Uptake and incorporation of choline and ethanolamine into lipoteichoic acid and teichoic acid by the...
5
Uptake and incorporation of choline and ethanolamine into lipoteichoic acid and teichoic acid by the choline-independent mutant JY2190 of Streptococcus pneumoniae Klaus Leopold, Werner Fischer * Institut fu ºr Biochemie, Med. Fak., Universita ºt Erlangen-Nu ºrnberg, Fahrstrasse 17, D-91054 Erlangen, Germany Received 13 October 1998; accepted 20 October 1998 Abstract Lipoteichoic acid- and teichoic acid-containing muropeptides were isolated from choline- or ethanolamine-grown cells of the choline-independent mutant JY2190 of Streptococcus pneumoniae. Choline was taken up and incorporated into lipoteichoic acid and teichoic acid with 81% efficiency, compared with the parent strain Rx1. With similar efficiency, ethanolamine was incorporated. Accordingly, the mutant is a valuable tool for identifying the individual genes encoding the enzymes of choline utilisation, because any of these genes can be deleted without affecting viability and growth rate. z 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved. Keywords : Choline-independent pneumococcus ; Choline phosphate ; Pneumococcal lipoteichoic acid ; Pneumococcal teichoic acid 1. Introduction For wild-type pneumococci, choline (Cho) is an absolute nutritional requirement [1,2]. It is incorpo- rated as choline phosphate (ChoP) into the repeats of lipoteichoic acid (LTA) and teichoic acid (TA) (Fig. 1). For growth, Cho can be substituted by etha- nolamine (Etn) [6]. Although Etn is incorporated as EtnP into LTA and TA in place of ChoP, it cannot replace ChoP functionally (for references see [7]). The major and minor pneumococcal autolysin, N- acetylmuramyl-L-alanine-amidase (LytA) and endo- L-1,4-N-acetylglucosaminidase, require both TA- linked ChoP residues on their cell wall substrate for activity [7,8]. LytA is involved in cell separation at the end of cell division and responsible for stationary phase and L-lactam induced cell lysis [6,8]. Recently, we characterised a Cho-independent mutant strain JY2190 of Streptococcus pneumoniae Rx1 which was isolated by serial passage of strain Rx1 in chemi- cally de¢ned medium (CDM), containing decreasing concentrations of Etn [9]. The change from a strict requirement of choline to the ability to dispense with it was explained by the hypothesis that the mutation in strain JY2190 eliminated a regulatory site which in wild-type strains ensures that only ChoP- (EtnP-) substituted TA is incorporated into peptidoglycan. In this model, growth in the absence of amino alco- hols leads to accumulation of polyprenol-linked non- 0378-1097 / 98 / $19.00 ß 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved. PII:S0378-1097(98)00509-6 * Corresponding author. Tel.: +49 (91) 140-5957; Fax: +49 (91) 3185-4605; E-mail: [email protected] FEMS Microbiology Letters 169 (1998) 355^359
-
Upload
klaus-leopold -
Category
Documents
-
view
212 -
download
0
Transcript of Uptake and incorporation of choline and ethanolamine into lipoteichoic acid and teichoic acid by the...

Uptake and incorporation of choline and ethanolamine intolipoteichoic acid and teichoic acid by the choline-independent
mutant JY2190 of Streptococcus pneumoniae
Klaus Leopold, Werner Fischer *Institut fuër Biochemie, Med. Fak., Universitaët Erlangen-Nuërnberg, Fahrstrasse 17, D-91054 Erlangen, Germany
Received 13 October 1998; accepted 20 October 1998
Abstract
Lipoteichoic acid- and teichoic acid-containing muropeptides were isolated from choline- or ethanolamine-grown cells of thecholine-independent mutant JY2190 of Streptococcus pneumoniae. Choline was taken up and incorporated into lipoteichoicacid and teichoic acid with 81% efficiency, compared with the parent strain Rx1. With similar efficiency, ethanolamine wasincorporated. Accordingly, the mutant is a valuable tool for identifying the individual genes encoding the enzymes of cholineutilisation, because any of these genes can be deleted without affecting viability and growth rate. z 1998 Federation ofEuropean Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.
Keywords: Choline-independent pneumococcus; Choline phosphate; Pneumococcal lipoteichoic acid; Pneumococcal teichoic acid
1. Introduction
For wild-type pneumococci, choline (Cho) is anabsolute nutritional requirement [1,2]. It is incorpo-rated as choline phosphate (ChoP) into the repeatsof lipoteichoic acid (LTA) and teichoic acid (TA)(Fig. 1). For growth, Cho can be substituted by etha-nolamine (Etn) [6]. Although Etn is incorporated asEtnP into LTA and TA in place of ChoP, it cannotreplace ChoP functionally (for references see [7]).The major and minor pneumococcal autolysin, N-acetylmuramyl-L-alanine-amidase (LytA) and endo-
L-1,4-N-acetylglucosaminidase, require both TA-linked ChoP residues on their cell wall substrate foractivity [7,8]. LytA is involved in cell separation atthe end of cell division and responsible for stationaryphase and L-lactam induced cell lysis [6,8]. Recently,we characterised a Cho-independent mutant strainJY2190 of Streptococcus pneumoniae Rx1 whichwas isolated by serial passage of strain Rx1 in chemi-cally de¢ned medium (CDM), containing decreasingconcentrations of Etn [9]. The change from a strictrequirement of choline to the ability to dispense withit was explained by the hypothesis that the mutationin strain JY2190 eliminated a regulatory site whichin wild-type strains ensures that only ChoP- (EtnP-)substituted TA is incorporated into peptidoglycan.In this model, growth in the absence of amino alco-hols leads to accumulation of polyprenol-linked non-
0378-1097 / 98 / $19.00 ß 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.PII: S 0 3 7 8 - 1 0 9 7 ( 9 8 ) 0 0 5 0 9 - 6
FEMSLE 8508 3-12-98
* Corresponding author. Tel. : +49 (91) 140-5957;Fax: +49 (91) 3185-4605;E-mail: [email protected]
FEMS Microbiology Letters 169 (1998) 355^359

substituted TA, rendering polyprenolphosphate un-available for peptidoglycan synthesis [9].
Severin et al. [10] described another Cho-inde-pendent S. pneumoniae strain, R6Cho3. It was re-covered from a heterologous cross with DNA fromStreptococcus oralis which also incorporates ChoPinto TA but, in contrast to pneumococci, grows inthe absence of amino alcohols. Both choline-inde-pendent strains had not acquired the capability tosynthesise Cho or Etn, because both amino alcoholswere absent from TA and LTA when the bacteriahad been grown in the absence of amino alcohols.Under these conditions, JY2190 and R6Cho3
formed long chains and were resistant to all kindsof autolysis, similar to Etn-grown cells of the parentstrains. Since the properties of the parent strainswere in part re-established by growth in the presenceof Cho, it was suggested that both strains retainedthe ability of incorporating Cho into LTA and TA.The present study is aimed to prove this for themutant JY2190 by analysis of LTA and TA isolatedfrom Cho- and Etn-grown cells.
2. Materials and methods
2.1. Materials
CPL, a muramidase of pneumococcal bacterio-phage Cp-1 [11], was kindly provided by ErnestoGarc|èa (Centro de Investigaciones Bioloègicas, Ma-drid, Spain). Mouse monoclonal IgAU (TEPC-15, as-cites £uid) was purchased from Sigma, Germany.
2.2. Bacteria and growth
Pneumococcal strains Rx1, Rx1/Al3 (autolysin de-fective), and JY2190 were from previous work [9].Bacteria were grown at 37³C on a rotary shaker inCDM containing either Cho (5 Wg ml31) or Etn (200Wg ml31). Rx1/Al3 was used in the experiments withCho, because like Cho-grown JY2190 it does not lysein the stationary phase, thus providing intact wallsfor analysis.
2.3. Preparation of LTA- and TA-containingmuropeptides
Late exponential phase cells (OD578V1) were har-vested, suspended in 0.1 M sodium acetate, pH 4.7(bu¡er A), mechanically disintegrated and extractedin a Bligh Dyer monophasic system as previouslydescribed [4,9]. Extracted cell walls were collectedby centrifugation. To the supernatant, appropriatevolumes of bu¡er A and CHCl3 were added, toachieve phase partition. From the resulting MeOHlayer, LTA was puri¢ed by hydrophobic interactionchromatography as described [4]. The cell walls werefurther puri¢ed by successive treatment with nucle-ases and trypsin, extraction with SDS (1%), followedby washing in sequence with H2O, 5.6 M LiCl, and100 mM Na2 EDTA [12]. Puri¢ed cell walls weretreated with CPL, a muramidase (22 Wg ml31), in5 mM ammonium acetate (pH 6.0) and the reactionwas followed by the decrease of OD578 [9]. Sampleswere taken to dryness under reduced pressure withseveral additions of water.
2.4. Radiolabelling
The mutant was grown in CDM containing [meth-yl-3H]choline (5 Wg ml31 ; 1.1U104 Bq Wg31). At timeintervals, samples were withdrawn precipitated inice-cold 10% TCA, collected on glass ¢bre ¢lters(Sartorius, Germany), washed with 10% TCA, and,after drying, the radioactivity was measured in Ro-tiszint plus (Roth, Germany) on a Wallac 1410 liquidscintillation counter (Pharmacia, Finland).
2.5. A¤nity chromatography
For ChoP-speci¢c a¤nity chromatography, mouse
FEMSLE 8508 3-12-98
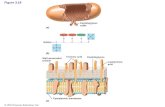
Fig. 1. Identical repeat of LTA and TA [3,4]. One ChoP per re-peat, as shown, is characteristic for S. pneumoniae Rx1 and itsnon-lytic derivative Rx1/Al3 [5]. R, lipid anchor or attachmentto peptidoglycan.
K. Leopold, W. Fischer / FEMS Microbiology Letters 169 (1998) 355^359356

myeloma protein TEPC-15 was used [13,14]. Thecolumn (11 ml) was prepared by binding mouseIgAU (5 mg, ascites £uid) to CNBr-activated Sephar-ose B (2 mgw8 ml gel) as recommended by the man-ufacturer. Samples (180^200 nmol phosphorus) wereapplied in 0.04 M ammonium acetate, pH 6.0 (1 ml)and the column was eluted ¢rst with this bu¡er (24ml), followed by 0.5 M triethylammonium hydrogencarbonate pH 6.0 (50 ml). In the case of LTAs, bothbu¡ers contained 0.1% Triton X-100 in order to de-compose LTA-micelles and incorporate single LTAmolecules into Triton micelles [15]. For analysis, Tri-ton was extracted from combined fractions withCHCl3, salts were removed by lyophilisation withrepeated addition of water.
2.6. Analytical procedures
Choline, ethanolamine and phosphorus werequanti¢ed as in previous work [9].
3. Results
3.1. Incorporation of choline
Fig. 2 shows the incorporation of [methyl-3H]choline by mutant cells, which parallels thegrowth curve. LTA and TA were isolated fromJY2190 and Rx1/Al3 grown in the presence ofnon-labelled choline (5 Wg ml31). As shown in Fig.
3, the LTA from the mutant eluted with the propa-nol gradient from octyl-Sepharose as a double peak.Peak A (62% of the phosphorus) contained a chol-ine/phosphorus molar ratio of 0.39. Peak B (choline/phosphorus = 0.19) was approximately 50% contami-nated with membrane lipids (shown by TLC afterBligh^Dyer extraction) and was not further studied.LTA from Rx1/Al3 eluted as a single peak fromoctyl Sepharose and contained a choline/phosphorusratio of 0.52.
Puri¢ed cell walls from both strains were solubi-lised by hydrolysis with CPL, a muramidase, pro-duced by the pneumococcal bacteriophage Cp-1[11]. This hydrolysis yields in addition to non-sub-stituted muropeptides, TA-substituted muropeptides,essentially TA-(GlcNAc-MurNAc)-tripeptide andcrosslinked TA-(GlcNAc-MurNAc)2-tri-tetrapeptide(Fischer, W. and Leopold, K., unpublished results).
LTA samples and muropeptide preparations werechromatographed on an a¤nity column, containingTEPC-15, a ChoP-speci¢c Mab [13,14]. Sampleswere applied in ammonium acetate pH 6.0, ChoP-containing muropeptides were eluted through com-petitive release by triethylammonium ions. The elu-tion pro¢les are depicted in Fig. 4, and analyticaldata are summarised in Table 1. Ten to twenty per-cent of the phosphate was not retained (peak I). This
FEMSLE 8508 3-12-98
Fig. 2. Incorporation of [methyl-3H]choline into mutant cellsgrowing in CDM. Growth was followed photocally. Radioactiv-ity (Bq) was measured as described in Section 2.
Fig. 3. Hydrophobic interaction chromatography on octyl-Seph-arose of crude LTA from Cho-grown mutant cells. Fractions of1 ml were collected and elution was monitored by phosphate de-termination. U5, Magni¢cation factor.
K. Leopold, W. Fischer / FEMS Microbiology Letters 169 (1998) 355^359 357

material did not contain TA, when tested after acidhydrolysis for GalNAc as a marker. LTA- and TA-containing muropeptides eluted in peak II and III.Since peak II showed almost the same Cho/P ratio aspeak III, it contains presumably shorter TA chains.The Cho/P ratios of LTA and TA of each strainwere close to each other and the Cho/P ratios ofLTA and TA from the mutant were 81 and 86%compared with those of Rx1/Al3.
3.2. Incorporation of ethanolamine
The mutant and Rx1 were grown in Etn-contain-ing CDM. Bacteria were processed and cell wallswere prepared and puri¢ed as described previously[12]. Etn was released from cell wall-linked TA byhydrolysis with HF (48% by mass, room tempera-ture, 18 h). After drying and suspending the samplesin 10 mM lithium acetate pH 4, cell walls were re-moved by centrifugation. In the supernatant, Etnwas identi¢ed and quanti¢ed by reverse-phaseHPLC [9]. The Etn/P ratio was 0.47 þ 0.2 (n = 6)for the mutant TA which compares to 0.56 þ 0.06(n = 6) for the TA of the parent strain.
4. Conclusions
The data presented indicate that the Cho-inde-pendent mutant JY2190 retained the capability toincorporate nutritional Cho and Etn as phospho-diester into LTA and TA with a similar e¤ciencyas the parent strain. The capability to incorporateCho into LTA and TA renders the mutant an idealsystem to ¢gure out the pathway of Cho incorpora-tion because any of the presumably four genes, en-coding a Cho transporter and the enzymes of thispathway [16], can be deleted without a¡ecting viabil-ity and growth rate. A number of studies have re-vealed the signi¢cance of ChoP for pneumococcalphysiology. It labels the cell wall substrate for auto-lytic enzymes [8], is necessary for genetic transforma-
FEMSLE 8508 3-12-98
Table 1Analyses of peaks I^III from TEPC-15 a¤nity chromatographyof LTA and TA from the choline-grown mutant and strain Rx1/Al3
Designationa of samplesand characterisation
Peak
I II III
(A) Mutant LTAPhosphorus % 20 20 60Cho/P molar ratio 0.09 0.45 0.43
(B) Mutant TAb
Phosphorus % 21 10 69Cho/P molar ratio n.d.c 0.44 0.49
(C) Rx1/Al3 LTAPhosphorus % 10 0 90Cho/P molar ratio n.d.c 0 0.54
(D) Rx1/Al3 TAb
Phosphorus % 14 0 86Cho/P molar ratio n.d.c 0 0.56
aThe capital letters refer to the panels in Fig. 4.bMuropeptide-linked TA (see text).cn.d., not determined.
Fig. 4. Phosphocholine-speci¢c a¤nity chromatography on aTEPC-15 column. Elution pro¢les of LTA (A) and TA muropep-tides (B) from choline-grown mutant cells in comparison withLTA (C) and TA muropeptides (D) from Rx1/Al3.
K. Leopold, W. Fischer / FEMS Microbiology Letters 169 (1998) 355^359358

tion [6], and serves to anchor a number of ChoP-recognizing proteins to the surface of pneumococciwhich act as virulence factors and adhesins [9,17,18].ChoP residues also play a direct role in pneumococ-cal disease for colonisation of the nasopharynx andthe entrance of pneumococci into lung and vascularendothelial cells [19,20].
Acknowledgments
This work was supported by the Bundesministe-rium fuër Bildung und Forschung (Grant 01KI9401/2/3).
References
[1] Badger, E. (1944) The structural speci¢city of choline for thegrowth of type III pneumococcus. J. Biol. Chem. 153, 183^191.
[2] Rane, L. and Subbarow, Y. (1940) Nutritional requirementsof the pneumococcus. 1. Growth factors for types I, II, V, VII,VIII. J. Bacteriol. 40, 695^704.
[3] Behr, T., Fischer, W., Peter-Katal|ènic, J. and Egge, H. (1992)The structure of pneucoccal lipoteichoic acid. Improved prep-aration, chemical and mass spectrometric studies. Eur. J. Bio-chem. 207, 1063^1075.
[4] Fischer, W., Behr, T., Hartmann, R., Peter-Katal|ènic, J. andEgge, H. (1993) Teichoic acid and lipoteichoic acid of Strep-tococcus pneumoniae possess identical chain structures. A re-investigation of teichoic acid (C polysaccharide). Eur. J. Bio-chem. 215, 851^857.
[5] Fischer, W. (1997) Pneumococcal lipoteichoic acid and tei-choic acid. Microb. Drug Resist. 3, 309^325.
[6] Tomasz, A. (1968) Biological consequences of the replacementof choline by ethanolamine in the cell wall of pneumococcus:chain formation, loss of transformability, and loss of autoly-sin. Proc. Natl. Acad. Sci. USA 59, 86^93.
[7] Hoëltje, J.-V. and Tomasz, A. (1975) Speci¢c recognition ofcholine residues in the cell wall of teichoic acid by the N-acetylmuramyl-L-alanine amidase of pneumococci. J. Biol.Chem. 250, 6072^6076.
[8] Lopez, R., Garc|èa, E., Garc|èa, P. and Garc|èa, J.L. (1997) The
pneumococcal cell wall degrading enzymes: a modular designto create new lysines? Microbial Drug Res. 3, 199^211.
[9] Yother, J., Leopold, K., White, J.A. and Fischer, W. (1998)Generation and properties of a Streptococcus pneumoniae mu-tant which does not require choline or analogs for growth.J. Bacteriol. 180, 2093^2101.
[10] Severin, A., Horne, D. and Tomasz, A. (1997) Autolysis andcell wall degradation in a choline-independent strain of Strep-tococcus pneumoniae. Microb. Drug Resist. 3, 391^400.
[11] Garcia, J.L., Garcia, E., Arraraès, A., Garcia, P., Ronda, C.and Loèpez, R. (1987) Cloning, puri¢cation and biochemicalcharacterization of the pneumococcal bacteriophage Cp-1 ly-sine. J. Virol. 61, 2573^2580.
[12] Heumann, D., Barras, C., Severin, A., Glauser, M.P. andTomasz, A. (1994) Gram-positive cell walls stimulate synthesisof tumor necrosis factor alpha and interleukin-6 by humanmonocytes. Infect. Immun. 62, 2715^2721.
[13] Potter, M. and Leon, M.A. (1968) Three IgA myeloma im-munoglobulins from the BALB/c mouse: precipitation withpneumococcal C polysaccharide. Science 162, 369^371.
[14] Chesebro, B. and Metzger, H. (1972) A¤nity labeling of aphosphocholine binding mouse myeloma protein. Biochemis-try 11, 766^771.
[15] Fischer, W. (1996) Molecular analysis of lipid macroamphi-philes by hydrophobic interaction chromatography. J. Micro-biol. Methods 25, 129^144.
[16] Weiser, J.N., Shchepetov, M. and Chong, S.T.H. (1997) Dec-oration of lipopolysaccharide with phosphorylcholine: aphase-variable characteristic of Haemophilus in£uenzae. Infect.Immun. 65, 943^950.
[17] Yother, J. and White, J.M. (1994) Novel surface attachmentmechanism of the Streptococcus pneumoniae protein PspA.J. Bacteriol. 176, 2962^2985.
[18] Rosenow, C., Ryan, P., Weiser, J.N., Johnson, S., Fontan, P.,Ortqvist, A. and Masure, H.R. (1997) Contribution of novelcholine-binding proteins to adherence, colonisation and im-munogenicity of Streptococcus pneumoniae. Mol. Microbiol.25, 819^829.
[19] Cundell, D.R., Gerard, N.P., Gerard, C., Idanpaan-Helklia,I. and Tuomanen, E.I. (1995) Streptococcus pneumoniae an-chor to activated human cells by the receptor for platelet-activating factor. Nature 377, 435^438.
[20] Kim, J.O. and Weiser, J.N. (1998) Association of intrastrainphase variation in quantity of capsular polysaccharide andteichoic acid with the virulence of Streptococcus pneumoniae.J. Infect. Dis. 177, 368^377.
FEMSLE 8508 3-12-98
K. Leopold, W. Fischer / FEMS Microbiology Letters 169 (1998) 355^359 359