
Whole-mount confocal imaging of nuclei in giant feeding cells induced by root-knot nematodes in...
9
Methods Whole-mount confocal imaging of nuclei in giant feeding cells induced by root-knot nematodes in Arabidopsis Paulo Vieira 1,2,3 , Gilbert Engler 1,2,3 * and Janice de Almeida Engler 1,2,3 * 1 Institut National de la Recherche Agronomique, UMR 1355 ISA, 400 route des Chappes, Sophia-Antipolis, France; 2 Centre National de la Recherche Scientifique, UMR 7254 ISA, 400 route des Chappes, Sophia-Antipolis, France; 3 Universite ´ de Nice-Sophia Antipolis, UMR ISA, 400 route des Chappes, Sophia-Antipolis, France Author for correspondence: Janice de Almeida Engler Tel: +33 492 386 459 Email: [email protected] Received: 22 March 2012 Accepted: 6 April 2012 New Phytologist (2012) 195: 488–496 doi: 10.1111/j.1469-8137.2012.04175.x Key words: Arabidopsis roots, giant cell, nuclei, Kip-related protein 4 (KRP4), Meloidogyne. Summary • Excellent visualization of nuclei was obtained here using a whole-mount procedure adapted to provide high-resolution images of large, irregularly shaped nuclei. The procedure is based on tissue clearing, and fluorescent staining of nuclear DNA with the dye propidium iodide. • The method developed for standard confocal imaging was applied to large multicellular root swellings, named galls, induced in plant hosts by the root-knot nematode Meloidogyne incognita. • Here, we performed a functional analysis, and examined the nuclear structure in giant feed- ing cells overexpressing the cell cycle inhibitor Kip-related protein 4 (KRP4). Ectopic KRP4 expression in galls led to aberrant nuclear structure, disturbing giant cell expansion and nema- tode reproduction. In vivo live-cell imaging of GFP-KRP4 demonstrated that this protein co-localizes to chromosomes from prophase to late anaphase during cell cycle progression. • The data presented here suggest the involvement of KRP4 during mitotic progression in plant cells. The detailed results obtained using confocal analysis also demonstrate the potential utility of a rapid, easy-to-use clearing method for the analysis of the nuclei of certain Arabidopsis mutants and other complex plant nuclei. Introduction Root-knot nematodes (Meloidogyne spp.) are plant-parasitic nem- atodes capable of inducing large feeding sites within the root vas- cular tissue of plants. Feeding sites contain five to seven hypertrophied giant cells resulting from the substantial develop- mental reprogramming of host root cells caused by the pathogen. Giant cells undergo multiple rounds of nuclear division without cytokinesis, giving rise to large multinucleated cells (Sijmons et al., 1994; de Almeida Engler et al., 1999). These cells are highly metabolically active, and provide the nematode with nutri- ents necessary for its development and life cycle completion (Jones & Payne, 1978; Davis et al., 2000). As the development of the nematode feeding site (NFS) progresses, multiple rounds of DNA synthesis occur in the nuclei of giant cells, coupled with nuclear and cellular expansion (de Almeida Engler et al., 1999). Parallel with giant cell development, neighbouring root cells pro- liferate to form a multilayered cellular coat surrounding the giant feeding cells. Hyperplasia of neighbouring cells and hypertrophy of giant cells form the typical root-knot gall enclosing the nema- todes (Jones & Northcote, 1972). Plant–nematode interactions are often studied by performing detailed cytological analyses of transgenic lines compared with wild type in order to better understand major changes occurring during NFS development. Most of these cytological analyses are based on the physical production of gall sections using classical microtomy techniques. Although these sections can be easily stained with various dyes and microscopically observed, the method remains limited. A major drawback when using sectioned gall tissues for studying nuclei in giant cells is the loss of spatial perspective, especially when studying complex-shaped nuclei within feeding cells. Furthermore, reconstructing a 3D view from individual tissue sections from a complex architecture remains quite difficult and time consuming. In Arabidopsis, whole-mount (WM) techniques are well estab- lished for studying the nuclei of different plant tissues (e.g. root meristems, endosperm or leaves) as well, for DNA quantification, nuclei in situ hybridizations, and others (Bauwens et al., 1994; Boisnard-Lorig et al., 2001; Willemse et al., 2008; Tirichine et al., 2009). These methods, coupled with confocal microscopy and computer-aided modelling software, provide detailed quali- tative and quantitative information on the nuclear changes occur- ring in different plant tissues. Alternatively, an elegant WM procedure has been established for the model plant Arabidopsis *These authors contributed equally to this work. Research 488 New Phytologist (2012) 195: 488–496 www.newphytologist.com ȑ 2012 INRA New Phytologist ȑ 2012 New Phytologist Trust
-
Upload
paulo-vieira -
Category
Documents
-
view
212 -
download
0
Transcript of Whole-mount confocal imaging of nuclei in giant feeding cells induced by root-knot nematodes in...

Methods
Whole-mount confocal imaging of nuclei in giant feeding cellsinduced by root-knot nematodes in Arabidopsis
Paulo Vieira1,2,3, Gilbert Engler1,2,3* and Janice de Almeida Engler1,2,3*1Institut National de la Recherche Agronomique, UMR 1355 ISA, 400 route des Chappes, Sophia-Antipolis, France; 2Centre National de la Recherche Scientifique, UMR 7254 ISA, 400 route
des Chappes, Sophia-Antipolis, France; 3Universite de Nice-Sophia Antipolis, UMR ISA, 400 route des Chappes, Sophia-Antipolis, France
Author for correspondence:Janice de Almeida Engler
Tel: +33 492 386 459
Email: [email protected]
Received: 22 March 2012
Accepted: 6 April 2012
New Phytologist (2012) 195: 488–496doi: 10.1111/j.1469-8137.2012.04175.x
Key words: Arabidopsis roots, giant cell,nuclei, Kip-related protein 4 (KRP4),Meloidogyne.
Summary
• Excellent visualization of nuclei was obtained here using a whole-mount procedure adapted
to provide high-resolution images of large, irregularly shaped nuclei. The procedure is based
on tissue clearing, and fluorescent staining of nuclear DNA with the dye propidium iodide.
• The method developed for standard confocal imaging was applied to large multicellular root
swellings, named galls, induced in plant hosts by the root-knot nematode Meloidogyne incognita.
• Here, we performed a functional analysis, and examined the nuclear structure in giant feed-
ing cells overexpressing the cell cycle inhibitor Kip-related protein 4 (KRP4). Ectopic KRP4
expression in galls led to aberrant nuclear structure, disturbing giant cell expansion and nema-
tode reproduction. In vivo live-cell imaging of GFP-KRP4 demonstrated that this protein
co-localizes to chromosomes from prophase to late anaphase during cell cycle progression.
• The data presented here suggest the involvement of KRP4 during mitotic progression in
plant cells. The detailed results obtained using confocal analysis also demonstrate the potential
utility of a rapid, easy-to-use clearing method for the analysis of the nuclei of certain
Arabidopsis mutants and other complex plant nuclei.
Introduction
Root-knot nematodes (Meloidogyne spp.) are plant-parasitic nem-atodes capable of inducing large feeding sites within the root vas-cular tissue of plants. Feeding sites contain five to sevenhypertrophied giant cells resulting from the substantial develop-mental reprogramming of host root cells caused by the pathogen.Giant cells undergo multiple rounds of nuclear division withoutcytokinesis, giving rise to large multinucleated cells (Sijmonset al., 1994; de Almeida Engler et al., 1999). These cells arehighly metabolically active, and provide the nematode with nutri-ents necessary for its development and life cycle completion(Jones & Payne, 1978; Davis et al., 2000). As the developmentof the nematode feeding site (NFS) progresses, multiple roundsof DNA synthesis occur in the nuclei of giant cells, coupled withnuclear and cellular expansion (de Almeida Engler et al., 1999).Parallel with giant cell development, neighbouring root cells pro-liferate to form a multilayered cellular coat surrounding the giantfeeding cells. Hyperplasia of neighbouring cells and hypertrophyof giant cells form the typical root-knot gall enclosing the nema-todes (Jones & Northcote, 1972).
Plant–nematode interactions are often studied by performingdetailed cytological analyses of transgenic lines compared withwild type in order to better understand major changes occurringduring NFS development. Most of these cytological analyses arebased on the physical production of gall sections using classicalmicrotomy techniques. Although these sections can be easilystained with various dyes and microscopically observed, themethod remains limited. A major drawback when using sectionedgall tissues for studying nuclei in giant cells is the loss of spatialperspective, especially when studying complex-shaped nucleiwithin feeding cells. Furthermore, reconstructing a 3D view fromindividual tissue sections from a complex architecture remainsquite difficult and time consuming.
In Arabidopsis, whole-mount (WM) techniques are well estab-lished for studying the nuclei of different plant tissues (e.g. rootmeristems, endosperm or leaves) as well, for DNA quantification,nuclei in situ hybridizations, and others (Bauwens et al., 1994;Boisnard-Lorig et al., 2001; Willemse et al., 2008; Tirichineet al., 2009). These methods, coupled with confocal microscopyand computer-aided modelling software, provide detailed quali-tative and quantitative information on the nuclear changes occur-ring in different plant tissues. Alternatively, an elegant WMprocedure has been established for the model plant Arabidopsis*These authors contributed equally to this work.
Research
488 New Phytologist (2012) 195: 488–496
www.newphytologist.com� 2012 INRA
New Phytologist � 2012 New Phytologist Trust

based on the specific fluorescent staining of plant cell walls, usingcovalently attached propidium iodide (PI) fluorochromes. Thismethod allows visualization of the plant cellular organization inWM to depths of > 200 lm, and was successfully used to gener-ate excellent 3D reconstructions of protophloem development(Truernit & Haseloff, 2008). Although excellent for staining thecell walls of WM samples, this procedure does not stain nuclearDNA, even in the presence of the double-stranded DNA interca-lating dye PI. Therefore, to stain nuclear DNA with PI in clearedWMs, we took advantage of established protocols (Bauwenset al., 1994; Willemse et al., 2008), and adapted the methodol-ogy to visualize nuclei in galls induced by root-knot nematodes.This allowed us to study a cell cycle inhibitor, Kip-related protein4 (interactor ⁄ inhibitor of CDK (inhibitor/interactor of cyclin-dependent kinase, ICK7/KRP4)), in Arabidopsis tissue thatshowed altered nuclear morphology. The WM method presentedfacilitates the study of nuclear organization in complex plant tis-sues, such as galls, while avoiding laborious tissue sectioning.Although the molecular mechanism behind the formation anddevelopment of the NFSs is still not well understood, the charac-terization of several core cell cycle genes highly expressed duringNFS development implies the involvement of the host plant cellcycle machinery during this plant-pathogen interaction (Niebelet al., 1996; de Almeida Engler et al., 1999, 2011, and deAlmeida Engler et al., unpublished data).
One of the gene families involved in the regulation of thecyclin-dependent kinase (CDK) ⁄ cyclin (CYC) complexesduring the cell cycle are the ICK ⁄ KRPs (Wang et al., 1997;De Veylder et al., 2001; Inze & De Veylder, 2006). ICK ⁄ KRPgenes have been associated with the switch from mitosis to en-docycle and have been suggested to be involved in the varioussteps of cell division by binding to and inactivatingCDK ⁄ CYC complexes. Previous studies have shown thatectopic expression of ICK ⁄ KRP genes affects nuclear ploidy lev-els. Low ICK ⁄ KRP levels block cell division and induce en-doreduplication, while overexpression of ICK ⁄ KRP genesnegatively affects cell division and endoreduplication cycles,resulting in a decrease in nuclear DNA content (Wang et al.,2000; De Veylder et al., 2001; Zhou et al., 2002; Verkestet al., 2005). Here we present data illustrating that expressionof ICK7 ⁄ KRP4 (hereafter termed KRP4) in galls stronglyaffects nuclear morphology in giant cells. This phenotype isunique among members of the ICK ⁄ KRP family in Arabidop-sis. We show that a simplified WM protocol can be efficientlyused to study nuclear morphology throughout giant celldevelopment, permitting the study of nuclear rearrangementsthat occur in complex organs such as galls induced byroot-knot nematodes in plant roots.
Materials and Methods
Nematode inoculation tests
For both transgenic line and wild-type Arabidopsis thaliana (L.)Heynh Columbia (Col-0), c. 60 sterilized seeds were placed insterile Petri dishes containing 1% Murashige and Skoog (MS)
germination medium (Duchefa, Haarlem, the Netherlands), 1%sucrose and 0.8% plant cell culture-tested agar (Sigma-Aldrich),supplemented with the appropriate antibiotics. After 2 d of coldtreatment in the dark, seeds were kept in a growth chamber witha 16-h light : 8-h dark photoperiod at 21�C : 18�C, respectively.Fourteen-day-old plants were transferred to MS medium, andfive plants per plate were placed in eight replica plates per line.One week later, root tips were inoculated with c. 100 surface-sterilized, freshly hatched, second-stage juveniles (J2s) of Meloidogyneincognita. All infection tests were performed in three independentexperiments for both transgenic and wild-type lines. Transgenic T1lines overexpressing ICK7 ⁄KRP4, driven by the cauliflower mosaicvirus (CaMV) 35S promoter (35S::GFP-ICK7 ⁄KRP4, designatedKRP4OE hereafter), were kindly provided by E. Russinova (Depart-ment of Plant Systems Biology, Flanders Institute for Biotechnology,Gent, Belgium). Kanamycin-resistant T3 KRP4OE lines were gener-ated and evaluated based on KRP4 gene expression levels, proteinlocalization and plant phenotype.
RNA isolation and semiquantitative RT-PCR amplification
To examine the expression of KRP4 in wild-type and KRP4OE
Arabidopsis plants, total RNA was extracted with Trizol reagent(Invitrogen) from 7-d-old whole seedlings, using the RNeasyPlant Mini Kit (Qiagen) according to the manufacturer’s instruc-tions. The RNA was treated with RQ1 RNase-free DNase (Pro-mega) before reverse transcription. One microgram of treatedRNA was added to RT reactions using a Bio-Rad iScriptfirst-strand synthesis kit with random hexamer primers to makecDNA, according to the manufacturer’s instructions. Semiquan-titative RT-PCR amplification was performed using KRP4 for-ward (Fw, 5¢-AACCCCGGGATCGTCTAC-3¢) and reverse(Rv, 5¢-GCACCCGAGAAAAACTCG-3¢) primers. The ArabidopsisElongation Factor 1-a (EF1-a) gene was used as a control (Fw,5¢-GCTCTATGGAAGTTCGAGACCAC and Rv, 5¢-GGTGTGGCAATCGAGAACTG).
Whole-mount analysis of giant cell nuclei
Nuclear analyses of cleared feeding sites were performed basedon a WM mRNA in situ hybridization protocol described byBauwens et al. (1994) and Willemse et al. (2008), and adaptedwith the following modifications. Four-week-old, uninfectedand nematode-infected roots from 1 to 40 d after inoculation(DAI) were fixed in phosphate-buffered saline (PBS), pH 7.2,containing 1% formaldehyde and 10% dimethyl sulfoxide(DMSO) for 30 min at 20�C. Plant material was subsequentlydehydrated twice with absolute methanol, and four times withabsolute ethanol for 10 min for each step. Samples were storedat )20�C for 2–4 d for tissue clearing. Subsequently, seedlingswere rinsed twice for 10 min with ethanol and for 40 min inethanol : xylene (1 : 1). Samples were then washed twice withabsolute ethanol and absolute methanol, for 5 min each, andpost-fixed for 5 min in PBS, containing 0.1% Tween 20 (PBT)and 1% formaldehyde. Following post-fixation, roots werewashed five times with PBT for 5 min. Samples were then
NewPhytologist Research 489
� 2012 INRA
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 195: 488–496
www.newphytologist.com

washed twice for 5 min with PBS containing 50% formamide,and boiled twice for 3 min followed by quenching on ice. Aftersamples had been stored overnight at room temperature (RT),they were washed four times in PBS for 15 min and stainedwith 0.5 lg ml)1 propidium iodide (PI) in PBS for 30 min atRT. Stained samples were mounted in 90% glycerol on amicroscope slide and cover-slipped.
Confocal microscopy
Image acquisition was performed using a Zeiss LSM 510META confocal laser scanning microscopy. The excitationwavelength for PI-stained samples was 543 nm, and the fluores-cence emission was collected between 560 and 650 nm.High-resolution optical sections were acquired to distances >200 lm inside the cleared galls using a ·40 1.3 oil objectivelens. By combining the high fluorescence levels of the PI-stainednuclei and a high refraction index mounting medium, it waspossible to collect extended z-series images with good resolu-tion, allowing appropriate 3D projection of nuclei. Z-stackswere processed using the LSM 510 software to create maximumbrightness projections and 3D view representations of nuclei.To visually improve image contrast, the red fluorescence signalof PI was converted to white.
Morphological analysis
For observation of the morphology of gall tissues, infectedroots were collected at 14, 28 and 40 DAI, and fixed in 2%glutaraldehyde in 50 mM PIPES buffer, pH 6.9. Subsequently,samples were dehydrated and embedded in Technovit 7100(Heraeus Kulzer, Wehrheim, Germany), as described by themanufacturer. Embedded roots and gall tissues were sectioned(3 lm) and stained with 0.05% toluidine blue and mountedin Depex (Sigma-Aldrich). Microscopic observations wereperformed using bright-field optics and images taken with adigital camera (AxioCam HRc; Zeiss).
In vivo observations of fresh gall root slices andwhole-mount roots
Observations of nuclei in galls induced by M. incognita wereperformed in infected Arabidopsis roots harbouring free GFP(35S::GFP), and in the KRP4OE line. Galls at various time-pointsafter infection (7–40 DAI) were dissected from roots and embed-ded in 5% agar. Fresh thick sections of 50–100 lm (7–14 DAI)or 150–200 lm (14–40 DAI) were obtained with a HM650Vvibratome (Microm, Walldorf, Germany). Whole roots and freshslices were observed using an inverted confocal microscope(model LSM510 META; Zeiss). As a consequence of thepresence of tissue autofluorescence, GFP fluorescence specificitywas monitored by spectral fingerprinting, using the Lambdamode tool. Standard GFP fluorescence was captured upon488-nm excitation using a 500–530-nm emission band passfilter. All observations were performed in three independentexperiments.
Results
Protocol set-up for whole-mount clearing and PI staining ofroots and galls
The intrinsic thickness and opacity of plant tissues, which canproduce inadequate staining and imaging, limit visualization ofnuclear morphology in whole galls induced by nematodes. Gallsinduced by root-knot nematodes in Arabidopsis roots can reachup to 500 lm (or more) in diameter, creating a challenge for theuse of WM samples for nuclear imaging.
The principal changes to the original protocols (Bauwenset al., 1994; Willemse et al., 2008) are described hereafter.Whole roots of uninfected 4-wk-old Arabidopsis roots and gallsat different developmental stages were tested. In the first series ofexperiments, we performed the clearing process by leaving thesamples in ethanol at )20�C for a minimum of 2 d for thinnerroot samples, and a maximum of 4 d for thicker galls. Differentconcentrations of formamide were applied during the differentwashing steps, and 50% formamide in PBS (pH 7.2) gave thebest clearing quality of root tissues. Formamide, also known asmethanamide, can be used as a solvent to facilitatedouble-stranded DNA denaturation upon heating. In order toachieve good preservation and to avoid damage to the root mate-rial, a series of alternate short steps of heating and cooling in50% formamide were tested. Depending on the root thickness,two to four series of 2–3 min at 100�C were applied and thenthe samples were immediately placed on ice for 5 min. Thisprocedure was found to be best for tissue softening, transparencyand nuclear staining. Staining of nuclei was performed by adding0.5 lg ml)1 of PI in 90% glycerol and roots were mounted ontoglass slides.
Slides were kept for several days in this staining mixture at4�C. Repeated DNA heat denaturation, in the presence offormamide, followed by fast quenching, resulted in the prefer-ential staining of nuclei along the entire root and gall tissue. Asseen in Supporting Information Fig. S1, this specificity was lostwhen the denaturation ⁄ quenching step was omitted from theprotocol. In Fig. S1(d) nuclei are more clearly visible because ofthe lack of background autofluorescence in cells filled with alarge vacuole, such as epidermal and cortical root cells. By con-trast, cells with a dense cytoplasm, such as root meristematiccells and giant cells, are more prone to PI-induced cytoplasmicbackground fluorescence. We observed that a long incubationin 90% glycerol leads to progressively more translucent roots orgall tissues.
Observation of galls induced by Meloidogyne incognita
Using WM preparations of infected roots, we were able to fol-low the migratory path of the second-stage nematode juveniles(J2s) within Arabidopsis roots. Nuclei of both roots and migrat-ing nematodes were labelled with PI, whereas a weak PIcounterstaining of the cell wall provided information on rootcell morphology (Fig. 1a). During NFS induction, the firstnuclear divisions occurring in giant cells were detected
490 Research
NewPhytologist
� 2012 INRA
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 195: 488–496
www.newphytologist.com
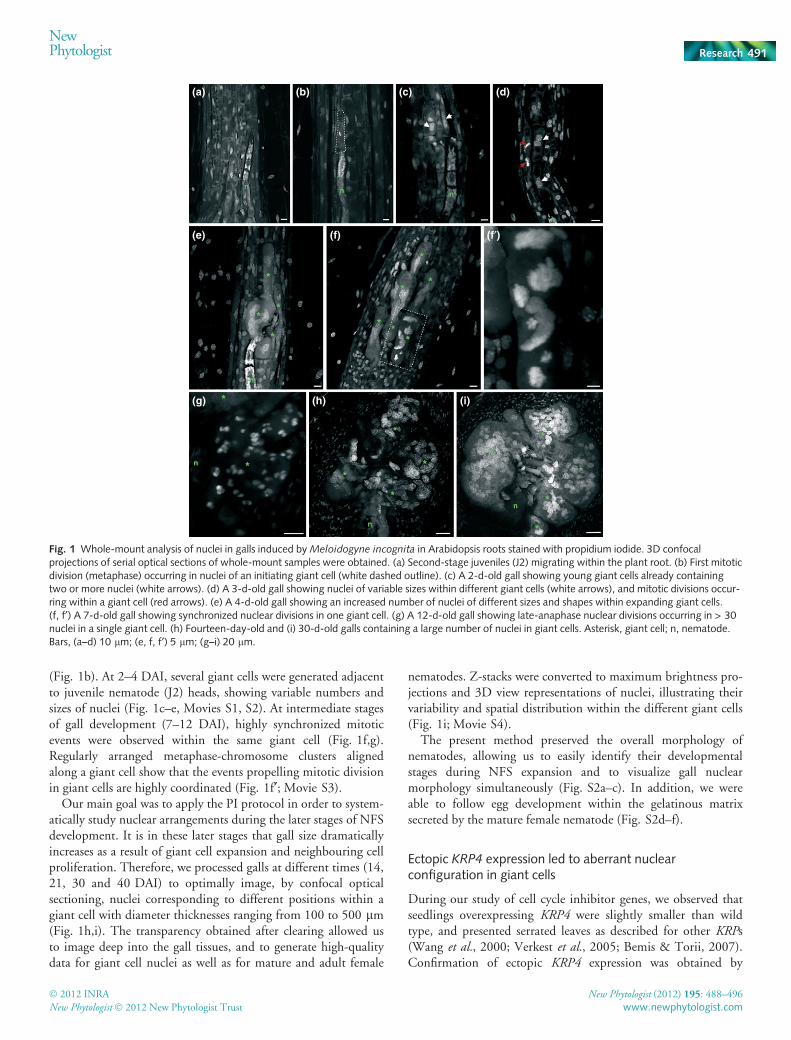
(Fig. 1b). At 2–4 DAI, several giant cells were generated adjacentto juvenile nematode (J2) heads, showing variable numbers andsizes of nuclei (Fig. 1c–e, Movies S1, S2). At intermediate stagesof gall development (7–12 DAI), highly synchronized mitoticevents were observed within the same giant cell (Fig. 1f,g).Regularly arranged metaphase-chromosome clusters alignedalong a giant cell show that the events propelling mitotic divisionin giant cells are highly coordinated (Fig. 1f¢; Movie S3).
Our main goal was to apply the PI protocol in order to system-atically study nuclear arrangements during the later stages of NFSdevelopment. It is in these later stages that gall size dramaticallyincreases as a result of giant cell expansion and neighbouring cellproliferation. Therefore, we processed galls at different times (14,21, 30 and 40 DAI) to optimally image, by confocal opticalsectioning, nuclei corresponding to different positions within agiant cell with diameter thicknesses ranging from 100 to 500 lm(Fig. 1h,i). The transparency obtained after clearing allowed usto image deep into the gall tissues, and to generate high-qualitydata for giant cell nuclei as well as for mature and adult female
nematodes. Z-stacks were converted to maximum brightness pro-jections and 3D view representations of nuclei, illustrating theirvariability and spatial distribution within the different giant cells(Fig. 1i; Movie S4).
The present method preserved the overall morphology ofnematodes, allowing us to easily identify their developmentalstages during NFS expansion and to visualize gall nuclearmorphology simultaneously (Fig. S2a–c). In addition, we wereable to follow egg development within the gelatinous matrixsecreted by the mature female nematode (Fig. S2d–f).
Ectopic KRP4 expression led to aberrant nuclearconfiguration in giant cells
During our study of cell cycle inhibitor genes, we observed thatseedlings overexpressing KRP4 were slightly smaller than wildtype, and presented serrated leaves as described for other KRPs(Wang et al., 2000; Verkest et al., 2005; Bemis & Torii, 2007).Confirmation of ectopic KRP4 expression was obtained by
(a) (b) (c) (d)
(e) (f) (f′)
(g) (h) (i)
Fig. 1 Whole-mount analysis of nuclei in galls induced by Meloidogyne incognita in Arabidopsis roots stained with propidium iodide. 3D confocalprojections of serial optical sections of whole-mount samples were obtained. (a) Second-stage juveniles (J2) migrating within the plant root. (b) First mitoticdivision (metaphase) occurring in nuclei of an initiating giant cell (white dashed outline). (c) A 2-d-old gall showing young giant cells already containingtwo or more nuclei (white arrows). (d) A 3-d-old gall showing nuclei of variable sizes within different giant cells (white arrows), and mitotic divisions occur-ring within a giant cell (red arrows). (e) A 4-d-old gall showing an increased number of nuclei of different sizes and shapes within expanding giant cells.(f, f¢) A 7-d-old gall showing synchronized nuclear divisions in one giant cell. (g) A 12-d-old gall showing late-anaphase nuclear divisions occurring in > 30nuclei in a single giant cell. (h) Fourteen-day-old and (i) 30-d-old galls containing a large number of nuclei in giant cells. Asterisk, giant cell; n, nematode.Bars, (a–d) 10 lm; (e, f, f¢) 5 lm; (g–i) 20 lm.
NewPhytologist Research 491
� 2012 INRA
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 195: 488–496
www.newphytologist.com
RT-PCR analysis (Fig. S3a) and by nuclear localization of theKRP4 protein (Fig. S3b,c). The effect of KRP4 overexpressionduring nematode infection was analysed in detail by microscopicanalysis of galls. The cellular morphology of both the KRP4OE
line and the wild type was analysed and followed by WM clearingand PI staining of KRP4OE galls in order to investigate the overallnuclear morphology. Ultimately, data for fixed cleared tissuewere validated by in vivo GFP-KRP4 analysis.
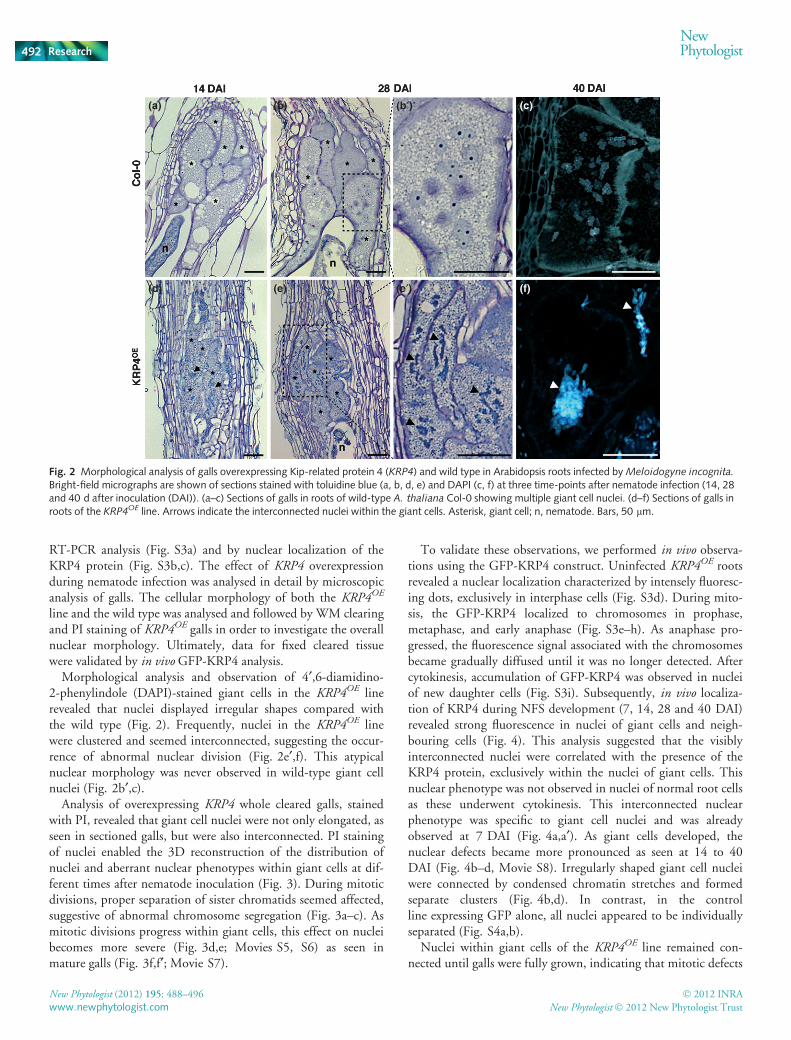
Morphological analysis and observation of 4¢,6-diamidino-2-phenylindole (DAPI)-stained giant cells in the KRP4OE linerevealed that nuclei displayed irregular shapes compared withthe wild type (Fig. 2). Frequently, nuclei in the KRP4OE linewere clustered and seemed interconnected, suggesting the occur-rence of abnormal nuclear division (Fig. 2e¢,f). This atypicalnuclear morphology was never observed in wild-type giant cellnuclei (Fig. 2b¢,c).
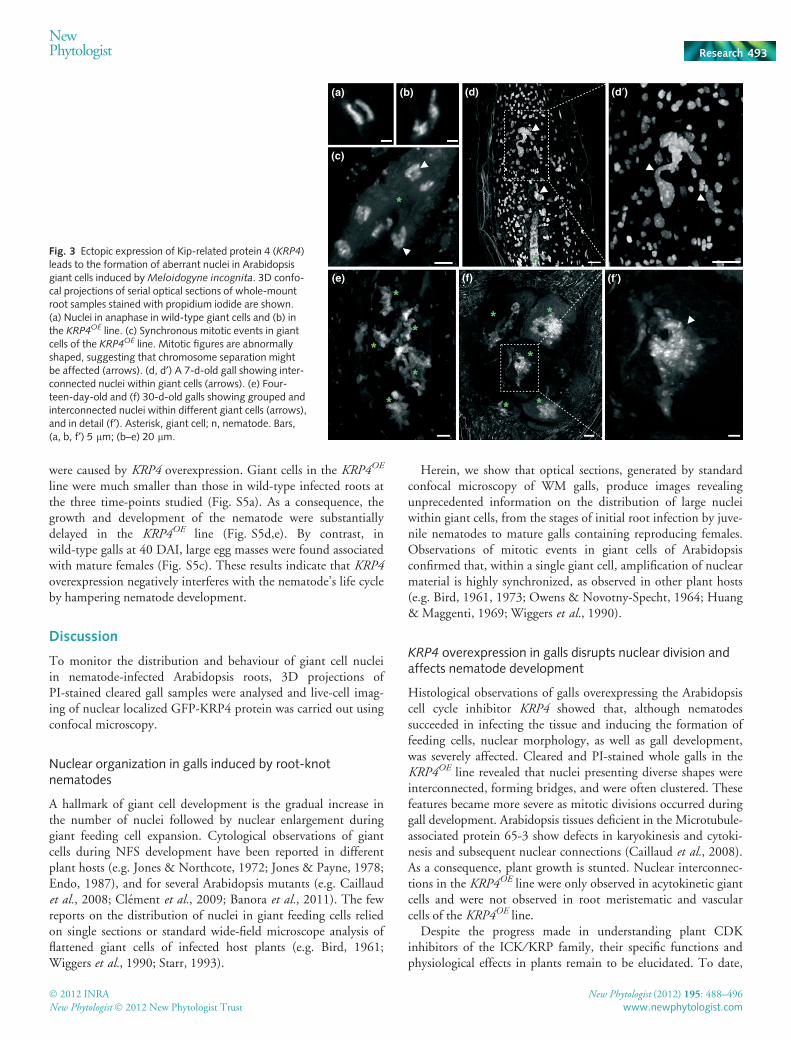
Analysis of overexpressing KRP4 whole cleared galls, stainedwith PI, revealed that giant cell nuclei were not only elongated, asseen in sectioned galls, but were also interconnected. PI stainingof nuclei enabled the 3D reconstruction of the distribution ofnuclei and aberrant nuclear phenotypes within giant cells at dif-ferent times after nematode inoculation (Fig. 3). During mitoticdivisions, proper separation of sister chromatids seemed affected,suggestive of abnormal chromosome segregation (Fig. 3a–c). Asmitotic divisions progress within giant cells, this effect on nucleibecomes more severe (Fig. 3d,e; Movies S5, S6) as seen inmature galls (Fig. 3f,f¢; Movie S7).
To validate these observations, we performed in vivo observa-tions using the GFP-KRP4 construct. Uninfected KRP4OE rootsrevealed a nuclear localization characterized by intensely fluoresc-ing dots, exclusively in interphase cells (Fig. S3d). During mito-sis, the GFP-KRP4 localized to chromosomes in prophase,metaphase, and early anaphase (Fig. S3e–h). As anaphase pro-gressed, the fluorescence signal associated with the chromosomesbecame gradually diffused until it was no longer detected. Aftercytokinesis, accumulation of GFP-KRP4 was observed in nucleiof new daughter cells (Fig. S3i). Subsequently, in vivo localiza-tion of KRP4 during NFS development (7, 14, 28 and 40 DAI)revealed strong fluorescence in nuclei of giant cells and neigh-bouring cells (Fig. 4). This analysis suggested that the visiblyinterconnected nuclei were correlated with the presence of theKRP4 protein, exclusively within the nuclei of giant cells. Thisnuclear phenotype was not observed in nuclei of normal root cellsas these underwent cytokinesis. This interconnected nuclearphenotype was specific to giant cell nuclei and was alreadyobserved at 7 DAI (Fig. 4a,a¢). As giant cells developed, thenuclear defects became more pronounced as seen at 14 to 40DAI (Fig. 4b–d, Movie S8). Irregularly shaped giant cell nucleiwere connected by condensed chromatin stretches and formedseparate clusters (Fig. 4b,d). In contrast, in the controlline expressing GFP alone, all nuclei appeared to be individuallyseparated (Fig. S4a,b).
Nuclei within giant cells of the KRP4OE line remained con-nected until galls were fully grown, indicating that mitotic defects
(a) (b) (b′) (c)
(d) (e) (e′) (f)
Fig. 2 Morphological analysis of galls overexpressing Kip-related protein 4 (KRP4) and wild type in Arabidopsis roots infected by Meloidogyne incognita.
Bright-field micrographs are shown of sections stained with toluidine blue (a, b, d, e) and DAPI (c, f) at three time-points after nematode infection (14, 28and 40 d after inoculation (DAI)). (a–c) Sections of galls in roots of wild-type A. thaliana Col-0 showing multiple giant cell nuclei. (d–f) Sections of galls inroots of the KRP4OE line. Arrows indicate the interconnected nuclei within the giant cells. Asterisk, giant cell; n, nematode. Bars, 50 lm.
492 Research
NewPhytologist
� 2012 INRA
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 195: 488–496
www.newphytologist.com
were caused by KRP4 overexpression. Giant cells in the KRP4OE
line were much smaller than those in wild-type infected roots atthe three time-points studied (Fig. S5a). As a consequence, thegrowth and development of the nematode were substantiallydelayed in the KRP4OE line (Fig. S5d,e). By contrast, inwild-type galls at 40 DAI, large egg masses were found associatedwith mature females (Fig. S5c). These results indicate that KRP4overexpression negatively interferes with the nematode’s life cycleby hampering nematode development.
Discussion
To monitor the distribution and behaviour of giant cell nucleiin nematode-infected Arabidopsis roots, 3D projections ofPI-stained cleared gall samples were analysed and live-cell imag-ing of nuclear localized GFP-KRP4 protein was carried out usingconfocal microscopy.
Nuclear organization in galls induced by root-knotnematodes
A hallmark of giant cell development is the gradual increase inthe number of nuclei followed by nuclear enlargement duringgiant feeding cell expansion. Cytological observations of giantcells during NFS development have been reported in differentplant hosts (e.g. Jones & Northcote, 1972; Jones & Payne, 1978;Endo, 1987), and for several Arabidopsis mutants (e.g. Caillaudet al., 2008; Clement et al., 2009; Banora et al., 2011). The fewreports on the distribution of nuclei in giant feeding cells reliedon single sections or standard wide-field microscope analysis offlattened giant cells of infected host plants (e.g. Bird, 1961;Wiggers et al., 1990; Starr, 1993).
Herein, we show that optical sections, generated by standardconfocal microscopy of WM galls, produce images revealingunprecedented information on the distribution of large nucleiwithin giant cells, from the stages of initial root infection by juve-nile nematodes to mature galls containing reproducing females.Observations of mitotic events in giant cells of Arabidopsisconfirmed that, within a single giant cell, amplification of nuclearmaterial is highly synchronized, as observed in other plant hosts(e.g. Bird, 1961, 1973; Owens & Novotny-Specht, 1964; Huang& Maggenti, 1969; Wiggers et al., 1990).
KRP4 overexpression in galls disrupts nuclear division andaffects nematode development
Histological observations of galls overexpressing the Arabidopsiscell cycle inhibitor KRP4 showed that, although nematodessucceeded in infecting the tissue and inducing the formation offeeding cells, nuclear morphology, as well as gall development,was severely affected. Cleared and PI-stained whole galls in theKRP4OE line revealed that nuclei presenting diverse shapes wereinterconnected, forming bridges, and were often clustered. Thesefeatures became more severe as mitotic divisions occurred duringgall development. Arabidopsis tissues deficient in the Microtubule-associated protein 65-3 show defects in karyokinesis and cytoki-nesis and subsequent nuclear connections (Caillaud et al., 2008).As a consequence, plant growth is stunted. Nuclear interconnec-tions in the KRP4OE line were only observed in acytokinetic giantcells and were not observed in root meristematic and vascularcells of the KRP4OE line.
Despite the progress made in understanding plant CDKinhibitors of the ICK ⁄ KRP family, their specific functions andphysiological effects in plants remain to be elucidated. To date,
(a) (b)
(c)
(d) (d′)
(e) (f) (f′)
Fig. 3 Ectopic expression of Kip-related protein 4 (KRP4)leads to the formation of aberrant nuclei in Arabidopsisgiant cells induced by Meloidogyne incognita. 3D confo-cal projections of serial optical sections of whole-mountroot samples stained with propidium iodide are shown.(a) Nuclei in anaphase in wild-type giant cells and (b) inthe KRP4OE line. (c) Synchronous mitotic events in giantcells of the KRP4OE line. Mitotic figures are abnormallyshaped, suggesting that chromosome separation mightbe affected (arrows). (d, d¢) A 7-d-old gall showing inter-connected nuclei within giant cells (arrows). (e) Four-teen-day-old and (f) 30-d-old galls showing grouped andinterconnected nuclei within different giant cells (arrows),and in detail (f¢). Asterisk, giant cell; n, nematode. Bars,(a, b, f¢) 5 lm; (b–e) 20 lm.
NewPhytologist Research 493
� 2012 INRA
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 195: 488–496
www.newphytologist.com

detailed functional analyses of the KRP1 and KRP2 Arabidopsisgenes have been carried out (Wang et al., 2000; De Veylderet al., 2001; Schnittger et al., 2003; Zhou et al., 2003; Verkestet al., 2005; Weinl et al., 2005; Jakoby et al., 2006; Roederet al., 2010; Sanz et al., 2011), whereas for the other ICK ⁄ KRPfamily members little is known (Bemis & Torii, 2007; Kimet al., 2008; Liu et al., 2008; Anzola et al., 2010). Strong overex-pression of KRPs (KRP1, KRP2, KRP6 and KRP7) in Arabidopsisseedlings affects cell proliferation and consequently plant growth.
However, different Arabidopsis ICK ⁄ KRP members may havedistinct roles, as their expression patterns vary within differentplant tissues (de Almeida Engler et al., 2009 inhttp://www.psb.ugent.be/ishi/) and also during the cell cycle(Menges et al., 2005). KRP4 transcripts seem to be preferentiallyexpressed in mitotically dividing cells such as root and shoot mer-istems (Ormenese et al., 2004; Marjanac et al., 2009). KRP4promoter activity or transcripts are not detected in vascular roottissue of Arabidopsis (http://www.psb.ugent.be/ishi/, SampleID:152103) or in NFSs induced within the vascular cylinder (P.Vieira et al. and de Almeida et al., unpublished data). At anyevent, KRP4 overexpression in NFSs revealed that giant cellnuclei were unable to divide properly. Multiple defects in giantcell nuclei, such as irregular shape, size and interconnections,were observed during our microscopic observations. In wild-typegiant cells, nuclei tend to group, appearing to be interconnected,in sectioned galls. Here our PI analyses showed that nuclei wereunconnected within mature giant cells of Arabidopsis. However,we cannot exclude the possibility that occasionally abnormal divi-sion of genetic material might occur (Wiggers et al., 1990),leading to the formation of aberrant nuclei. The occurrence ofcumulative defects in nuclei within a single giant cell overexpress-ing KRP4 suggests the occurrence of abnormal chromosomesegregation leading to an abnormal nuclear phenotype. There-fore, ectopic KRP4 expression might cause a mitotic instability,possibly hampering the proper separation of sister chromatids.KRP4 degradation, during sister chromatid separation, appearsto be a prerequisite for normal mitotic cell division (Boruc et al.,2010 and our data). Therefore, we propose that the continuousmisexpression of KRP4 in giant cells will drive a cumulativenegative effect on mitotic events, resulting in clustered and inter-connected nuclei.
Despite the effects of KRP4 overexpression on mitotic eventsin giant cells, resulting in an aberrant nuclear morphology, thesefeeding cells could be induced and maintained during nematodeparasitism. The competence of the parasitic nematode to alternormal cellular processes might allow mitosis to progress evenunder abnormal conditions. Nevertheless, under ectopic KRP4expression, giant cells often contained little cytoplasm, andexpansion was hampered. Giant cells constitute the main sourceof nutrient supply for nematode development and most femalesassociated with these galls were small and did not produce eggs.
The precise functions of KRP4 in plant cells are still unknown.Co-localization of KRP4 with chromosomes during mitosis innormal cells suggests its regulatory role during cell division: forexample, mediating the association of CDKA;1 with chromosomesduring mitosis (Boruc et al., 2010). Also, Boruc et al. (2010)suggested that KRP4 binds to the CDK ⁄ cyclin ⁄ CKS (CDKsubunit) complex to modulate its activity in a concentra-tion-dependent manner.
Concluding remarks
Despite numerous studies on plant–nematode interactions,morphological and cellular studies have relied mainly on thestudy of single histological sections of infected roots. Herein,
(a)
(b) (b′)
(c) (c′)
(d) (d′)
Fig. 4 In vivo nuclear localization of GFP-KRP4 in Arabidopsis gallsinduced by Meloidogyne incognita. These images are 3D confocalprojections. Left panel, GFP fluorescent images; right panel, overlays ofGFP fluorescence with differential interference contrast. (a, a¢) A 7-d-oldgall showing abnormal nuclear division (red arrows) and interconnectednuclei within giant cells (white arrows). (b, b¢) A 14-d-old gall showinginterconnected nuclei within giant cells (arrows). (c, c¢) A 30-d-old gallshowing grouped and interconnected nuclei within giant cells (arrows).(d, d¢) Isolated nuclei of 40-d-old giant cells showing bridges ofinterconnected nuclei. Asterisk, giant cell; n, nematode. Bars, (a–c) 20 lm;(d) 5 lm.
494 Research
NewPhytologist
� 2012 INRA
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 195: 488–496
www.newphytologist.com

we present a simple procedure that allows the 3D imaging ofnuclei in giant feeding cells induced by parasitic root-knot nem-atodes. This methodology provides new insights into the organi-zation and morphology of complex organelles, such as nuclei, inlarge biological samples, such as nematode-induced galls. Thisapproach is complementary to histological and in vivo studies.The WM procedure applied here also allowed us to easily iden-tify the distinct nematode life cycle stages associated with eachgall. The identification of the precise developmental stage ofnematodes is difficult and often impossible when solely based onstandard tissue sectioning. Our method allowed us to study thenuclear structure of a cell cycle mutant compared with the wildtype and to follow the parasite stage simultaneously. Our dataprovide insights into KRP4 function and validate functional pre-dictions for this cell cycle inhibitor. We present further evidencethat KRP4 is probably involved in chromatid separation duringchromosome segregation (Boruc et al., 2010), also illustratingthat the WM method proposed, combined with other studies,can help to reveal gene function.
Several strategies are currently used to target nematodeparasitism genes as well as the genes involved in nematodemetabolism, via RNAi constructs, to inhibit gall formation ornematode development (e.g. Bakhetia et al., 2005; Huang et al.,2006). The WM method presented here has the potential to bea very useful tool for readily acquiring insights into the cellularand nuclear changes occurring in complex multicellular biologi-cal specimens, such as nematode-induced galls.
Acknowledgements
We especially thank Eugenia Russinova for providing the KRP4OE
line. We also thank Vanesa Sanchez for help during our initialexperiments, and Nathalie Marteu for the production of biologicalmaterial and Erin Matticola for language correction. P.V. was sup-ported by a doctoral scholarship from Fundacao para a Ciencia epara a Tecnologia, Portugal (SFHR\BD\41339\2007).
References
de Almeida Engler J, De Veylder L, De Groodt R, Rombouts S, Bouldouf V, De
Meyer B, Hemerly A, Ferreira P, Beeckman T, Karimi M et al. 2009.
Systematic analysis of cell cycle gene expression during Arabidopsisdevelopment. Plant Journal 59: 645–660.
de Almeida Engler J, De Vleesschauwer V, Burssens S, Celenza JLJ, Inze D, Van
Montagu M, Engler G, Gheysen G. 1999. Molecular markers and cell cycle
inhibitors show the importance of cell cycle progression in nematode-induced
galls and syncytia. The Plant Cell 11: 793–808.
de Almeida Engler J, Engler G, Gheysen G. 2011. Unravelling the plant cell
cycle in nematode induced feeding sites. In: Jones J, Gheysen G, Fenoll C, eds.
Genomics and molecular genetics of plant–nematode interactions. Dordrecht, the
Netherlands: Springer Science+Business Media, 349–368.
Anzola JM, Sieberer T, Ortbauer M, Butt H, Korbei B, Weinofer I, Mullner
AE, Lusching C. 2010. Putative Arabidopsis transcriptional adaptor protein
(PROPORZ1) is required to modulate histone acetylation in response to auxin.
Proceedings of the National Academy of Sciences, USA 107: 10308–10313.
Bakhetia M, Charlton W, Atkinson HJ, McPherson MJ. 2005. RNA
interference of dual oxidase in the plant nematode Meloidogyne incognita.
Molecular Plant–Microbe Interactions 18: 1099–1106.
Banora MY, Rodiuc N, Baldacci-Cresp F, Smertenko A, Bleve-Zacheo T,
Mellilo MT, Karimi M, Hilson P, Evrard J-L, Favery B et al. 2011. Feeding
cells induced by phytoparasitic nematodes require c-tubulin ring complex for
microtubule reorganization. PLoS Pathogens 12: e1002343.
Bauwens S, Katsanis K, Van Montagu M, Van Oostveldt P, Engler G. 1994.
Procedure for whole mount fluorescence in situ hybridization of interphase
nuclei on Arabidopsis thaliana. Plant Journal 6: 123–131.
Bemis SM, Torii KU. 2007. Autonomy of cell proliferation and developmental
programs during Arabidopsis aboveground organ morphogenesis.
Developmental Biology 304: 367–381.
Bird AF. 1961. The ultrastructure and histochemistry of a nematode-induced
giant cell. The Journal of Biophysical and Biochemical Cytology 11: 701–715.
Bird AF. 1973. Observations on chromosomes and nucleoli in syncytia induced
by Meloidogyne javanica. Physiological and Molecular Plant Pathology 3:
387–391.
Boisnard-Lorig C, Colon-Carmona A, Bauch M, Hodge S, Doerner P,
Bancharel E, Dumas C, Haseloff J, Berger F. 2001. Dynamic analyses of
the expression of the HISTONE::YFP fusion protein in Arabidopsis show
that syncytial endosperm is divided in mitotic domains. Plant Cell 13:
495–509.
Boruc J, Mylle E, Duda M, De Clercq R, Rombauts S, Geelen D, Hilson P, Inze
D, Van Damme D, Russinova E. 2010. Systematic localization of the
Arabidopsis core cell cycle proteins reveals novel cell division complexes. PlantPhysiology 152: 553–565.
Caillaud M-C, Lecomte P, Jammes F, Quentin M, Pagnotta S, Andrio E, de
Almeida Engler J, Marfaing N, Gounon P, Abad P et al. 2008. MAP65-3
microtubule-associated protein is essential for nematode-induced giant cell
ontogenesis in Arabidopsis. The Plant Cell 20: 423–437.
Clement M, Ketelaar T, Rodiuc N, Banora MY, Smertenko A, Engler G, Abad
P, Hussey PJ, de Almeida Engler A. 2009. Actin-depolymerizing
factor2-mediated actin dynamics are essential for root-knot nematode infection
of Arabidopsis. The Plant Cell 21: 2963–2979.
Davis EL, Hussey RS, Baum TJ, Bakker J, Schots A, Rosso M-N, Abad P. 2000.
Nematode parasitism genes. Annual Review of Phytopathology 38: 365–396.
De Veylder L, Beeckman T, Beemster GT, Krois L, Terras F, Landrieu I, Van
Der Schueren E, Maes S, Naudts M, Inze D. 2001. Functional analysis of
cyclin-dependent kinase inhibitors of Arabidopsis. The Plant Cell 13:
1653–1667.
Endo BY. 1987. Histopathology and ultrastructure of crops invaded by
certain sedentary endoparasitic nematodes. In: Veech JA, Dickson DW, eds.
Vistas on nematology. A commemoration of the twenty-fifth anniversary of thesociety of nematologists. Hyattsville, MD, USA: Society of Nematologists,
196–210.
Huang G, Allen R, Davis EL, Baum TJ, Hussey RS. 2006. Engineering broad
root-knot resistance in transgenic plants by RNAi silencing of a conserved and
essential root-knot nematode parasitism gene. Proceedings of the NationalAcademy of Sciences, USA 103: 14302–14306.
Huang CS, Maggenti AR. 1969. Mitotic aberrations and nuclear changes of
developing giant cells in Vicia faba caused by root knot nematode Meloidogynejavanica. Phytopathology 59: 447–455.
Inze D, De Veylder L. 2006. Cell cycle regulation in plant development. AnnualReview of Genetics 40: 77–105.
Jakoby MJ, Weinl C, Pusch S, Kuijt SJ, Merkle T, Dissmeyer N, Schnittger A.
2006. Analysis of the subcellular localization, function, and proteolytic control
of the Arabidopsis cyclin-dependent kinase inhibitor ICK1 ⁄ KRP1. PlantPhysiology 141: 1293–12305.
Jones MGK, Northcote DH. 1972. Multinucleate transfer cells induced in coleus
roots by the root-knot nematode, Meloidogyne arenaria. Protoplasma 75:
381–395.
Jones MGK, Payne HL. 1978. Early stages of nematode-induced giant-cell
formation in roots of Impatiens balsamina. Journal of Nematology 10: 70–84.
Kim HJ, Oh SA, Brownfield L, Hong SH, Ryu H, Hwang I, Twell D, Nam
HG. 2008. Control of plant germline proliferation by SCFFBL17 degradation of
cell cycle inhibitors. Nature 455: 1134–1137.
Liu J, Zhang Y, Qin G, Tsuge T, Sakaguchi N, Luo G, Sun K, Shi D, Aki S,
Zheng N et al. 2008. Targeted degradation of the cyclin-dependent kinase
inhibitor ICK4 ⁄ KRP6 by RING-type E3 ligases is essential for mitotic cell
NewPhytologist Research 495
� 2012 INRA
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 195: 488–496
www.newphytologist.com

cycle progression during Arabidopsis gametogenesis. The Plant Cell 20:
1538–1554.
Marjanac G, Karimi M, Naudts M, Beeckman T, Depicker A, De Buck S. 2009.
Gene silencing induced by hairpin or inverted repeated sense transgenes varies
among promoters and cell types. New Phytologist 184: 851–864.
Menges M, de Jager SM, Gruissem W, Murray JAH. 2005. Global analysis of
the core cell cycle regulators of Arabidopsis identifies novel genes, reveals
multiple and highly specific profiles of expression and provides a coherent
model for plant cell cycle control. Plant Journal 41: 546–566.
Niebel A, de Almeida Engler J, Hemerly A, Ferreira P, Van Montagu M,
Gheysen G. 1996. Induction of cdc2a and cyc1At expression in Arabidopsisduring early phases of nematode-induced feeding cell formation. Plant Journal10: 1037–1043.
Ormenese S, de Almeida Engler J, de Groot R, de Veylder L, Inze D, Jacqmard A.
2004. Analysis of the spatial expression pattern of seven Kip related proteins
(KRPs) in the shoot apex of Arabidopsis thaliana. Annals of Botany 93: 575–580.
Owens RG, Novotny-Specht H. 1964. Root-knot histogenesis. Contribibutionsfrom Boyce Thompson Institute 22: 39–50.
Roeder AHK, Chickarmane V, Cunha A, Obara B, Manjunath BS, Meyerowitz
EM. 2010. Variability in the control of cell division underlies sepal epidermal
patterning in Arabidopsis thaliana. PLoS Biology 8: e1000367.
Sanz L, Dewitte W, Forzani C, Patell F, Nieuwland J, Wen B, Quelhas P, De
Jager S, Titmus C, Campilho A et al. 2011. The Arabidopsis D-type cyclin
CYCD2;1 and the inhibitor ICK ⁄ KRP2 modulate auxin-induced lateral root
formation. The Plant Cell 23: 641–660.
Schnittger A, Weinl C, Bouer D, Schobinger U, Hulskamp M. 2003.
Misexpression of the cyclin-depedent kinase inhibitor ICK1 ⁄ KRP1 in
single-celled Arabidopsis trichomes reduces endoreduplication and cell size and
induces cell death. The Plant Cell 15: 303–315.
Sijmons PC, Atkinson HJ, Wyss U. 1994. Parasitic strategies of root nematodes
and associated host cell responses. Annual Review of Phytopathology 32:
235–259.
Starr JL. 1993. Dynamics of the nuclear complement of giant cells induced by
Meloidogyne incognita. Journal of Nematology 25: 416–421.
Tirichine L, Andrey P, Biot E, Maurin Y, Gaudin V. 2009. 3D fluorescent insitu hybridization using Arabidopsis leaf cryosections and isolated nuclei. PlantMethods 5: 1–7.
Truernit E, Haseloff J. 2008. A simple way to identify non-viable cells within
living plant tissue using confocal microscopy. Plant Methods 4: 1–6.
Verkest A, Manes CL, Vercruysse S, Maes S, Van Der Schueren E, Beeckman T,
Genschik P, Kuiper M, Inze D, De Veylder L. 2005. The cyclin-dependent
kinase inhibitor KRP2 controls the onset of the endoreduplication cycle during
Arabidopsis leaf development through inhibition of mitotic CDKA;1 kinase
complexes. The Plant Cell 17: 1723–1736.
Wang H, Fowke LC, Crosby WL. 1997. A plant cyclin-dependent kinase
inhibitor gene. Nature 386: 451–452.
Wang H, Zhou Y, Gilmer S, Whitwill S, Fowke LC. 2000. Expression of the
plant cyclin-depedent kinase inhibitor ICK1 affects cell division, plant growth
and morphology. Plant Journal 24: 613–623.
Weinl C, Marquardt S, Kuijt SJH, Nowack MK, Jakoby MJ, Hulskamp M,
Schnittger A. 2005. Novel functions of plant cyclin-dependent kinase
inhibitors, ICK1 ⁄ KRP1, can act non-cell autonomously and inhibit entry into
mitosis. The Plant Cell 17: 1704–1722.
Wiggers RJ, Starr JL, Price HJ. 1990. DNA content and variation in
chromosome number in plant cells affected by Meloidogyne incognita and
M. arenaria. Phytopathology 80: 1391–1395.
Willemse J, Kulikova O, de Jong H, Bisseling T. 2008. A new whole-mount
DNA quantification method and the analysis of nuclear DNA content in the
stem-cell niche of Arabidopsis roots. Plant Journal 55: 886–894.
Zhou Y, Fowke LC, Wang H. 2002. Plant CDK inhibitors: studies of
interactions with cell cycle regulators in the yeast two-hybrid system and
functional comparisons in transgenic Arabidopsis plants. Plant Cell Reports 20:
967–975.
Zhou Y, Li G, Brandizzi F, Fowke LC, Wang H. 2003. The plant
cyclin-dependent kinase inhibitor ICK1 has distinct functional domains for invivo kinase inhibition, protein instability and nuclear localization. Plant Journal35: 476–489.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Whole-mount analysis of uninfected 40-d-old Arabidop-sis roots stained with propidium iodide.
Fig. S2 Whole-mount analysis of nuclei stained with propidiumiodide in galls and nematodes during parasitism.
Fig. S3 Kip-related protein 4 (KRP4) expression and localizationin Arabidopsis roots.
Fig. S4 In vivo localization of free GFP in galls induced byMeloidogyne incognita.
Fig. S5 Ectopic Kip-related protein 4 (KRP4) expression delaysgiant cell expansion and nematode development.
Movie S1 3D confocal projections of serial optical sections of a3-d-old gall induced by Meloidogyne incognita in Arabidopsiswild-type roots.
Movie S2 Z-stack confocal optical sections of a 4-d-old gallinduced by Meloidogyne incognita in Arabidopsis wild-type roots.
Movie S3 Z-stack of confocal optical sections of a 7-d-old gall,showing arranged metaphase-chromosome clusters aligned alonga giant cell.
Movie S4 3D confocal projections of serial optical sections of a30-d-old gall induced by Meloidogyne incognita in Arabidopsiswild-type roots.
Movie S5 3D confocal projections of serial optical sections of a5-d-old gall induced by Meloidogyne incognita in the KRP4OE
line, showing abnormally connected nuclei within the giant cells.
Movie S6 3D confocal projections of serial optical sections of a7-d-old gall induced by Meloidogyne incognita in the KRP4OE
line, showing abnormally connected nuclei within the giant cells.
Movie S7 3D confocal projections of serial optical sections of a30-d-old gall induced by Meloidogyne incognita in the KRP4OE
line, showing abnormally connected nuclei within the giant cells.
Movie S8 In vivo localization of GFP-KRP4 in nuclei of a30-d-old gall induced by Meloidogyne incognita, showing groupedand interconnected nuclei within giant cells.
Please note: Wiley-Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
496 Research
NewPhytologist
� 2012 INRA
New Phytologist � 2012 New Phytologist Trust
New Phytologist (2012) 195: 488–496
www.newphytologist.com

















