centre på 1 ha. Priser er indhentet fra bl.a Hvalpsund Net og Vónin.
Vareforbrug 1 år 2 år NOTER
Seed lines 1 ha Spoler a 220m Pris
Liner 4000 m 18 245 4410 4410
Hvide kar 4 2500 10000 1 l havvand 1 m line
Projektør 10 1500 15000 Lysintensitet > 25 mmol fotoner/m2/sek.
Beluftning Akvariepumper 8 500 4000 *Køb af sterilfiltreret havvand
Vandbehandling* 5000 5000
Laboratorievægt 2000 ml/l Pr. gang I alt ml l
Drift laboratorie (kemikalier, filtre, dunke) 4000 4000 ESI 20 80000 240000 240
El 2000 2000 Vareforbrug i alt 46410 15410
Havanlæg 1 ha stks/m pris (DKK) Eurosteel tov 12mm 2000mtr = 3.500,00 DKK
Cementblokke 1.5 tons (2 tons) 8 2000 16000 Eurosteel tov 14mm 2000mtr = 4.895,00 DKK
Cementblokke 5 tons (4 tons) 4 4000 16000
Wire 14mm 260 2447,5 2447,5 8 x dybde + 20 m i mellem dobbelt-blokke i hjørnerne Reb 16mm 2000 2,5 5000 Rebpriser: (dybde 30 m) Reb 18 mm 200 707 16mm DKK 550/220 2,5Reb til ramme 50mm 400 22,5 9000 Bøjer 1000 l 4 1500 6000 18mm DKK 707/220m 3,213636Bøjer 200 l 4 860 3440 10mm DKK 245/220 1,113636Bøjer 25 l 30 146 4380 50mm DKK 4954/220 m 22,51818
Havanlæg i alt 28527 Bøjer B 40 DAN-HILL 23 liter = 146,00 DKK pr stk.
Bøjer B 100 DAN-HILL 280 liter = 860,00 DKK pr stk.
Bøjer B 135 DAN-HILL 675 liter = 1.500,00 DKK pr stk.
Omkostninger i alt 74937 15410
Top Related