







([WUDFHOOXODUQXFOHDVHVRI Streptococcus equi VVS ... · $%675$&7 7KHSDWKRJHQ Streptococcus equi VVS...
45
1 Extracellular nucleases of Streptococcus equi ssp. zooepidemicus degrade host neutrophil 1 extracellular traps and impair macrophage activity 2 3 4 Fang Ma, a Xiao Guo, a Hongjie Fan a,b, # 5 6 College of Veterinary Medicine, Nanjing Agricultural University, Nanjing, China a ; 7 Jiangsu Co-innovation Center for Prevention and Control of Important Animal Infectious 8 Diseases and Zoonoses, Yangzhou, China b 9 10 Running Head: Extracellular nucleases of SEZ 11 12 #Address correspondence to Hongjie Fan, [email protected]. 13 14 AEM Accepted Manuscript Posted Online 4 November 2016 Appl. Environ. Microbiol. doi:10.1128/AEM.02468-16 Copyright © 2016, American Society for Microbiology. All Rights Reserved. on March 20, 2020 by guest http://aem.asm.org/ Downloaded from
Transcript of ([WUDFHOOXODUQXFOHDVHVRI Streptococcus equi VVS ... · $%675$&7 7KHSDWKRJHQ Streptococcus equi VVS...

1
Extracellular nucleases of Streptococcus equi ssp. zooepidemicus degrade host neutrophil 1
extracellular traps and impair macrophage activity 2
3
4
Fang Ma,a Xiao Guo,a Hongjie Fana,b, # 5
6
College of Veterinary Medicine, Nanjing Agricultural University, Nanjing, China a; 7
Jiangsu Co-innovation Center for Prevention and Control of Important Animal Infectious 8
Diseases and Zoonoses, Yangzhou, China b 9
10
Running Head: Extracellular nucleases of SEZ 11
12
#Address correspondence to Hongjie Fan, [email protected]. 13
14
AEM Accepted Manuscript Posted Online 4 November 2016Appl. Environ. Microbiol. doi:10.1128/AEM.02468-16Copyright © 2016, American Society for Microbiology. All Rights Reserved.
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

2
ABSTRACT 15
The pathogen Streptococcus equi ssp. zooepidemicus (SEZ) is associated with a 16
wide range of animals, including humans, and outbreaks frequently occur in pigs, equines 17
and goats. Thus far, few studies have assessed interactions between the host immune 18
system and SEZ and how these interactions explain the wide host spectrum of SEZ. 19
Neutrophils, the first line of innate immunity, possess a defense mechanism called 20
neutrophil extracellular traps (NETs), which primarily consist of DNA and granule 21
proteins that trap bacteria via charge interactions. Extracellular nucleases play important 22
roles in the degradation of DNA backbone of NETs. Here, two related extracellular 23
nucleases, nuclease and 5’-nucleotidase (named ENuc and 5Nuc in this study), were 24
identified as being encoded by SESEC_RS04165 gene and SESEC_RS05720 gene (named 25
ENuc and 5Nuc, respectively), and three related gene deletion mutants, specifically, 26
ΔENuc, Δ5Nuc and double mutant ΔENucΔ5Nuc, were constructed. The ΔENuc, Δ5Nuc 27
and ΔENucΔ5Nuc strains demonstrated lower virulence than wild-type SEZ when the 28
mouse survival rate was evaluated post-infection. Furthermore, wild-type SEZ more 29
frequently traversed the bloodstream and transferred to other organs. Wild-type SEZ 30
induced fewer NETs and was able to survive in NETs, whereas only 40% of 31
ΔENucΔ5Nuc cells survived. SEZ degraded NETs DNA backbone and produced 32
deoxyadenosine, primarily through the action of ENuc and/or 5Nuc. However, double 33
mutant ΔENucΔ5Nuc lost the ability to degrade NETs into deoxyadenosine. 34
Deoxyadenosine decreased RAW264.7 cell phagocytosis to 40% of that of normal 35
macrophages. 36
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

3
IMPORTANCE 37
Streptococcus equi ssp. zooepidemicus (SEZ) cause serious bacteremia in its hosts. 38
However, little is known about how SEZ interacts with the host innate immune system, 39
particularly innate cells found in the blood. SEZ is capable of evading NETs-mediated 40
killing via the actions of its potent extracellular nucleases, ENuc and 5Nuc, which 41
directly degrade NETs DNA backbone to deoxyadenosine. In previous studies, other 42
pathogens have required the synergism of nuclease and 5’-nucleotidase to engage in this 43
self-protective process; however, ENuc and 5Nuc both possess nuclease activity and 44
5’-nucleotidase activity, highlighting the novelty of this discovery. Furthermore, 45
deoxyadenosine impairs phagocytosis but not the intracellular bactericidal activity of 46
macrophages. For the first time, we describe a novel mechanism for SEZ extracellular 47
nucleases in NETs degradation, which may provide new insights into pathogen immune 48
evasion mechanism and the prevention and treatment of bacterial disease. 49
50
Keyword: zoonoses, bacteremia, host-pathogen interactions, immune evasion, 51
pathogenic mechanism 52
INTRODUCTION 53
Streptococcus equi ssp. zooepidemicus (SEZ) is a zoonotic pathogen belonging to 54
Lancefield group C, which includes Streptococcus equi ssp. equi and Streptococcus equi 55
ssp. ruminatorum. Streptococcus equi ssp. equi is an important horse pathogen causing 56
strangles, a serious and highly contagious disease of the upper respiratory tract (1). SEZ 57
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

4
is thought to be the ancestor of Streptococcus equi ssp. equi and infects a wild range of 58
animal species, including humans, who contract the disease through contact with infected 59
animals and via the consumption of contaminated dairy products (2, 3). Recent SEZ 60
outbreaks in Sweden (2009/10) and Brazil (1997/98) led to significant suffering and 61
economic loss (4). The widespread nature of SEZ may be related to its strong ability to 62
evade the immune systems of its hosts. 63
Neutrophils are regarded as the first line of immune defense against bacterial 64
infection (5) and kill microbes by releasing antimicrobial proteins into extracellular space 65
via degranulation or by phagocytosing invading microbes (6, 7). However, neutrophils 66
also kill pathogens by unleashing neutrophil extracellular traps (NETs) (8). NETs respond 67
to bacteria, fungi, viruses, other chemical compounds, and even host factors such as 68
activated platelets and IL-8. Upon neutrophil activation, the nuclear envelope 69
disintegrates, and DNA fibers burst into extracellular space (9). 70
To survive in the host, many bacteria produce extracellular DNases to degrade NETs 71
DNA backbone such as Sda1 of Streptococcus pyogenes and EndA of Streptococcus 72
pneumoniae (10, 11). These enzymes allow bacteria to evade the host immune system. In 73
recent years, the involvement of many extracellular nucleotidases in the interaction 74
between NETs and bacteria has been identified. For example, adenosine synthase A 75
(AdsA) of Staphylococcus aureus acts as a nucleotidase to convert the NETs degradation 76
products into deoxyadenosine, which activates caspase-3-mediated apoptosis in 77
macrophages and monocytes (12). However, few studies have assessed the interaction 78
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

5
between NETs and SEZ, and the mechanism underlying SEZ-mediated bacteremia 79
remains unclear. 80
Therefore, in this study, we aimed to assess the interaction between NETs and SEZ 81
to determine whether the actions of extracellular nucleases contribute to NETs evasion 82
and the spread of SEZ in the host. Here, we found two genes, SESEC_RS04165 gene and 83
SESEC_RS05720 gene (named ENuc and 5Nuc in this study), encoding putative 84
extracellular nucleases. ENuc are predicted to encode nuclease (ENuc) and 5Nuc are 85
predicted to encode 5’-nucleotidase (5Nuc). The properties of the two related 86
extracellular nuclease genes are characterized and their role in immune evasion are 87
confirmed. 88
MATERIALS AND METHODS 89
Bacterial strains, cells and experimental animals. The bacterial strains and 90
plasmids used in this study are listed in Table 1. The wild-type SEZ strain ATCC35246 91
(hereafter referred to as WT SEZ) was purchased from the American Type Culture 92
Collection (ATCC) and grown in Todd-Hewitt broth (THB) medium (Difco, BD, 93
Franklin, NJ, USA) at 37°C on a gentle rocking shaker. Escherichia coli strains were 94
grown in Luria-Bertani (LB) broth at 37°C. When necessary, the antibiotic were used at 95
the following concentrations to screen transformants: 100 µg/ml spectinomycin (Sigma 96
Aldrich, St. Louis, Mo, USA) for SEZ, 50µg/ml spectinomycin for E. coli, 50 µg/ml 97
ampicillin (Sigma Aldrich) for E. coli, 50 µg/ml kanamycin (Sigma Aldrich) for E. coli. 98
RAW264.7 cells (ATCC® TIB-71TM) were purchased from ATCC. The cells were 99
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

6
cultured in Dulbecco’s modified Eagle’s medium (DMEM; Wisent, Canada) 100
supplemented with 10% fetal bovine serum at 37°C in 5% CO2. 101
Four-week-old female ICR (Institute of Cancer Research) specific-pathogen-free 102
mice were purchased from the Comparative Medicine Center of Yangzhou University. 103
All experimental protocols were approved by the Laboratory Animal Monitoring 104
Committee of Jiangsu Province. Animals showing signs of illnesses were humanely 105
euthanized with 100% CO2. 106
Quantitative reverse transcription polymerase chain reaction (qRT-PCR) 107
analysis. Total RNA was extracted from WT SEZ in the logarithmic phase using TRIzol 108
reagent (Gibco-BRL, Thermo Fisher Scientific, altham, MA, USA). Briefly, the bacteria 109
were washed 3 times with phosphate-buffered saline (PBS). One milliliter of TRIzol 110
solution was added to the bacterial precipitate, which was shaken for 15 s to lyse the 111
bacterial cells; after that, 200 μl of chloroform was added. The mixture stood at room 112
temperature for 2 min and was then centrifuged at 12000 × g for 15 min. Two hundred 113
microliters of the upper phase (avoiding the middle layer containing DNA) was collected, 114
and 500 μl of isopropyl alcohol was added to precipitate the RNA. The suspension was 115
centrifuged to collect the RNA and washed twice with 75% ethanol. Seven genes 116
predicted to be extracellular nucleases were chosen based on genomic information for 117
Streptococcus equi ssp. zooepidemicus ATCC 35246 (NC_017582.1) in the NCBI 118
database. qRT-PCR was performed to determine the transcript concentrations of these 7 119
predicted extracellular nucleases genes: ENuc, 5Nuc, SESEC_RS02695, SESEC_RS03220, 120
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

7
SESEC_RS04065, SESEC_RS04270 and SESEC_RS05625 (Table 2). All primers used are 121
listed in Table 3. The results were analyzed using the 2−ΔΔCt method. 122
Cloning and purification of recombinant ENuc and 5Nuc. Cloning and 123
purification of the recombinant protein ENuc (rENuc) were performed as previously 124
described (13). ENuc was cloned by PCR using the primers ENucF and ENucR (Table 3) 125
and the WT SEZ genome as a template. The cloned ENuc was digested with BamHI and 126
XhoI and cloned into the pET-32a (+) vector (Invitrogen, Thermo Fisher Scientific, 127
altham, MA, USA). After sequencing (Shanghai Sunny Biotechnology Co., Ltd., China), 128
the recombinant plasmid pET32a-ENuc was transformed into E. coli Rosetta cells with 129
ampicillin selection. The recombinant E. coli was incubated at 37°C until reaching the 130
exponential growth phase, and then protein expression was induced for 6 h with the 131
addition of 0.1 mM isopropyl-β-D-thiogalactopyranoside (Sigma Aldrich). Cells were 132
harvested, washed and suspended in ice-cold binding buffer (20mM Tris, 200mM NaCl, 133
pH7.4). 134
Protein purification procedures were performed at 4°C. Bacteria were homogenized 135
by ultrasonication for 30 min. The resulting crude extract was centrifuged at 10000 × g 136
for 30 min. Recombinant rENuc were purified using a 1 ml Ni-Sepharose™ 6 Fast-Flow 137
column (GE Healthcare, Uppsala, Sweden). 5Nuc was cloned into pET-28a (+) vector 138
(Invitrogen) to improve its expression; purification of 5Nuc recombinant protein (r5Nuc) 139
were performed as mentioned above. 140
Construction of gene deletion mutants. The construction of ENuc and 5Nuc single 141
gene deletion mutants (hereafter called ΔENuc and Δ5Nuc) and double gene deletion 142
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

8
mutant (ΔENucΔ5Nuc) was performed as previously described (14). Briefly, the upstream 143
and downstream sequences surrounding ENuc were amplified from WT SEZ 144
chromosomal DNA using the primer combinations ENuc-L-F/ENuc-L-R and 145
ENuc-R-F/ENuc-R-R (Table 3). The upstream and downstream flanking sequences of 146
ENuc were ligated between the SalI and BamHI sites of the thermosensitive suicide 147
vector pSET4s to yield the recombinant plasmid pSET4s-ΔENuc, which was 148
electroporated into WT SEZ (15). The plasmid pSET4s-ΔENuc and the bacterial genome 149
exchanged genetic fragments twice via intermolecular recombination. Putative deletion 150
mutants were screened by PCR performed with the primer combination ENucF/ENucR 151
(Table 3). The mutant Δ5Nuc was constructed on the WT SEZ background using the 152
process mentioned above, and the double mutant ΔENucΔ5Nuc was constructed using the 153
same process on the ΔENuc background. 154
Isolation and purification of neutrophils. Murine neutrophils were isolated from 155
the fresh, heparinized venous blood of healthy ICR mice as previously described, with 156
some modifications (16). Briefly, whole blood was centrifuged at 400 × g for 30 min, and 157
then the top layer of plasma was discarded. Red blood cells were treated with lysis buffer 158
(Tris-NH4Cl). White blood cells were suspended in 0.85% NaCl. Three milliliters of 65% 159
Percoll were gently overlaid in a 15-ml tube (BD Falcon), followed by a 3-ml layer of 55% 160
Percoll; finally, the cell suspension was carefully overlaid as the top layer. The Percoll 161
gradient was centrifuged at 1000 × g for 30 min, and the cells at the plasma interface 162
were carefully removed and discarded. The cells between the 55% and 65% Percoll layers 163
were collected, washed in RPMI 1640 medium (Gibco) and resuspended in RPMI 1640 164
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

9
medium. Neutrophil purity was confirmed by performing flow cytometry, and cell 165
viability was detected by performing Trypan blue staining and counting cells in a 166
chamber slide (Invitrogen). 167
Isolation and purification of monocytes from blood. Blood from the ICR mice 168
was diluted in PBS and overlaid on a Histopaque® 1077 solution (Sigma Aldrich) via 169
gradient centrifugation at 400 × g for 20 min (17). The upper layer was aspirated, and 170
monocytes were enriched in the interphase. Then, the cells were washed with PBS, 171
suspended in RPMI 1640, and seeded into 24-well plates for further study. 172
Mouse survival assay. A mouse survival assay was carried out to assess the 173
virulence of WT SEZ, ΔENuc, Δ5Nuc and double mutant ΔENucΔ5Nuc. Fifty ICR mice 174
were divided into 5 groups, with 10 mice in each group. The positive group was 175
challenged with WT SEZ at a dose of 105 colony forming units (CFU) via intraperitoneal 176
injection. The experimental groups were challenged with 105 CFU of ΔENuc, Δ5Nuc and 177
ΔENucΔ5Nuc via intraperitoneal injection. The negative control group was challenged 178
with PBS. The symptoms of the mice were documented for 7 days post-infection. 179
Bacterial dissemination assay. Bacterial dissemination was evaluated as described 180
previously (18). Mice were inoculated with 104 CFU of WT SEZ, ΔENuc, Δ5Nuc and 181
double mutant ΔENucΔ5Nuc, with 5 mice in per group, via intravenous injection into the 182
caudal vein to ensure the direct interaction of bacteria and neutrophils; this dose of 183
bacteria causes asymptomatic infection. Bacterial burdens were enumerated in the blood, 184
liver, lung and spleen at 24 h, 48 h, 72 h and 96 h to evaluate bacterial proliferation. At 185
each time point, mice were euthanized. Livers, spleens, lungs, and blood were harvested, 186
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

10
weighed and homogenized in 1 ml of PBS. After that, the mixture was serially diluted 187
and plated on THB agar plates. 188
NETs bactericidal assays. Bactericidal assays were carried out to compare the 189
survival abilities of WT SEZ, ΔENuc, Δ5Nuc and double mutant ΔENucΔ5Nuc in the 190
presence of NETs, as described elsewhere (19). For the NETs bactericidal assays, 191
neutrophils were activated with Phorbol-myristate-acetate (PMA; Sigma-Aldrich) for 3 h, 192
after which cytochalasin B (Sigma Aldrich) was added to a final concentration of 10 193
μg/ml to suppress cell phagocytosis. WT SEZ bacteria and the three mutant strains were 194
grown to the logarithmic phase in THB, washed 3 times with sterile PBS, and added to 195
neutrophils at an MOI of 100 at 37°C under a 5% CO2 atmosphere. After incubation for 196
90 min, The remaining neutrophils were lysed with 0.02% Triton X-100 (Sigma Aldrich), 197
and the contents were serially diluted and plated on THB agar plates to determine the 198
number of viable bacteria. For the blank control, the same number of CFU for each 199
bacterial strain were plated without incubation. The percent bactericidal activity was 200
calculated as ([CFU/ml in blank control] – [CFU/ml in experimental group])/ [CFU/ml in 201
blank control] × 100%. 202
Nuclease activity assay. To detect the function of ENuc and 5Nuc, nuclease assays 203
were performed as described previously (20, 21). Briefly, 10 μl of calf thymus DNA 204
(Sigma Aldrich) at a dose of 100 ng/μl was mixed with reaction buffer (300 mM Tris; 3 205
mM MgCl2; 3 mM CaCl2, pH 7.0) in a total volume of 200 μl. The reaction was initiated 206
by adding 0.1 µM rENuc or r5Nuc protein and DNase I (Sigma Aldrich). DNase I and 207
reaction buffer were used as a positive control and negative control, respectively. After 208
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

11
incubation at 37°C for 1 h, the reaction was halted with the addition of EDTA at a final 209
concentration of 50 mM. The remaining DNA was visualized using GoldView Nucleic 210
Acid Stain (Vazyme Biotech Co., Ltd, Nanjing, China) under UV light. To further study 211
the function of ENuc and 5Nuc in SEZ, WT SEZ and the three mutant strains were 212
collected in the logarithmic phase of growth and washed 3 times with PBS. Thereafter, 10 213
μl of bacterial suspension at a dose of 106 CFU/ml was incubated with 1 μg of calf 214
thymus DNA in reaction buffer in a total volume of 50 μl at 37°C for 3 h. The remaining 215
DNA was detected as described above. DNase I and reaction buffer were used as a 216
positive control and negative control, respectively. 217
Nucleotidase activity and reverse phase high-performance liquid 218
chromatography (rpHPLC). To digest nucleotides and DNA, reaction buffer (300 mM 219
Tris; 3 mM MgCl2; 3 mM CaCl2, pH 7.0) was mixed with 1 mM nucleotides or 1 µg of 220
calf thymus DNA (Sigma Aldrich) and 0.1 µM rENuc, r5Nuc protein or bacteria at dose 221
of 106 CFU/ml in a total volume of 500 µl, followed by incubation at 37°C for 1 h. The 222
enzymatic reaction was stopped by adding EDTA at a final concentration of 50 mM. To 223
assess NETs digestion, 0.5 µM rENuc, r5Nuc or bacteria at a dose of 107 CFU/ml was 224
incubated with NETs for 3 h. The solution was centrifuged at 800 × g for 10 min, and the 225
supernatant was collected. 226
The production of deoxyadenosine and adenosine was determined by rpHPLC. 227
Samples underwent chromatography on a 250-mm × 4.6-mm column (BDS Hypersil C18, 228
5-µm particle size; Thermo Fisher Scientific). The mobile phase was a solution of 70% 229
65 mm K3PO4 mixed with 30% methanol, pH 6.8. The reaction samples were eluted with 230
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

12
the mobile-phase solution for 20 min for deoxyadenosine or 15 min for adenosine at a 231
flow rate of 0.8 ml/min. Peaks were detected at a UV absorbance of 260 nm. Commercial 232
nucleosides (Sigma-Aldrich) were used as a standard sample. 233
Phagocytosis assay. A phagocytosis assay was performed according to a previously 234
described method with modifications (22). RAW264.7 cells in 24-well plates were 235
pretreated with 50 μM deoxyadenosine for 1 h. WT SEZ was cultured to the 236
mid-exponential phase and washed 3 times with PBS. The RAW264.7 cells were then 237
washed 3 times with DMEM without fetal bovine serum. Semi-confluent monolayers 238
were infected with 100-μl suspensions containing 106 CFU/ml WT SEZ. The control 239
group comprised RAW264.7 cells without deoxyadenosine pretreatment. Each 24-well 240
plate was concentrated at 800 × g for 10 min and incubated for 2 h at 37°C in 5% CO2. 241
Cells were washed 3 times with DMEM, followed by treatment with 242
penicillin-streptomycin at a concentration of 100 µg/ml for 1 h to kill extracellular 243
bacteria. The cells were again washed 3 times with DMEM, and 100 μl of trypsin and 900 244
μl of sterile deionized water were added to release all bacteria. CFU numbers were 245
determined by plating serial dilutions. WT SEZ without incubation was plated on THB 246
agar as the initial concentration, and the percent bactericidal activity in the control group 247
was set as 100%. The percent bactericidal activity in the treatment group was calculated 248
as (bactericidal CFU in the treatment group)/ (bactericidal CFU in the control group) × 249
100%. 250
To study the effects of deoxyadenosine on macrophages, we designed the following 251
assay. Briefly, cells were pretreated with or without 50 μM deoxyadenosine for 1 h and 252
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

13
then incubated with WT SEZ for 2 h; following this, extracellular bacteria were killed 253
with 100 µg/ml penicillin-streptomycin for 1 h, 2 h, 3 h and 4 h, and the number of viable 254
bacteria on THB agar was calculated for each time point. The number of surviving 255
bacteria after antibiotic killing for 1 h was considered the total number of viable 256
intracellular bacteria. RAW264.7 cells without treatment were used as controls. The 257
bactericidal percentage was calculated as follows: ([CFU/ml at n h] - [CFU/ml at n+1 h])/ 258
(CFU/ml at 1h) × 100%. 259
Neutrophil and monocyte bactericidal assays. Cells isolated from murine blood 260
were pretreated with or without 50 μM deoxyadenosine; following this, WT SEZ bacteria 261
cultured to the mid-exponential growth phase were washed 3 times with PBS and added 262
to murine blood cells. The mixture was incubated for 90 min and lysed with 0.02% Triton 263
X-100 on ice. The CFU of viable bacteria were determined by plating serial dilutions on 264
THB agar. 265
Statistical analysis. All experiments were repeated at least 3 times. The GraphPad 266
Prism 5 Software package (GraphPad Software, La Jolla, CA, USA) was used to perform 267
statistical analyses. Values of p <0.05 were considered statistically significant. 268
RESULTS 269
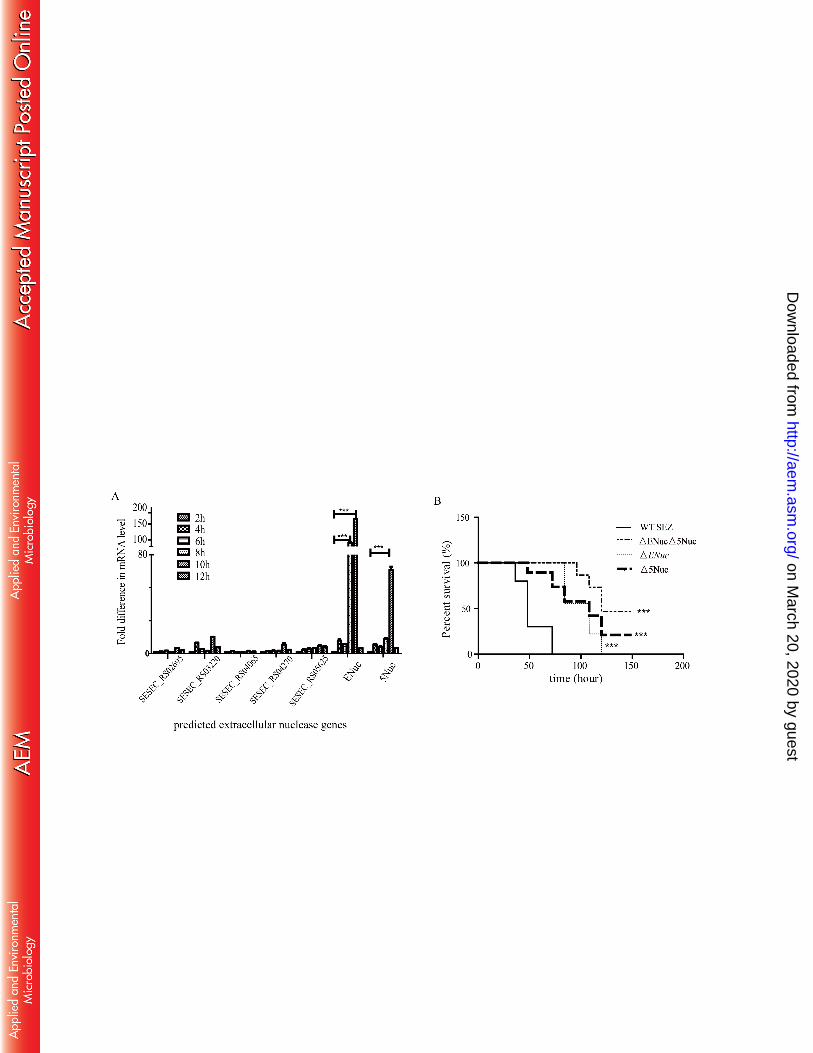
Determination and identification of ENuc and 5Nuc. Seven genes in WT SEZ are 270
predicted to encode related extracellular nucleases according to a literature search in the 271
NCBI database (listed in Table 2); however, it is unknown whether these genes are 272
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

14
transcribed. Therefore, qRT-PCR was performed to study the transcriptional levels of 273
these genes. mRNA expression levels at 2 h were set as 1-fold, and all 7 genes were 274
transcribed during growth, particularly by 10 h (early in the stationary phase). However, 275
the expression of ENuc and 5Nuc was significantly higher by 10 h (p <0.001) than that of 276
other nuclease genes. At 10 h, ENuc and 5Nuc transcription levels were 165 times and 67 277
times higher, respectively, than their transcription levels at 2 h (Fig. 1A). 278
ENuc (2781-bp ORF; NCBI Gene ID, SESEC_RS04165) and 5Nuc (2007-bp ORF; 279
NCBI Gene ID, SESEC_RS05720) are uncharacterized genes encoding a nuclease and a 280
5’-nucleotidase, and their corresponding proteins are designated ENuc and 5Nuc. ENuc 281
contains a Gram-positive cocci surface protein LPXTG motif, a signal peptide, and three 282
predicted extracellular nuclease domains, including two motifs similar to the OB fold of 283
Bacillus subtilis YhcR: a sugar-nonspecific nuclease (YhcR_OBF_like; accession, 284
cd04486; E-value = 2.72e-13 and 2.13e-12) and an exonuclease-endonuclease-phosphatase 285
domain (accession, cl00490; E-value = 4.48e--30). 5Nuc contains an LPXTG motif, a 286
peptide and two 5’-nucleotidase metallophosphatase domains in the amino-terminal 287
region (accession, cl13995; E-value = 1.26e--121) and a 5’-nucleotidase domain (accession, 288
pfam02872; E-value = 3.18e-32) in the carboxyl-terminal region. All data available from 289
the NCBI were analyzed. 290
Virulence comparison. The virulence of the three gene deletion mutants decreased 291
to varying degrees compared with WT SEZ (p<0.001). Half of the mice challenged with 292
double mutant ΔENucΔ5Nuc survived for 7 days post-infection. All mice in the WT SEZ 293
group died between 24 h and 72 h post-infection. In contrast, mice challenged with 294
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

15
ΔENuc and ΔENucΔ5Nuc exhibited no obvious symptoms during the first 3 days 295
post-infection; however, all mice challenged with ΔENuc eventually died between 84 h 296
and 120 h post-infection, suggesting ENuc contributes to bacterial survival during the 297
early stage of infection. Mice challenged with Δ5Nuc began to die at 48 h post-infection, 298
and 20% of mice in the Δ5Nuc group survived, indicating an important role for 5Nuc in 299
bacterial proliferation during infection (Fig. 1B). Clearly, ENuc and 5Nuc are important 300
virulence factors during SEZ infection, and ENuc may play a role in early infection, 301
while 5Nuc appears to be more active than ENuc. All mice challenged with PBS survived, 302
and the data are not shown in Fig. 1B. 303
Roles of ENuc and 5Nuc in NETs entrapment evasion. To study the functions of 304
ENuc and 5Nuc, we cloned these two genes from the genome of WT SEZ and ligated 305
them into the vectors pET-32a (+) and pET-28a (+), respectively. Recombinant proteins 306
were purified from E. coli Rosetta. rENuc and r5Nuc both degraded calf thymus DNA 307
within 1 h at 37°C. Notably, double mutant ΔENucΔ5Nuc lost its ability to degrade calf 308
thymus DNA compared with WT SEZ, ΔENuc and Δ5Nuc (Fig. 2). Thus, the expression 309
of ENuc and 5Nuc allows WT SEZ to acquire the ability to degrade extracellular DNA. 310
Deoxyadenosine derived from the degradation of NETs DNA backbone is toxic to 311
host immune cells. To determine whether ENuc and 5Nuc degrade DNA into 312
deoxyadenosine, deoxyadenosine production was analyzed using rpHPLC. Recombinant 313
rENuc and r5Nuc degraded calf thymus DNA, which mimics the DNA backbone of 314
NETs, and NETs DNA and produced deoxyadenosine (Fig. 3A–G). WT SEZ, ΔENuc and 315
Δ5Nuc also produced deoxyadenosine following NETs interaction, but the double mutant 316
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

16
ΔENucΔ5Nuc did not (Fig. 3H-K). Additionally, WT, ΔENuc and Δ5Nuc degraded calf 317
thymus DNA to produce deoxyadenosine (see Fig. S1 in the supplemental material). Thus, 318
ENuc and 5Nuc, which possess both nuclease activity and 5’-nucleotidase activity, 319
appear to be the primary nucleases that allow SEZ to evade entrapment of NETs. 320
Recombinant rENuc and r5Nuc both exhibited 5’-nucleotidase activity and 321
hydrolyzed adenosine triphosphate (ATP), adenosine diphosphate (ADP) and adenosine 322
monophosphate (AMP) to produce adenosine (Fig. 4). Adenosine is an 323
immunomodulatory molecule that suppresses neutrophil functions such as degranulation. 324
Therefore, the ability of ENuc and 5Nuc to generate adenosine enhances the virulence of 325
SEZ. 326
Survival in different organs. NETs degradation contributes to SEZ-induced 327
bacteremia and spread to the organs from the blood. In particular, WT SEZ showed a 328
greater tendency to break through blood barrier and cause bacteremia. At 24 h 329
post-infection, only mice challenged with WT SEZ demonstrated bacterial loads in the 330
blood, lung, spleen and liver, indicating that the mutant strains were less able to survive 331
and cross the blood barrier. Mice challenged with ΔENuc showed bacterial loads at 48 h 332
and 72 h, but mice challenged with Δ5Nuc only showed bacteria loads at 48 h, suggesting 333
an increased tendency for ΔENuc to spread to the organs from the blood. The nuclease 334
activity of 5Nuc appear to be more potent than ENuc, which is consistent with the 335
virulence of ΔENuc and Δ5Nuc. Importantly, at 72 h post-infection, mice challenged with 336
ΔENuc demonstrated bacterial loads equivalent to those in mice challenged with WT SEZ; 337
however, at 96 h post-infection, WT SEZ was better able to survive in organs such as the 338
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

17
lung, liver and spleen. Thus, not only dose WT SEZ demonstrate a superior ability to 339
transfer to organs but also impedes clearance by the host. Mice challenged with double 340
mutant ΔENucΔ5Nuc did not exhibit bacterial loads from 24 h to 96 h, suggesting 341
bacteria were killed by the host immune system at an early stage in the blood due to the 342
lack of ENuc and 5Nuc (Fig. 5). 343
Bactericidal capacity of NETs. WT SEZ resists killing by NETs. The bactericidal 344
activity of NETs after incubation with WT SEZ was negative, facilitating bacterial 345
replication and increased numbers of WT SEZ. However, approximately 30% of ΔENuc, 346
50% of Δ5Nuc and 60% of ΔENucΔ5Nuc were killed following incubation with NETs for 347
90 min (Fig. 6). In addition, fibrous structure of NETs DNA backbone was only found in 348
gene deletion mutant strains and NETs concentrations induced by ΔENuc and Δ5Nuc 349
were 2 and 5 times higher than that induced by WT SEZ. Importantly, NETs 350
concentrations induced by ΔENucΔ5Nuc were more than 7 times higher than that induced 351
by WT SEZ (see Fig. S2 in the supplemental material). Weaker ability to induce NETs 352
formation by WT SEZ compared with gene deletion mutants may attribute to the DNA 353
degradation activities of ENuc and 5Nuc, which contributes to bacterial survival in NETs. 354
Therefore, ENuc and 5Nuc play important roles in protecting bacteria against killing by 355
NETs, aiding SEZ proliferation and spread to deep tissues. 356
The effects of deoxyadenosine on RAW264.7 phagocytosis. To evaluate the 357
cytotoxic effects of the NETs degradation product deoxyadenosine on immune cells, 358
RAW264.7 cells and neutrophils and monocytes obtained from blood were employed. 359
Phagocytosis and the bactericidal capacity of RAW264.7 pre-treated with 360
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

18
deoxyadenosine decreased by almost 60% (Fig. 7A). Surprisingly, in an intracellular 361
killing assay, the intracellular killing activity of RAW264.7 cells pre-treated with 362
deoxyadenosine was equivalent to that of RAW264.7 cells without pre-treatment (Fig. 363
7B). Thus, deoxyadenosine primarily impairs the phagocytosis of macrophages but does 364
not affect the intracellular killing process. Furthermore, deoxyadenosine did not influence 365
the bactericidal activity of monocytes (Fig. 7C). 366
The effects of deoxyadenosine on neutrophil function. To evaluate the effects of 367
deoxyadenosine on neutrophils, two assays were designed. In the first, we tested the 368
influence of deoxyadenosine on bacterial killing activity, and in the second we tested the 369
effects of deoxyadenosine on NETs formation induced by PMA. Based on the results of 370
both assays, deoxyadenosine did not affect neutrophil function (Fig. 8). Purified 371
neutrophils comprised greater than 90% of total cells, and viability was nearly 90% over 372
4 h in vitro in study (see Fig. S3 in the supplemental material). Thus, our experiments 373
were carried out with active and pure neutrophils, lending credibility to the results. 374
DISCUSSION 375
This study was initially designed to identify the functions of extracellular nucleases 376
of SEZ in the context of immune evasion and provide new ideas for future studies 377
examining bacterial zoonosis. We assessed the interactions between SEZ and NETs and 378
observed the ability of SEZ to evade entrapment and killing by NETs. Importantly, SEZ 379
possesses a potent ability to degrade NETs into deoxyadenosine, negatively influencing 380
macrophage phagocytic activity. The production of extracellular nucleases by bacteria is 381
a significant mechanism utilized by bacteria to escape NETs (23). In this work, we 382
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

19
confirmed the identities of two primary extracellular nucleases by conducting a literature 383
search of the NCBI database and performing qRT-PCR. Interestingly, the recombinant 384
proteins rENuc and r5Nuc demonstrated both nuclease and nucleotidase activities 385
allowing both to degrade NETs and produce deoxyadenosine. To further study the 386
functions of these two enzymes, three gene deletion mutant strains—ΔENuc, Δ5Nuc and 387
double mutant ΔENucΔ5Nuc—were constructed. These mutants exhibited lower 388
virulence and a weaker ability to spread from the blood to organs compared to the WT 389
SEZ strain; this was particularly the case for the two-gene deletion mutant ΔENucΔ5Nuc. 390
Half of the mice challenged with ΔENucΔ5Nuc survived, while all of the mice in the WT 391
SEZ group were dead within three days. Thus, it appears difficult for ΔENucΔ5Nuc to 392
exit the blood-stream. Despite the degradation of NETs by SEZ, deoxyadenosine, a NETs 393
degradation product, may contribute to bacterial survival in the host; according to our 394
results, deoxyadenosine suppresses macrophage phagocytosis. These two characteristics 395
of SEZ strongly contribute to its pathogenicity and proliferation in the host. 396
NETs constitute an important and novel innate immune defense system that 397
influences pathogen invasion. Recently, this phenomenon was noted in several bacteria, 398
including Staphylococcus aureus and Streptococcus pyogenes (24). In this study, when 399
the genes ENuc and 5Nuc were knocked down simultaneously, SEZ was less able to 400
survive in NETs and better able to stimulate neutrophils to produce NETs which 401
indicated that ENuc and 5Nuc influenced SEZ virulence. PMA is generally 402
acknowledged to be an appropriate positive control when stimulating neutrophils to 403
produce NETs and was employed in this study in vitro (25). In previous studies 404
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

20
examining the interactions between pathogens and NETs in the context of 405
deoxyribonuclease, extracellular deoxyribonucleases protected pathogens by allowing 406
them to evade entrapment by NETs; these include EndA produced by Streptococcus 407
pneumoniae and Sda1 produced by Streptococcus pyogenes (26, 27-29). The recombinant 408
rENuc and r5Nuc proteins both degraded calf thymus DNA, a common substitute for the 409
NETs DNA backbone in laboratory research. Thus, ENuc and 5Nuc likely protect SEZ 410
from NETs entrapment. In addition, high bacterial loads were only observed in mice 411
challenged with WT SEZ at 24 h post-infection; this may be attributable to the functions 412
of ENuc and 5Nuc such that WT SEZ not only survived in the host but was able to spread 413
to organs better than the mutant strains. Moreover, Δ5Nuc and ΔENucΔ5Nuc 414
demonstrated difficulty in spreading to other organs from the blood, probably due to the 415
deletion of 5Nuc. Thus, 5Nuc may be more active than ENuc. Importantly, mice 416
challenged with ΔENucΔ5Nuc demonstrated no bacterial loads, highlighting the 417
importance and virulence of ENuc and 5Nuc. In summary, SEZ degrades NETs and 418
protects itself by evading entrapment by the host immune system, which may account for 419
the differences in virulence between SEZ, ΔENuc, Δ5Nuc and double mutant 420
ΔENucΔ5Nuc. 421
Extracellular 5′-nucleotidases, such as AdsA of Staphylococcus aureus, produce 422
adenosine, which is an immunomodulatory molecule (30, 31). However, Thammavongsa 423
V and colleagues noted cooperation between AdsA and a nuclease, facilitating the 424
production of deoxyadenosine through the degradation of NETs DNA. Deoxyadenosine 425
induces macrophage apoptosis via caspase-3 activation, thereby restricting macrophage 426
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

21
survival and promoting the establishment of infection (12). In this study, SEZ degraded 427
NETs DNA backbone and produced deoxyadenosine to suppress the phagocytosis of 428
macrophages but did not negatively influence neutrophils and monocytes. Unexpectedly, 429
both ENuc and 5Nuc demonstrated the ability to convert calf thymus DNA and NETs 430
DNA into deoxyadenosine as directly detected by rpHPLC. This mechanism differs from 431
that observed in other pathogens such as Staphylococcus aureus or Streptococcus 432
pyogenes because these pathogens possess NETs-cleaving nucleases in addition to 433
5′-nucleotidases (30, 32, 33). In this study, SEZ secreted two major extracellular enzymes, 434
ENuc and 5Nuc, that degraded DNA backbone of NETs and produced deoxyadenosine. 435
Even with the deletion of either of these two genes, SEZ was also able to degrade NETs 436
into deoxyadenosine. However, when both genes were knocked out, SEZ virulence 437
decreased and its abilities to degrade NETs and generate deoxyadenosine were lost. 438
Additionally, ENuc and 5Nuc possess the ability to hydrolyze AMP, ADP and ATP to 439
produce the immunomodulatory substrate adenosine, similar to Nt5e produced by 440
Streptococcus sanguinis and AdsA produced by Staphylococcus aureus but unlike S5nA 441
produced by Streptococcus pyogenes and Nud P produced by Streptococcus agalactiae, 442
which demonstrate limited specificity for AMP and ADP (31, 34, 35). Adenosine 443
suppresses neutrophil activation by occupying the A2a receptor on neutrophils and 444
promoting bacterial survival in the host bloodstream (36). Our data reveal a novel and 445
potent mechanism in which extracellular nucleases degrade NETs and produce 446
deoxyadenosine. 447
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

22
Deoxyadenosine is cytotoxic to macrophages. During infection with Staphylococcus 448
aureus, deoxyadenosine production is sufficient to restrict macrophage survival and 449
promote the establishment of persistent infection (12). The production of deoxyadenosine 450
through the degradation DNA backbone of NETs is an important mechanism enabling 451
bacterial survival. However, only a few studies have examined the functions of 452
deoxyadenosine in immune cells. In this study, we investigated the influence of 453
deoxyadenosine on macrophages, neutrophils and monocytes and only observed negative 454
effects on macrophage phagocytosis; however, deoxyadenosine did not affect the 455
bactericidal activity of macrophages. Additionally, deoxyadenosine exerted no influence 456
on the functions of neutrophils and monocytes. The molecular mechanism of 457
deoxyadenosine underlying immune cell impairment will be explored in a future study. 458
In summary, SEZ secretes two potent extracellular enzymes, ENuc and 5Nuc, each 459
demonstrating nuclease and 5′-nucleotidase activities, which allow SEZ to evade 460
entrapment by degrading NETs and producing the cytotoxic substrate deoxyadenosine to 461
impair macrophage phagocytosis. This mechanism differs from a previously reported 462
mechanism in which bacteria utilize nuclease in collaboration with 5′-nucleotidase to 463
degrade NETs DNA into deoxyadenosine. Our novel mechanism represent an ingenious 464
method for bacteria to utilize host components for survival and simultaneously perturb 465
host immune defenses. In addition, the two enzymes hydrolyze ATP, ADP and AMP to 466
produce the immunomodulatory substrate adenosine. Thus, ENuc and 5Nuc are novel and 467
important virulence factors for SEZ, and their identification will aid in the further study 468
of immune evasion by zoonotic pathogens and contribute to the design of therapeutics. 469
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

23
ACKNOWLEDGEMENTS 470
We thank high-performance liquid chromatography unit and flow cytometer unit 471
(Laboratory Center of Life Science, Nanjing Agriculture University). 472
FUNDING INFORMATION 473
This work was supported by grants from the National Transgenic Major Program 474
(2014ZX0800946B), the Special Fund for Agro-scientific Research in the Public Interest 475
(201403054), the National Natural Science Foundation of China (31272581, 31672574) 476
the Jiangsu Agriculture Science and Technology Innovation Fund (CX(15)1056) and the 477
Project Funded by the Priority Academic Program Development of Jiangsu Higher 478
Education Institutions (PAPD). 479
REFERENCES 480
1. Moloney E, Kavanagh KS, Buckley TC, Cooney JC. 2013. Lineages of 481
Streptococcus equi ssp. equi in the Irish equine industry. Ir Vet J 66:1. 482
2. Ma Z, Geng J, Yi L, Xu B, Jia R, Li Y, Meng Q, Fan H, Hu S. 2013. Insight 483
into the specific virulence related genes and toxin-antitoxin virulent pathogenicity 484
islands in swine streptococcosis pathogen Streptococcus equi ssp. zooepidemicus 485
strain ATCC35246. BMC Genomics 14:377. 486
3. Holden MTG, Heather Z, Paillot R, Steward KF, Webb K, Ainslie F, 487
Jourdan T, Bason NC, Holroyd NE, Mungall K, Quail MA, Sanders M, 488
Simmonds M, Willey D, Brooks K, Aanensen DM, Spratt BG, Jolley KA, 489
Maiden MCJ, Kehoe M, Chanter N, Bentley SD, Robinson C, Maskell DJ, 490
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

24
Parkhill J, Waller AS. 2009. Genomic evidence for the evolution of 491
Streptococcus equi: Host restriction, increased virulence, and genetic exchange 492
with human pathogens. PLOS pathogens doi:e1000346. 493
4. Lindahl SB, Aspan A, Baverud V, Paillot R, Pringle J, Rash NL, Soderlund R, 494
Waller AS. 2013. Outbreak of upper respiratory disease in horses caused by 495
Streptococcus equi subsp. zooepidemicus ST-24. Vet Microbiol 166:281-285. 496
5. Wartha F, Beiter K, Normark S, Henriques-Normark B. 2007. Neutrophil 497
extracellular traps: casting the NET over pathogenesis. Curr Opin Microbiol 498
10:52-56. 499
6. Borregaard N. 2010. Neutrophils, from marrow to microbes. Immunity 500
33:657-670. 501
7. Fuchs TA, Abed U, Goosmann C, Hurwitz R, Schulze I, Wahn V, Weinrauch 502
Y, Brinkmann V, Zychlinsky A. 2007. Novel cell death program leads to 503
neutrophil extracellular traps. J Cell Biol 176:231-241. 504
8. Brinkmann V, Zychlinsky A. 2012. Neutrophil extracellular traps: is immunity 505
the second function of chromatin? J Cell Biol 198:773-783. 506
9. Yost CC, Cody MJ, Harris ES, Thornton NL, McInturff AM, Martinez ML, 507
Chandler NB, Rodesch CK, Albertine KH, Petti CA, Weyrich AS, 508
Zimmerman GA. 2009. Impaired neutrophil extracellular trap (NET) formation: 509
a novel innate immune deficiency of human neonates. Blood 113:6419-6427. 510
10. Buchanan JT, Simpson AJ, Aziz RK, Liu GY, Kristian SA, Kotb M, 511
Feramisco J, Nizet V. 2006. DNase expression allows the pathogen group A 512
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

25
Streptococcus to escape killing in neutrophil extracellular traps. Curr Biol 513
16:396-400. 514
11. Puyet A, Greenberg B, Lacks SA. 1990. Genetic and structural characterization 515
of endA: A membrane-bound nuclease required for transformation of 516
Streptococcus pneumoniae. J Mol Biol 213:727-738. 517
12. Thammavongsa V, Missiakas DM, Schneewind O. 2013. Staphylococcus 518
aureus degrades neutrophil extracellular traps to promote immune cell death. 519
Science 342:863-866. 520
13. White MJ, Boyd JM, Horswill AR, Nauseef WM. 2014. 521
Phosphatidylinositol-Specific Phospholipase C Contributes to Survival of 522
Staphylococcus aureus USA300 in Human Blood and Neutrophils. Infect Immun 523
82:1559-1571. 524
14. Miyoshi-Akiyama T, Takamatsu D, Koyanagi M, Zhao J, Imanishi K, 525
Uchiyama T. 2005. Cytocidal effect of Streptococcus pyogenes on mouse 526
neutrophils in vivo and the critical role of streptolysin S. J Infect Dis 527
192:107-116. 528
15. Takamatsu D, Osaki M, Sekizaki T. 2001. Thermosensitive suicide vectors for 529
gene replacement in Streptococcus suis. Plasmid 46:140-148. 530
16. Brinkmann V, Laube B, Abu Abed U, Goosmann C, Zychlinsky A. 2010. 531
Neutrophil extracellular traps: how to generate and visualize them. J Vis Exp 532
doi:10.3791/1724. 533
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

26
17. Bernatowicz P, Kowal K. 2016. Soluble CD163 modulates cytokine production 534
by peripheral blood mononuclear cells stimulated by Dermatophagoides 535
pteronyssinus allergens in vitro. Adv Med Sci 61:249-254. 536
18. Ermert D, Shaughnessy J, Joeris T, Kaplan J, Pang CJ, Kurt-Jones EA, Rice 537
PA, Ram S, Blom AM. 2015. Virulence of Group A Streptococci is enhanced by 538
human complement inhibitors. PLOS Pathogens 11. 539
19. Liu P, Pian Y, Li X, Liu R, Xie W, Zhang C, Zheng Y, Jiang Y, Yuan Y. 2014. 540
Streptococcus suis adenosine synthase functions as an effector in evasion of 541
PMN-mediated innate immunit. J Infect Dis 210:35-45. 542
20. Moon AF, Gaudu P, Pedersen LC. 2014. Structural characterization of the 543
virulence factor nuclease A from Streptococcus agalactiae. Acta Crystallogr D 544
Biol Crystallogr 70:2937-2949. 545
21. de Buhr N, Stehr M, Neumann A, Naim HY, Valentin-Weigand P, von 546
Kockritz-Blickwede M, Baums CG. 2015. Identification of a novel DNase of 547
Streptococcus suis (EndAsuis) important for neutrophil extracellular trap 548
degradation during exponential growth. Microbiology 161:838-850. 549
22. Mitterstiller AM, Haschka D, Dichtl S, Nairz M, Demetz E, Talasz H, Soares 550
M, Einwallner E, Esterbauer H, Fang FC, Geley S, Weiss G. 2016. Heme 551
oxygenase 1 controls early innate immune response of macrophages to Salmonella 552
typhimurium infection. Cell Microbiol doi:10.1111/cmi.12578. 553
23. Altincicek B, Stotzel S, Wygrecka M, Preissner KT, Vilcinskas A. 2008. 554
Host-derived extracellular nucleic acids enhance innate immune responses, induce 555
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

27
coagulation, and prolong survival upon infection in insects. J Immunol 556
181:2705-2712. 557
24. Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, 558
Weinrauch Y, Zychlinsky A. 2004. Neutrophil extracellular traps kill bacteria. 559
Science 303:1532-1535. 560
25. Daigo K, Hamakubo T. 2012. Host-protective effect of circulating pentraxin 3 561
(PTX3) and complex formation with neutrophil extracellular traps. Front Immunol 562
3:378. 563
26. de Buhr N, Neumann A, Jerjomiceva N, von Kockritz-Blickwede M, Baums 564
CG. 2014. Streptococcus suis DNase SsnA contributes to degradation of 565
neutrophil extracellular traps (NETs) and evasion of NET-mediated antimicrobial 566
activity. Microbiology 160:385-395. 567
27. Sumby P, Barbian KD, Gardner DJ, Whitney AR, Welty DM, Long RD, 568
Bailey JR, Parnell MJ, Hoe NP, Adams GG, Deleo FR, Musser JM. 2005. 569
Extracellular deoxyribonuclease made by group A Streptococcus assists 570
pathogenesis by enhancing evasion of the innate immune response. Proc Natl 571
Acad Sci U S A 102:1679-1684. 572
28. Zhu L, Kuang Z, Wilson BA, Lau GW. 2013. Competence-independent activity 573
of pneumococcal EndA mediates degradation of extracellular DNA and NETs and 574
Is important for virulence. PLoS One 8. 575
29. Chatellier S, Ihendyane N, Kansal RG, Khambaty F, Basma H, 576
Norrby-Teglund A, Low DE, McGeer A, Kotb M. 2000. Genetic relatedness 577
and superantigen expression in Group A Streptococcus serotype M1 isolated from 578
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

28
patients with severe and nonsevere invasive diseases. Infect Immun 579
68:3523-3534. 580
30. Zheng L, Khemlani A, Lorenz N, Loh JM, Langley RJ, Proft T. 2015. 581
Streptococcal 5'-nucleotidase A (S5nA), a novel Streptococcus pyogenes 582
virulence factor that facilitates immune evasion. J Biol Chem 290:31126-31137. 583
31. Thammavongsa V, Schneewind O, Missiakas DM. 2011. Enzymatic properties 584
of Staphylococcus aureus adenosine synthase (AdsA). BMC Biochem 12:56. 585
32. Fan J, Zhang Y, Chuang-Smith ON, Frank KL, Guenther BD, Kern M, 586
Schlievert PM, Herzberg MC. 2012. Ecto-5'-nucleotidase: a candidate virulence 587
factor in Streptococcus sanguinis experimental endocarditis. PLoS One 7:e38059. 588
33. Thammavongsa V, Kern JW, Missiakas DM, Schneewind O. 2009. 589
Staphylococcus aureus synthesizes adenosine to escape host immune responses. J 590
Exp Med 206:2417-2427. 591
34. H Z. 1992. 5'-Nucleotidase: molecular structure and functional aspects. Biochem 592
J 285:345-365. 593
35. Firon A, Dinis M, Raynal B, Poyart C, Trieu-Cuot P, Kaminski PA. 2014. 594
Extracellular nucleotide catabolism by the Group B Streptococcus 595
ectonucleotidase NudP increases bacterial survival in blood. J Biol Chem 596
289:5479-5489. 597
36. Haskó G. 2004. Adenosine: an endogenous regulator of innate immunity. Trends 598
Immunol 25:33-39. 599
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

29
FIG 1 The major nucleases and their virulence as determined by qRT-PCR and in a 600
mouse survival assay 601
(A) Changes in the transcription of predicted extracellular nuclease genes measured by 602
qRT-PCR during different growth phases of wild-type SEZ. SEZ GADPH 16S rRNA was 603
chosen as the internal control. The results are depicted as the mean ±SD (n = 3). (B) The 604
survival rate of mice challenged with wild-type SEZ and 3 gene deletion mutants. During 605
the first 72 h, all mice challenged with wild-type SEZ died. At 84 h, mice challenged 606
ΔENuc began to die, and all mice were dead by 120 h. Mice challenged with Δ5Nuc 607
began to die at 48 h post-infection, and almost 20% of mice survived by 148 h. Mice 608
challenged with double mutant ΔENucΔ5Nuc showed low mortality; approximately 50% 609
survived by 148 h. Virulence was compared between the three mutants and wild-type 610
SEZ. ***, p<0.001. 611
FIG 2 DNase activity of ENuc and 5Nuc. 612
(A) ENuc and 5Nuc degrade calf thymus DNA. Banding indicates the remaining DNA, 613
and lanes from left to right represent DNase I, reaction buffer, rENuc and r5Nuc 614
incubated with calf thymus DNA. (B) Wild-type SEZ and the 3 mutants degrade calf 615
thymus DNA. Lanes from left to right indicate remaining DNA after calf thymus DNA 616
incubated with wild-type SEZ, ΔENuc, Δ5Nuc, ΔENucΔ5Nuc, reaction buffer and DNase 617
I. 618
FIG 3 Production of deoxyadenosine detected by rpHPLC 619
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

30
(A) A standard deoxyadenosine sample (Sigma Aldrich) was evaluated by rpHPLC. (B) 620
Calf thymus DNA was evaluated by rpHPLC. (C) and (D) Deoxyadenosine production 621
following the incubation of rENuc and r5Nuc with calf thymus DNA. (E) NETs derived 622
from neutrophils induced by PMA were measured by rpHPLC. (F) to (K) 623
Deoxyadenosine production after the incubation of NETs with rENuc, r5Nuc, wild-type 624
SEZ, ΔENuc, Δ5Nuc and ΔENucΔ5Nuc was determined by rpHPLC. The arrows indicate 625
deoxyadenosine. 626
FIG 4 Hydrolysis of AMP, ADP and ATP by ENuc and 5Nuc 627
(A) and (B) Standard adonosine and AMP sample were evaluated by rpHPLC. (C) and (D) 628
AMP was hydrolyzed by rENuc and r5Nuc to generate adenosine. (E) Standard ADP 629
sample. (F) and (G) Adenosine evaluated by rpHPLC as a product of rENuc and r5Nuc 630
following incubation with ADP. (H) Standard ATP sample. (I) and (J) ATP was 631
hydrolyzed by rENuc and r5Nuc to generate adenosine. The arrows indicate the produced 632
adenosine. 633
FIG 5 Survival of wild-type SEZ and the 3 mutant strains in different organs 634
Rows represent the time points for sample collection. From top to bottom: 24 h, 48 h, 72 635
h and 96 h post-infection. Columns represent organs collected from infected mice. From 636
left to right: blood, liver, lung and spleen. The results are depicted as the mean ±SD (n = 637
5). *, p<0.05; **, p<0.01; ***, p<0.001; ns, no difference between groups. 638
FIG 6 NETs survival capacity of wild-type SEZ and its 3 mutants 639
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

31
Nearly all wild-type SEZ survived and proliferated in the presence of NETs, whereas 640
30%, 50% and 60% of ΔENuc, Δ5Nuc and ΔENucΔ5Nuc, respectively, were killed in the 641
presence of NETs. The results are depicted as the mean ±SD (n = 5); *, p<0.05; **, 642
p<0.01. 643
FIG 7 Effects of deoxyadenosine on RAW264.7 phagocytosis 644
Deoxyadenosine(-) indicates control groups that were not treated with deoxyadenosine; 645
deoxyadenosine(+) indicates groups that were pretreated with deoxyadenosine. (A) The 646
percent bactericidal activity of the control group was set to 100%. The bactericidal 647
activity of RAW264.7 cells treated with deoxyadenosine decreased to 40%. (B) △1 h, 648
△2 h and △3 h indicate the intracellular bactericidal rates of RAW264.7 cells between 1 649
and 2 h, 2 and 3 h and 3 and 4 h after extracellular bacteria were killed. Bactericidal 650
activity for each time range was compared. After 4 h, all intracellular bacteria were killed, 651
both in the control and treatment groups. (C) The bactericidal activity of monocytes is 652
evaluated by determining the CFU of viable bacteria after incubation. The CFU of viable 653
bacteria in the control group was set to 100%. The results are depicted as the mean ±SD 654
(n = 5); ***, p<0.001; ns, no differences between groups. 655
FIG 8 Measurement of the effects of deoxyadenosine on neutrophil function 656
Deoxyadenosine(-) indicates control groups that were not treated with deoxyadenosine; 657
deoxyadenosine(+) indicates treatment groups that were pretreated with deoxyadenosine. 658
(A) The bactericidal activity of neutrophils was evaluated by determining the CFU of 659
surviving bacteria after incubation. (B) PMA-stimulating NETs formation by neutrophils 660
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

32
was measured as the concentration of extracellular DNA, which was assessed with Pico 661
green and represented as relative fluorescence units (RFU). Resulted are depicted as the 662
mean ±SD n=5; ns indicates no differences between groups. 663
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 1. Strains and plasmids used in this study 1
Bacteria strain and plasmid Notable characteristic (s) Source of reference
SEZ strains
SEZ ATCC35246 Wild-type isolated from infected pig Purchased from ATCC
ΔENuc The SESEC_RS04165 (ENuc) deletion mutant of SEZ
ATCC35246
This study
Δ5Nuc The SESEC_RS05720 (5Nuc) deletion mutant of SEZ
ATCC35246
This study
ΔENucΔ5Nuc The ENuc and 5Nuc double gene deletion mutant of SEZ
ATCC35246
This study
E. coli strains
E. coli DH5a Plasmid cloning host and Gram-negative replicon Purchased from Invitrogen
E. coli Rosetta E. coli - plasmid expressing host Purchased from Invitrogen
E. coli Rosetta 32a-ENuc E. coli Rosetta carrying plasmid pET32a-ENuc, Ampr This study
E. coli Rosetta 28a-5Nuc E. coli Rosetta carrying plasmid pET28a-5Nuc, Kmr This study
plasmids
pSET4s Streptococcus thermosensitive suicide vector, Spcr Takamatsu, 2001
pSET4s-ΔENuc pSET4s containing the upstream and downstream sequences This study
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

surrounding ENuc for deleting ENuc, Spcr
pSET4s-Δ5Nuc pSET4s containing the upstream and downstream sequences
surrounding 5Nuc for deleting 5Nuc, Spcr
This study
pET-32a (+) E. coli – gene expressing vector, Ampr Purchased from Invitrogen
pET32a-ENuc pET-32a containing ENuc gene, Ampr This study
pET-28a (+) E. coli – gene expressing vector, Kmr Purchased from Invitrogen
pET28a-5Nuc pET-28a containing 5Nuc gene, Kmr This study
Ampr, ampicillin resistance cassette; Kmr, kanamycin resistance cassette; Spcr, 2
spectinomycin resistance cassette. 3
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 2. Predicted genes encoding extracellular nucleases in SEZ 1
Gene ID Producta Abbreviation
SESEC_RS04165
SESEC_RS05720
nuclease
5’-nucleotidase
ENuc
5Nuc
SESEC_RS02695 CRISP-associated protein Cas4
SESEC_RS03220 DNA/RNA non-specific endonuclease
SESEC_RS04065 DNAse
SESEC_RS04270 Competence associated endonuclease
SESEC_RS05625 DNA/RNA non-specific endonuclease
aData information referred to genomic information for Streptococcus equi ssp. 2
zooepidemicus ATCC 35246 (NC_017582.1) in the NCBI database. 3
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 3. Primers used in this study 1
Primer Sequence (5ʹ-3ʹ) Restriction
enzyme
Fl-ENucF TCACTGGCTGCTGAGTTTATT
Fl-ENucR CGGCTGAACACGTCCATATAA
Fl-5NucF TCAGTCAAGGTGGGCTTTATC
Fl-5NucR CCACTCTAGGCAACCATCTTC
Fl-Nuc1F GCTTGGGTCTGTATGGGATTAT
Fl-Nuc1R CCACTCTAGGCAACCATCTTC
Fl-Nuc2F CTGGAGTGCTTTCGTCTGTAATA
Fl-Nuc2R TACCAATATGGCGGCATCAG
Fl-Nuc3F GCATTCCTGTGCGGTAATCTA
Fl-Nuc3R CAAATTGGGAGGCAACAAAGAA
Fl-Nuc4F CTTCTAGGCTATGCGCTTGTT
Fl-Nuc4R CTCTCGTTAGCTGCTTGGTTAG
Fl-Nuc5F ATTACCAATCCAGCCAGAAGG
Flu-Nuc5R CAGGTCGCAAGAGGAACAATA
ENucF CGCGGATCCCAGTCTTCAGCTCAGCATGTTTATG BamHI
ENucR GCCCTCGAG G CTTCCTTTCCTGACGTTTCCTA XhoI
ENuc-L-F GCCGGATCCAGCTTAGCTCGTTATGGAAGAGTGT BamHI
ENuc-L-R AAAGAAATCCTTTACTTCCTTTCCTTTGACATAAAAACTCCTTTTTAATG
ENuc-R-F CATTAAAAAGGAGTTTTTATGTCAAAGGAAAGGAAGTAAAGGATTTCTTT
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from

2
Restrict enzyme sites were underlined. 3
ENuc-R-R GCCGTCGACTCTACCACGTTTGTAATATGCTTGG SalI
5NucF CGCGGATCCGATCAGGTAGACGTGCAATTTCTTG BamHI
5NucR GCCCTCGAGTTATAAGGCAGCATTAGCTTGCTTT XhoI
5Nuc-L-F GCCGGATCCGGAATAATAAAAGCAATCATACCAC BamHI
5Nuc-L-R
5Nuc-R-F
5Nuc-R-R
GCCGGTTTTATTGGCCTTCCTTTTTAAACAAAATGCTTTTTCATAAACGC
GCGTTTATGAAAAAGCATTTTGTTTAAAAAGGAAGGCCAATAAAACCGGC
GCCGTCGACTCATCGATTGTATGACATCATGAAG
SalI
on March 20, 2020 by guest
http://aem.asm
.org/D
ownloaded from


















