
Ventilation and hypoxic ventilatory response of Tibetan...
21
Ventilation and Hypoxic Ventilatory Response of Tibetan and Aymara High Altitude Natives CYNTHIA M. BEALL, 1 * KINGMAN P. STROHL, 1 JOHN BLANGERO, 2 SARAH WILLIAMS-BLANGERO, 2 LAURA A. ALMASY, 2 MICHAEL J. DECKER, 1 CAROL M. WORTHMAN, 3 MELVYN C. GOLDSTEIN, 1 ENRIQUE VARGAS, 4 MERCEDES VILLENA, 4 RUDY SORIA, 4 ANA MARIA ALARCON, 4 AND CRISTINA GONZALES 4 1 Case Western Reserve University, Cleveland, Ohio 44106-7125 2 Southwest Foundation for Biomedical Research, San Antonio, Texas 78227-0147 3 Emory University, Atlanta, Georgia 30322 4 Instituto Boliviano de Biologia de Altura, La Paz, Bolivia KEY WORDS high altitude adaptation; quantitative genetics; heritability; microevolution ABSTRACT Newcomers acclimatizing to high altitude and adult male Tibetan high altitude natives have increased ventilation relative to sea level natives at sea level. However, Andean and Rocky Mountain high altitude natives have an intermediate level of ventilation lower than that of newcom- ers and Tibetan high altitude natives although generally higher than that of sea level natives at sea level. Because the reason for the relative hypoventila- tion of some high altitude native populations was unknown, a study was designed to describe ventilation from adolescence through old age in samples of Tibetan and Andean high altitude natives and to estimate the relative genetic and environmental influences. This paper compares resting ventila- tion and hypoxic ventilatory response (HVR) of 320 Tibetans 9–82 years of age and 542 Bolivian Aymara 13–94 years of age, native residents at 3,800–4,065 m. Tibetan resting ventilation was roughly 1.5 times higher and Tibetan HVR was roughly double that of Aymara. Greater duration of hypoxia (older age) was not an important source of variation in resting ventilation or HVR in either sample. That is, contrary to previous studies, neither sample acquired hypoventilation in the age ranges under study. Within populations, greater severity of hypoxia (lower percent of oxygen saturation of arterial hemoglo- bin) was associated with slightly higher resting ventilation among Tibetans and lower resting ventilation and HVR among Aymara women, although the associations accounted for just 2–7% of the variation. Between populations, the Tibetan sample was more hypoxic and had higher resting ventilation and HVR. Other systematic environmental contrasts did not appear to elevate Tibetan or depress Aymara ventilation. There was more intrapopulation genetic variation in these traits in the Tibetan than the Aymara sample. Thirty-five percent of the Tibetan, but none of the Aymara, resting ventilation variance was due to genetic differences among individuals. Thirty-one percent of the Tibetan HVR, but just 21% of the Aymara, HVR variance was due to *Correspondence to: Cynthia M. Beall, Ph. D., Department of Anthropology, 238 Mather Memorial Building, Case Western Reserve University, Cleveland, Ohio 44106-7125. E-mail: [email protected] Received 18 September 1996; revised 5 June 1997; accepted 4 August 1997. AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 104:427–447 (1997) r 1997 WILEY-LISS, INC.
Transcript of Ventilation and hypoxic ventilatory response of Tibetan...

Ventilation and Hypoxic Ventilatory Response of Tibetanand Aymara High Altitude Natives
CYNTHIA M. BEALL,1* KINGMAN P. STROHL,1 JOHN BLANGERO,2SARAH WILLIAMS-BLANGERO,2 LAURA A. ALMASY,2MICHAEL J. DECKER,1 CAROL M. WORTHMAN,3MELVYN C. GOLDSTEIN,1 ENRIQUE VARGAS,4MERCEDES VILLENA,4 RUDY SORIA,4 ANA MARIA ALARCON,4AND CRISTINA GONZALES4
1Case Western Reserve University, Cleveland, Ohio 44106-71252Southwest Foundation for Biomedical Research,San Antonio, Texas 78227-01473Emory University, Atlanta, Georgia 303224Instituto Boliviano de Biologia de Altura, La Paz, Bolivia
KEY WORDS high altitude adaptation; quantitative genetics;heritability; microevolution
ABSTRACT Newcomers acclimatizing to high altitude and adult maleTibetan high altitude natives have increased ventilation relative to sea levelnatives at sea level. However, Andean and Rocky Mountain high altitudenatives have an intermediate level of ventilation lower than that of newcom-ers and Tibetan high altitude natives although generally higher than that ofsea level natives at sea level. Because the reason for the relative hypoventila-tion of some high altitude native populations was unknown, a study wasdesigned to describe ventilation from adolescence through old age in samplesof Tibetan and Andean high altitude natives and to estimate the relativegenetic and environmental influences. This paper compares resting ventila-tion and hypoxic ventilatory response (HVR) of 320 Tibetans 9–82 years of ageand 542 Bolivian Aymara 13–94 years of age, native residents at 3,800–4,065m. Tibetan resting ventilation was roughly 1.5 times higher and Tibetan HVRwas roughly double that of Aymara. Greater duration of hypoxia (older age)was not an important source of variation in resting ventilation or HVR ineither sample. That is, contrary to previous studies, neither sample acquiredhypoventilation in the age ranges under study. Within populations, greaterseverity of hypoxia (lower percent of oxygen saturation of arterial hemoglo-bin) was associated with slightly higher resting ventilation among Tibetansand lower resting ventilation and HVR among Aymara women, although theassociations accounted for just 2–7% of the variation. Between populations,the Tibetan sample was more hypoxic and had higher resting ventilation andHVR. Other systematic environmental contrasts did not appear to elevateTibetan or depress Aymara ventilation. There was more intrapopulationgenetic variation in these traits in the Tibetan than the Aymara sample.Thirty-five percent of the Tibetan, but none of the Aymara, resting ventilationvariance was due to genetic differences among individuals. Thirty-one percentof the Tibetan HVR, but just 21% of the Aymara, HVR variance was due to
*Correspondence to: Cynthia M. Beall, Ph. D., Department of Anthropology, 238 Mather Memorial Building, Case Western ReserveUniversity, Cleveland, Ohio 44106-7125. E-mail: [email protected]
Received 18 September 1996; revised 5 June 1997; accepted 4 August 1997.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 104:427–447 (1997)
r 1997 WILEY-LISS, INC.

genetic differences among individuals. Thus there is greater potential forevolutionary change in these traits in the Tibetans. Presently, there are twodifferent ventilation phenotypes among high altitude natives as comparedwith sea level populations at sea level: lifelong sustained high restingventilation and a moderate HVR among Tibetans in contrast with a slightlyelevated resting ventilation and a low HVR among Aymara. Am J PhysAnthropol 104:427–447, 1997. r 1997 Wiley-Liss, Inc.
Increased ventilation in response to hypo-baric hypoxia is a crucial component ofnewcomers’ acclimatization to high altitude(e.g., Lenfant and Sullivan, 1971; West,1982). The puzzling findings that adult highaltitude residents in the South AmericanAndes and North American Rocky Moun-tains had relatively low ventilation com-pared to newcomers led to studies of theirventilatory sensitivity to hypoxia and foundsmall ventilatory response to additional ex-perimentally-induced hypoxia comparedwith low altitude residents (e.g., Chiodi,1957; Severinghaus et al., 1966; Weil et al.,1971). Some evidence suggested that therelative hypoventilation and ventilatory in-sensitivity were gradually acquired afteryears of exposure to hypoxia during child-hood or adulthood. Andean high altitudebabies breathed and responded to experi-mentally-induced hypoxia no differently thansea level babies (Mortola et al., 1990; Mor-tola et al., 1992). Andean children 12 yearsof age and living at 3,900 m and Coloradochildren 9–10 years of age at 3100 m stillresponded normally to experimentally-in-duced hypoxia (Lahiri et al., 1976; Byrne-Quinn et al., 1972). Thus the small adultAndean and Colorado ventilatory responseto experimentally-induced hypoxic stress,referred to as ‘blunting’, was apparentlyacquired between 10 or 12 years of age andyoung adulthood. Adult migrants in theAndes with periods of 3–42 years of resi-dence at 3,990 and 4,515 m (Chiodi, 1957)also hypoventilated relative to newcomers.Adult migrants to 3,100 m in Colorado devel-oped depressed ventilatory response to ex-perimental hypoxia in proportion to theirlength of residence at high altitude (Weil etal., 1971). The inference was that prolongedexposure to hypoxia during childhood oradulthood leads to hypoventilation relative
to newcomers under everyday conditionsand upon exposure to additional experimen-tally-induced hypoxia. Acquired hypoventil-ation appeared to be the single pattern ofventilatory response to prolonged exposureto hypobaric hypoxia because it was re-ported in the Andean high altitude popula-tion with a long microevolutionary history ofresidence, Colorado residents with just 1–3generations of residence, and long-term mi-grants with only years of residence at highaltitude.
However, it was subsequently demon-strated that the ventilation of adult maleTibetans (including Sherpas1) was similar tothat of newcomers (Ge et al., 1994a and b;Hackett et al., 1980; Zhuang et al., 1993).Although there were no data on children,childhood acquisition of reduced ventilationseemed unlikely because adult ventilationwas high. However, there was some evi-dence that an increased level of ventilationmight not be sustained throughout adult-hood. Lower ventilatory response to addi-tional experimentally-induced hypoxia wascorrelated with greater altitude exposure(Hackett et al., 1980) and age (Curran et al.,1995) among Tibetan men. Similarly, mennative to H 4,400 m had a lower ventilatoryresponse to additional experimentally-in-duced hypoxia than residents at 3,658 m(Curran et al., 1995). This was interpretedas due to ‘‘ . . .increased severity of hypoxicexposure due to their lifelong residence at ahigher altitude.’’ (Curran et al., 1995). Theauthors suggested that future studies distin-guish between duration and severity of expo-sure to hypoxia.
The reasons for the Andean-Tibetan con-trasts in level of adult ventilation and re-
1Sherpas migrated from Eastern Tibet to Nepal in the mid-1500s (Stevens, 1993) and are therefore considered as Tibetans.
428 C.M. BEALL ET AL.

sponse to experimentally-induced hypoxiaand apparently different childhood and adultdevelopmental susceptibilities to acquiredhypoventilation were unknown. The possi-bilities included substantive reasons such asdifferences in genes influencing level of ven-tilation, susceptibility to blunting or thetiming of blunting, differences in other envi-ronmental factors influencing ventilation aswell as methodological reasons such as differ-ences in sampling, altitudes of residenceand ages of individuals in previous studies.None of the high altitude populations hadbeen studied across a broad age range andthere were no available data analyzing thegenetic and environmental sources of varia-tion in these traits at high altitude.
This paper presents the results of a studydesigned to provide such information. Itcompares resting ventilation and ventila-tory response to additional experimentally-induced hypoxia (hypoxic ventilatory re-sponse or HVR) in 320 Tibetans 9–82 yearsof age and 542 Bolivian Aymara 13–94 yearsof age, native residents at 3,800–4,065 m.Tibetan resting ventilation was roughly 1.5times higher and Tibetan HVR roughlydouble that of Aymara, confirming previousreports in small samples of men and extend-ing these to females, adolescents, middle-aged and old adults. Neither sample exhib-ited age differences indicative of acquiredhypoventilation in the age ranges studied.More of the variation in both traits could beattributed to genetic factors in the Tibetanthan in the Aymara sample.
MATERIALS AND METHODSPopulation and sample
The study was conducted in rural sites at3,800–4,065 m in the Tibet AutonomousRegion (TAR), China and Bolivia. The Ti-betan study site was Pen-Dri, two ruralagropastoral villages with a population of773 ethnic Tibetans living at 3,800–4,065 min Lhasa Municipal District, TAR. The aver-age barometric pressure was 479 Torr. Eachhousehold was contacted in May, June, Octo-ber or November 1993 in order to invitehousehold members and their biological rela-tives nine years of age and older to partici-pate in a study collecting interview, genea-logical, anthropometric and physiological
data. Ninety-six percent of the householdscontributed one or more participants and68% of those eligible by age participated toyield a total sample of 428 people age 9–82.Age was verified with reference to reportedanimal year of birth which was then trans-lated into the equivalent Western calendaryear. Nearly all were residents of the twocommunities. All were lifelong high altitudenative residents at 3,600 m or higher (withthe exception of one low altitude native whohad lived in the village since 1950).
The Bolivian study site was composed offour dispersed communities in Provincia Mu-rillo, Departamento La Paz, Bolivia with apopulation of 1,175 ethnic Aymara living at3,900–4,000 m. The average barometric pres-sure was 478 Torr. Each household wascontacted between May and August 1994 inorder to invite household members and theirbiological relatives 14 years of age and olderto participate in the study. Age was verifiedby birth certificates or identity cards for 64%of the sample and by reference to historicalevents for a few elderly people. Seventy-seven percent of all households participatedand 57% of the residents eligible by ageparticipated to yield a total sample of 608people age 13–94. Seventy percent of thesample resided in the four communities andthe rest were relatives residing elsewhere(one at low altitude). All were Aymara na-tives (with the exception of one Quechua) ofthis or nearby high altitude communities.
Measurement
Measurements were conducted in indoorfield laboratories established in central loca-tions in the study communities. Demo-graphic, lifestyle and health informationwere obtained by questionnaire in the na-tive language. Andean highlanders wereasked to abstain from coca chewing on theday of the test. Anyone who indicated thathe had chewed coca or had fresh coca leavesin his mouth was asked to return anotherday. Physiological measurements and mostanthropometrics were taken by the firstauthor working with a local assistant. Aftercollection of the interview data, anthropo-metric measurements were taken accordingto standard protocols (Cameron et al., 1981)and then resting percent of oxygen satura-
429VENTILATION AT HIGH ALTITUDE

tion of arterial hemoglobin (SaO2) and pulsewere measured with a Criticare model 5011pulse oximeter. The instrument updates anddisplays SaO2 and pulse every four to sixheartbeats (approximately one to three sec-onds). The noninvasive sensor was placed onthe index finger (occasionally on anotherfinger if a reading were not obtainedpromptly on the index finger or it was miss-ing), the investigator waited ten to fifteenseconds after a reading appeared on theoutput screen and then recorded the SaO2and pulse readings every ten seconds for atotal of six measurements each. The averageof the six measurements is reported for SaO2(Beall and Goldstein, 1990; Beall et al.,1994; Beall et al., 1997) and pulse. Therepeatability of this measure of SaO2, as-sessed by the mean and standard deviationof the difference between measurements ofthe same individuals on two different days(Bland and Altman, 1986) was 11.3 6 2.7%,n 5 22 in the Tibetan and 10.3 6 2.9%, n 522 in the Aymara sample. The repeatabilityof this measure of pulse was 12 6 12 f/min(frequency per minute), n 5 22 in the Ti-betan and 14 6 9 f/min, n 5 22 in theAymara sample. The pulse oximeter read tothe nearest percent of SaO2 and integer ofheart rate.
Individuals who reported that they werenot pregnant and were free of symptoms ofrespiratory disease then provided ventila-tion measurements consisting of a three-minute resting ventilation determination fol-lowed by a three to five minute rest and thenthree HVR determinations separated by tenminute rests. At least an hour had elapsedsince the last meal and the study participanthad been seated for ten to fifteen minutesbefore the beginning of the ventilation mea-surements. A complete set of measurementstook place in the following order: interview,anthropometry, resting pulse and SaO2 (un-attached to the ventilation testing appara-tus), resting ventilation, and three HVRdeterminations.
The following instrumention and physi-ological measurement recordings were usedfor the ventilation determinations. Inspira-tory and expiratory carbon dioxide (CO2)were measured in real time at the mouth-piece by a Biochem Lifespan CO2 monitor
validated with a two point calibration (roomair and a gas of known concentration) everytwo to three study participants or, for part ofthe Aymara sample, a Biochem Capnographvalidated with a one point calibration (roomair) prior to testing each person. SaO2 andpulse were measured continuously with aCriticare 5011 pulse oximeter whose analogoutput was validated by a self-calibrationroutine. Inspiratory and expiratory airflowvelocity and volume and ventilatory fre-quency were measured by a Fleisch pneumo-tachograph connected to a 13.0 cm H2OValidyne pressure transducer that was elec-tronically balanced daily. The voltage outputof the pressure transducer was amplified bya Validyne carrier demodulator and ampli-fier.
The Biochem Lifespan CO2 monitor failedpart way through the Aymara data collec-tion and the Biochem Capnograph was sub-stituted. The latter had only a one pointcalibration capability and therefore the end-tidal CO2 values of the two machines are notcomparable. Only the values obtained withthe Biochem Lifespan are reported here.However, the Biochem Capnograph voltageoutput was used to directly monitor end-tidal CO2 during the ventilation tests.
The ventilation measurements were ad-ministered carefully to seated study partici-pants whose noses were occluded with nose-clips as they breathed through a rubbermouthpiece attached to a large bore, low-resistance Collins valve attached to theFleisch pneumotachograph. The Collinsvalve was open to room air during theresting ventilation determinations and opento the rebreathing circuit during the HVRdeterminations. Large bore, low resistanceone-way valves directed exhaled gas throughthe rebreathing circuit which had a T-piececonnector at the mid-point. A variable speedblower drew a portion of the exhaled gasfrom the T-piece through a cannister contain-ing CO2 absorbing crystals. The blower wasadjusted as needed in order to maintainconstant end-tidal CO2. The exhaled gas(partially cleansed of CO2) was directed intoa seven-liter weather balloon which wasinflated with three liters of room air prior toeach rebreathing trial and emptied betweentrials. As the study participant inspired, gas
430 C.M. BEALL ET AL.

was drawn from the balloon, directedthrough one-way valves through the pneu-motachograph and to the mouthpiece. Thus,the individual rebreathed previously ex-haled gas, some of which had been scrubbedof some CO2, that became increasingly hy-poxic as the oxygen was consumed in theclosed circuit. An external pump infusedsupplemental room air into the rebreathingcircuit to regulate the rate of arterial hemo-globin oxygen desaturation to approximately1 to 2% every 15 seconds.
Isocapnic HVR was measured by a modi-fied Rebuck and Campbell (1974) rebreath-ing technique. The modifications were a)initiation of the rebreathing trial from athree-liter mixture of 5% CO2 and 20–21%O2 (rather than 5–6% CO2 and 24% O2), b)termination of rebreathing trials at 75%SaO2 (rather than 65%) and c) slowing therate of descent of SaO2 by adding supplemen-tal room air. Isocapnia was maintained dur-ing the HVR determinations at or near theindividual’s average end-tidal CO2 duringthe resting ventilation determination. Theaverage and standard deviation of the differ-ence between end-tidal CO2 measured dur-ing the resting ventilation determination.The average and standard deviation of thedifference between end-tidal CO2 measuredduring the resting ventilation and that mea-sured during the HVR determination was20.3 6 2.2 Torr, n 5 303 in the Tibetansample and 10.3 6 2.8 Torr, n 5 149 in theAymara sample. The repeatability of thisdifference was 20.10 6 4.1 Torr, n 5 21 inthe Tibetan sample. The repeatability couldnot be calculated for the Aymara samplebecause the repeat determinations were con-ducted after the change of capnometers. TheCO2 monitors read to the nearest integerTorr.
HVR was calculated on a breath-by-breath basis as the change in ventilationdivided by the change in SaO2 determinedby the regression of ventilation on SaO2(DL/min/D%SaO2). HVR is usually a nega-tive value because SaO2 falls during the testand ventilation usually rises. In the presentpaper HVR values were multiplied by 21 sothat stronger responses have higher values.
A resting ventilation or HVR determina-tion was judged to be technically unaccept-
able for inclusion in the analyses if one ormore of the following occurred: a) less than90 seconds of data were collected for aresting ventilation determination, b) breath-by-breath end-tidal CO2 fluctuated morethan 4 Torr from the baseline, c) the ampli-tude of the airflow signal decreased morethan 40% from one breath to the next (indi-cating pneumotachograph failure), d) SaO2fell abruptly to less than 80% after themouthpiece was in place, e) if breathingwere erratic or f) the investigator stoppedthe measurement due to observed discom-fort of the study participant. Ninety-onepercent of all Tibetan HVR determinationsand 95% of all Aymara determinations weretechnically acceptable. An individual’s HVRwas the average of all his or her technicallyacceptable determinations. Eighty percentof the Tibetan and 91% of the Aymara HVRvalues analyzed were the average of threedeterminations.
The repeatability of the resting ventila-tion determinations was 3.5 6 1.6 L/min,n 5 25 in the Tibetan sample and 20.83 60.43 L/min, n 5 20 in the Aymara sample.The repeatability of the HVR determina-tions was 20.05 6 0.19 DL/min/%SaO2, n 525 in the Tibetan sample and 10.12 6 0.09DL/min/%SaO2, n 5 20 in the Aymara sam-ple. These small differences indicate thatthe measurement techniques had good reli-ability in both sites.
Steps were taken to ensure that studyparticipants were at ease during the ventila-tion testing. The investigators carefully ex-plained and demonstrated the instrumentsand procedures in the native language. Eachstudy participant practiced wearing nose-clips and breathing through a mouthpieceunattached to the Fleisch pneumotacho-graph before practicing on the rebreathingcircuit itself. If he or she did not appear tobreathe comfortably and naturally on therebreathing circuit then the explanations,demonstration, and practice were repeated.Evidence suggesting that the study partici-pants were equally at rest before and duringthe ventilation determinations is providedby pulse and SaO2 measurements taken atthe two times. Resting pulse and SaO2 mea-sured while resting and unattached to the
431VENTILATION AT HIGH ALTITUDE

rebreathing circuit, as described above, dif-fered little from pulse and SaO2 measuredwhile breathing room air with the mouth-piece and noseclips in place during the rest-ing ventilation determination. The averagedifference between pulse measured whilequietly resting and unattached to the re-breathing circuit and pulse measured dur-ing the resting ventilation determinationswas 3.4 6 5.0 f/min, n 5 18 in the Tibetanand 4.2 6 11.9 f/min, n 5 17 in the Aymarasample. The average and standard deviationof the difference between SaO2 measuredwhile resting and unattached to the re-breathing circuit and that maintained whilebreathing with the noseclips and mouth-piece during the three-minute determina-tion of resting ventilation was 10.8% 6 3.4,n 5 298 in the Tibetan sample and 11.2% 62.6, n 5 383 in the Aymara sample.
Additional evidence that the study partici-pants were not particularly anxious aboutthe testing procedure was provided by thegood repeatability of pulse, SaO2, and end-tidal CO2 measured during resting ventila-tion and the repeatability of the restingventilation and HVR measurements. Famil-iarity with the tests gained during the firstmeasurement would have lessened any testassociated anxiety and could have resultedin recording different values during the sec-ond test; however, the readings were stable.The average repeatability of pulse duringthe resting ventilation determination was24.3 6 15.9 f/min, n 5 16 in the Tibetan and16.2 6 10.2 f/min, n 5 13 in the Aymarasample. The average repeatability of theSaO2 maintained during the resting ventila-tion determination was 20.5 6 3.7%, n 5 28in the Tibetan and 20.7 6 4.0, n 5 20 in theAymara sample. The average repeatabilityof the resting ventilation end-tidal CO2 was20.4 6 1.9 Torr, n 5 20 and that duringHVR determination was 10.7 6 7.2 Torr,n 5 20 in the Tibetan sample. Thus we didnot detect physiological variability that couldbe attributed to anxiety or learning (alterna-tively, the study participants learned noth-ing during the first ventilation determina-tions and were equally anxious during thesecond).
Analysis
The analyses were conducted with samplesof 320 Tibetans and 542 Aymara males andfemales 9–94 years of age who reported thatthey were healthy and not pregnant andwho had a technically acceptable restingventilation measurement and/or one or moretechnically acceptable HVR measurementswithin three standard deviations of the re-spective sample mean. Individuals 20 yearsof age and older are considered adults be-cause height growth is completed by then inboth sexes in both populations. Table 1describes the characteristics of Tibetan chil-dren 9–12 years of age, Tibetan and Aymaraadolescents 13–19 years of age and adults 20years of age and older. Body mass index wascalculated as weight in kilograms divided bythe square of height in meters.
Means and standard deviations are re-ported. Bivariate correlations, multiple re-gression analyses, analyses of variance andcovariance, and t-tests address various hy-potheses about resting ventilation and HVR.A significance level of 0.05 is used.
Quantitative genetic analyses tested aseries of hypotheses regarding sources ofvariation in resting ventilation and HVRusing maximum likelihood variance decom-position methods (Hopper and Mathews,1982; Lange and Boehnke, 1983) availablein the computer programs FISHER (Langeet al., 1988). These analyses provided infor-mation regarding the relative importance ofgenetic, shared environmental, and randomenvironmental effects for resting ventilationand HVR variation.
Quantitative resting ventilation HVRvariation was modeled based on the follow-ing linear function for the vector of pheno-types in a pedigree of size n (bold lower caseletters denote vectors and bold upper caseletters denote matrices):
y 5 µln 1 (X 2 lns8)b 1 g 1 h 1 e (1)
where y is the n x 1 vector of phenotypes, µ isthe grand mean of the trait, X is an n x kmatrix containing k covariates, ln is a vectorof n ones, s represents a vector of baselinecovariates (e.g. 0 for qualitative covariatesand x for continuous covariates), b is a k x 1vector of regression coefficients, g is the
432 C.M. BEALL ET AL.

vector of additive genetic values, h is avector of shared household effects, and e is avector of random environmental deviations.Given this model, the expected variance/
covariance matrix for y is written:
Var(y) 5 V 5 2Fsg2 1 Zsd
2 1 Inse2 (2)
where F is the n x n matrix of kinshipcoefficients, Z is an indicator matrix whoseij-th element is 1 if the i-th and j-th individu-als live in the same household and is 0otherwise, and In is an identity matrix oforder n. The variance terms in Eq. (2) in-clude the additive genetic variance (sg
2), thevariance due to shared household effects(sh
2), and the random environmental vari-ance (se
2). Assuming multivariate normalityof y, the likelihood of the pedigree is easilycalculated and optimization methods can beused for parameter estimation. Subsequenthypothesis testing is performed using likeli-hood ratio tests. As implied in Eq. (1), theeffects of potential covariates (such as age,sex, BMI) were simultaneously estimated inall analyses. The effects of covariates weretested using likelihood ratio tests.
Residual heritability (h2) was calculatedas the additive genetic variance/(1-covariatevariance). Residual household variance (c2)was calculated as the shared household ef-fects variance/(1-covariate variance). Thevariance component analyses of resting ven-tilation used 295 individuals in nine pedi-grees (groups of individuals with a commonancestor) from the Tibetan sample and 537individuals in 59 pedigrees from the Aymarasample. The variance component analyses ofHVR used 319 individuals in nine pedigreesfrom the Tibetan sample and 522 individu-als in 58 pedigrees from the Aymara sample.
RESULTS
Figures 1 and 2 illustrate the higher Ti-betan resting ventilation and HVR valuesand wider Tibetan variation throughout theage range. There were no notable age trendsfrom childhood to old age in either sample.Tibetan resting ventilations were about 1.4to 1.7 times higher than the Aymara andHVR were about double theAymara, depend-ing upon age and sex (Table 2). The higherTibetan resting ventilation was associatedwith tidal volumes roughly 1.4 to 1.7 timeshigher than Aymara. Tibetan end-tidal CO2during rest breathing ambient air and dur-ing HVR determinations were 2–6 Torr lowerthan Aymara. End-tidal CO2 during resting
TABLE 1. Characteristics of Tibetan andAymara samples1
Tibetan Aymara
Mean S.D. n Mean S.D. n
MalesChildren, 9–12 yrs
Age 11 1 22 — — —Height, cm 131 8 22 — — —Weight, kg 24.5 36 22 — — —BMI, kg/m2 14.2 0.8 22 — — —Chest width, cm 21.4 1.2 22 — — —Chest depth, cm 15.7 0.9 21 — — —SaO2 91.1 2.3 22 — — —Pulse, f/min 92 14 22 — — —
Adolescents, 13–19 yrsAge 16 2 34 16 2 55Height, cm 154 11 34 153 9 55Weight, kg 39.2 8.5 34 45.4 8.8 54BMI, kg/m2 16.4 1.6 34 19.2 2.2 54Chest width, cm 25.0 1.9 34 26.5 2.1 55Chest depth, cm 18.0 1.3 33 18.2 1.8 55SaO2% 89.7 2.6 34 93.3 2.2 39Pulse, f/min 81 14 33 74 12 55
Adults, 201 yrsAge 38 14 88 41 16 232Height, cm 165 7 87 160 5 229Weight, kg 51.9 6.1 87 59.2 8.5 229BMI, kg/m2 19.1 1.4 87 23.0 2.8 228Chest width, cm 28.0 1.5 86 29.1 1.8 228Chest depth, cm 20.7 1.3 87 21.1 2.0 230SaO2% 88.5 3.3 87 92.2 2.8 167Pulse, f/min 73 13 88 67 11 231
FemalesChildren, 9–12 yrs
Age 11 1 15 — — —Height, cm 128 10 15 — — —Weight, kg 23.1 3.7 15 — — —BMI, kg/m2 14.1 0.7 15 — — —Chest width, cm 20.5 1.1 15 — — —Chest depth, cm 15.1 0.9 15 — — —SaO2% 91.8 3.0 15 — — —Pulse, f/min 93 14 15 — — —
Adolescents, 13–19 yrsAge 16 2 39 16 2 52Height, cm 148 10 39 149 5 52Weight, kg 36.4 8.6 38 47.6 8.8 50BMI, kg/m2 16.4 2.2 38 21.3 3.3 50Chest width, cm 23.8 1.9 39 26.8 1.9 50Chest depth, cm 17.1 1.8 39 17.9 1.7 51SaO2% 89.8 2.5 38 92.9 2.1 36Pulse, f/min 84 12 39 80 11 52
Adults, 201 yrsAge 38 14 122 40 15 212Height, cm 153 5 122 149 5 210Weight, kg 44.9 5.6 115 52.2 8.6 200BMI, kg/m2 19.2 1.8 115 23.6 3.5 198Chest width, cm 25.6 1.6 121 28.1 1.8 208Chest depth, cm 19.3 1.4 120 19.0 1.7 208SaO2% 89.3 3.2 87 91.2 2.9 100Pulse, f/min 77 12 122 73 10 212
1 BMI 5 body mass index, SaO2% 5 percent of oxygen saturationof arterial hemoglobin.
433VENTILATION AT HIGH ALTITUDE

ventilation averaged within one Torr of thatduring HVR determination in both sites.HVR determinations took less time amongthe Tibetans, consistent with starting at alower resting SaO2.
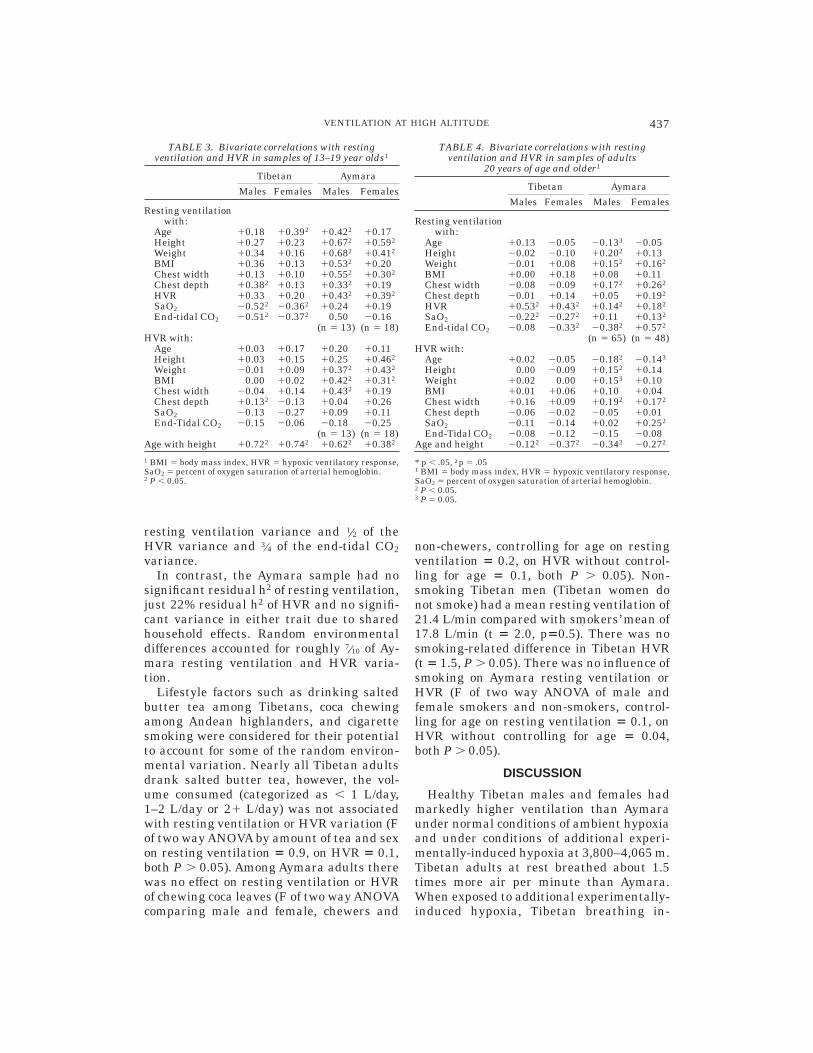
There was no evidence for adolescent ac-quisition of hypoventilation (i.e., no negativeassociation between age and ventilation) ineither sample. Adolescent development ofventilation was analyzed in those 13–19years of age (Table 3). There was a signifi-cant, moderate, positive correlation betweenage and resting ventilation among Tibetanfemales and Aymara males. There was noconsistent pattern of association betweenbody size measured as height, weight, BMI,or chest depth and resting ventilation orHVR among the Tibetan adolescents. Ti-betan males had a significant, moderate,positive association between chest depthand resting ventilation and a significant,small, positive association between chestdepth and HVR. In contrast, there was aconsistent significant, moderate, positive as-sociation between body size measures and
resting ventilation and HVR among theAymara adolescent males and females.
Tibetan adolescents exhibited significant,moderate, negative associations betweenSaO2 and resting ventilation and betweenend-tidal CO2 and resting ventilation, butnot HVR. Aymara adolescents exhibited in-significant associations between the SaO2and resting ventilation in the opposite, posi-tive, direction. There were no significantassociations between end-tidal CO2 and ven-tilation in the Aymara sample, perhaps dueto the small sample size. There was a signifi-cant, moderate, positive correlation betweenresting ventilation and HVR in the Aymarasample.
There was scant evidence for adult acqui-sition of hypoventilation. The only negativeassociation between age and ventilationamong adults was a trend (P 5 0.05) towardlower resting ventilation with age amongAymara men that explained less than 2% ofthe variation and a slight, significant, nega-tive correlation of HVR with age that ex-plained just 3% of the variation (Table 4).
Fig. 1. Resting ventilation (L/min) of Tibetan and Aymara males and females 9–94 years of age.
434 C.M. BEALL ET AL.

There was no association between body sizeand ventilation in Tibetan adults. Height,weight and chest width had small, signifi-cant, positive correlations with resting venti-lation and HVR among the Aymara adults.
Resting ventilation had a small, signifi-cant, negative correlation with SaO2 amongthe Tibetan adults and a small, significant,positive correlation with SaO2 among Ay-mara women. The associations with SaO2account for just 4–7% of the variation inresting ventilation. HVR had a small, signifi-cant, positive association with SaO2 onlyamong Aymara women that explained lessthan 2% of the variation (Table 4).
In both samples, adults with higher HVRhad higher resting ventilation. There was asignificant, moderate, positive correlationamong Tibetans and a significant small posi-tive correlation among Aymara. Individualswith both higher HVR and lower SaO2 wouldbe expected to have higher resting ventila-tion. This hypothesis was explored withmultiple regression analyses. The multipleR2 of age, SaO2 and HVR with resting venti-lation was significant for Tibetan men and
women, but not Aymara (Table 5). The addi-tional contribution of body size variableswas assessed because they were associatedwith ventilation among the Aymara. Heightadded significantly to the explained varia-tion only among Aymara men. Chest widthadded significantly to the explained varia-tion among Aymara women and chest depthdid so among Tibetan women. The multipleR2 for the six variable model accounted formore of the Tibetan than the Aymara varia-tion, yet these traits explained just 10–36%of the resting ventilation variation.
The contribution of genetic factors to rest-ing ventilation and HVR variation in eachsample was evaluated by estimating therelative importance of genetic effects, sharedhousehold environment and random environ-ment effects using maximum likelihood vari-ance decomposition methods to analyze thefamilial patterning of these traits. The re-sidual heritability (h2) of Tibetan restingventilation was 0.317 and of Tibetan HVRwas 0.345 (Table 6). That is, about 1/3 of thevariance in both traits in the Tibetan samplewas attributable to additive genetic factors.
Fig. 2. HVR (DL/min/D%SaO2) of Tibetan and Aymara males and females 9–94 years of age.
435VENTILATION AT HIGH ALTITUDE

There was a significant high genetic correla-tion of 0.67 between Tibetan resting ventila-tion and HVR indicating that some pleiotro-pic genes influence both traits. The residualheritability of Tibetan end-tidal CO2 was
0.218. The residual variance due to sharedhousehold effects (c2) was 0.303 for restingventilation and 0.153 for HVR and 0 forend-tidal CO2. Random environmental differ-ences accounted for roughly 1⁄3 of the Tibetan
TABLE 2. Ventilatory characteristics of Tibetan and Aymara high altitude natives1
Tibetan Aymara t, sitedifferencesMean S.D. n Mean S.D. n
MalesChildren, 9–12 yrs
Resting ventilation, L/min 16.4 7.2 20 — — — —Tidal volume, L/min 1.1 0.5 22 — — — —Breathing frequency, f/min 17.9 4.2 22 — — — —HVR, DL/min/D%SaO2 0.46 0.39 22 — — — —Length of HVR determination, sec 82 32 22 — — — —Resting end-tidal CO2, Torr 32.8 5.2 20 — — — —HVR end-tidal CO2, Torr 32.7 6.1 22 — — — —Resting-HVR difference in end-tidal CO2, Torr 20.03 2.1 20 — — — —
Adolescents, 13–19 yrsResting ventilation, L/min 17.3 8.0 30 12.2 1.8 54 3.52
Tidal volume, L/min 1.1 0.6 30 0.7 0.1 54 3.32
Breathing frequency, f/min 17.8 4.5 31 17.1 2.0 54 0.8HVR, DL/min/D%SaO2 0.84 0.94 34 0.42 0.20 53 2.62
Length of HVR determination, sec 110 47 34 143 37 53 23.72
Resting end-tidal CO2, Torr 29.5 5.2 31 35.2 2.8 13 23.82
HVR end-tidal CO2, Torr 29.2 5.9 34 34.3 2.7 13 24.12
Resting-HVR difference in end-tidal CO2, Torr 20.3 2.1 30 1.0 1.5 13 22.0Adults, 201 yrs
Resting ventilation, L/min 19.7 8.4 84 13.4 2.4 230 6.82
Tidal volume, L/min 1.4 0.6 85 0.9 0.2 222 8.62
Breathing frequency, f/min 14.8 3.4 86 15.5 2.8 225 21.8HVR, DL/min/D%SaO2 0.93 0.85 87 0.45 0.25 222 5.22
Length of HVR determination, sec 136 50 88 194 50 225 29.12
Resting end-tidal CO2, Torr 29.7 4.6 86 33.8 4.1 66 25.82
HVR end-tidal CO2, Torr 30.4 4.7 87 33.3 5.1 67 23.62
Resting-HVR difference in end-tidal CO2 Torr 20.6 2.0 85 0.3 2.4 65 22.62
FemalesChildren, 9–12 yrs
Resting ventilation, L/min 17.5 4.7 15 — — — —Tidal volume, L/min 1.1 0.3 15 — — — —Breathing frequency, f/min 18.5 5.2 15 — — — —HVR, DL/min/D%SaO2 0.60 0.5 15 — — — —Length of HVR determination, sec 79 24 15 — — — —Resting end-tidal CO2, Torr 28.8 8.9 15 — — — —HVR end-tidal CO2, Torr 28.5 8.1 15 — — — —Resting-HVR difference in end-tidal CO2, Torr 10.2 2.6 15 — — — —
Adolescents, 13–19 yrsResting ventilation, L/min 19.2 7.1 36 11.4 1.9 51 6.42
Tidal volume, L/min 1.0 0.5 36 0.7 0.1 51 5.02
Breathing frequency, f/min 19.7 4.1 36 17.9 2.2 50 2.52
HVR, DL/min/D%SaO2 0.76 0.72 38 0.36 .16 49 3.42
Length of HVR determination, sec 105 42 39 146 47 50 24.32
Resting end-tidal CO2, Torr 29.7 5.2 36 32.9 3.6 18 22.42
HVR end-tidal CO2, Torr 29.6 5.3 39 31.6 5.2 19 21.4Resting-HVR difference in end-tidal CO2, Torr 20.3 1.8 36 0.3 0.9 17 21.6
Adults, 201 yrsResting ventilation, L/min 17.7 7.8 111 11.5 2.1 207 8.32
Tidal volume, L/min 1.2 0.5 114 0.7 0.1 203 9.32
Breathing frequency, f/min 16.4 3.1 112 17.0 2.8 204 21.6HVR, DL/min/D%SaO2 0.70 0.59 121 0.37 0.20 197 6.02
Length of HVR determination, sec 131 58 122 156 48 200 23.92
Resting end-tidal CO2, Torr 29.2 5.5 113 34.0 4.5 49 25.52
HVR end-tidal CO2, Torr 29.4 4.9 119 34.0 4.6 51 25.72
Resting-HVR difference in end-tidal CO2, Torr 20.3 2.1 114 0.0 2.0 49 20.81 HVR 5 hypoxic ventilatory response.2 P , 0.05.
436 C.M. BEALL ET AL.
resting ventilation variance and 1⁄2 of theHVR variance and 3⁄4 of the end-tidal CO2variance.
In contrast, the Aymara sample had nosignificant residual h2 of resting ventilation,just 22% residual h2 of HVR and no signifi-cant variance in either trait due to sharedhousehold effects. Random environmentaldifferences accounted for roughly 7⁄10 of Ay-mara resting ventilation and HVR varia-tion.
Lifestyle factors such as drinking saltedbutter tea among Tibetans, coca chewingamong Andean highlanders, and cigarettesmoking were considered for their potentialto account for some of the random environ-mental variation. Nearly all Tibetan adultsdrank salted butter tea, however, the vol-ume consumed (categorized as , 1 L/day,1–2 L/day or 21 L/day) was not associatedwith resting ventilation or HVR variation (Fof two way ANOVA by amount of tea and sexon resting ventilation 5 0.9, on HVR 5 0.1,both P . 0.05). Among Aymara adults therewas no effect on resting ventilation or HVRof chewing coca leaves (F of two way ANOVAcomparing male and female, chewers and
non-chewers, controlling for age on restingventilation 5 0.2, on HVR without control-ling for age 5 0.1, both P . 0.05). Non-smoking Tibetan men (Tibetan women donot smoke) had a mean resting ventilation of21.4 L/min compared with smokers’ mean of17.8 L/min (t 5 2.0, p50.5). There was nosmoking-related difference in Tibetan HVR(t 5 1.5, P . 0.05). There was no influence ofsmoking on Aymara resting ventilation orHVR (F of two way ANOVA of male andfemale smokers and non-smokers, control-ling for age on resting ventilation 5 0.1, onHVR without controlling for age 5 0.04,both P . 0.05).
DISCUSSION
Healthy Tibetan males and females hadmarkedly higher ventilation than Aymaraunder normal conditions of ambient hypoxiaand under conditions of additional experi-mentally-induced hypoxia at 3,800–4,065 m.Tibetan adults at rest breathed about 1.5times more air per minute than Aymara.When exposed to additional experimentally-induced hypoxia, Tibetan breathing in-
TABLE 3. Bivariate correlations with restingventilation and HVR in samples of 13–19 year olds1
Tibetan Aymara
Males Females Males Females
Resting ventilationwith:
Age 10.18 10.392 10.422 10.17Height 10.27 10.23 10.672 10.592
Weight 10.34 10.16 10.682 10.412
BMI 10.36 10.13 10.532 10.20Chest width 10.13 10.10 10.552 10.302
Chest depth 10.382 10.13 10.332 10.19HVR 10.33 10.20 10.432 10.392
SaO2 20.522 20.362 10.24 10.19End-tidal CO2 20.512 20.372 0.50 20.16
(n 5 13) (n 5 18)HVR with:
Age 10.03 10.17 10.20 10.11Height 10.03 10.15 10.25 10.462
Weight 20.01 10.09 10.372 10.432
BMI 0.00 10.02 10.422 10.312
Chest width 20.04 10.14 10.432 10.19Chest depth 10.132 20.13 10.04 10.26SaO2 20.13 20.27 10.09 10.11End-Tidal CO2 20.15 20.06 20.18 20.25
(n 5 13) (n 5 18)Age with height 10.722 10.742 10.622 10.382
1 BMI 5 body mass index, HVR 5 hypoxic ventilatory response,SaO2 5 percent of oxygen saturation of arterial hemoglobin.2 P , 0.05.
TABLE 4. Bivariate correlations with restingventilation and HVR in samples of adults
20 years of age and older1
Tibetan Aymara
Males Females Males Females
Resting ventilationwith:
Age 10.13 20.05 20.133 20.05Height 20.02 20.10 10.202 10.13Weight 20.01 10.08 10.152 10.162
BMI 10.00 10.18 10.08 10.11Chest width 20.08 20.09 10.172 10.262
Chest depth 20.01 10.14 10.05 10.192
HVR 10.532 10.432 10.142 10.182
SaO2 20.222 20.272 10.11 10.132
End-tidal CO2 20.08 20.332 20.382 10.572
(n 5 65) (n 5 48)HVR with:
Age 10.02 20.05 20.182 20.143
Height 0.00 20.09 10.152 10.14Weight 10.02 0.00 10.153 10.10BMI 10.01 10.06 10.10 10.04Chest width 10.16 10.09 10.192 10.172
Chest depth 20.06 20.02 20.05 10.01SaO2 20.11 20.14 10.02 10.252
End-Tidal CO2 20.08 20.12 20.15 20.08Age and height 20.122 20.372 20.342 20.272
* p , .05, †p 5 .051 BMI 5 body mass index, HVR 5 hypoxic ventilatory response,SaO2 5 percent of oxygen saturation of arterial hemoglobin.2 P , 0.05.3 P 5 0.05.
437VENTILATION AT HIGH ALTITUDE

creased roughly twice as much as the Ay-mara. For example, a 10% fall in SaO2produced an average 9.3 L/min increaseamong Tibetan men compared to a 4.5 L/minincrease among Aymara men. The popula-tion difference occurred from the secondthrough the ninth decade of life.
The study was designed and implementedto exclude differences in recruitment andmeasurement as sources of error. With re-spect to recruitment, the method of identify-ing and contacting study participants wasthe same and the rate of family and indi-vidual participation was high in both studysites. Sample selection in the present studybegan with a complete census of the localcommunity and eventually included 96% ofthe Tibetan and 77% of the Aymara house-holds; 68% of the Tibetans eligible by ageand 57% of the Aymara eligible by age. Thelarge proportion of households and individu-als participating makes it unlikely that theresults are influenced by selection bias forventilatory traits.
With respect to measurement, several linesof evidence indicate that the sample differ-ences are substantive rather than an arti-fact of methodology. First, the same investi-gator used the same equipment (with thenoted exception of the capnometer) and care-fully administered the same testing protocolin both study sites. The same criteria forexcluding determinations from analyseswere applied in both sites. (We found noother published criteria or rate of exclusion.)Second, the repeatability of the resting ven-tilation and HVR determinations was goodand was similar in the two study sites. Areview of 33 published studies of restingventilation found no reports of repeatabilitywith which to compare these findings. Areview of 23 published studies of HVR foundthree mentions of HVR repeatability. Thosethree reported the average of two trials asthe HVR and reported that the two trialswere ‘‘not different’’ in a US sample (Chap-man and Cherniack, 1987), and differed byan average of 20.03 DL BTPS/min/D%SaO2
TABLE 5. Multiple regression analysis of adult resting ventilation: R2 and F of R2 differences with theaddition of predictor variables1,2
Multiple R2
of age, SaO2and HVR
R2 changewith height
added
R2 changewith chest
width added
R2 changewith chest
depth added
Multiple R2 with age,SaO2, height, chest
width and chest depth
Tibetan men 0.3243 0.001 0.012 0.005 0.3603
Tibetan women 0.2473 0.000 0.005 0.0383 0.2903
Aymara men 0.047 0.0363 0.018 0.001 0.1013
Aymara women 0.068 0.032 0.0723 0.019 0.1933
1 SaO2 5 percent of oxygen saturation of arterial hemoglobin, HVR 5 hypoxic ventilatory response.2 The statistical test was an F test of the increase in the amount of explained variance with the addition of each predictor variable. Themultiple correlation coefficient for the three variable model was 0.59 for Tibetan men, 0.50 for Tibetan women (both P , 0.05), and 0.21for Aymara men and 0.22 for Aymara women (both P . 0.05). The multiple correlation coefficient for the six variable model was also0.60 for Tibetan men, 0.54 for Tibetan women, and 0.32 for Aymara men and 0.44 for Aymara women (all P , 0.05).3 Significance of F , 0.05.
TABLE 6. Relative variance components of ventilation in Tibetan and Aymara high altitude native samples1
Variables
Proportion of total phenotype variance
Total phenotypic S.D. Residual h2Covariates2 Genes Household Environment
TibetanResting ventilation 0.0144 0.3134 0.3035 0.370 7.556 0.317HVR 0.000 0.3453 0.1533 0.502 0.734 0.345End tidal CO2 0.0293 0.2183 0.000 0.753 5.538 0.225
AymaraResting ventilation 0.2036 0.044 0.073 0.680 2.378 0.055HVR 0.0516 0.2094 0.025 0.715 0.252 0.220
1 h2 5 heritability, HVR 5 hypoxic ventilatory response.2 Covariates for Tibetan resting ventilation are height and male age, for Tibetan end-tidal CO2 they are female age and age2, forAymara resting ventilation they are sex and height, for Aymara HVR it is height.3 P , 0.05.4 P , 0.01.5 P , 0.001.6 P , 0.0001.
438 C.M. BEALL ET AL.

(Curran et al., 1995) and an average of10.02 DL BTPS/min/D%SaO2 (Zhuang etal., 1993) in two samples of Tibetan males.Those calculations differ from the presentrepeatability measures (the average differ-ence between two days of HVR calculated asthe average of three determinations eachday). Yet, they accord well with the repeat-ability of 20.05 and 10.12 DL/min/D%SaO2found in the present study and indicatesome consistency of HVR repeatability acrossstudies. Third, the possibility that responsesto testing might account for the sampledifferences can be discounted. If the Tibet-ans, but not the Aymara, were anxious dur-ing the ventilation determinations, then thiscould result in a systematic elevation ofpulse and oxygen saturation in the ventila-tion testing situation compared with a non-ventilation testing situation in the Tibetan,but, not the Aymara sample. This was notobserved. Instead, the differences betweenpulse measured during quiet rest and thatmeasured during the resting ventilation de-termination was 3–4 f/minute in both sitesand the difference between SaO2 measuredduring quiet rest and that measured duringthe resting ventilation determination wasabout 1% in both sites. The two studysamples appear to have responded similarlyto ventilatory testing (alternatively, theywere equally anxious).
Fourth, the possibility that the Tibetansparticipating in this study were unique canbe discounted by comparing the results ofthe present study with those of previouslypublished studies of Tibetan men (we arenot aware of any on Tibetan children orwomen). The differences of 0.8% (Tibetan)and 1.2% (Aymara) between resting andtesting SaO2 found in the present studywere very similar to the 0.7% and 2.1%reported for Tibetan men measured withand without mouthpieces (Zhuang et al.,1993; Curran et al., 1995). The resting heartrates of Tibetan men in the present studyare 73 f/min which is in the range of 71–87f/min reported for young adult Tibetan menmeasured at their habitual altitudes of resi-dence (Ge et al., 1994 a; Zhuang et al, 1993;Sun et al., 1990); as well as the range of56–92 f/min reported for Tibetan men mea-sured while visiting at altitudes below their
habitual altitudes of residence (Curran etal., 1995; Ge et al., 1994b). The standarderrors of the mean (SEM) of resting heartrates were similar also (ranging from 1.4 inthe present study to 3 in Curran et al. 1995).The breathing frequency of the present sam-ple of Tibetan men was 15 f/min which isslightly lower than the 17–21 f/min reportedfor Tibetan and Sherpa men measured at ornear their habitual altitudes of residence(Sun et al., 1990; Ge et al., 1994 a and b;Hackett et al., 1980; Zhuang et al., 1993)and the 21 f/min reported for Tibetan menmeasured while visiting at lower altitudes(Curran et al, 1995; Ge et al, 1994b). TheSEM of breathing frequencies were similarin the various studies (ranging from 0.4 inthe present study to 1.1 in Hackett et al.1980). The adult male resting end-tidal CO2of the present sample of Tibetan men is 30.1Torr which is a bit higher than the 29 Torrreported for Sherpas at 1,700 m who live andwork at a variety of altitudes and lower thanthe 32 Torr reported for men at their ha-bitual altitude of residence (Zhuang et al.,1993) and 33 Torr reported for Tibetan menvisiting at lower altitude (Curran et al.,1995). The SEM of end-tidal CO2 were alsosimilar in the various studies (ranging from0.5 in Sun et al., 1990 and Zhuang et al.,1993, to 0.9 in the present study, to 1.1 in Geet al., 1994a). The similarity of the meansand standard errors indicates that the studysubjects were equally at rest during thepresent and published studies (or that allTibetans are anxious).
In summary, small physiologic differencesbetween resting and testing situations areevidence that the testing apparatus did notinduce anxiety. The good repeatability indi-cates that study participants were not anx-ious during their first ventilation tests andthat the measurement techniques were pre-cise. The similar size of these differences inthe two sites suggests that the Tibetan andAymara study participants were equally com-fortable during ventilation testing and thatthe measurement techniques did not differ.The repeatability findings and resting physi-ological data accord with other publishedstudies of Tibetans and indicate similarrepeatability of measurement technique andsimilar relaxed state of the study partici-
439VENTILATION AT HIGH ALTITUDE

pants. These diverse lines of evidence arguestrongly that the findings of the present studyare substantive, rather than methodological.
The adult findings of the present studyare quantitatively consistent with previousreports of resting ventilation and HVR onsmaller and less representative samples ofTibetans and Andean highlanders. Figure 3summarizes reports of mean values of rest-ing ventilation of 1,116 natives and long-term residents above 2,000 m, including 632from the present study, and contrasts thesewith reports from low altitude.2 There is a
wide range of variation at all altitudes.There is a 7.3 L/min difference between thelowest and highest North American meanresting ventilation and there is an 8.5–8.6L/min difference between the lowest andhighest Tibetan and Andean resting ventila-tions. The wider span of values at highaltitudes probably reflects the range of alti-tudes of the studies. The lowest Tibetanresting ventilations overlap only with thehighest sea level values. In contrast, themeans of eleven of the fifteen Andeansamples lie in the sea level range and justfour are above. That is, Andean high alti-tude natives generally breathe at rest about2Studies reported values in L if a flow measurement (e.g.
pneumotachograph) were used and in L BTPS if a volumemeasurement (e.g. weather balloon) were used. To determinewhether the two types of measurements gave similar results, weconducted analyses of variance comparing the published meansof low altitude samples reporting resting ventilation in L mea-sured by pneumotachograph, ventilation in L measured by otherflow devices (hot-wire flowmeter or bidirectional turbine) andventilation in L BTPS measured from a volume. There was nosignificant effect of measurement technique on resting ventila-tion or HVR. Therefore we report the published means on thesame figures. Reports of studies employing pneumotachographsinclude those by Filuk et al., 1988; Georgopoulos et al., 1989 aand b; Holtby et al., 1988; Littner et al., 1984; Peterson et al.,1981; Sato et al., 1992. The studies by Schoene, 1982 and
Schoene et al., 1990 using pneumotachographs were corrected toBTPS. Studies employing other flow devices include Nishimuraet al., 1987; Nishimura et al., 1989; Poulin et al., 1993; andSuzuki et al., 1989. Studies reporting volume measure in L BTPSinclude Altman and Dittmer, 1971; Beall et al., 1992; Chapmanet al., 1987; Chiodi, 1957; Cruz, 1973; Cudkowicz et al., 1972;Curran et al., 1995 ; Ge et al., 1994 a and b; Ge et al. , 1995;Hackett et al., 1980; Huang et al., 1984; Hultgren et al., 1965;Hurtado et al., 1964; Kryger et al., 1978; Levine et al., 1992;Moore et al., 1986. The Huang et al., 1981 study did not specifymeasurement technique and it is assumed to be BTPS.
Fig. 3. Summary of studies of resting ventilation at various altitudes.
440 C.M. BEALL ET AL.
the same as or slightly more than low alti-tude natives at low altitude despite thedifference in hypoxic stimulus to breathe.
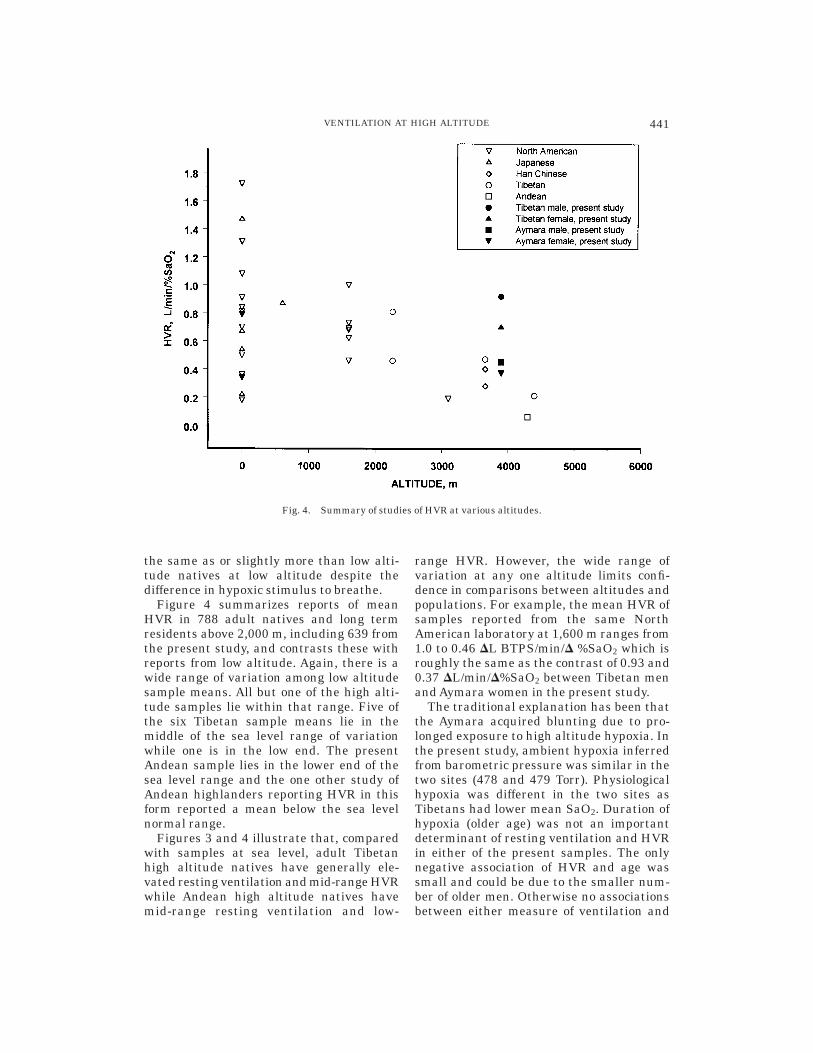
Figure 4 summarizes reports of meanHVR in 788 adult natives and long termresidents above 2,000 m, including 639 fromthe present study, and contrasts these withreports from low altitude. Again, there is awide range of variation among low altitudesample means. All but one of the high alti-tude samples lie within that range. Five ofthe six Tibetan sample means lie in themiddle of the sea level range of variationwhile one is in the low end. The presentAndean sample lies in the lower end of thesea level range and the one other study ofAndean highlanders reporting HVR in thisform reported a mean below the sea levelnormal range.
Figures 3 and 4 illustrate that, comparedwith samples at sea level, adult Tibetanhigh altitude natives have generally ele-vated resting ventilation and mid-range HVRwhile Andean high altitude natives havemid-range resting ventilation and low-
range HVR. However, the wide range ofvariation at any one altitude limits confi-dence in comparisons between altitudes andpopulations. For example, the mean HVR ofsamples reported from the same NorthAmerican laboratory at 1,600 m ranges from1.0 to 0.46 DL BTPS/min/D %SaO2 which isroughly the same as the contrast of 0.93 and0.37 DL/min/D%SaO2 between Tibetan menand Aymara women in the present study.
The traditional explanation has been thatthe Aymara acquired blunting due to pro-longed exposure to high altitude hypoxia. Inthe present study, ambient hypoxia inferredfrom barometric pressure was similar in thetwo sites (478 and 479 Torr). Physiologicalhypoxia was different in the two sites asTibetans had lower mean SaO2. Duration ofhypoxia (older age) was not an importantdeterminant of resting ventilation and HVRin either of the present samples. The onlynegative association of HVR and age wassmall and could be due to the smaller num-ber of older men. Otherwise no associationsbetween either measure of ventilation and
Fig. 4. Summary of studies of HVR at various altitudes.
441VENTILATION AT HIGH ALTITUDE

age were found during adulthood. Thus thepopulation difference was not due to ac-quired blunting during adulthood in thepresent study.
This finding contrasts with some but, notall, investigations of acquired blunting. OneAndean study compared five long term maleresidents at 3,990 and 20 at 4,515 m withnewcomers and found a low resting ventila-tion (Chiodi, 1957). However, another An-dean study of sea level natives at highaltitude failed to find any longer term mi-grants with low HVR (Sorenson and Sever-inghaus, 1968). One study of Tibetans re-ported no change in HVR with age across thespan from 21 to 31 years of age (Zhuang etal., 1993).Another noted a significant moder-ate decrease in HVR from 20 to 30 years ofage in a sample of 20 Tibetan males native toH4,400 m measured while visiting at 3,658m (Curran et al., 1995). That finding may bedue to the small sample sizes at the ends ofthe age range that captured a restrictedportion of the range of variation rather thanto a biological phenomenon in the popula-tion. Another noted a decrease in HVR withincreasing altitude exposure assessed as amultiple of altitude of birth and years ofresidence at that altitude (Hackett et al.,1980). That sample of elite Sherpa maleclimbers was native to a range of altitudesand had a variety of exposures to altitudeincluding the summit of Everest and wasmeasured while visiting at 1,377 m. Inspec-tion of the published figure reveals that theimpression of an association is mainly due totwo Sherpas who were outliers on the alti-tude exposure variable and who had nega-tive HVRs. Three of 25 Sherpas (12%) mea-sured had negative HVRs, yet, just ten of the418 men (2%) in the present study hadnegative HVRs. These observations suggestthat the sample may not have been represen-tative of high altitude native Sherpa andthat the conclusion that the reported resultsare due to duration of altitude exposure maybe confounded by self-selection for the occu-pation of high-altitude climber. These differ-ences in sample size and/or sample charac-teristics may account for the differentconclusions of published studies suggestingan association between blunting and longerduration of high altitude exposure and the
present study which finds no such associa-tion.
Nor is severity of hypoxia an importantdeterminant of resting ventilation and HVRin the present sample. Within populationsthere was a small negative Tibetan associa-tion between SaO2 and resting ventilation,but not HVR, that accounted for very littlevariation. Apart from the small positiveassociation of SaO2 and resting ventilationin Aymara females, associations of SaO2with ventilation were not found in the Ay-mara sample. Between populations, the morehypoxic Tibetans had higher ventilations.This also contrasts with a previous study.One previous report measured a sample ofTibetan men native to H4,400 m measuredwhile visiting at 3,658 m and compared itwith a sample of residents at 3,658 m (Cur-ran et al., 1995). It found a lower HVR, butnot resting ventilation, in the sample visit-ing from high altitude and concluded that itwas due to the more severe lifelong hypoxiaat H4,400 m compared with 3,658 m. Thepresent study found the opposite: Tibetanswith more severe hypoxia had slightly higherresting ventilation, but not HVR, and be-tween samples, the more hypoxic Tibetanshad higher ventilations and HVR. Possiblereasons for the different findings include thefollowing. The sample from H4400m in thatstudy was selected to match a sample from aprevious study by many of the same authorsand thus may not be representative of itspopulation of origin. Furthermore, informa-tion on lifelong physiological hypoxia wasnot provided for those from H4,400 m. Thereis a wide range of physiological hypoxia atany altitude, partly genetically determinedin Tibetans (Beall et al., 1994; Beall et al.,1997). For example, some individuals at4,850–5,450 m have SaO2 no different fromsome individuals at 3,800–4,065 m (Beall etal., 1994a; Beall et al., 1997 ). These consid-erations make it difficult to evaluate theinference that the lower mean HVR in thesample from H4,400 m was due to greaterhypoxia at the higher altitude of residence.
The adolescent findings from the presentstudy do not confirm previous studies sug-gesting that Andean hypoventilation is ac-quired during adolescence (Lahiri et al.,
442 C.M. BEALL ET AL.

1976). The published data on 58 Andeanhigh altitude natives between 13 and 20years of age were reported categorically asnormal, intermediate and blunted, ratherthan numerically, in the single study avail-able (Lahiri et al., 1976). Therefore it is notpossible to examine these data and to sug-gest a reason for the contrast with theresults of the present study of 105 Aymarabetween 13 and 19 years of age. Generally, afunctional ventilatory decrease seems un-likely to develop during adolescence. Circu-lating levels of estrogen, progesterone andtestosterone, all associated with greater rest-ing ventilation and HVR (e.g. Regensteineret al., 1989; Tatsumi et al., 1991; Tatsumi etal., 1994), increase during adolescence gener-ally (Tanner, 1978). Plasma testosterone con-centration increased as expected during ado-lescence in the present two samples (Bealland Worthman, unpublished observations).Aymara and European adolescents exhibitfunctional increase in cardiorespiratory char-acteristics during adolescence at high alti-tude including an age related increase inventilation during maximal exercise (Greksaet al., 1985). If hypoxia were the causalagent for the Aymara hypoventilation rela-tive to Tibetans in the present study sample,it would have to have exerted its influenceentirely before thirteen years of age. A defin-itive test of the absence of blunting wouldrequire longitudinal studies and studies ofyounger age groups.
If blunting during adolescence in the Ay-mara sample does not explain the popula-tion differences in ventilation found in thepresent study, then other factors must beconsidered. Larger body size could possiblyexplain the greater ventilation among thetaller Tibetans. However, Tibetans also hadsignificantly greater ventilation relative tostature. Among men, the average restingventilation relative to stature was 12.0 6 5.1L/meter among Tibetans compared with 8.46 1.5 L/meter among Aymara (t 5 6.2, P ,0.05). The average HVR relative to staturewas 0.58 6 0.55 DL/min/D%SaO2/meteramong Tibetans compared with 0.29 6 0.18DL/min/D%SaO2/meter among Aymara (t 58.8, P , 0.05). The same magnitude contrastwas found among women. The average rest-ing ventilation relative to stature was 11.6 6
5.2 L/min/meter among Tibetan women com-pared with 7.8 6 1.5 L/min/meter amongAymara (t 5 7.7, P , 0.05). The HVRrelative to stature was 0.48 6 0.42 DL/min/D%SaO2/meter among Tibetan women com-pared with 0.26 6 0.15 DL/min/D%SaO2/meter among Aymara (t 5 5.6, P , 0.05).Comparisons using BMI rather than statureyielded (data not shown) even greater popu-lation contrast because the Tibetans havelower BMI. Thus, the shorter stature of theAymara does not account for their low venti-lation compared to Tibetans.
Other factors that might cause Aymarahypoventilation include endurance trainingand a family history of chronic obstructivepulmonary disease (Saunders et al., 1976;Kawakami et al., 1981). It seems unlikelythat either occurs on a population-wide ba-sis from adolescence through old-age in bothsexes. Because testosterone raises HVR (Tat-sumi et al., 1994), lower testosterone concen-trations could hypothetically account for thelower Aymara HVR, but this is also unlikelybecause the population difference was foundamong males and females. Furthermore, theAymara and Tibetan adult male meanplasma testosterone concentrations do notdiffer (Worthman et al., 1977). There isevidence that intrauterine cocaine exposureis associated with blunted HVR in infanthumans and rabbits (Ward et al., 1992;Weese-Mayer et al., 1992). However, thenormal Andean newborn resting ventilationand HVR (Mortola et al., 1990; Mortola etal., 1992) argue against any influence ofmaternal coca chewing. Overall, there wasno evidence to suggest that known system-atic environmental factors account for thelow Aymara mean and variance of restingventilation and HVR.
On the other hand, the population differ-ence could be due to elevation of Tibetan,rather than depression of Aymara, ventila-tion. Factors that might cause Tibetan hyper-ventilation include systematically elevatedmetabolic rate (Huang et al., 1984; Regen-steiner et al., 1990). Because both Tibetanand Andean highlanders have basal meta-bolic rate similar to those predicted by sealevel equations (Beall et al., 1996), this is anunlikely explanation. Early hypertension,reported to increase resting ventilation
443VENTILATION AT HIGH ALTITUDE

(Tafil-Klawe et al., 1989), is common amongTibetans simply because there is a highprevalence of hypertension (Beall et al.,under submission a). Reasoning that hyper-tensives under 25 years of age are likely tobe in the early phases of hypertension, ananalysis of variance compared the ventila-tion of Tibetan normo-and hypertensives.Seven of 43 males and eight of 54 females inthe 13-24 year age range were hypertensivebut did not differ from the normotensives inresting ventilation or HVR. Just one of 177Aymara in that age range was hypertensive.Caffeine and xanthines are respiratorystimulants in tea (Howell, 1992; Larrick,1991). Daily intake of a liter or more of blacktea flavored with salt and butter was nearlyuniversal among Tibetan adults. Tea con-sumption was much lower in the Aymarasample where individuals drank just 1–3cups of black or herbal tea/day. BecauseTibetan children and teenagers drink lesstea than adults and the population differ-ence was found throughout the age range,tea drinking probably does not account forthe Tibetan hyperventilation.
Thus there was little evidence to indicatethat sustained lifelong Tibetan hyperventila-tion and relative Aymara hypoventilationwere due to the systematic influence ofenvironmental factors. Population differ-ences in the frequencies of alleles influenc-ing the level of ventilation could account forsome of the differences according to thefollowing logic. Quantitative genetic analy-ses revealed evidence of significant geneticvariation in resting ventilation only in theTibetan sample and more genetic variationin HVR in the Tibetan than the Aymarasample. Because the potential for naturalselection is a function of the amount ofgenetic variance, this means that only theTibetan population has the potential fornatural selection on resting ventilation andit also has more potential for natural selec-tion on HVR. The present data suggest ahypothesis explaining how this came about.With respect to resting ventilation, the com-bination of high Tibetan values compared tosea level and the existence of intrapopula-tion genetic variation suggest the hypoth-esis that natural selection and/or randomgenetic drift has acted to increase the fre-
quency of alleles for high ventilation. Thesimilarity ofAymara resting ventilation com-pared to sea level values, the absence ofAymara genetic variation, and the absenceof evidence for environmental blunting ofAymara ventilation suggest the hypothesisthat natural selection has not acted on rest-ing ventilation in Andean populations be-cause there was no heritable variation uponwhich to act. With respect to HVR, thecombination of normal Tibetan values com-pared to sea level, the existence of Tibetangenetic variation, and of genetic variation inHVR of sea level populations (Collins et al.,1978; Kawakami et al., 1984; Kawakami etal., 1981; Moore et al., 1976; Saunders et al.,1976) suggest the hypothesis that naturalselection has acted to maintain existinggenetic variation in the Tibetan sample. Thelow normal to below sea level Aymara HVRand the relatively low genetic variance sug-gest the hypothesis that genetic variationhas been lost in the Andean population dueto natural selection and/or random geneticdrift and that alleles for high values havebeen lost. Current knowledge of the geneticsof these traits does not permit direct evalua-tion of these models positing different micro-evolutionary histories leading to differentventilation patterns in these two popula-tions with long histories of high altitudeexposure.
The conclusion of this paper differs funda-mentally from previous studies emphasizingthe importance of acquired hypoventilationand blunting of HVR. One methodologicalreason for this may be the combination ofthe small samples often reported, some-times of specialized occupational groups,sometimes not measured at their habitualaltitudes of residence, and usually of unspeci-fied representativeness. Another method-ological reason for the different conclusionmay be the broad age range sampled in thepresent study which provided the opportu-nity to observe trends or, more importantlyas it turned out, their absence. Anotherreason may be the extraordinarily widerange of normal variation in these two traits.The coefficient of variation of HVR was 0.56among the Aymara and 0.90 among Tibetanadults in the present study while the coeffi-cient of variation in height was 0.05 in both
444 C.M. BEALL ET AL.

samples. Others have noted that normalHVR varies eight-fold (e.g. Reeves et al.,1993). That much variation in a trait such asheight would be unimaginable. With such avariable trait, there is great potential formisleading findings from small samples thatinadvertently capture just a fraction of thenormal range.
It is not clear whether the ventilationdifferences are relevant to successful high-altitude adaptation. Lower ventilation wouldseem likely to increase the chances of poorerSaO2 and oxygen delivery, but, the Aymarasample had the higher mean SaO2. Bothpopulations deliver adequate oxygen to tis-sues under a range of conditions as demon-strated by normal basal metabolic rates andmaximal physical work capacities, (Baker,1976; Beall, et al., 1996; Lahiri et al., 1976;Mazess et al., 1969; Sun et al., 1990; Tufts etal., 1985). Both populations are successfulby a number of criteria generally used forevolutionary analyses. Both are growingpopulations. The Tibetan population had a15.2/1,000 rate of natural increase in 1990(Xi Zang Tong Ji Jian Jian, 1993) and theAndean region had a 20.5/1,000 rate ofnatural increase in 1990–1995 (Pan-Ameri-can Health Organization, 1994). Both havepersisted for millennia in their high altitudeenvironments. The earliest chronometri-cally dated evidence for occupation of theTibetan Plateau above 2,500 m is a farmingsite that dates to 4,000–5,500 years ago(Chang, 1992). The earliest evidence foroccupation of the Andean Plateau above2,500 m dates to 11,000 years ago (Aldender-fer, in press 1997). These functional anddemographic lines of evidence suggest thatboth populations are well-adapted.
Thus the relatively low ventilation doesnot appear to have particularly adverse con-sequences for the Aymara and the hyperven-tilation does not appear to be particularlyadvantageous for the Tibetans. However,these traits must be viewed in a broadercontext. The Tibetan and Aymara samplesdiffer on more than ventilation. The Tibet-ans also have significantly lower hemoglo-bin concentration and SaO2 than the Ay-mara (Beall et al., 1997, submitted). Thereis evidence for significant genetic variationin Tibetan resting ventilation, HVR, SaO2
(for which there is a major gene) and hemo-globin concentration. There is evidence foran absence of genetic variance in Aymararesting ventilation and SaO2, small geneticvariance in HVR, and substantial geneticvariance in hemoglobin concentration (Beallet al., 1994; Beall et al., under submission).The results of the present paper add evi-dence to the hypothesis that natural selec-tion has acted on a suite of traits to producequantitatively different, well-adapted pheno-types in these two populations with long,successful histories of high altitude resi-dence.
ACKNOWLEDGMENTS
We are grateful to the residents of Pen-Driand Ventilla and their relatives for theirgenerous participation in this study. Wethank the Tibet Academy of Social Science,Lhasa, Tibet Autonomous Region for hostingthe Tibetan portion of this field study. Weare grateful to Mr. Ben Jiao and Mr. TseringPuntsoh for assistance in the Tibetan datacollection and to Mr. Augustin Huaynoca,Mr. Marcelino Mamani, Mr. Rene Condori,and Mr. Augustin Mamani for assistance inthe Bolivian data collection. Mr. DennisSepulveda performed some of the Aymaraanthropometry and interviews. We are espe-cially indebted to Tod Flak, Ph.D. for writingand customizing the Vital Signals r soft-ware. An abbreviated report of the findingsdescribing adults was presented in 1997 atthe Tenth Biennial International Sympo-sium on Hypoxia and Mountain Medicineand will appear in those proceedings. Thisresearch was supported by National ScienceFoundation award no. SBR92-21724 to CMB.
LITERATURE CITED
Aldenderfer M (In press, 1997) The Pleistocene/Ho-locene Transition in Peru and its Effects Upon HumanUse of the Landscape. Quarternary International .
Altman PL, and Dittmer DS, eds. (1971) Respirationand circulation. Bethesda, Md., FASEB.
Baker PT (1976) Work performance of highland natives.In Baker PT and MA Little (eds.): Man in the Andes: Amultidisciplinary study of high-altitude Quechua.Stroudsburg, Pennsylvania: Dowden, Hutchinson, &Ross, Inc., pp. 300–314.
Beall CM, and Goldstein MC (1990) Hemoglobin Concen-tration, Percent Oxygen Saturation and Arterial Oxy-gen Content of Tibetan Nomads at 4,850 to 5,450 M.In Sutton JR, G Coates and JE Remmers (eds.):Hypoxia: The Adaptations. Toronto, B.C. Decker, Inc.,pp. 59–65.
445VENTILATION AT HIGH ALTITUDE

Beall CM, Strohl KP, Gothe B, Brittenham GM, Barra-gan M, and Vargas E (1992) Respiratory and hemato-logical adaptations of young and older Aymara mennative to 3,600 m. Am. J. Hum. Biol. 4:17–26.
Beall CM, Blangero J, Williams-Blangero S, and Gold-stein MC (1994) A major gene for percent of oxygensaturation of arterial hemoglobin in Tibetan highland-ers. Am. J. Phys. Anthropol. 95:271–276.
Beall CM, Henry J, Worthman C, and Goldstein MC(1996) Basal metabolic rate and dietary seasonalityamong Tibetan nomads. Am. J. Hum. Biol. 8:361–370.
Beall CM, Strohl K, Blangero J, Williams-Blangero S,Brittenham GM, and Goldstein MC (1997) Quantita-tive genetic analysis of arterial oxygen saturation inTibetan highlanders. Hum. Biol. 69: 597–604.
Beall CM, Brittenham GM, Strohl KP, Blangero J,Williams-Blangero J, Almasy LA, Goldstein MC, Var-gas E, Villena M, Alarcon AM, and Gonzalez C (undersubmission) Blood pressures of Tibetan and Aymarahigh-altitude natives. Am. J. Hum. Biol.
Beall CM, Brittenham GM, Strohl KP, Blangero J,Williams-Blangero S, Goldstein MC, Decker MJ, Var-gal E, Villena M, Soria R, Alarcon AM, and Gonzales C(under submission) Lower hemoglobin concentrationof iron sufficient Tibetan compared to Bolivian Ay-mara highlanders. Am. J. Phys. Anthropol.
Beall CM, Almasy LA, Blangero J, Williams-Blangero S,Brittenham GM, Strohl KP, Decker M, Vargas E,Villena M, Soria R, Alarcon A, and Gonzales C (undersubmission) Percent oxygen saturation of arterialhemoglobin of Bolivian Aymara at 4,000 m. Am. J.Phys. Anthropol.
Bland JM, and Altman DG (1986) Statistical methodsfor assessing agreement between two methods ofclinical measurement. Lancet 1:307–310.
Byrne-Quinn E, Sodal IE, and Weil JV (1972) Hypoxicand hypercapnic ventilatory drives in children nativeto high altitude. J. Appl. Physiol. 32:44–46.
Cameron N, Hiernaux J, Jarman S, Marshall WA,Tanner JM, and Whitehouse RH (1981) Anthropom-etry. In Weiner JS and JA Lourie (eds.): PracticalHuman Biology. New York: Academic Press, pp. 27–52.
Chang K-C (1992) China. In Ehrich RW (ed.): Chronolo-gies in Old World Archaeology. Chicago and London:The University of Chicago Press.
Chapman KR, and Cherniack NS (1987) Aging effects onthe interaction of hypercapnia and hypoxia as ventila-tory stimuli. J. Gerontol. 42:202–209.
Chiodi H (1957) Respiratory adaptations to chronic highaltitude hypoxia. J. Appl. Physiol. 10:81–87.
Collins DD, Scoggin CH, Zwillich CW, and Weil JV(1978) Hereditary aspects of decreased hypoxic re-sponse. J. Clin. Invest. 62:105–110.
Cruz JC (1973) Mechanics of breathing in high altitudeand sea level subjects. Respir. Physiol. 17:146–161.
Cudkowicz L, Spielvogel H, and Zubieta G (1972) Respi-ratory studies in women at high altitude (3,600 m or12,200 ft and 5,200 m or 17,200 ft). Respiration29:393–426.
Curran LS, Zhuang J, Droma T, Land L, and Moore LG(1995) Hypoxic ventilatory responses in Tibetan resi-dents of 4,400 m compared with 3658 m. Respir.Physiol. 100:223–230.
Filuk RB, Berezanski DJ, andAnthonisen (1988) Depres-sion of hypoxic ventilatory response in humans bysomatostatin. J. Appl. Physiol. 65:1050–1054.
Ge R, Qiuhong C, Lungao H, and Hailing L (1994)Characteristics of hypoxic ventilatory response inTibetan living at moderate and high altitudes. Chi-nese J. Tuberculosis and Respiratory Diseases 17:364–366,388.
Ge R-L, Chen Q-H, Wang L-H, Gen D, Yang P, Keishi K,Fujimoto K, Matsuzawa Y, Yoshimura K, Takeoka M,and Kobayashi T (1994) Higher exercise performanceand lower VO2max in Tibetan than Han residents at4,700 m altitude. J. Appl. Physiol. 77:684–691.
Ge R, He Lun G, Chen Q, Li HL, Gen D, Kubo K,Matsuzawa Y, Fujimoto K, Yoshimura K, Takeoka M,and Kobayashi T (1995) Comparisons of oxygen trans-port between Tibetan and Han residents at moderatealtitude. Wilderness and Environmental Medicine6:391–400.
Georgopoulos D, Walker S, and Anthonisen NR (1989)Increased chemoreceptor output and ventilatory re-sponse to sustained hypoxia. J.Appl. Physiol. 67:1157–1163.
Georgopoulos D, Berezanski D, and Anthonisen NR(1989) Effects of CO2 breathing on ventilatory re-sponse to sustained hypoxia in normal adults. J. Appl.Physiol. 66:1071–1078.
Greksa LP, Spielvogel H, and Paredes-Fernandez L(1985) Maximal exercise capacity in adolescent Euro-pean and Amerindian high-altitude natives. Am. J.Phys. Anthropol. 67:209–216.
Hackett PH, Reeves JT, Reeves CD, Grover RF, andRennied D (1980) Control of breathing in Sherpas atlow and high altitude. J. Appl. Physiol. 49:374–379.
Holtby SG, Berezanski DJ, and Anthonisen NR (1988)Effect of 100% O2 on hypoxic eucapnic ventilation. J.Appl. Physiol. 65:1157–1162.
Hopper JL, and Mathews JD (1982) Extensions tomultivariate normal models for pedigree analysis.Ann. Hum. Genet. 46:373–383.
Howell OL (1993) Effects of adenosine agonists onventilation during hypercapnia, hypoxia in Rhesusmonkeys. J. Pharmacol. Exp. Ther. 265:971–978.
Huang ZR, Zhu SC, Ba ZF, and Hu XC (1981) Ventila-tory control in Tibetan highlanders. In Liu DS (ed.):Geological and ecological studies of Qinghai-XizangPlateau. New York: Gordon and Beach, pp. 1363–1369.
Huang SY, Alexander JK, Grover RF, Maher JT, McCul-lough RE, McCullough RG, Moore LG, Weil JV, Samp-son JB, and Reeves JT (1984) Increased metabolismcontributes to increased resting ventilation at highaltitude. Respir. Physiol. 57:377–385.
Hultgren HN, Kelly J, and Miller H (1965) Pulmonarycirculation in acclimatized man at high altitude. J.Appl. Physiol. 20:233–238.
Hurtado A (1964) Animals in high altitudes: residentman. In Dill DB (ed.): Handbook of Physiology. Section4: Adaptation to the environment. Washington, D.C.:American Physiological Society, pp. 843–859.
Kawakami Y, Yoshikawa T, Shida A, and Asanuma Y(1981) Relationships between hypoxic and hypercap-nic ventilatory responses in man. Jap. J. Physiol.31:357–368.
Kawakami Y, Yamamoto H, Yoshikawa T, and Shida A(1984) Chemical and behavioral control of breathingin adult twins. Am. Rev. Respir. Dis. 129:703–707.
Kryger M, McCullough R, Doekel R, Collins D, Weil JV,and Grover RF (1978) Excessive polycythemia of highaltitude: role of ventilatory drive and lung disease.Am. Rev. Respir. Dis. 118:659–667.
Lahiri S, Delaney RG, Brody JS, Simpser M, VelasquezT, Motoyama EK, and Polgar C (1976) Relativ role ofenvironmental and genetic factors in respiratory adap-tation to high altitude. Nature 261:133–135.
Lange K, and Boehnke M (1983) Extensions to pedigreeanalysis. IV. Covariance components models for multi-variate traits. Am. J. Med. Genet. 14:513–524.
Lange K, Weeks D, and Boehnke M (1988) Programs forpedigree analysis: Mendel, Fisher, and dGene. Genet.Epidemiol. 5:471–472.
446 C.M. BEALL ET AL.

Larrick JW (1991) The methyl xanthine hypothesis:does tea consumption by Tibetan natives blunt theeffects of high altitude? Med. Hypotheses 34:99–104.
Lenfant C, and Sullivan K (1971) Adaptation to highaltitude. New Eng. J. Med. 284:1298–1309.
Levine BD, Friedman DB, Engfred K, Hanle B, Kjaer M,Clifford PS, and Secher NH (1992) The effect ofnormoxic or hypobaric hypoxic endurance training onthe hypoxic ventilatory response. Med. Sci. SportsExerc. 24:769–775.
Littner M, Young E, McGinty D, Beahm E, Riege W, andSowers J (1984) Awake abnormalities of control ofbreathing and of the upper airway: occurrence inhealthy older men with nocturnal disordered breath-ing. Chest 86:573–579.
Mazess RB (1969) Exercise performance at high altitudein Peru. Fed. Proc. 28:1301–1306.
Moore GC, Zwillich CW, battaglia JD, Cotton EK, andWeil JV (1976) Respiratory failure associated withfamilial depression of ventilatory response to hypoxiaand hypercapnia. N. Eng. J. Med. 295:861–865.
Moore LG, Brodeur P, Chumbe O, D’Brot J, HofmeisterS, and Monge C (1986) Maternal hypoxic ventilatoryresponse, ventilation, and infant birth weight at 4,300m. J. Appl. Physiol. 60:1401–1406.
Mortola JP, Rezzonico R, Fisher JT, Villena-Cabrera N,Vargas E, Gonzales R, and Pena F (1990) Complianceof the respiratory system in infants born at highaltitude. Am. Rev. Respir. Dis. 142:43–48.
Mortola JP, Frappell PB, Frappell DE, Villena-CabreraN, Villena-Cabrera M, and Pena F (1992) Ventilationand gaseous metabolism in infants born at highaltitude, and their responses to hyperoxia. Am. Rev.Respir. Dis. 148:1206–1209.
Nishimura M, Suzuki A, Nishiura Y, Yamamoto H,Miyamoto K, Kishi F, and Kawakami Y (1987) Effectof brain blood flow on hypoxic ventilatory response inhumans. J. Appl. Physiol. 63:1100–1106.
Nishimura M, Miyamoto K, Suzuki A, Yamamoto H,Tsuji M, Kishi F, and Kawakami Y (1989) Ventilatoryand heart rate responses to hypoxia and hypercapniain patients with diabetes mellitus. Thorax 44:251–257.
Pan American Health Organization (1994) Health Con-ditions in theAmericas, 1994. Washington, DC: PAHO.
Peterson DD, Pack AI, Silage DA, and Fishman AP(1981) Effects of aging on ventilatory and occlusionpressure responses to hypoxia and hypercapnia. Am.Rev. Respir. Dis. 124:387–391.
Poulin MJ, Cunningham DA, Pateerson DH, KowalchukJM, and Smith WDF (1993) Ventilatory sensitivity toCO2 in hyperoxia and hypoxia in older aged humans.J. Appl. Physiol. 75:2209–2216.
Rebuck AS, and Campbell EJM (1974) A clinical methodfor assessing the ventilatory response to hypoxia. Am.Rev. Respir. Dis. 109:345–350.
Reeves JT, McCullough RE, Moore LG, Cymerman A,and Weil JV (1993) Sea-level PCO2 relates to ventila-tory acclimitization at 4,300 m. J. Appl. Physiol.75:1117–1122.
Regensteiner JG, Woodard WD, Hagerman DD, Weil JV,Pickett CK, Bender PR, and Moore LG (1989) Com-bined effects of female hormones and metabolic rateon ventilatory drives in women. J. Appl. Physiol.66:808–813.
Regensteiner JG, McCullough RG, McCullough RE,Pickett CK, and Moore LG (1990) Combined effects offemale hormones and exercise on hypoxic ventilatoryresponse. Respir. Physiol. 82:107–114.
Sato M, Severinghaus JW, Powell FL, Xu F, and SpellmanMJ (1992) Augmented hypoxic ventilatory response inmen at altitude. J.Appl. Physiol. 73:101–107.
Saunders MA, Leeder SR, and Rebuck AS (1976) Ventila-tory response to carbon dioxide in young athletes: afamily study. Am. Rev. Respir. Dis. 113:497–502.
Schoene RB (1982) Control of ventilation in climbers toextreme altitude. J. Appl. Physiol. 53:886–890.
Schoene RB, Roach RC, Lahiri S, Peters RM, HackettPH, and Santolaya R (1990) Increased diffusion capac-ity maintains arterial saturation during exercise inthe Quechua Indians of Chilean altiplano. Am. J.Hum. Biol. 2:663–668.
Severinghaus JW, Bainton CR, and Carcelen A (1966)Respiratory insensitivity to hypoxia in chronicallyhypoxic man. Respir. Phys. 1:308–334.
Sorenson SS, and Severinghaus JW (1968) Respiratorysensitivity to acute hypoxia in man born at sea levelliving at high altitude. J. Appl. Physiol. 25:211–216.
Stevens SF (1993) Claiming the high ground: Sherpas,subsistence, and environmntal change in the highestHimalaya. Berkeley: University of California Press.
Sun SF, Huang SY, Zhang JG, Droma TS, Banden G,McCullough RE, McCullough RG, Cymerman A,Reeves JT, and Moore LG (1990) Decreased ventila-tion and hypoxic ventilatory responsiveness are not re-versed by naloxone in Lhasa residents with chronicmountain sickness.Am. Rev. Respir. Dis. 142:1294–1300.
Suzuki A, Nishimura M, Yamamoto H, Miyamoto K,Kishi F, and Kawakami Y (1989) No effect of brainblood flow on ventilatory depression during sustainedhypoxia. J. Appl. Physiol. 66:1674–1678.
Tafil-Klawe M, Raschke F, Kublik A, Stoohs R, and vonWichert P (1989) Attenuation of augmented ventila-tory response to hypoxia in essential hypertension inthe course of aging. Respir. 56:154–160.
Tanner JM (1978) Fetus into man. Physical growth fromconception to maturity. Cambridge: Harvard Univer-sity Press.
Tatsumi K, Hannhart B, Pickett CK, Weil JW, andMoore LG (1991) Influences of gender and sex hor-mones on hypoxic ventilatory response in cats. J.Appl. Physiol. 71:1746–1751.
Tatsumi K, Hannhart B, Pickett CK, Weil JV, and MooreLG (1994) Effects of testosterone on hypoxic ventila-tory and carotid body neural responsiveness. Am. J.Respir. Crit. Care Med. 149:1248–1253.
Tufts DA, Haas JD, Beard JL, and Spielvogel H (1985)Distribution of hemoglobin and functional conse-quences of anemia in adult males at high altitude.Am. J. Clin. Nutr. 42:1–11.
Ward SLD, Bautista DB, Woo MS, Chang M, Schuetz S,Wachsman L, Sehgal L, and Bean X (1992) Responsesto hypoxia and hypeircapna in infants of substance-abusing mothers. J. Pediatr. 121:704–709.
Weese-Mayer DE, Klemka-Walden LM,. Bardov A andGingras JL (1992) Effects of prenatal cocaine on theventilatory response to hypoxia in newborn rabbits.Dev. Pharmacol. Ther. 18:116–124.
Weil JV, Byrne-Quinn E, Sodal IE, Filley GF, and GroverRF (1971) Acquired attenuation of chemoreceptorfunction in chronically hypoxic man at high altitude.J. Clin. Invest. 50:186–195.
West JB (1982) Respiratory and circulation control athigh altitudes. J. Exp. Biol. 100:147–157.
Worthman CM, Beall CM, and Stallings JF (1997)Population variation in reproductive function of men.Am. J. Phys. Anthropol. Supplement 24:246.
Xi Zang Tong Ji Nian Jian (1993) The Statistical Year-book of Tibet 1993. Bejing: China Statistical Publish-ing House.
Zhuang J, Droma T, Sun S, Janes C, McCullough RE,McCullough RG, Cymerman A, Huang SY, Reeves JT,and Moore LG (1993) Hypoxic ventilatory responsive-ness in Tibetan compared with Han residents of 3,658m. J. Appl. Physiol. 74:303–311.
447VENTILATION AT HIGH ALTITUDE


















