
Vol. 27 February 2004 No. 1 CONTENTS Volume 27, Number 1 ... · — Volume 27, Number 1 —...
14
Vol. 27 February 2004 No. 1 CONTENTS (Continued on inside back cover) ANGUS FERGUSON, BRADLEY EYRE, AND JENNITA GAY . Nutrient Cycling in the Sub-tropical Brunswick Estuary, Australia .......................................................................................................... 1 ALISON M. FEATHERSTONE, EDWARD C. V. BUTLER, AND BARRY V. O’GRADY . Seasonal Cycling of Arsenic Species in a Stratified, Microtidal Estuary (Huon River, Tasmania) ................................ 18 J. A. HALE, T. K. FRAZER, D. A. TOMASKO, AND M. O. HALL. Changes in the Distribution of Seagrass Species along Florida’s Central Gulf Coast: Iverson and Bittaker Revisited ................................ 36 VICTOR H. RIVERA-MONROY , ROBERT R. TWILLEY , ERNESTO MEDINA, E. BARRY MOSER, LEONOR BOTERO, ANA MARTA FRANCISCO, AND EVELYN BULLARD. Spatial Variability of Soil Nutrients in Disturbed Riverine Mangrove Forests at Different Stages of Regeneration in the San Juan River Estuary, Venezuela .......................................................................................................................... 44 KENNETH W. ABLE, DAVID M. NEMERSON, AND THOMAS M. GROTHUES. Evaluating Salt Marsh Restoration in Delaware Bay: Analysis of Fish Response at Former Salt Hay Farms .................. 58 GAIL L. CHMURA AND GRACE A. HUNG. Controls on Salt Marsh Accretion: A Test in Salt Marshes of Eastern Canada .......................................................................................................................... 70 CYNTHIA H. P AQUETTE, KAREN L. SUNDBERG, ROELOF M. J. BOUMANS, AND GAIL L. CHMURA. Changes in Saltmarsh Surface Elevation Due to Variability in Evapotranspiration and Tidal Flooding .......................................................................................................................................... 82 J ANE M. CAFFREY . Factors Controlling Net Ecosystem Metabolism in U.S. Estuaries .................... 90 CHRISTOPHER N. ROOPER, DONALD R. GUNDERSON, AND DAVID A. ARMSTRONG. Application of the Concentration Hypothesis to English Sole in Nursery Estuaries and Potential Contribution to Coastal Fisheries ............................................................................................................................ 102 MARY DOWNES GASTRICH, JUSTINE A. LEIGH-BELL, CHRISTOPHER J. GOBLER, O. ROGER ANDERSON, STEVEN W. WILHELM, AND MARTHA BRYAN. Viruses as Potential Regulators of Regional Brown Tide Blooms Caused by the Alga, Aureococcus anophagefferens ................................................ 112 MAURICIO ESCAPA, OSCAR IRIBARNE, AND DIEGO NAVARRO. Effects of the Intertidal Burrowing Crab Chasmagnathus granulatus on Infaunal Zonation Patterns, Tidal Behavior, and Risk of Mortality 120 E. W. NORTH, S.-Y. CHAO, L. P. SANFORD, AND R. R. HOOD. The Influence of Wind and River Pulses on an Estuarine Turbidity Maximum: Numerical Studies and Field Observations in Chesapeake Bay .................................................................................................................................................. 132 HONGBIN LIU, MICHAEL DAGG, LISA CAMPBELL, AND JUANITA URBAN-RICH. Picophytoplankton and Bacterioplankton in the Mississippi River Plume and its Adjacent Waters .................................. 147 Technical Notes and Comments BRIAN E. LAPOINTE AND PETER J. BARILE.Comment on J. C. Zieman, J. W. Fourqurean, and T. A. Frankovich. “Seagrass Die-off in Florida Bay: Long-term Trends in Abundance and Growth of Turtle Grass, Thalassia testudinum.” 1999. Estuaries 22:460–470. .............................................. 157
Transcript of Vol. 27 February 2004 No. 1 CONTENTS Volume 27, Number 1 ... · — Volume 27, Number 1 —...

—V
olume
27,N
umber
1—
February2004
Vol. 27 February 2004 No. 1
CONTENTS
(Continued on inside back cover)
ANGUS FERGUSON, BRADLEY EYRE, AND JENNITA GAY. Nutrient Cycling in the Sub-tropical Brunswick Estuary, Australia .......................................................................................................... 1
ALISON M. FEATHERSTONE, EDWARD C. V. BUTLER, AND BARRY V. O’GRADY. Seasonal Cycling of Arsenic Species in a Stratified, Microtidal Estuary (Huon River, Tasmania) ................................ 18
J. A. HALE, T. K. FRAZER, D. A. TOMASKO, AND M. O. HALL. Changes in the Distribution of Seagrass Species along Florida’s Central Gulf Coast: Iverson and Bittaker Revisited ................................ 36
VICTOR H. RIVERA-MONROY, ROBERT R. TWILLEY, ERNESTO MEDINA, E. BARRY MOSER, LEONOR
BOTERO, ANA MARTA FRANCISCO, AND EVELYN BULLARD. Spatial Variability of Soil Nutrients inDisturbed Riverine Mangrove Forests at Different Stages of Regeneration in the San Juan River Estuary, Venezuela .......................................................................................................................... 44
KENNETH W. ABLE, DAVID M. NEMERSON, AND THOMAS M. GROTHUES. Evaluating Salt Marsh Restoration in Delaware Bay: Analysis of Fish Response at Former Salt Hay Farms .................. 58
GAIL L. CHMURA AND GRACE A. HUNG. Controls on Salt Marsh Accretion: A Test in Salt Marshes of Eastern Canada .......................................................................................................................... 70
CYNTHIA H. PAQUETTE, KAREN L. SUNDBERG, ROELOF M. J. BOUMANS, AND GAIL L. CHMURA.Changes in Saltmarsh Surface Elevation Due to Variability in Evapotranspiration and Tidal Flooding .......................................................................................................................................... 82
JANE M. CAFFREY. Factors Controlling Net Ecosystem Metabolism in U.S. Estuaries .................... 90
CHRISTOPHER N. ROOPER, DONALD R. GUNDERSON, AND DAVID A. ARMSTRONG. Application of theConcentration Hypothesis to English Sole in Nursery Estuaries and Potential Contribution to Coastal Fisheries ............................................................................................................................ 102
MARY DOWNES GASTRICH, JUSTINE A. LEIGH-BELL, CHRISTOPHER J. GOBLER, O. ROGER ANDERSON,STEVEN W. WILHELM, AND MARTHA BRYAN. Viruses as Potential Regulators of Regional Brown Tide Blooms Caused by the Alga, Aureococcus anophagefferens ................................................ 112
MAURICIO ESCAPA, OSCAR IRIBARNE, AND DIEGO NAVARRO. Effects of the Intertidal Burrowing Crab Chasmagnathus granulatus on Infaunal Zonation Patterns, Tidal Behavior, and Risk of Mortality 120
E. W. NORTH, S.-Y. CHAO, L. P. SANFORD, AND R. R. HOOD. The Influence of Wind and River Pulseson an Estuarine Turbidity Maximum: Numerical Studies and Field Observations in Chesapeake Bay .................................................................................................................................................. 132
HONGBIN LIU, MICHAEL DAGG, LISA CAMPBELL, AND JUANITA URBAN-RICH. Picophytoplankton and Bacterioplankton in the Mississippi River Plume and its Adjacent Waters .................................. 147
Technical Notes and Comments
BRIAN E. LAPOINTE AND PETER J. BARILE. Comment on J. C. Zieman, J. W. Fourqurean, and T. A.Frankovich. “Seagrass Die-off in Florida Bay: Long-term Trends in Abundance and Growth of Turtle Grass, Thalassia testudinum.” 1999. Estuaries 22:460–470. .............................................. 157
ISSN: 0160-8347
AN INTERNATIONAL JOURNAL OF COASTAL SCIENCEPUBLISHED BY THE
ESTUARINE RESEARCH FEDERATION
Volume 27, Number 1—February 2004

SCOPE
ESTUARIES is a bimonthly interdisciplinary journal focused oncoastal science published by the Estuarine Research Federation.The journal ESTUARIES publishes manuscripts presenting origi-nal data, ideas, and interpretations on the coastal zone. The coastalzone refers to the interface between land and ocean, and is broad-ly defined to include areas within estuaries, lagoons, wetlands,tidal rivers, watersheds that include estuaries, and adjacent coastaland shelf waters.
Manuscripts may present original research, reviews, tech-niques, or technical comments, but in all cases will be judged onthe basis of their contribution and broad relevance to coastal sci-ence. Manuscripts must be written in English but may have a sec-ond abstract in an alternate language when appropriate. Onlymaterial intended for publication in ESTUARIES will be consid-ered. Manuscripts will be reviewed by a member of the EditorialBoard and by at least two anonymous referees.
JOSEPH C. ZIEMAN, JAMES W. FOURQUREAN, AND THOMAS A. FRANKOVICH. Reply to B. E. Lapointeand P. J. Barile (2004). Comment on J. C. Zieman, J. W. Fourqurean, and T. A. Frankovich.“Seagrass Die-off in Florida Bay: Long-term Trends in Abundance and Growth of Turtle Grass, Thalassia testudinum.” 1999. Estuaries 22:460–470. .................................................................... 165
TODD M. KANA AND DAVID L. WEISS. Comment on “Comparison of Isotope Pairing and N2:ArMethods for Measuring Sediment Denitrification” By B. D. Eyre, S. Rysgaard, T. Dalsgaard, and P. Bondo Christensen. 2002. Estuaries 25:1077–1087. .................................................................. 173
BRADLEY D. EYRE, SØREN RYSGAARD, TAGE DALSGAARD, AND PETER BONDO CHRISTENSEN. Reply to Comment on Our Paper “Comparison of Isotope Pairing and N2:Ar Methods for MeasuringSediment Denitrification.” .............................................................................................................. 177
(USPS 405-130)
JOURNAL OF THE ESTUARINE RESEARCH FEDERATION
ESTUARIES traces its origin to the late Dr. Romeo J. Mansueti, who conceived and nurtured it as CHESAPEAKE SCIENCE, a journal devoted to presenting research results and management studies on natural resources of the Chesapeake Bay region. CHESAPEAKE SCIENCE was pub-lished and partially subsidized by the Chesapeake Biological Laboratory during its eighteen year history.
The contents of the articles contributed to ESTUARIES, including any opinions or technical information included or referred to, are the sole responsibility of the author(s). The Estuarine Research Federation disclaims any influence over or liability for any inaccuracies in or omissionsfrom such articles or the results of their use by any person.
Copyright © 2004 Estuarine Research Federation.
ESTUARIES is issued on a calendar-year basis. Delivery outside the U.S.A. is not guaranteed, but most subscriptions will beshipped air mail. Correspondence concerning back issues and library or institutional subscriptions should be addressed to theEstuarine Research Federation, P. O. Box 1897, Lawrence, KS 66044-8897, U.S.A. Telephone (785)843-1235. A membershipapplication and subscription form is available at the back of this issue. ESTUARIES (ISSN 0160-8347) is published bimonthly atthe membership rate of $110.00, which covers both membership to the Federation and a subscription to the journal, and a rateof $470.00 for nonmember, library or institutional subscriptions in the United States, Canada and Mexico and of $490.00 forthose in all other countries by the Estuarine Research Federation, 2018 Daffodil Ln., Port Republic, MD 20676. Periodicalspostage paid at Port Republic, MD 20676 and additional mailing offices. POSTMASTER: send address changes to ESTUARIES,P. O. Box 1897, Lawrence, KS 66044-8897.
a This paper meets the requirements of ANSI/NISO Z39.48-1992 (Permanence of Paper).
AFFILIATE SOCIETIES
NEW ENGLAND ESTUARINE RESEARCH SOCIETYPAM ARNOFSKY NEUBERTENSR Consulting EngineeringWoods Hole, Massachusetts540/457-7900; [email protected] ESTUARINE RESEARCH SOCIETYTHEODORE DEWITTU.S. Environmental Protection AgencyNewport, Oregon541/867-4029; [email protected] ESTUARINE RESEARCH SOCIETYMICHAEL A. MALLINUniversity of North Carolina Center for Marine ScienceWilmington, North Carolina910/962-2358; [email protected]
ATLANTIC CANADA COASTAL ESTUARINE SCIENCE SOCIETYSYLVIE ST-JEANNational Water Research TrustBurlington, Ontario506/851-3195; [email protected]
ATLANTIC ESTUARINE RESEARCH SOCIETYCHRIS SWARTHJug Bay Wetlands SanctuaryLothian, Maryland410/741-9330; [email protected]
CALIFORNIA ESTUARINE RESEARCH SOCIETYRANDALL L. BROWN4258 Brookhill DriveFair Oaks, California916/961-5449; [email protected]
GULF ESTUARINE RESEARCH SOCIETYCHRISTOPHER ONUFU.S. Geological Survey/NWRCCorpus Christi, Texas361/985-6266; [email protected]
LINDA SCHAFFNER, PresidentSchool of Marine ScienceThe College of William and MaryGloucester Point, Virginia804/684-7366; [email protected]
DENNIS ALLEN, Past PresidentBaruch Marine LabUniversity of South CarolinaGeorgetown, South Carolina843/546-6219; [email protected]
ROBERT CHRISTIAN, President ElectEast Carolina UniversityGreenville, North Carolina252/328-1835; [email protected]
LISA V. LUCAS, SecretaryU.S. Geologcial SurveyMenlo Park, California605/329-4588; [email protected]
CAROLYN KEEFE, TreasurerChesapeake Biological LabSolomons, Maryland410/326-7293; [email protected]
HOLLY GREENING, Member-at-LargeTampa Bay Estuary ProgramSt. Petersburg, Florida727/893-2675; [email protected]
MERRYL ALBER, Member-at-LargeUniversity of GeorgiaAthens, Georgia706/542-5966; [email protected]
JAMES W. FOURQUREAN, Member-at-LargeFlorida International UniversityMiami, Florida305/348-4084; [email protected]
JORGE A. HERRERA-SILVEIRA, Member-at-LargeCINVESTAV-IPNMeridia, Yucatan, [email protected]
JOY BARTHOLOMEW, Executive DirectorEstuarine Research FederationPort Republic, Maryland410/586-0997; [email protected]

58Q 2004 Estuarine Research Federation
Estuaries Vol. 27, No. 1, p. 58–69 February 2004
Evaluating Salt Marsh Restoration in Delaware Bay: Analysis of
Fish Response at Former Salt Hay Farms
KENNETH W. ABLE*, DAVID M. NEMERSON†, and THOMAS M. GROTHUES
Marine Field Station, Institute of Marine and Coastal Sciences, Rutgers University, 800 c/o 132Great Bay Boulevard, Tuckerton, New Jersey 08087-2004
ABSTRACT: In a continuing effort to monitor the fish response to marsh restoration (resumed tidal flow, creation ofcreeks), we compared qualitative and quantitative data on species richness, abundance, assemblage structure and growthbetween pre-restoration and post-restoration conditions at two former salt hay farms relative to a reference marsh in themesohaline portion of Delaware Bay. The most extensive comparison, during April–November 1998, sampled fish pop-ulations in large marsh creeks with otter trawls and in small marsh creeks with weirs. Species richness and abundanceincreased dramatically after restoration. Subsequent comparisons indicated that fish size, assemblage structure, andgrowth of one of the dominant species, Micropogonias undulatus, was similar between reference and restored marshes 1and 2 yr post-restoration. Total fish abundance and abundance of the dominant species was greater, often by an orderof magnitude, in one of the older restored sites (2 yr post-restoration), while the other restored site (1 yr post-restoration)had values similar to the reference marsh. The success of the restoration at the time of this study suggests that returnof the tidal flow and increased marsh area and edge in intertidal and subtidal creeks relative to the former salt hayfarms contributed to the quick response of resident and transient young-of-the-year fishes.
IntroductionIt is well known that salt marshes serve as im-
portant habitat for marine fishes and invertebrates(Weinstein 1979; Rountree and Able 1992; Kneib1997; Deegan et al. 2000; Minello et al. 2003).Threats to salt marshes (Thayer 1992; Dionne etal. 1999; Minton 1999; Thomas 1999) have causedconcern for the long-term health of fish speciesthat use them (Boesch and Turner 1984) and haveraised interest in the restoration and creation ofsalt marshes expressly to increase fish production(Seneca and Broome 1992; Minello et al. 1994;Zedler and Lindig-Cisneros 2000; Zedler 2001).
Delaware Bay, one of the largest estuaries on theEast Coast of the United States, is currently the siteof a 4,050 ha salt marsh restoration project aimedat increasing fish production in the bay to offsetthe potential for any adverse effects on fish popu-lations caused by operations at Public Service En-terprise Group’s (PSEG) Salem Generating Sta-tion. Much of the restoration effort is designed torestore diked salt hay farms to normal salt marshstructure and function (Weinstein et al. 1997,2000, 2001). Human activities affecting the floraand fauna of Delaware Bay over the last 200 yr (Se-bold 1992) include water level regulation, bulk-heading, diking, and alteration of marsh plant
* Corresponding author; tele: 609/296-5260 x230; fax: 609/296-1024; e-mail: [email protected]
† Current Address: National Aquarium in Baltimore, Pier 3/501 E. Pratt Street, Baltimore, Maryland 21202-3194.
communities. The few studies that have examinedjuvenile fish use of Delaware Bay marshes (as dis-tinct from open-water habitats in the bay proper)generally concluded that marshes serve as habitatto a number of economically and ecologically im-portant species. Collections in low-salinity saltmarsh creeks characterized juvenile fish assemblag-es, and recorded 55 species representing 43 genera(Smith 1971). Another analysis synthesized the in-formation available for the first year in the life ofestuarine species in the Middle Atlantic Bight, in-cluding several of the dominant forms that usemarshes in Delaware Bay (Able and Fahay 1998).The changes in salt marsh fish species compositionacross the salinity gradient have been examined(Able et al. 2001). A few studies address responseof fishes to salt marsh restoration. In the restora-tion addressed here, construction of a network ofchannels or creeks opened sites to tidal circulationin former salt hay farms (Weinstein et al. 1997,2000) with the long-term goal of reestablishmentof Spartina alterniflora-dominated communities anda concomitant restoration of fish populations.
We evaluate the status of salt hay farm restora-tions by comparing different measures of habitatquality in the context of Essential Fish Habitat—i.e., those waters and substrate necessary for spawn-ing, breeding, feeding, or growth to maturity (Able1999; Schmitten 1999) for the dominant fishes inrestored and reference marshes. We compare pre(1995) and post (1998) restoration measures (spe-cies composition and relative abundance) and

Fish Response to Salt Marsh Restoration 59
Fig. 1. Sampling locations for restored (Commercial and Dennis townships) and reference (Moores Beach) marshes in the me-sohaline portion of lower Delaware Bay.
post-restoration quantitative measures (fish speciesabundance, composition, and growth) across hab-itat types (large and small marsh creeks) in tworestored and one reference marsh over the studyperiod from April–November 1998. A prior analy-sis addressed preliminary findings over 18 mo ofmonitoring at one of these restored sites and onereference site (Able et al. 2000). Another set ofobservations examined habitat use, movements,growth, and production of Fundulus heteroclitus atone of the restored sites (Teo and Able 2003a,b),while another study determined the response ofCallinectes sapidus at the same restored and refer-ence sites as in this study ( Jivoff and Able 2003).We continue to follow the fish response to the res-toration in order to determine if the prior respons-es are persistent features, an important step to ef-fective evaluation of restoration success (Senecaand Broome 1992; Simenstad and Thom 1996;Zedler and Lindig-Cisneros 2000; Zedler 2001).
Materials and Methods
STUDY SITES
Salt hay farm sites at Dennis and Commercialtownships (Fig. 1) were subject to ditching and dik-ing aimed at controlling the frequency and dura-tion of tidal inundation to promote the growth ofsalt hay (Spartina patens), an agricultural crop.There was no regular tidal flow and little water onthe marsh surface except during the winter whenthe dikes were opened to flood the marsh for ashort period. At each of the restored marshes,creeks were created as part of the restoration. Cre-ated creeks were typically 700 to 1,300 m long fromtheir intersection with the nearest natural creek tothe upper limit of the created creek. The mainchannels were 500 to 700 m in length with 1 or 2branches of 100 to 400 m length. Each channel was6 to 10 m in width from bank to bank, and 2 to 3m deep at high tide. Creek dimensions are withinthe range of those encountered for natural creeks

60 K. W. Able et al.
in Delaware Bay and are similar to those in thereference creeks. Some of the linear ditches thatwere still present in portions of each salt hay farmwere incorporated into the new flood-drain systemwhen the dikes were opened during the restora-tion process. The series of created creeks and ex-isting ditches in the restored marshes were de-signed to provide the proper hydroperiod for re-vegetation by Spartina alterniflora (Weinstein et al.1997, 2001).
A portion of the reference marsh at MooresBeach (Fig. 1) was an operating salt hay farm untilstorms breached the perimeter dikes in 1979 andtidal flow was restored (Talbot et al. 1986). In theensuing years regular marsh vegetation, dominatedby Spartina alterniflora, returned to the site. The521 ha reference site is composed of an upper areawith natural creek drainage density (UpperMoores Beach) and a lower area (Lower MooresBeach) that was part of the original salt hay farm.All data collected from both upper and lowerMoores Beach have been combined here. Both thereference marshes at Moores Beach and the re-stored marsh at Dennis Township are bordered bylarger, natural creeks that connect the marshes toDelaware Bay. At Commercial Township the creat-ed creeks are connected directly to the bay.
In order to make pre-restoration versus post-res-toration comparisons, it was essential to identifythe point in time when the sites were restored. Thispoint occurred when dikes were breached and tid-al flow was restored to the marsh via the newly cre-ated channels. Breaching occurred in August 1996at Dennis Township, and all samples collected afterAugust 1996 at this site are considered post-resto-ration. At the Commercial Township restorationsite, one dike was breached in March 1997 and asecond in June 1997. We consider all collections in1998 to be post-restoration. We designated MooresBeach (see Able et al. 2000 for details) as a refer-ence site due to its close proximity (Fig. 1) andsimilar creek morphometry to the restored marsh-es, and due to its relatively natural condition.
SAMPLING TECHNIQUES
Prior to restoration at Commercial Township,fishes were sampled with traps and seines duringMay–August 1995. The habitats sampled were pre-viously non-tidal ditches typical of salt hay farms(Talbot et al. 1986). To make extensive post-res-toration comparisons, the two restoration (Dennisand Commercial townships) and one reference site(Moores Beach), all located on the New Jersey sideof Delaware Bay (Fig. 1), were sampled monthly,April–November in 1998. Following restoration,large marsh creeks were sampled using a 4.9 motter trawl with 6 mm cod end mesh at two stations
in two creeks at each site. At the Moores Beachreference site, the stations were located in creeksof the same approximate width and depth as thecreated creeks at the restored marshes, i.e., creeksfrom 5 to 10 m wide. Start and end points for eachtrawl were recorded using Global Positioning Sys-tem (GPS) coordinates to ensure that identical ar-eas were sampled each month. Sampling tookplace around high tide, and consisted of four 2-min tows per station. All tows were made againstthe current and at a constant engine RPM of 2,500.Depth was measured at each site using a Hummin-bird Wideye depth recorder. The scope of tow lineto water depth was usually maintained at 5:1. Mi-nor adjustments were occasionally made to com-pensate for current speed and tidal flow. Tows ter-minated early due to obstructions in the samplingpath were eliminated from analyses if less than 1.5min. All fish were identified and enumerated. Fish-es to be measured were randomly selected fromwithin species and measured separately to thenearest millimeter for each replicate tow (up to 20indiv of each species). Fork length (FL) was re-corded for fish species with forked tails; totallengths (TL) were recorded for all other fish.
Small intertidal marsh creeks were sampled fol-lowing restoration using weirs (2.0 m 3 1.5 m 31.5 m, with 5.0 m 3 1.5 m wings, 6.0 mm mesh)set at high tide and recovered at low tide, approx-imately six hrs later. At each intertidal creek sam-pled, a net was stretched across the channel withsupport poles embedded vertically in the sediment.Wings were extended onto the marsh surface fromeach end of the net to form a funnel-shaped weir.The bottom of the net was buried in the sedimentto eliminate gaps in the weir. Local topography oc-casionally prevented the complete draining ofcreeks. Fish remaining in standing pools of waterwere seined into the block net but some escapedbecause they buried in the mud. Predation by wad-ing birds frequently occurred before the net wasretrieved; predation occurred at all sites. As a re-sult, weir catches represent a minimum of fish ac-tually present in the area sampled. Weirs were de-ployed during the day at two intertidal creeks emp-tying into one of the marsh creeks adjacent to anotter trawling site. Weir locations were similar butdifficult to match across all sites, as local differenc-es in creek morphology affected the volume of wa-ter that drained through the weirs. Fish were iden-tified and enumerated, and up to 50 individualsper species per sample were measured, using thesame techniques as for the trawl collections.
In samples from both large and small marshcreeks, small specimens of Morone spp. were iden-tified to the species level (Morone americana or Mo-rone saxatilis) using pterygiophore-interdigitation

Fish Response to Salt Marsh Restoration 61
patterns (Olney et al. 1983). All specimens ,30mm (n 5 190) were x-rayed at 16 KV for 5 minand then species identification was determined bythree independent observers. Specimens that werenot able to be definitively identified to species weredesignated as Morone sp.
Three abundant species were present as multipleyear classes; M. americana, Anchoa mitchilli, andMenidia menidia. In order to provide the best dis-crimination for age classes within species in theassemblage analysis, we created monthly (all sitescombined) length-frequency distributions for eachof these species and examined the distributions formultiple modes indicative of the presence of mul-tiple year classes. In every case, a clear mode rep-resenting the young-of-the-year (YOY) was present,so we divided each of these species into two classes:YOY and age 11. To do so, we examined themonthly length-frequency distributions and visual-ly determined the maximum length for the YOYclass for each species and month and then com-pared them to available monthly size estimates(Able and Fahay 1998). We determined the pro-portion measuring less than or equal to this cutoff(YOY) and greater than the cutoff (age 11) foreach species in each month at each site. We mul-tiplied the total of each species caught in eachmonth at each site by the appropriate proportionsto determine the total YOY and age 11 caught foreach species in a given month and site. The YOYand age 11 classes of the three species were thentreated separately in multivariate analyses. We es-timated the growth rate of M. undulatus using themonthly progression of median lengths divided bythe number of days between sampling events.
Physical and chemical variables were measuredat the end (otter trawls in the large marsh creeks)or beginning (weir samples in the small marshcreeks) of each sample. Temperature, dissolved ox-ygen concentration, and salinity were measuredwith a hand-held salinity, temperature, oxygen me-ter (YSI Model 85), by lowering the probe into thewater and recording surface and bottom values.Water transparency was measured by lowering aSecchi disc in the water column until it was nolonger visible, and recording the correspondingdepth in 0.1 m increments.
Ordination by principal components analysis(PCA; Jongman et al. 1995) reduced fish assem-blage variation among monthly samples at each ofthe three study sites into the four most importantdimensions (principle components). Ordinationwas repeated separately for large (otter trawl) andsmall (weir) marsh creeks. Log(y 1 1) transfor-mation of catch-per-unit-effort-standardized abun-dance (CPUE) kept the most abundant speciesfrom dominating the ordination. Species occur-
ring at very low abundance were removed from theanalysis because we doubted the validity of drawinginferences from their distribution. A minimalabundance cut-off was determined relative to gearand sensitive to the potential seasonal periodicityin fish utilization. In the analysis of weir samples,a species had to occur at a CPUE .1.0 in at leastone marsh type over all months; for trawls, a spe-cies had to occur in at least two sites in any monthor have had a total CPUE not less than 0.14 fishper tow, all tows combined. We performed PCAwith the program group CANOCO (Ter Braak andSmilauer 1998).
Fish assemblages may vary seasonally in accor-dance with larval supply, and a response to specificenvironmental factors could be evident only on aseasonal basis. It is desirable to test differencesamong sites in a 2-way crossed design. Because theabundance of all included species at all three sitesdetermined the amplitude of monthly site samplesalong principal component axes (i.e., samplescores), the scores along any one axis provide aunivariate proxy to multi-species assemblage stateand allow assemblage differences to be easily testedfor differences within or among site and month byunivariate methods, or by multivariate methods byusing scores from more than one principal com-ponent, each weighted by the eigenvalues. We usedthe analysis of similarity (2-way crossed ANOSIM,Clarke and Warwick 1994), which is a non-para-metric analog of ANOVA or MANOVA (dependingon the number of components along which samplescores are considered), to test the null hypothesisthat there were no differences in fish assemblageamong sites (3 classes) or months (8 classes). TheANOSIM provided for significance testing by com-paring the Spearmans’s statistic (Rho) derivedfrom ranking classified sample scores against thedistribution of Rho derived from 999 random(Monte Carlo) permutations of sample scores; thenull hypothesis is rejected when Rho from the sam-ple equals or exceeds 95% of the Rhos generatedby 999 Monte Carlo iterations. The ANOSIM is fa-vored over the MANOVA and ANOVA for this ap-plication because it is more robust to deviationsfrom the assumptions of MANOVA and ANOVA,such as normality. We performed ANOSIM in thePrimer 5 software group.
Results and Discussion
PHYSICAL CHARACTERISTICS
The opening of the former salt hay farm to reg-ular tidal inundations at Dennis (Able et al. 2000)and Commercial townships drastically increasedflooding of the sites via created creeks, connectingditches and the marsh surface. After restoration, a

62 K. W. Able et al.
TA
BL
E1.
Ph
ysic
alan
dsa
mp
ling
char
acte
rist
ics
ofD
elaw
are
Bay
mar
shst
ud
ysi
tes
du
rin
gm
onth
lysa
mp
ling
Ap
ril–
Nov
embe
r19
98.S
eeFi
g.1
for
loca
tion
sof
ind
ivid
ual
site
s.
Site
Typ
eD
ate
Res
tora
tion
Com
ple
ted
Are
a(h
a)
Mea
nD
ista
nce
ofT
raw
ling
Stat
ion
sfr
omB
ay(k
m)
Mea
nD
epth
Ran
geat
Tra
wlin
gSt
atio
ns2
(m)
Mea
nSu
rfac
eSa
linit
y1(‰
)M
ean
Surf
ace
D.O
.1(m
g12
1 )
Mea
nSu
rfac
eT
emp
erat
ure
1
(8C
)M
ean
Secc
hi
Dep
th1
(m)
Den
nis
Tow
nsh
ipM
oore
sB
each
Com
mer
cial
Tow
nsh
ip
Res
tora
tion
Ref
eren
ceR
esto
rati
on
Au
gust
1996
— Mar
ch–J
un
e19
97
227
521
1,61
9
0.97
2.6
2.7
1.5–
2.2
1.7–
2.4
1.7–
2.5
17.0
18.1
17.0
5.7
5.9
6.8
20.2
20.1
20.3
0.3
0.5
0.4
1In
dic
ates
the
salin
ity,
D.O
.,te
mp
erat
ure
,an
dSe
cch
iva
lues
wer
eco
mp
ute
dby
fin
din
ga
mon
thly
aver
age
for
each
par
amet
eran
dth
enav
erag
ing
acro
ssth
em
onth
ssa
mp
led
(Ap
ril–
Nov
embe
r).
2In
dic
ates
valu
esre
por
ted
are
the
min
imu
mto
the
max
imu
md
epth
atea
chtr
awlin
gst
atio
n,
aver
aged
acro
ssth
esi
xtr
awlin
gst
atio
ns
atea
chsi
te.
tidal range of 1–2 m was typical and much of themarsh surface at both sites was consistently floodedon every high tide. Most morphological and waterquality characteristics measured in 1998 were sim-ilar between the restored and reference marshes(Table 1). Sampling depths for otter trawl samplesoverlapped, but distances from the bay at Com-mercial Township and Moores Beach were slightlygreater than at Dennis Township. Average salinity,temperature, water clarity, and dissolved oxygenwere very similar across all sites. These same pa-rameters were also similar in our earlier evalua-tions in 1996 and 1997 at the restored marsh atDennis Township and the reference marsh atMoores Beach (Able et al. 2000).
COMPARISON OF PRE-RESTORATION VERSUS POST-RESTORATION MARSHES
Prior to restoration the fish species diversity andabundance was very low at the Commercial Town-ship salt hay farm. Fishes collected in the dominantaquatic habitats at this site (i.e., standing water inditches and pools) were captured with small meshseines and traps in May–August 1995 (n 5 1,722indiv). These were primarily F. heteroclitus (92.3%)and fewer numbers of Cyprinodon variegatus (2.1%)and Menidia beryllina (2.1%). The first two speciesalso made up the vast majority of individuals col-lected at the Dennis Township site prior to resto-ration (Able et al. 2000), and at other salt hayfarms (Talbot et al. 1986). Other species collectedin the salt hay farm prior to restoration includedAnguilla rostrata, Fundulus luciae, Fundulus diaphan-us, M. menidia, M. beryllina, and Pogonias cromis,with most represented by less than six indiv.
After creek creation and restoration to tidal in-undation at Commercial Township, species rich-ness increased dramatically in 1998 such that a rel-atively diverse assemblage of fish species (n 5 28)was collected with otter trawls and weirs in large(n 5 27 species, n 5 22,475 indiv) and small (n 518 species, n 5 106,398 indiv) marsh creeks (Fig.2). It is difficult to make more detailed diversitybecause of the large-scale alteration of habitats andresulting change in sampling gears prior to andafter restoration but abundances (CPUE) of fishesafter the restoration was great (Tables 2 and 3; Fig.2). The fish response, as measured by quantitativecomparison in fish species composition and abun-dance only 1 yr after restoration, was similar to theresponse at the restored marsh at Dennis Township(Able et al. 2000). The fish response was immedi-ate and dramatic relative to the prerestoration con-dition in both of these former salt hay farms.
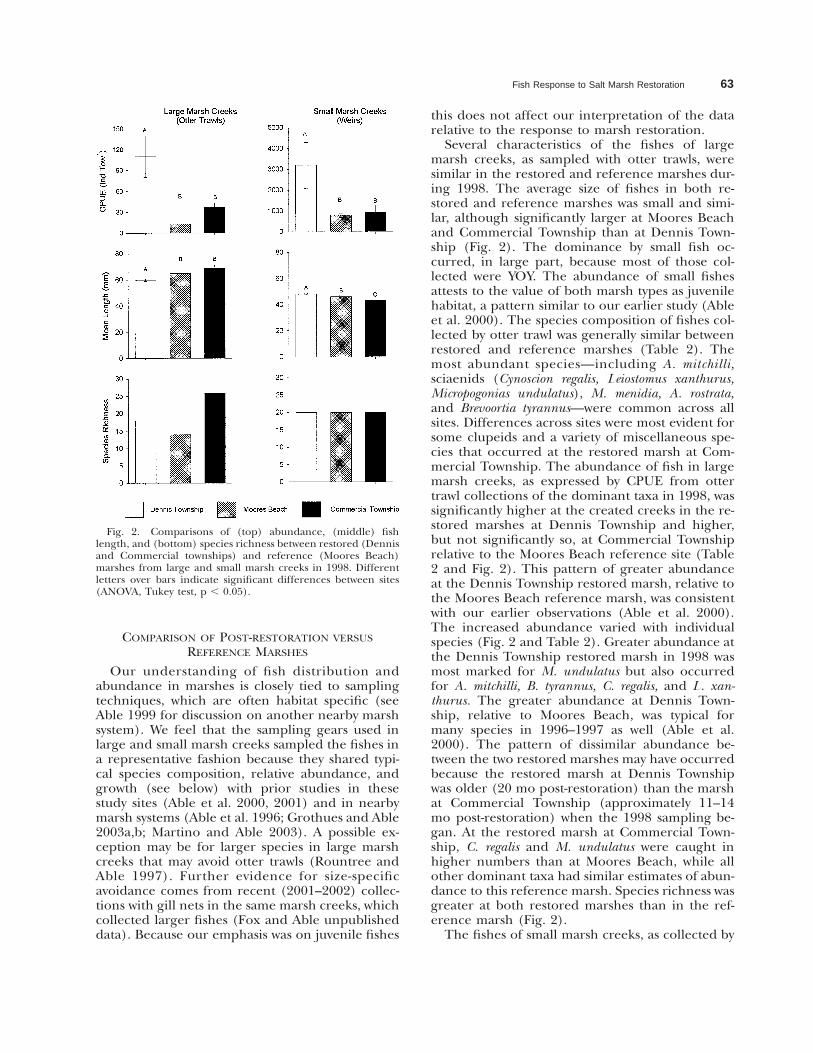
Fish Response to Salt Marsh Restoration 63
Fig. 2. Comparisons of (top) abundance, (middle) fishlength, and (bottom) species richness between restored (Dennisand Commercial townships) and reference (Moores Beach)marshes from large and small marsh creeks in 1998. Differentletters over bars indicate significant differences between sites(ANOVA, Tukey test, p , 0.05).
COMPARISON OF POST-RESTORATION VERSUSREFERENCE MARSHES
Our understanding of fish distribution andabundance in marshes is closely tied to samplingtechniques, which are often habitat specific (seeAble 1999 for discussion on another nearby marshsystem). We feel that the sampling gears used inlarge and small marsh creeks sampled the fishes ina representative fashion because they shared typi-cal species composition, relative abundance, andgrowth (see below) with prior studies in thesestudy sites (Able et al. 2000, 2001) and in nearbymarsh systems (Able et al. 1996; Grothues and Able2003a,b; Martino and Able 2003). A possible ex-ception may be for larger species in large marshcreeks that may avoid otter trawls (Rountree andAble 1997). Further evidence for size-specificavoidance comes from recent (2001–2002) collec-tions with gill nets in the same marsh creeks, whichcollected larger fishes (Fox and Able unpublisheddata). Because our emphasis was on juvenile fishes
this does not affect our interpretation of the datarelative to the response to marsh restoration.
Several characteristics of the fishes of largemarsh creeks, as sampled with otter trawls, weresimilar in the restored and reference marshes dur-ing 1998. The average size of fishes in both re-stored and reference marshes was small and simi-lar, although significantly larger at Moores Beachand Commercial Township than at Dennis Town-ship (Fig. 2). The dominance by small fish oc-curred, in large part, because most of those col-lected were YOY. The abundance of small fishesattests to the value of both marsh types as juvenilehabitat, a pattern similar to our earlier study (Ableet al. 2000). The species composition of fishes col-lected by otter trawl was generally similar betweenrestored and reference marshes (Table 2). Themost abundant species—including A. mitchilli,sciaenids (Cynoscion regalis, Leiostomus xanthurus,Micropogonias undulatus), M. menidia, A. rostrata,and Brevoortia tyrannus—were common across allsites. Differences across sites were most evident forsome clupeids and a variety of miscellaneous spe-cies that occurred at the restored marsh at Com-mercial Township. The abundance of fish in largemarsh creeks, as expressed by CPUE from ottertrawl collections of the dominant taxa in 1998, wassignificantly higher at the created creeks in the re-stored marshes at Dennis Township and higher,but not significantly so, at Commercial Townshiprelative to the Moores Beach reference site (Table2 and Fig. 2). This pattern of greater abundanceat the Dennis Township restored marsh, relative tothe Moores Beach reference marsh, was consistentwith our earlier observations (Able et al. 2000).The increased abundance varied with individualspecies (Fig. 2 and Table 2). Greater abundance atthe Dennis Township restored marsh in 1998 wasmost marked for M. undulatus but also occurredfor A. mitchilli, B. tyrannus, C. regalis, and L. xan-thurus. The greater abundance at Dennis Town-ship, relative to Moores Beach, was typical formany species in 1996–1997 as well (Able et al.2000). The pattern of dissimilar abundance be-tween the two restored marshes may have occurredbecause the restored marsh at Dennis Townshipwas older (20 mo post-restoration) than the marshat Commercial Township (approximately 11–14mo post-restoration) when the 1998 sampling be-gan. At the restored marsh at Commercial Town-ship, C. regalis and M. undulatus were caught inhigher numbers than at Moores Beach, while allother dominant taxa had similar estimates of abun-dance to this reference marsh. Species richness wasgreater at both restored marshes than in the ref-erence marsh (Fig. 2).
The fishes of small marsh creeks, as collected by
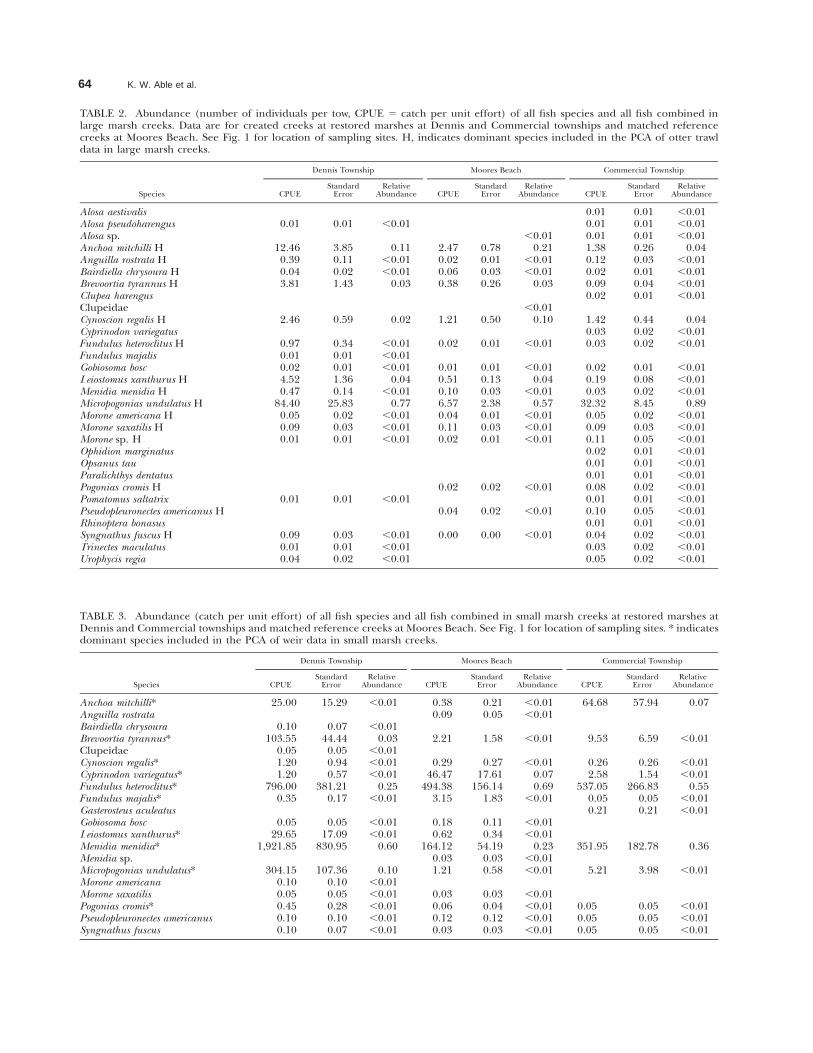
64 K. W. Able et al.
TABLE 2. Abundance (number of individuals per tow, CPUE 5 catch per unit effort) of all fish species and all fish combined inlarge marsh creeks. Data are for created creeks at restored marshes at Dennis and Commercial townships and matched referencecreeks at Moores Beach. See Fig. 1 for location of sampling sites. H, indicates dominant species included in the PCA of otter trawldata in large marsh creeks.
Species
Dennis Township
CPUEStandard
ErrorRelative
Abundance
Moores Beach
CPUEStandard
ErrorRelative
Abundance
Commercial Township
CPUEStandard
ErrorRelative
Abundance
Alosa aestivalisAlosa pseudoharengusAlosa sp.Anchoa mitchilli HAnguilla rostrata H
0.01
12.460.39
0.01
3.850.11
,0.01
0.11,0.01
2.470.02
0.780.01
,0.010.21
,0.01
0.010.010.011.380.12
0.010.010.010.260.03
,0.01,0.01,0.01
0.04,0.01
Bairdiella chrysoura HBrevoortia tyrannus HClupea harengusClupeidaeCynoscion regalis HCyprinodon variegatus
0.043.81
2.46
0.021.43
0.59
,0.010.03
0.02
0.060.38
1.21
0.030.26
0.50
,0.010.03
,0.010.10
0.020.090.02
1.420.03
0.010.040.01
0.440.02
,0.01,0.01,0.01
0.04,0.01
Fundulus heteroclitus HFundulus majalisGobiosoma boscLeiostomus xanthurus HMenidia menidia H
0.970.010.024.520.47
0.340.010.011.360.14
,0.01,0.01,0.01
0.04,0.01
0.02
0.010.510.10
0.01
0.010.130.03
,0.01
,0.010.04
,0.01
0.03
0.020.190.03
0.02
0.010.080.02
,0.01
,0.01,0.01,0.01
Micropogonias undulatus HMorone americana HMorone saxatilis HMorone sp. HOphidion marginatus
84.400.050.090.01
25.830.020.030.01
0.77,0.01,0.01,0.01
6.570.040.110.02
2.380.010.030.01
0.57,0.01,0.01,0.01
32.320.050.090.110.02
8.450.020.030.050.01
0.89,0.01,0.01,0.01,0.01
Opsanus tauParalichthys dentatusPogonias cromis HPomatomus saltatrixPseudopleuronectes americanus HRhinoptera bonasus
0.01 0.01 ,0.010.02
0.04
0.02
0.02
,0.01
,0.01
0.010.010.080.010.100.01
0.010.010.020.010.050.01
,0.01,0.01,0.01,0.01,0.01,0.01
Syngnathus fuscus HTrinectes maculatusUrophycis regia
0.090.010.04
0.030.010.02
,0.01,0.01,0.01
0.00 0.00 ,0.01 0.040.030.05
0.020.020.02
,0.01,0.01,0.01
TABLE 3. Abundance (catch per unit effort) of all fish species and all fish combined in small marsh creeks at restored marshes atDennis and Commercial townships and matched reference creeks at Moores Beach. See Fig. 1 for location of sampling sites. * indicatesdominant species included in the PCA of weir data in small marsh creeks.
Species
Dennis Township
CPUEStandard
ErrorRelative
Abundance
Moores Beach
CPUEStandard
ErrorRelative
Abundance
Commercial Township
CPUEStandard
ErrorRelative
Abundance
Anchoa mitchilli*Anguilla rostrataBairdiella chrysouraBrevoortia tyrannus*Clupeidae
25.00
0.10103.55
0.05
15.29
0.0744.440.05
,0.01
,0.010.03
,0.01
0.380.09
2.21
0.210.05
1.58
,0.01,0.01
,0.01
64.68
9.53
57.94
6.59
0.07
,0.01
Cynoscion regalis*Cyprinodon variegatus*Fundulus heteroclitus*Fundulus majalis*Gasterosteus aculeatusGobiosoma bosc
1.201.20
796.000.35
0.05
0.940.57
381.210.17
0.05
,0.01,0.01
0.25,0.01
,0.01
0.2946.47
494.383.15
0.18
0.2717.61
156.141.83
0.11
,0.010.070.69
,0.01
,0.01
0.262.58
537.050.050.21
0.261.54
266.830.050.21
,0.01,0.01
0.55,0.01,0.01
Leiostomus xanthurus*Menidia menidia*Menidia sp.Micropogonias undulatus*Morone americana
29.651,921.85
304.150.10
17.09830.95
107.360.10
,0.010.60
0.10,0.01
0.62164.12
0.031.21
0.3454.190.030.58
,0.010.23
,0.01,0.01
351.95
5.21
182.78
3.98
0.36
,0.01
Morone saxatilisPogonias cromis*Pseudopleuronectes americanusSyngnathus fuscus
0.050.450.100.10
0.050.280.100.07
,0.01,0.01,0.01,0.01
0.030.060.120.03
0.030.040.120.03
,0.01,0.01,0.01,0.01
0.050.050.05
0.050.050.05
,0.01,0.01,0.01

Fish Response to Salt Marsh Restoration 65
Fig. 3. Comparison of fish assemblage at small marsh creeksat the Dennis Township, Moores Beach, and Commercial Town-ship sites based on PCA of weir collections in 1998. Ordinationspace is the same for both sites (top) and species (bottom).Axes are scaled for optimal depiction of sample similarity. Num-bers in top refer to sampling month. A zero after species namein bottom indicates YOY; a 1 indicates age 1+, if no designationall year classes are combined.
weirs, shared many characteristics among restoredand reference marshes and differed in others (Fig.2 and Table 3). While there were statistically sig-nificant differences in size of fish among all marshtypes, the absolute difference was small. Fishes insmall marsh creeks averaged somewhat smallerthan those in large marsh creeks (Fig. 2). Speciesrichness in small marsh creeks was identical acrossall restored and reference marshes (Fig. 2) andgenerally similar to that in large marsh creeks. Thespecies composition in the small marsh creeks wassimilar for the dominant species such as A. mitchilli,B. tyrannus, F. heteroclitus, M. menidia, and M. un-dulatus (Table 3). Differences in species composi-tion were most evident for some of the less abun-dant species. A pattern of greater abundance insmall marsh creeks in the restored marsh at DennisTownship relative to the restored marsh at Com-mercial Township and the reference marsh atMoores Beach, as observed in this study (Fig. 2),was also observed in previous sampling in 1996–1997 (Able et al. 2000). The dominant fishes insmall marsh creeks at the restored sites were moreabundant than in the reference marsh (Table 3).B. tyrannus, F. heteroclitus, M. menidia, M. undulatus,C. regalis, and L. xanthurus were all most abundantat the Dennis Township site. Only A. mitchilli wasmore abundant at Commercial Township than atDennis Township. The only species that was moreabundant at the reference marsh than at the tworestored sites was C. variegatus.
In a further attempt to evaluate the success ofthe marsh restoration at former salt hay farms, wecompared the fish assemblage structure across re-stored and reference marshes. Large and smallmarsh creeks were analyzed independently (Figs. 3and 4). Of 36 species encountered during sam-pling of large marsh creeks, we retained 17 foranalysis (Tables 2 and 3). We retained 13 of the 23species collected in small marsh creeks weirs (Ta-bles 2 and 3).
Month and site had similarly strong and signifi-cant effects on the assemblage of both small andlarge marsh creeks as explained by PCA, leadingto rejection of all null hypotheses. The site effectwas stronger than the month effect in both largeand small marsh creeks (ANOSIM for weirs Rho 50.732, p 5 0.01 site effect, Rho 5 0.474, p 5 0.01mo effect; for trawls Rho 5 0.732, p 5 0.02 siteeffect, Rho 5 0.457, p 5 0.001 mo effect). Bothmarsh creek types are characterized by an increasein the abundance of individual species beginningfrom a low in early spring. In small marsh creeks,assemblages were most differentiated along bothof the first two principal components (together ex-plaining 68% of the variation) by separation of pri-marily Dennis Township samples and secondarily
Commercial Township samples in all months fromthose of the Moores Beach reference site (Fig. 3top) by an increased abundance, especially of tran-sient species (Fig. 3 bottom). The abundant resi-dent F. heteroclitus contributed relatively little to var-

66 K. W. Able et al.
Fig. 4. Comparison of fish assemblages at large marsh creeksat the Dennis Township, Moores Beach, and Commercial Town-ship site based on PCA of otter trawl collections in 1998. Or-dination space is the same for both sites (top) and species (bot-tom). Axes are scaled for optimal depiction of sample similarity.Numbers in A refer to sampling month. A zero after speciesname in B indicates YOY; a 1 indicates age 1+, if no designationall year classes are combined. Coordinates for Syngnathus fuscus(not shown) coincide closely with those for Leiostomus xanthurusand Anguilla rostrata.
TABLE 4. Growth rate of Micropogonias undulatus based on progression of monthly median lengths for individuals caught in 1998 atDelaware Bay marsh sampling sites.
SiteSite
StatusDays
Elapsed
May
Length(mm) n
August
Length(mm) n
GrowthRate
(mm/day)
Dennis TownshipCommercial TownshipMoores Beach
RestoredRestoredReference
909091
47.052.545.0
27819843
123.0132.0122.0
12811
105
0.870.840.85
iation while the transients, as represented by adultM. menidia and A. mitchilli, and YOY M. menidia andM. undulatus, most strongly determined variation.In general, the adults of these species (except M.undulatus) were more prevalent in spring at therestored marshes at Dennis and Commercial town-ships, while their YOY dominated late in the sea-son. Young-of-the-year M. undulatus, together withF. heteroclitus and YOY of C. regalis, M. menidia andA. mitchilli, weakly discerned summer DennisTownship collection variability (the first principlecomponent factor explained 40.7% of the vari-ance, while the second, which was influencedheavily by F. heteroclitus, explained 27.4%).
Some transient species occurred in large marshcreeks in a similar seasonal pattern as for smallmarsh creeks, and together with other mostly tran-sient species, drove a similar seasonal variation(Fig. 4). The strong M. undulatus influx to all sitesin October served to minimize any site distinction
in that month, as did a relative lack of all speciesother than M. menidia and B. chrysoura in August.Otherwise, Dennis Township was again distin-guished from the similarly constituted CommercialTownship and Moores Beach sites by a relativelygreater abundance of most species in the monthsduring which they occurred, essentially magnifyingthe seasonal effects evident at other sites.
Seasonally spawned recruits, returning juvenilesfrom the previous fall, and large adult predatorsuse large marsh creeks seasonally as dictated byspawning and migration times and by permissivetemperature (Able and Fahay 1998; Able et al.2000, 2001; Tupper and Able 2000). YOY M. un-dulatus, M. saxatilis, and Anchoa sp., as well as allL. xanthurus, A. rostrata, B. tyrannus, and Syngnathusfuscus also accounted for strong seasonal and weaksite separation, especially at Dennis Township inthe spring (Fig. 4). Both YOY M. undulatus and C.regalis drove variability along the second axisamong large marsh creeks, but opposed each oth-er in distribution without discernable regard to siteor month. Assemblages varied more at the DennisTownship site over time (evident as distributionalong the first axis) than at the Commercial Town-ship restored site and the Moores Beach referencesite; it is possible that correlates to time since res-toration (e.g., creek profile or slope) make thesemore or less important to specific transient fishesor that some other as yet unidentified factors in-fluence fish use of the Dennis Township site.
GROWTH
We compared the growth of one of the domi-nant species (M. undulatus; Tables 2 and 3), in re-stored and reference marshes as an independentmeasure of habitat quality and Essential Fish Hab-itat (see Able 1999) in the comparison of referenceand restored marshes (Table 4). Species that im-migrate completely into the study sites over a shortperiod, have strong site fidelity, and simultaneousemigration make the best candidates for modalprogression growth analysis. M. undulatus ap-peared to conform to these criteria. In 1998, age1 individuals grew at very similar rates at all threesites (0.84 to 0.87 mm d21), based on the progres-sion of median lengths between the May and Au-

Fish Response to Salt Marsh Restoration 67
gust collecting events. In other studies of this samespecies, using tag/recapture with coded wire tags,the summer growth rates in 1998 were very similarto each other; at Dennis Township they averaged0.69 mm d21 and at Moores Beach they averaged0.63 mm d21 (Miller and Able 2002). Throughoutthe summer and continuing into October, this spe-cies averaged 5–15 mm longer at the restoredCommercial Township site compared to the othertwo sites. This difference, if it reflects differencesin habitat quality, may not be biologically signifi-cant unless there is a strong size-selective overwin-tering effect. Regardless, with growth as a measureof habitat quality, the restored marshes are at leastcomparable to the reference marsh.
In general, the response of fishes at the largeand small marsh creeks at the recent marsh res-toration at Commercial Township was immediatewith no important differences in species composi-tion, abundance and growth relative to the refer-ence marsh. This pattern of immediate responsewas identical to that at Dennis Township in an ear-lier study (Able et al. 2000). The positive responseat both of the restored marshes may have been duein part to adequate larval supply at these locations,a factor not often considered when marshes arecreated in an inappropriate portion of the land-scape (see Simenstad et al. 2000 for other exam-ples of the need for a landscape scale approach torestoration). The fast response of fishes to marshrestoration, as observed in this study, was consis-tent with the fastest responses observed in Gulf ofMaine marshes (1–5 yr) that were restored aftertidal restrictions were removed (Dionne et al.1999).
Marsh restoration at former salt hay farms inDelaware Bay included several factors that may ac-count for the positive fish response. The return oftidal flow to the marshes accounts for the dramaticincrease in the number of transient fish speciesthat once again have access to the marsh. A similarresponse is apparent by blue crabs, C. sapidus ( Ji-voff and Able 2003). There are also large amountsof intertidal and subtidal edge in these restoredmarshes because of the creation of large creeksand the connection of these creeks to existingditches from the former salt hay farm. This is con-sistent with the recognition that these edges arethe sites of extensive secondary production by fish-es (Wiegert and Pomeroy 1981; Minello et al. 1994;Peterson and Turner 1994; Minton 1999). Thesemarsh habitats are important to productive (abun-dant) resident species that reproduce and grow inthe marsh such as F. heteroclitus (Teo and Able2003a,b), which in turn provide prey for other spe-cies (Nemerson and Able 2003) as occurs in nat-ural marsh systems (Kneib 1997). The YOY of an-
other species, M. undulatus, are abundant, residentin the created creeks at Dennis Township for muchof the summer, and have high growth rates there(this study; Miller and Able 2002). As a result, theycan consume relatively large amounts of F. hetero-clitus and thus transfer this source of marsh pro-duction to deeper waters as they get larger andleave the marshes (Miller et al. 2003); this furtherdemonstrates the value of these restored marshes.While most of this evaluation is based on YOY fish-es, it appears that a large predator, M. saxatilis, inthe restored marsh at Dennis Township and thereference marsh at Moores Beach has similar dis-tributions and tide-based behaviors and uses simi-lar small marsh fishes and crustaceans as prey(Tupper and Able 2000; Nemerson and Able2003).
As a result of the above studies, we have a clearerunderstanding of how marsh restoration contrib-utes to fish habitat quality, in general, and specifi-cally to Essential Fish Habitat. One approach toevaluation of Essential Fish Habitat has been toidentify levels of increasingly complex types of fishresponse that may be used to address issues of hab-itat quality (Able 1999). Level 1 is the simplest andis based on presence and absence data. We used aLevel 1 approach in the comparison of preresto-ration and post-restoration marshes and thesecomparisons clearly indicated that species richnesswas much greater after restoration in both restoredmarshes. Level 2 information is based on habitatspecific abundance and our examination clearlydemonstrates fish abundance for individual specieswas typically equal to, or greater, in both the re-stored marshes relative to the reference marsh.Further analysis of relative abundance by exami-nation of fish assemblage structure also deter-mined that the assemblages were similar among re-stored and reference marshes. We attempted tobring Level 3 information—i.e., habitat-relatedmeasures of growth—to this comparison; in thatcase, growth of Age 1 M. undulatus was similar be-tween restored and reference marshes. All of themeasures of habitat quality used in our evaluationto date suggest that the restored salt marshes shareselected structural and functional characteristics ofthe reference marsh and thus the restoration isproceeding well. While our analysis is promising,future efforts should focus even more on function-al significance of the restored marshes over moreyears in order to completely address the success ofthe restoration relative to fish habitat quality (Able1999; Minello 1999; Zedler and Lindig-Cisneros2000; Swamy et al. 2002).
ACKNOWLEDGMENTS
Numerous individuals from the Rutgers University MarineField Station participated in the field sampling or helped with

68 K. W. Able et al.
data analysis. We would particularly like to thank R. Bush, B.Lemasson, S.L.H. Teo, and J. Chitty. J. Balletto, K. Strait, and J.Klein provided background information and logistical support.Financial support was provided by the Estuary EnhancementProgram of Public Service Enterprise Group. This is Contribu-tion No. 2004-2 from the Rutgers University Institute of Marineand Coastal Sciences.
LITERATURE CITED
ABLE, K. W. 1999. Measures of juvenile fish habitat quality: Ex-amples from a National Estuarine Research Reserve, p. 134–147. In L. R. Benaka (ed.), Fish Habitat: Essential Fish Habitatand Rehabilitation. American Fisheries Society, Symposium22, Bethesda, Maryland.
ABLE, K. W. AND M. P. FAHAY. 1998. The First Year in the Lifeof Estuarine Fishes in the Middle Atlantic Bight. Rutgers Uni-versity Press, New Brunswick, New Jersey.
ABLE, K. W., D. M. NEMERSON, R. O. BUSH, AND P. R. LIGHT.2001. Spatial variation in Delaware Bay (U.S.A.) marsh creekfish assemblages. Estuaries 24:441–452.
ABLE, K. W., D. M. NEMERSON, P. R. LIGHT, AND R. O. BUSH.2000. Initial response of fishes to marsh restoration at a for-mer salt hay farm bordering Delaware Bay, p. 749–773. In M.P. Weinstein and D. A. Kreeger (eds.), Concepts and Contro-versies in Tidal Marsh Ecology. Kluwer Academic Publishers,Dordrecht, The Netherlands.
ABLE, K. W., D. A. WITTING, R. S. MCBRIDE, R. A. ROUNTREE,AND K. J. SMITH. 1996. Fishes of polyhaline estuarine shoresin Great Bay—Little Egg Harbor, New Jersey: A case study ofseasonal and habitat influences, p. 335–353. In K. F. Nords-trom and C. T. Roman (eds.), Estuarine Shores: Evolution,Environments and Human Alterations. John Wiley and Sons,Chichester, U.K.
BOESCH, D. F. AND R. E. TURNER. 1984. Dependence of fisheryspecies on salt marshes: The role of food and refuge. Estuaries7:460–468.
CLARKE, K. R. AND R. M. WARWICK. 1994. Similarity-based testingfor community pattern: The 2-way layout with no replication.Marine Biology 118:167–176.
DEEGAN, L. A., J. E. HUGHES, AND R. A. ROUNTREE. 2000. Saltmarsh ecosystem support of marine transient species, p. 333–365. In M. P. Weinstein and D. A. Kreeger (eds.), Conceptsand Controversies in Tidal Marsh Ecology. Kluwer AcademicPublishers, Dordrecht, The Netherlands.
DIONNE, M., F. T. SHORT, AND D. M. BURDICK. 1999. Fish utili-zation of restored, created, and reference salt-marsh habitatin the Gulf of Maine, p. 384–404. In L. R. Benaka (ed.), FishHabitat: Essential Fish Habitat and Rehabilitation. AmericanFisheries Society, Symposium 22, Bethesda, Maryland.
GROTHUES, T. M. AND K. W. ABLE. 2003a. Response of juvenilefish assemblages in tidal salt marsh creeks treated for Phrag-mites removal. Estuaries 26:563–573.
GROTHUES, T. M. AND K. W. ABLE. 2003b. Discerning vegetationand environmental correlates with subtidal marsh fish assem-blage dynamics during Phragmites eradication efforts: Inter-annual trend measures. Estuaries 26:574–586.
JIVOFF, P. R. AND K. W. ABLE. 2003. Evaluating salt marsh resto-ration in Delaware Bay: The response of blue crabs, Callinectessapidus, at former salt hay farms. Estuaries 26:709–719.
JONGMAN, R. G. H., C. J. F. TER BRAAK, AND O. F. R. VAN TON-GEREN. 1995. Landscape Ecology. Cambridge University Press,Cambridge, U.K.
KNEIB, R. T. 1997. The role of tidal marshes in the ecology ofestuarine nekton, p. 163–220. In A. D. Ansell, R. N. Gibson,and M. Barnes (eds.), Oceanography and Marine Biology: AnAnnual Review 1997. UCL Press, London, U.K.
MARTINO, E. AND K. W. ABLE. 2003. Fish assemblages across themarine to low salinity transition zone of a temperate estuary.Estuarine, Coastal and Shelf Science 56:967–985.
MILLER, M. J. AND K. W. ABLE. 2002. Movements and growth oftagged young-of-the-year Atlantic croaker, Micropogonias un-dulatus, in restored and reference marsh creeks in DelawareBay. Journal of Experimental Marine Biology and Ecology 267:15–38.
MILLER, M. J., D. N. NEMERSON, AND K. W. ABLE. 2003. Seasonaldistribution, abundance, and growth of young-of-the-year At-lantic croaker, Micropogonias undulatus, in Delaware Bay andadjacent marshes. Fishery Bulletin 101:100–115.
MINELLO, T. J. 1999. Nekton densities in shallow estuarine hab-itats of Texas and Louisiana and the identification of essentialfish habitat, p. 43–75. In L. R. Benaka (ed.), Fish Habitat:Essential Fish Habitat and Rehabilitation. American FisheriesSociety, Symposium 22, Bethesda, Maryland.
MINELLO, T. J., K. W. ABLE, M. P. WEINSTEIN, AND C. G. HAYS.2003. Salt marshes as nurseries for nekton: Testing hypotheseson density, growth, and survival through meta-analysis. MarineEcology Progress Series 246:39–59.
MINELLO, T. J., R. J. ZIMMERMAN, AND R. MEDINA. 1994. The im-portance of edge for natant macrofauna in a created saltmarsh. Wetlands 14:184–198.
MINTON, M. D. 1999. Fish utilization of restored, created, andreference salt-marsh habitat in the Gulf of Maine, p. 405–420.In L. R. Benaka (ed.), Fish Habitat: Essential Fish Habitat andRehabilitation. American Fisheries Society, Symposium 22, Be-thesda, Maryland.
NEMERSON, D. M. AND K. W. ABLE. 2003. Spatial and temporalpatterns in the distribution and food habits of Morone saxatilis(Walbaum), striped bass, in marsh creeks of Delaware Bay,USA. Fisheries Management and Ecology 20:337–348.
OLNEY, J. E., G. C. GRANT, F. E. SCHULTZ, C. L. COOPER, AND J.HAGEMAN. 1983. Pterygiophore-interdigitation patterns in lar-vae of four Morone species. Transactions of the American FisheriesSociety 112:525–531.
PETERSON, G. W. AND R. E. TURNER. 1994. The value of saltmarsh edge vs. interior as a habitat for fish and decapod crus-taceans in a Louisiana tidal marsh. Estuaries 17:235–262.
ROUNTREE, R. A. AND K. W. ABLE. 1992. Fauna of polyhalinesubtidal marsh creeks in southern New Jersey: Composition,abundance and biomass. Estuaries 15:171–185.
ROUNTREE, R. A. AND K. W. ABLE. 1997. Nocturnal fish use ofNew Jersey marsh creek and adjacent bay shoal habitats. Es-tuarine, Coastal and Shelf Science 44:703–711.
SCHMITTEN, R. A. 1999. Essential fish habitat: Opportunities andchallenges for the next millennium, p. 3–10. In L. R. Benaka(ed.), Fish Habitat: Essential Fish Habitat and Rehabilitation.American Fisheries Society, Symposium 22, Bethesda, Mary-land.
SEBOLD, K. R. 1992. From Marsh to Farm: The Transformationof Coastal New Jersey. New Jersey Coastal Heritage Trail. Na-tional Park Service, U.S. Department of the Interior, Wash-ington, D.C.
SENECA, E. C. AND S. W. BROOME. 1992. Restoring tidal marshesin North Carolina and France, p. 53–78. In G. W. Thayer(ed.), Restoring the Nation’s Marine Environment. MarylandSea Grant Program, College Park, Maryland.
SIMENSTAD, C. A., W. G. HOOD, R. M. THOM, D. A. LEVY, AND D.L. BOTTOM. 2000. Landscape structure and scale constraintson restoring estuarine wetlands for Pacific coast juvenile fish-es, p. 597–630. In M. P. Weinstein and D. A. Kreeger (eds.),Concepts and Controversies in Tidal Marsh Ecology. KluwerAcademic Publishers, Dordrecht, The Netherlands.
SIMENSTAD, C. A. AND R. M. THOM. 1996. Functional equivalencytrajectories of the restored Gog-Le-Hi-Te estuarine wetland.Ecological Applications 6:38–56.
SMITH, B. A. 1971. The fishes of four low-salinity tidal tributariesof the Delaware Bay estuary. M.S. Thesis, Cornell University,Ithaca, New York.
SWAMY, V., P. E. FELL, M. BODY, M. B. KEANEY, M. K. NYAKU, E.

Fish Response to Salt Marsh Restoration 69
C. MCILVAIN, AND A. L. KEEN. 2002. Macroinvertebrate andfish populations in a restored impounded salt marsh 21 yearsafter the reestablishment of tidal flooding. Environmental Man-agement 29:516–530.
TALBOT, C. W., K. W. ABLE, AND J. K. SHISLER. 1986. Fish speciescomposition in New Jersey salt marshes: Effects of marsh al-terations for mosquito control. Transactions of the AmericanFisheries Society 115:269–278.
TEO, S. L. H. AND K. W. ABLE. 2003a. Habitat use and movementof the mummichog (Fundulus heteroclitus) in a restored saltmarsh. Estuaries 26:720–730.
TEO, S. L. H. AND K. W. ABLE. 2003b. Growth and productionof the mummichog (Fundulus heteroclitus) in a restored saltmarsh. Estuaries 26:51–63.
TER BRAAK, C. J. F. AND P. SIMLAUER. 1998. CANOCO for Win-dows: Software for Canonical Community Ordination, version4. Microcomputer Power, Ithaca, New York.
THAYER, G. W. (ED.). 1992. Restoring the Nation’s Marine En-vironment. Maryland Sea Grant College, College Park, Mary-land.
THOMAS, R. G. 1999. Fish habitat and coastal restoration in Lou-isiana, p. 240–251. In L. R. Benaka (ed.), Fish Habitat: Essen-tial Fish Habitat and Rehabilitation. American Fisheries So-ciety, Symposium 22, Bethesda, Maryland.
TUPPER, M. AND K. W. ABLE. 2000. Movements and food habitsof striped bass (Morone saxatilis) in Delaware Bay salt marshes:Comparison of a restored and a reference marsh. Marine Bi-ology 137:1049–1058.
WEINSTEIN, M. P. 1979. Shallow marsh habitats as primary nurs-eries for fish and shellfish, Cape Fear River, North Carolina.Fishery Bulletin 77:339–357.
WEINSTEIN, M. P., J. H. BALLETTO, J. M. TEAL, AND D. F. LUDWIG.1997. Success criteria and adaptive management for a large-scale wetland restoration project. Wetlands Ecology and Man-agement 4:111–127.
WEINSTEIN, M. P., S. Y. LITVIN, K. L. BOSLEY, C. M. FULLER, AND
S. C. WAINRIGHT. 2000. The role of tidal salt marsh as anenergy source for marine transient and resident fishes: A sta-ple isotope approach. Transactions of the American Fisheries So-ciety 129:797–810.
WEINSTEIN, M. P., J. M. TEAL, J. H. BALLETTO, AND K. A. STRAIT.2001. Restoration principles emerging from one of theworld’s largest tidal marsh restoration projects. Wetlands Ecol-ogy and Management 9:387–407.
WIEGERT, R. G. AND L. R. POMEROY. 1981. The salt-marsh eco-system: A synthesis, p. 219–230. In L. R. Pomeroy and R. G.Weigert (eds.), The Ecology of a Salt Marsh. Springer-Verlag,New York.
ZEDLER, J. B. 2001. Handbook for Restoring Tidal Wetlands.CRC Press LLC, Boca Raton, Florida.
ZEDLER, J. B. AND R. LINDIG-CISNEROS. 2000. Functional equiva-lency of restored and natural salt marshes, p. 565–582. In M.P. Weinstein and D. A. Kreeger (eds.), Concepts and Contro-versies in Tidal Marsh Ecology. Kluwer Academic Publishers,Dordrecht, The Netherlands.
SOURCE OF UNPUBLISHED MATERIALS
FOX, D. A. Unpublished Data. Delaware State University, JamesBaker Building Room 9, 1200 N. Dupont Highway, Dover, Del-aware 19901.
Received, January 17, 2003Accepted, May 16, 2003


















