
Volatile fatty acid cycling in organic-rich marine sediments
15
I Geochimico et Cosmochimlco Acta Vol. 46. pp. 157510 1589 0016·7037/82/091575-15$03.00/0 e Pergamon Pr.... Ltd. 1982. Printed in U.S.A. Volatile fatty acid cycling in organic-rich marine sediments FRANCIS J. SANSONE 1 • 2 • and CHRISTOPHER S. MARTENS 1 Marine Sciences Program, University of North Carolina at Chapel Hill, 12-5 Venable Hall 045A, Chapel Hill, NC 27514, U.S.A. 2 Institute of Marine Sciences, University of North Carolina at Chapel Hill, Morehead City, NC 28557, U.S.A. (Received November 4, 1981; accepted in revised form April 22, 1982) Abstract-Volatile fatty acid (VFA) apparent turnover rates were determined by measuring whole sediment VFA concentrations and the corresponding reaction rate constants. The following ranges of VFA concentrations were measured in Cape Lookout Bight, N.C. sediments (/lmole ·1,-1): acetate 54- 660, propionate 1-24, butyrate <0.5-22, iso-butyrate <0.5-6. Apparent turnover rates measured over a one-year period ranged from 18-600 /lmole·l,-I·h- I for acetate and 0.7-7 /lmole·I,-I·h- 1 for the carboxyl carbon of propionate. Methane production was observed only with acetate and only in sulfate- depleted sediments; total acetate turnover attained approximately the same maximum value in both sulfate-reducing and sulfate-depleted sediments. Apparent turnover rates for acetate and propionate appeared to be controlled by similar factors: in sulfate-reducing (surface) sediments the turnover rates were stimulated by autumn storm-mediated deposition/resuspension events; in deeper sulfate-depleted sediments the turnover rates followed changes in the ambient temperature. Changes in VFA poolsizes were proportionally much larger than changes in corresponding rate constants. The ratio of CO 2 to CH. produced from acetate vs. depth suggested that non-methanogenic bacteria accounted for 60% of the acetate turnover in sulfate-depleted sediments. VFA concentrations were much lower in N.C. continental slope mud than in Cape Lookout sediments; acetate was the only VFA detectable throughout the top 40 cm of the slope sediments. The estimated production rate of CO 2 from acetate decreased rapidly with depth. The surface rate was approximately 20 times less than that measured at similar temperatures in sulfate-reducing Cape Lookout sediments. INTRODUCTION Carolina (U.S.A.). In addition, the turnover rates of acetate were measured in anoxic continental slope DETRITUS decomposition reactions are important in sediments. global and local carbon budgets in that they recycle complex organic molecules originally fixed by auto- trophs into small molecules such as CO 2 and CH 4 • Biogeochemical zonation and organic Important sites for detritus decomposition include matter decomposition organic-rich marine sediments, where the availability of readily degraded organic matter can be viewed as Competition between decomposer microorganisms the "master variable" (Goldhaber and Kaplan, 1974, results in a depth-dependent zonation of biogeochem- 1975). The input of organic carbon provides the en- ical respiration processes in which inorganic electron ergy source for these heterotrophic systems and ul- acceptors are sequentially removed from the sedi- timately controls the rates and spatial distribution ment with depth. That is, oxygen is consumed at the of respiration and fermentation reactions during dia- top of the sediment column and when it is depleted genesis. These reactions control, and in turn are in- organic matter oxidation will continue using the next fluenced by, changes in pH (Berner et al., 1970; Ben- most efficient oxidant, and so on until either all the Yaakov, 1973), Eh (Thorstenson, 1970) and the con- inorganic electron acceptors are depleted or all the centra.tion of the organic and inorganic reactants and utilizable organic matter is oxidized (Claypool and products of organic matter decomposition (e.g., Kaplan, 1974; Mechalas, 1974). In addition to these Zehnder, 1978). anaerobic respiration processes that result in the deg- This research examined the role of volatile fatty radation of short-chain organics, fermentation pro- acids (VFAs), particularly acetate and propionate, cesses also occur simultaneously in these systems in sedimentary decomposition processes occurring in (Zehnder, 1978). Such fermentations by the so- sulfate-containing and sulfate-depleted zones of or- called "acid-forming bacteria" involve the use of the ganic-rich coastal sediments. VFA turnover rate organic end products themselves as the electron ac- measurements were compared with previous mea- ceptors of specific oxidations (Thauer et al., 1977). surements of sulfate reduction and methane and dis- The end products of fermentations can become sub- solved inorganic carbon (C0 2 ) production at our ex- strates for bacteria involved in inorganic anaerobic perimental site in Cape Lookout Bight, North respirations. Unlike aerobic organisms, which can decompose 'complex organic substrates directly to CO 2 independently of other organisms, individual • Present address: Department of Oceanography and Hawaii Institute of Geophysics, University of Hawaii at anaerobes can only catalyze specific oxidations; they Manoa, Honolulu, HI 96822, U.S.A. must rely on other species to carry out other steps 1575
Transcript of Volatile fatty acid cycling in organic-rich marine sediments

I
Geochimico et Cosmochimlco Acta Vol. 46. pp. 157510 1589 0016·7037/82/091575-15$03.00/0 e Pergamon Pr.... Ltd. 1982. Printed in U.S.A.
Volatile fatty acid cycling in organic-rich marine sediments
FRANCIS J. SANSONE1•2
• and CHRISTOPHER S. MARTENS 1
Marine Sciences Program, University of North Carolina at Chapel Hill, 12-5 Venable Hall 045A, Chapel Hill, NC 27514, U.S.A.
2 Institute of Marine Sciences, University of North Carolina at Chapel Hill, Morehead City, NC 28557, U.S.A.
(Received November 4, 1981; accepted in revised form April 22, 1982)
Abstract-Volatile fatty acid (VFA) apparent turnover rates were determined by measuring whole sediment VFA concentrations and the corresponding reaction rate constants. The following ranges of VFA concentrations were measured in Cape Lookout Bight, N.C. sediments (/lmole ·1,-1): acetate 54660, propionate 1-24, butyrate <0.5-22, iso-butyrate <0.5-6. Apparent turnover rates measured over a one-year period ranged from 18-600 /lmole·l,-I·h- I for acetate and 0.7-7 /lmole·I,-I·h- 1 for the carboxyl carbon of propionate. Methane production was observed only with acetate and only in sulfatedepleted sediments; total acetate turnover attained approximately the same maximum value in both sulfate-reducing and sulfate-depleted sediments.
Apparent turnover rates for acetate and propionate appeared to be controlled by similar factors: in sulfate-reducing (surface) sediments the turnover rates were stimulated by autumn storm-mediated deposition/resuspension events; in deeper sulfate-depleted sediments the turnover rates followed changes in the ambient temperature. Changes in VFA poolsizes were proportionally much larger than changes in corresponding rate constants. The ratio of CO2 to CH. produced from acetate vs. depth suggested that non-methanogenic bacteria accounted for 60% of the acetate turnover in sulfate-depleted sediments.
VFA concentrations were much lower in N.C. continental slope mud than in Cape Lookout sediments; acetate was the only VFA detectable throughout the top 40 cm of the slope sediments. The estimated production rate of CO2 from acetate decreased rapidly with depth. The surface rate was approximately 20 times less than that measured at similar temperatures in sulfate-reducing Cape Lookout sediments.
INTRODUCTION Carolina (U.S.A.). In addition, the turnover rates of acetate were measured in anoxic continental slope
DETRITUS decomposition reactions are important in sediments.
global and local carbon budgets in that they recycle complex organic molecules originally fixed by autotrophs into small molecules such as CO2 and CH4• Biogeochemical zonation and organic
Important sites for detritus decomposition include matter decomposition
organic-rich marine sediments, where the availability of readily degraded organic matter can be viewed as Competition between decomposer microorganisms the "master variable" (Goldhaber and Kaplan, 1974, results in a depth-dependent zonation of biogeochem1975). The input of organic carbon provides the en ical respiration processes in which inorganic electron ergy source for these heterotrophic systems and ul acceptors are sequentially removed from the seditimately controls the rates and spatial distribution ment with depth. That is, oxygen is consumed at the of respiration and fermentation reactions during dia top of the sediment column and when it is depleted genesis. These reactions control, and in turn are in organic matter oxidation will continue using the next fluenced by, changes in pH (Berner et al., 1970; Ben most efficient oxidant, and so on until either all the Yaakov, 1973), Eh (Thorstenson, 1970) and the con inorganic electron acceptors are depleted or all the centra.tion of the organic and inorganic reactants and utilizable organic matter is oxidized (Claypool and products of organic matter decomposition (e.g., Kaplan, 1974; Mechalas, 1974). In addition to these Zehnder, 1978). anaerobic respiration processes that result in the deg
This research examined the role of volatile fatty radation of short-chain organics, fermentation proacids (VFAs), particularly acetate and propionate, cesses also occur simultaneously in these systems in sedimentary decomposition processes occurring in (Zehnder, 1978). Such fermentations by the sosulfate-containing and sulfate-depleted zones of or called "acid-forming bacteria" involve the use of the ganic-rich coastal sediments. VFA turnover rate organic end products themselves as the electron acmeasurements were compared with previous mea ceptors of specific oxidations (Thauer et al., 1977). surements of sulfate reduction and methane and dis The end products of fermentations can become subsolved inorganic carbon (C02 ) production at our ex strates for bacteria involved in inorganic anaerobic perimental site in Cape Lookout Bight, North respirations. Unlike aerobic organisms, which can
decompose 'complex organic substrates directly to CO2 independently of other organisms, individual• Present address: Department of Oceanography and
Hawaii Institute of Geophysics, University of Hawaii at anaerobes can only catalyze specific oxidations; they Manoa, Honolulu, HI 96822, U.S.A. must rely on other species to carry out other steps
1575

1576 F. J. Sansone and C. S. Martens
in the decomposition of organic substrates to CO2
(Fenchel and J0rgensen, 1977; Zehnder, 1978). The sequence of successive oxidant depletion pre
ceding sulfate reduction can be seen in pelagic sediments because the low organic content of the accumulating sediment allows the zones of lowconcentration oxidants, such as NO), to be spread out over a relatively large vertical distance (Froelich et al., 1979). Typically, only zones of aerobic respiration, sulfate reduction, and methanogenesis are observed in organic-rich coastal sediments because of the relatively large concentrations of fermentable substrates available (Claypool and Kaplan, 1974; Martens, 1978).
The dominant source of detritus in shallow coastal environments is macrophyte litter from both marsh and sea grasses, especially along the middle and southeastern Atlantic coast of the United States (Tenore, 1977). This predominance is due to the high productivity of marine macrophytes, as well as their tendency toward generating detritus rather than entering decomposer food chains via herbivore grazing (Mann, 1972; Tenore, 1977). Water-soluble, labile organics are readily leached from leaf litter shortly after senescence. The residual material is composed primarily of recalcitrant cellulose and ligno-cellulose complexes (Mann, 1972; Fenchel and J0rgensen, 1977).
Volatile fatty acids, which are generally defined as aliphatic carboxylic acids with six or fewer carbon atoms per molecule, have been found to be intermediates in the anaerobic decomposition of model lignin degradation products (Healy et al., 1980) and of cellulose (Alexander, 1961). VFAs are known to be produced by members of the genus Clostridia during the anaerobic decomposition of cellulose in soils (Brock, 1979) and during the decomposition of complex organic substrates in anaerobic digestors (Toerien and Hattingh, 1969). VFAs thus formed by fermentations are substrates for sulfate-reducing bacteria (SRB) and methanogens that are responsible for the terminal steps of the anaerobic decomposition of organic matter to CO2 and CH4 • (For a review of SRB see Postgate (1979); methanogens have been reviewed by Mah et al. (1977), Zeikus (1977), Zehnder (1978), Balch et al. (1979).)
For a complex organic substrate to be completely oxidized to CO2 it must be at least partially respired; in anaerobic environments an electron acceptor such as sulfate or CO2 must be used. The bacteria responsible for these respirations (the so-called "terminal organisms"), however, utilize only a limited number of substrates and therefore must rely on fermentative microbes to provide them with suitable substrates (Hungate, 1966, 1975). The terminal organisms are thus able to undergo respiration and therefore remove reducing equivalents from the system, as well as lower the concentration of end products of the fermenters. The result of this mutualism
is the complete anaerobic oxidation of organic material and the resultant optimization of the energy yield for the entire anaerobic community (Hungate, 1966, 1975).
Our work at Cape Lookout to date has emphasized the dynamics of the terminal end-products of organic matter decomposition, CH4 and CO2, In this paper we present work on the cycling of key organic intermediates that are the precursors of these end-products. Because this research was conducted in separate biogeochemical environments dominated by sulfate reduction and methanogenesis, the results should be applicable to similar organic-rich marine sediments.
METHODS
Field sites
The primary field site was station A-I, located in Cape Lookout Bight, N.C., an organic-rich coastal basin of approximately 2 km2 area located 115 km SW of Cape Hatteras on the North Carolina Outer Banks (Martens, 1976; Martens and Klump, 1980; Martens et al., 1980). The water column in Cape Lookout Bight is well oxygenated (> 130 I'M 0,) (Bartlett, 1981) and low in methane all year «5 I'M) (Sansone and Martens, 1978; Martens and Klump, 1980); it is underlain by a fine-grained mud with an organic carbon content ranging from approximately 3% by weight at the surface to 2% at several meters depth (C. S. Martens, unpublished data). The fine-grained sediment and, presumably, much of its organic content is derived rrom the flushing of nearby back barrier island lagoons (Martens and Klump, 1980). The sediment porosity decreases from 0.94 to 0.87 cm' of pore water per cm' of whole wet sediment in the top 25 cm and remains approximately at the latter value to a depth of 50 cm (Klump, 1980).
Cape Lookout Bight has proven to be a good site for studying organic decomposition reactions. The sedimentation rate of approximately 8-12 cm' y-I (ChantoD, 1979) results in a large organic material burial rate and, consequently, high rates of decomposition. Macroinfaunal bioturbation is limited to only the upper 1-3 cm of the sediment during winter months, at which time the sediment is oxidized and beige to light brown in color (Bartlett, 1981). In addition, in situ fluxes of the terminal carbon end-products CO2 and CH4 across the sediment-water interface have been measured (Martens and Klump, 1980; Martens and Klump, unpublished data). It has recently been shown that the methane flux exhibits the same seasonal pattern as does the production of methane from acetate in underlying, sulfatedepleted sediments (Sansone and Martens, 1981 a).
For the purpose of comparing VFA cycling in organicrich nearshore (i.e., Cape Lookout Bight) and open ocean environments, samples were also taken from the North Carolina continental slope in 655 m of water. The sampling site was located approximately 150 km NNE of Cape Hatteras (36 0 20.3'N, 74 0 44.2'W; R/V Eastward station 36438). The sediment was an anoxic silt-clay with porosity decreasing from approximately 0.76 to 0.69 in the top 40 cm.
Apparent turnover rate measurements
The methodology used to measure apparent VFA turnover rates has been described in detail (Sansone and Martens, 1981 b). Briefly, both the whole sediment substrate concentrations and the corresponding reaction rate constants were measured periodically and these parameters multiplied to obtain apparent production rates:
1577 Sediment fatty acids
Q)
0 30
Sulfate (mM)
40
Apparent production rate (/lmole·I;'· h-')
= Whole sediment concentration (/lmole ·1;')
x Rate constant (h-'), (I)
where I, is the volume in liters of whole wet sediment. Use of the term "apparent" is based on our measurements
of whole sediment VFA concentrations (pool size). Our methodology allows for determination of total "extractable" pools of VFAs; however, these pools should be carefully distinguished from the "microbiologically available" pools. These latter pools may be less than the total because of partitioning of VFAs between sediment pore water (both free and complexed), adsorption sites on sediment particle surfaces, and microbial cells. The actual microbial production rates may therefore be less than the apparent production rates (see below).
In order to measure VFA concentrations, bulk wet sediment samples were treated with base, freeze-dried, extracted with methanol, derivatized with methanoJic-HCI to form methyl esters of the VFAs, and analyzed by packed column gas chromatography with hexanoic acid as an internal standard. The analytical column was packed with two materials: the first half contained SP-2100 (to separate the VFAs); the second half contained THEED (tetrahydroxyethylenediamine) to retard the methanol solvent. The detection limits for acetate, propionate, butyrate, and isobutyrate were 1.0,0.4, O. 2, and 0.2/lmole '1;', respectively, for 600-ml samples.
VFA turnover rate constants were measured by adding tracer quantities of 14C-Iabeled compounds to sediment s~~ples, incubating anaerobically at in situ temperatures, killing the samples, and then recovering the metabolized label (a modification of the method of Christian and Wiebe 1978). Pre-existing whole-sediment VFA concentration~ were increased less than 1.5% by label additions, and during the incubations less than 5% of the added label was allowed to be utilized before termination. It was thus assumed that the rates measured were not affected by changes in substrate or end-product concentrations. Activity was recovered as follows: (a) "C02 on phenethylamine-treated filter paper; (b) "CH. by the method of Zehnder et al. (1979); (c) labeled particulate (including cellular) material; (d) labeled water-soluble, but not ether-soluble, material; and (e) labeled ether-soluble material (i.e., non-utilized substrate). Typically, 85-90% of the added label was recovered at the end of each experiment. Abiotic controls, which were
10 ,.... E 0......,.
20 r. .... a.
killed before the addition of labeled substrate, were used to correct for non-biological phenomena. The activities recovered in fractions (a), (b), (c), and (d) were summed to calculate the "total" turnover rate constant (Sansone, 1980). Labeled compounds used were: 1,2-"C-sodium acetate, 53.5 mCi/mmol (New England Nuclear, Boston, MA); I-"C-sodium propionate, 56.7 mCi/mmol (lCN, Irvine, CA).
The turnover rate constants measured here are not true first order rate constants determined over a range of substrate concentrations. Rather they are a measure of the instantaneous turnover rate at the in situ concentration. These rate constant measurements assume that the experimental system achieves nearly instantaneous partitioning between added substrates and the endogenous (intracellular) pools of the compounds. Incomplete partitioning during the course of our 5 to 60 minute incubations would lead to measured rate constants lower than the actual in situ constants. This effect may be counteracted by the adsorption of these compounds by sediment solid phase. If a significant portion of the whole sediment VFA concentration is quickly bound to surfaces so as to limit their bioavailability then the poolsizes measured in our samples would overestimate the pools available to the microbes. Turnover rates calculated by using such Whole sediment concentrations would thus result in overestimates of the actual turnover rate. It is not known to what degree these two effects compensate for each other.
Sulfate
Dissolved sulfate was determined gravimetrically as BaSO. from 3-5 ml pore water samples obtained by centrifuging sediment samples. The BaSO. was filtered with 25-mm diameter 0.4 /lm pore size Nucleopore filters aDd then weighed. Analytical precision of replicate samples was ±0.2 mM (Iu) (Martens and Klump, 1980).
RESULTS
Temperature and sulfate
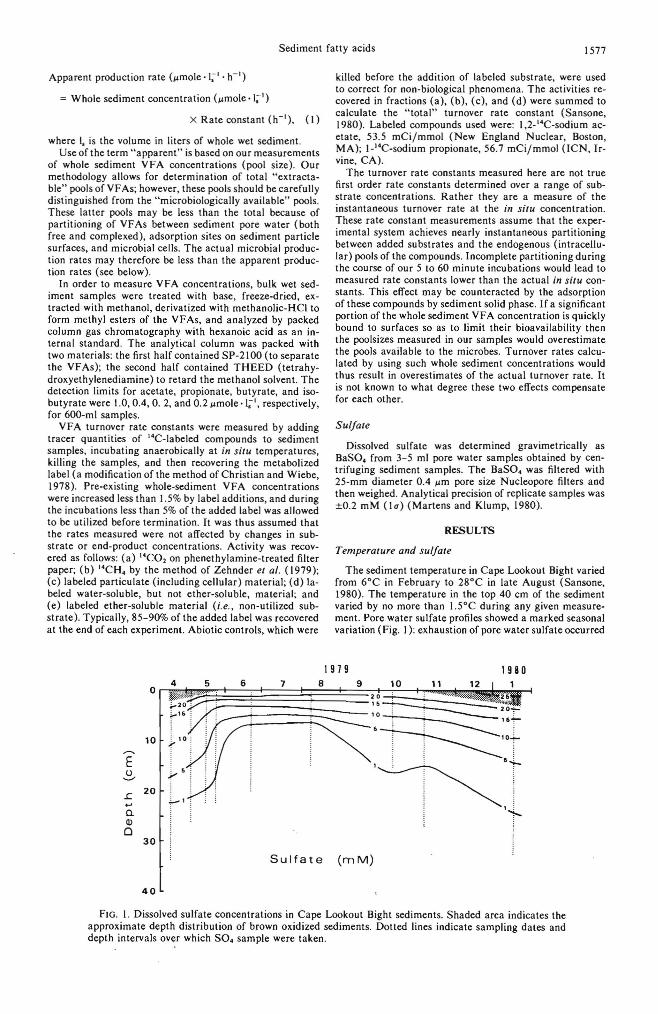
The sediment temperature in Cape Lookout Bight varied from 6°C in February to 28°C in late August (Sansone, 1980). The temperature in the top 40 cm of the sediment varied by no more than 1.5°C during any given measurement. Pore water sulfate profiles showed a marked seasonal variation (Fig. I): exhaustion of pore water sulfate occurred
FIG. I. Dissolved sulfate concentrations in Cape Lookout Bight sediments. Shaded area indicates the approximate depth distribution of brown oxidized sediments. Dotted lines indicate sampling dates and depth intervals over which SO. sample were taken.

00
c
V. -J
A 1979 1980 B 1979 1980
01 ' : I • " .~_ ~ '", 0'1 '." 0 ~ .,ooJ 3-' . I •
10
E .£ r. 20 ~
a. Gl o
30
40
I. E ~
.r: 20
a. III o
3.
f.
I. E ~
.r: 20 ~
a. " III o
'00 ,.
Acetate. (umole.I~')
f.
1979 1980
'0 '2
-'--, .~ /' ' .....
/~
, ~>'2.
'J' J (: /
'0 /
/
20~ )3.10 ,
"., /""
E3
~ .r: a. III o
!Butyrate
( 30
f. (umolel~)
, I. II " ,o~·,,·,·,
_~ ,_,' 10"""l. ~_,~J , 1 " ..........', \ \.:.....-,.' ~
./ "". ,"---------'0".....,.,· . ./;/ /
/
-'----~._-"
Propionate :-'1 ~(umole· I~') CIl
'" ~ '" '" " '" Co '" (')o 1979 1980
.0 II .2 ~
.....~,- ~ .,
_e_ _ _ ,3 '(::.;;.<'_ • '"\. \' , .-----:-' ' ,C '"
/ n I ,
/. , ':'-, -, ·so-Butyrate. •I • •~,I A • ~ __~// 0 ,.~ '(umQI,e'I~I~
'"
_/)
/~,' 1\ /
3
FIG. 2. Whole sediment VFA concentrations in Cape Lookout Bight sediments: (A) Acetate, (B) Propionate, (C) Butyrate, (D) iso-Butyrate. Dashed lines indicate the l-mM SOf isopleth.

1579 Sediment fatty acids
at a depth of approximately 30 cm during winter and approximately 10 cm during summer.
VFA concentrations
VFA concentration profiles were measured at station AI approximately every six weeks for a year; the results are shown as contour plots in Fig. 2. Acetate concentrations, by far the highest of the four VFAs measured, showed a pronounced seasonal and spatial pattern. The maximum concentration measured occurred in the summer at a depth corresponding to the level of sulfate depletion (i.e. sulfate concentration less than I mM). Propionate, butyrate, and isobutyrate concentrations showed different patterns.
Apparent VFA turnover rates
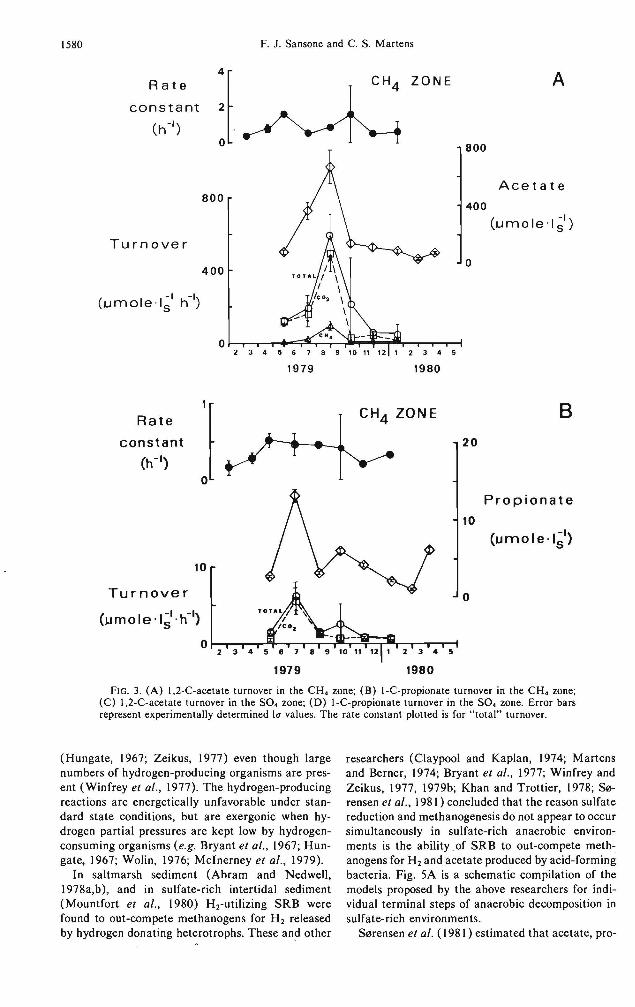
Turnover rate constants were measured for 1,2-"C-acetate and I-"C-propionate over the course of a year in two zones of station A-I sediments: the top 1-6 cm of the reduced portion of the sediment ("SO. zone"), and the top 5 cm of the sulfate-depleted « I mM SO.) portion of the sediments (''CH. zone"). The total rate constants measured (i.e., the sum of the CO2, CH., particulate material, and water soluble material production rate constants) are plotted in Fig. 3 along with the corresponding in situ substrate concentrations and the calculated turnover rates.
The turnover of 1,2-"C-acetate was measured at four depths during mid-summer in order to examine the depth distribution of CH. and CO2 production from acetate (Fig. 4). The upper two depths corresponded to the "SO. zone" and "CH. zone" studied in the previous experiments. CO2
production was the highest in the top depth interval; CH. production was very low in the top sample and was highest in the depth interval 7-12 cm below the point of sulfate depletion (depth = 15-20 cm).
The respiration index (RI) from 1,2-"C-sodium acetate is also shown in Fig. 4;
RI = "C02 Production/("C02 Production
+ "CH. Production). (2)
RI values ranged from 1.00 ± 0.04 between 0-5 cm in the sulfate reduction zone to 0.79 ± 0.07 at 30-35 cm depth in the methane production zone. RI values were invariant within experimental error from 8-35 cm implying that the proportion of the methyl group of acetate oxidized to CO2
did not vary significantly with depth within the methane production zone.
Continental slope VFA experiments
Volatile fatty acid concentrations measured in August 1979 in continental slope mud (Table 1) were much lower than in Cape Lookout Bight sediments. The low rate of anaerobic decomposition in these sediments, as indicated by the lack of sulfate depletion with depth, was reflected in the low VFA concentrations found; acetate was the only VFA detectable in all portions of the sediment examined. Below 6 cm, acetate decreased monotonically with depth; the other VFAs showed variable distribution with depth.
Rate constants for the production of CO2 from acetate in these sediments were measured in September 1980. These data were multiplied by the corresponding acetate concentrations measured in 1979 to obtain the apparent rates of CO2 production from acetate (Table 2). Rates and rate constants both decreased monotonically with depth. Continental slope apparent CO2 production rates were 20-3000 times slower than rates measured at similar temperatures in sulfate-containing Cape Lookout Bight sediments (Fig. 3C). Rate constants were most variable in the upper portion of the sediment, presumably because of the effects of bioturbation and irrigation by the abundant macroinfaunal
community dominated by polychetes (M. Watzin, pers. commun., 1980).
DISCUSSION
These data, which represent the first seasonal study of VFA cycling in organic-rich marine sediments, can be used to test important hypotheses concerning the role of specific groups of bacteria which mediate both decomposition processes and fluxes of small molecules such as acetate, CH4, and CO2 in nonbioturbated anoxic marine sediments.
The 6°-28°C temperature range observed in Cape Lookout Bight during 1979-1980 reflects the dynamic nature of the system. This temperature fluctuation seems to be the major factor affecting the activity of SRB controlling the distribution of dissolved sulfate in these sediments (Fig. I), as previously reported by Klump ( 1980) and Bartlett (1981 ).
The depth distribution of biogeochemical zones (i.e., oxidized, anaerobic sulfate-reducing, and anaerobic sulfate-depleted) varied seasonally (Fig. I). The deepest penetration of dissolved sulfate ranged from 8 cm in summer months to 30 cm in winter. Saturation methane concentrations, presumably delineating the top of the CH4 zone, did not occur at depths above the point of sulfate depletion (Martens and Klump, 1980). Because of the seasonal changes in the biogeochemical zonation of Cape Lookout sediments, changes would be expected in the concentrations of VFAs and their reaction rates as a function of sediment depth, respiration mode, and season. In addition to the effects of biogeochemical zonation of respiration modes, interactions between bacteria must be considered before depth-dependent differences in reaction mechanisms and rates can be interpreted. These interactions may take such forms as the competition for organic substrates or the mutualistic transfer of metabolic end products between species.
Biogeochemistry of terminal decomposition
Biogeochemists historically have viewed acidforming bacteria (AFB) (e.g., the genus Clostridium) as degraders of large organic molecules to smaller molecules such as VFAs. In addition to providing suitable organic substrates for other anaerobes, these fermentative bacteria are also able to transfer hydrogen derived from the oxidation of organic matter to other bacteria capable of anaerobic respirations such as sulfate or CO2 reduction (Hungate, 1966; Tezuka, 1966; Iannotti et al., 1973; Mountfort et al., 1980; McInerney et al., 1981). Hydrogen has also .been observed to be transferred from SRB to methanogens under conditions of low sulfate concentration (Bryant et al., 1977).
The above processes, both termed interspecies hydrogen transfer, lead to rapid consumption of hydrogen in organic-rich anaerobic environments. This leads to generally undetectable H 2 concentrations
1580 F. J. Sansone and C. S. Martens
CH 4 ZONE ARat e
constant
(h -I) :[ 800
Turnover
400
(umole'ls-I h- I)
0 2
Rate
'Iconstant
(h- I)
0
10
Turnover
( -I -I)JJmole'l ·hs
0 2 3 4
3 4
1979 1980
CH4 ZONE
800
Acetate
400
(u mol e . I ~I )
0
B 20
Propionate
10
(umole'I~I)
0
1979 1980
FIG. 3. (A) 1,2-C-acetate turnover in the CH. zone; (8) I-C-propionate turnover in the CH. zone; (C) 1,2-C-acetate turnover in the SO. zone; (D) l-C-propionate turnover in the SO. zone. Error bars represent experimentally determined [(1 values. The rate constant plotted is for "total" turnover.
(Hungate, 1967; Zeikus, 1977) even though large numbers of hydrogen-producing organisms are present (Winfrey et al., 1977). The hydrogen-producing reactions are energetically unfavorable under standard state conditions, but are exergonic when hydrogen partial pressures are kept low by hydrogenconsuming organisms (e.g. Bryant et al., 1967; Hungate, 1967; Wolin, 1976; McInerney et al., 1979).
In saltmarsh sediment (Abram and Nedwell, 1978a,b), and in sulfate-rich intertidal sediment (Mountfort et al., 1980) H 2-utilizing SRB were found to out-compete methanogens for H 2 released by hydrogen donating heterotrophs. These and other
researchers (Claypool and Kaplan, 1974; Martens and Berner, 1974; Bryant et al., 1977; Winfrey and Zeikus, 1977, 1979b; Khan and Trottier, 1978; S0rensen et al., 1981) concluded that the reason sulfate reduction and methanogenesis do not appear to occur simultaneously in sulfate-rich anaerobic environments is the ability. of SRB to out-compete methanogens for H 2 and acetate produced by acid-forming bacteria. Fig. 5A is a schematic compilation of the models proposed by the above researchers for individual terminal steps of anaerobic decomposition in sulfate-rich environments.
S0rensen et al. (1981) estimated that acetate, pro

1581
4
o
Sediment fatty acids
IIJ' / CO,
0234567
1979
FtG. 3. (Continued).
6
Rate
constant (h- I )
800
Turnover 400
(umole·I~I.h-I)
Rate
constant
(h -I)
Turnover
1979
pionate, butyrate, and H2 were used as electron donors for 40-65%, 5-20%, 5-20%, and 5-10% of the sulfate reduction, respectively, in coastal sediment slurries. These estimates were from studies on the accumulation of these substrates following the inhibition of SRB by molybdate; since these experiments did not account for concurrent inhibition of acetate and propionate production by SRB, the percentages may be underestimates for these two VFAs.
In intertidal sediments with low sulfate, Mountfort et al. (1980) found sulfate reduction rate potential (as measured by reduction of added 35S04) to be comparable to that measured in overlying sulfaterich sediments. This suggested that SRB were active
504 ZONE c
200 Acetate
(umole'I~I)
100
o
6 9 10 11 12 1 2 3 4 5
1980
504 ZOI\JE D
20
Propionate
10 -I(umole'l s )
o
6
1980
in the sulfate-depleted sediments, presumably supplying H2 and/or acetate to methanogens. Desulfovibrio have been co-cultured with methanogens in a sulfate-free medium that would not support either organism separately (Bryant et al., 1977; McInerney and Bryant, 1981). Fermentative (non-sulfate reducing) bacteria also have been shown to transfer hydrogen t9 methanogens (e. g. Bryant et al., 1967; Winfrey et al., 1977; Boone and Bryant, 1980) with the result that the fermentative bacteria produced 'products of a higher oxidation state than in the absence of the methanogens (Reddy et al., 1972). Jeris and McCarty (1965) found that, during active fermentation in a (low sulfate) anaerobic
30
2 3 4

1582 F. J. Sansone and C. S. Martens
o
10
E u
20
.t:
0. Ql
o 30
40 ~0-~--:I::-0-~-----::2'=-0-~----;;30
Sulfate (mM)
o
Martens (1981 a).
digestor, more than two-thirds of the starting organic carbon was converted to acetate. Fig. 5B is a schematic compilation of the relationships proposed by the above researchers for bacteria responsible for the terminal steps of degradation in sulfate-depleted environments, including the proposed transfer of reducing equivalents from AFB and SRB to methanogens.
In summary, interspecies hydrogen transfer results in (a) increased substrate utilization, (b) different proportions of reduced end products, (c) more ATP synthesized by the hydrogen-producing organisms, and (d) displacement of unfavorable reaction equilibria (Zeikus, 1977; Thauer et al., 1977).
Figure 5 will be used as a conceptual framework for interpreting the Cape Lookout Bight results. If this model is applicable to Bight sediments several processes can be predicted to occur in the two zones studied there. In the sulfate zone, acetate should be oxidized to CO2 with little or no CH4 being produced. In the CH4 zone, acetate should be disproportionated to CO2 and CH4 by methanogens and/or oxidized to CO2 by SRB.
Propionate would be expected to follow pathways similar to that of the hydroxy~acid lactate (Fig. 5). Both of these compounds are produced by acid-form-
Table 1. Volatile fatty acid and sulfate concentrations in box core from R/V Eastward station 36438 (360 20.3'N, 74° 44.2'WT.19Au9ust 1979. Sediment temperature was 5°C
Depth Vo1ati le Fatty Acid Cone. (umole·ls -1) S04 Interval
(em) Acetate Propionate 8utyrate iso-8utyrate (nt1)
0-6 61.4 1.7 <2.2 <.41 29.7 6-12 69.0 11.4 <3.3 3.4 29.0
12-18 46.1 3.1 3.1 0.86 28.5 20-26 35.8 3.0 <1.9 1.4 29.3 34-40 10.7 <1.2 5.4 0.86 28.7
Resp. Index
+ FIG. 4. Depth variation of apparent 14CH. production from 1,2-14C-acetate (broken line), dissolved
sulfate concentration (fl.), and 1,2-14C-acetate respiration index. Data are from 13-15 July 1980 (26.5°C). Vertical bars indicate the depth interval from which samples were taken. Horizontal bars indicate the standard deviation (lu) of triplicate acetate utilization experiments. From Sansone and
ing bacteria (Zehnder, 1978), and both are utilized by SRB (Postgate, 1979; Widdel, 1980) but not by methanogens (Zeikus, 1977). Propionate would thus be oxidized to CO2 in both zones. If acetate is released as an intermediate of this reaction in low-sulfate sediments, or if CO2 reduction is occurring, then methane may be formed as a result.
Rates of sulfate reduction, nutrient regeneration, and CO2 regeneration are highest at the sedimentwater interface and decrease exponentially with depth in Cape Lookout Bight sediments (Klump, 1980; Bartlett, 1981) as in other organic-rich environments (e.g. J0rgensen, 1977; Aller and Yingst, 1980). Seasonal variation in the input of labile organic materials might therefore be expected to be a major control in microbially mediated processes in the upper 10 cm. Below this depth, temperature and the availability of specific organic substrates and inorganic hydrogen acceptors would be expected to be the major factors controlling seasonal changes in the rates of decomposition.
In the following sections seasonal variations in VFA concentrations and turnover rates will be utilized to examine these hypotheses.
Table 2. Calculation of the apparent production rate of CO2from acetate in continental slope sediments. Values in parentheses are standard deviations (la). From Sansone and Martens (1981b)
Whole-Sediment Apparent Rate Depth
Interval (em)
CO2 from Acetate Rate Canst.
(h-1 )a
Acetate Cone.
(umolefls)b
of CO2 Production from Aceta te (umo1efl/h)
0-5 .3102 (.4878) 8-13 .0654 (.0162)
17-22 .0232 (.0090) 20-25 .0148 (.0078) 28-33 .0106 (.0024) 35-40 .0092 (.0124)
aOata from 11-13 September 1980 bOata from 19 AU9ust 1979.
60 (10.2) 18.62 (29.4l 67l11.4) 4. 38 ~ 1. 32 40 6.8) .928 .394) 33 ~5.6) .488 (.270) 19 3.23) .202 (.058l12 (2.04) .110 (.150

1583 Sediment fatty acids
A. Sulfate Present B. Sulfate Absent
cO:! CC7
FIG. 5. A model of the terminal steps of anaerobic decomposition in (A) sulfate-containing, and (B) sulfate-depleted environments. AFB = acid forming bacteria. SRB = sulfate reducing bacteria. MB = methanogenic bacteria.
VFA concentrations
In general, all four of the fatty acids measured in Cape Lookout Bight sediments show an increase in concentration in the sulfate-depleted portions of the sediment in warmer months as compared to colder months (Fig. 2). This increase may mean that higher decomposition rates due to higher temperatures result in higher pools of VFA intermediates. Alternatively, it may be the result of a depletion of sulfate during summer months; such a situation may cause a reduction in the activity of SRB and a resultant increase in the pools of their VFA substrates. However, the fact that acetate and propionate turnover rates in the CH4 zone show maxima during the summer (Figs. 3A,B) supports the hypothesis that the summer VFA concentrations are high due to high production rates rather than low consumption rates.
The concentrations of acetate and propionate are higher in the surface sediments as compared to deeper sediments during colder months, but are lower in the surface sediment during the summer. This near-surface fall/winter enrichment of acetate and propionate may be due to increases in the available organic matter content during this time (see below). Turnover rates for acetate and propionate show increases during this time (Figs. 3C,D), thereby again supporting the hypothesis that these increases are due to increases in production rates rather than due to decreases in consumption rates.
VFA concentrations measured in Cape Lookout Bight sediments are similar to values reported from other anaerobic sediments (Table 3). At present it is not known what fraction of whole-sediment, extractable VFA concentrations is due to VFAs dissolved in pore water, and how this fraction may vary among different sedimentary regimes; comparisons
such as those shown in Table 3 are thus difficult to interpret. In a single experiment on 13 July 1980, less than I% of the acetate measured in whole sediment from Cape Lookout Bight (0-30 cm) was found to be dissolved in pore water recovered by centrifugation. The remainder was presumably bound to the solid phase and also present in microbial cellular material (Sansone, 1980).
Acetate cycling in Cape Lookout Bight sediments
Significant methane production from acetate was observed in the CH4 zone (Fig. 3A) but not in the S04 zone (Fig. 3C). This result agreed with previous findings (e.g. Martens and Berner, 1974, 1977; Winfrey and Zeikus, 1977; Abram and Nedwell. 1978a) that significant methane production did not occur in the presence of appreciable sulfate concentrations. The present study did not, however, determine whether there was a significant amount of CO2 reduction to CH4 in sulfate-containing sediments. S0rensen et al. (1981) reported that CO2 reduction accounted for approximately 10% of the total H 2
utilization in sulfate-reducing sediment slurries. The rate of CH4 production, however, was approximately 50 times lower than that observed in similar sulfatedepleted slurries, and is thus consistent with the previous studies cited above.
Oremland and Taylor (1978) found small amounts of methane produced in homogenized sulfate-containing samples from the top 30 cm of Biscayne Bay, Florida sediments: These authors suggested a model similar to Fig. 5 to explain their results: SRB outcompete methanogens for H 2 by a factor of 1000 when sulfate is present.
In Santa Barbara Basin, methane production from labeled acetate and bicarbonate was observed in sul

1584 F. J. Sansone and C. S. Martens
Table 3. Comparison of reported VFA sediment concentrations. Tr = trace amount; NR not reported
Sediment VFA Concentrations
Location Site
Description Acetate Propionate Butyrate iso-Butyrate Units Reference
Cape Lookout Bight
Coas ta 1 Mari ne:
S04 Zone
CH4 Zone
55
40
-
-
210
660
-
-
16
13
<0.5
<0.5
-
- 8
<0.5
<0.5
- 4
-~mole/ls (whole sed.)
This work
Loch E;l, Scotland
Fjord Receiving Pulp Waste
D.9 - 12a Tr - D.2a Tr - 0.2a NR ~mo 1e/ 1s(whole sed.)
Miller et al. (1979)
(0-12 cm depth)
Limfjorden, Denmark
High Sulfate Coastal Marine
0.1 - 6.0 <0.1 - 6.0 <0.1 - 0.5 NR ~mole!lpw (pore water)
Ansbaek and Blackburn (1980)
(2-7°C, 0-9 em depth)
Santa Brbara Basin
High Su lfa te 33 - 471 <0.1 <0.1 - 549b ~mole/lpw (pore wa ter)
Barcelona (1980)
Newport Canyon
High Sulfate 51 - 1150 <0.1 <0.1 - 347b ~mole/lpw (pore water)
Barcelona (1980)
Lake Vech ten, The Ne ther 1ands
Low Sulfate (1-5 cm depth)
5 - 10 NR 1m ~mole/1pw (pore water)
Cappenberg and Jongejan (1977)
aAssuming 0.4 g dry weight/Is' bButyrate and lso-butyrate combi ned.
fate-reducing sediments (Kosiur and Warford, 1979; Warford et al., 1979). The rates measured were very low (5-42 mole·l-I,y-'), approximately 105_106
times lower than the rates of CH. production from acetate reported here for Cape Lookout Bight. Those authors concluded that, despite the fact that acetate was oxidized readily at all depths, bicarbonate and not acetate was the preferred substrate for methanogens.
From the results of the above-mentioned studies it can be concluded that, while small amounts of CH. may be produced, in sulfate-containing sediments methanogens are out-competed by SRB for the substrates acetate and/or H2 (Fig. 5). The rate of methane production in such environments seems to be several orders of magnitude less than the rate of sulfate reduction
Warford et al. (1979) observed the production of large amounts of I'C02 from 2- '·C-acetate added to sulfate-depleted Santa Barbara Basin sediments; production of only I·CH. would be expected if acetate were being cleaved to CO2 and CH. from the carboxyl and methyl carbons, respectively (Stadtman and Barker, 1949; Pine and Barker, 1956; Mah et al., 1978):
*CH3COO- + H 20" *CH. + HCOj, (3)
This observation is consistent with the results reported here that the respiration index of 1,2-I·C-acetate utilization in the CH. zone was approximately 0.8 (Fig. 4) instead of the value of 0.5 that would
be expected if the acetate were being cleaved only by methanogens. These observations can be explained by the hypothesis that sulfate-reducing bacteria are responsible for a significant portion of the CH. zone acetate utilization and consequently transfer hydrogen to methanogens because there is no sulfate present (Fig. 5). The methane produced from this hydrogen transfer would not have been detected in these experiments because the methane produced would have come from the reduction of unlabeled CO2 . This hypothesis is supported by the observation of Mountfort et ai, (1980) that the rate of reduction of added 35S0~- was similar in both sulfate-containing and sulfate-depleted sediments. They concluded that there were active populations of SRB in both environments. These data are also consistent with the observation by Winfrey and Zeikus (1979a) that the methyl carbon of acetate was converted to CO2 and CH. in the ratio 1:4 in anoxic lakewater with and without the addition of I mM SO•. Such a ratio would result in a RI value of 0.6 if the carboxyl carbon were oxidized solely to CO2,
The acetate turnover rate constant in the SO. zone shows maxima in early spring and in late summer (Fig. 3C). In addition, acetate concentrations diminish through the spring and summer and then rise drastically in the fall. These fluctuations result in a maximuQ1 in the apparent turnover rate in the fall, and are consistent with a model in which seasonal variations in organic matter input influence degradation rates in the upper 10 cm of the sediment.

1585 Sediment fatty acids
500 iment bacteria (Tison et al., 1980), no methanogens A with optimal activity in the psychrophilic range0
100
CH4 Zone""'~0
'..c:: • '0 l_fIJ
OJ 10 ~~ 0
::J '-../
Q)
E . "'" .~
-co c:: c
1000 B 0 0
0u 0 :> 0"0 0 0 804 Zone
100 0
00
10 '---...I..---I.-.----'-----'--.l----J 3.3 3.4 3.5 3.6
1/T (10-3 . K-1 )
FIG. 6. Arrhenius plots of CO2 (0) and CH. (.) production from 1,2-"C-acetate in (A) the CH. zone, and (B) the SO. zone of Cape Lookout Bight sediments.
During the fall the rate of sedimentation appears to increase dramatically due to the resuspension of nearby lagoonal sediments by strong autumn storms. This increase presumably results in the deposition of relatively labile organic matter. Such fresh input of substrate would, under summer conditions, be followed by higher decomposition rates; however, because of falling temperatures this organic matter is evidently not depleted until the following spring when temperatures rise again. Alternatively, the fall maximum in the apparent acetate turnover may result from some other factor such as an increase in the whole sediment acetate concentration due to a decrease in the rate constant. Ansbaek and Blackburn (1980) reported a similar seasonal variation in the acetate turnover rate constant in high-sulfate Limfjorden sediments (i.e., a spring minimum and a later summer maximum).
The acetate turnover rate constant in the CH. zone (Fig. 3A) did not vary much over the course of the year; there was, however, a large increase in the acetate concentration during late summer. As a result there was a maximum in the apparent turnover rate during late summer. Because the CH. zone was always at least 7 cm below the sediment surface it is assumed that the rate of sedimentation onto the surface sediments had little direct effect on the microbial processes in this zone. Instead, it is hypothesized that the temperature effect on bacteria was the dominant factor controlling acetate turnover. It is significant that, unlike many other groups of heterotrophic sed
«20°C) have been isolated (Zehnder, 1978). As a result it would be expected that methanogenic activity would decrease with decreasing temperature.
Acetate apparent turnover rates in both zones were examined to determine whether they followed the Arrhenius relationship (i.e., whether the log of the rate was a linear function of l' (OK-I); Giese, 1979). In the CH. zone the apparent production rates of CO2 and CH. from acetate showed good agreement with the Arrhenius relationship (Fig. 6A), thereby supporting the hypothesis that temperature was the dominant factor controlling the observed seasonal variations of acetate utilization in the CH. zone. Conversely, the data from the SO. zone (Fig. 6B) did not show an Arrhenius relationship between apparent rate and temperature, thus suggesting that factors other than temperature are limiting acetate turnover in this zone during at least part of the year.
In summary, the maximum apparent acetate turnover rates observed in the SO. and CH. zones were approximately the same. It is concluded, however, that the rates were principally controlled by substrate availability in the SO. zone and by temperature in the CH. zone.
There are no other reported seasonal studies of sediment VFA turnover rates with which to compare the above data, but two other sets of sediment acetate turnover rates have been reported (Table 4). The difference between these turnover rates was primarily due to differences in measured acetate pool sizes: the turnover rate constants measured in methane-producing sediments of Lake Vechten (0.07-0.24 h- 1
)
were similar to those in the CH. zone of Cape Lookout (0.36-1.6 h- 1
), and the turnover rate constants measured in the sulfate-reducing sediments of the Limfjorden (1.5-2.5 h- 1
) were similar to those in the SO. zone of Cape Lookout (0.74-4.2 h- 1
). If indeed I% of the whole sediment acetate in Cape Lookout sediments is dissolved in the pore water (see "Discussion-VFA Concentrations," above) then there is excellent agreement between the apparent rates presented here and those of the other two studies. It is likely that the microbially available acetate concentrations are smaller than the solvent-extractable whole-sediment concentrations reported here and larger than the pore water concentrations measured by Cappenberg and Jongejan (1977) and Ansbaek and Blackburn (1980). The latter effect would be due to the presumed bioavailability of some of the VFAs bound or otherwise associated with surfaces.
Continental slope acetate mineralization
Sediment VFA concentrations measured at the continen~al slope site were much lower. than those measured in sulfate-containing Cape Lookout sediments (Table I vs. Fig. 2). The lack of a gradient in the continental slope sulfate profile below the

1586 F. J. Sansone and C. S. Martens
Table 4. Comparison of reported sedimentary acetate turnover rates
Reported Values
Site Acetate Turnover Acetate Loca t i on Description Ra te Cons t. (h-1) Turnover Ra te Units Reference
Cape Lookout Coastal Marine: Bight
S04 Zone 0.74-4.2
CH 4 Zone 0.36 - 1. 6 50 - 600
Limfjorden, Denmark
Coastal Marine:
(2-70 C, 0-10 cm, S04" 22 mM)
1.5 - 2.5 1 - 12 (~mole·rl .h-l) pore water
Ansbaek & Blackburn (1980)
Lake Vechten, Low Sulfate The Netherlands
(0-5 cm depth) 0.07 - 0.24 0.3-1.7 (~mole'l-l pore
.h- 1) water
Cappenberg&Jongejan (1977)
98 - 540
heavily bioturbated upper 10 cm suggests that decomposition rates were slower at that site.
The rate constants for CO2 production from acetate decreased rapidly with depth at the slope station (Table 2); the 0-5 cm rate constant was approximately 6 times smaller than that measured in the S04 zone of Cape Lookout sediments (11.9°C, January 1980; Fig. 3C).
The apparent rates of CO2 production from acetate were estimated by using rate constants and whole sediment concentrations from separate years (Table 2). The surface sediment rates were approximately twenty times smaller than those measured in the S04 zone of Cape Lookout sediments at similar temperatures (l1.9°C, January 1980; Fig. 3C). Rates decrease rapidly with depth, thereby suggesting that substrate limitation was important.
Rate constant results were most variable in samples from the top 13 cm (Table 2). This was probably due to the activity of the macroinfauna, which appeared to be most abundant in the surface 0-20 cm of sediment. The better reproducibility of the deeper rate constant measurements was presumably due to the greater sediment homogeneity with depth that was, in turn, a result of lower levels of infaunal activity. Future measurements will be needed to determine how representative this site is of deep-ocean sedimentary environments.
Propionate cycling
In both the S04 and CH4 zones of Cape Lookout sediments the l- 14C-propionate turnover rate constants generally followed the variation in the sediment temperature (Figs. 3B,D). Propionate concentrations, however, showed different patterns in each case. Depletion of propionate in the summer in the S04 zone resulted in a bimodal apparent turnover rate with a minimum in the summer similar to that seen with acetate in this zone. The accumulation of propionate in the CH4 wne during the summer re-
suited in an apparent turnover rate with a single major maximum in the summer, again paralleling the situation observed with acetate.
Sansone (1980) determined that the apparent production of CO2 from all carbons of propionate in Cape Lookout sediments was always less than 10% of that of acetate. Thus, he concluded that propionate was not the major source of acetate (or acetyJ-CoA) for microbes in this environment, assuming that similar fractions of the acetate and propionate pools were available to the microbes. Instead, it is likely that lactate and/or pyruvate were the most important acetate precursors (Fig. 5).
Unlike the case of acetate, I-C-propionate apparent CO2 production rates did not show well-defined Arrhenius-type temperature dependencies in either zone. Significant methane production from 1_ 14C_ propionate was not observed in either zone (Figs. 3B,D); however, measurable methane was produced from the other two propionate carbon atoms at 20°C in the CH4 zone (Sansone, 1980). No significant methane production from any propionate carbon was measured at 8°C. The apparent methane ;:>roduction from all carbons of propionate was found to be less than I% of that from acetate at 20°C in the CH4 zone. This is not surprising because propionate is not a known substrate for methanogens (Mah et al., 1977; Zehnder, 1978). Any methane produced from propionate carbon is presumed to be derived from acetate and/or CO2 produced from propionate oxidation.
Butyrate and iso-butyrate cycling
The large butyrate concentration maximum (Fig. 2) at the depth of sulfate depletion during the summer suggests that butyrate is an intermediate in the degradation of organic matter in the CH4 zone. Butyrate has been found to be a major intermediate in the anaerobic decomposition of cellulose in the rumen ecosystem (Hungate, 1975); bacteria of the genera

1587 Sediment fatty acids
Clostridium and Butyrvibrio were primarily responsible for the butyrate formation. Butyrate has been shown to be a substrate for acetate production via ,a-oxidation in anaerobic digestor sludge (McInerney et al., 1979; Mcinerney et al., 1981), and as such may be a significant precursor for CO2and CH4 production (Fig. 5).
The low concentrations of iso-butyrate in Cape Lookout sediments (Fig. 2), along with the lack of distinct seasonal maxima in the concentrations, may indicate that iso-butyrate is not an important intermediate in organic matter degradation in these sediments. As in the case of butyrate, it will be necessary to have data on its rate of utilization before its importance can be determined. Hungate (1975) found that a number of rumen bacteria require iso-butyrate in small concentrations as a growth factor (i.e., not used as a source of energy or as a major source of cell carbon). The fact that iso-butyrate was only used as a growth factor suggests it may not be important in anaerobic decomposition processes.
CONCLUSIONS
In Cape Lookout Bight sediments acetate and propionate turnover rates were strongly influenced by effects of biogeochemical zonation, temperature, and availability of organic substrates. Of primary importance were sediment temperature and the availability of inorganic electron acceptors (i.e., biogeochemical zonation). It was hypothesized that the apparent rates of acetate and propionate turnover in the S04 zone also were strongly influenced by seasonal episodes of sedimentation/resuspension during the fall. The seasonal variations in apparent turnover rates in the CH4 zone seemed to be controlled primarily by temperature, especially in the case of acetate.
Variations in these calculated apparent turnover rates were due mainly to changes in the respective whole sediment VFA concentrations rather than the relatively small changes in the turnover rate constants. The high acetate and propionate concentrations observed in the summer appeared to be due to high production rates rather than low consumption rates.
Turnover rates had been expected to be higher in the S04 zone than in the CH4 zone due to (I) the closer proximity of the S04 zone to the sedimenting organic matter, and (2) the higher theoretical free energy yield of S04 reduction compared to that of methanogenesis. The apparent turnover rates for propionate were higher in the S04 zone than in the CH4 zone, but in the case of acetate the rates were comparable in both zones. This may have been due to the utilization of acetate by both SRB and methanogens, and the utilization of propionate only by SRB.
Acetate molecules were oxidized to two molecules of CO2 in the S04 zone but were split to form, on the average, 1.6 molecules of CO2and 0.4 molecules
of CH4 in sulfate-depleted sediments (Fig. 4). Thus, in the latter case, acetate oxidizers accounted for 60% of the acetate turnover and acetate fermenters (methanogens) accounted for 40%. It was concluded that other precursor molecules besides propionate were important in the production of acetate in these sediments.
The data presented here also support the conclusion of previous studies (e.g. Winfrey and Zeikus, 1977) that methanogenesis is not important in sulfate-rich anoxic environments due to the ability of SRB to out-compete methanogens for acetate and/ or H 2•
The whole sediment concentrations of butyrate and iso-butyrate were measured seasonally in Cape Lookout sediments, and were interpreted as indicating that butyrate may have been a significant fermentation intermediate in this system but iso-butyrate probably was not.
North Carolina continental slope sediments showed much lower VFA concentrations than Cape Lookout sediments, probably because of a lower input rate of utilizable organic substrates to the slope sediments. Acetate mineralization was approximately twenty times slower than that in the S04 zone of Cape Lookout sediments. Large variations in the measured acetate mineralization rate in the upper 15 cm of continental slope sediments was attributed to sediment reworking and irrigation by macroinfaunal organisms.
Finally, it is apparent that there is a great need to combine geochemical analytical techniques with microbiological methods in order to understand the dynamics of organic matter degradation in these complex systems. Indeed, the factor most restricting our knowledge of the cycling rates of organic intermediates is the reliable measurement of the microbially available pool sizes of these compounds in sediments.
Acknowledgements-We thank the members of team CH,.A02S for their much appreciated support and assistance. We especially thank George Kipphut for his extensive help with field operations and laboratory analyses, and for extensive discussions. Jeff Chanton, Patrick Crill, Karen Gruebel, and Val Klump also provided expert field assistance.
The staff of the UNC Institute of Marine Science in Morehead City, N.C. provided support in many phases of this study, as well as laboratory and office space. Dr. T. Rice and the National Marine Fisheries Service, Beaufort, N.C. provided convenient access to equipment not otherwise available. This research was supported by NSF grants OCE78-09485 and OCE80-09245 (C.S.M.) and OCE7809507 (F.J.S.) from the Marine Chemistry Program, Oceanography Section. Hawaii Institute of Geophysics Contribution No. 1260.
REFERENCES
Abram 'J. W. and Nedwell D. B. (l978a) Inhibition of methanogenesis by sulfate-reducing bacteria competing for transferred hydrogen. Arch. Microbiol. 117, 89-92.
Abram J. W. and Nedwell D. B. (l978b) Hydrogen as a substrate for methanogenesis and sulfate reduction in

1588 F. J. Sansone and C. S. Martens
anaerobic saltmarsh sediments. Arch. Microbiol. 117, 93-97.
Alexander M. (1961) Introduction to Soil Microbiology, John Wiley & Sons, Inc., New York.
Aller R. C. and Yingst J. Y. (1980) Relationship between microbial distributions and the decomposition of organic mailer in surface sediments of Long Island Sound, U.S.A. Mar. Bioi. 56,29-42.
Ansbaek J. and Blackburn T. H. (1980) A method for the analysis of acetate turnover in a coastal marine sediment. Microbiol. Ecol. 5, 253-264.
Balch W. E., Fox G. E., Magrum L. J., Woese C. R. and Wolfe R. S. (1979) Methanogens: Reevaluation of a unique biological group. Microbiol. Rev. 43, 260-296.
Barcelona M. J. (1980) Dissolved organic carbon and volatile fally acids in marine sediment pore waters. Geochim. Cosmochim. Acta 44, 1977-1984.
Bartlett K. B. (1981) Seasonal variation in sulfate reduction and macrofaunal irrigation rates in an organic-rich coastal sediment. M.S. Thesis, Univ. of North Carolina at Chapel Hill.
Ben-Yaakov S. (1973) pH buffering of pore water of recent anoxic marine sediments. Limnol. Oceanogr. 18, 86-94.
Berner R. A., Scoll M. R. and Thomlinson C. (1970) Carbonate alkalinity in the pore waters of anoxic marine sediments. Limnol. Oceanogr. 15, 544-549.
Boone D. R. and Bryant M. P. (1980) Propionate-degrading bacterium Syntrophobacter wolinii sp. nov. gen. nov., from methanogenic ecosystems. Appl. Environ. Microbioi. 40, 626-632.
Brock T. D. (1979) Biology of Microorganisms, PrenticeHall, Englewood Cliffs, N.J.
Bryant M. P., Campbell L. L., Reddy C. A. and Crabill M. E. (1977) Growth of Desulfovibrio in lactate or ethanol media low in sulfate in association with H2-utilizing bacteria. Appl. Environ. Microbiol. 33, 11621169.
Bryant M. P., Wolin E. A., Wolin M. J. and Wolfe R. S. (1967) Methanobacillus omelianskii, a symbiotic association of two species of bacteria. Arch. fur Mikrobiol. 59,20-31.
Cappenberg Th. E. and Jongejan E. (1977) Microenvironments for sulfate reduction and methane production in freshwater sediments. In Environmental Biogeochemistry and Geomicrobiology (ed. W. E. Krumbein), Proc. Third Int. Symp. Environ. Biochem., pp. 129-138. Ann Arbor Science, Ann Arbor, Mich.
Chanton J. P. (1979) Pb-210 geochronology of a changing environment: Cape Lookout Bight, N.C. M.S. Thesis, Univ. of North Carolina at Chapel Hill.
Christian R. R. and Wiebe W. J. (1978) Anaerobic microbial community metabolism in Spartina alterniflora soils. Limnol. Oceanogr. 23, 328-336.
Claypool G. E. and Kaplan I. R. (1974) The origin and distribution of methane in marine sediments. In Natural Gases in Marine Sediments (ed. I. R. Kaplan), pp. 99139. Plenum Press, N.Y.
Fenchel T. M. and J"rgensen B. B. (1977) Detritus food chains of aquatic ecosystems: The role of bacteria. Adv. Microbiol. Ecol. 1, I-58.
Froelich P. N., Klinkhammer G. P., Bender M. L., Leudtke N. A., Heath G. R., Cullen D., Dauphin P., Hammond D., Hartman B. and Maynard V. (1979) Early oxidation of organic matter in pelagic sediments of the eastern equatorial Atlantic: Suboxic diagenesis. Geochim. Cosmochim. Acta 43, 1075-1090.
Giese A. C. (1979) Cell Physiology, 5th edn., W. B. Saunders, Philadelphia, pp. 234-235.
Goldhaber M. B. and Kaplan I. R. (1974) The sulfur cycle. In The Sea (ed. E. D. Goldberg), Vol. 5, pp. 569-655. Wiley and Sons, N.Y.
Goldhaber M. B. and Kaplan I. R. (1975) Controls and
consequences of sulfate reduction rates in recent marine sediments. Soil Sci. 119, 42-45.
Healy J. B. Jr., Young L. Y. and Reinhard M. (1980) Methanogenic decomposition of ferrulic acid, a model lignin derivative. Appl. Environ. Microbiol. 39, 436-444.
Hungate R. E. (1966) The Rumen and Its Microbes, Academic Press, N.V.
Hungate R. E. (1967) Hydrogen as an intermediate in the rumen fermentation. Arch. fur Mikrobiol. 59, 158-164.
Hungate R. E. (1975) The rumen microbial ecosystem. Ann. Rev. Ecol. Systematics 6, 39-66.
Iannotti E. L., Kafkewitz D., Wolin M. J. and Bryant M. P. (1973) Glucose fermentation products of Ruminococcus albus grown in continuous culture with Vibrio succinogenes. Changes caused by interspecies transfer of H 2• J. Bacteriol. 114, 1231-1240.
Jeris J. S. and McCarty P. L. (1965) The biochemistry of methane fermentation using 14C tracers. J. Water Pollut. Control Fed. 37, 179-192.
J"rgensen B. B. (1977) The sulfur cycle of a coastal marine sediment (Limfjorden, Denmark). Limnol. Oceanogr. 22, 814-832.
Khan A. W. and Trottier T. M. (1978) Effect of sulfurcontaining compounds on anaerobic degradation of cellulose to methane on mixed cultures obtained from sewage sludge. Appl. Environ. Microbiol. 35, 1027-1034.
Klump J. V. (1980) Benthic nutrient regeneration and mechanisms of chemical sediment-water exchange in an organic-rich coastal marine sediment. Ph.D. Thesis, Univ. of North Carolina at Chapel Hill.
Kosiur D. R. and Warford A. L. (1979) Methane production and oxidation in Santa Barbara Basin sediments. Estuarine Coastal Mar. Sci. 8, 379-385.
McInerney M. J. and Bryant M. P. (1981) Anaerobic degradation of lactate by syntrophic associations of Methanosarcina barkeri and Desulfovibrio species and effect of H 2 on acetate degradation. Appl. Environ. Microbiol. 41, 346-354.
McInerney M. J., Bryant M. P., Hespell R. B. and Costerton J. W. (1981) Syntrophomonas wolfei gen. nov. sp. nov., an anaerobic syntrophic, fatty acid-oxidizing bacterium. Appl. Environ. Microbiol. 41, 1029-1039.
McInerney M. J., Bryant M. P. and Pfennig N. (1979) Anaerobic bacterium that degrades fatty acids in syntrophic association with methanogens. Arch. Microbiol. 122,129-135.
Mah R. A., Smith M. R. and Baresi L. (1978) Studies on an acetate-fermenting strain of Methanosarcina. Appl. Environ. Microbiol. 35, 1174-1184.
Mah R. A., Ward D. M., Baresi L. and Glass T. L. (1977) Biogenesis of methane. Ann. Rev. Microbiol. 31, 309341.
Mann K. H. (1972) Macrophyte production and detritus food chains in coastal areas. Mem. 1st. Ital. Idrobiol. Suppl. 29, 353-382.
Martens C. S. (1976) Control of methane sediment-water bubble transport by macroinfaunal irrigation in Cape Lookout Bight, North Carolina. Science 192,998-1000.
Martens C. S. (1978) Some or the chemical consequences of microbially mediated degradation of organic materials in estuarine sediments. In Biogeochemistry of Estuarine Sediments (ed. E. D. Goldberg), Proc. UNESCO/SCOR Workshop, pp. 266-278. Melreux, Belgium. UNESCO.
Martens C. S. and Berner R. A. (1974) Methane production in the interstitial waters of sulfate-depleted marine sediments. Science 185,1167-1169.
Martens C. S. and Berner R. A. (1977) Interstitial water chemistry of anoxic Long Island Sound sediments. I. Dissolved gases. Limnol. Oceanogr. 22, 10-25.
Martens 'c. S., Kipphut G. W. and Klump J. V. (1980) Sediment-water chemical exchange in the coastal zone traced by in situ Radon-222 flux measurements. Science 208, 285-288.

1589 Sediment fatty acids
Martens C. S. and Klump J. V. (1980) Biogeochemical cycling in an organic-rich coastal marine basin. I. Methane sediment-water exchange processes. Geochim. Cosmochim. Acta 44, 471-490.
Mechalas B. J. (1974) Pathways and environmental requirements for biogenic gas production in the ocean. In Natural Gases in Marine Sediments (ed. I. R. Kaplan), pp. 11-25. Plenum Press, N.Y.
Miller D., Brown D. M., Pearson T. H. and Stanley S. O. (1979) Some biologically imponant low molecular weight organic acids in the sediments of Loch Eil. Mar. Bioi. 50, 375-383.
Mountfort D.O., Asher R. A., Mays E. L. and Tiedje J. M. (1980) Carbon and electron flow in mud and sandflat intertidal sediments at Delaware Inlet, Nelson, New Zealand. Appl. Environ. Microb. 39,686-694.
Oremland R. S. and Taylor B. F. (1978) Sulfate reduction and methanogenesis in marine sediments. Geochim. Cosmochim. Acta 42,209-214.
Pine M. J. and Barker H. A. (1956) Studies on the methane fermentation. XII. The pathway of hydrogen in the acetate fermentation. J. Bact. 71, 644-648.
Postgate J. R. (1979) The Sulphate-reducing Bacteria, Cambridge University Press, Cambridge.
Reddy C. A., Bryant M. P. and Wolin M. J. (1972) Characteristics of S organism isolated from Methanobaci//us omelianskii. J. Bacteriol. 109, 539-545.
Sansone F. J. (1980) Volatile fatty acid cycling in anoxic coastal sediments. Ph.D. dissertation, Univ. of North Carolina at Chapel Hill.
Sansone F. J. and Martens C. S. ( 1978) Methane oxidation in Cape Lookout Bight, N.C. Limnol. Oceanogr. 23,349355.
Sansone F. J. and Martens C. S. (l98Ia) Methane production from acetate and associated methane fluxes from anoxic coastal sediments. Science 211, 707-709.
Sansone F. J. and Martens C. S. (l98Ib) Determination of volatile fatty acid turnover rates in organic-rich marine sediments. Mar. Chem. 10,233-248.
Serensen J., Christensen D. and Jergensen B. B. (1981) Volatile fatty acids and hydrogen as substrates for sulfate-reducing bacteria in anaerobic marine sediment. Appl. Environ. Microbiol. 42, 5-11.
Stadtman T. C. and Barker H. A. (1949) Studies on the methane fermentation. VII. Tracer experiments on the mechanism of methane formation. Arch. Biochem. 21, 256-264.
Tenore K. R. (1977) Food chain pathways in detrital feeding benthic communities: A review, with new observations on sediment resuspension and detrital recycling. In Ecol
ogy of Marine Benthos (ed. B. C. Coull), pp. 37-53. Univ. South Carolina Press, Columbia, S.C.
Tezuka Y. (1966) A commensalism between sulfate-reducing bacterium Desulfovibrio desulfuricans and other heterotrophic bacteria. Bot. Mag. 79, 174-178.
Thauer R. K., Jungermann K. and Decker K. (1977) Energy conversation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 41, 100-180.
Thorstenson D. C. (1970) Equilibrium distribution of small organic molecules in natural waters. Geochim. Cosmochim. Acta 34, 745-770.
Tison D. C., Pope D. H. and Boylen C. W. (1980) Influence of seasonal temperature on the temperature optima of bacteria in sediments of Lake George, New York. Appl. Environ. Microbiol. 39, 676-677.
Toerien D. F. and Hattingh W. H. J. (1969) Anaerobic digestion. I. The microbiology of anaerobic digestion. Wat. Res. 3, 385-408.
Warford A. L., Kosiur D. R. and Doose P. R. (1979) Methane production in Santa Barbara basin sediments. Geomicrob. J. 1,117-137.
Widdel F. (1980) Anaerober Abbau von Fettsauren und Benzoesauer durch neu isolierte Arten von Sulfat-reduzierenden Bakterien. Doctoral Thesis, Univ. of Gottingen, F.D.R.
Winfrey M. R., Nelson D. R., Klevickis S. C. and Zeikus J. G. (1977) Association of hydrogen metabolism with methanogenesis in Lake Mendota sediments. Appl. Environ. Microbiol. 33, 312-318.
Winfrey M. R. and Zeikus J. G. (1977) Effects of sulfate on carbon and electron flow during microbial methanogenesis in freshwater sediments. Appl. Environ. Microbioi. 33, 275-281.
Winfrey M. R. and Zeikus J. G. (l979a) Microbial methanogenesis and acetate metabolism in a meromictic lake. Appl. Environ. Microbiol. 37, 213-221.
Winfrey M. R. and Zeikus J. G. (l979b) Anaerobic metabolism of immediate methane precursors in Lake Mendota. Appl. Environ. Microbiol. 37, 244-253.
Wolin M. J. (1976) Interactions between H2-producing and methane-producing species. In Microbial Formation and Utilization of Gases (ed. H. G. Schlegel, G. Gottschalk and N. Pfennig), pp. 141-150. E. Goltze, Gottingen.
Zehnder A. J. B. (1978) Ecology of methane formation. In Water Pollution Microbiology (ed. R. Mitchell), Vol. II. Wiley, N.Y.
Zehnder A. J. B., Huser B. and Brock T. D. (1979) Measuring radioactive methane with the liquid scintillation counter. Appl. Environ. Microbiol. 37, 897-899.
Zeikus J. G. (1977) The biology of methanogenic bacteria. Bact. Rev. 41, 514-541.


















