ww2.biol.sc.eduww2.biol.sc.edu/~elygen/biol303/2014 term papers/Ling... · Web viewOkuma M, Mochida...
20
Campbell 1 Ling Campbell Dr. Ely Biology 303 31 October 2014 Regeneration at the Genetic Level Tissue engineering and regenerative medicine are becoming increasingly popular. The purpose of tissue engineering is to restore, maintain, or improve damaged tissues or even whole organs through looking at the functional units of the body. Regenerative medicine is a field that includes tissue engineering, but also looks to see how bodies can naturally self-heal. Advances in this field could positively affect millions of lives. Tissue engineering and regenerative medicine tend to focus on the basic unit of life, cells. Cells are the building blocks of tissues, and understanding how cells work is the main framework for any regenerative medicine advances. (“Tissue Engineering and Regenerative Medicine”) The central dogma of molecular biology states that DNA is transcribed to mRNA, which is then translated to proteins. Thus, every functional unit of the cell begins with the content of the DNA. Genes are the units of hereditary information that make up DNA. By studying the genes associated with tissue regeneration, scientists will be able to see what is causing the tissue degradation at a molecular level. Understanding how the problem starts is the
Transcript of ww2.biol.sc.eduww2.biol.sc.edu/~elygen/biol303/2014 term papers/Ling... · Web viewOkuma M, Mochida...

Campbell 1
Ling Campbell
Dr. Ely
Biology 303
31 October 2014
Regeneration at the Genetic Level
Tissue engineering and regenerative medicine are becoming increasingly popular.
The purpose of tissue engineering is to restore, maintain, or improve damaged tissues or
even whole organs through looking at the functional units of the body. Regenerative
medicine is a field that includes tissue engineering, but also looks to see how bodies can
naturally self-heal. Advances in this field could positively affect millions of lives. Tissue
engineering and regenerative medicine tend to focus on the basic unit of life, cells. Cells
are the building blocks of tissues, and understanding how cells work is the main
framework for any regenerative medicine advances. (“Tissue Engineering and
Regenerative Medicine”)
The central dogma of molecular biology states that DNA is transcribed to mRNA,
which is then translated to proteins. Thus, every functional unit of the cell begins with the
content of the DNA. Genes are the units of hereditary information that make up DNA. By
studying the genes associated with tissue regeneration, scientists will be able to see what
is causing the tissue degradation at a molecular level. Understanding how the problem
starts is the first step to solving it. Possible treatments can range from determining how
the body naturally solves a problem and capitalizing on it, to artificial regeneration of the
loss tissue. This paper will examine three articles that focus on genes specific to
regeneration of varying types of tissues.
According to “Limb Loss Statistics,” about 2 million people in the United States
are living with some form of amputation. The major causes of limb loss are vascular
diseases (such as diabetes), trauma, and cancer. Although natural, complete limb
regeneration is not possible for humans, other animals such as lizards and zebra fish
possess this amazing capability. In a paper by Elizabeth D. Hutchins et al. (2014),
scientists used a transcriptomic-wide analysis to study which genes are involved in tail
regeneration of the Lizard Anolis carolinensis. Lizards were chosen for this experiment
because of the models capable of regenerating limbs, lizards are the most closely related

Campbell 2
to humans evolutionarily. By studying the genetic regulation of regeneration, scientists
will have an advanced understanding of the process, which is relevant to developing
human therapies. In order to study the genes, sample cells were necessary. To obtain their
samples, autonomy (tail removal) of 5 reptile tails was performed by applying pressure to
the tails until it was separated. Sample sections of the regenerated tails were collected 25
days post autonomy (dpa).
RNA-Seq analysis was used to determine the levels of different genes’ expression
within the regenerating tails. A total of 326 genes were clustered in two distinctive
groups: genes whose expression increased towards the proximal base (Cluster I) and
genes whose expression increased towards the distal tip (Cluster 2) (Figure 1).
The RNA-Seq analysis also investigated Gene Ontology (GO) groups related to tissue
differentiation, specifically skeletal muscle growth, myogenesis, and biological adhesion,
in the base of the tail. The analysis found an increase in the expression of myogenic
regulatory factors related to muscle growth and repair. Some of these transcription
factors, such as pax, mowhawk (mkw), and lcf15, are typically found in myogenic stem or
progenitor cells (Anderson et al. 2009, Fan et al. 2012, Takahashi et al. 2007) while
others like the TGF family member myostain (msln) are involved in modifying muscles
mass (Manceau et al 2008). In particular, the increase of mkx, which demonstrated that
growth is possible with coordination between tendons and muscles in the tail cell, is
promising because this transcription factor is required for growth and repair in mammal
Figure 1. Heatmap of the 326 genes. The 25 dpa tissues were divided into 5 equal segments (S1-S5). Tissue was taken from each of the segments to perform RNA-Seq. There are significant differences in the density of certain genes along the tail. 129 genes had a higher level of expression distally towards the tip (Cluster II; S2 and S1). 197 genes had increased expression at the proximal base of the tail (Cluster I; S5, S4, S5).

Campbell 3
cells, indicating a similarity that could useful to human regeneration (Anderson et al.
2012) (Hutchins et al. 2014). The increased level of expression of these types of genes
and regulatory factors at the base of the tail is logical because the base of the tail is the
first area to grow and differentiate.
Additionally, RNA-Seq analysis identified different levels of expressed genes in
the tip of the tail in GO groups associated with wound response, immune response,
hormonal regulation, and embryonic morphogenesis. The analysis found increased
expression of thyroid hormone generation genes, such as cga and dio2 that are central in
neuromuscular development, both in normal growth and after injury. Because there is a
need for a higher level of tissue differentiation and function at the tip of the tail, the the
higher level of expression of genes that code for these types of traits at this location is
fitting. The high levels of the wnt5a ligand and its receptor ror2 were especially
promising. Recent studies of regeneration have identified that Wnt signaling stimulates
differentiation of cells from skeletal muscles, osteogenic, and cardiogenic lineages
(Cruciat and Niehrs 2013).
At the tip and middle regions of the tail there was a high density of Wnt inhibitors such as
dkk2, igfbp4, wif1, and sgfrp2 (Figure 2). It is possible that the increased distribution of
these inhibitors at the end and center of the tail could create a gradient of Wnt signaling
throughout the tail that is required to constantly sustain the growth region of the
regenerating tail (Hutchins et al. 2014).
Figure 2. Heatmaps based on RNA-Sequence Analysis. Shows the 9 Wnt pathway genes. These genes and their inhibitors have a higher level of distribution at the tip and middle of regions of the regenerating tail, suggesting that the Wnt signaling can be transmitted throughout the entire tail.
Dr. Bert Ely, 11/01/14,
The paper says otherwise. Rewrite this section and the figure legend.

Campbell 4
In order to determine the source of cells for tail regeneration, Hutchins and her
colleagues compared the cells from the regenerating tail to lizard satellite and embryo
cells. They found that the satellite and embryo cells have distinct patterns of gene
expression. Conversely, in the tail cells these genes are expressed at low levels without
any patterns along the length of the tail, suggesting that the tail stem cells are evenly
distributed throughout the tail (Figure 3A,B).
These findings are in accordance with the patterns of cell proliferation and specification
because it has been found by analyzing the distribution of minichromosome maintenance
complex component 3 (MCM3) and proliferating cell nuclear antigen (PCNA), two
markers of proliferation, that growth cells are evenly distributed throughout the
regenerated tail. The end result is that there is no single growth zone within the
regenerating tail (Hutchins et al. 2014). The information gained by this part of the study
is applicable to humans because satellite cells act as stem cells in mammals because some
have the ability to differentiate into skeletal muscle after injury (Askura et al. 2001).
Through the transcriptomic analysis of the lizard tail regeneration, Hutchins et al.
was able to identity genes, transcription factors, and signal pathways that play a pivotal
role in developmental roles such as myogenesis, chondrogenesis, and neurogenesis along
with processes reserved for adults such as responses to the immune system and wound
healing. The model of regeneration discovered in lizards shows a complex process of
formation of advanced developmental processes and specified wound repair that seem to
be unique to this amniote vertebrate. Some of the genes discovered have similar purposes
Figure 3. The expression of Satellite Cell Markers (A) and Embryonic Stem Cell markers (B). Expression of embryonic and satellite cell genes vary with position in the regenerating tail.

Campbell 5
in the human genome, which creates a bridge to applying the knowledge learned from
this study to the development of human therapies.
The study of genes for regenerative therapy does not have to involve regenerating
an entire entity. Instead, it is possible that the regeneration of one small area of tissue can
make a big difference in improving someone’s quality of life. For example, one of the
major causes of lower back pain is degenerative disc disease (DDD) (Luoma et al. 2000).
DDD is linked to intervetebral disc degeneration (IVDD), which decreases the function
and number of working cells in the disc and a reduction of extracellular matrix (ECM)
components such as aggrecan and type II collagen (Zhao et al. 2007). The current
treatments for the disease might temporarily alleviate pain, but in the end they do not fix
the actual problem. Studies have found that Nucleus pulposus (NP) cell transplantation
has been effective to slow down IVDD (Okuma et al. 2000). Unfortunately, it is not
possible to obtain NP cells from healthy normal disks because it might cause
degeneration in the original disc. But, NP cells have a chondrocyte-like phenotype similar
to bone marrow mesenchymal stem cells (BMSCs), which means BMSCs can also
differentiate to have the chondrocyte-like phenotype. Thus, BMSCs should be an
adequate cell source for IVD regeneration (Richardson et al. 2006, Steck et al. 2005,
Hiyama et al. 2008, Sakai et al. 2006, Sakai et al. 2003). Studies have also found that a
decrease in the transcription factor Sox9 found in NP cells, which is pivotal for
chondrogenesis and type II collagen synthesis, is linked to IVDD (Gruber et al. 2009).
Other research has determined that Sox9 gene transfer could stimulate cartilage growth of
the BMSCs (Paul et al. 2003, Tsuchiya et al. 2003). The paper by Sun et al. (2014)
hypothesizes that Sox9-induced BMSCs will have positive effect on IVD repair and
regeneration.
To study the effects of the Sox9 transcription factor, BMSCs were collected from
white rabbits and then cultured. These cells were then either transduced with just green
fluorescent protein (GFP) or both GFP and Sox9 genes. The results of the Sox-9 goups
showed the expression of Sox9, Col II, and Aggrecan increased substantially without
causing hypertrophy, cell enlargement. The BMSCs (both with the Sox9 and without)
were then embedded into chitosan-glycerophosphate (C/Gp) gels. The scientists found a
significant increase in the expression levels of Col II, Aggrecan and Sox9 in Sox9-

Campbell 6
transduced BMSCs in C/Gp gels compared to the increase on GFP-transduced BMSCs at
21 days after transduction. These findings show that Sox-9 transduction can induce
chondrogenic differentiation of BMSCs using the C/Gp gels as a growth medium.
The 50 rabbits in this study were induced with disk degeneration two weeks
before the beginning of treatment using the puncture method. The animals were then
divided into 5 different groups according to their treatments: rabbits with no surgery
(normal), rabbits who had the puncture surgery (degeneration), rabbits with the puncture
surgery and a chitosan-glycerophosphate (C/Gp) injection (C/Gp), rabbits with the
puncture surgery and then GFP-transduced BMSCs on C/Gp gels, and rabbits with Sox-9
transduced BMSCs using the C/Gp transplantation (Sox9). Sun et al. (2014) used
Magnetic resonance imaging (MRI) to visualize the effects of different induced BMSCs
on disk degeneration, using the amount of NP cells as an indicator (Figure 4).
In Figure 4A, the T2-weighted MRI images show that there was a significant decrease in
the size of the NP area for untreated degenerative cells and for the C/Gp groups that had
no additional BMSCs. Conversely, the Sox9 and GFP group showed decreased amounts
of NP degradation. In Figure 4B, the average T2 of the NP cells was higher for the Sox9
Figure 4. Imaging Studies. C/Gp group (A) T2- weighted MRI images at 6 and 12 weeks. (B) The mean of NP at 6 and 12 weeks.

Campbell 7
group than the other three groups. These findings suggest that the addition of Sox9 to
BMSCs can significantly slow down the amount of IVDD.
Through histological evaluations Sun et al. (2014) evaluated the amount of
chondrocyte-like cells and the NP structures. After 6 weeks of treatment, in the
degeneration group and C/Gp group the distinction between the disks was hard to see, but
still present and the number of BMSCs had decreased (Figure 5A-J). In contrast, the
distinction between disks was easily identifiable in the GFP and Sox9 group, and the NP
structure was comparatively well intact. 12 weeks after treatment, the degeneration and
C/Gp groups had no distinct border between disks and there was an inflow of connective
tissue in the gap. The GFP and Sox9 groups managed to maintain divided disks, with the
Sox9 group being slightly more successful. The results from this analysis augment the
idea that Sox9 transduced BMSCs are more capable of preventing disk degeneration.
The results of this experiment suggest that BMSCs exposed to higher levels of
Sox9 can decrease IVDD, meaning that successful stem therapies for this disease could
be possible with the right implantation procedure. Still, there are some limitations to this
study. For example, this injury model might not be similar enough to IVDD in humans.
But although the results from this study are not the absolute cure, Sun et al. were able to
demonstrate that Sox9 gene transfer could enhance the repair effects of BMSCs through
an increase in ECM production. Further studies will be necessary to determine NP-
specific markers in order to test which additional phenotypes should be targeted.
Furthermore, additional studies will be necessary to test if this or any therapy is plausible
long-term.
Another way gene expression can help scientists study regeneration is through the
examination of the genes used to monitor a certain pathway within the cell. The paper by
Zhang et al. (2014) looks at how the Il-18 pathway positively stimulates liver
regeneration. Hepatocyte cells make up the majority of the liver (Sell 2013). When the
liver has undergone a partial hepatectomy (PH) and therefore has lost liver cell mass, the
hapatocytes are activated and replenish this cell mass through liver regeneration (LR)
(Taub 2004). The LR process is divided into three phases: the priming phase that is
typically a half hour to six hours after liver cell removal, the progressing phase that is 6
hours to 72 hours after PH, and the terminal phase that is 72 hours to 168 hours after PH

Campbell 8
(Ma et al. 2009). Interleukin-18 (Il-18), typically associated with defense against
infection, has been linked to liver regeneration. Shiraki et al. (2007) found that the
concentration of Il-18 increased significantly after PH. Other studies have shown that Il-
18 can increase the amount of NF-B and X-linked inhibitors of apoptosis (Yamamoto et
al. 2008).
In order to determine what pathways were used to stimulate Il-18, J. Zhang et al.
(2014) performed PCR. They determined that there were three branches consisting of 13
genes involved with Il-18 signaling. The genes associated with these pathways can be
seen in Table 1. By looking at these results, it is possible to see that the genes associated
with regeneration were upregulated.
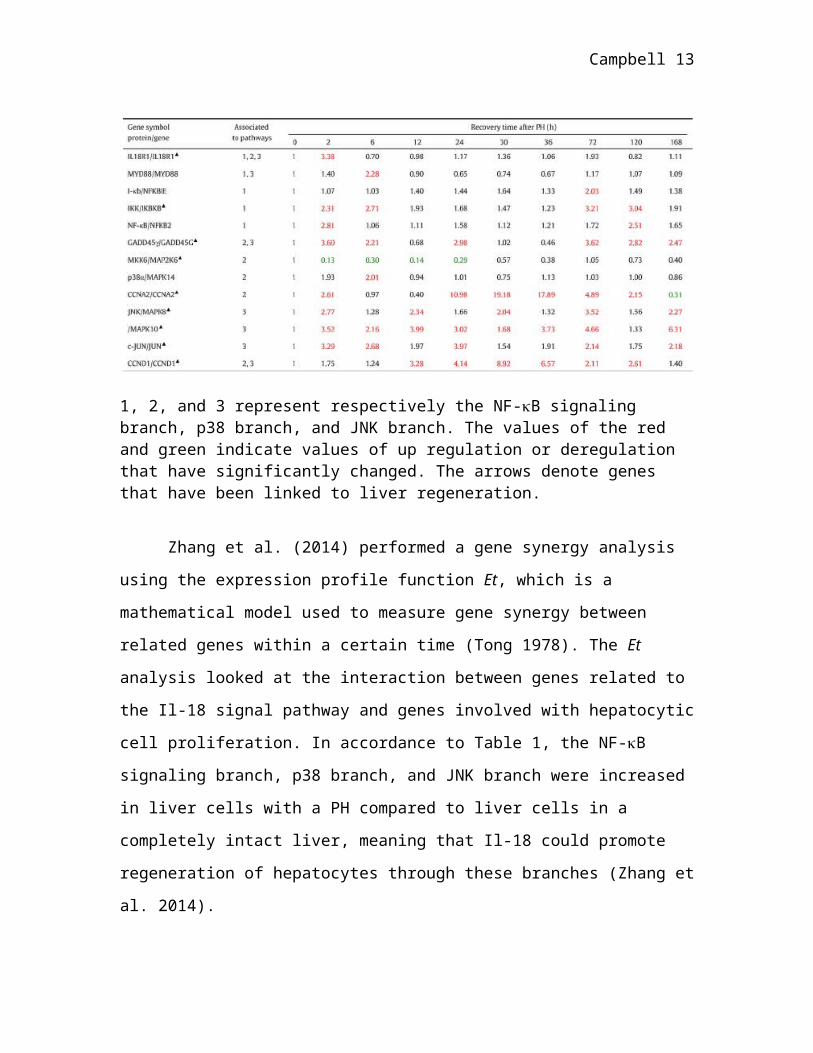
Table 1.Change in genes related to the Il-18 pathway during liver regeneration.
1, 2, and 3 represent respectively the NF-B signaling branch, p38 branch, and JNK branch. The values of the red and green indicate values of up regulation or deregulation that have significantly changed. The arrows denote genes that have been linked to liver regeneration.
Zhang et al. (2014) performed a gene synergy analysis using the expression
profile function Et, which is a mathematical model used to measure gene synergy
between related genes within a certain time (Tong 1978). The Et analysis looked at the
interaction between genes related to the Il-18 signal pathway and genes involved with
hepatocytic cell proliferation. In accordance to Table 1, the NF-B signaling branch, p38

Campbell 9
branch, and JNK branch were increased in liver cells with a PH compared to liver cells in
a completely intact liver, meaning that Il-18 could promote regeneration of hepatocytes
through these branches (Zhang et al. 2014).
Additionally, Zhang et al. treated his rats with injections of rrIl-18, an activated
form of recombinant Il-18. There were no significant differences in the mitotic indexes of
rats treated with low dosages of 0.5l or 1l rrIl-18. But, once the amount of rrIl-18 was
increased to 4l, there was a significant increase in mitotic activity (Figure A, B),
suggesting that high concentrations of Il-18 can increase the rate of hepatocyte
regeneration after PH.
In this study, Zhang and his colleagues determined that the Il-18 pathways
associated with increased LR can be viewed at a genetic level through the upregulation
and downregulation of associated genes. By understanding the genetic component driving
the Il-18 pathway in rabbits, scientists can seek similarities in the human genome.
Regulation of Il-18 production and signaling pathways has potential for liver failure and
regeneration treatments.

Campbell 10
In conclusion, the papers by Hutchins et al. (2014), Sun et
al. (2014), and Zhang et al. (2014) has offered new insight into
the genetic components of different types of tissue regeneration. The paper by Hutchins
et al. identified genes, regulatory factors, and signal pathways that were linked to lizard
tail regeneration. Sun et al. investigated the effect of increased quantities of Sox-9 on
decreasing the progress of IVDD. Finally, Zhang et al. determined that controlling the Il-
18 pathway and production has a direct correlation to liver regeneration. Unfortunately,
all these studies have limited application because the scientists are using animal models,
not human subjects. But, identifying the genes, transcription factors, and signal pathways
(in addition to any other genetics regulators) responsible for specific cell proliferation can
increase understanding of how degeneration occurs and provide insight into future
therapies.
Literature Cited
Anderson DM, Beres BJ, Wilson-Rawls J, Rawls A (2009) The homeobox geneMohawk represses transcription by recruiting the sin3A/HDAC co-repressorcomplex. Dev Dyn 238: 572–580. doi:10.1002/dvdy.21873.
Anderson DM, Beres BJ, Wilson-Rawls J, Rawls A (2009) The homeobox geneMohawk represses transcription by recruiting the sin3A/HDAC co-repressorcomplex. Dev Dyn 238: 572–580. doi:10.1002/dvdy.21873.
Asakura A, Komaki M, Rudnicki M (2001) Muscle satellite cells are
Figure 5. Effects of IL-18 treatment on rat hepatocytic regeneration.(A) Rats were prepared for treatment with rrIL-18 and sacrificed after 24 hours of treatment. Sections of the liver were taken and stained. (B) IL-18 levels in plasma of rat livers were taken and tested at 1 hour and 24 hours.

Campbell 11
multipotential stem cells that exhibit myogenic, osteogenic, and adipogenicdifferentiation. Differentiation 68: 245–253.
Cruciat C-M, Niehrs C (2013) Secreted and transmembrane wnt inhibitors andactivators. Cold Spring Harb Perspect Biol 5: a015081. doi:10.1101/cshperspect.a015081.
Fan C-M, Li L, Rozo ME, Lepper C (2012) Making skeletal muscle fromprogenitor and stem cells: development versus regeneration. Wiley Interdisc RevDev Biol 1: 315–327. doi:10.1002/wdev.30.\
Gruber HE, Norton HJ, Ingram JA, Hanley EN Jr. (2005) The SOX9transcription factor in the human disc: decreased immunolocalization with age
and disc degeneration. Spine (Phila Pa 1976) 30: 625–63
Hiyama A, Mochida J, Iwashina T, Omi H, Watanabe T, et al. (2008)Transplantation of mesenchymal stem cells in a canine disc degeneration model.J Orthop Res 26: 589–600.
Hutchins ED, Markov GJ, Eckalbar WL, George RM, King JM, et al. (2014) Transcriptomic Analysis of Tail Regeneration in the Lizard Anolis carolinensis Reveals Activation of Conserved Vertebrate Developmental and Repair Mechanisms. PLoS ONE 9(8): e105004. doi:10.1371/journal.pone.0105004. <http://web.a.ebscohost.com.pallas2.tcl.sc.edu/ehost/detail/detail?vid=5&sid=8f8f3635-a43e-4ff2-ad06-cf3e03952392%40sessionmgr4005&hid=4201&bdata=JnNpdGU9ZWhvc3QtbGl2ZQ%3d%3d#db=a2h&AN=97802213>.
"Limb Loss Statistics." Amputee Coalition. N.p., n.d. Web. 31 Oct. 2014. <http://www.amputee-coalition.org/limb-loss-resource-center/limb-loss-statistics/>
Luoma K, Riihimaki H, Luukkonen R, Raininko R, Viikari-Juntura E, et al.(2000) Low back pain in relation to lumbar disc degeneration. Spine (Phila Pa1976) 25: 487–492.
Ma, X.M., Dong, X.S., Zhao, H.L., 2009. Research progress of liver regeneration-relatedmolecules. Int. J. Surg. 36, 760–763.
Manceau M, Gros J, Savage K, Thome´ V, McPherron A, et al. (2008) Myostatinpromotes the terminal differentiation of embryonic muscle progenitors. GenesDev 22: 668. doi:10.1101/gad.454408.
Okuma M, Mochida J, Nishimura K, Sakabe K, Seiki K (2000) Reinsertion ofstimulated nucleus pulposus cells retards intervertebral disc degeneration: an invitro and in vivo experimental study. J Orthop Res 18: 988–997.

Campbell 12
Paul R, Haydon RC, Cheng H, Ishikawa A, Nenadovich N, et al. (2003)Potential use of Sox9 gene therapy for intervertebral degenerative disc disease.Spine (Phila Pa 1976) 28: 755–763.
Richardson SM, Walker RV, Parker S, Rhodes NP, Hunt JA, et al. (2006)Intervertebral disc cell-mediated mesenchymal stem cell differentiation. StemCells 24: 707–716.
Sakai D, Mochida J, Iwashina T, Hiyama A, Omi H, et al. (2006) Regenerativeeffects of transplanting mesenchymal stem cells embedded in atelocollagen to thedegenerated intervertebral disc. Biomaterials 27: 335–345.
Sakai D, Mochida J, Yamamoto Y, Nomura T, Okuma M, et al. (2003)Transplantation of mesenchymal stem cells embedded in Atelocollagen gel to theintervertebral disc: a potential therapeutic model for disc degeneration.Biomaterials 24: 3531–3541.
Sell, S., 2003. The hepatocyte: heterogeneity and plasticity of liver cells. Int. J. Biochem.Cell Biol. 35 (3), 267–271.
Shiraki, K., Yamaguchi, Y., Uemoto, S., 2007. Seruminterleukin-18 during liverregeneration.Hepatogastroenterology 54 (76), 1.
Sive JI, Baird P, Jeziorsk M, Watkins A, Hoyland JA, et al. (2002) Expression ofchondrocyte markers by cells of normal and degenerate intervertebral discs. MolPathol 55: 91–97.
Steck E, Bertram H, Abel R, Chen B, Winter A, et al. (2005) Induction ofintervertebral disc-like cells from adult mesenchymal stem cells. Stem Cells 23:403–411.
Sun W, Zhang K, Liu G, Ding W, Zhao C, et al. (2014) Sox9 Gene Transfer Enhanced Regenerative Effect of Bone Marrow Mesenchymal Stem Cells on theDegenerated Intervertebral Disc in a Rabbit Model. PLoS ONE 9(4): e93570. doi:10.1371/journal.pone.0093570.<http://web.b.ebscohost.com.pallas2.tcl.sc.edu/ehost/pdfviewer/pdfviewer?sid=49a108a9-f7dc-4b54-a7ef-85b61fcc2014%40sessionmgr111&vid=12&hid=110>.
Takahashi Y, Takagi A, Hiraoka S, Koseki H, Kanno J, et al. (2007)Transcription factors Mesp2 and Paraxis have critical roles in axialmusculoskeletal formation. Dev Dyn 236: 1484–1494. doi:10.1002/dvdy.21178.
Taub, R., 2004. Liver regeneration: from myth to mechanism. Nat. Rev. Mol. Cell Biol. 5(10), 836–847.
Campbell 13
"Tissue Engineering and Regenerative Medicine." National Institute of Biomedical Imaging and Bioengineering. N.p., n.d. Web. 31 Oct. 2014. <http://www.nibib.nih.gov/science-education/science-topics/tissue-engineering-and-regenerative-medicine>.
Tong, H., 1978. On a threshold model, pattern recognition and signal processing. In:Chen,C.H. (Ed.), NATO ASI series E: Applied Sc No. 29. Sijthoff & Noordhoff,Alphen aan den Rijn, The Netherland.
Tsuchiya H, Kitoh H, Sugiura F, Ishiguro N (2003) Chondrogenesis enhancedby overexpression of sox9 gene in mouse bone marrow-derived mesenchymalstem cells. Biochem Biophys Res Commun 301: 338–343.
Yamamoto, N., et al., 2008. Protective role of interleukin-18 against Fas-mediated liverinjury.Int. J. Mol. Med. 22 (1), 43–48.
Zhang Jihong, Chengkai Ma, Yunqing Liu, Gang Yang, Yun Jiang, Cunshuan Xu, et al. (2014) Interleukin 18 Accelerates the hepatic cell proliferation in rat liver regeneration after partial hepatectomy. Gene 537 : 230-237. DOI: 10.1016/j.gene.2013.12.062
Zhao CQ, Wang LM, Jiang LS, Dai LY (2007) The cell biology of intervertebraldisc aging and degeneration. Ageing Res Rev 6: 247–261.












![Six Quartets for Trumpet eBook - Paul Ayick Vintage Brass 2/Trumpet... · 2 37 af dkk k k ekek k k k n [4] j j j j dj k k 42 af j k k k k kz k t dk k k j kkk k j j jz k k dkk k k](https://static.fdocuments.in/doc/165x107/60a22fff1284953eff4aa254/six-quartets-for-trumpet-ebook-paul-ayick-vintage-2trumpet-2-37-af-dkk-k.jpg)