
Wk2_Ch02,03_F09
11
9/8/09 1 Chapter 2 Transport of ions and small molecules across membranes By Stephan E. Lehnart & Andrew R. Marks Cell Biology, Roosevelt University, Fall 2009 2.1 Introduction • The membrane potential across a cell membrane is due to: – an electrochemical gradient across a membrane – a membrane that is selectively permeable to ions • Cells maintain a negative resting membrane potential with the inside of the cell slightly more negative than the outside. • The membrane potential is a prerequisite for electrical signals and for directed ion movement across cellular membranes. 2.4 Electrochemical gradients across the cell membrane generate the membrane potential Action Potentials • Action potentials enable rapid communication between cells. • Na + , K + , and Ca 2+ currents are key elements of action potentials. • Membrane depolarization is mediated by the flow of Na + ions into cells through voltage- dependent Na + channels. • Repolarization is shaped by transport of K + ions through several different types of K + channels. • The electrical activity of organs can be measured as the sum of action potential vectors. • Alterations of the action potential can predispose for arrhythmias or epilepsy. 2.12 Action potentials are electrical signals that depend on several types of ion channels
Transcript of Wk2_Ch02,03_F09

8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 1/11
9/8/09
1
Chapter 2
Transport of ions and small molecules
across membranesBy
Stephan E. Lehnart & Andrew R. Marks
Cell Biology, Roosevelt University, Fall 2009
2.1 Introduction
• The membrane potential across a cell
membrane is due to:
– an electrochemical gradient across a membrane
– a membrane that is selectively permeable to ions
• Cells maintain a negative resting membrane
potential with the inside of the cell slightly
more negative than the outside.
• The membrane potential is a prerequisite for
electrical signals and for directed ion
movement across cellular membranes.
2.4 Electrochemical gradients across the cell membrane generate the membrane potential
Action Potentials
• Action potentials enable rapid communicationbetween cells.
• Na+, K+, and Ca2+ currents are key elementsof action potentials.
• Membrane depolarization is mediated by theflow of Na+ ions into cells through voltage-dependent Na+ channels.
• Repolarization is shaped by transport of K+
ions through several different types of K+
channels.
• The electrical activity of organs can be
measured as the sum of action potential
vectors.
• Alterations of the action potential can
predispose for arrhythmias or epilepsy.
2.12 Action potentials are electrical signals that depend on several types of ion channels

8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 2/11

8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 3/11
9/8/09
3
• The selectivity filter of K+ channels is an
evolutionarily conserved structure.
• The K+ channel selectivity filter catalyzes
dehydration of ions, which:
– confers specificity
– speeds up ion permeation
2.5 K+ channels catalyze selective and rapid ion permeation 2.5 K+ channels catalyze selective and rapid ion permeation
• Gating is an essential
property of ion
channels.
• Different gating
mechanisms definefunctional classes of K+
channels.
• The K+ channel gate is distinct from the
selectivity filter.
• K+ channels are regulated by the membrane
potential.
2.6 Different K+ channels use a similar gate coupled to different activating or inactivating mechanisms.
• The inwardly directed Na+ gradient maintained by the
Na+/K+-ATPase is required for the function of Na+
channels.
• Electrical signals at the cell membrane activate
voltage-dependent Na+ channels.
• The pore of voltage-dependent Na+ channels is
formed by one subunit, but its overall architecture is
similar to that of 6TM/1P K+ channels.
• Voltage-dependent Na+ channels are inactivated by
specific hydrophobic residues that block the pore.
2.7 Voltage-dependent Na+ channels are activated by membrane depolarization and translate electrical signals

8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 4/11
9/8/09
4
• Cell surface Ca2+ channels translate membrane
signals into intracellular Ca2+ signals. Muscle Contraction
• The process of excitation-contraction
coupling, which is initiated by membrane
depolarization, controls muscle contraction.
• Ryanodine receptors and inositol 1,4,5-
trisphosphate receptors are Ca2+ channels
that release Ca2+ into the cytosol from
intracellular stores
• Intracellular Ca2+ release through ryanodine
receptors in the sarcoplasmic reticulummembrane stimulates contraction of the
myofilaments.
• Several different types of Ca2+ transport
proteins, including the Na+/Ca2+-exchanger
and Ca2+-ATPase are important for
– decreasing the cytosolic Ca2+ concentration
– controlling muscle relaxation
2.13 Cardiac and skeletal muscles are activated by excitation-contraction coupling 2.13 Cardiac and skeletal muscles are activated by excitation-contraction coupling
Aquaporins
• Aquaporins allow rapid
and selective water
transport across cellmembranes.
• Aquaporins aretetramers of four
identical subunits, with
each subunit forming a
pore.
• The aquaporin selectivity filter has three
major features that confer a high degree of
selectivity for water:
– size restriction
– electrostatic repulsion
– water dipole orientation
2.11 Selective water transport occurs through aquaporin channels
8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 5/11
9/8/09
5
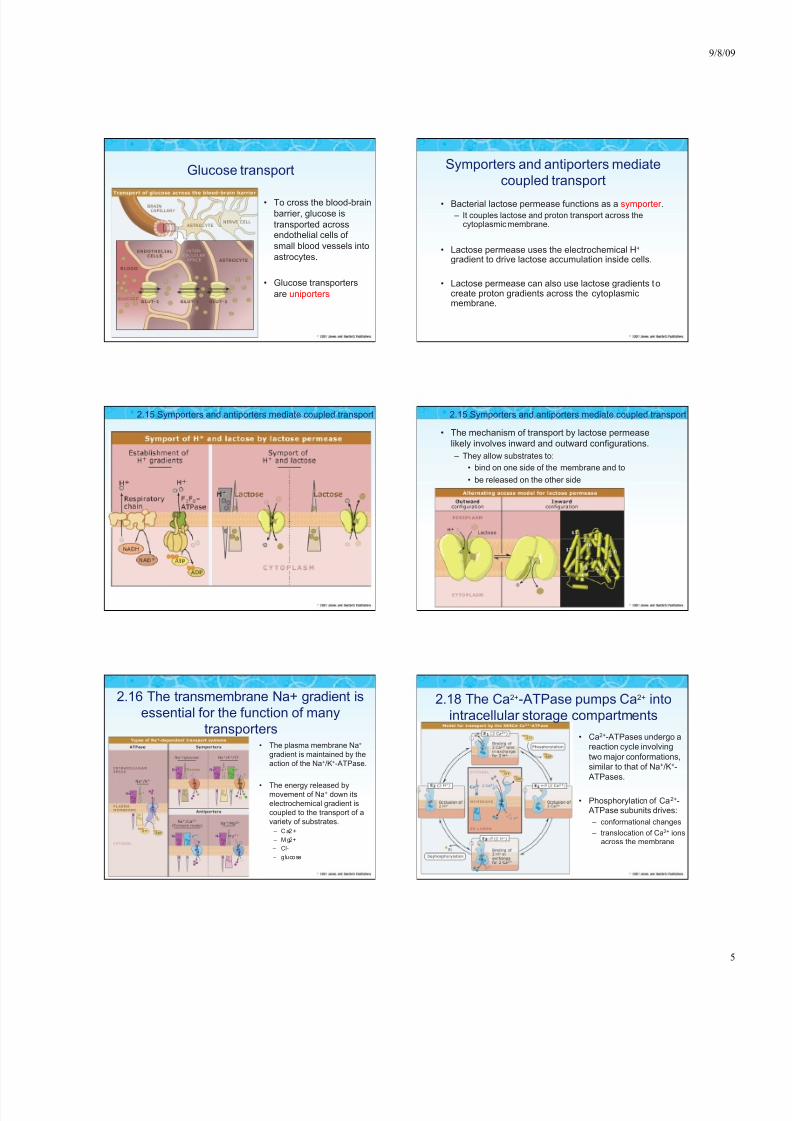
Glucose transport
• To cross the blood-brain
barrier, glucose is
transported acrossendothelial cells of
small blood vessels into
astrocytes.
• Glucose transporters
are uniporters
Symporters and antiporters mediate
coupled transport
• Bacterial lactose permease functions as a symporter .
– It couples lactose and proton transport across thecytoplasmic membrane.
• Lactose permease uses the electrochemical H+ gradient to drive lactose accumulation inside cells.
• Lactose permease can also use lactose gradients tocreate proton gradients across the cytoplasmicmembrane.
2.15 Symporters and antiporters mediate coupled transport
• The mechanism of transport by lactose permease
likely involves inward and outward configurations.
– They allow substrates to:
• bind on one side of the membrane and to
• be released on the other side
2.15 Symporters and antiporters mediate coupled transport
2.16 The transmembrane Na+ gradient is
essential for the function of many
transporters • The plasma membrane Na+
gradient is maintained by the
action of the Na+/K+-ATPase.
• The energy released bymovement of Na+ down its
electrochemical gradient is
coupled to the transport of a
variety of substrates.
– Ca2 +
– Mg2+
– Cl-
– glucose
2.18 The Ca2+-ATPase pumps Ca2+ into
intracellular storage compartments
• Ca2+-ATPases undergo areaction cycle involving
two major conformations,
similar to that of Na+/K+-
ATPases.
• Phosphorylation of Ca2+-ATPase subunits drives:
– conformational changes
– translocation of Ca2+ ionsacross the membrane

8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 6/11
9/8/09
6
2.19 The Na+/K+-ATPase maintains the
plasma membrane Na+ and K+ gradients
• The Na+/K+-ATPase is a P-type ATPase that is
similar to the Ca2+-ATPase and the H+-ATPase.
• The Na+/K+-ATPase maintains the Na+ and K+
gradients across the plasma membrane.
• The plasma membrane Na+/K+-ATPase is
electrogenic:
– it transports three Na+ ions out of the cell for every two K+
ions it transports into the cell.
2.20 The F1Fo-ATP synthase couples H+
movement to ATP synthesis or hydrolysis
• The F1Fo-ATP synthase is a
key enzyme in oxidative
phosphorylation.
• The F1Fo-ATP synthase is a
multisubunit molecular
motor.
– It couples the energy
released by movement of protons down their
electrochemical gradient toATP synthesis.
2.21 H+-ATPases transport protons out of
the cytosol
• Proton concentrations affectmany cellular functions.
• Intracellular compartments areacidified by the action of V-ATPases.
• V-ATPases are proton pumpsthat consist of multiplesubunits, with a structure
similar to F1Fo-ATP synthases.
• V-ATPases in the
plasma membraneserve specialized
functions in:
– acidification of
extracellular fluids
– regulation of cytosolicpH
2.21 H+-ATPases transport protons out of the cytosol
Chapter 3
Membrane targeting of proteinsBy
D. Thomas Rutkowski & Vishwanath R. Lingappa
• Cells must localize proteins to
specific organelles and
membranes.
• Proteins are imported from the
cytosol directly into several types
of organelles.
• The endoplasmic reticulum (ER):
– is the entry point for proteinsinto the secretory pathway
– is highly specialized for that
purpose
• Several other organelles and theplasma membrane receive their
proteins by way of the secretory
pathway.
3.1 Introduction

8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 7/11
9/8/09
7
• Signal sequences targetnascent secretory and
membrane proteins to theER for translocation.
• The only feature commonto all signal sequences is
a central, hydrophobic
core that is usually
sufficient to translocateany associated protein.
• Secretory proteins translocate completely across the ERmembrane; – transmembrane proteins are integrated into the membrane.
• Before leaving the ER, proteins are modified and folded byenzymes and chaperones in the lumen.
3.2 Proteins enter the secretory pathway by translocation across the ER membrane
• Docking of SRP with its
receptor brings the
ribosome and nascentchain into proximity with
the translocon.
• Docking requires theGTP binding and
hydrolysis activities of
SRP and its receptor.
• Proteins translocate through
an aqueous channel
composed of the Sec61complex, located within the
ER membrane.
• Numerous accessoryproteins that are involved in:
– Translocation
– Folding
– Modification associate with thechannel
• An interaction
between the
translocon and thesignal sequence
causes the channel
to open and initiates
translocation.
• The exact
mechanism of
translocation mayvary from one
protein to another.
• An interaction
between the
translocon and thesignal sequence
causes the channel
to open and initiates
translocation.
• The exact
mechanism of
translocation mayvary from one
protein to another.
8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 8/11
9/8/09
8
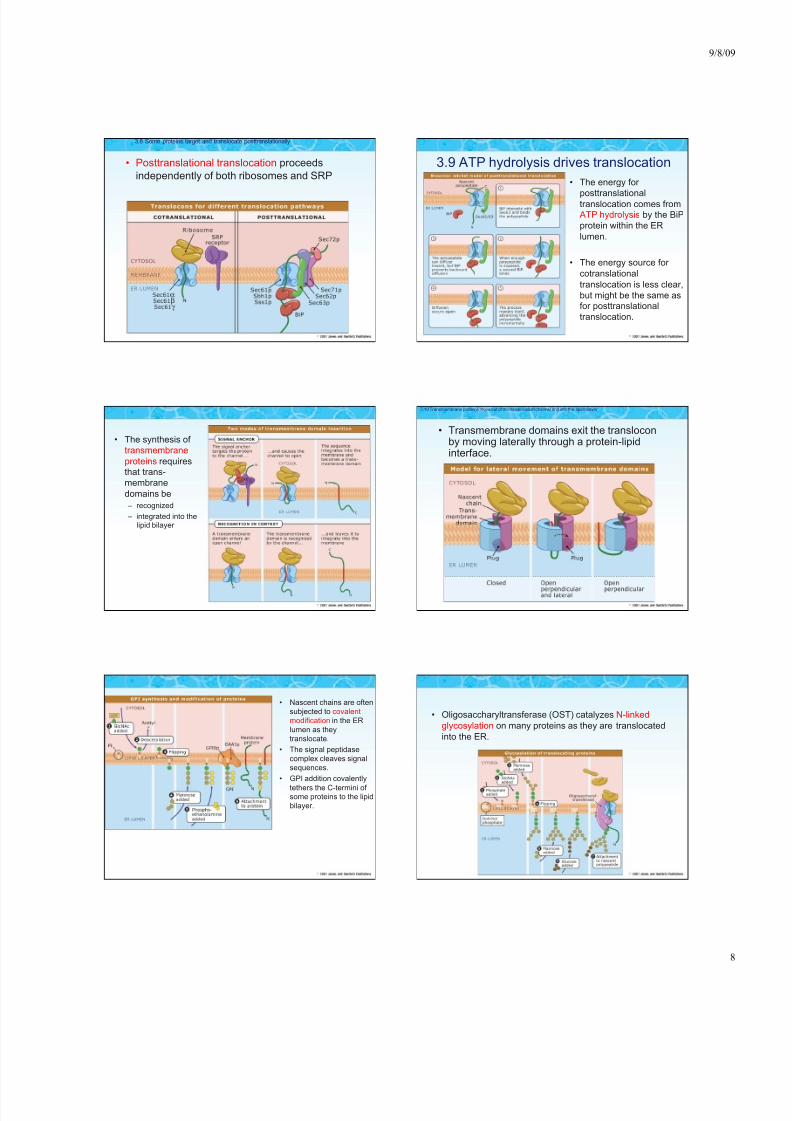
• Posttranslational translocation proceeds
independently of both ribosomes and SRP
3.8 Some proteins target and translocate posttranslationally
3.9 ATP hydrolysis drives translocation
• The energy for
posttranslationaltranslocation comes fromATP hydrolysis by the BiP
protein within the ER
lumen.
• The energy source for
cotranslational
translocation is less clear,
but might be the same asfor posttranslational
translocation.
• The synthesis of
transmembrane
proteins requiresthat trans-
membrane
domains be
– recognized
– integrated into thelipid bilayer
• Transmembrane domains exit the transloconby moving laterally through a protein-lipidinterface.
3.10 Transmembrane proteins move out of the translocation channel and into the lipid bilayer
• Nascent chains are often
subjected to covalent
modification in the ER
lumen as they
translocate.
• The signal peptidase
complex cleaves signal
sequences.
• GPI addition covalently
tethers the C-termini of some proteins to the lipid
bilayer.
• Oligosaccharyltransferase (OST) catalyzes N-linked
glycosylation on many proteins as they are translocated
into the ER.

8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 9/11
9/8/09
9
• Molecular chaperones associate with proteins in the
lumen and assist their folding.
• Subunits that have not yet assembled into complexes are
retained in the ER by interaction with chaperones.
• Protein disulfide isomerases (PDI) catalyze disulfide
bond formation and rearrangement in the ER.
• Calnexin and
calreticulin escortglycoproteins
through repeated
cycles of
chaperoning.
– The cycles arecontrolled by
addition and
removal of
glucose.
• Proteins are returned to the cytosol by the
process of retrograde translocation.
3.19 Terminally misfolded proteins in the ER are returned to the cytosol for degradation
• The major cellular
phospholipids are
synthesized predominantly
on the cytosolic face of the
ER membrane.
• The localization of enzymes
involved in lipid
biosynthesis can be
controlled by the cell to
regulate the generation of
new lipids.
• Cholesterol biosynthesis is
regulated by proteolysis of
a transcription factor
integrated into the ER
membrane.
• Each organelle has a unique composition of lipids.
– This requires that lipid transport from the ER toeach organelle be a specific process.
• The mechanisms of lipid transport betweenorganelles are unclear.
– They might involve direct contact between the ERand other membranes in the cell.
• Transbilayer movement of lipids establishesasymmetry of membrane leaflets.

8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 10/11

8/8/2019 Wk2_Ch02,03_F09
http://slidepdf.com/reader/full/wk2ch0203f09 11/11


















