
University of Ghana SCHOOL OF ...
238
SCHOOL OF PUBLIC HEALTH, COLLEGE OF HEALTH SCIENCES UNIVERSITY OF GHANA, LEGON THE ROLE OF GENETIC AND EPIGENETIC FACTORS IN ENDOTHELIAL DAMAGE AND REPAIR AMONG GHANAIAN CHILDREN WITH CEREBRAL MALARIA BY DANIEL AMOAKO-SAKYI (10174124) THIS THESIS IS SUBMITTED TO THE UNIVERSITY OF GHANA, LEG ON IN PARTIAL FULFILLMENT FOR THE REQUIREMENT FOR THE AWARD OF DOCTOR OF PHILOSOPHY (PHD) DEGREE IN PUBLIC HEALTH. MARCH 2019 University of Ghana http://ugspace.ug.edu.gh
Transcript of University of Ghana SCHOOL OF ...

SCHOOL OF PUBLIC HEALTH, COLLEGE OF HEALTH SCIENCES
UNIVERSITY OF GHANA, LEGON
THE ROLE OF GENETIC AND EPIGENETIC FACTORS IN
ENDOTHELIAL DAMAGE AND REPAIR AMONG GHANAIAN CHILDREN
WITH CEREBRAL MALARIA
BY
DANIEL AMOAKO-SAKYI
(10174124)
THIS THESIS IS SUBMITTED TO THE UNIVERSITY OF GHANA, LEG ON
IN PARTIAL FULFILLMENT FOR THE REQUIREMENT FOR THE
AWARD OF DOCTOR OF PHILOSOPHY (PHD) DEGREE IN PUBLIC
HEALTH.
MARCH 2019
University of Ghana http://ugspace.ug.edu.gh

DECLARATION
I hereby declare that this thesis is the result of my own original research, except for
areas where specific references have been made and duly acknowledged. I also affirm
that the studies reported in this docwnent were carried out by me under the supervision
of my team of academic supervisors. Lastly, I declare that this work has not been
submitted, either in part or in whole, to any other institution for an award of a degree.
Prof. Isabella A. Quakyi (PhD) (Academic Supervisor)
Prof.&(PbDl (Academic Supervisor)
Prof. Julius Fobil (PhD) (Academic Supervisor)
Dr Kwadwo Asamoah Kusi (PhD) (Academic Supervisor)
Dr John Arko-Mensah (PhD) (Academic Supervisor)
IJ'V-'~I" Date
\q \ \\J \ (...cJ\,\ Date
University of Ghana http://ugspace.ug.edu.gh

DEDICATION
To Reggie, Eno and Kobby
ii
University of Ghana http://ugspace.ug.edu.gh

ACKNOWLEDGEMENTS
I am eternally grateful to God for the gift of life, strength and wisdom for this pursuit.
I also thank. my employers, the University of Cape Coast for offering me a career
development opportunity that enabled me to enrol in this prestigious doctoral program.
My time with the Department of Biological, Environmental, and Occupational Health
Sciences (BEOHS), School of Public, Health (SPH) has been worthwhile and I cherish
the stimulating academic environment at BEOHS. I thank the Chair of BEOH, Prof.
Julius Fobil and his team for this opportunity.
Majority of the work described in this thesis was conducted in the laboratories of the
Immunology Department, Noguchi Memorial Institute for Medical Research (NMIMR)
and I am thus, indebted the Chair of the Department for granting me access to the
facility and resources. To my academic supervisors, Prof. Isabella Quakyi, Prof. Ben
Gyan. Prof. Julius Fobil, Dr Asamoah Kusi and Dr John Arko-Mensah, I say a big thank
you. Your expert advice, critiques, frank confrontations and love will continue to shape
my life and career.
I thank all students and staff on the EPCmal Study for taking time to train and equip
me with the various skills and techniques I needed for this work. My special
appreciation goes to Thomas Addison for being there whenever I needed him. I thank
the staff of the health facilities that partnered us in this study, your time and efforts are
appreciated. Finally, to all the children and parents who participated in this study, I say
thank you.
iii
University of Ghana http://ugspace.ug.edu.gh

TABLE OF CONTENTS
DECLARATION ............................................................................................................ i
DEDICATION ................................... .. .................................................................... ii
ACKNOWLEDGEMENTS ......................................................................................... iii
TABLE OF CONTENTS .............................................................................................. iv
LIST OF FIGURES ...................................................................................................... ix
LIST OF TABLES ........................................................................................................ xi
LIST OF ABBREVIATIONS .................................................................................... xiii
DEFINITION OF TERMS ........................................................................................ xvii
ABSTRACT .............................................................................................................. xxvi
CHAPTER ONE ............................................................................................................ 1
1.0 INTRODUCTION ................................................................................................... 1
1.1. Malaria in a global health perspective ................................................................. I
1.2. The malaria pathophysiology nexus .................................................................... 2
1.3. An emerging pathophysiologic model for CM ................................................... 5
1.4. Host genetic and epigenetic factors in the pathogenesis of CM ......................... 7
1.5. Problem statement ............................................................................................... 9
1.6. Conceptual Framework ..................................................................................... 10
1.7 Justification of study .......................................................................................... 14
1.8 General objective ................................................................................................ 15
1.9 Specific objectives .............................................................................................. 15
1.10 Hypothesis ........................................................................................................ 16
CHAPTER TWO ......................................................................................................... 17
2.0 LITERATURE REVIEW ...................................................................................... 17
2.1. Malaria as a global health problem ................................................................... 17
2.1.1 Malaria in Ghana: epidemiology ................................................................. 23
2.1.2 Malaria in Ghana: the socioeconomic burden ............................................. 25 iv
University of Ghana http://ugspace.ug.edu.gh

2.2. Malaria: the parasite, vector, and disease ..................................... · .. · .. · ... ········ ... 27
2.2.1. The parasite: exploring Plasmodium parasite biology through the lifecycle
.............................................................................................................................. 29
2.2.2. The disease: pathogenic mechanisms and determinates of severe malaria 35
2.2.2.1 Pathogenesis of cerebral malaria .......................................................... 36
2.2.2.2 New paradigms in the pathogenesis of cerebral malaria ..................... .42
2.3. Markers of endothelial damage and repair ........................................................ 43
2.3.1 Marker of endothelial dysfunction .............................................................. 44
2.3.2 Markers of endothelial repair ...................................................................... 45
2.4 Malaria immunology .......................................................................................... 47
2.4.1 Immune responses during pre-erythrocytic stages of Plasmodium life cycle
.............................................................................................................................. 47
2.4.2 Immune responses during erythrocytic stages of Plasmodium lifecycle .... .48
2.4.2.1 Innate immune responses ...................................................................... 49
2.4.2.2 Adaptive immune responses: humoral immunity ................................. 50
2.4.2.3 Adaptive immune responses: cell-mediated immunity ......................... 52
2.5 Host genetics and epigenetics in malaria pathogenesis ofCM .......................... 53
2.5.1 Genetic disorders of erythrocytes and susceptibility to malaria .................. 54
2.5.2 Malaria immunogenetics ............................................................................. 55
2.5.3 Malaria host epigenetics .............................................................................. 58
CHAPTER THREE ..................................................................................................... 59
3.0 METHODS ............................................................................................................ 59
3.1 Study Site ........................................................................................................... 59
3.2. Study design and sample size estimations ......................................................... 60
3.3. Ethical Considerations ....................................................................................... 61
3.4 Inclusion criteria ................................................................................................. 61
3.4.1 Specific inclusion Criteria ........................................................................... 61
v
University of Ghana http://ugspace.ug.edu.gh

3.4.2 Exclusion criteria ......................................................... ·· .... · .. ·· .. • .................. 63
3.5 Blood S81llple collection ................................................ ·.· .......•.......................... 63
3.6. Sample processing and downstream analysis ..................................... ·· .. ·· .. ·· .... ·64
3.7 Measurement of angiogenic factors ................................................................... 66
3.8 SNP Genotyping ............................................................. · .. · ................................ 66
3.8.1 DNA isolation from whole blood ............................................. ··· ........ · ...... ·67
3.8.2 Pre-PeR: DNA and oligo pool preparation ................................................. 68
3.8.3 peR amplification of target loci .................................................................. 68
3.8.4 peR product clean-up with Shrimp alkaline phosphatase (SAP) protocol.69
3.8.5 iPLEX reaction ............................................................................................ 70
3.9. Other laboratory evaluations ............................................................................. 71
3.9.1 Haematological analysis .............................................................................. 72
3.9.2 Parasitological evaluation ............................................................................ 72
3.9.3 Bacteraemia evaluation ................................................................................ 72
3.10. The use of Gaussian mixture model ................................................................ 72
3.11. Statistical Analysis .......................................................................................... 73
3.12. Dealing with missing data ............................................................................... 75
CHAPTER FOUR ........................................................................................................ 76
4.0 RESULTS .............................................................................................................. 76
4.1. Demographic characteristics of study participants ............................................ 76
4.2 Haematological indices among study participants ............................................. 78
4.3. Parasitological indices among study participants .............................................. 80
4.4 Immunological indices among study population ............................................... 81
4.5 Angiogenic indices among clinical malaria phenotypes .................................... 82
4.6 Endothelial integrity, malaria phenotypes, and angiogenic factors ................... 82
4.7 Association of endothelial integrity with key haematological, parasitological.
immunological and angiogenic variables ................................................................. 86
vi
University of Ghana http://ugspace.ug.edu.gh

4.8. Genotyping results ............................................................................................. 87
4.9 Association ofmg SNPs with endothelial integrity and malaria ........................ 87
4.9.1. Tag SNPs on chromosome 1 ............................................................•......•.. 89
4.9.2. Tag SNPs on chromosome 2 ...................................................................... 92
4.9.3. Tag SNPs on chromosome 4 ...................................................................... 92
4.9.4. Tag SNPs on chromosome 6 ...................................................................... 95
4.9.5. Tag SNPs on chromosome 7 ...................................................................... 97
4.9.6. Tag SNPs on chromosome 9 .................................................................... 100
4.9.7. Tag SNPs on chromosome 10 .................................................................. 104
4.9.8. Tag SNPs on chromosome 16 .................................................................. 107
4.9.9. Tag SNPs on chromosome 20 .................................................................. 109
4.10 Global association plots ................................................................................. 112
4.11 In silico analysis ............................................................................................. 116
4.11.1 Linkage disequilibrium analysis .............................................................. 116
4.11.2. Epigenetic contexts ofSNPs ................................................................... 127
4.11.3. Potential effects of SNPs on Transcription Factors (TF) ....................... 127
4.11.4. Effects of trait- and malaria-associated SNPs on microRNA ................. 137
4.12 Prospecting biomarkers for cerebral malaria ................................................. 140
4.13 Association of SNPs with angiogenic factors ................................................ 145
CHAPTER FIVE ....................................................................................................... 146
5.0 DISCUSSION ...................................................................................................... 146
5.1 Association SNPs with malaria and endothelial integrity: ............................... 146
5.2. SNPs and epigenetic mechanisms: insights from in silico analysis ................ 149
5.3 Endothelial integrity and malaria phenotype ................................................... 150
5.4 Malaria in context: parasitological, immunological and haematological indices .
................................................................................................................................ 152
5.4.1 Parasitological indices ............................................................................... 152
vii
University of Ghana http://ugspace.ug.edu.gh

5.4.2 Immunological indices .............................................................................. 152
5.4.3 Angiogenic factors ..................................................................................... 153
CHAPTER SIX .......................................................................................................... 156
6.0 CONCLUSION AND RECOMMENDATIONS ................................................ 156
6.1 Conclusion ........................................................................................................ 156
6.2 Recommendations ............................................................................................ 157
REFERENCES .......................................................................................................... 159
APPENDICES ........................................................................................................... 197
Appendix I: Ethical Approval ................................................................................ 197
Appendix II: Scientific and Technical Committee ApprovaL .............................. 198
Appendix III: Inform Consent ................................................................................ 199
Appendix IV: Primer for iPLEX Reaction ............................................................. 208
Appendix V: iPLEX protocol. ................................................................................ 21 0
viii
University of Ghana http://ugspace.ug.edu.gh

LIST OF FIGURES
Figure
Figure 1. 1: Conceptual framework
Page
13
Figure 2. 1: Malaria cases by WHO regions ..........................•..................................... 19
Figure 2. 2: Access to ITN in Sub-Saharan Africa ...................................................... 22
Figure 2. 3: Confirmed malaria cases in Ghana 2005 - 2016 ...................................... 23
Figure 2. 4: Malaria admissions in Ghana (2005 - 2016) ............................................ 24
Figure 2.5: Source of funding for malaria interventions ............................................. 25
Figure 2. 6: Ghana government expenditure on malaria .............................................. 25
Figure 2. 7: Lifecycle of the malarial parasites ............................................................ 28
Figure 2. 8: Pre-erythrocytic stage life cycle ............................................................... 32
Figure 2. 9: Pathological differences in CMl and CM2 .............................................. 39
Figure 2. 10: Adaptive immunity at the pre-erythrocytic stage ................................... 48
Figure 3. I: Study site .................................................................................................. 59
Figure 3. 2: From field to data: a schematic flowchart of sample processing ............. 65
Figure 3. 3: iPLEX reaction ......................................................................................... 70
Figure 4. I: Age and malaria phenotypes .................................................................... 77
Figure 4.2: Haematological indices in study participants ........................................... 79
Figure 4.3: Parasitological indices among malaria phenotypes .................................. 80
Figure 4. 4: Histogram of EPCs in the study population ............................................. 83
Figure 4.5: Malaria phenotypes and endothelial integrity .......................................... 84
Figure 4. 6: Endothelial integrity and malaria phenotypes .......................................... 85
Figure 4. 7: Association ofSNPs with CM in a CM vs UM comparison ................. 113
Figure 4.8: Association ofSNPs with malaria in a malaria vs HC comparison ....... 114
ix
University of Ghana http://ugspace.ug.edu.gh

Figure 4. 9: Association of SNPs with ProDamage in a ProDamage vs ProRepair
comparison .......................................................................................................... 115
Figure 4. 10: A composite linkage disequilibrium plot for rsl0489181 .................... 117
Figure 4. 11: A composite linkage disequilibrium plot for rs2070744 ...................... 118
Figure 4. 12: A composite linkage disequilibrium plot for rs3918211 ...................... 119
Figure 4. 13: A composite linkage disequilibrium plot for rs3917419 ...................... 120
Figure 4. 14: A composite linkage disequilibrium plot for rs59055740 .................... 121
Figure 4. 15: A composite linkage disequilibrium plot for rs684951 ........................ 122
Figure 4. 16: A composite linkage disequilibrium plot for rs73422262 .................... 123
Figure 4. 17: Linkage disequilibrium for rs3818256 ................................................. 124
Figure 4. 18: A composite linkage disequilibrium plot for rs943082 ........................ 125
Figure 4. 19: A composite linkage disequilibrium for rs2304527 ............................ 126
Figure 4.20: SNPs and their Chromatin States ......................................................... 129
Figure 4. 21: Performance of angiogenic factors as a biomarker for CM ................. 141
Figure 4.22: Performance of angiogenic factors as a biomarker for endothelial
integrity ............................................................................................................... 142
Figure 4.23: Performance of parasitological indices as a biomarker for CM ........... 143
Figure 4.24: Comparison of3 parasitological variables as biomarkers for CM ....... 144
Figure 4.25: Comparison ofMMP9leveis among rs3918256 in a recessive model 145
x
University of Ghana http://ugspace.ug.edu.gh

LIST OF TABLES
Table
Table 3. 1: PCR cycling program for target amplification
Table 3. 2: PCR cycling program for iPLEX reaction
Table 3.3: Regression models for genetic analysis
Table 4. 1: Association of angiogenic factors with malaria phenotypes
Table 4.2: Association of key variables with endothelial integrity
Table 4.3: Summary ofgenotyped SNPs
Page
69
71
7S
82
86
88
Table 4. 4: Chromosome 1: Association of SNPs with CM in CM vs UM comparison
90
Table 4.5: - Chromosome 1: Association ofSNPs with malaria in malaria vs HC
comparison 91
Table 4. 6: Chromosome 2: Association ofSNPs with malaria in malaria vs HC
comparison. 93
Table 4. 7: Chromosome 4: Association of SNPs with malaria in a malaria vs HC
~ari~ M
Table 4. 8: Chromosome 6: SNPs with endothelial integrity 96
Table 4.9: Chromosome 7: Association ofSNPs with endothelial integrity 98
Table 4. 10: Chromosome 7: Association SNPs with malaria in a malaria vs HC
comparison 99
Table 4. 11: Chromosome 9: Association ofSNPs with endothelial integrity 101
Table 4.12: Chromosome 9: Association ofSNPs with CM in a CM versus UM
comparison 102
Table 4. 13: Chromosome 9: Association ofSNPs with malaria in a malaria versus
HC comparison 103
xi
University of Ghana http://ugspace.ug.edu.gh

Table 4. 14: Chromosome 10: Association ofSNPs with CM in a CM versus UM
comparison lOS
Table 4. 15: Chromosome 10: Association ofSNPs with malaria in malaria vs HC
comparison 106
Table 4. 16: Chromosome 16: Association ofSNPs with endothelial integrity 108
Table 4. 17: Chromosome 20: Association ofSNPs with endothelial integrity 110
Table 4. 18 Chromosome 20: Association ofSNPs with malaria in malaria vs HC
comparison III
Table 4. 19: Effect of SNPs on the binding affinity of transcription factors (TF) 130
Table 4.20: Top 5 transcription factors (TF) impacted by rs2304S27 _T/O 131
Table 4.21: Top 5 transcription factors (TF) impacted by rs39I 82S6_0/A 132
Table 4.22: Top 5 transcription factors (TF) impacted by rs3917419 _O/A 133
Table 4. 23: Top 5 transcription factors (TF) impacted by rs684951_T/O 134
Table 4.24: Top 5 transcription factors (TF) impacted by rs2070744_T/O 135
Table 4.25: Top 5 transcription factors (TF) impacted by rs59055740_T/O 136
rable 4. 26: microRNA-binding sites influenced by rs3918211 [O/A] variants 138
Table 4.27: MicroRNA-binding sites influenced by rs3918211 [O/A] variants 139
xii
University of Ghana http://ugspace.ug.edu.gh

LIST OF ABBREVIATIONS
AIC Akaike Infonnation Criterion
AMA-I Apical Membrane Antigen-l
Angl Angiopoietin-l
Ang2 Angiopoietin-l
ANOVA Analysis Of Variance
BCS Blantyre Coma Scale
CAl Carbonic Anhydrase 1
CBC Complete Blood Count
CD Cluster Of Differentiation
CelTOS Cell
cEPC Circulatory Endothelial Progenitor Cells
CHMI Controlled Human Malaria Infection
CM Cerebral Malaria
CpG S' -C-Phosphate-G-3
CS Cerebrospinal Fluid
CSP Circumsporozoite Protein
CXCLIO C-X-C MotifChemokine Ligand
DAMP Damage Associated Molecular Pattern
DARC Duffy AntigenlChemokine Receptor
DCs Dendritic Cells
DM Diabetes Miletus
DNA Deoxyribonucleic Acid
dNTP Deoxy Nucleotide Triphosphate
EDTA Ethylenediarninetetraacetic Acid
EDV Electron Dense Vesicle
ELISA Enzyme-Linked Immunosorbent Assay
eNOS Endothelial Nitric Oxide Synthase
xiii
University of Ghana http://ugspace.ug.edu.gh

EPCR Endothelial Protein C Receptor
EPCs Endothelial Progenitor Cells
EphA2 Ephrioe Type-A Receptor 2
G6PD Glucose 6-Phosphate Dehydrogenase Deficiency
gDNA Genomic DNA
GES Ghana Education Service
GEST Gamete Egress And Sporozoite Traversal Protein
GHS Ghana Health Services
GLURP Glutamine Rich Protein
G W AS Genome-Wide Association Studies
Hb Haemoglobin
HC Healthy Control
HIV Human Immunodeficiency Virus
HRP2 Histidine-Rich Protein-2
HUVEC Human Umblical Vein Endothelial Cell
ICAM-l Intercellular Adhesion Molecule 1
IE Infected Erthryocyte
IFN--, Interferon Gamma
IL Interleukins
IL-I~ Interleukin-l Beta
IRB Institutional Review Board
ITNs Insecticide-Treated Nets
let-7 Lethal-7
MAHRP2 Membrane Associated Histidine-Rich Protein-2
MgCl2 Magnesium Chloride
MI Multiple Imputation
miRNA Microma
MMP-9 Matrix Metalloproteinase-9
mRNA Messenger RNA
xiv
University of Ghana http://ugspace.ug.edu.gh

MSPI
MSP-119
MSP-3
NORl
NMIMR
NOS3
OPD
PAMP
pbP
PCR
PECAM
PjEMPI
PjHRPl
PL
PLPI
pRBC
qPCR
RBCs
RIFIN
RNA
ROC
RON
RT-PCR
SIOOB
SAO
SAP
SBE
Merozoite Surface Protein
Merozoite Surface Protein-I 19
Merozoite Surface Protein 3
Neuregulin I
Noguchi Memorial Institute For Medical Research
Nitric Oxide Synthase3
Out-Patient Department
Pathogen Associated Molecular Pattern
Peripheral Blood Parasitaemia
Polymerase Chain Reaction
Platelet Endothelial Cell Adhesion Molecule
P. Jalciparum Erythrocyte Membrane Protein-!
P. Jalciparum Erythrocyte Membrane Protein-l
Phospholipids
Perf orin-Like Protein 1
Parasitized Red Blood Cells
Real-Time Polymerase Chain Reaction
Red Blood Cells
Repetitive Interspersed Family Protein
Ribonucleic Acid
Receiver Operating Characteristics
Rhoptry Neck Protein
Reverse Transcription Polymerase Chain Reaction
S 1 00 Calcium-Binding Protein B
Southeast Asian Ovalocytosis
Shrimp Alkaline Phosphatase
Single Base Extension
xv
University of Ghana http://ugspace.ug.edu.gh

SDF-I
SMA
SNP
SPECT
spp
SR-BI
STAT6
STEVOR
SURFIN
TF
TFBS
Tie-2
TLP
TLR-2
TLR-4
TLR-9
TNF
TRAP
TREMl
TSR
UM
UTR
VEGFR2
VSA
WBC
WHO
Stromal Cell Derived Growth Factor 1
Severe Malarial Anaemia
Single Nucleotide Polymorphism
Sporozoite Microneme Protein Essential For Traversal
Species
Scavenger Receptor B 1
Signal Transducers And Activator Of Transcription
Subtelomeric Variable Open Reading Frame Proteins
Surface Associated Interspersed Gene Family Protein
Transcription Factor
Transcription Factor Binding Sites
Tyrosine-Protein Kinase Receptor
Trap-Like Proteins
Toll-Like Receptor-2
Toll-Like Receptor-4
ToU-Like Receptor-9
Tumour Necrosis Factors
Thrombospondin Related Anonymous Protein
Triggering Receptor Expressed On Myeloid Cells I
Type-l Throbospondin Repeat
Uncomplicated Malaria
Untranslated Region
Vascular Endothelial Growth Factor Receptor 2
Variant Surface Antigen
White Blood Cells
World Health Organisation
xvi
University of Ghana http://ugspace.ug.edu.gh

DEFINITION OF TERMS
Allele: One of the different forms of a gene or DNA sequence that can exist at a single
locus.
Apoptosis: Programmed cell death (peD); a process in which cellular DNA is
degraded and the nucleus condensed; then cell is then devoured by neighbouring cells
or phagocytes.
Artemisinin: A class of drugs used for the treatment (not prevention) of malaria usually
as a part ofa combination therapy, derived from the sweet wormwood or Qinghao plant
(Artemisia annua).
Atovaquone: A drug used against malaria. It is found in the combination atovaquone
proguanil which can be used for both prevention and treatment.
Carrier: In human genetics, an individual heterozygous for a mutant allele that
generally causes disease only in the homozygous state. More generally, an individual
who possesses a mutant allele but does not express it in the phenotype because of a
dominant allelic partner; thus, an individual of genotype Aa is a carrier of a if there is
complete dominance of A over a.
Cerebral malaria: A severe malaria syndrome in which infected red blood cells
obstruct blood circulation in the small blood vessels in the brain andlor release
cytokines that disrupt normal brain function.
xvii
University of Ghana http://ugspace.ug.edu.gh

Chi-square (2) test: A statistical test to determine the probability that an observed
deviation from the expected event or outcome occurs solely by chance.
Chromatid: One of the two side-by-side replicas produced by chromosome
duplication.
Chromosomes: Self-replicating structures of cells that carry in their nucleotide
sequences the linear array of genes.
Coma: A decreased state of consciousness from which a person cannot be roused.
Complementarity: The chemical affInity between specifIc nitrogenous bases as a
result of their hydrogen bonding properties. The property of two nucleic acid chains
having bast! sequences such that an antiparallel duplex can form where the adenines
and thymines (or uracils) are opposed to each other, and the guanines and cytosines are
opposed to each other.
Complex disease: A disorder in which the cause is considered to be a combination of
genetic effects and environmental influences.
Denaturation: The separation of the two strands ofa DNA double helix, or the severe
disruption of the structure of any complex molecule without breaking the major bonds
of its chains.
xviii
University of Ghana http://ugspace.ug.edu.gh

Dominance: The expression of a trait in the heterozygous condition.
Downstream Sequences proceeding farther in the direction of transcription, for
example, the coding region is downstream of the promoter.
Elimination: In the context of malaria. reducing all local transmission down to zero
cases within a defined geographic location.
Endonuclease: An enzyme that cleaves the phosphodiester bond within a nucleotide
chain
Epigenetics: Heritable changes to DNA structure that do not alter the underlying DNA
sequence, e.g., DNA methylation.
Epigenomics: The application of epigenetics to the whole genome.
Eradication: In the context of malaria, reducing the number of malaria parasites that
circulate in the natural world to zero.
Exoerythrocytic stage: A stage in the life cycle of the malaria parasite found in liver
cells (hepatocytes). Exoerythrocytic stage parasites do not cause symptoms.
Exoo: Any segment of an interrupted gene that is represented in the mature RNA
product. The protein-coding sequences of a gene.
xix
University of Ghana http://ugspace.ug.edu.gh

G6PD deficiency: An inherited abnormality that causes the loss of a red blood cell
enzyme. People who are G6PD deficient should not take the antimalarial drug
primaquine.
Gene: The basic unit of inheritance. A gene is a segment of DNA that specifies the
structure of a protein or an RNA molecule.
Genetic association: The non-random occurrence of a genetic marker (usually a
particular allele of a polymorphism) with a trait, which suggests an association between
the genetic marker (or marker close to it) and disease pathogenesis.
Genetic heterogeneity: A similar phenotype being caused by different mutations. Most
commonly used for a similar phenotype being caused by mutations in different genes.
Allelic heterogeneity refers to different mutations in the same gene.
Genome: The total genetic material of an organism, i.e. an organism's complete set of
DNA sequences.
Genome-w ide association study (GW AS): A test for the association between genetic
polymorphisms spread evenly over the entire genome, and a disease. Usually at least
300 000 markers are required to adequately cover the genome.
Genotype: The genetic constitution with respect to the alleles at one or more pairs of
genetic loci under observation. The genotype of an individual is the sum total of the
xx
University of Ghana http://ugspace.ug.edu.gh

· . . th hro as distinguished from the genetic mfonnatlOn contamed on e c mosomes,
individual's phenotype (idiotype).
Haploid: A single genome or set of chromosomes (e.g., in human)
Haplotype: A combination of alleles at closely linked gene loci that are inherited
together.
Heterogeneous trait: see Genetic Heterogeneity
Heterozygous: Having different alleles for one or more genes in homologous
chromosome segments, as opposed to being homozygous with identical alleles at these
loci.
Homozygote: An individual possessing a pair of identical alleles at a given locus on a
pair of homologous chromosomes.
Hybridization: The process of joining two complementary strands of DNA or one
each of DNA and RNA to fonn a double-stranded molecule.
Hypnozoite: Donnant fonn of malaria parasites found in liver cells. Hypnozoites occur
only with Plasmodium vivax and P. ovale. After sporozoites (inoculated by the
mosquito) invade liver cells, some sporozoites develop into dormant forms (the
hypnozoites). which do not cause any symptoms. Hypnozoites can become activated
months or years after the initial infection, producing a relapse.
xxi
University of Ghana http://ugspace.ug.edu.gh

Incubation period: The interval of time between infection by a microorganism and the
onset of the illness or the ftrst symptoms of the illness. In malaria, the incubation is
between the mosquito bite and the ftrst symptoms. Incubation periods range from 7 to
40 days, depending on species.
Indoor residual spraying (IRS): Treatment of houses where people spend night-time
hours, by spraying insecticides that have residual efficacy (i.e., that continue to affect
mosquitoes for several months). Residual insecticide spraying aims to kills mosquitoes
when they come to rest on the walls. usually after a blood meal.
Infection: The invasion of an organism by a pathogen such as bacteria, viruses, or
parasites. Some, but not all. infections lead to disease.
Introns: The DNA base sequences interrupting the protein-coding sequences ofa gene.
These sequences are transcribed into RNA but are cut out of the message before it is
translated into protein.
Linkage disequilibrium (LD): Alleles at different loci that are inherited together more
Linkage: Genetic linkage refers to the observation that two or more genes located on
the same chromosome are inherited together. The ratio of being transmitted together
versus being separated al10ws an estimate of their distance from each other
(recombination fraction).
Locus: A specific location on a chromosome.
xxii
University of Ghana http://ugspace.ug.edu.gh

Mutant allele: An allele differing from the allele found in the standard, or wild type.
Null hypothesis: The prediction that an observed difference is due to chance alone and
not due to a systematic cause; this hypothesis is tested by statistical analysis and
accepted or rejected.
Oligonucleotides: Small single-stranded segments of DNA typically 20-30 nucleotide
bases in size which are synthesized in vitro.
Parasitaemia: The presence of parasites in the blood. The tenn can also be used to
express the quantity of parasites in the blood (e.g., "a parasitaemia of 2%").
Phenocopy: A nonhereditary, phenotypic modification (caused by special
environmental conditions) that mimics a similar phenotype caused by a gene mutation.
Phenotype: Observable characteristics of an organism.
Pleiotropy: Genes or mutations that result in the production of multiple, apparently
unrelated, effects at the phenotypic level. For example, patients with phenylketonuria,
caused by mutations in the PAH (phenylalanine hydroxylase) gene, have reduced hair
and skin pigmentation in addition to mental retardation, resulting from toxic levels of
phenylalanine.
xxiii
University of Ghana http://ugspace.ug.edu.gh

Polymorphism (genetic): A chromosome or DNA variant that is observed in natural
populations. A gene locus is defined as polymorphic if a rare allele has a frequency of
0.01 (1%) or more.
Presumptive treatment: Treatment of clinically suspected cases without, or prior to,
results from confirmatory laboratory tests.
Primer: Short, pre-existing oligonucleotide or polynucleotide chain to which new
DNA can be added by DNA polymerase.
Promoter: A region of DNA involved in binding of RNA polymerase to initiate
transcription.
Restriction enzymes: Proteins that recognize specific, short nucleotide sequences in
DNA and catalyse cutting at those sites.
Silent mutation: Mutation in which the function of the protein product of the gene is
unaltered.
Single nucleotide polymorphism (SNP): Heritable polymorphism resulting from a
single base
Structural variant: Structural genomic variation includes any genetic variant that
alters chromosomal Structure, including inversions, translocations, duplications and
deletions.
xxiv
University of Ghana http://ugspace.ug.edu.gh

Synonymous nucleotide change/non-synonymous nucleotide change: A change in
the DNA sequence which does not result in the change in the amino acid sequence, e.g.,
GTf>QTC both code for Valine (Valor V). A nonsynonymous change results in the
coding of a different amino acid (e.g .• GTT>GAT results in Val>Asp).
Trait: Any detectable phenotypic variation of a particular inherited character.
Transcription unit: The distance between sites of initiation and termination by RNA
polymerase; may include more than one gene.
Vector (genetic): In cloning. the plasmid, phage, or yeast chromosomal sequences used
to propagate a cloned DNA segment.
Vector (infection transmission): An organism (e.g., Anopheles mosquitoes) that
transmits an infectious agent (e.g. malaria parasites) from one host to the other (e.g.,
humans).
Vector competence: The ability ofa vector (e.g., Anopheles mosquitoes) to transmit a
disease (e.g., malaria).
Wild type: The genotype or phenotype that is found most commonly in nature or in the
standard laboratory stock for a given organism.
Zoonosis: A disease that naturally occurs in animals that can also occur in humans.
xxv
University of Ghana http://ugspace.ug.edu.gh

ABSTRACT
The declining malaria burden in endemic regions is predicted to increase the proportion
of malaria infections that progress to cerebral malaria (CM). This epidemiologic
scenario appears ominous against the backdrop of a poor understanding of CM
pathogenesis, lack of effective adjunctive therapies, and poor prognosis after onset.
Thus, the need to better understand the pathogenesis of CM has become
more apparent. To better understand the pathogenesis of CM, this study explored both
genetic and epigenetic aspects of the emerging malaria pathophysiologic paradigm,
which pivots on imbalances in endothelial damage and repair in cerebral
microvasculature during P. falciparum infections.
The Sequenom MassARRA Y platform (iPLEX) was used to genotype a focused panel
of 27 single nucleotide polymorphisms (SNPs) in a cross-sectional study involving 221
children. In silico techniques were used to characterize the epigenetic context of SNPs
and assess their potential effect on microRNAs and transcription factors. Immune cells
and angiogenic factors were measured with Human Magnetic Luminex Assay and flow
cytometry, respectively.
A striking find of this study was the association of a CDH5 SNP (rs2304527) and an
MMP9 SNP (rs3918256) with CM and endothelial integrity respectively. CDH5 SNP
(rs2304527) offered protection from CM under the over-dominant inheritance model
assumption and children with the heterozygote T/G genotype were approximately three
times less likely to have CM relative to their colleagues with the IT -GG genotype. On
the other hand, MMP9 SNP (rs3918256) was a risk factor for endothelial damage.
Relative to the reference genotype (GG), children with the AA genotype ofrs3918256
xxvi
University of Ghana http://ugspace.ug.edu.gh

were approximately 4 times more likely to be classified as ProDamage under the
recessive inheritance model. These two SNPs were subsequently found to disrupt the
binding sites of several transcription factors involved in the angiopoietin and tie
signalling pathway. Several other SNPs were found to influence the binding affinity of
transcription factors but only two (rs3918211 and rs20544) affected micro RNA target
sites.
Receiver operating characteristic (ROC) analysis to test the ability of angiogenic factors
to discriminate between malaria and endothelial integrity phenotypes gave middling
results. The best performing angiogenic factor for discriminating eM from UM was
NGRI which had only a 66% chance of accurately discriminating eM from UM.
Similarly, all angiogenic factors performed poorly in discriminating endothelial
integrity phenotypes.
This is the first study to implicate rs2304527 and rs3918256 in the pathogenesis of eM.
Although in silico analysis suggests some epigenetic roles for these SNPs, future studies
may want to further explore their functional roles. Unfortunately, the prospects of using
angiogenic factors considered in this study to discriminate between malaria and
endothelial integrity phenotypes appear dim. Taken together, this study provides
valuable insights on the genetic and epigenetic aspects of endothelial damage and repair
during a P. Jalciparum malaria in Ghanaian children.
xxvii
University of Ghana http://ugspace.ug.edu.gh

CHAPTER ONE
1.0 INTRODUCTION
1.1. Malaria in a global health perspective
Malaria is a global health threat that has existed since antiquity. Its persistence from
prehistoric periods until now attests to the resilience of the etiologic agent, the
efficiency of its transmission and the subtleness of its pathogenic mechanisms. The
Plasmodia parasites responsible for human malaria have exploited anthropoid lifestyles
and survived successive malaria eradication attempts in the past (Carter and Mendis,
2002.2002; Kriefet aI., 2010; Liu et al., 2010; Rich et a1., 2009; White, 2004; World
Health Organization. 2016a). Today, over 200 million people in 91 different countries
are infected with malaria and although this is grim statistics, it actually represents a
drastic reduction in disease burden, especially, in the last two decades (World Health
Organization. 2016a). Ironically, insights from the mathematical modelling of malaria
epidemiology suggests that the current decline in malaria transmission may result in
new epidemiologic scenarios: (i) a shift in malaria burden from younger to older
children (Carneiro et at., 2010), (ii) an increase in the incidence of cerebral malaria
(CM) (O'Meara et at., 2008) and (iii) a change in the population at risk (Bouyou-Akotet
et aI., 2014). These predictions present malaria control stakeholders with novel
challenges that will require a paradigm shift to overcome. Thus, although the decline in
malaria transmission is desirable and should be pursued in earnest, its unintended
consequences should not escape malaria control managers.
University of Ghana http://ugspace.ug.edu.gh

Planning malaria control strategies is encumbered by the far-reaching and sometimes
obscured consequences of the disease burden. Apart from its potentially fatal
consequence, malaria affects the economy, education, child development, maternal
health and the generalliveliboods of endemic communities (Cormier, 2016; Gallup and
Sachs, 2001; Nonvignon et al., 2016; Tang et al., 2017). Thus, control efforts that do
not take cognizance of these complexities and harness expertise from across disciplines
risk failing (Hemingway et aI., 2016). In this regard, the coincidence of a drastic decline
in malaria transmission with the synergistic use of mUltiple malaria control
tools/interventions in the last two decades may be instructive. The cross-disciplinary
nature of malaria control notwithstanding, the role of biomedical research remains
conspicuous and vital. Several effective malaria control tools and interventions in the
past were made possible by breakthroughs in biomedical research and the success of
future strategies still hinges delicately on advances in the field (Baird, 2015;
Hemingway et al., 2016). This thesis focuses on a biomedical question within the
broader malaria problem. Empirical findings from this study may be relevant to current
malaria control efforts and future strategies.
1.2. The malaria pathopbysiology nexus
Malaria is characterised by a wide range of clinical syndromes and disease burdens that
can be partly explained in terms of the pathogenesis of the disease. Whereas some
children infected with Plasmodium Jalciparum remain asymptomatic, others develop
clinical malaria with varying degrees of severity. Clinical malaria may manifest as
uncomplicated malaria (UM) characterised by nonspecific symptoms akin to those seen
in minor systemic viral conditions (headache, lassitude, fatigue, abdominal discomfort
and muscle and joint aches, usually followed by fever, chills, perspiration, anorexia,
2
University of Ghana http://ugspace.ug.edu.gh

vomiting and worsening malaise) (World Health Organisation, 2015). A minority of
children with UM, however, develop severe fonns of the disease which are often
characterised by one or more of the following: coma (cerebral malaria), metabolic
acidosis, severe anaemia, hypo glycaemia, acute renal failure or acute pulmonary
oedema (World Health Organisation, 2015). The progression from UM to severe
malaria (SM) is usually occasioned by poor management of the disease at the initial
stages or a delay in the commencement of treatment (World Health Organisation,
2015). Host and parasite factors may also contribute to the onset and outcome of SM,
but the actual mechanisms involved have remained elusive. The pathophysiology of
severe malaria (SM), especially the CM phenotype, has captured the attention of
scientists for decades. This fixation is justified because, after onset, CM has an
unacceptably high case-fatality and significant functional deficits (Wahlgren et al.,
20 17a; Wassmer et al.. 2015; World Health Organization, 20 16b). Its dire consequences
notwithstanding, decades of research on CM is yet to culminate in a complete
understanding of the pathophysiology mechanisms involved.
The aetiology of CM is most likely multifactorial and the current body of evidence
identifies four key hallmarks: (a) sequestration of IE in the microvasculature, (b)
endothelial activation, (c) a pro-inflammatory immune response and (d) disruption of
BBB. These hallmarks have led malariologists to posit two main hypotheses for the
pathogenesis ofCM: vascular occlusion and inflammatory hypothesis. Both hypotheses
invoke endothelial activation in their respective mechanisms but differ on the cause
thereof. Whereas proponents of the inflammatory hypothesis point to systemic
inflammation akin that seen in sepsis as the main cause of the endothelial activation
seen CM, proponents of vascular occlusion blame sequestration of infected
3
University of Ghana http://ugspace.ug.edu.gh

erythrocytes (IE) for endothelial activation (Stonn and Craig, 2014). A synthesis of the
pathophysiologic events during a P. Jalciparum infection suggests that the two
hypotheses may not be mutually exclusive. During a P. Jalciparum infection
parasitized red blood cells (pRBC) try to avoid splenic clearance by adhering to
endothelial cells in the microvasculature with the help of various adhesion molecules
such as Intercellular Adhesion Molecule I (ICAM-I), CD36 and Endothelial protein C
receptor (EPCR)(Miller, Baruch, Marsh, & Doumbo, 2002). This sequestration is
believed to stimulate the adverse responses that characterise CM i.e. inflammation,
endothelial activation leading to vascular occlusion, disruption of the blood-brain
barrier and apoptosis of microvascular endothelium (Boehme, Werle, Kommerell, &
Raeth, 1994; Miller et aI., 2002; N'Dilimabaka et al., 2014). Although these pieces of
evidence seemingly lean towards vascular occlusion, studies showing endothelial
activation in the absence of microvascular sequestration (Manning et al., 2012; Yeo et
aI., 2010) support the inflammatory hypothesis. The reality of apparently healthy
individuals with high parasitaemia (Clark and Alleva, 2009) and the association of pro
inflammatory cytokines with CM are the other lines of evidence that raises valid
objections about the role of sequestration.
Although previous studies have unravelled several pieces of the CM pathophysiology
puzzle, substantial knowledge gaps persist. Addressing these persistent knowledge
gaps is undoubtedly imperative but doing so without recourse to various theoretical
frameworks may obscure important findings and hinder progress. Thus, whereas the
deciphering of the minute mechanistic process in CM pathogenesis is necessary, a
paradigm shifts in how we conceptualise CM pathogenesis may offer fresh insights into
the CM pathophysiology nexus.
4
University of Ghana http://ugspace.ug.edu.gh

1.3. An emerging pathophysiologic model for eM
An emerging pathophysiologic model for CM explains the development and recovery
from CM in the light of disequilibrium in cerebral microvascular damage and repair
(Gyan et aI., 2009). According to this model, a child's odds of progressing to CM or
recovering from it may be partly dependent on her ability to repair damaged endothelial
tissue in time to restore equilibrium. Although some studies lend credence to the
endothelial damage/repair equilibrium model, it is essentially an untested construct and
remains a hypothesis (Dickinson-Copeland et al., 2015; Gyan et al., 2009; Tetteh,
2014). This hypothesis is however interesting because it refocuses attention on the
details of malaria-induced endothelial damaged (endothelial dysfunction) and the host
mediated mechanisms for repairing damaged microvasculature (vasculogenesis). It
further pushes the frontiers of knowledge and asks several questions pertaining to
endothelial dysfunction and post-natal angiogenesis. It asks if there are yet
uncharacterised mediators of postnatal angiogenesis; whether children with CM have
dysfunctional or insufficient mediators; and whether there is a genetic or epigenetic
explanation for microvascular endothelium dysfunction. The latter of these questions is
particularly interesting because it provides the opportunity to interrogate the issue at
various levels of the genotype-phenotype continuum. To answer these questions, one
will have to first decipher how the global process of endothelial dysfunction and
postnatal angiogenesis play out in the specific context of malaria Subsequently, a
decryption of the roles of growth factors, receptors, adhesion molecules, proteases,
inhibitors. matrix proteins and cytokines will be necessary to gain deeper insights into
how these factors interact at the genomic, epigenomic and physiologic levels to
influence malaria pathogenesis.
5
University of Ghana http://ugspace.ug.edu.gh

This pathophysiologic model hinges on two main factors: endothelial damage and
repair. Whereas studies abound on the endothelial damage (dysfunction) arm of the
model (Desruisseaux, Machado, Weiss, Tanowitz, & Golightly, 2010; Gyan et aI.,
2009; Swanson et al., 2016), very little is known about postnatal angiogenesis or
vascular repair in the context of malaria. Besides the established roles of pre.existing
vascular wall endothelial cells in the repair of damaged endothelium, recent studies
have highlighted the role of other factors such as circulatory endothelial progenitor cells
(cEPC) in the repair of damaged microvasculature (Asahara et aI., 1999). It is now
known that cEPes are incorporated into the sites of microvasculature damage with the
help of stromal cell·derived growth factor 1 (SDF·I) and the matrix metalloproteinase·
9 (MMP·9) during the repair process (Asahara et aI., 1997; Hristov et aI., 2003; Rafii,
2000; Urbich and Dimmeler, 2004). Previous work on eM pathophysiology in
Ghanaian children aligns with the seminal work by Asahara et al on the role of cEPes
in the repair of damaged endothelium (Asahara et aI., 1999; Gyan et aI., 2009). The
study by Gyan et al found that compared to those with uncomplicated malaria,
asymptomatic parasitaemia, or healthy controls, eM patients had lowered cEPe levels
and increased SDF-I levels (Gyan et al., 2009). These findings have given impetus to
the hypothesis that eM develops due to insufficient or dysfunctional cEPe response to
malaria·induced microvascular damage. These studies do not, however, address the
genetic and epigenetic underpinnings of biological dysfunctions and insufficiencies.
Thus, it has become imperative to investigate and identify the genetic and epigenetic
factors that may influence endothelial dysfunction and repair in eM pathogenesis.
6
University of Ghana http://ugspace.ug.edu.gh

1.4. Host genetic and epigenetic factors in the pathogenesis of eM.
There is a wide body of evidence on the role(s) that host genetic factors play in malaria
susceptibility and severity. Majority of these studies report on haemoglobinopathies
with the best-known being sickle cell trait, a-thalassemia, Glucose-6-phosphate
dehydrogenase (G6PD) deficiency, and Duffy antigen receptor negativity (Aidoo et al.,
2002; Ayi et aI., 2008; Baird, 2015; Cholera et aI., 2008; Maier et al., 2003; May et aI.,
2007; Ruwende et aI., 1995; Taylor et aI., 2012; Williams et aI .• 2005). In addition to
haemoglobinopathies, several studies have investigated the role of host immunogenetic
factors and implicated some immunogens in the pathogenesis of CM (Amoako-Sakyi
et aI., 2016; Crompton et aI., 2014; Cserti-Gazdewich et aI., 2011; Hill, 1999; Mazier
et aI., 2000). Although immunogenetics and haemoglobinopathies appear to be the main
drivers of host genetics and malaria susceptibility, candidate gene studies and genome
wide association studies (GWAS) have reported on association between malaria
susceptibility and other host genes that are neither immunogens nor
haemoglobinopathies related (Manjurano et aI., 2015; Ravenhall et aI., 2018). Howbeit,
there is a dearth of knowledge on how host genetic factors may influence endothelial
dysfunction in the context of CM.
Its importance notwithstanding, host genetic factors do not tell the whole story. This is
because gene expression is partly regulated by changes in the DNA sequence at the
genetic level and partly by epigenetic mechanisms including DNA methylation,
chromatin and RNA modifications (Gupta et aI., 2017; Robertson, 2005; Shames,
Minna, & Gazdar, 2007). Modifications to chromatin architecture are classified as
either a non-pennissive (compact chromatin architecture that silence genes) or
permissive (relaxed chromatin architecture that enhances transcription). These
7
University of Ghana http://ugspace.ug.edu.gh

chromatin states play prominent roles in gene expression and can, therefore, affect the
pathophysiology of many disease models (Berger, Kouzarides, Shiekhattar, &
Shilatifard, 2009). Furthermore, aberrations to nucleosomes are possible, and when
they occur, they create epigenetic marks that specifically relates to pennissive or non-
permissive chromatin.
Conceivably, epigenetic mechanisms collaborate with genetic mechanisms to co
regulate gene function in several disease models including malaria (Berger et aJ., 2009)
and this is perhaps a more interesting aspect of epigenetic research. For instance, the
occurrence of single nucleotide polymorphisms (SNPs) within epigenetic marks can
affect chromatin structure at specific genomic locations by modifying methylation
patterns or histone type recruitment (Zaina et aI., 2010). Similarly, SNPs can affect
microRNA and transcription factors by influencing the target sites and binding
affinities respectively (Hu and Bruno, 2011; Moszytiska et aI., 2017; Wang etal., 2013).
Perhaps, the interactions between genetics and epigenetics may offer better
explanations for scenarios where disease-associated genetic variants lie outside
promoters or coding regions (Zaina, Perez-Luque, & Lund, 2010). So far, a
considerable number of studies have implicated host epigenetic mechanisms in the
pathophysiology of some human disease but very few have focused on malaria (Bell et
aI., 2011; Dayeh et a1., 2013a; Gupta et aI., 2017; Wagner et aI., 2014) and even fewer
looked at the interaction of SNPs and epigenetic marks in malaria pathogenesis.
Chromatin marks and/or architecture allows for the segmentation of the genome into
different chromatin states including enhancer, insulator, transcribed, repressed, inactive
and even CpG islands (Blackledge and Klose, 2011). Thus, this study characterised the
8
University of Ghana http://ugspace.ug.edu.gh

epigenetic contexts (chromatin states) of SNPs and explored its relationships with
endothelial integrity, angiogenic, immunological and haematological factors in
Ghanaian children with different phenotypes of P. falciparum malaria.
1.5. Problem statement
Several pathogenic mechanisms have been proposed for CM but none have been
conclusively established (Riggle et al., 2017). The rare nature of the disease and the
inherent limitations of human studies makes it difficult to investigate the
pathophysiology of CM. That said, animal models and in vitro studies offer some
insights into the hallmarks of CM. Existing knowledge suggests that the pathogenesis
of CM begins with the binding of pRBC to brain endothelium, which in tum activates
the endothelium and initiates parasite antigen cross-presentation to cytotoxic T
lymphocytes (CD8+ T cells). The CD8+ T cells are then recruited to sites of the binding
where they employ perf orin-dependent mechanisms to damage brain endothelium and
the blood-brain barrier (BBB) leading to swelling, micro-haemorrhaging and death
(Riggle et aI., 2017; Swanson et aI., 2016). Just as these mechanisms suggest a
prominent role for Cytotoxic T lymphocyte (CD8+ T cells), they also posit roles for
endothelial cell surface receptors and other factors that aid in the pRBC-endothelium
interaction (Hansen et aI., 2007; Chen et aI., 2000; Schumak et aI., 2015; Nitcheu et aI.,
2003).
Surviving a P. Jalciparum infection heavily hinges on a child's ability to repair
damaged endothelial cells in a timely manner to restore equilibrium and maintain
endothelial integrity (Gyan et al., 2009). Generally, damaged endothelial cells are
repaired through the replication of existing endothelial cells at the site of injury or by
9
University of Ghana http://ugspace.ug.edu.gh

bone marrow-derived circulating endothelial progenitor cells (Asahara et a1., 1997).
Thus, factors that affect the mobilization, release and eventual integration of EPCs into
sites of endothelial damage are important for CM and other diseases that involve
vasculopathology. So far only a few angiogenic factors have been studied in the context
of endothelial damage/repair in CM pathogenesis (Adukpo et al., 2016; Gyan et a1.,
2009). Furthermore, these studies have often failed to explore how host genetic and
epigenetic factors influence the production of these molecules and endothelial integrity.
This study investigates how dozens of factors grouped either as angiogenic,
haematological or immunological influence endothelial integrity during a P. Ja/ciparum
infection. The study further explores how SNPs and their epigenetic contexts influence
the production of these factors and subsequently, endothelial integrity. This study
overcomes the problem of in vivo assessment of endothelial integrity by fitting EPC
data to a Gaussian mixture model to create a binary endothelial integrity variable with
pro-damage and pro-repair as the possible outcomes. The study also used the concepts
of tagged SNPs to glean information from genomic locations in linkage disequilibrium
with genotyped variants. Thus, beyond studies that just seek to associate EPCs, and
angiogenic factors with malaria phenotypes, this study generates a wealth of data that
can be analysed at several points in the genotype-phenotype continuum. The fmdings
of this study could have implications for CM pathogenesis especially in the adjunctive
therapy efforts and the search for prognostic biomarkers for CM.
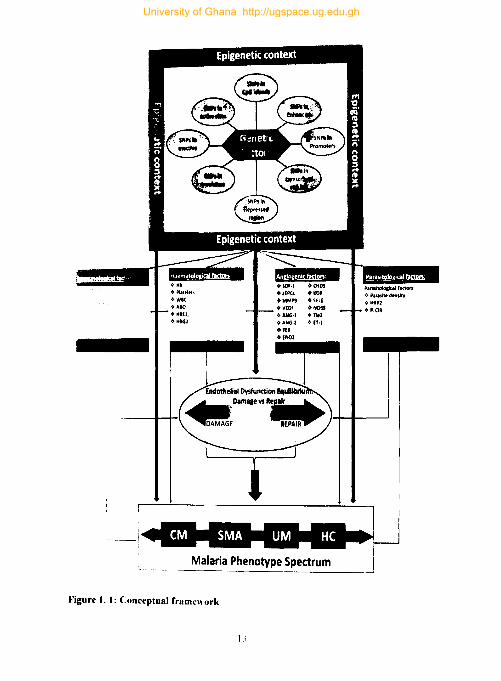
1.6. Conceptual Framework
The theoretical framework undergirding this study conceptualises CM as
cerebrovascular pathology and explores the notion that disequilibrium in damage/repair
10
University of Ghana http://ugspace.ug.edu.gh

of brain endothelium during P. falciparum malaria infection partly governs the
pathogenesis of CM. The discourse on this pathophysiologic model in the preceding
sections of the chapter unveils dozens of key mediators that are used as variables in this
conceptual framework. These variables loosely fall under four groups (angiogenic,
haematological, immunological or parasitological factors) and the framework explores
relationships between these factors, endothelial integrity and subsequently malaria
phenotypes. Endothelial integrity is at the heart of this study as a bivariate outcome
(pro-damage and pro-repair). This outcome variable is obtained by fitting empirically
measured EPCs to a Gaussian mixture model to allow for dichotomization. The use of
EPCs in this regard is reasonable because EPCs have been shown to be good markers
of endothelial dysfunction (Gyan et al., 2009; Taguchi et ai., 2008; Venna et al., 2017;
Werner et al., 2005).
While a number of studies report on associations of angiogenic, immunological, and
haematological factors with CM pathophysiology (Adukpo et aI., 2016; Boufenzer et
al.. 2012; Gyan et al., 2009; Machado et al., 2006), the genetic and epigenetic
mechanisms that regulate the productions of these factors are yet to be explored. In this
conceptual framework, genetic factors (SNPs) with specific epigenetic contexts are
hypothesized to influence microvascular endothelial integrity via influencing the
production of the biomolecules they encode (i.e. angiogenic factors, haematological
factors, and immunological factors). Thin blue lines in Figure 1 map these hypothesized
relationships. Red lines represent relationships that are plausible but not explored in
this study. The study also envisages the possibility of a direct association of genetic
factors with endothelial integrity and such associations are mapped with medium
11
University of Ghana http://ugspace.ug.edu.gh

weight blue lines. Associations between endothelial integrity and malaria phenotypes
are mapped with a thick blue line.
Malaria phenotype spectrum is an important outcome variable in this conceptual
framework. Study participants are categorized into four (4) groups - uncomplicated
malaria. severe malarial anaemia, cerebral malaria, and healthy controls. This allows
for case-case and case-control comparisons. Although the framework explores the
relationships malaria phenotypes may have with other variables, it does that with
caution when it comes to the association of SNPs with malaria phenotype because of
sample size constraints.
12
University of Ghana http://ugspace.ug.edu.gh
+ TEl +fNOI
Malaria Phenotype Spectrum
Figure 1. I: Conceptual framework
University of Ghana http://ugspace.ug.edu.gh

Taken together, explaining CM in the light of microvascular damage and repair is an
emerging pathophysiological model that needs further clarification. Using EPCs as
markers of endothelial dysfunction and a focused panel of SNPs with a well
characterised regulatory and epigenetic context, this study explored relationships
between SNPs, some key mediators of endothelial cell damage/repair, endothelial
integrity and malaria phenotypes in Ghanaian children with CM.
1.7 Justification ofstudy
Majority of children infected with P. Jalciparum malaria remain asymptomatic or
develop UM. However, a minority (about 2%) may progress to severe malaria which
sometimes manifests as SMA or CM (Greenwood et ai., 1991). Emerging empiric
evidence and mathematical modelling suggest that the decline in malaria transmission
in hitherto endemic regions will present fresh epidemiological scenarios that could alter
the status quo in malaria epidemiology and control (Nkumama et aI., 2017). One such
scenario instanced by the decline in malaria transmission is the change in malaria's
clinical epidemiology to favour higher incidence ofCM (Ceesay et aI., 2008; Flirnert et
aI., 2014; O'Meara et aI., 2008). This predicted increase in the proportion of CM is
ominous because, after onset, mortality from CM is high (15 - 25%) regardless of
treatment with anti-malarial drugs (World Health Organization, 2016b). Worse still,
25% of infected children who recover from CM may suffer deficits in cognition,
hearing, vision or develop epilepsy (Gupta et aI., 2017; Wahlgren et al., 20 17; Wassmer
et ai., 2015). The changing clinical epidemiology of malaria and its consequences
notwithstanding, reliable prognostic markers and effective adjunctive therapy for CM
are still unavailable. This situation brings to the fore the need for a better understanding
of CM pathogenesis.
14
University of Ghana http://ugspace.ug.edu.gh

I.S General objective
General objective
This aim of the study is to genotype a focused panel of 27 SNPs, characterized their
epigenetic context and explore their relationships with endothelial integrity, clinical
malaria phenotypes, angiogenic, and immunological factors.
1.9 Specific objectives
The objectives of this study are to:
1. Genotype and epigenetically characterize a focused panel of 27 SNPs in the study
popUlation.
2. Explore the relationships between the focused panel of 27 SNPs, endothelial
integrity and malaria phenotypes.
3. Use in silico tools to determine the potential influence of trait-associated SNPs on
the binding sites of microRNAs and transcription factors.
4. Determine the association of trait-associated SNPs with serum levels of the
angiogenic factors they encode.
5. Explore the relationship between malaria phenotypes, endothelial integrity, and key
clinical, haematological, immunological, parasitological and angiogenic factors.
IS
University of Ghana http://ugspace.ug.edu.gh

1.10 Hypothesis
This study hypothesises that mutant variants of a focused panel of27 SNPs compromise
brain endothelial integrity and subsequently increase the risk of eM in Ghana children
with P. Jalciparum malaria.
16
University of Ghana http://ugspace.ug.edu.gh

CHAPTER TWO
2.0 LITERATURE REVIEW
2.1. Malaria as a global health problem.
The term "global health" arguably suffers an identity crisis. Academics and
practitioners have contented its definition and struggled to distinguish it from allied
disciplines such as public health, international health, planetary health and the likes
(Bettcher and Lee. 2002; Hoffman and Cole, 2018; Lerner and Berg, 2017). In spite of
a contentious definition and blurry boundaries, there seem to be the consensus that a
global health issue should be one that is transnational and require a collaborative
research and health promotion efforts to curb (Beaglehole and Bonita, 2010; Koplan et
al.. 2009). Thus, a review that seeks to establish the status of any disease as a global
health problem should proffer insights into its transnational distribution and the need
for collaborative research efforts in curbing the disease. In a bid to justify the status of
malaria as a global health problem, section 2.1 of this thesis reviews the global
distribution of malaria, outlines international partnerships in malaria control efforts, and
highlights the threats to malaria elimination aspirations.
One of the most palpable features that characterise malaria as a global health problem
is its distribution. Latest malaria burden estimates show the persistence of the disease
in 91 countries with 219 million cases and 4510000 deaths (World Health Organisation,
2018). Compared with estimates from 2010 through to 2015, the 2017 estimates
represent a decline in the burden of malaria. This decline notwithstanding, an annual
disease burden of 219 cases in over 90 countries is enough to earn malaria the tag of
17
University of Ghana http://ugspace.ug.edu.gh

global health problem. More troubling, however, is the evidence suggesting that the
much-touted decline in malaria burden is actually beginning to stall. For instance, there
were about 2 million more cases of malaria in 2017 relative to 2016 (World Health
Organisation, 2018; World Health Organization and Global Malaria Programme, 2017).
The global status of malaria is self-evident and somewhat a mundane find; what is
striking, however, is the disproportionate global distribution of the disease. Currently,
90% of all malaria cases and deaths occur in the WHO African region and even within
this region. Sub Saharan Africa bears about 80% of the burden (World Health
Organization and Global Malaria Programme. 2017) (Fig 2.1 ). Ironically, the latest data
on malaria suggests that the largest gains in malaria decline did not occur in the WHO
African regions with the highest burden (World Health Organization and Global
Malaria Programme, 2017). This observation subtly suggests a possible misalignment
of efforts and endemicity.
Another feature that marks malaria as a global health problem is the nature and source
of investments made towards curbing the disease. Traditionally, the bulk of the funding
for malaria control programs activities in endemic regions has come from international
partners. In 2016 for instance, only 31 % of expenditure on malaria control activities
came from endemic countries (World Health Organization and Global Malaria
Programme, 2017).
18
University of Ghana http://ugspace.ug.edu.gh

University of Ghana http://ugspace.ug.edu.gh

Although funding for malaria has remained stable since 2010, the US$ 2.7 billion
invested in 2016 is less than half (41%) of the funding required for meeting Global
technical strategy for malaria 2016-2030 (GTS) targets. Worse still, the funding
available per person at risk has reduced to below US$ 2 in 34 out of the 41 high-burden
countries (World Health Organization and Global Malaria Programme, 2017).
Investment in proven interventions, tools, and strategies is the best way to ensure that
GTS is on track and thus, it worrying to see investments in malaria control dwindle.
Although individual malaria-endemic nations may face unique challenges in their
efforts to eliminate malaria, some of the challenges are common to all. These challenges
may include lack of sustainable funding, the emergence of drug and insecticide
resistance, climate change, and political instability. Although all these are formidable
threats, the generation and spread of drug- and insect resistant parasites and vectors
pose the most threat of rendering current effective control measures redundant. In this
regard, the increasing prevalence of histidine-rich protein-2 gene (HRP2) deletions in
parasites are potentially deleterious to the rapid diagnosis of malaria (Koita et aI., 2012).
Dealing with the aforementioned threats often requires a collaboration between
endemic countries and international partners which is another testament to the global
status of malaria.
The transnational nature of malaria control efforts is another aspect of malariology that
reflects the global health status of malaria The malaria control toolset has interventions,
tools and strategies that could prove instructive in the fight against malaria if deployed
effectively in endemic regions (Hemingway et aI., 2016). However, the deployment of
these interventions be they diagnostics, medicines, insecticides, or surveillance systems
20
University of Ghana http://ugspace.ug.edu.gh

is often suboptimal. For instance, insecticide-treated mosquito nets (ITNs) which form
the foundation of malaria prevention in Sub Saharan Africa reached only 54% of the
population at risk in 2016 (Fig 2.2). Although this shows ITN cover only about half of
the population at risk, it is noteworthy that this represents an increase from 30% in 2010
(Bhatt and Gething, 2014; World Health Organization and Global Malaria Programme,
2017). On the other hand, the proportion of the population protected by indoor residual
spraying (IRS), the only other vector control measure, shrunk by about 2.6% between
2010 and 2016 (pluess et al., 2010; World Health Organization and Global Malaria
Programme. 2017). Variations in country-level commitment and capacity to implement
malaria control strategies affect coverage of control strategies in endemic areas. The
resultant mosaic nature of malaria control efforts in these sub-regions eventually waters
down the efforts of compliant nations.
Several other factors that are not discussed here, such as, access to health facility and
medications, accurate diagnosis, robust surveillance systems and political will
contributes to making malaria global health issue (Eisele et al., 2010; World Health
Organization and Global Malaria Programme, 2017). This section has reviewed malaria
as a global health problem and in so doing touched on malaria epidemiology and control
strategies. Taken together, the review reveals that the burden of malaria may have
reduced drastically in the last decade, but warns of the risk of a possible reversal of
gains if investments in malaria control are not increased. In addition, the variable
disease distribution and uptake of interventions suggest that endemic countries must
play lead roles in malaria elimination efforts to bolster the odds of success.
21
University of Ghana http://ugspace.ug.edu.gh

~ N
'" N ;S < I: ~ r.. ~ .s: ~
rJ:J .s: = '" .5
~'~ z t: .s ~ QI
</" CJ CJ
<
t N N QI r.. = I:)Jl
i;:
University of Ghana http://ugspace.ug.edu.gh

2.1.1 Malaria in Ghana: epidemiolo~
Located along the Gulf of Guinea between latitudes 4° and 12°N and longitudes 4°W
and l°E. Ghana has a malaria epidemiological profile typical of the West African Sub-
region. It is a high malaria transmission area (> 1 case per 1000 population) with almost
all malaria cases caused by P. jaiciparllm and transmitted either by Anopheles gambiae.
Anopheles fllnesflls. or Anopheles arahiensis. Although malaria cases and deaths
reported at health facilities in Ghana in 2016 stood at 4,535,167 and 1.264 respectively,
the number is estimated to be 8,060.000 [5.300.000-11,950,000] and 12.880 [11.510-
14.2501 respectively (World Health Organization and Global Malaria Programme,
2017). fhere appears to be a steady rise in malaria cases in Ghana since 2005 with the
highest recorded in 2016 (Fig 2.3) but it is unclear whether the increase represents an
actual rise in the number of cases or a result of improved surveillance systems.
Whereas malaria admission in Ghanaian health facilities has increased in the last 12
years. mortality for the same lime period seems to have decreased (Fig 2.4) .
• • Figure 2. 3: Confirmed malaria cases in Ghana 2005 - 2016
23
University of Ghana http://ugspace.ug.edu.gh
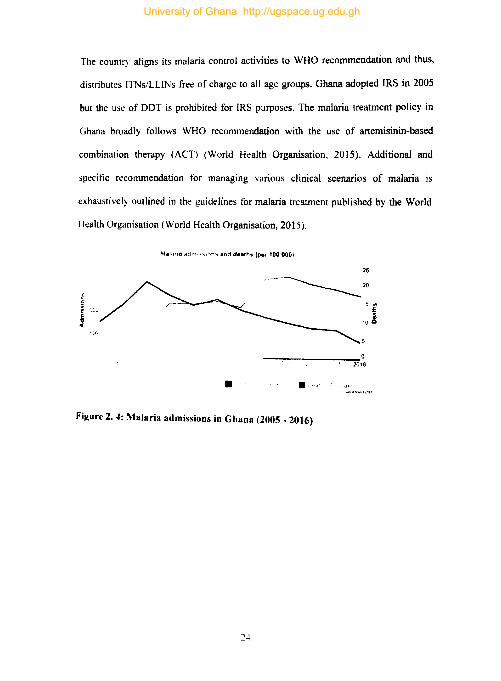
The count!") aligns its malaria control activities to WHO recommendation and thus,
distributes ITNslLLINs free of charge to all age groups. Ghana adopted IRS in 2005
but the us~ of DDT is prohibited for IRS purposes. The malaria treatment policy in
Ghana broadly foIlows WHO recommendation with the use of artemisinin-based
combination therapy (ACT) (World Health Organisation, 2015). Additional and
specific recommendation for managing various clinical scenarios of malaria is
exhaustively outlined in the guidelines for malaria treatment published by the World
Health Organisation (World Health Organisation, 2015).
lr1alartft Bdlfl'''I''' )'1'" and d.ath. (Pel 1000001
25
________ 20
2016
• . ... ,., Figure 2. 4: Malaria admissions in Ghana (2005 - 2016)
2-+
University of Ghana http://ugspace.ug.edu.gh

I • ,. • -':1.'.17 2tK>"
I I t J j I I I • ."1', .'01: 201 \ JII.t 201~ 2016
t:il!1 ' j"ll:' liMn",,' • lMiOlNCEF • Olhers
Source' world report 2017
Figure 2. 5: Source of funding for malaria inten'entions
Government financing of malaria control activities seemed to have increased from 2005
through to 2010 and stalled aftel"\\ards. Interestingly, all of the government's
imestment in malaria control activities goes in the purchase of antimalarial and
remuneration statT (Fig 2.6).
Ghana government expenditure by Intervention
• · '. • .' i.I
Figure 2. 6: Ghana go\'ernment expenditure on malaria
2.1.2 \talaria in Ghana: the socioeconomic burden
Besides its clinical burden. malaria imposes a myriad of direct and indirect socio-
economic burden in endemic countries. At the household level, it is estimated to cost
Ghanaian households about US$ 14 to treat an episode of malaria (Asante and Asenso-
Okyere, 2003: Dalaba et aI., 2014; Nonvignon et aI., 2016: fawiah et aI., 2016). On the
25
University of Ghana http://ugspace.ug.edu.gh

cooperate front, businesses in Ghana lost an estimated US$6.58 million to malaria in
2014 (Dalaba et al., 2014; Nonvignon et al., 2016; Tawiah et aI., 2016). Around the
same period, one of the foremost mining companies in Ghana reported that it incurs
US$S5,000 per month in the treatment of malaria in their employees and dependants
(Anglogold Ashanti, 2004). At the national level, a seminal work by Asenso-Okyere
and colleagues in the early 2000s posited that a percentage rise in the incidence of
malaria reduces productivity by 0.41% (Asante and Asenso-Okyere, 2003). Although
these studies offer some insights into the socioeconomic burden of malaria in Ghana,
the studies are old and heavily lopsided in favour of economics.
Epidemiologist and health economics often describe the socioeconomic burden of
disease in quantitative terms only and ignore the social and cultural dimension of
disease burden. On the contrary, "burden" is a sociocultural contrast and thus,
estimations of socioeconomic burdens that do not incorporate anthropological
perspectives could be narrow and misleading (Jones and Williams, 2004). From this
perspective, "burden" is not just a quantity; it has meaning too. The meaning of malaria
burden will differ among sociocultural contexts but in most African communities,
malaria phenotypes are perceived as different disease entities and not a continuum.
Variations in the sick roles for different malaria phenotypes and the associated social
vulnerabilities may differentially influence malaria interventions and control efforts in
different communities (Jones and Williams, 2004). This notwithstanding, there is a
dearth of knowledge in sociocultural dimensions of malaria burden in Ghana).
26
University of Ghana http://ugspace.ug.edu.gh

2.2. Malaria: tbe parasite, vector, and disease
Malaria is a vector-borne infectious disease transmitted by the bite of the female
anopheles mosquito. The human host, the insect vector, and the protozoan parasite are
pivotal factors in the transmission of malaria and a better understanding of the
interaction between these factors and the environment is instructive in the dash for
elimination. Although the schematics of how malaria is transmitted is now common
knowledge, the research that incrementally unravelled the lifecycle of malaria spanned
almost a century (Cox, 2010; Guillemin, 2002; King, 1883; Krotoski et al., 1982;
Lawrie, 1898; Manson, 1898; Shortt and Garnham, 1948). On the surface, the lifecycle
of malaria appears straightforward and well describe (Fig 2.7), but advances in
molecular biology, cell biology, genomics, and epigenomics has opened up a Pandora's
box of new knowledge, some of which could be instructive in the search for novel
therapies, vaccines and management of severe complications of malaria. Using the
schematic lifecycle as a guidepost, this section reviews the literature on parasite
biology, vector biology, and disease phenotypes in an integrated manner.
27
University of Ghana http://ugspace.ug.edu.gh

.~:.j :~
~~$: F Oocy.'
r"'-;"--;"equlto blood n10.'
~pr. Ory'hr;;cvtl!hU;"a:~-;"'~';CtlOn ~-r 'nl< , 'I""" t I ...... ,- VII.'..... I .t I ••
11: .... .,'"." .... , .,.
~ ... "It_ ... ~ ~
~ ". "-~ ... ~ pv_
.h ........... "" •. · , iL\1 • '\ I
JIm. : .. a q ... :{ " . .,. • ..-
~ "..,Idgut .1 .. ~ ..
_ .~ _____ ~1__~ __ ~
C Asexual ery'hrOCY~'C a'.ge ,. '.1. "'"~1.1''' ·IM '1(.""5
'" VII...... ....8 "'n,'~H
0- fItJl M_:~,
"'.-~ ....... ~~~ ~1,l
r In'I~3I-6 .. ylh..-O(_V1lc galTtetocyt_ devetopn"tent I r-- , ;.': .':.: ~'~ ':"·"L .. .'J:, ~ •.. u_.v .... ..~ ••• - E .. ..- II
t ,;.." (" v--:' .. 't": , .' r' .. "'~ti~J-.... :J- - . .,... --- ........ I - ... ----
(J\) Malaria infe( lion h initidltrd with the Injection of "'POI ol'oltes (!>opz,,) Into the del rni!\. by a fe~din& female anopheline mosquito. (B) The spzs enter the vascul.aturL' ... nd are transported to the liver. wht..~, e they eXit the' !>lnu!tOld5 throul" Kupffer or endothelial cell$ and enter a hepatocyte. Active Invasion IS. prt!t:\ .. ded by <.ellular lr.jver~dlllntil a s.uitable hepatocyte is. found. They form a PVM and under80 schlzoeonv until tens. af thousands of d .. u8hter nl(~r()~(1It(~5 are releas.ed In pdckets. of merosorne'!> Into the vasculature. (C) tIl( it' th.-v enc.ountel ~rythrocvtes dnd bp£ln .1 chronic cycte of as.exual schizogony in the bloodstream, (D) A pr L.portion of asexually rrproduc.tng merozoltes. are reproarammed to undergo gametocytoe.enesis. ([) Witliln a 15 ct.-Jy period. eametoc.ytes sequester and develop within the bone marrow and. once mature. enter the peripheral circulation for ingestion by a mosquito where thpv e-mprge as. extracE"lIular malE" and female gametes In the mideut. (f) Mato)&; Oleurs by fusion of nliero- clnd macrogamele to for-m a zYaote transforming over 24 hr into a ookinete that migrates throueh th~ mosquito m6dKut epithelium and encysts to become an oocyst where asexual sporogenic replication occurs. Motile s.por-ozoltes are released into the hemocoel by oocyst rupture and pass Into salivary glands where thry can be injected into the next human host.
Cowtnan" Healer, Marapana" & Marsh" 2016
Figure 2. 7: Lifecycle of the malarial parasites
28
University of Ghana http://ugspace.ug.edu.gh

2.2.1. The parasite: exploring Plasmodium parasite biology through the lifecycle
Human malaria is caused by protozoan parasites belonging the genus of Plasmodium.
There are over 100 species of Plasmodium but only five infect humans. Four of the five
Plasmodium spp. (P. ovale. P. malariae. P. Jalciparum. and P. vivax) are natural
pathogens of humans whilst the fifth (P. knowles;) is a natural pathogen to macaques
but can cause zoonotic malaria. Almost all malaria cases in African are caused by P.
Jalciparum but a significant fraction of malaria cases in the Americas, South East Asia,
Eastern Mediterranean Regions, and the Western Pacific Regions of are caused by P.
vivax. The other Plasmodium spp. have limited epidemiologic significance.
Pre-erythrocytic parasite/orms
During a blood meal. the female anopheles mosquito injects sporozoites into the dermis
of the skin of a human host. A minority of these sporozoites glide through the dermis
of the skin to locate and penetrate blood vessels but the fate of the majority of
sporozoites that remain in the dermis is not fully known (Prudencio et at., 2006).
Plausibly, inflammatory responses involving polymorphonuclear neutrophils and
inflammatory monocytes in the skin destroy these sporozoites and drain them through
the lymphatics (Mac-Daniel et at., 2014). The sporozoites that make it to the
bloodstream locate blood vessels through homing mechanisms involving factors such
as Trap-like proteins (TLP). Trap-like proteins (TLP) are believed to play significant
roles in this homing mechanism because TLP-mutant sporozoites are unable to enter
the bloodstream even though they have similar motility rates and patterns as TLP-wild
type sporozoites (Cowman et at., 2016). Once in the bloodstream, sporozoites do not
invade red blood cells but quickly move to the liver and traverse it. The process of
migrating through the sinusoidal barrier of the liver which comprises fenestrated
29
University of Ghana http://ugspace.ug.edu.gh

endothelial cells and kupffer cells is referred to as traversal (Tavares et ai., 2013).
Several proteins are required for successful traversals but the best-known are SPECT
(sporozoite microneme protein essential for traversal), phospholipase (PL), PLPI
(perforin-Iike protein I), CelTOS (cell traversal protein for ookinetes and sporozoites)
and GEST (gamete egress and sporozoite traversal protein) (Bhanot et al., 2005; Ishino
et aI., 2004; Risco-Castillo et al., 2015). Besides clues that PLPI uses a membrane
attack complex to punch holes in membranes, the specific roles of these proteins in
traversal are yet to be fully clarified (Cowman et ai., 2016). Some scientists have argued
that traversal somehow prepares sporozoites for hepatocyte invasion and as much as
that is plausible. the primary function of traversal, which is the crossing of the
sinusoidal barrier, cannot be downplayed. In P. jaiciparum, traversal is achieved
through transient vacuoles and regress is via pH and PLPI sensing (Risco-Castillo et
ai., 2015; Tavares et aI., 2013).
In the skin, sporozoites are in the "migratory mode" and have to be converted into
"invasion mode" before they can invade hepatocytes. The switching from migratory to
invasive modes is regulated by several signals but the roles of highly-sulfated HSPGs
and a calcium-dependent protein kinase are critical. Whiles the former activates
sporozoites for invasion, the latter is critical for the switch to an invasive phenotype
(Coppi et aI., 2007; Cowman et ai., 2016). The circumsporozoite protein (CSP), TYPE
I thrombospondin repeat (TSR), thrombospondin-related anonymous protein (TRAP)
and apical membrane antigen-l (AMA-l) are other key proteins involved in hepatocyte
invasion (Cowman et aI., 2016; Herrera et aI., 2015). Important receptors on hepatocyte
required for invasion include the tetraspanin CDS 1 and scavenger receptor B 1 (SR-B I)
but not the famed EphA2 which appears to have no role I'n th·· b e lOvaslon process ut
30
University of Ghana http://ugspace.ug.edu.gh

crucial for intra-hepatocytic development (Kaushansky et al .• 2015). After establishing
liver infection. sporozoites transform into liver-stage parasites or what is referred to as
exo-erythrocytic form. and finally. into merozoites that are budded off in vesicles called
merosomes into the hepatic portal system (Burda et al.. 2017; Graewe et aI.. 2011).
Taken together. it should be noteworthy that the transition from skin sporozoites to
merosome merozoites involves complicated mechanisms. signalling. gene expressions
we do not as yet fully understand. Traditionally. research has focused on blood-stage
infections because of its association with clinical symptoms but recent insights suggest
that liver-stage infections may hold the key to novel therapeutic and vaccinology
approaches (Derbyshire et al.. 2012).
31
University of Ghana http://ugspace.ug.edu.gh

A
);010 i I
:)
( , ,.:
...... ]Ia C) I )
.... _ ...................... .". .... ... '~, 1...") •• ",
J' ,..". :~o~ ~, £)
A Pla>rtlod,un. sporowlles (g"'"n) are depos,ted under the skIn of the human host through the bite of an infected fE-male ilnopheles mosquIto After InjectIOn mto the skin, the sporololtes move through the dermis until they contact blood vessels (red) and move into th" Circulatory system, which allows them to travel to the Itver, Majoritv of sporozoites remain In the dermIS and a small proportion of sporoloites <an enter the Iymphahc ,ystem (vellow),
Il Once th .. 'I'orOlolles (green) ... ach the lover "nusold" Ihey glid .. over the endotheloum and Interacl WIth heparan ,ulphate proteoglycans (HSPGs) from hepatocyte and strllate cells TheV then cross the sinusoidal laver, posslblV through Kupffer cells as shown
C Once the sporoloite (green) has crossed the sinusoidal layer and entered a hepatocyte, it subsequently traverses several hepatocytes until it becomes established in one, In which a parasitophorous vacuole is formed. Each Invading sporololte develop, and Inultlplies in, ide a hepatocyte. forming the ,chllont, which Is made up of thousands of merOlolt",
D. The final ,.II!p involve, til£' release of meroloites (green) Into the bloodstream. The signal(s, that trigger the release remain unknown, Plasmodium meroloites are released bV the formation of merozoite·filled vesicles (merosomes), which bud offfrom the infected hepatocytes into the sinusoidal lumen
- (Prudlnclo, RodrlfIW~, .. Mora, ZOO6J
Figure 2. 8: Pre-Cl),throcytie stage life cycle
32
University of Ghana http://ugspace.ug.edu.gh

Erythrocytic parasite forms
In broad strokes, the cardinal events in the erythrocytic phase of the lifecycle of
Plasmodium parasites are invasion of erythrocytes, schizogony, egress, and the
transitions from merozoites into gametocytes. Once in hepatic circulation, merozoites
invasion of erythrocytes is a fast and vigorous process. Although the invasion is a multi
step process, it takes under 2 minutes to complete. The first step in this process is the
pre-invasion interaction which is thought to involve the merozoites surface protein I
(MSPI) (Holder, 1994). However. studies showing that merozoites lacking MSPI can
also invade erythrocytes suggests that MSPI may have other functions that may even
tum to be more fundamental than erythrocyte invasion (Das et aI., 20 IS). The details of
pre-invasion interaction. the signalling and proteins involved are thoroughly reviewed
in Tham et al., 2012 but a synthesis of the literature portrays to fierce merozoites -
erythrocyte interaction that leads to the deformation of host cells (Gao et ai., 2013;
Singh et al.. 2010; Tham et al.. 2012; Weiss et aI., 2015). Merozoites reorient
themselves after the deformation and get ready for the active invasion phase which
involves an irreversible attachment of merozoites to erythrocytes through the formation
of tight junctions between AMA I and RON complex. After the active invasion phase,
the fusion of the posterior end of merozoites with erythrocyte membranes seals off the
parasite within parasitophorous vacuole within the now infected erythrocyte (Riglar et
al.. 20 II). After invasion, merozoites undergo schizogony leading to the egress of
between 16 - 32 merozoites after 48 hrs. Egression destroys infected erythrocytes and
the free merozoites look for another erythrocyte to repeat the process.
As schizogony continues in the erythrocytic phase of the life cycle a proportion of
merozoites make a developmental switch and become committed to gametogenesis.
33
University of Ghana http://ugspace.ug.edu.gh

The molecular events governing the switch to gametogenesis is not fully understood
but environmental stimuli such as parasitaemia and presence of anti-malarial have been
shown to increase gametocyte production. The role of epigenetic regulation of sexual
differentiation is also gradually coming to the fore and this has generated a lot of interest
in the possible role of transcription factor with AP2 domain (AP2-G) as a master
regulator of gametocytogenesis. (Kafsack et al., 2014; Waters, 2016). Although sexual
development from merozoites to gametes is crucial for the transmission of malaria from
humans to the insect vector, details of this transition for transmission are lacking.
Increased research in this area may offer clues for transmission-blocking vaccines or
therapies (Cowman et aI., 2016).
Akin to renovating a new apartment to suit your lifestyle, merozoites remodels host
cells invasions and equip it with organelles and structure that aids optimal development
and protect them from host immune responses (Boddey and Cowman, 2013). The
parasite does this by sorting exporting several proteins to specific subcellular locations
via a trafficking network characterised by Maurer's clefts, MAHRP2 (membrane
associated histidine-rich protein-2), EDV (electron dense vesicles), J-dots and other
membranous structures(Boddey and Cowman, 2013). The P. falciparum erythrocyte
membrane protein 1 (PfEMPI) family is one of the best study surface proteins on
infected erythrocytes. Although PfEMP 1 elicit strong humoral immune responses from
the host, the parasite can evade host immune responses and establish chronic infections.
This is made possible by the ability of the parasite to express antigenically distinct
isoforms of PfEMPl(Hviid and Jensen, 2015)
34
University of Ghana http://ugspace.ug.edu.gh

Parasite forms in the vector
The development of Plasmodium spp in the mosquito vector is intriguing and have
captured the attention of researchers since time immemorial. However, the push for a
better understanding of the developmental events in the mosquito has become urgent in
the era of heightened efforts towards transmission-blocking vaccine research.
Generally, development in the vector starts with the ingestion ofan infected blood meal
containing gametocytes. Gametocytes egress from blood cells and differentiate into
gametes; differentiation of the male gametocytes is most dramatic with the
transformation known as exflagellation. Microgametes searches and fertilise female
gametes giving rise to a diploid zygote that further differentiates to form a motile
ookinite. The ookinite then comes out of the blood bolus and invade the midgut
epithelium of the insect. The ookinete traverses the epithelium to the basal lamina and
differentiates into a round oocyst that grows and produce thousands of sporozoites
through sporogony. Matured oocysts release sporozoites into the haemolymph that
eventually invade the salivary gland and are deposited alongside with saliva in the
dermis of a human host on the next feeding. Reviews of the current understanding of
these processes are unequivocal in pointing out the need to go beyond schematics and
unravel the genetic and epigenetic regulation of the process (Smith et al., 2014)
2.2.2. The disease: pathogenic mechanisms and determinates of severe malaria
Malaria is a complex disease with a wide clinical spectrum. Infection with p.
/alciparum may result in asymptomatic parasitaemia or malaria with varying degrees
of severity. Clinical malaria may manifest as uncomplicated malaria or take the form
of life-threating syndromes such as severe malarial anaemia, cerebral malaria,
metabolic acidosis or even a multiorgan system failure (Wassmer et al., 2015).
35
University of Ghana http://ugspace.ug.edu.gh

Symptoms of uncomplicated malaria include fever, chills, and headaches and they
appear 10-15 days after an infective bite. Most of the symptoms in uncomplicated
malaria are due to erythrocytic schizogony and correlates with the rapture and release
factors and toxins from infected erythrocytes. If prompt treatment is delayed,
uncomplicated malaria may rapidly turn into severe malaria with dire consequences
(World Health Organisation, 2015).
Although only a minority (1 - 2 %) of P. Jalciparum infection ends up as severe
malaria(Marsh, 1992), mortality associated with severe malaria is unacceptably high
and has remained unchanged for decades (Wassmer and Grau, 2017). For instance, after
the onset of CM, mortality is between 15 - 25 % irrespective of antimalarial treatment
following standard protocols (Gupta et al., 2017; Idro et al., 2010; Wahlgren et aI.,
2017; Wassmer et aI.. 2015). Majority of stakeholders blame the current situation on
our poor understanding of the pathogenic mechanisms of CM, which severely hinders
efforts to find good prognostic markers and develop effective therapies. Considering
the imminent decline in malaria and the predicted concomitant rise in CM, scientists
are obliged to better understand the pathogenesis, find accurate prognostic markers and
developed effective therapies for CM. Adopting a conscious bias towards cerebral
malaria, this section reviews the current knowledge on malaria pathogenesis and
highlights the crucial gaps in our understanding of severe malaria.
2.2.2.1 Pathogenesis of cerebral malaria
The World Health Organization defines CM as a clinical syndrome characterized by
coma (at least 60 minutes after cessation of seizure and correction of hypoglycaemia),
presence of asexual P. Jaiciparum parasites in peripheral blood, and no other possible
36
University of Ghana http://ugspace.ug.edu.gh

cause coma (WHO. 2000). This definition is non-specific and can be confusing in
malaria-endemic regions where virial- and bacterial-encephalitis are common (Idro et
al .• 20 I 0). It might also lead to misdiagnosis in situations where malaria is incidental to
previously undiagnosed neurological conditions. This is epitomized in the landmark
Malawian post-mortem study where over 24% of children who were thought to have
died from CM. actually died of courses other than CM (Taylor et al., 2004). The
prognosis ofCM is often poor, especially when coma is accompanied by brain swelling,
intracranial hypertension. retinal changes, and other complications such as metabolic
acidosis. hypoglycaemia and shock (Idro et al., 20 10; Newton et al., 2000). Researchers
do not yet fully understand the pathogenic mechanism of CM but two hypotheses
(vascular occlusion and inflammatory), which might not be mutually exclusive, have
been proposed. Both hypotheses invoke endothelial activation in their respective
mechanisms. the contention, however, bothers on whether the endothelial activation is
a result of systemic inflammation akin to sepsis or endothelial sequestration of infected
erythrocytes (IE) in cerebral microvasculature (Storm and Craig, 2014).
Proponents of the vascular occlusion hypothesis posits that IE sequestration in the
microvasculature via PI EMP I binding to endothelial surface proteins (ICAM-I,
VCAM-l. CD36 or EPCR) leads to obstruction in microvascular blood flow. ischemia,
tissue hypoxia and the coma seen in CM (Chen et aI., 2000; Dunst et aI., 2017; Rowe
et aI., 2009). Rosettes formation and clumps may further worsen microvascular
obstruction and heighten cerebral pathology (Dunst et al., 2017; Rowe et aI., 2009). On
the other hand. proponents of the inflammatory posit that imbalance in pro- and anti
inflammatory immune responses trigger immune-induced pathology that might lead to
CM (Clark et aI., 2006; Clark. and Alleva, 2009). Although both pathogenic
37
University of Ghana http://ugspace.ug.edu.gh

mechanisms haves some loose ends, each is backed by evidence from several good
quality studies and reviews.
The most convincing evidence in support for IE sequestration (and for that matter
vascular occlusion) as the pathogenic mechanism for eM has come autopsy studies.
Despite the inherent limitation in autopsy studies, histological analysis of eM autopsy
samples has provided invaluable insights into the pathogenesis of eM. First, they
clearly demonstrated that sequestration of IE is a hallmark of eM (Dorovini-Zis et aI.,
2011; MacPherson et al.. 1985; Maneerat et aI., 1999; Taylor et aI., 2004). Secondly,
they provided evidence that IE sequestration in brain microvasculature causes
microvascular obstruction, which in turn leads to microvascular pathology, coma and
ultimately death (Dorovini-Zis eta1., 2011; Idro et aI., 2005; Postels and Birbeck, 2013).
Finally. they give a strong indication of the possible role of monocytes and fibrin
platelet thrombi in IE sequestration and microvascular congestion (Ponsford et aI.,
2012). Perhaps. the most important study in this arena is one that compared the
histopathology of Malawian children dying with clinically defined eM to the
histopathology in malaria-infected children with non-eM causes of death. Apart from
finding that approximately 25% of children clinically diagnosed as eM had died from
non-eM causes. the study more profoundly found true eM (i.e. CM patients with
evidence of IE sequestration) to have two distinct pathological patterns: CMI and CM2.
Whereas a one-third of the true eM patients had histologic evidence of IE sequestration
only (CM I), the remaining two-thirds had intra- and perivasCUlar pathology in addition
to IE sequestration (fig) (Taylor et aI., 2004).
38
University of Ghana http://ugspace.ug.edu.gh

,; ... ,
eM?
"~~.-• -J A ., .:--
~--'~-. "'-,-. -,;
Sourr. Tiylorft aI~lOO4
Figure 2. 9: Pathological differences in CM 1 and CM2
e
~ l.-
I
39
SchematIc lepresentahon of thp pathologIcal dIfferences between cerebral malaria CM 1 and CM2 Autopsy studies In chIldren have dIvided CM cases Into two groups based on histologIcal features. eM 1 cases have Infected erythrocyte sequestratIon In the cerebral mIcrovasculature and no associated vascular pathology CM2 cases are defined by cerebral sequestration plus Intra- and penvascular pathology. Including nng hemorrhages flbrtn-platelet thrombi. and Intravascular monocytes In tlH' CM2 group Infected erythrocyte (IE') sequestratIon IS frequently assocIated wIth f,brtn-platelet thrombi in both capIllaries and postcapillary venules Insets provIde examples of pathologIcal features observed In CM2. (A) shows a small branching capIllary In whIch the upstream region IS filled with sequestered IEs and one of the branches i8 occluded by a thrombus This fOvent IS associated with a rtng henlorrhage In which the mlcrove:,sL"1 I~ partially denuded of endothelial cC'lIs and is surrounded by a zone of neClO:'15 and a ring of unmtected red blood cells In the white matter, Inset (B) shows a small vessel packed with sequestered IEs and surrounded by extravasated fltxinogen Indicating i"creased permeability of the blood-brain barrier Inset (C) shOWS a micovessel filled wIth monocytes containing phagocytosed hemozoln piglTlent Intravascular pigmented monocytes are found adherent to the microvessel wall. but do not transverse across the blood-bulln barrier.
University of Ghana http://ugspace.ug.edu.gh

Thus. although it appears the body of evidence supports a CM pathogenic mechanism
that involves IE sequestration leading microvascular obstruction, data from Taylor et
at caution that this pathogenic mechanism is not a histologically uniform syndrome
(Taylor et al., 2004).
The data discussed so far. particularly fmdings of Taylor et ai., stokes the age-old
debate of whether sequestration, and therefore microvascular congestion, is the main
cause of coma in CM (Berendt et al., 1994; Cunnington et aI., 2013; Hanson et al.,
2013; Ponsford et al., 2012; White et al.. 2013). The arguments of the proponents of
sequestration are well known: (a) correlation of sequestration and disease severity, (b)
correlations between obstruction induced lactate production and severity of disease, (c)
association of abnormal distribution or decreased expression of PfEMP 1 with disease
severity and (d) poor performance of adjunctive therapies based the alternative CM
pathogenic mechanisms (Storm and Craig, 2014). On the other side of the aisle,
proponents of the inflammatory (cytokine) hypothesis argue on the following points:
(a) activation of endothelium in the absence microvascular sequestration (Manning et
aI., 2012; Yeo et a1., 2010), (b) the reality of apparently healthy individuals with high
parasitaemia (Clark and Alleva, 2009) and (c) association of pro-inflammatory
cytokines with CM.
Whereas the preceding paragraphs give an audience to the sequestration side of the
debate, very little has been adduced in favour of an inflammatory hypothesis. Results
of studies on the role of cytokine in CM pathogenesis has been difficult to intemperate
for a number of reasons. Firstly, the lack of consistency in the choice of control groups
poses huge interpretability challenges. Control groups for these studies could be drawn
40
University of Ghana http://ugspace.ug.edu.gh

from patients categorised as non-CM, severe malarial anaemia (SMA), uncomplicated
malaria (UM), encephalitis, febrile illnesses or even healthy individuals (Storm and
Craig, 2014). Secondly, the prevalence of co-infections means that whatever cytokine
profile described among CM is probably not due to malarial alone. These limitations
notwithstanding, the role of cytokines in the pathogenesis is patent and extensively
reviewed by Dunst and colleagues (Dunst et aI., 2017). The dominant idea emerging
from a synthesis of the role of cytokines in CM pathogenesis suggests that
disequilibrium in pro- and anti-inflammatory immune responses initiate immune
induced pathology that might play a leading role in CM pathogenesis (Clark and
Rockett, 1994; Dunst et al., 2017). Early studies in this arena focused on the roles of
individual cytokines but it quickly became clear that a complex interplay of several
cytokines is more conceivable. Thus, the focus now is to determine common
cytokine/chemokine signatures for CM and although unambiguous cytokine signature
is yet to be identified, the body of evidence suggests roles for cytokines in endothelial
activation, blood-brain barrier integrity and neuroinflammation (Bakmiwewa et al.,
2016; Combes et aI., 2010; Dunst et aI., 2017; Obermeier et aI., 2013; Renia et aI.,
2012).
Regardless of the equivocations in the literature on a pathogenic mechanism for CM,
certain cardinal features of the syndrome are undisputed: (a) sequestration of IE in the
microvasculature, (b) endothelial activation, (c) a pro-inflammatory immune response
and (d) disruption of BBB. The dispute appears to be on the lines of causality is
endothelial activation caused by IE sequestration or pro-inflammatory cytokines; do
pro-inflammatory cytokines aid IE sequestration; can cytokines on their own induce
coma without IE sequestration; etc. Some of these questions will be difficult to answer
41
University of Ghana http://ugspace.ug.edu.gh

within the limitations of autopsy studies. Although some imaging technologies are on
the horizon, the application of these technologies in CM pathogenesis studies is in its
infancy (Dunst et al., 2017; Wassmer and Grau, 2017). The potential of robust
biomarkers for endothelial activation becomes apparent in the backdrop of the
limitation of autopsy study studies.
2.2.2.2 New paradigms in the pathogenesis of cerebral malaria
Despite the methodological challenges in studying the pathogenesis of CM, this
research arena is active and bourgeoning with new knowledge. Recent studies shed
light on several aspects of CM pathogenesis including parasite-brain microvascular
specificity, new causes and repercussions of endothelial dysfunction, and the
consequences of rosette formation and clumping (Riggle et al., 2017; Wassmer et al.,
2015). Other areas where further insights have emerged include BBB integrity, novel
parasites pathogenic factors, new methodological approaches and adjunct therapies
(Wassmer and Grall, 2017). But perhaps, the most interesting twist is this bourgeoning
research arena is the novel pathogenic mechanism emerging from the work of Gyan
and colleagues which propose a pathophysiological model that does not only look at
microvascular damage but considers hosts capacity to repair damaged endothelium
(Gyan et ai., 2009). This pathophysiological model is intriguing because it balances the
details of malaria-induced endothelial damage with host-mediated mechanisms for
repairing damage microvasculature (Desruisseaux et al., 2010; Dickinson-Copeland et
al., 2015; Gyan et aI., 2009; Tetteh, 2014). Thus, unlike the two main
pathophysiological models for eM that only focuses on IE sequestration in endothelial
microvasculature, endothelial activation, inflammation and microvascular pathology,
42
University of Ghana http://ugspace.ug.edu.gh

this model additionally focuses on factors that influence repairs such as post-natal
angiogenesis and its regulation during malaria infections.
In this pathogenic model, the prognosis of eM is hypothesized to partly hinge on a
child's ability to promptly repair damaged microvasculature and restore equilibrium.
Although this appears to be an elegant hypothesis, evidence in support of this
pathophysiological model is scanty. Repair of damaged endothelium was thought to
occur through replication of cells adjacent to the site of injury but a seminal work
highlighted the role of other factors such as circulatory endothelial progenitor cells
(cEPC) in the repair of damaged microvasculature almost a decade ago (Asahara et aI.,
1999). The role of cEPes in repairing endothelial damage has since become clearer and
scientists agree that the incorporation of cEPes into sites of microvasculature damage
with the help of stromal cell-derived growth factor 1 (SDF-l) and the matrix
metalloproteinase-9 (MMP-9) is crucial (Asahara et aI., 1997; Hristov et aI., 2003;
Rafii, 2000; Urbich and Dimmeler, 2004). Interesting, a study in Ghana found
significantly lower cEPe levels and higher SDF-l in eM patient relative to healthy
controls and non-eM malaria phenotypes (Gyan et aI., 2009). These findings suggest
that the repair of endothelial damage could play significant roles in eM pathogenesis.
These finding notwithstanding, this eM pathophysiologic model will require further
research.
2.3. Markers of endothelial damage and repair
There are structural and functional differences in the endothelia of the vascular tree and
the endothelium lining the microvasculature is distinguishable by the absence of
fenestrations, extensive tight junctions, and sparse pinocytotic vesicular transport
43
University of Ghana http://ugspace.ug.edu.gh

(Poggesi et aI., 2016a). When its integrity is intact, the cerebral endothelium serves as
a barrier that strictly controls the exchange of solutes and circulating cells between the
plasma and the interstitial space (poggesi et al., 2016b). However, the integrity of this
barrier is breached in certain pathological situations leading to a cascade in
microvascular pathology. In P. falciparum infection, this breach may be due to the
sequestration of IE and/or inflammation - be it localised or systemic. An accurate
assessment of cerebral endothelial damage or dysfunction is not always possible during
malaria infection and thus, the availability of reliable biomarkers of microvascular
endothelial health would be invaluable. This section reviews the biomarkers of
endothelial damage and repair with the aim of identifying those with the greatest
potential of becoming proxy markers. Although subtle differences discriminate
between tenns endothelial damage, endothelial activation and endothelial dysfunction,
they are used interchangeably in this thesis.
2.3.1 Marker of endothelial dysfunction
Generally, markers that consistently characterise endothelial dysfunction include von
WilIebrand factor (Hollestelle et aI., 2006), angiopoietin-l and -2 (Lovegrove et aI.,
2009). and soluble endothelial receptors (Turner et aI., 1998). Physiologically.
angiopoietin-I functions to stabilise the endothelium whiles angiopoietin-2, its
antagonist, is released in response to inflammatory stimuli to regulate vascular
penneability and the expression of endothelial receptors (Stonn and Craig, 2014). In
the context ofCM pathogenesis, studies found increasing Ang-2/Ang-l ratio, which in
reality, represents either a decrease in Ang-l or an increased in Ang-2, to be associated
CM (Conroy et aI., 2010, 2009; Jain et aI., 2011; Lovegrove et aI., 2009; Prapansilp et
al., 2013). Other studies have found low bio-availability of nitric oxide to be associated
44
University of Ghana http://ugspace.ug.edu.gh

with malaria severity; but this is unsurprising considering the fact that nitric oxide
regulates the actions of Ang-lIAng-2, causes vasorelaxation and downregulates the
expression of endothelial adhesive molecules (Anstey et aI., 1996; Lopansri et al., 2003;
Yeo et al., 2008, 2007). In addition to the angiopoietins, von Willebrand factor, and
nitric oxide, studies have implicated other related molecules as vasoactive substances
but their role in malaria appears insignificant relative to Ang-l and Ang-2 (Bergmark
et aI., 2012; Canavese and Spaccapelo, 2014; Kim et aI., 2013; Starke et al., 2011).
Soluble endothelial receptors such as ICAM-l, V-CAM, E-selectin and EPCR play
vital roles in the characterisation of endothelial dysfunction, however, heterogeneous
study designs tend to obscure their impact in the context of malaria. For instance,
soluble ICAM-l which did not appear to correlate with CM in Ghanaian children was
associated with SM when compared among UM and other SM cases (Adukpo et aI.,
2013a; Cserti-Gazdewich et aI., 2010; Erdman et aI., 2011). This situation is further
complicated by discordant findings in children and adults (Combes et aI., 2010; Sahu
et aI., 2013). Apart from ICAM-I, EPCR, another receptor for PtEMPI has been
associated with 8M including CM. Interestingly, seven newly described receptors for
PfEMP I expressed in the brain failed to show any association with malaria severity
(Esser et al., 2014; Turner et aI., 2013). Taken together, angiopoietin-l, angiopoietin-2
and EPCR appear to be a promising candidate as biomarkers for endothelial
dysfunction.
2.3.2 Markers of endothelial repair
Sequestration of IE in cerebral microvasculature and/or inflammation, be it localised in
the brain or systemic, can lead to endothelial activation and damage. The extent of the
45
University of Ghana http://ugspace.ug.edu.gh

injury is detemrined by the degree of IE sequestration and inflammation, however, the
role of biologic capacity to repair damage endothelium cannot be discounted (Idro et
ai., 2010; Turner et ai., 1998). Thus, a review of the repair mechanisms of endothelial
offers great insights into potential biomarkers of endothelial repair.
Two main repair mechanisms have been described. The first mechanism assumes an
endothelium with terminally differentiated cells, low proliferative turnover and an
inherently weak self-repair capacity. In this scenario, the repair of damaged
endothelium is through the replication of mature endothelial cells surrounding the
injured site (Avogaro et ai., 2011; Dzau et aI., 2005). However, it is now known that
circulating EPCs recruited from the bone marrow and other tissues can embed
themselves at the injured site and differentiate into mature endothelial cells (Asahara et
aI., 1999; Ribatti, 2007). This establishes the importance of cEPCs for the maintenance
and repair of endothelial cells.
The precise characterisation of EPCs is complicated by the heterogeneous nature and
shared surface markers among that group of cells (Asahara et aI., 20 11; Resch et aI.,
2012; Ribatti. 2007). Currently, cells positive for both a hematopoietic stem cell marker
such as CD34 and an endothelial marker protein such as VEGFR2 are considered as
EPCs (Resch et aI., 2012). Several factors including nitric oxide, VEGF, SDF-I,
erythropoietin, and oestrogens are crucial for the mobilisation, migration and homing
of EPCs into sites of endothelial injury. Thus, quantitative or qualitative deficits in any
of these factors may influence the repair of damaged endothelium. Accordingly, recent
evidence shows overexpression ofVEGF in brain tissues ofCM patients (Canavese and
Spaccapelo, 2014). Similarly, SDF-l, nitric oxide and EPCs have been implicated in
46
University of Ghana http://ugspace.ug.edu.gh

the pathogenesis CM (Anstey et al., 1996; Gyan et aI., 2009). Taken together, VEGF,
SDF-l and EPCs are "front-liners" in the search of biomarkers for endothelial repair.
The role of EPCs as a biomarker of endothelial repair is further bolstered by studies
that show higher EPCs levels to be associated with a better endothelial function
(assessed by brachial artery flow-mediated dilatation (FMO) (Verma et al., 2017).
2.4 Malaria immunology
2.4.1 Immune responses during pre-erythrocytic stages of Plasmodium life cycle
Pre-erythrocytic stage infections with P. jalciparum are clinically silent and thus, host
immune response against parasite forms at this stage remains largely unexplored.
However, it is has been suggested that interaction of the immune system with pre-
erythrocytic phase parasites primes the innate immune system (Offeddu et aI., 2012).
Most of our understanding of the immune responses at this stage come from murine
studies and the data suggests an interaction between innate immune cells and liver
sporozoites of P. berghei (Liehl et al., 2014). These studies suggest a type I interferon
(lFN) response and the subsequent recruitment of a number of cytokines and
chemokines (Crompton et al., 2014; Liehl et aI., 2014; Liehl and Mota, 2012; Miller et
al..2014). In human studies, however, the evidence for potent innate immune responses
in the skin and liver is lacking, and unsurprisingly, antibody, C04+ and CD8+ T cell
responses to the infecting sporozoites are also lacking (Offeddu et aI., 2012). The
clinical silence and relatively weak innate and adaptive immunity to pre-erythrocytic
stages of the Plasmodium are poorly understood. Perhaps, this reflects the low inoculum
dose of sporozoites per infective bite (10 - 100 sporozoites) andlor the parasite's
exploitation of the inherent immune-regulatory environment of the skin and liver
(Crispe, 2009; Honda et al., 2011). Thus, although parasites interact with the immune
47
University of Ghana http://ugspace.ug.edu.gh
I'ti 1-' mune responses against pre-. h erythrocytic stages of the 1 e cyc c, lffi system In t e pre-
erythrocytic parasites are not potent enough to limit the further development of the
parasite and the transition to the erythrocytic stage (Kordes et aL 2011).
I(::zo-._J' • • e. 'f .~. _.
Adaptive Immune responses to preerythrocytic antigens.
-. -. -- ---e---'l~ ~~~j ~.'~~I •
B
A.;.., to.
Figure 2. 10: AdaptiH immunit~ at the pre-erythrocytic -;tage
2.4.2 Immune responses during erythrocytic stages of Plasmodium lifecycle
Almost all the clinical symptoms associated with malaria can be attributed to the
erythrocytic phase of Plasmodium's Iifecyclc which stans with the release of
merozoites into the bloodstream. Merozoites releast: inll \ the bloodstream are only
brietly (- 60 S) exposed to the immune system before they infect erythrocytes. The
characteristic recurrent fever seen in malaria coincide~ with synchronized rupture of IE
which introduces parasite- and host-deri\ ed molecules (PAMP and DAMPs) capable
of inducing T"F (Evans et aI., 2015: Kv.iatkowski ct al.. 1990, 1989). Studies found
levels of T!\F to coincide with fever suggesting that fever is induced by the repeated
exposure to PAMPs and DAMPs (Kwiatk'l\\ ski et al .. 1990. 1989; Oakley et aI., 20] I).
48
University of Ghana http://ugspace.ug.edu.gh

Although generally associated with the resolution of infection, uncontrolled fever can
lead to a dysregulation of the immune system and ultimately severe malaria (Oakley et
aI., 2011). These findings are consistent with the textbook understanding offever as a
physiologic innate response that precedes the involvement cells of the innate immune
system.
2.4.2.1 Innate immune responses
Cells of the innate immune system are thought to playa crucial role in providing the
first line of defence against malaria. Thus, macrophages, dendritic cells (DC) and other
non-professional immune cells interact with merozoites and mount strong pro
inflammatory responses during the erythrocytic phase. For instance, toll-like receptor
2 (TLR2) interacts with PAMPs such as GPI to produce a strong TNF response from
murine macrophages (Krishnegowda et al., 2005; Schofield and Hackett, 1993;
Tachado et aI., 1996; Zhu et aI., 2011, 2009). The importance of GPI was however
questioned when studies failed to show that TLR deficiency impaired immune
responses in mice (Lepenies et aI., 2008; Togbe et al., 2007). Hemozoin, which is
released during erythrocyte rapture is another potential malaria PAMP that also induce
the expression of TNF and IL-I ~ from monocytes, human monocyte-derived DCs and
murine macrophages, probably, via binding to TLR9 (Bujila et aI., 2016; Coban et aI.,
2005; Olivier et al., 2014). Interestingly, the responsiveness of macro phages and DCs
to TLR-mediated signalling has been shown to be impaired by phagosomal
acidification upon phagocytosis (Wu et aI., 2015). Thus, regardless of phagocytosis
being an important feature in innate immune response that helps in the uptake ofIE and
increases serum levels of pro-inflammatory cytokine, it counterbalances PRR-mediated
signalling. Another contributory factor to the pro-inflammatory environment created
49
University of Ghana http://ugspace.ug.edu.gh

during malaria infection is host-derived DAMPs such as urate crystals, heme, and
macro vesicles release upon rapturing of host cells. In line with this thought, studies
have found higher levels plasma microparticles in severe malaria cases relative to
uncomplicated malaria (Campos et aI., 2010; Nantakomol et aI., 2011; Sahu et aI.,
2013). It is important to reiterate here again that the inflammatory environment created
during infection also influences the severity of malaria and several cytokines and
chemokines, including CXCLlO and lNF have been associated with CM (Kwiatkowski
et aI., 1990; Wilson et aI., 20 II). Levels oflNF are thought to be particularly important
in influencing malaria severity and even though more recent studies failed to find an
association between cerebrospinal fluid (CSF) lNF levels and CM, among CM cases,
it was associated with longer-term neurologic deficits (Shabani et aI., 2017).
2.4.2.2 Adaptive immune responses: humoral immunity
The role B cells and humoral immunity is most pronounced during the erythrocytic
phase but almost six decades after showing the central role of antibodies in immunity
against blood-stage parasites, we still do not fully understand the parasite proteins
targeted, the actual mechanisms of protection, and acquired immunity albeit nonsterile
(Cohen et aI., 1961; Gardner et aI., 2002a). By showing that IgG from immune adults
could reduce parasitaemia in subjects with clinical malaria, the passive transfer studies
fundamentally demonstrated the importance of antibodies in malaria immunity.
Additionally, these studies demonstrated antibody cross-reactivity, the role of B cells
in immune protections against malaria and the induction of memory in B cells
(Hirunpetcharat et aI., 1999; Meding and Langhorne, 1991; Ndungu et aI., 2012, 2009;
Sabchareon et aI., 1991; Wykes et aI., 2005). These studies and the many that followed
have however failed to identify the targets of protective immunity using the immune
50
University of Ghana http://ugspace.ug.edu.gh

correlates of protection approach and/or growth inhibition assays. Although yet to be
fully clarified, variant surface antigens expressed on IE and merozoites surface antigens
used for invasion have emerged as the most likely target of protective immunity (Wykes
et al .• 2017). Four classes ofVSA have been identified in P.falciparum malaria namely:
P. jalciparum erythrocyte membrane protein 1 (PfEMP1), repetitive interspersed
family proteins (RIFIN), subtelomeric variable open reading frame proteins
(STEVOR), and surface-associated interspersed gene family proteins (SURFIN). Of
these 4 groups, studies have found that antibodies targeting PfEMP 1 and RIFIN show
the strongest association with protection (Abdel-Latif et aI., 2003; Gardner et al.,
2002b; Kraemer and Smith, 2003; Tan et al., 2016). Antibodies targeting some
merozoites surface proteins involved in erythrocyte invasion have also been shown to
be associated with protection. The best-studied are the apical membrane antigen 1
(AMA1), glutamine-rich protein (GLURP), merozoites surface protein (MSP)-3, and
MSP-119 (Fowkes et aI., 2010). Selection of antigens for candidate vaccine studies
have been based on these studies but findings of a recent study in Mali that found 49
potential vaccine candidates not been considered as possible vaccine antigens in
ongoing vaccine candidate studies points to gapping gaps in knowledge or lack of
methodological rigour (Davies et aI., 2015). For instance, most GIA studies using heat
inactivated purified sera would have inadvertently excluded the effects of compliment,
lymphocytes and monocytes all of which work in concert in natural situations
(Bouharoun-Tayoun et aI., 1990; Boyle et aI., 2015b; Kennedy et aI., 2016; Rosa et aI.,
2016).
51
University of Ghana http://ugspace.ug.edu.gh

2.4.2.3 Adaptive immune responses: cell-mediated immunity
Malaria T cell immunity is usually under-researched, nonetheless, it appears to play an
important role in malaria immunity. Most of what we know about T cell immunity
comes from murine studies and these studies confirm the crucial role of CD4+ T cells.
Besides aiding B cells and promoting phagocytosis, CD4+ T cells help control
parasitaemia through the induction oflFN-y and TNF -a both of which, contributes to
a vast and intricate network of protective immunity and immunopathology (Langhorne
et aI., 1989; Mota et al., 1998; Podoba and Stevenson, 1991). In a recent controlled
human malaria infections (CHMO study that was not confounded by the prolonged
effects of atovaquone in vivo, multiple immunization with live sporozoites under drug
cover induced sporozoites and blood-stage-specific T cells responses that were
polyfunctional (Edstein et al., 2005; Pombo et aI., 2002; Roestenberg et aI., 2009).
Subsequently. it was found that although the immunised individual was protected
against sporozoite challenge, they were no such protection when they were challenged
with blood stage parasites (Roestenberg et aI., 2009).
Early IFN-y responses to P. Jalciparum is a good correlate of better anti-parasitic
immunity but this has been difficult to measure in many studies. However, a
longitudinal study in Uganda found malaria-specific CD4+ T cells responses in almost
all children with majority producing of the CD4+ T producing IFN-y and interleukin-
10 (IL-IO) in response to IE (Jagannathan et aI., 2014). Interestingly, IFN-r responses
to MSP-I antigens were commonly associated recent exposure to malaria but offered
no protection from subsequent malaria challenge and inversely, IFN-y responses to pre
erythrocytic antigens were uncommon but protected against subsequent infections
(Boyle et aI., 2015a). Other studies have reported differences between adults and
52
University of Ghana http://ugspace.ug.edu.gh

children in high and low transmission regions (Boyle et aJ., 2015a, 2015b). The
contribution of CD8+ T cell-mediated immunity against blood-stage malaria parasites
has been difficult to decipher because research on this has been limited to their effect
on liver-stage infection, the pathogenesis of cerebral malaria, and damage to splenic
architecture (Beattie et aJ., 2006; Hafalla et aI., 2006). However, several studies point
to the importance of CD8+ T lymphocytes in the immunity against blood-stage
parasites. For instance, it has been shown in animal experimental studies that the
depletion of CD8+ T lymphocytes during blood-stage P. chabaudi infections delayed
the clearance of infection (McCall and Sauerwein, 2010; Suss et aI., 1988). In another
experimental study, naIve mice transfused with CD8+ T lymphocytes derived from
infected mice survived lethal blood-stage challenge (Imai et aI., 2010). Besides the
possibility that immature erythrocytes can activate CD8+ T lymphocytes, there is
evidence to suggest that severe malarial anaemia is mediated by CD8+ T lymphocytes
dependant parasite clearance (Imai et aI., 2013; Safeukui et aI., 2015).
2.5 Host genetics and epigenetics in malaria pathogenesis of eM
The interplay between host genetics and susceptibility to malaria has been known for
almost a century (Allison, 1954; Bryceson et al., 1976; Fleming et aI., 1979). In fact,
malaria is believed to have acted as an evolutionary force to keep otherwise deleterious
genetic mutations in the population (Allison, 1954; Hill et aI., 1997). This fact is
epitomised by the prevalence and distribution of genetic diseases like sickle cell trait,
a-thalassemia, Glucose-6-phosphate dehydrogenase (G6PD) deficiency, and Duffy
antigen receptor negativity (Hill et al., 1997). Similarly, the fact of interethnic
differences in malaria susceptibility among the Fulani and their sympatric neighbours
further confirms the importance of host genetic factors in the epidemiology of malaria
53
University of Ghana http://ugspace.ug.edu.gh

(Arama et ai., 2015; Cherif et aI., 2016; Fleming et aI., 1979). The mechanisms of
genetic susceptibility are not fully understood, however, deciphering these mechanisms
could further elucidate pathogenic mechanisms and lead to therapeutic breakthroughs
(Handel and Horuk, 2010; Kidson et aI., 1981). This section of the thesis reviews the
role of host genetics and epigenetic factors in the pathogenesis of malaria with a
conscious bias on CM.
2.5.1 Genetic disorders of erythrocytes and susceptibility to malaria
The clinical symptoms of malaria are associated with the erythrocytic stage of the life
cycle and thus, genetic disorders of erythrocytes (be it membranes-, enzymes-, or
haemoglobin-related) can potentially influence the pathogenesis of malaria. For
instance. studies have shown membrane-related genetic disorders such as
spherocytosis, ovalocytosis, elliptocytosis, pyropoikilocytosis, and acanthocytosis to be
either associated with lower parasitaemia or resistance to invasion (Hadley and Miller,
1988; Schulman et al.. 1990). The protection conferred by a particular type of Southeast
Asian ovalocytosis (SAO) is of interest because heterozygous individuals do not show
clinical symptoms or have haemolysis upon infection (Kidson et aI., 1981). Similarly,
a SNP (rs2814778) that causes a glycine to asparagine change in the Duffy
antigenlchemokine receptor (DARC) and another polymorphism (-33T>C) in the
promoter region of the same antigen are associated with a lower risk of viva x malaria
(King et aI., 2011; Maestre et aI., 2010). Polymorphisms in other erythrocyte surface
receptors use by P. !a[ciparum such as glycophorin, protein band 3 and others have
been shown confer resistance in Papua New Guinea and Brazil (Tarazona-Santos et
aI., 2011). A recent study reported a novel malaria resistance locus close to glycophorin
54
University of Ghana http://ugspace.ug.edu.gh

gene and a haplotype at this locus provided 33% protection against severe malaria
(Malaria Genomic Epidemiology Network, 2015).
One of the commonest erythrocyte genetic disorder with implication for malaria
pathogenesis affects the metabolic enzyme G6PD. The G6PD deficiency which affects
_ 400 million people in tropics and sub-tropics is thought to reduce intracellular parasite
growth, probably through enhanced phagocytosis by monocytes (Ayi et aI., 2008;
Cappadoro et aI., 1998). Erythrocyte genetic disorder may also involve haemoglobin
altemtions generally referred to as haemoglobinopathies. Haemoglobinopathies may
involve structural alterations leading to variants ofHb, such as HbS, HbC, and HbE; or
a defect in the synthesis of the globin chain in Hb (alpha- and beta-thalassemia). An
SNP (rs334) in the beta-globin gene (HBB) is responsible for the famed HbS variant
whose heterozygosity is consistently associated with protection from severe fonns of
malaria including CM (reviewed in (Mendon~a et aI., 2012). The other Hb variants
(HbC and HbE) have been shown to protect against severe malaria phenotypes
(Agarwal et aI., 2000; Mendon~a et aI., 2012; Nagel et ai., 1981). Although variants of
thalassaemia and HHB have been associated with malaria severity in different parts of
the world, the patterns of resistance are mosaic in nature and difficult to generalise
(Arese et aI., 2015; Daou et aI., 2015; Mendon~a et ai., 2012; Para et al., 2018).
2.5.2 Malaria immunogenetics
Surviving a P. /alciparum infection requires a well-coordinated and finely tuned
immune responses involving several immune cells, antibodies, cytokines, chemokines,
receptors, transcription factors etc. Genetic and/or epigenetic modifications of any of
these factors can derail the response and lead to an inappropriate response. Studies over
55
University of Ghana http://ugspace.ug.edu.gh

the years have identified several polymorphisms in these factors that can potentially
influence the immune response and affect the pathogenesis of malaria. Toll-like
receptors play important roles in innate immune response and TLR2, TLR4 and TLR9
have been found to particularly important in malaria (Coban et al., 2005; Krishnegowda
et al.. 2005; Parroche et aI., 2007). Majority of functional SNPs describe in TLRs affects
ligand recognition and intracellular signalling and thus, it is unsurprising that these TLR
SNPs are implicated in parasitic disease (SchrOder and Schumann, 2005). Similarly,
several polymorphisms in TLR2, TLR4 and TLR9 have been associated with malaria
but the most interesting is a 22-base pair deletion in the UTR in TLR2 which is
associated with protection from CM (Greene et al., 2012). Polymorphisms in other
TLRs are inconsistently associated with malaria in Ghana, Brazil and Malawi (Omar et
aI .• 2012; Zakeri et aI., 2011). However, a meta-analysis of all TLR SNPs found the
association of TLR9T -123 7C with severe malaria to be the most robust (Dhangadamajhi
et al.. 2017).
Cytokines play an important role in the pathogenesis of malaria and an imbalance in
pro-inflammatory. anti-inflammatory and regulatory cytokines is believed to contribute
to the development of severe forms of malaria including CM. Tumour Necrosis Factor
Q is a key cytokine in malaria immunity and immunopathology and several
polymorphisms have been described in the TNF -Q gene that can influence the
pathogenesis of malaria. A study in Gambian children associated rs 1800628 and
rs361525, two of the best known TNF-Q SNPs, with CM and SMA respectively (Clark
et aI., 2009; McGuire et aI .• 1994). These same SNPs were associated with protection
from mild (rs361525) malaria and the frequency of malaria infection (rs1800629)
(Clark et aI., 2009; Meyer and Astor, 2002). Other studies on these SNPs and others
56
University of Ghana http://ugspace.ug.edu.gh

(rs 1799724 and rs 1799964) have been associated with severe malaria phenotypes in
different malaria-endemic regions (Wattavidanage et al., 2001). Several other studies
have finds SNPs in TNF-a to be associated with eM but these studies are froth with
heterogeneous study designs and mixed results (Mendon~a et al., 2012). Other
cytokines and transcription factor gene polymorphisms, as well as some chromosomal
regions (5q31-33), have been shown to influence malaria in different settings (Furini
et aI., 2016; Naka et aI., 2009). Of note is the role of an intronic SNP and the STAT6
gene that was shown to protect against CM in Ghanaian children (Amoako-Sakyi et aI.,
2016).
Polymorphisms in receptors for the Fc fragment of IgG (FcyRs) which serve as a link
between humoral and cellular immune responses have also been shown to influence
malaria pathogenesis. Besides the famed FcyRIIA H 131 R polymorphism, several
studies have reported on SNPs that could influence malaria pathogenesis (Adu et al.,
2012; Braga et al" 2005; Mendon~a et aI., 2012; Munde et aI., 2017, 2012). A recent
study among the Fulani who are naturally resistant from severe fOnDS malaria suggests
that the mechanism of protection may be FcyRs-mediated (Cherifet aI., 2016)
Polymorphism in cells, molecules and receptor that are not exactly immune-related but
involved in the pathogenic mechanisms can also influence the severity of malaria. Here
again, polymorphisms in ICAM-l (rs5491, rs5498), C036 (rs321 1938 and GI439C),
PECAM-I (rs668, rsl2953 and rsl131012) and CRI (rs9429942) have been associated
with malaria severity (Adukpo et aI., 2013b; Kikuchi et aI., 2001). Taken together, the
role of host genetics on malaria pathogenesis cannot be discounted, however,
57
University of Ghana http://ugspace.ug.edu.gh

methodological heterogeneity, regional differences in malaria transmission. variance in
parasite virulence and other factor has led to a mosaic result that is difficult to interpret.
2.5.3 Malaria bost epigenetics
Apart from heterogeneity in study designs, regional differences in malaria transmission
and variable parasite virulence, host epigenetics can also cofound results of genetic
susceptibility studies. The regulation of gene expression is not entirely genetic and
epigenetic modifications such as DNA methylation, histone modifications and RNA
base modification can influence the pathogenesis of several diseases including malaria
(Wagner et aI., 2014; Zeng et aI., 2014). Interestingly, there is evidence to suggest that
epigenetic mechanisms collaborate with genetic mechanisms to co-regulate gene
function in several disease models (Berger et aI., 2009). This possibility is exemplified
in the occurrence of SNPs within epigenetic marks. Such occurrences affect chromatin
structure at specific genomic locations by modifying methylation patterns or histone
type recruitment (Dayeh et aI., 20 13b). Some researchers posit that interactions between
genetics and epigenetics may offer better explanations for scenarios where disease
associated genetic variants lie outside promoters or coding regions (Zaina, Perez
Luque, and Lund 2010). The literature on the effects of epigenetic mechanisms on the
pathophysiology of human disease are growing but few have focused on malaria or
looked at the interaction ofSNPs and epigenetic marks in malaria pathogenesis (Bell et
aI., 2011; Dayeh et aI., 2013a; Gupta et aI., 2017). However, the advancements in
bioinformatics and the availability of new tools have opened the prospects of analysing
such interactions in silico.
S8
University of Ghana http://ugspace.ug.edu.gh
CHAPTER THREE
3.0 METHODS
3.1 Study Site
This study recruited volunteers from five partner health facilities (Figure 3.1) within
Accra and It:ma metropolitan areas in Ghana. The partner hospitals were referral
facilities. and thus. they served the communities in which they are situated as well as
attend to cases referred from smaller satellite facilities such as Health Centres and
CHiPS zones. Thc~c facilities were selected based on their sentinel nature and
proximit~ to Noguchi Memorial Institute for Medical Research where laboratory
analysis was conducted.
a .\ .... 1 •. ••
Locations of 5 stUdy hOSPI;~~ In relation to NMIMA
Figure 3. 1: Study site
University of Ghana http://ugspace.ug.edu.gh

3.2. Study design and sample size estimations
Participants for this cross-sectional study comprised three different malaria phenotypes
(CM. UM and SMA) collectively referred to as cases and a control group are referred
to as healthy controls (HC). The cases were recruited from partner hospitals whilst
healthy controls were recruited from basic school within the vicinity of the partner
hospitals. All partner hospitals were within a reasonable distance from NMIMR to
ensure that samples got to the laboratory on time for analysis. Recruitment of study
participants was done between 2013 to 2016.
This study used G*Power 3.9.1.2 for all sample size estimations. This study had
multiple objectives with each requiring different sample size considerations. For
instance. in comparing malaria phenotypes and controls (i.e. 4 groups), G*Power
3.9.1.2 estimated that a total sample size of 280 will be enough to detect a moderate
effect size 0.25, at a power of 0.95 and a - level of 0.05. However, in comparisons
among malaria phenotypes only (3 groups), a total sample size of 240 was thOUght to
be enough to give the same effect size and power at an a-level of 0.05. In comparing 2
groups under the same conditions, a total of 21 0 was deemed to be enough. This study
ended up using a total of 221 study participants, thus, whilst the study was slightly
underpowered (0.85) in four-group comparisons, it was adequately powered and
slightly overpowered in detecting differences in three-group and two-group
comparisons respectively. This study prioritised detecting the difference between CM
and other non-eM groups and in all these two-group comparisons. the study was
adequately powered (> 0.95) in detecting an effect size 0.1.
60
University of Ghana http://ugspace.ug.edu.gh

Hong & Park, 2012 have shown that sample size for candidate gene studies is dependent
on the inheritance model assumed (Hong and Park, 2012). This study's sample size of
221 is deemed adequate for all inheritance models except for the recessive inheritance
model. Correcting for multiple testing for the 27 SNPs. a Bonferroni approach
suggested a conservative significance threshold for this study (p < 0.002). Thus, genetic
testing was deemed significant p < 0.002.
3.3. Ethical Considerations
This study secured ethical clearance from the Institutional Review Board (lRB) of the
Noguchi Memorial Institute for Medical Research (NMIMR). Both the Ghana Health
Service (GHS) and the Ghana Education Service (GES) gave approval for study
participants to be recruited from their respective facilities. Parents and legal guardians
consented to the study before their wards were enrolled in the study. Parents had the
right to discontinue the study anytime during the study.
3.4 Inclusion criteria
Children reporting at the GPO or emergency departments of partner hospitals with
suspected cases of malaria. Parental consent and a child's accent were considered as
an inclusion criterion.
3.4.1 Specific inclusion Criteria
Study participants were categorized as either cerebral malaria (CM), uncomplicated
malaria (UM), severe malarial anaemia (SMA) or healthy controls (HC) based on
specific and stringent clinical case definitions:
61
University of Ghana http://ugspace.ug.edu.gh

Uncomplicated malaria (UM):
.:. History of fever within the last 48 hours of fever (axillary temperature ~
37.5°C) .
• :. Five or more parasite per HPF (approx. 2S00/~I)
.:. No other obvious cause for fever.
Cerebral malaria (CM):
.:. Inclusion criteria for UM .
• :. Unconsciousness with coma score of < 3 for the duration of> 60 minutes on
the Blantyre coma scale .
• :. No record of recent severe head trauma and other cause of coma or neurological
diseases .
• :. Haemoglobin of> SgldL
Severe malaria anaemia (SMA):
.:. Inclusion criteria for UM .
• :. Haemoglobin of < SgldL,
.:. Fully conscious,
.:. No cases of severe bleeding reported or observed
.:. No convulsion.
Healthy controls:
.:. No fever
.:. No malaria parasitaemia.
62
University of Ghana http://ugspace.ug.edu.gh

3.4.2 Exclusion criteria
Potential study participants with concomitant infections such as bacteraemia and
meningitis/encephalitis (ascertained from CSF testing conducted by a trained
technician) at the time of recruitment were excluded from the study. Children with HIV
infections and other medical conditions that can potentially influence EPC levels or
microvascular damage were also excluded from the study. Other conditions and
medical procedures that excluded potential study participants from the study included
cardiovascular diseases, diabetes mellitus, hypercholesterolemia, surgery in the last one
month, bone fracture in the last three months, major trauma in the last three month (e.g.,
road traffic accident (RT A» and blood transfusion in the last three months. Study
clinicians in charge of recruitment used information in the folder of study participants
and clinical judgement to exclude a participant from the study.
3.5 Blood sample collection
Each study participant provided 2ml of venous blood in EDT A tubes for a complete
blood count (CBC), blood culture, sickling test, and blood smear for malaria parasite
estimation. Additional blood samples were collected into heparin tubes (lml), EDTA
tubes (lml) and blood culture bootless (1 ml) for downstream laboratory analysis,
which included immunoassays, flow cytometry, and genetic analysis. Healthy controls
provided venous blood in heparin (lml) and EDT A (2ml) tubes. All blood samples were
collected by a trained phlebotomist.
63
University of Ghana http://ugspace.ug.edu.gh

3.6. Sample processing and downstream analysis
Blood samples for routine clinical procedures were taken immediately to the hospital
laboratories for evaluation, whiles those for the research study were transported in cold
ice chests to the laboratories of the Immunology Department, NMIMR. Cerebrospinal
fluid samples were transported to either the Korle Bu Teaching Hospital or the Lancet
laboratory for analysis. At NMIMR, a 400ul aliquot of EDT A treated blood were used
for flow cytometry and the rest were separated into RBCs and plasma for storage at -
30°C. Heparinized blood was also processed by centrifugation and separated into RBCs
and platelet-free plasma This was done by initial centrifugation at lOOO x g for 15
minutes and separated plasma at 10000 x g for 10 minutes. Both RBCs and platelet free
plasma were stored at -30oe for further analysis.
64
University of Ghana http://ugspace.ug.edu.gh
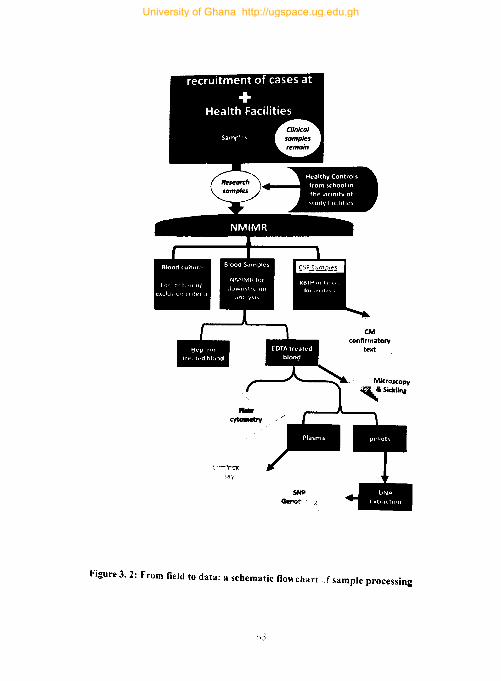
Figure 3.2: From field to data: a schematic flowchart "f sample processing
University of Ghana http://ugspace.ug.edu.gh

3.7 Measurement of angiogenic factors
Serum levels ofSDF.I, NGRI, TREMl, ANGI, ANG2, CAl, TEK. SIB and MMp·9
were multiplexed and measured with a Human Magnetic Luminex Assay (Luminex
Cooperation, Texas) according to manufacturer's protocol. Briefly, calibrator diluent
RD6.52 (provided by the manufacturer) was used to dilute plasma samples and standard
cocktail 10·fold and 3-fold respectively whilst microparticles cocktail concentrate was
diluted to I X by adding 5ml of RD2-1 diluent. The I X microparticles cocktail was then
added to wells of the microplate at 501l1/well before the addition of 50 III of sample and
standards following the predesign ELISA plate template. The microplate was then
incubated for two 2 hours on a horizontal orbital microplate shaker at 800rpm at room
temperature. After incubation, the microplate attached to a magnet and washed thrice.
The attachment of the plate to a magnet ensured that microparticles were not
accidentally washed away. Fifty microliters (50 Ill) IX Biotin-Ab cocktail was then
added to each well of the microplate and incubated for an hour on a microplate shaker
at 800rpm before washing thrice again. Streptavidin-PE diluted 24-folds was then
added to the microplate at 50 Ill/well and incubated on a shaker at room temperature
for 30 minutes after which the previous wash step was repeated thrice. A hundred
microliters (100 Ill) of wash buffer was then added to each well of the microplate and
incubated on a shaker for 2 minutes before reading with a Luminex 200 analyser not
later than 90 minutes after the last incubation step.
3.8 SNP Genotyping
All 27 SNPs were genotyped using the commercially available Sequenom
MassARRA Y iPLEX platform. This assay comprises several different experiments or
operations starting with DNA extraction from whole blood followed by an initial locus-
66
University of Ghana http://ugspace.ug.edu.gh

specific PCR reaction, a single base extension, and finally mass spectrometry that
identifies the SNP allele. Assay design and preparation of DNA samples were done at
Noguchi Memorial Institute for Medical Research, University of Ohana. Robotics of
the final iPLEX reaction and base calling was however performed on the commercial
Sequenom MassARRA Y platform at Inqaba Biotec, South Africa.
3.8.1 DNA isolation from whole blood
Genomic DNA was extracted from whole blood samples using Quick-gDNATM
MidiPrep from Zymo Research Corp. DNA isolation was done following
manufacturers protocol with minor modification (Zymo Research Corporation, 2014).
Exactly 0.6 ml of genomic lysis butTer provided by manufacturer was added - 150 ul
of whole blood and vortex for 4 - 6 second to mix completely. The mixture was allowed
to stand for 5 minutes at room temperature and before transferring it to the Zymo
Spin™ V-E ColumniZymo-Midi Filter™ provided by the manufacturer. The Zymo
Spin™ V-E ColumnlZymo-Midi Filter™ assemblage with its content was then
centrifuged at 1,000 x g for 5 minutes. The Zymo-SpinTM V-E Column was
disconnected from the assemblage and transferred to a collection tube and spun at
10,000 x g for 1 minute to remove any residue from the column. Exactly 0.3 mlofDNA
Pre-Wash ButTer provided by the manufacturer was added to the column and spun at
10,000 x g for 1 minute and the flow-through discarded. The previous step was repeated
twice with 0.4 ml of g-DNA Wash ButTer before transferring the column to a 1.5 ml
centrifuge tube. DNA Elution ButTer was then added directly to the column matrix and
allowed to stand for 2 minutes. The column matrix with its content was spun at 10,000
x g for I minute elute the DNA. The concentration of eluted was measured with
67
University of Ghana http://ugspace.ug.edu.gh

NanoDropTM 8000 Spectrophotometer (Thermo Fisher Scientific) and then store at -
SO°C.
3.8.2 Pre-PCR: DNA and oligo pool preparation
The concentration of gDNA isolated from whole blood was diluted to 2.5 nglut using
TE buffer and then divided into two aliquots of 2 J.ll/well in a 384-well PCR reaction
plate from (Marsh Biomedical). PCR plates with DNA a1iquots were kept at 4°C whilst
awaiting downstream reactions. Unmodified and standard purified Oligonucleotides
(oligos) (Integrated DNA Technologies) for PCR and iPLEX reaction were ordered for
al127 SNPs. Oligos for PCR were ordered at final equimolar concentrations of240 J.lM
(vendor ensured) in 96-well plates, however, they were used at a working concentration
of 1 IlM (table 3.1). Probes for iPLEX extension were ordered unmixed in a 96-well
deep plate at 250 to 450 J.lM (vendor ensured). To allow for multiplexing, oligo pooling
for PCR and extension was performed following the pre-designed assay pool plex and
oligo plate map. The oligos were divided into 3 groups based on their masses with the
highest mass group (Le. group I) diluted to 15 J.lM, medium mass group (i.e. group 2)
diluted to 10 11M. and the low mass group (Le. group 3) diluted to 5 J.lM. These
concentration adjustments were done to ensure that the peak. intensity of extension
primers is uniform. Oligos diluted to a working concentration were stored at 40C.
3.8.3 peR amplification of target loci
Exactly 2 III of prepared gDNA was dispensed into each well of a 384-well PCR plate
before applying the assay pool. Thus, each well received a different sample but the
same assay pool. Four microliters (4 J.ll) of PCR master mix was then added to each of
the wells to make a total reaction volume of 6 J.lUwell. The PCR reaction was then
68
University of Ghana http://ugspace.ug.edu.gh

perfonned with a PC-controlled thennal cycler (Thermo Scientific Hybaid 384-well
blocks) following the cycling program described in table 3.2.
Denaturation Annealing Extension Final extension Hold
1 cycle Final step
30 sec 1 min 3 min indefinite
3.8.4 PCR product clean-up with Shrimp alkaline phosphatase (SAP) protocol
The resultant PCR product was the "cleaned up" to remove all unincorporated dNTPs
using the SAP protocol. The SAP cocktail solution (akin to a master mix in a PCR
reaction) was prepared following manufactures protocol and added to the PCR plates
containing the PCR products at 2 Ill/well. The PCR reaction plate with its content was
then placed on a carrier and then mounted on the stackers in the post-PCR Multimek
SpectroPREP machine. The SAP program was then implemented in StakNet. The plate
was removed after the SAP operation, vortexed and then centrifuged at 425 x Ig at
room temperature to bring the solution to the bottom. The SAP reaction was then
incubated using the following program in a thennal cycler: 1 cycle: 40 min 37°C; 1
cycle: 10 min 85°C; final step: indefinite 4°C. The mixture was centrifuged at 425 x Ig
for I minute and then stored away at 4°C until needed for the downstream process.
69
University of Ghana http://ugspace.ug.edu.gh
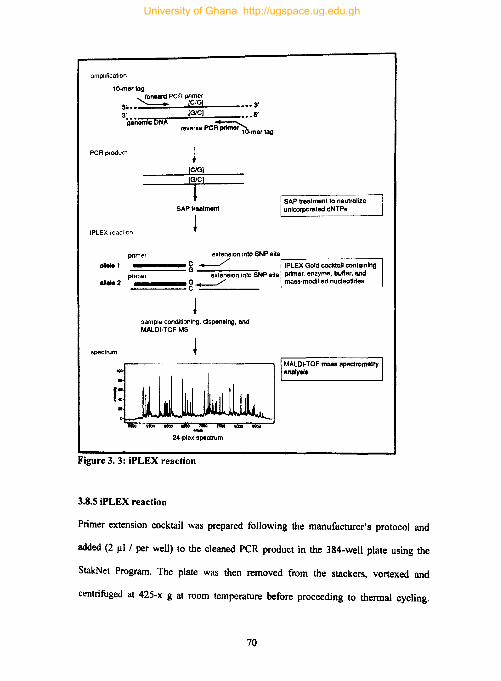
amplificatIOn
10·mertog --..!0rwan: PCR prtmer
5~ .. (C/G] _ -- 3' 3' [GtC) ___ S'
ganomlCDNA _ reverse PeR primei~mer log
PCR producl
[C/G)
(GlC)
I SAPlr8lllment
iPLEX reneltOn ~
SAP trealmenllo neulmlize unlcorpol'llted dNTPs
primer extension imo SNP alte,....... ________ ,
~~~~~ C ~ IPLEX Gold cocktail containing allele 1
a1 .... 2 - G extension Into SNP aite primer. enzyme, buller. and p~n~m~er~~~~ g ~ m;l8s.modlfled nucleotldes
spectrum
sample conditioning. dispensing. and MALOf· TOF MS
~
f~~1 24·plax spectrum
Figure 3. 3: iPLEX reaction
3.8.5 iPLEX reaction
MALOI-TOF malll spectrometry analytl.
Primer extension cocktail was prepared following the manufacturer's protocol and
added (2 Ill/per well) to the cleaned peR product in the 384-well plate using the
StakNet Program. The plate was then removed from the stackers, vortexed and
centrifuged at 425-x g at room temperature before proceeding to thermal cycling.
70
University of Ghana http://ugspace.ug.edu.gh

Thennal cycling for the primer extension reaction was done in Hybaid 384-well block
(Thenno Scientific) with the following cycling program (Table 3.3)
ili'R .Iii "'lim for iPLEX re.ction
initial denaturation 1 cycle sec denaturation 40 cycles 5 sec 940 C annealing 5 cycles 5 sec 52·C extension 5 sec 800 C final extension I cycle 3 min 720 C hold indefinitely 40 C
The primer extension product was cleaned up using SpectroCLEAN (Sequenom) to
remove salts such as Na+, K+, and Mg2+ ions and optimise for mass spectrometry
analysis. Resin slurry was prepared following the manufacturer's protocol and added
(16 Ill/well) to the post-PCR plate containing the product of primer extension with the
help of the SpectroPREP Multimek. The plate was removed after resin addition and
rotated in plate rotator before centrifuging at 425 x g for 3 minutes at room temperature.
The primer extension product was then spotted on SpectroCHIPs (Sequenom) and
primer extension detection was done with Compact mass spectrometer (Sequenom).
The resulting spectra are analysed by SpectroTyper software©
3.9. Other laboratory evaluations
Routine laboratory analysis and diagnostic tests were done in the laboratories of the
partner hospitals and results were made available to the research team. However, some
of these tests were repeated to meet the a priori stringent laboratory analysis of this
study.
71
University of Ghana http://ugspace.ug.edu.gh

3.9.1 Haematological analysis: Complete blood count (CaC) was done for all study
participants on successive visits using a 5part - Sysmex haematological analyser
(Sysmex Corporation. Japan). This operation outputs many haematological indices but
those of interest to this study included haemoglobin levels (Hb), total white blood cell
(WBC), platelets.
3.9.2 Parasitological evaluation: Blood films for malaria diagnosis was done in the
hospital laboratories following established protocols. Parasite density estimations were
done at NMIMR following the WHO protocol ("Malaria diagnosis," 1988; World
Health Organization. 2010). Total parasite biomass (Ptot) was estimated using a
formula proposed by Dondord and colleagues Ptot = 7.3·PtHRP2 ·(I-Hct (%»·body
weight (Kg)·1013, with PtHRP2 in giL (Dondorp etal., 2005; Hendriksen et aI., 2012).
On the other hand. peripheral blood parasitaemia (PbP) was estimated using the
formula: parasites/ilL· 1 06·blood volume, with blood volume defined as 0.08· body
weight [kg] (Dondorp et al., 2005; Hendriksen et aI., 2012). The difference between
Ptot and pbP makes up the sequestered parasites (Pseq).
3.9.3 Bacteraemia evaluation: bacteraemia was assessed in children recruited in the
CM group using a protocol described by Cheesbrough (Cheesbrough, 1984).
3.10. The use of Gaussian mixture model
Classification of endothelial integrity was based on secondary data from an earlier study
on the same samples which measured serum levels ofcEPC (Oduro, 2015). The need
to convert continuous biomarker data into discrete variables is common and although
some studies have used the mean- or median-split approach, that approach has come
72
University of Ghana http://ugspace.ug.edu.gh

under heavy criticism in recent times (DeCoster et al., n.d.; Iacobucci et aI., 2015;
Rucker et a\., 2015). Mixture models are preferred because unlike the mean- or median
split. it estimates cut-off points using the distribution of the variable or by optimizing
the correlation with outcomes (Budczies et al., 2012; Trang et aI., 20 IS). This study
used a Gaussian mixture model to covert secondary cEPC data on the same sample into
a binary variable called endothelial integrity with ProRepair (PR) and ProDamage (PO)
being the two possible states. This operation was implemented in Cutoff Finder
(Budczies et aI., 2012)
3.11. Statistical Analysis
Data were first tested for normality. skewness and kurtosis to determine whether they
were best suited for parametric or non-parametric statistical approaches. Analysis of
variance (ANOV A) was used in the comparing means across groups in situations where
the data were normally distributed and its non-parametric equivalents such as Mann
Whitney or Kruskal-Wallis was used in situations where the data was not normally
distributed. Receiver operating characteristic (ROC) curve was used in prospecting
biomarkers that may discriminate between CM and non-CM phenotypes.
Association ofSNP with trait was done using SNPstats software (Sole et aI., 2006). The
SNPstats analysis rubrics starts with the determination of allele and genotypic
frequencies followed by a test for Hardy-Weinberg equilibrium. Logistic regression
models were used to assess the association between SNPs and trait or disease because
they allowed interaction between SNPs and other factors. The estimation of the OR
(odds ratio) for each genotype was done with respect to a reference genotype. The
general logistic regression model used was:
73
University of Ghana http://ugspace.ug.edu.gh

109(.2..) = a + fJG + yZ 1-p
P = probability; G = SNP turned into a categorical variable and codified based on the
inheritance model assumed: Z = variables to adjust the model.
This equation, however, changed depending on the inheritance model assumed. Table
3.4 shows the logistic regression model used in each of the five inheritance models
assumed in this study. The Akaike information criterion (AIC) was used to detennine
the best inheritance model: the lower the score, the better.
In silico analysis to characterise the epigenetic context of genotyped SNPs was done
using the ChroMoS (Chromatin Modified SNPs), sTRAP, and MicroSNiPer web tools
(Barenboim et aI., 2010; Barenboim and Manke, 2013; Manke et aI., 2010). ChroMoS
was used to predict the chromatin state of the SNPs whilst sTRAP was used to predict
the effect of these SNPs on the binding affinity of transcription factors. MicroSNiPer
was used to predict the effect of the SNPs microRNA target sites.
74
University of Ghana http://ugspace.ug.edu.gh

Codominant model
Dominant model
Recessive model:
Over-dominant model:
Additive model:
IOg( L )=a+ ,8He+ r Z 1-p
tog( L )=a+ {JDo+ yZ 1-p
tog( l )=a+.BRe+yZ 1-p
tog( L )=a+PAd+YZ 1-p
3.12. Dealing with missing data
This is the most general model. It allows every genotype to give a different and non-additive risk.
A single copy of C is enough to modify the risk. Thus, heterozygous and homozygous genotypes have the same risk. A single copy of the allele is enough to modify the risk. Thus, heterozygous and homozygous genotypes have the same risk. Heterozygous is compared to a pool of both allele homozygous Each copy of the recessive allele modifies the risk in an additive manner.
Comparison
TIC vs TIT
CIC vs TIT
T/C-CC vs TIT
CIC vs TITTIC
TIC vs TITCIC
Like most studies, this study was beset with missing data problems that needed to be
addressed. Listwise deletion is the traditional way of dealing with missing data but this
approach has been criticised lately for two main reasons: the introduction of bias into
the data and the loss of statistical power. Fortunately, current advances in theoretical
and computational statistics have led to statistically sound techniques, such as mUltiple
imputations (MI), for handling missing data such (Deng et aI., 2016). This study
implemented a multiple imputation (MI) algorithm in SPSS. Downstream data analysis
highlighted differences in instances where MI datasets produced results that differed
from original data.
7S
University of Ghana http://ugspace.ug.edu.gh

CHAPTER FOUR
4.0 RESULTS
4.1. Demographic characteristics of study participants
This study involved 221 children (68 healthy controls, 60 uncomplicated malaria, 25
severe malarial anaemia and 68 cerebral malaria) aged between 11 months and 12 years
with a mean age of 5 .126 years. The mean age of study participants was highest in the
healthy controls (HC) and lowest in the severe malarial anaemia (SMA) groups (Table
4.1). Whereas the mean age ofHC was significantly higher in all pairwise comparisons,
that of UM was only higher than SMA (p = 0.012) but not CM (p = 1). Figure 4.1
summarises age differences among malaria phenotypes and healthy controls with a post
hoc pairwise comparison adjusted for multiple testing.
Sex ratio in this study population expressed as the number of males per 100 females
was found to be 135.106. Although the perfect sex ratio of 100 was only seen in healthy
controls, the distribution of males and females was not significantly different among
the different malaria phenotypes and healthy controls (p = 0.192).
76
University of Ghana http://ugspace.ug.edu.gh

Age distribution among malarial Phenotypes and post hoc comparisons
CM
~SMA ~ o c CII
f UM .!!
:; I ~ He l
Comparison
SMAvsCM
SMA'IUM
SMA 'IS He CM 'IS UM
eM 'IS HC UM vs He
6 10 12
Age (yrs)
P value
Unadjusted Adjusted
0.002 0.015 0.002 0.012
< 0.0001 < 0.001 0.873 1 0.001 0.005 0.002 0.010
/-jo/in plot.1 representing age ,hstrihlll/,,'; among malaria phenotypes. The black bars represent tht' first and third quartile ullil rhe white dots represent the median. The table bel/mth tile \/Olin plots shows mliltip/e PI' ,lIle comparison among malaria phenotypes. Each roll' /1/ Ih" ,uhlc lest Ihe nl/It Inpolhe.I/\· Ih,,: rhe age is the same in the malarial phenotypes being wlI1pared. Significance Ine! is {J.05.
Fi:!ufl' ... I: Age and malaria phenotypes
University of Ghana http://ugspace.ug.edu.gh

4.2 Haematological indices among study participants
Haematological indices provide valuable insight into the mechanisms of disease and
are therefore central to malaria pathogenesis studies. This study reports on the
association of haemoglobin (Hb) levels, platelets, and white blood cell counts (WBC)
with malaria phenotypes. Since Hb levels were invoked in the categorization of SMA,
a lower Hb level in this group was an insipid finding that only served as a confirmatory
test for appropriate disease categorization. A more interesting observation was the
finding of lower Hb levels in the CM group relative to HC (p = 0.004) and the lack of
significant differences between Hb levels in UM relative to CM (p = 0.280) and UM
relative HC (P = 1.000).
The comparison of white blood cell (WBC) counts among malaria phenotypes was done
using a non-parametric statistic (Kruskal-Wallis H test) and thus mean ranks are
reported. WBC counts in the SMA group was ranked highest whiles WBC counts in
HC was ranked the least with mean ranks of 149.86 and 87.33 respectively. Pairwise
comparison revealed a significantly lower WBC count in HC relative to CM (p = 0.013)
and SMA (p < 0.0001). Further pairwise comparisons revealed a significantly higher
WBC counts in the SMA group relative to UM (p < 0.0001). Mean platelet levels were
consistently higher in HC in all case-control companions at a significance level ofp <
0.0001.
78
University of Ghana http://ugspace.ug.edu.gh
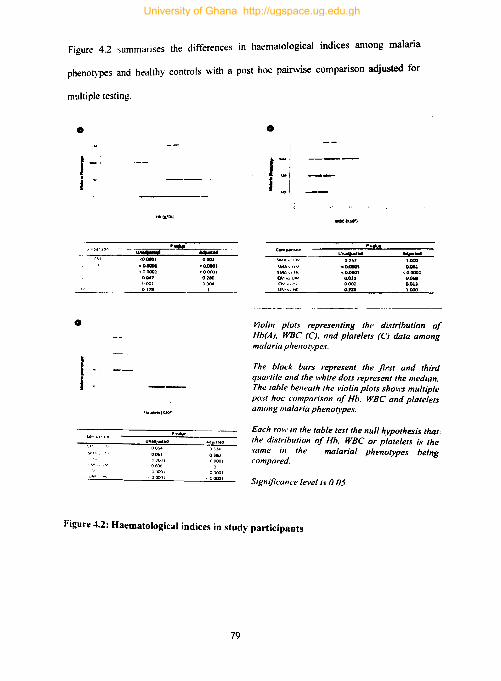
Figure 4.2 summarises the differences in haematological indices among malaria
phenotypes and healthy controls with a post hoc pairwise comparison adjusted for
multiple testing .
•
I i v
o
I .. J
<00001 0001
<O.CllOl cO...,1 ,,00001 <00001
0017 0280 0001 0004
0171 1
(..100'))
-00001
00001 I
•
f I
SM,. y~ ~H_ c;,... ... ~ UM CMv',tH
UMv\HC
woe, .... ,
0.157 .~
<0.0001 0.011 0.002 0.226
1.000
~ <o.oocu
O.OiII D.OU 1.000
Violin plot.~ representing the distribution of Hb(A). WBe (C). and platelets (e) data among malaria phenotypes.
The black bars represent the first and third quartile and the white dots represent the median. The table belleath the violin plots shows multiple post hoc comparison of Hh. WBe and platelets among malaria phenotypes.
Each row in the table test the null hypothesis that the distrihution of Hh. WBe or platelets is the .\·ame in the malarial phenotypes being compared.
Significance level is 0.05
Figure 4.2: Haematological indices in study participants
79
University of Ghana http://ugspace.ug.edu.gh
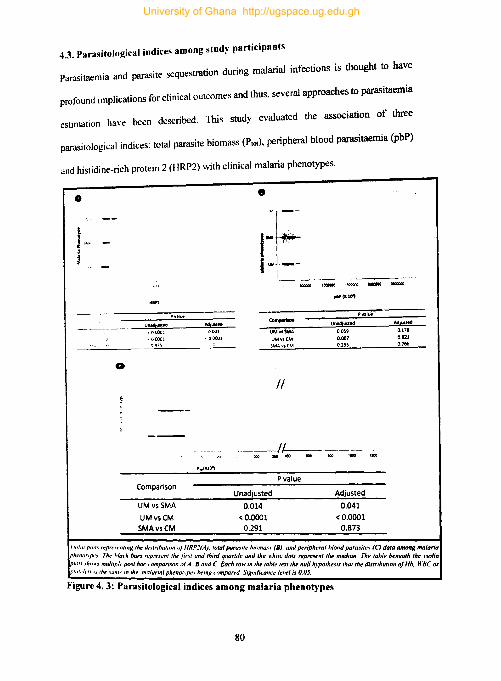
4.3. Parasitoiogiclil indices among study participants
Parasitaemia and parasite sequestration during malarial infections is thought to have
profound implications for clinical outcomes and thus, several approaches to parasitaemia
estimation have been described. This study evaluated the association of three
parasitological indices: total parasite biomass (PUll). peripheral blood parasitaemia (PbP)
and histidine-rich protein 2 (HRP2) with clinical malaria phenotypes .
• •
- -
•
-
~ 00001
-00001
Comparison
UMvsSMA
UMvsCM SMAvsCM
0001
·00001 , UMVISMA UM'IICM
SMAV10CM
II
UNdlus,od
00~9
0.007 0.2~~
pval\.if:'
----/~------lOCI 110" IOC
Unadjusted
0.014 < 0.0001
0.291
P value
Adjusted
0.041 <0.0001
0.873
Adlusted
0.178
0.021 0.766
l/fl/", pltll,~ Il'pn.·,,:nttng th~ ,1J.lilnhulllln orHRP'(A) lolul puru~ I h !~~'.nunp.., Th,. hiade ban "'1',.. .. '"0/ Ih,. jirsl ,,:d I;'inl quartii: :1I;(I,~a.".:B). ;nd peripheral h/o"d paru.tlle., (e) dala uMong malaria I'''''''. ,h" .. , mull/p,' •• po." hot: wmpomon "rA, B Gild C Each row i I ,f! \I III' ols I"PpI"Fsenllhr melltun. The table beneallr lire ",olin 1P1ul•I,.,., " lire 'am, rn Ih" malaria/ plrenol\'~' ""lnO wm"" d S n .Ir,~ ,able lesllh" null "rpolhes,., Ihallhe distrlbullOn of Hb WSC ar
• . r" ~ r-I"F 'lint 'cum'e lel-e/,.' O.[}S. .
Figure 4. 3: Parasitological indices among malaria phenotypes
80
University of Ghana http://ugspace.ug.edu.gh

PlOIranked highest in CM (93.57) and least in UM (54.58). Pairwise comparison revealed
PlOt to be significantly higher in CM relative to UM (p < 0.0001) but not SMA (p = 0.873).
Further pairwise comparison revealed Plol to be higher in SMA relative UM (p = 0.041).
Comparison of HRP2 and pbP revealed similar trends albeit at higher a-levels (Fig 4.3).
4.4 Immunological indices among study population
Immunology plays an important role in malaria pathogenesis and several studies have
explored the roles immunological parameters play in the pathogenesis of malaria. This
study explored the relationships between four cellular immunity parameters (neutrophils,
lymphocytes, CD4 and CD8), malaria phenotypes, and endothelial damage. A Kruskal
Wallis H test found lower neutrophil levels in HC relative to UM (P < 0.0001), CM (P <
0.0001), and SMA (P = 0.017). Neutrophils levels were also higher in UM relative to
SMA but comparable with levels in CM (p =1). Lymphocytes levels were highest in HC
(46.5) and least in UM (25.70). All pairwise comparisons of lymphocytes levels across
malaria phenotypes revealed significant differences at p < 0.0001 except in the
comparison between CM and UM where lymphocytes levels in CM were significantly
higher at a borderline p-value (p = 0.043). Similarly, CD4+ T cells were highest in the
HC (14.30) and least among the UM (4.6), here again. all pairwise comparisons were
significant at a p < 0.0001 except for the CM and SMA comparison which differed at p
= 0.022. Levels of CD8+ T cells followed similar trends save that the CM and UM
comparison which did not reveal significant differences p = 0.078.
81
University of Ghana http://ugspace.ug.edu.gh

4.5 Angiogenic indices among clinical malaria phenotypes.
Table 4. 1: Association of angiogenic factors with malaria phenotypes
Angiogenic Factors
SDF -I hlrnmal ccll-deri,ed laclor I)
NG R I (nclireguhn 1 ) TREM I \nl!ll'nn~ ''"1''''' cxprnscd myelOId ,dl, II
Ang-I (anglOpmellnl)
Ang-2 (anglopmelin21
Ang-2/Ang-1 (ang-2,ang-1 mho)
CAl ,,:arbon I, dIlhydrase I)
Tie2Ianglop",ctinrcceplOr) SI 008 1<'100 ,.iclum-b,ndons protem HI
M M P9r rrtatr" mctallopepl,dasc 9)
Mean Rank Endothelial Integrity • 62.28 74.39 59.85 71.01 61.94 58.57 71.18 65.06 67.41 64.53
67.02 53.29 69.77 57.12 64.40 71.22 56.93 63.87 61.20 64.47
P-value
0.471 0.001 0.131 0.034 0.41 0.47
0.030 0.856 0.344 0.992
Angiogenic factors as used in this study refers to a loose collection of receptors, cells.
molecules, enzymes and proteins that are directly or indirectly involved in the process of
angiogenesis. They comprise S 1 00 calcium-binding protein B (S 1 OOB), Stromal cell-
derived factor 1 (SDF-l), Neuregulin 1 (NRGl). Triggering receptor expressed on
myeloid cells 1 (TREM-l), Angiopoietin-l (Ang-l), Angiopoietin-2 (Ang-2), Carbonic
anhydrase (C1\ 1 ), Tyrosine-protein kinase receptor (Tie-2) and Matrix
metalloproteinase-9 (MMP9). The levels of these factor among malaria phenotypes are
presented in Tables 4.1. The comparison was limited to excluded SMA because of the
potential of anaemia to influence angiogenic factors to obscure the findings of this study
(Dunst et al.. 1999).
4.6 Endothelial integrity, malaria phenotypes, and angiogenic factors.
Secondary cEPe data on the samples used in this study was fitted to a Gaussian mixture
model to dichotomise endothelial integrity into Pro-Repair and Pro-Damage. To this
end, a histogram of cEPC data was plotted and then fitted with a mixture model of two
Gaussian distributions. This was implemented in "Cutoff Finder", a web tool that uses
82
University of Ghana http://ugspace.ug.edu.gh

the tlemix function from the R package flemix (Budczies et al.. 2012; Leisch, 2004).
An optimal cut-off value of 0.07234 was returned after implementation and study
participants who recorded cEPe levels less than 0.07234 were classified as Pro
Damage and those with cEPe level above the optimal cut-off were classified as Pro
Repair. After this classification. 41 % of the study participants were classified as Pro
Repair and 59% classified as Pro-Damage.
Post categorization and to address objective five of this study, a series of amenable
statistical tests were conducted. To this end. a chi-square test revealed a significant
association between endothelial integrity and malaria phenotypes (X2
= 13.331, df = 3,
p = 0.004, Fig 4.5) with over 70% of the eM cases occurring among the Pro-Damage
phenotype.
Figure 4.4: Histogram of EPCs in tbe study popUlation
83
University of Ghana http://ugspace.ug.edu.gh
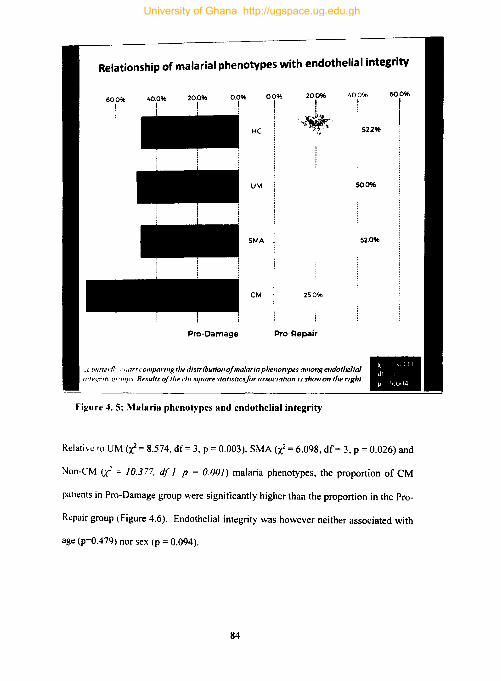
Relationship of malarial phenotypes with endothelial integrity
20.0% 400%
I I HC
.... ~ . .... '. .. t, 52296
UM 50.0%
SMA 52.0%
CM 250%
Pro-Damage Pro-Repair
.1 bll!Terf/, , lIor' comparlllg 'he distribuNollo!lIIatnria phellotypes alllong e"dolhe/iat 111'"~''''' <:' UIII'I Resu/'s of tlte clll sqllare statistics!or assoc/aholl /s shaw 011 the right.
figure 4. 5: Malaria phenotypes and endothelial integrity
60.0%
I
Relative to UM <x: = 8.574, df= 3, p = 0.003), SMA (x: = 6.098, df= 3, p = 0.026) and
Non-eM Li = 10.377. df I. P = 0.0(1) malaria phenotypes, the proportion of eM
patients in Pro-Damage group were significantly higher than the proportion in the Pro-
Repair group (Figure 4.6). Endothelial integrity was however neither associated with
age (p=0.479) nor sex (p = 0.094).
84
University of Ghana http://ugspace.ug.edu.gh

~
ProDamage
ProRepair
'i: DO Pro-! Damage c
.!! Qj .c Pro+' o Repair "C
Jf = 8.574, d/ =1, P = 0.003
0; >i', 3b.l%
.UM .CM
Frequency (%)
• SMA .CM
C L&.I
-'----L-...--L..-..L----'--'--.L.---'---'--':--:t~:x, Frequency (%)
ProDamage
ProRepair
Jf = 10.317, dl =1, p = 0.001
/I 1 lIS ~%
Frequency (%)
Figure .... 6: Endothelial integrity and malaria pbenotypes
85
_NON-CI\ _CM
Frequency (%)
University of Ghana http://ugspace.ug.edu.gh

4.7 Association of endothelial integrity with ke~ haematological, parasitological,
immunological and angiogenic variables.
Although endothelial integrity was neither associated with haematological nor
parasitological variables, it was found to be associated with CD4+ levels and a few
angiogenic factors (Table 4.2)
Table .'-2: Association of key variables with endothelial integrity
Ke~ variable Mean Rank Mann-Whitney
U Test P-value
1~!I~~!I!lDlIllIllIllr:~~""""""""" ",jij.lijjMiiMli; Hb (g/dL) mean 94 0.295 102.59 4944.5 PLT(Xlo-') 103.62 0.067 88.59 3839
WBe (X 10")
IQi""iMi!igll· P. den\ity (x 103) P. burden (1110") P. CIR (x IOJ
)
HRP2
I mmunulogll'al L~mphocyte
""culrophils CD4+ T cells CD8+ T cells
lflNi!liMM"" ~DF-I hlrprn;JI \.ell~dl!n\C'J tal..:tor I) NGRI ,""",'.',tion I)
TRE M II fn~'!-, _ !ljo: ~,..~ptOf c .. prc·ucd m\clPld ~(1I~ I!
Ang-I,angillpmeunil Ang-2 (angloJl<1I<lon21
Ang-2/Ang-1 l.mg-2iang-1 ratio,
C A I I,arb"m, anh) drase I ,
Tie2 (angillpolctm receptor)
SIOOB ''''''tI calCium bIRding rrtlkJlI HI
101.17
93.98 81.92 74.69 85.71
90.71 102.13 88.10 94.55
78.05 88.86
81.72
90.15 83.36 109.10 90.35 91.94
83.96
92.16
95.23 69.37 80.58 80.15
102.53 89.60 109.85 100.54
86.93 69.68
78.72
69 75.02 113.67 71.42 70.57
79.19
4121
4347 2332 3005 3079
4892 3918.3 5518 4782
3460 3894 2972
2626 2721
6179.5 2510 2454 3028
0.273
0.876 0.087 0.422 0.466
0.146 0.126 0.008 0.464
0.240 0.010
0.688
0.023 0.361 0.600 0.012 O.OOS
0.526 M MP9(maim metaliopcPllda.,c Y, 85.06 91.13 3894 0.436
Children with the Pro-Damage endothelial integrity phenotype had significantly lower
median CDot.,.. relative to the Pro-Repair phenotype (p = 0.008). As shown in Table 4.2,
all angiogenic factors associated with endothelial integrity were higher in the Pro-
Damage phenotype (Tie-2: p = 0.005; NGRl: p = 0.010; CAl: p = 0.012; ANGI: p =
0.023).
86
University of Ghana http://ugspace.ug.edu.gh

4.8. Genotyping results.
To achieve objectives 2, this study genotyped a focused panel of 27 SNPs (tag SNPs)
in 16 different loci on 10 different chromosomes. Conformity to Hardy-Weinberg
equilibrium (HWE) is used as an indicator of proper sampling and genotyping in most
genetic studies, however. a deviation from HWE can also be an indicator of the
existence ofa gene-disease association (Lee. 2003; Salanti et al., 2005). Twelve of the
SNPs genotyped in this study deviated from HWE at a significance threshold of 0.05,
however, only 8 SNPs deviated from HWE after correcting for multiple testing (Table
4.3).
4.9 Association of tag SNPs with endothelial integrity and malaria.
Detail results of tag SNPs and their association with endothelial integrity and malaria
phenotypes are shown in Tables 4.4 -18. These tables are organised by chromosomes
and for each chromosome, several case-case and case-control comparisons were done
to determine the association of tag SNPs with malaria phenotypes and endothelial
integrity. Data analysis for this section generated a lot of results, some of which are
presented in the appendix as supplementary data. The tables presented here are a
summary of the most interesting findings. This study prioritised the discrimination of
eM from UM and thus associations of SNPs in CM versus UM and ProDamage versus
Pro Repair comparisons are prioritised.
87
University of Ghana http://ugspace.ug.edu.gh
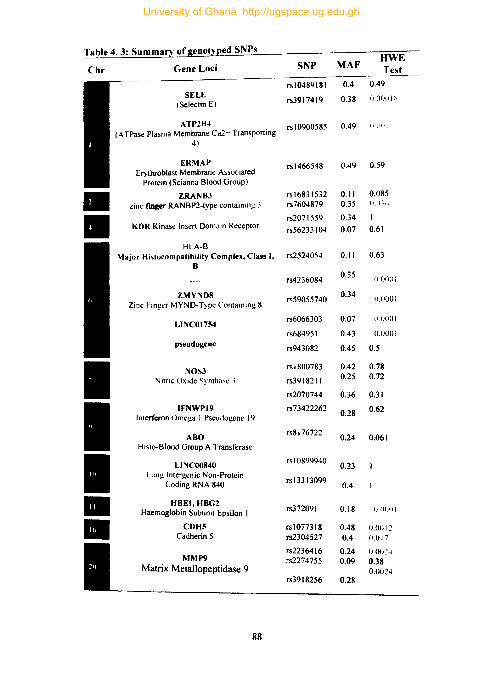
Table 4. 3: Summarv ofgenotyped SNPs "WE
Chr Gene Loci SNP MAF Test
rsl0489181 0.4 0.49 SELE
rs3917419 0.38 0.0f)(l1 ~ (Selectin E)
ATP2B" rs I 0900585 0.49 (1(11 1
(ATPas~ Plasma Membrane Ca2'" Transporting 4)
ERMAP rs I 466548 0.49 0.59 El)throblast Membrant: Associated
Protein (Scianna Blood Group) • ZRANB3 rsl6831532 0.11 0.085
zinc rmger RANBP2-typt: containing ~ rs7604879 0.35 0.1);"
• rs2071559 0.34 KDR Kinase Insert Domain Receptor rs56233 104 0.07 0.61
HLA-B Major Histocompatibility Complex. Class I. rs2524054 0.11 0.63
B
rs4236084 0.35
II (JOOI
ZMYND8 rs59055740 0.34 f).OOOI
Zinc Finger MYND-T)'pe Containing 8
LlNCOl754 rs6066303 0.07 1I.IJOOI
rs68495I 0.43 0.0001
pseudogene rs943082 0.45 0.5
I NOS3 rs I 800783 0.42 0.78
Nitric Oxide Synthase, rs3918211 0.25 0.72
rs2070744 0.36 0.31
I IFNWP19 rs73422262 0.62
Interferon Omega I Pseudogene 19 0.28
ABO rs8176722
0.24 0.061 Hislo-Blood Group A Transferase
I L1NCOO840 rsl0899940
0.23 L.ung Intergenic Non-Protein
rsl3313099 Coding RNA 840 0.4
• HBEI. HBG2 rs372091 Haemoglobin Subunit Epsilon I 0.18 i) (lfilli
• CDH5 rs1077318 0.48 0.0012 Cadherin 5 rs2304527 0.4 0.017
rs2236416 0.24 ().O(j2~ MMP9
Matrix Metallopeptidase 9 rs2274755 0.09 0.38
0.OO~4 rs39 I 8256 0.28
88
University of Ghana http://ugspace.ug.edu.gh

4.9.1. Tag SNPs on chromosome I
Four of the SNPs assayed in this study were located on chromosome 1. Two of these
SNPs, rs10489181 and rs3917419, were in the gene encoding E-selectin (SELE) whiles
the third (rs10900585) and a fourth (rs1466548) were in the genes encoding ATPase
Plasma Membrane Ca2+ Transporting 4 (A TP2B4) and Erythroblast Membrane
Associated Protein (ERMAP) respectively. Although none of the SNPs on chromosome
I were associated with endothelial integrity, variants ofrs3917419 were associated with
an increased risk of CM in the CM versus UM comparison (Table 4.4). In the less
prioritised case-control comparisons, all SNPs on chromosome I were associated with
reduced risk of malaria except a variant of rs3917419, which was associated with
increased risk of malaria in the recessive inheritance model (Table 4.5). However, AIC
score suggested that the over-dominant inheritance model, rather than the recessive
model, was most probable.
89
University of Ghana http://ugspace.ug.edu.gh

Table 4.4: Chromosome J: Association ofS!,/Ps l\'i~(:l\lin _('M n lll\1 eumpa"hull
Loci SNP IM* Genotype Malaria Phenotype
OR (95% CI) P-value Ale liM eM
II'I.'IIIH:UII:J_ co 1 ( 34 (56.700' 35 (51.5%) 0.66 (0.30-1.44) 0.42 11l1.2 CC 10 (16.70'0) 8(11.8%) 0.51 (0.17-1.57)
0 TIC-CC 44 (73.3%) 43 (63.2%) 0.63 (0.29-1.33) 0.22 179.4 R CIC 10(16.7%) 8(11.8%) 0.67 (0.24-1.82) 0.43 ISO.3
00 TIC' 34 (56.7~0) 35 (51.5%) O.SI (0.40-1.63) 0.56 180.6 LA 0.70 (0.41-1.20) 0.19 179.3
CO GIA 43 (71.7%) 53 (77.9%) 1.90 (II XI-~ -<'I) O.u2:' 175.6 AlA o (0° .. ) 4(5.9%) NA
0 G/A-AIA 43 (71.7%) 57 (S3.8%) 2.05 (0.8704.S2) 0.096 178.2 R AlA 0(0%) 4(5.9%) NA 175.8
00 G'A 43 (71.7%) 53 (77.9%) 1.40 (0.63-3.12) 0.41 ISO.3 LA 2.39 ( I.OS-5 .28) 0.027 176
II'I!II!~ CD CIG 33 (55%) 40(58.8%) O.SI (0.32-2.03) 0.42 ISI.2 GIG 17 (2S.3%) \3 (19.1%) 0.51 (0.17-1.50)
0 C/G-G/G 50 (S3.3%) 53 (77.9%) 0.71 (0.29-1.72) 0.44 180.4 R GIG 17(28.3%) 13 (19.1%) 0.60 (0.26-1.36) 0.22 179.4
00 CG 33 (55%) 40 (58.8%) 1.17 (0.58-2.36) 0.66 180.S
• LA 0.71 ~0.41-\.21) 0.21 179.3 Model (1M) Codominal1l (CD): Dominant (D); Recessil'e (R): Overdominant (OD); Log-additive (LA).
AssuminK bial/elic SNPs comprising X and Yalleles. the reference genotypes/or the CD. D, Rand OD inheritance models are XX. xx. XY-YY and XX-rr respectively. Red highlight: Susceptibility SNP. Green highlight: protective SNP
90
University of Ghana http://ugspace.ug.edu.gh
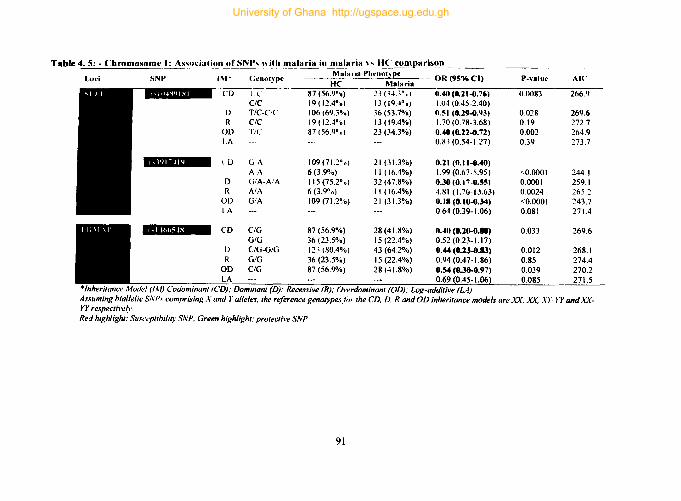
Table 4. 5: - Chromosome I: As,ul'iati_'.!n of SN~, l\ HI1 malaria in malaria n 11(' comparison .----- ------~ .. Loci SNP IM* (;enotype
Malaria Pht'notlpe - OR(9S%CI) P-value AI('
HC Malaria --.~-
CD I.l 81 (56.9°'0) 23 (1.t.3°,u) 0.40 (0.21-0.76) 0.0083 266.Q ClC 19 (12.4°0) 13 (19,4° 0 ) 1.04 (0.45-2.40)
D T/C-ClC 106 (69.3%) 36(53.1%) O.~I (0.29-0.93) 0.028 269.6 R CIC 19 (12,4°'0) 13 (19.4%) 1.70 (0.18-3.68) 0.\9 212.7
OD T,C 87 (56.9°0) 23 (34.3%) 0.40 (8.22-0.72) 0.002 264.9 LA 0.S.1 (0.54-1.27) 0.39 273.7
~ CO GA 109(71.2°0) 21 (31.3%) 0.21 (0.11-0.40) AA 6(3.9%) 11 (16.4%) 1.99 (0.61-5.95) <0.0001 244.1
D G/A-NA 115 (75.2%) 32 (47.8%) 0.30(0.17-0.~) 0.0001 259.1 R AlA 6 (3.90.) II (16.4%) ·I.S I (1.70- 13.63) 0.0024 265.2
OD G/A 109 (71.2%) 21 (31.3%) 0.11 (0.10-0.34) <0.0001 243.7 LA 0.64 (0.39-1.06) 0.081 271.4
..... l!rII&1!~ CD C/G 87 (56.9%) 28 (4U!%) 0.40 (8.20-0.88) 0.033 269.6 GIG 36 (23.5%) 15(22.4%) 0.52 (0.23-1.17)
0 C/G-G/G 12:> (80.4%) 43 (64.2%) 0.44 (8.23-0.13) 0.012 268.1 R GIG 36 (23.5%) 15 (22.4%) 0.94 (0.47-1.86) 0.85 274.4
OD C/G 87 (56.9%) 28 (41.8%) 0.54 (0.30-0.97) 0.039 270.2 LA 0.69 ~0.45-1.06} 0.085 271.5
·Inheritance M-;;del (IA/) Codominant (CD); Dominant (D); Recessive (R); Overdominant (OD); Log-additive (LA) Assuming biallelic SN P.I comprising X and r alleles, the reference genotypesjiJr the CD, D. R and OD inheritance models are XX, xx. X}"-)'Y and XX-YY respectil'e~\·. Red highlight: Sust't!plibiliry SNP, Green highlight: protective SNP
91
University of Ghana http://ugspace.ug.edu.gh

4.9.2. Tag SNPs on chromosome 2
Two of the SNPs considered in this study occurred in the locus encoding zinc finger
RANBP2-type containing 3 (ZRANB3). None of the variants of rs16831532 and
rs7604879 in ZRANB3 were associate with endothelial integrity or eM. However,
rs7604879 was found to be associated with an increased risk of malaria in the case
control comparison (Table 4.6). Although this SNP was associated with malaria in two
inheritance models. an Ale score of 266 indicates that the over dominant inheritance
model is the most probable (4.12).
4.9.3. Tag SNPs on chromosome 4
The locus encoding Kinase Insert Domain Receptor (KDR) harboured two of the SNPs
(rs2071559 and rs56233 104) assayed in this study. These SNPs were not associated
with any of the malaria phenotypes in case-case comparisons. However, a variant of
rs2071559 was associated with protection from clinical malaria in general in the
recessive inheritance model (OR = 0.27,95% eI = 0.10 - 0.75, P = 0.012, Table 4.7)
92
University of Ghana http://ugspace.ug.edu.gh
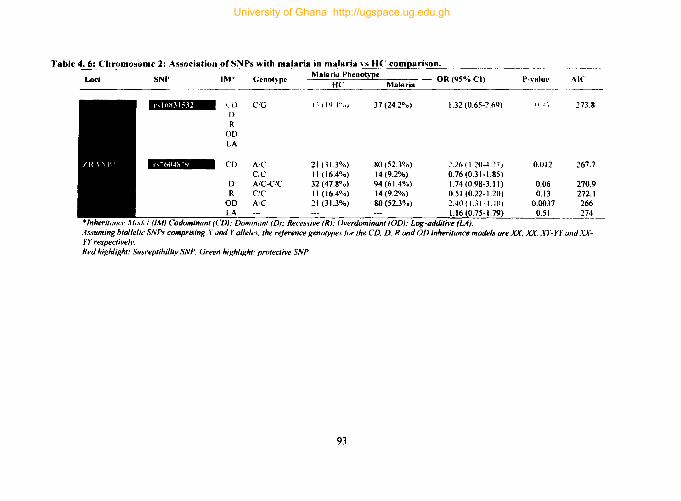
Table 4. 6: Chromosome 2: Association of SNPs with malaria in malaria, \s H(' comparison. , ______ ,
Loci SNP IM* Gcnot~'Jlc
~ C'j) CIG I)
R OD LA
CD Air CC AfC-ClC ("IC
A'C
Malaria Phcnotypc _ OR (95% el) He ~M'!.'a~18~r:.!:ia~ _____ _
1 ~ I 1'1 -to oj
21 (31.3%) II (16.4%) 32 (47.80 0)
11 (16.4%) 21 (31.3%)
37 (24,20 .)
80(52.3%) 14(9.2%) 94 (6],4%) 14(9.2%) 80(52.3%)
1.32 (0.65-~,69)
~ ,26 ( 1.20-427 J
0.76 (0.31-1,85) 1.74 (0.98-3.11) 0,51 (0.22-1.20) 2.40 (U I--UO) 1.16 (0.75-1.
: Dominant (0): Reces,~ive (R): Overdominanl (OD): Log-additive (LA),
P-\alue
Ill;
0.012
0.06 0.13
0.0037 1
Ale
273.8
267.7
270.9 272.1 266 274
Assuming bial/elic SNPs comprising .r LInd r alleles, the reference genotype.\jor the CD, D, Rand OD inheritance models are n, XX, XY-YY and AXrr respective(l', Red highlight: SUJceptihility SNP, Green highlight: protective SNP
93
University of Ghana http://ugspace.ug.edu.gh

Table 4.7: Chromosome 4: Association ofSNPs "ith malaria in a malari~'-~ .. li}IC compar:~'..o.!'.. ....
Loci SNP 1M" Genot,pe Malaria Phenotype
OR (95% ('I) P-\alul' Ale fI( Malaria
CD (jIA 29 (43.~%) 84 (54.90 0) 1.31 (0.71-2.42) 0.029 269 D G/A-A/A 39 (~1!.2%) 91 (59.5%) 1.05 (0.59-1.89) 0.86 274 R A'A 10(1 .. 1.'1%) 7 (4.60 '0) 0.27 (o.l~.75) 0.012 268
()[) (j,'A 29(43.3%) 114 (54.9"0) 1.60 (0.89-2.85) 0.11 272 LA 0.79 (0.49-1.25) 0.31 273
.atIII&I.IDl! CD Te 10 (14.9°0) 19(12.4%) 0.81 (0.35-1.85) 0.62 274 D R
OD
Model (1M) Codominant (CD): Dominant W); ReL'essive (R). Owrdominant (aD): Log-additive (LA). Assuming biallelic SNPs comprising X and r al/e1es. the re{l!rt'ncc 1!.en(}~ype.'for the CD. D, R and aD inheritance models are XX, XX. XY-YY and XXYY respectil·ely. Red highlight: Susceptibility SNP. Green highlight: pmleclh'e SNP
94
University of Ghana http://ugspace.ug.edu.gh

4.9.4. Tag SNPs on chromosome 6
Six of the 27 tag SNPs considered in this study were located on chromosome 6. Three
of these SNPs were in pseudogenes (rs684951) or noncoding regions (rs4236084 and
rs943082) of the human genome. The other SNPs: rs2524054, rs59055740, and
rs6066303 were in the loci encoding Major Histocompatibility Complex, Class I, B
(HLA-B), Zinc Finger MYND-Type (ZMYND8), and Long Intergenic Non-Protein
Coding RNA 1754 (LINCOI754) respectively. Apart from rs684951 which afforded
some protection from endothelial damage in the recessive inheritance model (OR =
0.45,95% CI = 0.24 - 0.87, p = 0.014, Table 4.8), none of the SNPs on chromosome 6
were associated with the outcomes of interest.
95
University of Ghana http://ugspace.ug.edu.gh
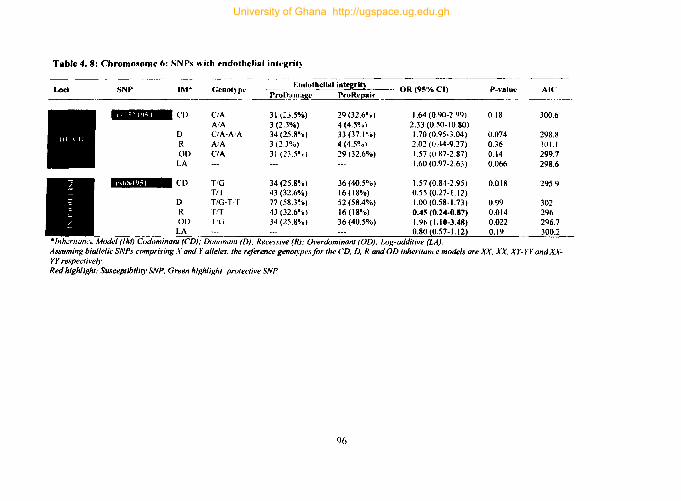
Table 4. 8: Chromollome 6: SN Ps ~ ith endothelial inh'gri~
Loci SNP 1M· Genotype Endothelial intrgritl OR (95%CI)
ProJ}_arnage ProRe(!air
~Cf) CiA 31 (2".5%) 29 (32.6~.) 1.64 (0.90-2.99) AlA 3(2.3%) 4 (4.5'10) 2.33 (0.50-10.80)
f) CIA-AlA 34 (25.8°'0) 33 (37.10 0) 1.70 (0.95-3.04) R AlA 3 (2.3%) 4 (4.5~0) 2.02 (0.44-9.27) Of) CIA 31 (23.5°0) 29 (32.6%) 1.57 (U.87-2.87) LA 1.60 (0.97-2.63)
nElDA CD T/G 34 (25.8%) 36 (40.5%) 1.57 (0.84-2.95) Trr 43 (32.6%) 16 (18%) 0.55 (0.27-1.12)
D T/G-TT 77 (58.3%) 52 (58.4%) 1.00 (0.58-1.73) R TiT 43(32.6%) 16(18%) 0.45(0.24-0.17) Of) I'(j 34(25.8'10) 36(40.5%) 1.96(1.10-3.48)
LA 0.80 *lnherilal1ct: Model (1M) Codominant (CD): Doml1lanl (D): Recessive (R): Overdominant (aD): Log-additive (LA).
----_ .. -P-valuc
0.18
0.074 0.36 0.14 0.066
0.018
0.99 0.014 0.022 0.19
AIC
300.6
298.8 .10 I.! 299.7 298.6
295.9
302 296 296.7 300.2
Assuming biallelic SNPs comprising .r and Yalleles. the reference genotypes for the CD. D. R and aD inheritance models are n. XX; XY-YY and X\'YY respectil·ely. Red highlight: Susceptibility SNP. Green highlight: protective SNP
96
University of Ghana http://ugspace.ug.edu.gh

4.9.5. Tag SNPs on chromosome 7
The SNPs on chromosome 7 were all located in the gene encoding Nitric Oxide
Synthase 3 (NOS3) and comprised rs1800783, rs3918211, and rs2070744. Although
variants of rs 1800783 were associated with an increased risk of endothelial damage
(OR = 2.38,95% CI = 1.17 - 4.85, P = 0.054, Table 4.15), none of the SNPs were
associated with CM (Table 4.9). A summary of the association of variants ofrs3918211
and rs2070744 with clinical malaria is shown in Table 4.10.
97
University of Ghana http://ugspace.ug.edu.gh

Table 4. 9: Chromosome 7: Association ofSNPs with endothelial int~ri~'
Loci SNP IM* Genot~pe F:ndothelial Inte~rit~
- OR (95%('1) P-value Ale _---.!!(IDall.!.a~ ProRe~air
rs180078~ CD r'A 70(53%) 40(44.~0) 0.97 (0.53-1.80) 0.054 298.1 AlA 16(12.1%) 22 (24.7%) 2.34 (1.0~-5.22)
0 T/A-A/A 86 (65.2%) 62 (69.7°0) 1.23 (0.69-2.19) 0.48 .101.) R AlA 16(12.1%) 22 (24.7'lo) 2.311 (1.17-4."<;) 0.016 296.1 00 TIA 70 (53°.) 40 (44.9°;0) 0.72 (0.42-1.24) 0.24 300.6 LA 1.43 (0.97-2.13) 0.071 298.7
rs3918211 CO TIC 52 (39.4%) 43 (48.3%) 1.60 (0.89-2.88) 0.26 301.3 CIC 18(13.6%) 14 (15.7%) 1.51 (0.66-3.42) TIC-CiC 70 (53%) 57 (64%) 1.58 (0.91-2.74) 0.1 299.3
R CIC 18 (13.6°'0) 14 (15.7%) 1.18 (0.55-2.52) 0.67 301.8 00 TIC 52 (39.4%) 43 (48.3%) 1.44 (0.84-2.48) 0.19 300.2 LA 1.30 (0.89-1.92) 0.17 )00.1
rs2070744 CD TIC 46 (34.9°0) 40(44.9%) 1.62 (0.92-2.84) 0.19 300.6 CIC 6(4.5%) 6(6.7%) 1.86 (0.57-6.12)
0 T/C-C/C 52 (39.4%) 46(51.7%) 1.65 (0.96-2.83) 0.071 298.7 R C'C 6 (4.5%) 6(6.7%) 1.52 (0.47-4.87) 0.48 301.5 00 TIC 46 (34.9%) 40(44.9%) 1.53 (0.88-2.65) 0.13 299.7 LA 1.49 ~0.95-2.34~ 0.079 298.9
el (1M) Codominanl (CD). Dominant (D): Recessil'e (R): Overdominanl (OD): Log-additive (LA). A.uuming biallelic SNPs comprising X and r alleles. the reference genotypes for the CD. D, Rand OD inheritance mndels are XX, Xx. XY-YY and XX-YY respectively. Red highlight: Susceptihility SNP. Green highlight: protectiw SNP
98
University of Ghana http://ugspace.ug.edu.gh

Table 4. )0: Chromosome 7: Association SNJ', "ith malarht ill a malaria " II<' comparison
Loci SNP
rs1H0078.1
rs3918211
rs2070744
I~" Genol~pe Malaria Phenotype
OR(9!'%CI) Malaria HC ---------
III
f)
R Of)
IA
CD
D R
on LA
CD
()
R
I \ i7(50.~Il,d "l~lj 20/0) 1.11 (0.5R-2 151
AlA 24 (15.7001 14PII'l"iJ) 1.52 (0.M-~5\11 r:A-A-A 101 (660'0) 47 (711.2%) 1.2\ (0.65·2.251 AlA 24 (15.7°01 14 (20.(1''<» 1.42 (0.68·2.95) riA 77 (SOYo) 33 (·t<l2%) \I % (0.54-1.70)
\ .22 (0.80-1.84)
)'II.' \0(14.9%) 84 (54.9%) 5.45 (2.51-11.84) OC 20 (29.9~0) 12 (7.X%) 0.39 (0.17-0.19) T/C-Ot' 30 (44.8°0) 96(62.8%) 2.OS (1.16-3.72) CIC :!0(29.9%) 12(7.8%) 0.20 (0.09-0.44) \'1(' 10(14.9%) 84 (54.9%) 6.94 (J.JO-14.60)
0.92 (0.61-1.38)
TIC 84(54.9%) 10(\4.9%) 0.18(0.08 ... .40) CIC 12 (7.8%) 20 (29.9%) 2.57 (1.12.!'.87) T/C-CiC 96 (62.8°0) 30 (448%) 0.48 (0.27-0.86) CIC 12 (7.8%) 20 (29.9%) 5.00 (2.27-11.00) TIC 84 (54.9%
) 10 (14.9%) 0.14 (O.87 ... .l8)
A I. -Inheritance Model (1M) Codominant (CD); Dominant (D): Recessive (R): Overdominant (OD); Log-additive (LA).
P-"alue
0.62
(l.5S n.35 0.88 0.36
<0.0001
0.013 <0.000\ <0.0001 0.7
<0.0001
0.013 <0.0001 <0.0001 0.7
AI(
---~---
275.5
274.\ 2736 2744 273.6
238.1
268.3 2S7.x 241.2 274.3
238,1
26!!.3 257.8 241.2 274.3
Assuming bial/elic ,\'NPs comprising X and r alleles. the reference genotypesfor the CD. D. R andOD inheritance models are XX. XX, XY-YYandXX-Yr respectively. Red highlight: Susceptibili~I' S:\'p, Green highlight. protective SNP
99
University of Ghana http://ugspace.ug.edu.gh

4.9.6. Tag SNPs on chromosome 9
Two SNPs located in the genomic regions encoding Interferon Omega 1 Pseudogene
19 (lFNWPI9) and Histo-Blood Group A Transferase (ABO) were assayed on
chromosome 9. The Histo-Blood Group A Transferase (ABO) SNP (rs8176722) was
associated with an increased risk ofCM (OR = 2.19, 95% CI = 1.08-4.45, P = 0.029,
Table 4.11) and endothelial damage (OR = 2.14, 95% CI = 1.24-3.71, P = 0.006, Table
4.) 1) in the dominant inheritance model. However, in the case-control comparison, it
was associated with clinical malaria in the codominant model (Table 4.12). The
IFNWPI9 SNP (rs73422262) was associated with reduced risk of clinical malaria in
the case-control comparison (Table 4.13).
100
University of Ghana http://ugspace.ug.edu.gh
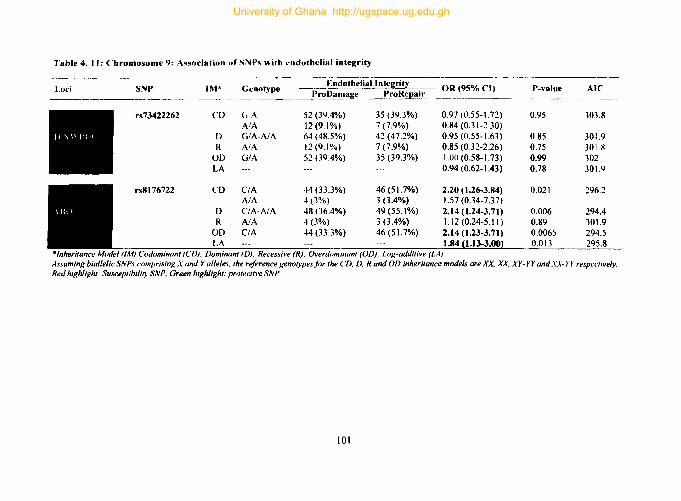
Table 4. J J: Chromosome 9: Assodatiun uf SNPs with l'ndothcJial integrity
Loci SNP IM* Genotype ~dothelial Integrity OR (95% ('I) P-value AI(' ProDan~ ProRepair -------.
rs73422262 CD (j A 52 (3Q.4%) 35 (39.3%) 0.97 (0.55-1.72) 0.95 303.8 AlA 12(9,1%) 7 (7.9%) 0.84 (0.31-2.30)
D G/A-A/A 64 (48.5%) 42 (47.2%) 0.95 (0.55-1.63) 0.85 301.9 R AlA 12(9.1%) 7 (7.9%) 0.85 (0.32-2.26) 0.75 301.X
00 G/A 52 (39.4%) 35 (39.3%) 1.00 (0.58-1.73) 0.99 302 LA 0.94 (0.62-1.43) 0.78 301.9
rs8J76722 CD CIA 44 (33.3%) 46 (51.7%) 2.20 (1.26-3.84) 0.021 296.2 AlA 4(J%) 3 (3.4%) 1.57 (0.34-7.37)
D CIA-AlA 48 (J6.4%) 49 (55.1%) 2.14 (1.24-3.71) 0.006 294.4 R AlA 4(3%) 3 (3.4%) 1.12 (0.24-5.11) 0.89 301.9
00 CIA 44 (33.3%) 46 (51.7%) 2.14 (1.23-3.71) 0.0065 294.5 LA 1.84 {1.13-3.00! 0.013 295.8
-Inheritance Model (1M) Codominunt (CD): Dominant (D); Recessive (R): Overdominunt (OD); Log-additive (LA) Assuming biallelicSNPs comprising X and Yalleles. the reference genotypes/or the CD. D. Rand OD inheritance models are xx. X¥. XY-YY andXX-l'l respel·/ively. Red highlight: Susceptibility SNP. Green highlight: protective SNP
101
University of Ghana http://ugspace.ug.edu.gh
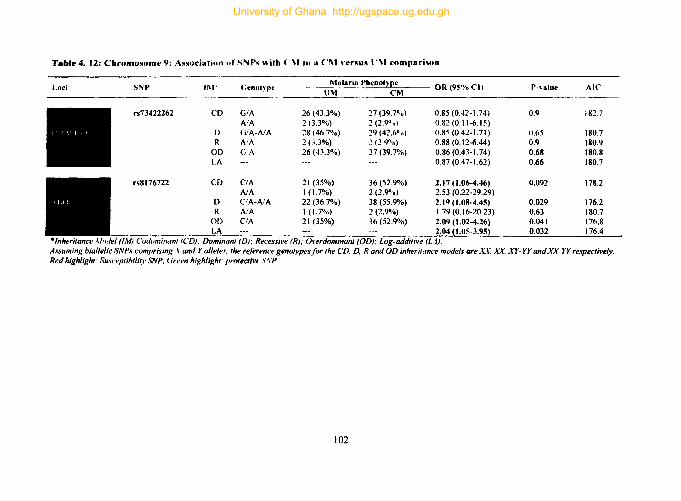
T.b'e 4. 12: Chromosome 9: AssociatioJl of SNPs with ('1\1 in a ('M \"Crsus t '1\1 comparison
Loci SNP 11\1' Genol~pc Malaria Phenot}pe
OR (95% CI) P-\alue AIC UM CM
~------.-.
rs73422262 CO G/A 26 (43.3%) 27 (39.70'0) 0.85 (0.42-1.74) 0.9 182.7 AlA 20.3%) 2 (2.9°0) 0.82 (0.11-6.15)
0 G/A-A/A 28 (46.70,.0) 29 (42.6°0) (j.85 (0.42-1.71) 0.65 180.7 R A'A 20.3%) :! (2.9";0) 0.88 (0.12-6.44) 0.9 180.9
00 GA 26 (43.3%) 27 (39.7%) 0.86 (0.43-1.74) 0.68 180.8 LA 0.87 (0.47-1.62) 0.66 180.7
rs8176722 CD CIA 21 (35%) 36(52.9%) 2.17 (1.06-4.46) 0.092 178.2 AlA 1(1.7%) 2 (2.9"/0) 2.53 (022-29.29)
0 CIA-AlA 22 (36.7%) 38 (55.9"/0) 2.19(1.08-4.45) 0.029 176.2 R AlA 1(1.7%) 2 (2.9"/0) 1.79 (0.16-20.23) 0.63 180.7
01) CIA 21 (35%) 36 (52.9"/0) 2.0'1 (1.02-4.26) 0.041 176.8 LA 2.04 (1.05-3.95\ 0.032 I
(1M) Codominalll (CD); Dominant (0); Recessive (R); Ol'erdominant (OD); Log-additive (LA). Assuming biallelicSNPs comprising X and Yalleles. the reference genotypes for the CD. D. R andOD inheritance modelll are AX. xr. XY-YY andXX-YY respectively. Red highlight: Susceptibility SNP. Green highlight. protectivl' s.1\'P
102
University of Ghana http://ugspace.ug.edu.gh

Table 4.13: Chromosome 9: Associatiun ofSNPs with malaria in a malaria \l'r'u~ U(, l'Itn11)arison
Loci SNP 1M· Genot~pe Malaria Phenotype __
OR (95% CI) P-vslue AIC ~-- Malaria
rs73422262 CD G!A 20 (29.9° 0) 67 (43.8°0) 1.41 (0.74-2.67) 0.0004 260.7 AlA 13(1'1.4%) 5 {3.3%) 0.16 (0.05-41.49)
D GIA-AIA 33 (49.2°0) 72 (47.1%) 0.92 (0.52-1.63) 0.76 274.4 R A'A 13 (1'I.4°u) 5(3.3%) U.I"(O.O~."I) 0.0004 259.8
00 G/A 20 (29.9%) 67 (43.8%) 1.83 (0<)'1·3.38) 0.049 270.6 LA 0.65 (0.42·1 .0 I) 0.053 270.7
rs8176722 CD CIA 19 (28,4%) 71 (46.4%) 2.08 (1.11-3.89) 0.021 268.8 A/A 4(6%) 3 (2%) 0.42 (0.09·1.95)
D CIA-AlA 23 (34.3%) 74 (48.4%) 1.79 (0.99-3.25) 0.052 270.7 R A'A 4(6%) 3(2%) 0.32 (0.07-1.45) 0.14 272.2
OD CIA 19 (28.4%) 71 (46.4%) 2.11) (1.18-4.06) 0.011 268 LA 1.39 (0.82-2.36) 0.22 272.9
·Inheritance Model (1M) Codominant (CD): Dominant (D): Recessive (R); Overdominant (OD); Log-additive (LA). Assuming biallelic SNPs comprisinK X and Yalleles. the reference genotypes for the CD. D. Rand OD inheritance models are XX. xr. XY -YY and XX- YY respect ive~)' Red highlighJ: Susceptibility SNP. <ireen highlight: protective SNP
103
University of Ghana http://ugspace.ug.edu.gh

4.9.7. Tag SNPs on chromosome 10
The SNPs assayed on chromosome 10 were in the loci encoding Long Intergenic Non
Protein Coding RNA 840 (LINCOO840). Case - case comparisons found variants of
rsl0899940 and rs13313099 to be associated with increased risk ofCM (Table 4.14)
and clinical malaria (Table 4.15). None of the SNPs was associated with endothelial
damage. Tables 4.14-15 summarises the association of chromosome 10 SNPs with
malaria phenotypes and endothelial damage.
104
University of Ghana http://ugspace.ug.edu.gh

Table 4.14: Chromosome 10: Association ofSNPs ni.h eM in a (,M \'erSliS liM comparison
Loc SNP IM* (;l'no~pe Malaria Pbenoo'~e
OR (95% ('I) P-\'alue AI(' 11M eM -----.
C'D ('IG 15 (25%) 31 (45.6%) 2.78 (1.29-~.98) 0.017 174.8 GIG 2 (3.3%) 5 (7.3%) 3.36 (0.61-18.44)
D ('rG-G/G 17 (28.3%) 36 (52.9%) 2.85 ( 1 36-5, tJ4) 0.0045 172.9 R GIG 2 (3.3%) 5 (7.3%) 2.30 (0.43-12.33) 0.31 179.9 OD ('IG 15(25%) 31 (45.6%) 2.51 (1.1 8-5 .. '4) 0.015 175 LA 2.35 ( 1.24-4.4") 0.006 173.4
rs 133 13099 CD TIC' 26 (43.3%) 38 (55.9%) 2.28 (1.02-5.09) 0.088 178.1 ('Ie 9 (15%) 14 (20.6%) 2.43 (0.85-6.92)
D pc-e/C' 35 (58.3%) 52 (76.5%) 2.32 (1.09-4.96) 0.028 176.1 R ell' 9 (15%) 14 (20.6%) 1.47 (0.59-3.69) 0.41 180.3 OD TIC' 26 (43.3%) 38 (55.9%) 1.66 (0.82-3.34) 0.16 178.9 LA 1.66 (0.99-2.78) 0.052 177.2
Model (1M) Codominant (CD); Duminant (0); Recessive (R); Overdominant (OD); Log-additn'e (LA). Assuming hialleli,· SYPs cumprisillfo:.r and Yalleles. the reference genotypes far the CD. D, Rand OD inheritance models are XX, X\' .. \T-YY and X\,-Yr I'l'spl'clively. Red highlight: Susceptihility SNP. Green highlight: protective SNP
105
University of Ghana http://ugspace.ug.edu.gh

T.b'e 4. J5: Chromosome 10: Association of SNP, "ith malaria in malaria \"S HC cnmparison ----"-- ~---"--- ----------
Loci SNP IM* Geno~'pe Malari~l Phenot)'pe
OR (95% ('I) P-\'alue AI(' He Malaria
rs]0899940 CD (. (j 15 (22.4%) 62 (40.5%) 2.44 (1.25-4.75) 0.024 269 GIG 3 (4.5%) 8 (5.2%) 1.57 (0.40-6.21)
D C/n-G/G 18 (26.9%) 70 (45.8%) 2.W ().B-4.~0) 0.0075 267.3 R G/(j 3 (4.5%) 8 (5.2%) I . I g (0.30-4.58) 0.81 274.4 OD C/G 15 (22.4%) 62 (40.5%) 2.3(. (1.22-·t56) 0.0079 267.4
LA 1.86 ( 1.08-3.] 9) 0.019 269
rs13313099 CD llC 24 (35.8%) 80 (52.3%) 2.18 (1.15-4.]2) 0.054 270.6 CIC II (16.4%) 24 (15.7%) 1.42 (0.61-3.30)
D T/C-C/C 35 (52.2%) 104 (68%) 1.94 (1.08-3.49) 0.027 269.6 R ('IC' II (16.4%) 24 (15.7%) 0.95 (0.43-2.07) 0.89 274.4 OD TIC 24 (35.8%) 80 (52.3%) 1.96 (1.09-3.55) 0.024 269.3 LA 1.37 (0.90-2.10) 0.14 272.3
(1M) Codominant (CD); Dominant (D); Recessive (R); Owrdominant (OD): Log-additive (LA). Assllming biallelicSNPs comprisinx X and Yalleles. the reference genotypesfor the CD. D. Rand OD inheritance models are XX, Xr. XY-YY andXX-Y> respectively. Red highlight. SlIsceptibility SNP. Green highlight; protective SNP
106
University of Ghana http://ugspace.ug.edu.gh

4.9.8. Tag SNPs on chromosome 16
This study assayed two SNPs located in the gene encoding Cadherin 5 (CDH5) on
chromosome 16. Although none of these SNPs were associated with endothelial
integrity, both SNPs were associated with protection from CM in a CM vs UM
comparison. Tables 4.16 summarises the association of chromosome 16 SNPs with CM.
Unlike other SNPs assayed in this study, neither rsl077318 nor rs2304527 was
associated with malaria in the case-control comparison.
107
University of Ghana http://ugspace.ug.edu.gh

Table 4. 16: Chromosome 16: Association ofSNPs with endothelial integrit)
Loci SNP 1'\1* Genotype Malaria Pheno~'~e OR (95% ('I) P-\'Blue AI(' eM l1M
--'-
~CD A/C 48 (70.6%) 29 (48.3°/0) 0.30 (0.12-0.76) 0.024 175.5 ClC II (\6.2%) 13 (21.7%) 0.59 (0.19-1.84)
D AlC-ClC 59 (S6.S%) 42 (70%) 0.36 (0.1 ~-O.87) 0.02 175.5
R ClC 11 (16.2%) 13 (21.7%) 1.43 (0.59-3.49) 0.43 IS0.3 OD A/C 48 (70.6%) 29 (4S.3%) 0.39 (0.19-0.81) 0.01 174.3 LA 0.75 (0.43-1.31) 0.31 179.9
~(,D T/G 46 (67.7%) 24 (40%) 0.32 (0.J~-0.71) 0.0069 173 GIG 6 (8.8%) 10 (16.7%) 1.03 (0.31-3.37)
D T/G-G/G 52 (76.5%) 34 (56.7%) 0.40 (0.19-0.86) 0.017 175.2 R Glti 6(8.8%) 10 (16.7%) 2.07 (0.70-6.08) O.IS 179.1 OD T/G 46 (67.7%) 24 (40%) 0_'2 (0.15-0.66) 0.0016 171 LA 0.75 (0.43-1.29) 0.29 179.8
(1M) Codominanl (CD); Dommall{ (D); Recessive (R); Overdominant (OD); Log-additive (LA), Assuming biaflelic SNPs comprisinK X and r alleles. {he reference genotypes for the CD. D, R and OD inheritance models are X%', AX AT-IT and X\,rr respectively. Red highlight: Susceptibility SNP. Green highlight: protective SNP
108
University of Ghana http://ugspace.ug.edu.gh

4.9.9. Tag SNPs on chromosome 20
The loci encoding Matrix Metallopeptidase 9 (MMP9) harboured 3 of the SNPs
genotyped in this study. A variant of rs2236416 was associated with a reduced risk from
endothelial damage (OR = 0.22, 95% CI = 0.08-0.59, p = 0.002, Table 4.17) and
rs3918256 was associated with increased risk of endothelial damage (OR = 3.93, 95%
CI = 1.63-9.50, P = 0.0015, Table 4.17). Association of other MMP9 SNPs with clinical
malaria is summarised in Table 4.17.
109
University of Ghana http://ugspace.ug.edu.gh

Table". 17: Chromosome 20: Asso('iation ofS~I', with t'lIllothdial integri~ ~ .. -- -------~"-.-
Loci SNI' IM* Genotype Endotheliallntegritv . OR (95% C1) ProDama~ ProRepalr ____ . __
P-\'alue AI('
rs2236·H6 CD AlG 15 (22.4%) 54 (35.3%) 1.57 (0.79-3.10) 0,0036 265,2 GIG 12(17.9%) 7(4.6%) 0.25 (0.09-0.69)
D AlG-G/G 27 (40.3%) 61 (39.9%) 0.98 (0.55-1.76) 0.95 274.4 R GIG 12 (17.9%) 7(4.6%) 0.22 (0.08-0.59) 0'()o2 264.9
OD A/G 15 (22.4%) 54 (35.3%) 1.89 (0.97-3.67) 0.053 270.7 LA 0.73 (0.47-1.12) 0.15 272.4
rs3918256 CD G/A 45 (34.1%) 25 (28.1%) 0.95 (0.52-1. 75) 0.0063 293.8 AlA 8(6.1%) 18 (20.2%) 3.86 (1.56-9.59)
D G/A-AiA 53 (40.1%) 43 (48.3%) 1.39 (0.81-2.40) 0.23 300.5 R AlA 8(6.1%) 18 (20.2%) 3.93 (1.63-9.50) 0.0015 291.8
OD GIA 45 (34.1%) 25 (28.l%) 0.76 (0.42-1.36) 0.35 301.1 LA 1.58 (1.07-2.34) 0.02 296.5
*Inheritance Model (lAO Codominant (CD); Dominant (D); Recessive (R); Overdominant (OD); Log-additive (LA). Assuming biallelic SNPs comprising X and Yalleles, the reference genotypes for the CD, D, Rand OD inheritance models are xx, xx, XY- YY and XX- YY respective~l'_
Red highlight: Susceptibility SSP, (Jreen highlight: protective SNP
110
University of Ghana http://ugspace.ug.edu.gh
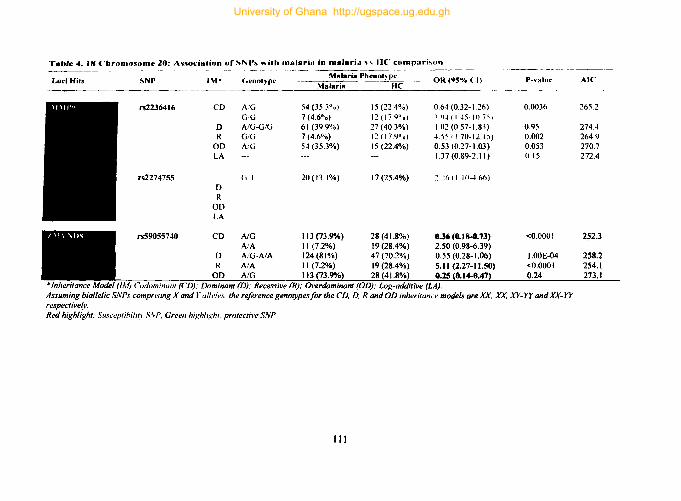
Tab'e". J8 Chromosoml' 20: Association of SN "s "ilh malaria in malaria" lie comfl~tri~on - .. --.~- -
Malaria Phmot~ P" Loci Hils SNf' 1M· Grnol)'pc OR (95% <I) P-"Hlu" Ale
Malaria HC
rs2236416 CO A/G 54 (35,3 0 0) 15 (22.4%) 0,64 (0.32- 1.26) 0,0036 265.2 GG 7 (4,6°'0) 12 (17,1)° 0 ) -; 'I,j (I 45-10 7~)
0 A/G-G/G 61 (39,9%) 27 (40,3%) ),02 (0,57- 1,83) 0,95 274.4 R GIG 7 (4,60",) I:? ()7,1)°0) 4,55 (1,70-12.15) 0.002 2649
00 A/G 54 (35.3%) 15 (22.4%) 0.53 (0,27-1.03) 0.053 270.7 LA 1.37 (0.89-2.11) 0.15 272.4
rs2274755 GI 20(13.1%) 17 (25.4%) ~:::h (I 10-4,66) 0 R
00 LA
rs59055740 CD NG 113 (73.9"AI) 28(41.8%) 0.36 (0. 1 8-0.73) <0.0001 252.3 AlA II (7.2%) 19(28.4%) 2.50 (0.98-6.39)
0 A!G-A/A 124(81%) 47 (70.2%) 0.55 (0.28-1.06) I.OOE-04 258.2 R AlA II (7.2%) 19 (28.4%) 5.11 (2.27-11.50) <0.0001 254.1
OD NG 113 p3.9"AI) 28 ~41.8%~ US {O.I4-0.471 0.24 273.1 (1M) lodominuflI (CD): Dominant (D): Recessive (R); Overdominant (aD); Log-additive (LA).
Assuming biaJlelic SNPs comprising X and r "fides, the reference genotypesfor the CD, D. R and aD inheritance models are AX, xx. xy-yy andXX-YY respective(v. Red highlight: Sliscepfibilifr S,.,P, Green highlight protective SNP
III
University of Ghana http://ugspace.ug.edu.gh

4.10 Global association plots
To have a global picture of the association of SNPs with endothelial integrity and
malaria phenotypes, a pseudo-GWAS approach was used to construct Manhattan plots
from LocusZoom's regional association plots (Pruim et al., 2010). The significance
threshold was adjusted to p < 0.002 to correct for multiple testing in this analysis.
A CDH5 SNP (rs2304527) emerged as the only SNP associated with eM in a UM vs
CM comparison under this conservative significance threshold. Under the over
dominant inheritance model assumption, children with the heterozygote riG genotype
were protected from CM relative to their colleagues with the TT -GG genotype (OR =
0.32,95% CI = (0.15-0.66), p=O.OOJ6. Fig 4.7). Several SNPs were associated with
malaria in the comparison between malaria and healthy controls (Fig 4.8) but the most
striking of these SNPs were Ts3917419, rs684951, rs2070744 and rs59055740 which
were associated with malaria at significance threshold comparable to that employed in
GWAS studies (i.e. p < 0.5 x 10-8) Fig 4.8. The MMP9 SNP (rs3918256) was the only
SNP associated with endothelial integrity at this studies significance threshold. Relative
to the reference genotype (GG), individuals with the AA genotype ofrs3918256 were
almost 4 times more likely to be classified as ProDamage under the recessive
inheritance model (OR = 3.93, 95% CI =1.63 - 9.50,p=O.OOJ5. Fig 4.9).
112
University of Ghana http://ugspace.ug.edu.gh

Association of tac SNPs with eM relative to UM
GWAS p-value threshold: < 5XIO·a - .. -.. - ---------------------------------------
~ ~ ~ i i~~~7
," ... n·fll.ISoO"'~
__ ~~~~~!..:~~~~~_~~:~~~!~ __ ::_..:_~~~~~__ ~ . -~~~---1 -------.. ------------~--------------=~=.:.!:::.----• • It. p< 0.05
· i • • ..
t i i J I • I .! • • ... • ... .."..... L....,......~ -..... Genehltj·
ci!1 J r MJ [5h;4- Jr~~r6j [c!:il M J ~1!~1 [Ch!t6...- ChrZO~"
Lit ... I i ( Chrom~som~ nu';'be~ In:rrn annotation key
~nt •
Dominant • Log-addiIi\Ie • <Nerdominant .. Recessive •
\ \t.nha"_n plol ,ho"ing ~,p, ."orialrd .. ilh C\I 81 differenl signifiesnrr Ihre.hold. \'.IIow, grHn and red lincs denole p.ulue Ihruhold at <0.05, < 0.002 and < 5.10""respeetlvdy
Figure ... 7: Association ofSNPs with eM in a eM vs UM comparison
113
University of Ghana http://ugspace.ug.edu.gh

~ c!. '0
~ 4
Association of tag SNPs with malaria relative to healthy controls
rs3917419 • r.;684951 • .r.;2070744
r.;S90SS740
c.WA\pnletJveeheld: s .1~
------------.---------------
• ___ ~-----------------------------!--------------------_______________________ ~~.;.~!!.a::_
• •••• • I • - • • • • J -, • • Ii Ie I
I • .
• ..
u..... --I.-\ Chromosome number I" • i. I •
___ 1.._ • • •
- ----~.-~
I
.... ~. Ch~_~.
amoIaIJon My
A Manhattan plot showing SNPs associated with malaria at different significance threshold. Yelhm. green and red lines denote p-value threshold at < 0.05, < 0.002 and < 5.10-8 respectively.
Figure 4. 8: Association of SNPs with malaria in a malaria vs HC comparison
114
University of Ghana http://ugspace.ug.edu.gh

~ ~
1'.)
1 I 241
Association of tag SNPs with endothelial damap
393 (1.63·9 SO)
.................... "'., .. -----------------_. -
rsJ918256 OR = 3.93, 95% CI ~ (1."9.501. p ... HI5
---------l
---- ~ -. ----------------------- ---------------------------------------9--------=--:::::.':.::.----t • • •
• • . •
I • a a • i
! I • J , i I •
U-...........i.1{ Chromosome number J-1L~
. ..... I - Genehll.
J annoIatlOfl key
Codonwlant • Dominant • ~ • • RecessIVe •
A Manhattan plot showing SNPs associated with endothelial integrity at different significance threshold. Yellow, green and red lines denote p-value threshold at < 0.05, < 0.002 and < 5.10-8 respectivel~.
Figure 4. 9: Association of SNPs with ProDamage in a ProDamage vs ProRepair comparison
115
University of Ghana http://ugspace.ug.edu.gh

4.11 In silko analysis
The functional significance of trait- or disease-associated SNPs are sometimes not
decipherable on their genetic attributes alone. To better appreciate the plausible roles
of disease-associated SNPs identified in this study, a number of in silico analysis were
conducted to: (a) find proxy SNPs for trait- and disease-SNPs identified in this study;
(b) characterised the epigenetic context of these SNPs (both tag and proxy SNPs); (c)
predict SNPs that may enhance or inhibit the binding affmity of transcription factors;
and (d) to identify SNPs that disrupts microRNA target sites.
4.11.1 Linkage disequilibrium analysis.
In line with the concept of tag or sentinel SNPs, this study did not only focus on SNPs
associated with malaria and endothelial integrity but also on the SNPs in linkage
disequilibrium with ''test'' SNPs. Using SNPs with significant associations as tag or
sentinel SNPs, linkage disequilibrium (LO) plots were generated using SNiPA - a tool
for functional annotations and linkage disequilibrium analysis for bi-allelic genomic
variants (Arnold et ai., 2015). LO plots were generated using the African population
database in Ensembl87 - grch37/1kgpp3vS with an LO threshold of 0.8. The list of
SNPs in linkage disequilibrium was incorporated in LO plots to create composite
figures (Figs 4.10 - 19).
116
University of Ghana http://ugspace.ug.edu.gh
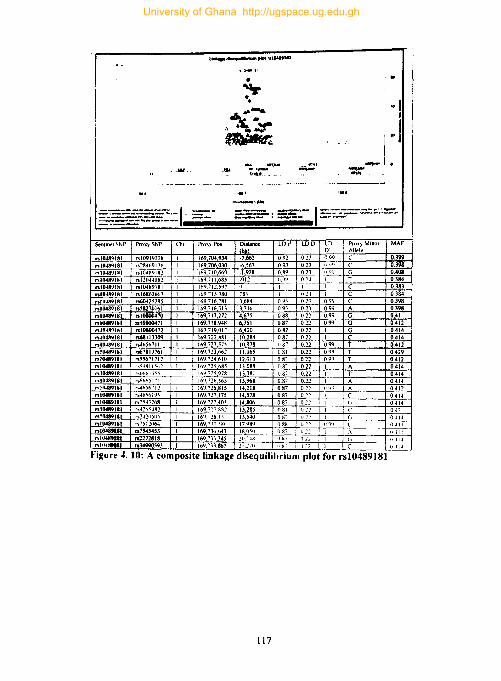
. -:;. . 11>-'"
~x~ .. ~ --.-----.--- I -_ .. _._-_ . ..-.- .... - ------""---- --.. _- ...... _ .. _--
Sem",el ~NP Proxy SNP Chi Pm') Po>
.. 10489181 "1091'(!21i 169.704 934
.. 10489181 f-s21Wt'C) I 16 169.706.030 n104K91~1 ,,10489182 169.710 669 ,,1(1489181 nl2044082 Ih9.71168~
"111-189181 ,,104891SI 169.712,5')7 nl0489181 n16862663 169.'1.1'80 .. 111489181 n6042~295 169.716.281
"10489181 ro82701bl 169.716.313
"10489181 ,,10800470 Ib9 717 272
"10411'1181 ,,10800471 169 718.948
,,1"4""181 ,,10800472 169719.01 7
,,111489181 1168113309 1697:!2.881
"10489181 no-I6S671 I 169723.575
"111-189181 007819761 169723662 .. 10489181 ,,~Sb71712 Ib9 7 24.610
"10489181 f .. ';IJX II ~I)~ Ih9,7B.h85 ,,10489181 r·.tlhfll'l~Ci Ih9715.978 ,,10489181 r~6()MI"1 IlN,72l1,5b5
1'11l41l'i181 r<o.-Jh~t"·J2 169,716815
"10489181 r~(''''bI9~ 169727175
"10489181 r .. "54.~248 Ib9 '27 403 nl0489111 r~4265482 16')."2'" kR2 nl0489181 r"lo~",:?n50"i Il,fj.-28,1'"' nt_181 r~ "'S I ~O64 169 i,(l "/0((,
"10489111 n7S45453 169 73u 1>47 nl0489.1 112272818 169.'" 345 ,,1114"'111 rs34990593 I 16').733.1167
~. _I}lMl _'I~ CI-'!!1 ___ ._ .
~s;- lOr'
·7663 092
-6 ~67 093 -1.928 089 .912 099
I 78.3
3.b84 09.l
3716 093 4675 088
6351 087 6420 087
10284 087 10978 I) 8"
11.065 081 12.013 () 8' IJOS8 08'
1>381 087
13968 0.87 14218 0.87 14578 0.87 14806 087 15285 OHI 15540 U8: J7 989 088 IR,O:liO () 87 20 -lH 'j ~7
21.270 ',s7
I-
LDD LD Pro,> Minor MAF D Allele
023 099 C 0.399 013 (' 0.398 0.13 G 0.408 o 2 ~ T 0.384 I C 0.383 u ~,l C 0384 02.1 O.9~ C 0398 o:n 099 A 0398 022 0.99 G 0.41
022 099 G 0.412 021 1 G OA14 022 C 0.414 021 099 T 0.412 0.22 099 T 0.429 022 099 T 0.412 022 I A 0.414 0.22 T 0.414 021 0414 () 2~ A 0.412 0.22 C 0414 0.:!2 0414 f) .:' ~ 043 IJ"; 0414 I) ~~ I) ()I) 0411 Il ~~ (III)
'122 \1114 1)!2 I (' (I II ~
Figure 4. 10: A composite linkage disequilihrium plot for rsl0489J81
117
University of Ghana http://ugspace.ug.edu.gh

Sentinel rs2070744 rs2070744
i i
8£Tlrl o_~
.-------o.iM"~p N[M'7'18 ACe1 ~ciNAP!. ~1'OA---
.505
lInup dl ........ rlum plot ~2070744
""""0,'4..)
*
• , '".1:1. L A • <..
;\t~l' "'i!!'"
"'!P ,"!'" H!5!! ~ ,¥t Q!,.f~
..J!!!!!!... 1!:.~ Mlf!'11 4Q!!!I Rt"lIt~'I' .....!!!L ~&. SMA.!'CCh_
.... t~ •. 'o W" 't!!!.. .....~I.'
.507 .1108
Chromoeome 7 iMb 1
"' ___ .n~ .......... ......, ....
40 f
i
____ .~_ .... I\,-'---...._. n.,y_ ,_ ~ .-.".-----......... '- 1 --- ........ -~ ........ ..... ~_(".v ........... .......,~ __ _
1
_____ ... ·_,,_ .... "'-~_~I.Jl.tf ........ ~ ~ .... ...,....., ..... ---... ... .... ",-""""<>,,,-...,~ ~ ___ cA .... 5NP'n..pIof""'"*"'oINdo __
~"'"~ __ tIO'I
Proxy ('hr. Proxy Pos. Distance (bp) LD r LD D LD D' Proxy Allele B rs1800779 7 150.689,943 -136 0.98 0.12 0.99 G rs2070744 7 150,690,079 0 1 I I C
Figure 4. 11: A composite linkage disequilibrium plot for rs2070744
118
MAF 0.139 0.138
University of Ghana http://ugspace.ug.edu.gh

~
J ~ 5
I
• ...... 8NP ( noe .. LD L LOI2'02
• LOr' , •• • lO" '01
TM~718 ~_
TM~lleA
150.5
Unkage disequilibrium plot for rs~918211
~118211
•
• ~ . q,w.,{:j \0 0 c 0 ~)., .\.~
COK5
-.P FASTl<
A8CII8 -na.e. I\IOS3 -- 9Lcw/.
--ATGIB ~,.12
<h CO
~ RPiI,.:,"""o • ~
150.7
at_e7(Mb)
CHI'F2
MiRe7' A8CFI &.w.cm
,:!!!!o --;u. .. ~ ... GBX1 ~L Rf'S.1~2
150.9
60
40
~ , 20 J
unuc·d,wq"",_ .... "_ .... _01_ Ii --. ~"wntlnll .... "t.l'Idtts~..,.,..ntsn."a.lt'\ _ tI'WIIICripI showt.llwocorrttt.noncoeffla.nrlrl).the .. -...,tI'ICM'I "'*'town ..... __ oI_h9<.""' .... _oI__ --------,nckatftlt, ....... 'ONf ....... tOn
_011 ..... ,~ ., ..-.noguIaIory ...... 11 ____ ..... .". ....... 71'........,.
_n~~';;:':;:-: ::==- I =:-=.,...-,,-_ ........ , ......
Figure 4. 12: A composite linkage disequilibrium plot for rs3918211
119
University of Ghana http://ugspace.ug.edu.gh

] I
API -.ota. ~ • ~C'Ma
Link .... dlsequtllbrium plot for n:J917419
.. • 6.Ml
6. • 6. 6....,;., ;-.Ce' ~ 4. ·&~'ir.l.f.~'
t.7
III:TTI. ••
RPI "'''''$ .. <: ..... , •
etvorno.om. 11Mb)
~"""'''"-tt. . L
~.!.':.s_ -
I
I~,'h,"~
I .----=:-:.. ..J
i ,f
!
==:7'..:==--:::.=.:';:;~~-:::. no. ="ec1 :.c'.If_..;::.;"'::'---: =---=... =.~OOftI ~--------u ............. ~_ ......... I ............. oI--""... _____ ~ ...... ........,. .. .- , ....... ___ ........ -...,.. ...... ,,,........,, ..
::::::--..:::::; of..m", n. ............ .,. _ ....
Sentinel Proxy Chr. Proxy Pos. Distance (bp) LD r' LD 0 LD D' Proxy Allele B MAF rs3917419 rs2076059 1 169.698.921 -898 0.99 0.19 I T 0.247 rs3917419 rs3917419 169.699.819 0 1 1 1 A 0249 rs3917419 rs753 1675 169,711,306 11,487 0.89 0.18 0.97 A 026 rs3917419 rs7543618 169.711.3~6 11,507 0.89 0.18 0.97 C 0261 rs3917419 r..12-l10806 169.718.236 18,417 0.93 0.18 0.97 A 0.248 rs3917419 rsl~n8631 169.725,635 25,816 0.88 0.18 0.96 A 0.256 rs3917419 rs7526645 169.728.916 29,097 0.88 0.18 0.96 T 0.256 rs3917419 rs7549412 169,729,250 29,431 0.88 0.18 0.96 G 0256 rs3917419 rsl2063022 169,731,865 32,046 0.92 0.18 0.97 G 0.247
Figure 4. 13: A composite linkage disequilibrium plot for rs3917419
120
University of Ghana http://ugspace.ug.edu.gh

Sentinel
rs59055740 rs59055740 rs59055740 rs59055740 rs59055740 rs59055740 B59055740 rs59055740 rs5<)055740 rs5'Xl55740
Un,,- .u..quIUbr,uR"I plot for ... 59055740
. '
I t .1
""~'In ~
. ,-...,-_ ...... __ .. - I -_, ... _. __ ...." •• ~--..n n....... ____ =_=~:::J: ... <+:.:.~ _ ;._.~~ .....
Pro~) ('hr. Proxy Pos.
rs6018446 20 46.007.753 rs6018447 20 46.011.511 ,."t.09-!o81 20 46.013,864 rs6125U05 20 46.014.346 rs59055740 20 46.014.727 ,,606629R 20 46.016.583 rs6066303 20 46.019.0·B rs6094689 20 40.033.012 rs1535172 20 46.033.609 rshl25012 20 4h.034.35I
.. . . • ' .
• ..... \.,
~.2'0(.., •
40 ('
'<:;. · i --- 1IIPs,..,..., •• ... ~p
=:-.. ~~: =-~ ... _ ......... _r ..... ___ • .---.. ............... _ ---- --- 1------_· __ · Oistanee LD LO LO
PrOlty Allele B !bl!l r' D 0' -6.974 0.98 0.25 0.99 C -3.216 0.95 0.24 0.98 G -863 0.98 0.25 0.99 T -381 0.96 0.24 0.99 G 0 1 I I G 1.856 0.88 0.23 I G 4,316 0,85 0.23 0.98 T 18.285 0.83 0.23 0.97 C 18.882 0.83 0.23 0.98 C 19.624 0,82 0.23 0.97 G
Figure 4.14: A composite linkage disequilibrium plot for rs59055740
121
MAF
0.492 0.498 0.492 0.488 0.494 0.461 0.463 0.459 0.457 0.457
University of Ghana http://ugspace.ug.edu.gh

~
i ~
J .c t 3
__ SNP
noftnLD '- LO?_-02 • LOr' ... 05
I ,- lOr' -08
Unkage disequilibrium plot for ~684951
r5684'.:151
...
... ,
~.
, '. ,
TREK. TREMl TREMLSP - 1NIe·e43P
TAUA2 RP1.i2.aoe· NFYA ... TREMl 2 RPI ~ .•
... --- - AOCYIOPI • TREML3P 'AP1-22IKmS NCII2 RP1·I."". __ -,,=OA~RO,,-,I_ .. _-- TR~"'" -- RNA5!iP207 - RP1~1 - API '4;8'"'' •
41.1 41_3
CIvorno8ome II 1Mb)
60
40 I I>
! j
~ I FOXr4-AIII
LlQlaN ~
41.5
l'n .... f'dl1eQuthbrtumplot'!howlht .. ~Cl-"""~.;~)<",.."or. I ---bflwff!l.SPn1inel.."...endlb ... .:h.'XI,'II' ... , .. '\t~~kW".· •• ,· _ &I~ipI
Ihow5n.CIIIfNIMtoncoe'fflo""('II.thfo'd"'~_ ......,.lIIIact c:~ .... _ of HCh SHtt nw Put "f'"bH at .. ~CII ..... ~·"n' ~1tItuncaan.a1flnCllMton
.....,. ... .:ton ...... 0 ~enec:tl r~_raV ........ Ion).fwIdIGnII5HP~lOns.,. __ ""~ ......... rogulolory_' 1-----lho . .-7fl ..... J!r5.
dl_rogoAokIry_ 0 ...... 001_..... J _ .. ·_r
Figure 4.15: A composite linkage disequilibrium plot for rs684951
122
University of Ghana http://ugspace.ug.edu.gh
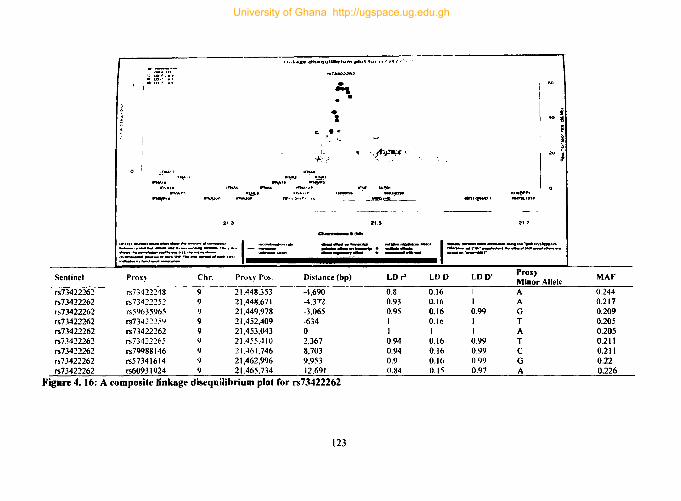
Sentinel
rs73422262 rs73422262 rs73422262 rs73422262 rs73422262 rs73422262 rs73422262 rs73422262 rs73422262
Link .... dt •• qulllbrium plot for r,,7"141/]' •
~:- LD~ ~O~
• &.O~ .' • lOr' -0.
....... ""-NAt.
.....,....
....... ...... ,3· -' I"~··
.~ ...... ----... - ... -- 1 .-._-~:;-;-:-,,~~~..:::.. -= .. -Prox~ Chr. Proxy Po~.
rs73422248 9 21,448.353 rs73422252 9 21,448.671 rs59635965 9 21,449,978 rs73422254 9 21,452,409 rs73422262 9 21,453.043 rs73422.:!65 9 21.455.410 rs79988 146 9 21.461,746 rs57341614 9 21,462,996 rs6093 1924 9 21.465,734
~ \ ,. ~
~-:> ,.r,ti'~~,:
....... . "-. ~P2
"!'f .......
---_~~I_HO __ _
2'.5
~.jMb,
1ItI'~,., ~tP
2.7
i 'i! i ~ i
- . .... -----. ,......,. .......... .a.c. 1...., .......... --.. ...... · .... ,,, ......... · ........... __ "--. ........... ....-...-_r~ ........ 101'11 ~INI' ___ _ __ ._ ~~~ . -........cI...... tauM ... • ....... r
Distance (bp) LD r' LDD LDO' Proxy Minor Allele
-4,690 0.8 0.16 I A -4.372 0.93 0.16 1 A -3,065 0.95 0.16 0.99 G -634 1 0.16 1 T 0 1 1 1 A 2,367 0.94 0.16 0.99 T 8,703 0.94 0.16 0.99 C 9.953 0.9 0.16 0.99 G 12,691 0.84 0.15 0.97 A
Figure 4. 16: A composite linkage disequilibrium plot for rs73422262
123
MAF
0.244 0.217 0.209 0.205 0.205 0.211 0.211 0.22 0.226
University of Ghana http://ugspace.ug.edu.gh

i ~ ~
i " t 5
o I
• ......... SNI' ".. .. lO
C lO,J "04'
• lO,J 'O~ • loi
linkage disequilibrium plot for rs3818256
f'. J9182S6
~ J
A
• A
A
A A·
t:.&tf, l:.~ ~~rJ . ~ 0~ v .• ~
lSWlM.
UB~ ~. ~ ThINCII NEURL2
ONTT\P. -- ACCTa SPAT42S
WfDC3 -- 1Aotaz .. SRP ·ClSA
_!A~ ~,' RP3-~ ...
44.4
~!g
Pelf! FTlP, SlC.2A5 - ZNf33S . RPI11!!L~
44.11
Chrom_ 20 (Mb)
~ ~
RI'L!Jf'2
60
40 I ! ~
m i RP5-fI!'HU
alHI2
44.8
""..,.d .... _ ..... _ .... _01_ 1 ,,,,,om_oon"', bft...en.MftfIrw4 ... ,..... .... ttt."',.rouncfIl1C\lQftanb- .......... '" -......,.,.
"""" ... COI'TWIetIaft~{,.l}. ...... I'1ohawI unknownelt8d -........_ .. _"' ......... ..- .. --"'IftIts "-ct ........... 1Oft
<Inc',Hee'"""""",,, ...-~- '1 _____ . .... ....... 71JtupN" puIIIIve_ ... ,_ 0 ....... _ . __ .. r .... ~ ___ _ ..... ..-., ... ect 0 ___ c~ .. ·_
Figure 4. ) 7: Linkage disequilibrium for rs3818256
124
University of Ghana http://ugspace.ug.edu.gh
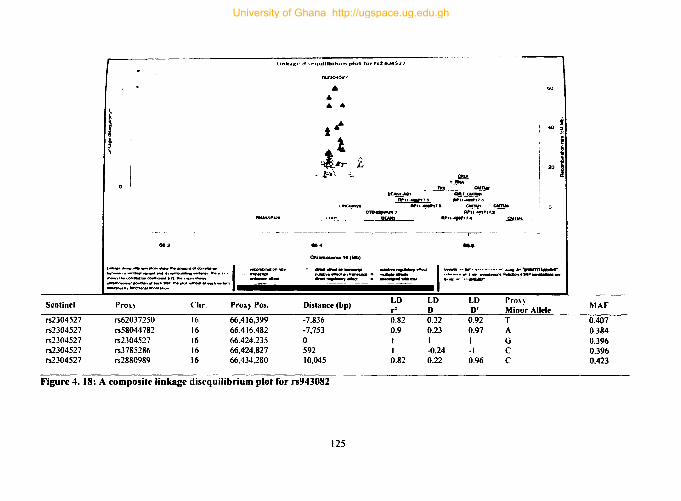
Sentinel
rs2304527 rs2304527 rs2304527 rs2304527 rs2304527
llnk .. gf> dl·.rqt .. mbrtum plot for rs2Ja.Sl1
i I
e412
,_ ..... -_._ ... _"'- I -_ .... ~r- .......... ___ eftIIII .. '---. __ .. n..."l., _.,.,..,.. ~1 .. d"..t.aon~I,lll................. ~.n.d
~ru:==n..P'cR.,.....-"vatt.m ~
Pro~) Chr. Proxy Pos.
rs62037250 16 66.416,399 rs58044782 16 66,416.482 rs2304527 16 66,424,235 rs3785286 16 66,424,827 rs2880989 16 66,434.280
• • • • t··
1 ~~ t
- .t-~\ .• ".
11114
ChromOSOl'lle 1. CUb)
_. Y-r!iA
CUlW 8UH1., ... ---QG.f a,rn.h-
j ~ ! I
20 1 ~u =:'':;''7.8 .. '1-;6 CI!!!M ! 0
CTD.~3 APll~P1'.t
~ PUt.'....,.,7.. ~_
_.11
,....,.,..n.d .... ...".,...,.,........ · ........... e_' .... ~~SIItP......-.. ... cINd............... ..........,.....oryef1.c1 I ~ .. <IfIn!I' ..... ~ •. ~.oot •• ...-!'~"""'-....n/1 ......... ... ,..,..".a.cr 0 ............... »-...... -.... .....,.
Distance (bp) LO LO LO Pn)\~ rl 0 0' Minor Allele
-7.836 0.82 0.22 0.92 T -7,753 0.9 0.23 0.97 A 0 I I I G 592 I -0.24 -I C 10,045 0.82 0.22 0.96 C
Figure 4.18: A composite linkage disequilibrium plot for rs943082
125
MAF
0.407 0.384 0.396 0.396 0.423
University of Ghana http://ugspace.ug.edu.gh

Sentinel
rs3918256 rs3918256 rs3918256 rs3918256 rs3918256 rs3918256 rs3918256
i t
lInkaae d.seqvtUbrtum plot for "94~2
... ,.. ... . ,- •
,
~~~
-!!!!:!L_~~ ~..,.,...~'.,....'. .. _. API ".I=I;a.I~. ~.r!'
"P'l~"
......... ~---- ... .,. .............. ..... _1·41·1'-· .. ••• 1_- - ... - ::::'7':":::'::--: ..... --=~
..• _SNP'-_ .. ~.
Proxy Chr. Proxy Pos. Distance LD LD ~b~) ... D
rs1169680~ 20 44,628.380 -12.,579 0.8 0.13 rs3933239 20 44,628.668 -12,291 0.8 0.13 rsl1698788 20 44,632,032 -8,927 0.85 0.13 rs3918253 20 44,639,511 -1,448 I 0.13 rs39 I 8256 20 44,640.959 0 I 1 rs20544 20 H645,010 4,051 0.95 0.\3 rs35567443 20 44.648,361 7.402 0.92 0.13
Figure 4. 19: A composite linkage disequilibrium for rs2304527
126
I
i ~ta. ~.~-----------i
1------· .. ·-·,-· ........... r __ ............ '·---__ ---LD Proxy Minor Allele MAF D' 0.98 T 0.\85 0.98 C 0.185 I T 0.181 I T 0.159 I A 0.159 0.98 T 0.157 0.97 T 0.157
University of Ghana http://ugspace.ug.edu.gh

4.11.2. Epigenetic contexts of SNPs
The epigenetic context of SNPs significantly associated with endothelial damage.
malaria phenotype and angiogenic factors were detennined using ChroMoS which is a
web tool that combines genetic and epigenetic data to classify and prioritise SNPs.
Using data based on Human Umbilical Vein Endothelial Cells (HUVEC), ChroMoS
found SNPs and their proxies to occur in 6 different human chromatin states. Figure
4.20 shows the specific chromatin state each SNP belonged to and their proportions.
As expected, the majority of the SNPs (50.7%) occurred in the low signal
heterochromatin. Just over 5% of SNPs occurred in strong enhancers and
transcriptional elongation chromatin state. Only 2.8% of SNPs occurred in insulators.
Sixteen per cent of SNPs occurred in weak transcribe chromatin states.
4.11.3. Potential effects ofSNPs on Transcription Factors (TF)
Based on the fact that noncoding SNPs in specific chromatin states affect various pre
and post-transcriptional mechanisms including the activity transcriptional factors, this
study assessed the potential effect ofSNPs on transcription factors (TF). The study used
the sTRAP function in ChroMoS to assess the possible effects of these SNPs on the
binding affinity of transcription factors. Thus, 71 SNPs comprising nine trait- and
malaria-associated SNPs and 62 proxies were inputted into ChroMoS. Using all 9-cell
types available in ChroMoS, 22 SNPs that lay in specific chromatin state in at least 4
cell types were selected for sTRAP analysis (Figure 4.21). At a threshold of 1 (which
returns only significant results), 16 SNPs that potentially influence 49 different
transcription factors were identified (Table 4.19). Table 4.19 depicts transfac matrixes
grouped by SNPs: -transfac matrices with a lowered binding affinity have a negative
ratio and those with increased binding have positive ratios (Table 4.19). Furthennore,
127
University of Ghana http://ugspace.ug.edu.gh

and perhaps more importantly. the direct effects ofSNPs associated with either malaria
or endothelial integrity were evaluated for their impact on transcription factors using
atSNP. atSNP is a web tool for predicting which TF binding sites created on eliminated
by SNPs. Tables x-x reports the top 5 TFs impacted by the 6 SNPs.
128
University of Ghana http://ugspace.ug.edu.gh

Polycomb .. pressed _
18·3%
Hetrochrom_low Signal __ _
50·7%
Figure 4. 20: SNPs and their Chromatin States
Weak Transcrrbed
16·9% Trans.cnptlonal Elongation
5·6% Strong Enhancer
5·6%
--'---2~8%
129
rs4G5619S rs4656711 rs465671a
rs55671712 rs57341614 rs58270161 rs59635965 r559811562 rs60425295 rs6661955 r56665171
rs67819761 rs6S113309 rs73422259 r573422262 r57512064 r57526645 rs7543268 rs7545453 .,.7549412
.,.79988146
University of Ghana http://ugspace.ug.edu.gh
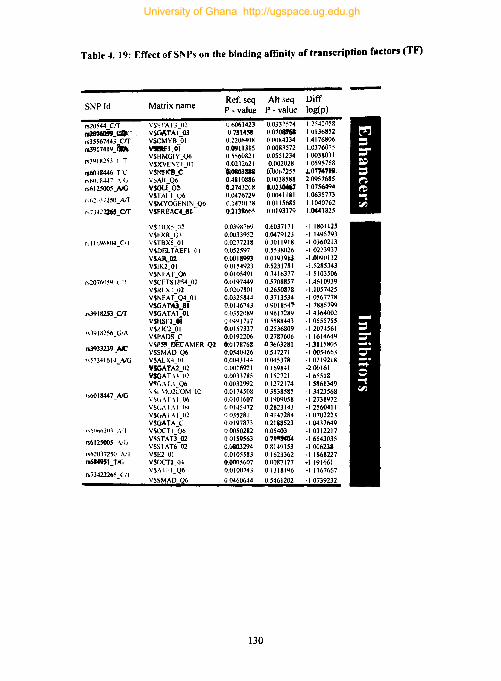
Table 4. 19: Effect of SNPs on the binding amnit)' of transcription factors (TF)
SNP Id Matrix name Ref. seq Alt seq Diff P - value P - value log(p)
1520544 crr \'S\TAfJ 02 06061423 0.0337574 1.2q~()~8
n.' .. OjU .. ·· VSQ,\TA(..03 0.731451 (1010&761 I 0136852
1535567443 crr V$CMYB 01 0.2206408 o IJ()H4334 14176806
1539 I 74 I 9)la V!lftF1_OI 0.0911315 0.0083572 1.0376075
153918253 l T V$HMGIY_Q6 05560821 0.0551234 1.0038031 VSXVENTI_OI 0.0232621 0.002028 1.0595758
n60I 8446_T"l' vSNfn_c ~31118 0:"0067255 ,1.0774719
,,1J1I18447 \/(J vSAR_Q6 0,4810886 0.0038588 2.0957685
rs6I 2500(AIG VSGLI_Q2 11.2743208 0.0230467 1'015M94
,,"~".1'250_AfT VSTAII_V6 u.0476729 0.0041181 1.0635773 VSMYOGENIN_Q6 0.1470118 0.0115685 1.1040762
,,:.142ll65_crr V$FRI!AC4_11 0.2138665 0.0193179 1.0641825
V$I BX5 O~ 0.03'1876'1 0.6037171 -I 1801125 VSfXR_Q3 0.0033952 0.047'1123 -I 1495793
"llo'loKfI4_cn vSTBX501 0.0277218 03011918 -1.0360213 V$DllLTAEFI_(JI 0.052597 0.5538026 -1.0223937 VSARJn 0.0018993 001939i3 -1.0()'1U II 2 VSIK2 01 0.0154923 0.5231781 -1.5285343 VSNFAT_Q6 0.0105491 03416377 -1.5103506
,,20701lW \ "I VSCETSI P54_02 0.UI97449 0.5708857 -14610939 VSRIXI 02 0.0207801 0.2650878 -1.1057425 VSNFAT~Q4_01 0.0325844 03713534 ·1.0567778 VSGAr,uJ" 001-16743 0.9018547 -1.7885199
r.3918253_crr VSGATAI 01 00352089 0.9617289 ·1.4364002 V5HSfl_oi 00491717 0.5588443 -105557SS
".N I 8256_ GI A VSZIC2 01 0.0157337 0.2536809 -I 2074561 VSPADS C 0.0192206 0.278160b -11614649
153933239 _NC VSP3'lJ>ECAMER_Q2 /);0178768 431J6n81 -1.3115805 VSSMAD_Q6 0.0540426 0.547271 -1.00;4663
",7341614_NG VSALX401 0.0043144 0.045378 -10219218 YSClATA2 02 U.OOl692 I 0.169841 -2.00161 VSGAT \.1-"2 0.0033785 0.152721 -165518 v~c .. ·\r.\_Q6 00032992 0.1272174 -1.S861349
"WI 8447_AIG \ ~I ~1U2COM_()2 0.0174508 0.3838585 -1.3423568 \ ~(,.\I" \1 06 0.0101607 0.1'109058 ·12738972 V$l •. \ I A 1-(14 00145472 0.2623143 -12560411 VSGAIAI-02 0035281 0.4147284 -10702223 VSOATA C 0.0197873 0.2188523 -1.0437649
r ... flllfltdlJ.l :\:, V$OCTI~Qb 00050282 0.05403 -10312217
"6125005_ -\:(i V$!>TATl 02 00159563 071~ -1.6543035 VSSTAT6:02 0.0803294 0814'1153 -100623&
156203725u AfT VSE2 01 00105583 0.1623362 -1.1868227 n614951_TK.; vroC'TI 114 0.()O(';607 0.11087117 -1.191661
"73422265_CfI VSAfFI_Q6 0.0100743 0.1318196 -l.lIb7667
VS~MAD ~6 00460644 0.5461202 -10739232
130
University of Ghana http://ugspace.ug.edu.gh

Table 4. 20: Top 5 transcription factors (TF) impacted by rs2304527 T/G . . SNP Impact
Rank Transcription Impact on Tf Motif Scan p-"alue Factor (Tf)
POU2f2
STAT
3 EP300
RORA
5 TFAP4
POUJfJ_' fIIdr ..... ,.,...,.
Loss of
'""''''''4 " ':'!'I~ SOlI'
Gain of function
function
.I'"
,-A-;;CGfAT ~44+~:r"4-T' cAAiT"f ~A~~CGtT,
.Acr::~n=fA. ·~L#T~+T~
~LuiLrrr Acy':'c;ATTA,
.. ~ - .A •. t;'GG-i:A·TA'GTC~ Gain of ' 1\ I \I I I', I\i \ l I
function , d ii-''jii- It \ l I
Cain Of~" functiun
- .A. T-r.';T.·A TA.llGTC~
TFAP'_I"'" 8 ... ,. ra2J01U7
.--:CAGCT~.
.\.} l'~ y <; T (:LM-,
0.0005
0.0017
0.0070
0.0127
0.0153
S\P ImpUd p-"alue,\ were used in ranking the top 5 TF impacted by'\'\I\ I S\ /' 11/01 lead to U Rain ur loss inJunction which was (!,I'aluated with the atSNP webtool, atSNP uses SNP infrJrmation and position weight mutricl!s (PWM) 10 predicllhe e{fe,·t ofSNP on TF. First-order Markov model and ImportanCI! .\(Jl1Ipling methods were used 10 detl!rl1lil1l! lhl! statistical !liRnifjcance q{ the best match betwl!('l1the PWM and Ihe subsequence overlapping the SNP position with both the reference and the S\P alleles. Finally. the impact ofSNPs un PWM and subsequence matche,\' werl! tested using the same statistical approach
131
University of Ghana http://ugspace.ug.edu.gh

Table 4.21: Top 5 transcription factors (TF) impacted by rs3918256 G/A
Rank
2
l
. SNP Impact TransactIOn Impact on TF Motif Scan p-value Factor (TF)
NR2C2
TBXI5
TBRI
ZNF345C
TEADS
Loss of function
Loss of functiun
Loss of function
I ,fJ" Il' funl"lion
Gain of function
~
<0.0001 ............. .tIa
CC<.JTACCA ~CCC~j..
0.0011
0.0020
0.0021 ___ ... " .. nw...,. .....
T~CC~(,
~Act~;.
Ref
I"
0.0033
S,\'" impact p-vallies were u\ed in ranking the lOp 5 Tf IIIlpacfed by SNPs, A SNP may lead to a Ka/n or I()~\ injunction which WQ.\' evaluated with the atSNP webtool, atSNP use., SNP information and pmilUJII weiKht matrices (PWA1) to predict the effect o/SNP on TF Fint-order Markov model and Impurtan,'e sampling methods were used to determine the statistical significance 0/ the best match h.:fllwn the PWM and the subsequence's overlapping the SNP position with both the reference and the SNP alleles, Finally, the impact o/SNPs on PWM and subsequence matches were tested using the ,\,Ime stallstieal approach.
132
University of Ghana http://ugspace.ug.edu.gh

Table 4. 22: Top 5 transcription factors (TF) impacted by rs3917419 s~~~mpact Rank Transaction Impact on TF Motif Scan p-value
Factor (TF!
ELK4
2 IRFI
3 API
NFATC2
HNF4
loss of function
Gain of function
Gain of function
l.1l'~ of IUllclion
Lo,~ of fUllction
~
• I~~~
..,' .. ' ..... lkMtfOf ..... "'.
M~:;;,.GMA!.~ . . ~ \l;~~ rr,~4-~~·
.r. A ,\.'l1fT(;A.UA( ~,;QAAk': .
NFATC2,.111oW ac. hw raUt'"'' .... cif:r;;-~r;rr+i
.......,"wRfI .......... ACITTTG
$NP .~CCtTT~ .
. AC'A;;G'G;C. \(. :~~'T(;',I" \
0.0004
0.0011
0,0025
0.0059
0.0100
\\f' Inl!'", 'Ii.values were used in ranking the top 5 TF impackti by S\P, A SVP may feud to a xam "r /"" in {unction which was evaluated with the atSNP webtool. atSNP uses SNP information and {,{).Iition weight matrices (PWM) to predict the effect ()/,SNP on TF. First·order Markov model and importance sampling methods were used to determme the statistical significance of the best match hetween the PWM and the subsequence overlapping the SNP position with hoth the reference and the SNP ullele.\'. Finally, the impact ofSNPs on PWM and subsequence matche.\' were tested using the ,Iume ,Iulwica! approach.
133
University of Ghana http://ugspace.ug.edu.gh

Table 4. 23: Top S transcription factors (TI) impacted by rs684951 T/G SNP Impact
Rank Transaction Impact on TF Motif Scan p-value Factor (IF) _
MEF2A Loss of function
0.00106196
• •
':-!O' .....• ~
P', ~
CtlXI Gain. fuDdi.
oat._ ..... -., ~"i";iinr~ . *r·~T.rAT·~T~:l ·~\Wf.rfH\·T ~~~.
0.00185737
I MEF2B
FOXGI
MYEF2
Lo~s of function
LII'~ of function
Lo\\ of function
..,.~' ............... 1
. ~C.r.Ai.";~AlAcr •. LU.~I·\P~
lAu;.\'($.\ I.~ .... (;IA,,~.IA.G..: ..
_ .. _---, .. TAAACA .. -:;::-f.:xAC, II ,,·1 I !'I .. r..1~T'1
II .,\ I .1~\j\1 .. 1 '.1 I.-} "TAAACA~ TGT",uCIi
1If&f1_ ........... ~
... -a';iA~, I 1 II\, \' I \ LH At. q ( r I T tt\-:-\..\f"n \1.r \ I
.... T ~~AAA T AACi ~ ..
0.00535357
0.00561902
0.01587156
S.\!' ""r,lL I {'-I'alue.~ were used in ranking Ihe lOp 5 TF impacted by SNPs. A SNP may lead to again or lo.u in Jimction which was L'\'aluated with the atSNP webtool. atSNP uses SNP information and po,ition weif{ht matrices (PWM) to predict the effect ofSNP on TF. First-order Markov model and importann! sampling methods were used to determine the statistical significance of the best match hefH'een the PWM and the suhsequence overlapping the !iNP position with both the rqerence and the SNP allele.1 Fina/~l'. the impacl ofSNPs on PWM and suhsequence matches were tested using the same stali.l'/ical approach.
134
University of Ghana http://ugspace.ug.edu.gh

Table 4024: Top 5 transcription factors (TF) impacted by rs2070744 T/G
Rank . SNP Impact
Transaction Impact on TF Motif Scan -value Factor (TF) p
NR3CI L.U\' of fUllclioll
0.00159594
.. Atohl 1.1"~ of
IlIlIdioll 0.00296101
~
3 NFY (.ain of function 0.00342305
-' ~
,::'1 (.;1111 "f
fllndiOIl ~". b.",'
CEBPB 0.00547063
~ ........ r_ ....... _ .....
Lo~\ of function
RORA
.t'CTGlIAT-.-.:rr;"?-·T <A.
~ll WI,1 ~ I \\ .. 0.00703981
,t. ' .~\ I j \t I~, ,-. -\
$t;.r" riA:~ - ,"CTG.GAT A· TGG-T. lAo. SNP Impact p-values were used in ranking the top 5 TF impacted by SNPs. A SNP may lead to a gam ur luss in functiun which was evaluated with the atSNP webfuol. atSNP uses SNP i"formation and position wt!ighf matri"es fPWM) to predict the effect of SNP on TF. First-order Marku\' model and importan,'e Jamplinf( methods were u.led lu determine the statistical significance of the best match between the PWM and the sub,~equen,'e overlapping the SNP position with both the reference and Ihe SNP allele-s. Finally. the impact of SNPs on PWM and subsequence matches were tested using the same statistical approach
135
University of Ghana http://ugspace.ug.edu.gh

Table 4. 25: Top 5 transcription factors (TF) impacted by rs59055740 T/G Transaction M 'f S SNP Impact
Rank Factor (TF) Impact 00 TF 011 can p-value
NR2C2
FOXGI
MLX J:1
TEADJ
ATF6
Gain of function
Loss of function
(.ain of
'"lIclion
Lo" of funelilm
Gain of function
~ , .-
;~
_.,_--.. ..-. -,W-uCAOh...
".1l.lt u.f \lIrit..-L . ~I,.
ll\J'c'~~h'CYvIJlI.I~ ,t;iGAC'CAAT '.
.TGAC(rfG~ ltLLy4~Y(~
o
0.00223591
0.00318962
0.00349957
0.00414664
S\ f' impu( f p-I('//lt',\ were used in ranking the top 5 TF impacted by the It!lf S,\{, , S,\P may /<!ad to "gain ur IusI' in function which was evaluated with the atSNP webtool. afSNP uses SNP information £Ind position weight matrices (PWM) for a transcription/actor to predict the effect o/SNP on TF. First-order Markov model and importunce ,wmpling n1<!thod are used to determine the statistical significance 0/ the best match hetween the PWM and the subsequen"e overlappinK the SNP pOJition "'ith both the reference and the SNP alleles. Finally, the impact o/S/vPs on PWM and subsequence matches are tested using the same statistical approach.
136
University of Ghana http://ugspace.ug.edu.gh

4.11.4. Effects of trait- and malaria-associated SNPs on microRNA
The MicroSNiPer tool in ChroMoS was used to check for SNPs that may affect target
sites ofmicroRNAs. Two of the SNPs, rs3918211 and rs20S44, affect microRNA target
sites: the former affected 5 microRNAs whiles the latter affected 14 (Table 4.20 -21).
The reference and alternate variants of rs20S44 [Crr] affected seven microRNA each
whilst that of the rs3918211 affected two and three microRNAs respectively.
137
University of Ghana http://ugspace.ug.edu.gh

Table 4. 26: microRNA-bindin~ ,ite~ influenced by rs3918211 IGIAI variants SNP microRN
microRNA Target Site \ariant A
hsa-miR- :... t60 fr:: 110 430 420 ~CATGlT.f~::: -;C7GCGATGTTACCATGGCAACCAACGTCCTGCAGA
rs39182I 409-5p AGGt1tIACCCGAGCAACUUOGCAO
lC 20
IIG/AI hsa-miR- He 'IS: HO 430 4:10
:~GJGCCACATGTTTGTCTG: ';GCGAT-GT fACCATGGCAACCAACGTC
4467 UGG"::;~':;':GGUAGUUAOGGGCUU
lC 20
hsa-miR- HO iSO HO 430 420 110 8'3'; JGCCACATGTTTGTCTG::GGCGAl'GT' ACCATGGCAACCAACGTCCTGCAGACCGT
767-5p OGCACCAOGGOOGOCOGAGCAOG 10 20
rs391821 hsa-miR- f70 He 450 110 430 420 CTCGA;C 3GGGCCACATGTTTGTCTGCGGCGATGTCACCATGGCAACCAACGTCCTGCAG
I 1(;1 \ I 2964a-5p AGAOGOCCAGCCACAAOOCOCG 10 20
h~a-miR- t'O '\10 430 420 no 118CaGGGCCAc'_: .. :-. -;::1 A .JGCGATGT(ACCATGGCAACCAACGTCCTGCAGACCG
1226-3p OCACCAGCCCOGOGOOCCCOAG 10 20
138
University of Ghana http://ugspace.ug.edu.gh
Table 4. 27: \licroRNA-binding sites influenced by rs3918211 IC/AI variants SNP \ ariant
rs20544 (errl
rs20S443 ICITI
microRNA microRNA Target Site
hsa-miR-6165 Tccc:rG~~ad;. - <::~~~~c= <7C
CAGC1.GGAGGUGAGGGGAG
_________ -'-:,I.,J.{ ... ;:_<~~-·. -, ;,c,~=::;:~ul hsa-miR-483-5p
hsa-miR-505-5p ~ A:;"" ;.~::Ac:~CAAA~CGGGA <: ... -
j'.Y·'·' ' ~I..oAcn......:A~QAD<il1 20
at ~ .... : ~·~(·A":TGCCAAAGCAGGACGGGAr.cC
hsa-miR-5591-5p - ________ -il:Iof.,~Ol'~.F': , UGGGAacu~""'()AUGG<;U"G
hsa-miR-920
hsa-miR-6090
hsa-miR-4300
hsa-miR-4436a
hsa-miR-468I
hsa-miR-76I
hsa-miR-184
hsa-miR-425-3p
hsa-miR-4471
hsa-miR-4422
c; ~GGAGCOG'7GGa aaa.G1D 20
30 20 '10 :iGGATTTACATGGCltCTGCCAAAGCAGGACGGGAr.cC
GGGGAGCGoAGGGGCGGGGC 10
:10 10 -;,~GATTTACATGGCACTGCCAAAGCAGGACGGGAGCC
UGGGAGCUGGACUACUOC 10
'I ~ J0 20 10 ~ ...,:- :.GGGATTT ACATGGCAC'IGCCAAAGCAOGACGGGAACC
GCAGG,.c;~"*.·GUGGAD
10 20
40 u 10 T, ,'· .... ATTTACATGGCACTGCCAAAGCAGGACGGGAA
:;::::1 AACClGGl.A.OCCA.G<JCtJGaAOetJ
10 20
1:0 30 20 to ": : CCAGTGGGGATTTACATGGCACl'GCCAAAGCAGGACGG<a.AcC
GCAGCAGG--GU~
10 20
30 20 10 ;":'-;G·JV.G.ATTrACATGGCACIGCO'AGCAGG.\CGG-GA.ACC
::::1: :::: tl~AtlAAGGGO
10 ----------
30 ao 10 .~(jGAI'TT ACATGGCAC'IGCCAAAGCAGGACGGGAACC
30 20' ~o
AOCCiGGAAOGUCGOGOCCGCCC 10
GGGATTTACJ;TaGCACTGCC1.AAGCAGGACGG<a.ACC
10 '0 ~o 10 ",. T ~ C :AGTGGGGAl'TTACAl'GGCACTGCCAAAGCAGGACGGGAIoCC
: ::1:
139
A.AAAGCA--~ACCCA 10
University of Ghana http://ugspace.ug.edu.gh

4.12 Prospecting biomarkers for .:erebral malaria.
This study evaluated the prospects of angiogenic and parasitological variable as
biomarkers of endothelial damage and CM. Variables that were evaluated were chosen
based on their association with endothelial integrity or malaria phenotypes. Receiver
operating characteristic (ROC) curves were used to assess how sensitive and specific
these variables were in discriminating CM from VM and ProDamage from Pro Repair
(Fig 4.21-24). The best performing angiogenic factor for discriminating CM from VM
was NORI with an AVC of 0.66 (p = 0.0013,95% CI = 0.6 - 0.76, Fig 4.21). Figure
4.22 shows the performance of angiogenic factors in discriminating ProDamage
individual from ProRepair. NORl, eAl and TEK (Tie-2) to perform comparably in
discriminating ProDamage from ProRepair, but all the factors had an AVC < 0.65.
Results on the performance of parasitological indices (total parasite biomass (Ptot),
HRP2 and parasite density) evaluated for similar discriminatory abilities is shown in
Fig 4.23. The similarities in the performance of Ptot and HRP2 in discriminating CM
from VM is noteworthy (fig4.23). However, the performance of the often parasite
density estimate was disappointing (AVe = 0.54, 95% eI = 0.45 - 0.65, P = 0.37).
Figure 4.24 shows that none of the parasitological indices was a good discriminator of
ProDamage from ProRepair (AVe = 0.434, 95% el = 0.34 - 0.53, P = 0.172); HRP2
(AUe = 0.535, 95% CI = 0.44 - 0.63, P = 0.471); Ptot (AVe = 0.582, 95% ci = 0.49-
0.68, P = 0.087).
140
University of Ghana http://ugspace.ug.edu.gh

Derived from 1 . Specificity 00 o~ 04 Oli o. 10
10 --~--• ,.i1' 0
08..-----t-- - --~---o~ 0
ri • :::1 <
_-1' • ~ 06
Q.
>
) (' ,06 :
E 3 lit C
104 ! • 04
,J ,/ en ! ,.,
iI / <
0: Ar 102 ~
K c= ' ---.-J OO 02 0.4 06 O. HI
1 • Specificity
Figure 4. 21: Performance of angiogenic factors as a biomarker for eM
141
mfnP_9 NGRI TI~':! CAl AngIopoltnl
- - Reference line
- ------'i!""'""--_----..... _ •• k~_ ~ •••
CAl
_~I
0*114 . "ot.{'l, Il,."IM
"11m OO)tn 71
0 . ...,.,. ~~'"
Performance of MMP9. NGR1. Tie-2. CAl and angiopoietin 1 as biomarkers for CM. None of the markers performs satisfactorily as a biomarker for CM. A satisfactory marker is expected to have an Area Under the Curve (AUe) > 8.
University of Ghana http://ugspace.ug.edu.gh

Derived from 1 - Specificity 0.0 02 0.4 06 08 10 .• '/
"O~[---. -.~'0 i ,,".'
0.8 1 . 10.8 0 ct :1
I ~ ~ 0.6 i 10.6 ~ ~ \ 0 • 3 c 0 ~ 0.4 (. 04 !
• a:
., .I-' % I., ~ --------~OO
0:2 0.4 0.6 0.8 '.0
1 • Specificity
Source of the Curve
mmp_9 NGRI
-Tie·] CAt Anglop01lO1
- - Reference Lme
mmp_9 0.46
NGRl 0.83 001
TEK 062 001
CAl 081 002 _A~opotinl 058 008
(0.54,072)
(0.53.0.70)
(0.52.0.70)
(0.49.0.69)
Performance of MMP9. NGR 1. TIe-2. CAl and angiopoietln 1 as biomarkers for eM. None of the rnalters performs satisfactorily as a biomarker for endotheHal integrity. A satisfactory marker is expected to have an Area Under the Curve IAUC) > 8.
Figure 4. 22: Performance of angiogenic factors as a biomarker for endothdial integri~
142
University of Ghana http://ugspace.ug.edu.gh

Oerlved from Sensitivity -o ~ 04 o ._~ 08 • 0
r" ' JlI 0 pPala5.1. Oenlrty
HRP_: Plot
0.' .5 O' li' Referenc. lJnt ::l. < •
~{ a.
U"', ' 06 a' E I -
u 3 pParaslte d4?'f"lS11V 0 546 0372 (0445,0648) • a. HRP 2 0.253 < 00001 (0 lea, 0,338) fI) 04 '04 fI
• D10t 0.2511 < 0.0001 (0173. 0 345) n
'r 3i n 02 ~
oo-~- Performance of parasite density, HRP 2, total parasite ----- - ~o.o
biomass in predicting CM. A satisfactory marker is 0.0 02 04 06 08 10
Sensitivity expected to have an Area Under the Curve (AUe) > 8.
Figure 4. 23: Performance of parasitological indices as a biomarker for eM
143
University of Ghana http://ugspace.ug.edu.gh

0.2 0.4 0'" 06 10
;.l10 r! pPafJSlte density
-J'/- J HRP~
-Ptot 0.81 ;1""/' 08 0 Refetenc. line
_. J' • ::2. <
I • 0,61 j-
10.6 ~ 0 pParlllllllt denSIty 0.434 O.ln (034.0.53)
3 HRP_2 0.535 0.471 (044.0.63)
'" Plot 0.582 0.087 (049. 068)
'" 1:
04 !
0." (.J •. .. tt <
10.2 ~ Performance of parasite density, HRP 2, total parasite I
I ~- biomass in predicting endothelial damage. A
0.0-- .--.. - '-------' 0.0 satisfactory marker is expected to have an Area
00 0:' 04 06 0.8 10 Under the Curve IAUC) > 8.
Figure 4. 24: Comparison of 3 parasitological variables as biomarker for eM
144
University of Ghana http://ugspace.ug.edu.gh

4.13 Association of SNPs with angiogenic factors
Although SNPs tested in this study were not selected based on overt functional
properties, those that were associated with malaria and/or endothelial integrity were
further tested to determine their association with the angiogenic factor they encode.
Comporiion 0/ MMP9 ,.... among r53918256 in a fffflU/1If! inh.,,"mc. mod,,',
Mo......whitMy _.-Jolla
• u_ :., _nl'llnllfllollM} ,aii Moon rank fOOl 74.0
P - v.lue .0046
200 400 600 800 1000
levels of MMP9 111IImL)
Figure 4. 25: Comparison of MMP9 levels among rs3918256 in a recessive model
The rs3918256 variant associated with almost a 4-fold risk of ProDamage was tested to
determine its association with the levels of MMP9 in the study population. Since
rs3918256 was associated with endothelial damage in the recessive inheritance model,
levels of MMP9 were compared between GAJAA and AA variants. With a mean rank
of 74.0 and 55.6. level in MMP9 in GG was significantly higher than GAJAA
respectively (p = 0.046. Fig X). Other SNPs in the coding regions of NGRI
(rs2466104). TEK (rs7023443 and rs7874391) and CD4 (rs2886398) were neither
associated with malaria, endothelial integrity nor their respective proteins.
145
University of Ghana http://ugspace.ug.edu.gh

CHAPTER FIVE
5.0 DISCUSSION
This study genotyped 27 SNPs, characterised their epigenetic context In silico, and
explored their interrelationships with endothelial integrity and malaria phenotypes. It
also explored the potential influence of SNPs on transcription factor and microRNA
binding sites. The study father explored the diagnostic value of various angiogenic and
parasitological variables. This section discusses the findings from this study in
integrated and thematic prose.
5.1 Association SNP! with malaria and endothelial integrity:
Discussions of SNP-disease/trait association analysis excludes associations that fell
within 0.002 < p > 0.05 because of the conservative significance threshold (p < 0.002)
the study adopts. The only SNP (rs2304527) found to be associated with CM in CM
versus UM comparisons was an intronic SNP located in the CDHS gene which encodes
cadherin 5 or VE-cadherin, a vital protein for cell-cell adhesion and intracellular
junction integrity. This SNP offered children with the heterozygous (T/G) genotype
protection that made them about 3 times less likely to have CM relative to their
colleagues with the IT -GG genotype. Regardless of the massive protection
rs2304527(T/G) affords, there is very little literature on this SNP (Schubert et aI., 2014).
Intronic SNPs can sometimes serve as tagged SNPs, and thus. their association with
diseases and/or traits could be due to their proximity to other SNPs in coding regions.
However, the four SNPs in linkage disequilibrium with rs2304527 do not occur in the
coding regions ofCDH5. Rather, they occur in introns in strong enhancer (rs620372S0
146
University of Ghana http://ugspace.ug.edu.gh

and rs58044782) and transcriptional elongation (rs3785286 and rs2880989) regions of
the genome. Once regarded as useless. introns are now known to play important roles
in gene expression. They harbour multiple functional elements such as intron splice
enhancer and silencers that control alternative slicing. trans splicing and other pre- and
post-transcriptional elements (Cooper. 2010). Mutations in these regions of the genome
can, therefore. lead to a cascade of downstream aberrations that culminates in
modifying clinical phenotypes. Taken together, although rs2304527 and it linked SNPs
are yet to be associated with disease or a clinical phenotype in literature, their location
and epigenetic context do not preclude them from playing a role in the pathogenesis of
CM.
Another SNP that breaks through this study's significance threshold is the splice region
variant (rs3918256) in MMP9 which was found to be associated with endothelial
integrity. In the recessive inheritance model, children with the homozygous recessive
genotype (AA) were almost 4 times more likely to be ProDamage. This SNP has been
evaluated for association with inflammatory diseases and angiogenesis in several
studies but no associations were found except in a study among non-Hispanics whites
women where it was associated with protection from pelvic prolapse (Amankwah et aI.,
2012; Haq et al., 2010; Nakashima et ai., 2006; Wu et aI., 2012). This is the first study
to implicate rs3918256 in endothelial integrity in the context of malaria pathogenesis.
Larger studies in this population and elsewhere will be required to shed more light on
the role of this SNP. The fact that rs3918256 was associated with endothelial integrity
and not CM in this study popUlation is noteworthy. The association between endothelial
integrity and CM has been hypothesised for decades but study design challenges and
the rare nature of CM has limited studies from directly evaluating the empirical
147
University of Ghana http://ugspace.ug.edu.gh

relationship between eM and endothelial integrity. (Bernabeu and Smith. 2017;
Graham et al., 2016; Souza et aI., 2015).
Although the case-control comparisons were not a priority in this study, it revealed
several interesting results. Four SNPs located inlor close to the SELE (rs3917419),
pseudogene (rs684951), ZMYND8 (rs590S5740) and NOS3 (rs3918211) loci were
associated with malaria at GWAS significance thresholds in malaria versus healthy
control comparisons. For instance, the intronic rs3917419 variant which has no
previous malaria-related mention in literature was found to increase the risk of malaria
by nearly 5 folds. Only two studies make reference to this variant associating it with
elevated levels of serum E-selectin and MMP9 (Montasser et al., 2010; Santos et al.,
2018). E-selectin is implicated in eM pathophysiology and thus, its association with
rs3917419 suggests that the SNP may have a role in eM pathogenesis (Renia et al.,
2012). Although Santos et al reported an association between rs3917419 and MMP9
levels, this study did not see a compelling biological reason to pursue that analysis
(Santos et a1.. 2018). All SNP in linkage disequilibrium with rs3917419 were intronic
or upstream gene variants except for two (rs7531675 and rs7543618) that were in a
regulatory sequence. There are a total of9 studies that make references to SNPs linked
to rs3917419 but none is in connection with malaria (Edwards et aI., 2011; Faruque et
aI., 2011; Hsieh et al., 2017; Montasser et al., 2010; Mullins et aI., 2011; Nasibullin et
aI., 2016; Solus et aI., 2015; Timasheva et aI., 2015; S. Wu et aI., 2012).
Another of the four SNPs associated with malaria at GWAS significance threshold is
the intergenic variant (rs68495I ) located in a pseudogene on chromosome 6. This SNP
and the SNPs in linkage equilibrium with it have not been associated with any clinical
148
University of Ghana http://ugspace.ug.edu.gh

phenotype yet. However. the synonymous variant (rs3918211) of NOS3 that was also
associated with malaria at GWAS significance threshold has some malaria-unrelated
mentions in literature (Blanton et al., 20 II; MacClellan et al., 2009; Shaw et al., 2009).
5.2. SNPs and epigenetic mechanisms: insights from in sill co analysis
In silico analysis found several SNPs that influenced transcription factor binding sites
(TFBS) by quantitatively influencing the binding affinity of transcription factors.
Interestingly, some SNPs associated with susceptibility to CM were predicted to
modulate the binding affinity of transcription factors. For instance, rs3917419,
rs3918256 and rs68495I were found to lower the binding affinities of some transfac
matrices. This study further evaluated which TFs gained or lost binding sites through
the impact of SNPs and found interesting results. For instance. rs3917419 in SELE
(gene encoding E-selectin) was found to influence two different families of
transcription factors (NFAT. and API). Whereas the rs3917419 mutation leads to the
loss of binding site for NFATC2, it created same for API transcription factors. The
NF AT family of transcription factors are known to enhance endothelial cell survival
via VEGF-mediated effects, (Hamik et aI., 2006a) and thus, the rs39I74I9-mediated
loss ofNFATC2 binding site could have implications for vasculopathies such as CM.
In the same breath, rs39I74I9 could lead to the creation of a binding site for API, a
transcription factor that regulates a wide variety of biological process including cell
differentiation. proliferation, apoptosis and angiogenic related processes (Hamik et al.,
2006a). Thus, although rs3917419 is an intronic SNP with yet an unknown function, it
can plausibly influence CM pathogenesis via altering angiogenic-related processes
during malaria.
149
University of Ghana http://ugspace.ug.edu.gh

Further analysis of the effects of the CDH5 (gene encoding Cadherin 5) SNP rs2304527
revealed that the SNP influences the STAT family of transcription factors. This
mutation creates a binding site for STAT, a transcription factor that promotes
angiogenesis by activating VEGF. Similarly, the rs3918256 SNP in the gene encoding
MMP9 disrupts the binding site for the ZnF family of transcription factors which are
among the most abundant DNA binding proteins (Hamik et al., 2006b; Miyashita et al.,
2004). Taken together, although there is a paucity of literature on these SNPs, In sllico
analysis provides a window for further insight into these SNPs and a basis for
developing future research hypotheses.
The finding that SNPs associated with CM and endothelial integrity disrupt or creates
microRNA target sites is interesting. There is an increasing body of evidence on the
importance of SNPs in miRNA and their biological consequences. In this study, two
SNPs (rs20544 and rs3918211) where found to disrupt target sites for 19 different
microRNAs, some of which, play pivotal roles in the post-transcriptional modifications
in various disease models Deciphering the role of miRNA in malaria is still in its
infancy (Chamnanchanunt et aI., 2017; Mantel et aI., 2016; van Loon et aI., 2019) and
it will be interesting to further explore the biological role ofrs20544 and rs3918211.
5.3 Endothelial integrity and malaria phenotype
Downstream analysis using the endothelial integrity variable produced results
consistent with conventional knowledge on CM pathogenesis (Postels and Birbeck,
2013). This finding is very instructive and constitutes a de facto validation of the
endothelial integrity variable used in this study. Although scientists have long known
that insults to cerebral microvasculature and concomitant endothelial damage could be
150
University of Ghana http://ugspace.ug.edu.gh

central to the pathogenesis of eM, limitations in the in vivo assessment of endothelial
integrity during a P. Jalciparum have hampered research efforts (Widmer and Lennan,
2014). Leveraging EPCs attributes as a good correlate of endothelial function (Dzau et
al., 2005; Venna et aI., 2017), this study converted EPC data into a binary variable
using a Gaussian mixture model. Traditionally, mean and median splits have been used
to convert continuous biomarker data into discrete variables but this approach has come
under heavy criticism in recent times (Iacobucci et al., 2015; Rucker et aI., 2015).
Mixture models are preferred because unlike the mean- or median-split, it estimates
cut-off points using the distribution of the variable or by optimizing the correlation with
outcomes (Budczies et al., 2012; Trang et al., 2015). Gaussian Mixture Models are not
without limitations but its major drawback of decreasing perfonnance with increasing
dimensionality was not apparent in this study (Bouveyron et ai., 2007). This study was
interested in creating and binary and thus fit the data to only two mixture models. This
user-defined operation can prevent the detection of other clusters that may exist in the
data (Leisch, 2004). Besides showing that children classified as ProDamage were more
likely to belong to the CM phenotype, this study provides evidence for the anticipated
but rarely demonstrated fact that individuals with non-CM phenotypes have better
endothelial integrity. In the pathophysiologic model underpinning this study, CM is
conceptualised to result from a disequilibrium between damage and repair and thus, the
ability to show here that non-CM phenotypes are Pro Repair is instructive.
151
University of Ghana http://ugspace.ug.edu.gh

5.4 Malaria in context: parasitological, immunological and baematological
indices.
5.4.1 Parasitological indices
Parasitaemia is an important but tricky malariometric index. The sequestration of
infected erythrocytes in the microvasculature of vital organs makes parasitaemia
assessment based on peripheral blood (PbP) unreliable (Berendt et al., 1994; Clark and
Alleva, 2009; Cunnington et al., 2013; World Health Organization, 2016c). This study
accessed three parasite estimation methods and found total parasite biomass (PlOt) and
HRP2 to better discriminate between UM and CM (fig 4.23). Scientists have debated
for decades on whether parasitaemia is a good predictor of CM but data from this study
and that from others cautions that the debate may be wrongly premised if peripheral
parasite density is used in estimating parasitaemia (Addison, 2017; Giha et aI., 2005;
Tangpukdee et al., 2012; Wilairatana et al., 2013). On the contrary, there appeared to
be very good concordance between Plot and HRP2 predictive abilities, and thus, the
prospects of developing HRP2-based quantitative diagnostic kits is promising. All the
parasitological indices considered in this study were poor in discriminating ProOarnage
from ProRepair (Fig 4.24). Thus, in spite of decades of research, the relationship
between the triad of parasitaemia, CM and endothelial integrity remains blurred. Future
studies may benefit from more robust study designs that aptly combine animal models,
in vitro cell culture, and human subjects to properly triangulate research questions
(Ghazanfari et aI., 2018).
5.4.2 Immunological indices
With the exceptions of C04+ T cells which was higher in the ProRepair group, none of
the immunological variables measured in this study differed between UM and CM.
152
University of Ghana http://ugspace.ug.edu.gh

Many aspects of the role ofT cells in malaria protection are still unclear but the role of
CD4+ T cells in stimulating B cells and promoting phagocytosis is well established
(Motaet al., 1998; Perez-Mazliah and Langhorne, 201S; Podobaand Stevenson, 1991).
Generally. CD4+ T cells are responsible for the production of IFN -y which plays an
important role in malaria immunity. especially, in controlling parasitaemia in acute
human infections (Wykes et al., 2017). However, in the specific case of endothelial
damage during P. /alciparum infections, CD8+ T cells rather than CD4+ T is thought
to playa leading role in the immune-mediated damage of the endothelium (Renia et al..
2012; Wykes et al.. 2017). Thus, the finding of comparable levels ofCD8+ T cells in
the ProRepair and Pro Damage groups was unexpected. Perhaps, this is due to a
discordance in CD8+ T cells levels circulating in peripheral blood and those
sequestered in brain endothelium (Dunst et aI., 2017). The higher levels of CD4+ T
cells seen in the ProRepair group may reflect a generally better immune response in the
ProRepair group. Taken together. endothelial damage during a P./a/ciparum is a multi
step process involving several immune players and the examinations of immune
signatures rather than individual immune correlates may provide better insights
(Valletta and Recker, 2017). Even though immunological parameters considered in this
study differed among malaria phenotypes, the pairwise comparisons between CM and
UM revealed no significant differences - a further testament for the need to consider
immune signatures in future studies.
5.4.3 Angiogenic factors
Angiogenic factors associated with endothelial integrity in this study included the
angiopoietin receptor Tie-2, neuregulin I (NRGI), carbonic anhydrase I(CAI), and
angiopoietin 1 (ANG I). Higher levels ofTie-2 in the Pro Damage portrays a more potent
IS3
University of Ghana http://ugspace.ug.edu.gh

endothelial activation in the Pro Damage group relative to their ProRepair counterparts.
These findings become interesting in the broader context of literature on Aug-Tie-2
signalling and CM pathogenesis, some of which, have proposed plasma concentrations
of Ang-2 and the Aug-2/Ang-1 ratio to be an independent predictors of death in
different populations (Conroy et al., 2012, 2010; Jain et al., 2011; Lovegrove et al.,
2009; Prapansilp et al., 2013; Yeo et al., 2008). Both systemic factors and local
vasoactive substances can induce endothelial activation in CM and further studies will
be required to decipher the actual role of Aug-Tie-2 signalling in CM pathogenesis.
Relative to the ProRepair group, the ProDamage group had higher levels ofNRGI and
CAl. These two angiogenic factors have not been extensively studied in the context of
malaria. however, considering their normal biological roles, it is conceivable that they
influence the pathophysiology ofCM and endothelial dysfunction. For instance, besides
CAl's role is the pathological remodelling in ischemic cardiomyopathy, it has been
shown to influence endothelial cell permeability and endothelial cell apoptosis in vitro
(ToreUa et al., 2014). On the other hand, NRG 1, which is a cell adhesion molecule has
been implicated in the pathogenesis of experimental CM (Liu et aI., 2018; Solomon et
aI., 2014). Interestingly, whereas higher levels ofNRGI was found among CM patients
relative to UM, levels of CA I was not significantly different between UM and CM.
These factors have not been traditional candidates in malaria pathogenesis but findings
from this study suggest that they could play important roles. Despite the significant
associations of some angiogenic factors with endothelial damage, ROC analysis to
prospect for CM biomarkers produced middling results. This could reflect the fact that
the development of CM is a multistep process with endothelial activation representing
only a snapshot of that spectrum.
154
University of Ghana http://ugspace.ug.edu.gh

An incidental finding from the data on general characteristics of study participants
broaches the changing maIaria epidemiology and its implications. The decline in
malaria burden may have several unintended consequences, one of which, is the
predicted shift in the age-specific malaria burden from younger to older children.
(Carneiro et aI., 2010; Pemberton-Ross et aI., 2015). With median ages that are
comparable to similar but older studies conducted in sympatric populations, evidence
for the predicted age shift in malaria burden was not apparent in this study (Dodoo et
al., 1999; Gyan et aI., 2009; Gyan et aI., 2004; Kusi et aI., 2008; Riley et aI., 2000).
Although data from this study is ill-suited for inferences on this subject, the observation
is nonetheless noteworthy.
This study was generally well planned and executed. That notwithstanding, it had some
limitation worth mentioning. First. the problem of missing data that often-beset
complex studies was a challenge for this study as well. Traditionally, researchers
attempt to address missing data with Listwise deletion but this is hardly a remedy for
missing data (Deng et aI., 2016; Goeij et aI., 2013). The use of multiple imputation
strategies are now accepted as the best approach in dealing with missing data (Deng et
aI., 2016; Goeij et aI., 2013; Pedersen et aI., 2017) and this was employed in this study
and implement in SPSS. Secondly, the lack of enough genetic material for additional in
vivo epigenetics analysis robs the study of additional empirical data to corroborate in
silico analysis.
155
University of Ghana http://ugspace.ug.edu.gh

CHAPTER SIX
6.0 CONCLUSION AND RECOMMENDATIONS
6.1 Conclusion
The major findings of this study distil into three main thematic conclusions. First,
Intronic CDH5 (rs2304527) and MMP9 (rs391S256) SNPs are associated with CM and
endothelial integrity respectively. This is the first study to implicate these SNPs in
malaria pathogenesis and endothelial damage, and thus, the dearth of knowledge on the
SNPs are not surprising. In silica analysis from this study suggests that some of the
SNPs may influence endothelial integrity and malaria pathogenesis via the modulation
of post-transcriptional or epigenetics mechanisms. This conclusion is undergirded by
the finding that the SNPs associated with malaria and endothelial integrity disrupts and
creates binding sites for important microRNAs and transcription factors. In the
particular case ofCDH5 (rs2304527) and MMP9 (rs3918256), the SNPs modified the
binding sites for transcription factors involved in various angiogenic processes.
Second, the prospects of using angiogenic factors considered in this study as a
biomarker for CM or endothelial integrity are slim. Whereas the binary endothelial
integrity variable created with a Gaussian mixture model behaved as expected, ROC
results on the performance of angiogenic factors in discriminating clinical malaria
phenotypes and endothelial integrity phenotypes were disappointing. On the other hand,
ROC results indicate that HRP 2 and PIO! estimates parasitaemia better than
conventional parasite density. Parasite density contributes significantly to clinical
decision-making and hence its inability discriminate between clinical malaria
phenotypes is worrying. All the parasite estimates measures considered in this study
156
University of Ghana http://ugspace.ug.edu.gh

performed poorly in discriminating endothelial integrity. This leaves the relationship
between the triad of eM, parasitaemia and endothelial integrity as blurry as ever.
Finally, immunological. parasitological, haematological and other malariometric
indices measured in this study provided the backdrop and context for the discussion of
findings of this study. Decades of malaria pathogenesis research have relied on such
indices for background and context. Thus. although the immunological. haematological
and parasitological indices measured in this study provided no fresh insights, it
triangulated the more interesting and novel findings of this study. Accordingly,
immunological. parasitological and haematological indices can be regarded as abiding
imperatives of malaria pathogenesis studies.
6.2 Recommendations
The recommendations put forth in this section ensues from the empirical findings of
this study and the reflections of the author on the design of this study and others in the
field. Firstly, the author recommends further studies in the following areas: The specific
recommendations are:
a) SNPs identified in this study should be further tested in G WAS to ascertain their
robustness. Previous studies with genomic data available may want to evaluate
these SNPs in their study popUlation. Such retrospective research endeavours
could be more meaningfully if SNPs are explored in a genomic and epigenomic
context. Functional studies of the biological roles of these SNPs will also be
crucial. The role of epigenetic mechanisms such as histone modifications and
DNA methylation must be explored in future studies.
157
University of Ghana http://ugspace.ug.edu.gh

b) Further exploration of Immunohaematologic and maIariometric signatures in a
community based longitudinal studies could provide valuable insights on the
development and resolution of eM.
Secondly, a critical appraisal of the findings from this study and the literature on eM
pathogenesis brings persistent loopholes in our conceptualisation of eM pathogenesis
to the fore. Despite the complex and interdisciplinary nature of eM, pathogenesis study
designs are often overly experimental and unamenable to transdisciplinary knowledge.
This sometimes leads to the exclusion of subpopulations that are thought of by
researchers as confounders, but, are integral to the population they try to study and help.
For instance, the exclusion ofindividual with haemoglobinopathies that protects against
malaria and those with bacteraemia and malaria coinfections can potentially rob
researchers of insights that may be more representative of the populations they study.
This study, therefore. recommends that malaria researchers explore other innovative
study designs that harness current advances in data sciences and statistics to enable
researchers to simultaneously focus on specific hypotheses without losing sight of the
big picture.
158
University of Ghana http://ugspace.ug.edu.gh

REFERENCES
Abdel-Latif, M.S., Dietz, K., Issifou, S., Kremsner, P.O., Klinkert, M.-Q., 2003.
Antibodies to Plasmodium falciparum rifin proteins are associated with rapid
parasite clearance and asymptomatic infections. Infect. Immun. 71,6229-6233.
Addison, T .K., 2017. The role of malaria parasite biomass and immunohaematologica1
indices in the pathogenesis of severe malaria among children in Accra
metropolis of Ghana 124.
Adu. B., Dodoo, D., Adukpo, S., Hedley, P.L., Arthur, F.K.N., Gerds, T.A., Larsen,
S.O., Christiansen, M., Theisen, M., 2012. Fc Gamma Receptor I1IB (FcyRIIIB)
Polymorphisms Are Associated with Clinical Malaria in Ghanaian Children.
PLoS ONE 7.
Adukpo, S., Gyan, B.A, Ofori, M.F., Dodoo, D., Velavan, T.P., Meyer, C.G., 2016.
Triggering receptor expressed on myeloid cells 1 (TREM-1) and cytokine gene
variants in complicated and uncomplicated malaria. Trop. Med. Int. Health TM
IH 21, 1592-1601.
Adukpo, S., Kusi, K.A, Ofori, M.F., Tetteh. J.K.A, Amoako-Sakyi, D., Goka, B.Q.,
Adjei, G.O., Edoh, D.A, Akarunori, B.D., Gyan, B.A., Dodoo, D., 2013a. High
Plasma Levels of Soluble Intercellular Adhesion Molecule (ICAM)-l Are
Associated with Cerebral Malaria. PLoS ONE 8, e84181.
Adukpo, S., Kusi, K.A, Ofori, M.F., Tetteh, J.K.A, Amoako-Sakyi, D., Goks. B.Q.,
Adjei, G.O., Edoh, D.A., Akanmori, B.D., Gyan, B.A, Dodoo, D., 2013b. High
plasma levels of soluble intercellular adhesion molecule (ICAM)-l are
associated with cerebral malaria. PI oS One 8, e84181.
Agarwal, A., Guindo, A., Cissoko, Y., Taylor, J.G., Coulibaly, D., Kone, A, Kayentao,
K., Djimde, A, Plowe, C.V., Doumbo, 0., Wellems, T.E., Diallo, D., 2000.
Hemoglobin C associated with protection from severe malaria in the Dogon of
Mali, a West African population with a low prevalence of hemoglobin S. Blood
96,2358-2363.
Aidoo, M., Terlouw, D.J., Kolczak, M.S., McElroy, P.D., ter Kuile, F.O., Kariuki, S.,
Nahlen. B.L., Lal, AA, Udhayakumar, V., 2002. Protective effects of the sickle
cell gene against malaria morbidity and mortality. Lancet Lond. Engl. 359,
1311-1312.
159
University of Ghana http://ugspace.ug.edu.gh

Allison. AC., 1954. Protection Afforded by Sickle-cell Trait Against Subtertian
Malarial Infection. Br. Med. J. 1,290-294.
Amankwah, E.K., Sellers, T.A, Park, J.Y .• 2012. Gene variants in the angiogenesis
pathway and prostate cancer. Carcinogenesis 33.1259-1269.
Amoako-Sakyi, D., Adukpo, S., Kusi, LA., Dodoo, D., Ofori, M.F., Adjei. 0.0., Edoh.
D.E., Asmah, R.H., Brown, C., Adu. B., Obiri-Yeboah, D., Futagbi, G ..
Abubakari, S.B., Troye-Blomberg, M., Akanmori, B.D., Goka, B.Q., Arko
Mensah, J., Gyan, B.A, 2016. A STAT6 Intronic Single-Nucleotide
Polymorphism is Associated with Clinical Malaria in Ghanaian Children.
Genet. Epigenetics 2016, 7-14.
Anglogold Ashanti, 2004. Anglogold Ashanti, AOA Report To Society, Annual Report.
Anstey, N.M., Weinberg, J.B., Hassanali, M.Y., Mwaikambo, E.D., Manyenga, D.,
Misukonis, M.A., Arnelle, D.R., Hollis, D., McDonald, M.L, Oranger, D.L.,
1996. Nitric oxide in Tanzanian children with malaria: inverse relationship
between malaria severity and nitric oxide production/nitric oxide synthase type
2 expression. J. Exp. Med. 184,557-567.
Apoorv, T.S., Babu, P.P., Meese, S., Oai, P.P., Bedu-Addo, 0., Mockenhaupt, F.P.,
2015. Matrix Metalloproteinase-9 Polymorphism 1562 C > T (rs3918242)
Associated with Protection against Placental Malaria. Am. J. Trop. Med. Hyg.
93,186-188.
Arama, C., Maiga, B., Dolo, A, Kouriba, B., Traore, B., Crompton, P.D., Pierce, S.K.,
Troye-Blomberg, M., Miller, L.H., Doumbo, O.K., 2015. Ethnic differences in
susceptibility to malaria: What have we learned from immuno-epidemiological
studies in West Africa? Acta Trop. 146, 152-156.
Arese, P., Pantaleo, A., Turrini, F., 2015. Protection from Malaria by
Hemoglobinopathies. In: Encyclopedia of Malaria. Springer, New York, NY,
pp.I-18.
Arnold, M., Ramer, 1., Pfeufer, A., Suhre, K., KastenmOller, 0., 2015. SNiPA: an
interactive, genetic variant-centered annotation browser. Bioinformatics 31,
1334-1336.
Asahara, T., Kawamoto, A., Masuda, H., 2011. Concise Review: Circulating
Endothelial Progenitor Cells for Vascular Medicine. STEM CELLS 29, 1650-
1655.
160
University of Ghana http://ugspace.ug.edu.gh

Asahara, T .• Masuda. H .• Takahashi. T., Kalka, C., Pastore, C .• Silver, M., Keame. M.,
Magner, M., Isner, I.M., 1999. Bone marrow origin of endothelial progenitor
cells responsible for postnatal vasculogenesis in physiological and pathological
neovascularization. Cire. Res. 85,221-228.
Asahara, T.. Murohara, T., Sullivan, A., Silver, M., van der Zee, R., Li, T.,
Witzenbichler, B., Schatteman, G., Isner, I.M .• 1997. Isolation of putative
progenitor endothelial cells for angiogenesis. Science 275,964-967.
Asante, D.F.A., Asenso-Okyere, K., 2003. Economic Burden of Malaria in Ghana 81.
Avogaro. A., Albiero, M., Menegazzo, L., Kreutzenberg, S. dc, Fadini, G.P .• 2011.
Endothelial Dysfunction in Diabetes: The role of reparatory mechanisms.
Diabetes Care 34, S285-S290.
Ayi, K., Min-Oo. G., Serghides, L., Crockett, M., Kirby-Allen, M., Quirt, I., Gros, P.,
Kain, K.C., 2008. Pyruvate Kinase Deficiency and Malaria. N. Engt. J. Med.
358,1805-1810.
Baird, K., 2015. Origins and implications of neglect of G6PD deficiency and
primaquine toxicity in Plasmodium vivax malaria. Pathog. Glob. Health 109,
93-106.
Bakmiwewa, S.M., Weiser, S .• Grey, M., Heng, B., Guillemin, G.l., Ball, H.J., Hunt,
N.H., 2016. Synergistic induction of CXCLlO by interferon-gamma and
lymphotoxin-alpha in astrocytes: Possible role in cerebral malaria. Cytokine 78,
79-86.
Barenboim, M., Manke, T., 2013. ChroMoS: an integrated web tool for SNP
classification, prioritization and functional interpretation. Bioinformatics 29,
2197-2198.
Barenboim, M., Zoltick, 8.1., Guo, Y., Weinberger, D.R .• 2010. MicroSNiPer: a web
tool for prediction of SNP effects on putative microRNA targets. Hum. Mutat.
31,1223-1232.
Beaglehole, R., Bonita, R., 2010. What is global health? Glob. Health Action 3.
Beattie, L., Engwerda, C.R., Wykes, M., Good, M.F., 2006. CD8+ T lymphocyte
mediated loss of marginal metallophilic macrophages following infection with
Plasmodium chabaudi chabaudi AS. l.lmmunol. Baltim. Md 1950 177.2518-
2526.
161
University of Ghana http://ugspace.ug.edu.gh

Bell, J.T., Pai, AA, Pickrell, J.K., Gaffney, 0.1., Pique-Regi, R., Degner, J.F., Gilad.
Y., Pritchard, J.K., 2011. DNA methylation patterns associate with genetic and
gene expression variation in HapMap cell lines. Genome BioI. 12, RIO.
Berendt. A.R., Ferguson, D.J., Gardner, J., Turner, G., Rowe, A., McConnick, C.,
Roberts, D., Craig, A., Pinches, R., Elford, B.C., 1994. Molecular mechanisms
of sequestration in malaria. Parasitology 108 Suppl, S 19-28.
Berger, S.L., Kouzarides, T., Shiekhattar, R" Shilatifard, A., 2009, An operational
definition ofepigenetics. Genes Dev. 23, 781-783.
Bergmark, B., Bergmark, R., Beaudrap, P,D., Boum, Y., Mwanga-Amumpaire, J.,
Carroll, R., Zapol, W., 2012. Inhaled nitric oxide and cerebral malaria: basis of
a strategy for buying time for phannacotherapy. Pediatr.lnfect. Dis. J. 31, e250-
254.
Bernabeu, M., Smith, J.D., 2017. EPCR and Malaria Severity: The Center ofa Perfect
Storm. Trends Parasitol. 33,295-308.
Bettcher, D., Lee, K., 2002. Globalisation and public health. J. Epidemiol. Community
Health 56, 8-17.
Bhanot. P .• Schauer, K., Coppens, I., Nussenzweig, V .• 2005. A surface phospholipase
is involved in the migration of plasmodium sporozoites through cells. J. BioI.
Chern. 280, 6752-6760.
Bhatt, S., Gething, P., 2014. Insecticide-treated nets (ITNs) in Africa 2000-2016:
coverage, system efficiency and future needs for achieving international targets.
Malar. J. 13,029.
Blackledge, N.P., Klose, R., 2011. CpO island chromatin: a platform for gene
regulation. Epigenetics 6, 147-152.
Blanton. S.H., Henry, R.R., Yuan, Q., Mulliken, J.B., Stal, S., Finnell, R.H., Hecht,
J.T., 2011. Folate pathway and nonsyndromic cleft lip and palate., The Folate
Pathway and Nonsyndromic Cleft Lip and Palate. Birth Defects Res. Part Clin.
Mol. Teratol. Birth Defects Res. Part Clin. Mol. Teratol. 91, 91, 50, 50-60.
Boddey, J.A, Cowman, A.F., 2013. Plasmodium nesting: remaking the erythrocyte
from the inside out. Annu. Rev. Microbiol. 67, 243-269.
Boehme, M.W., Werle, E., Kommerell, B., Raeth, U., 1994. Serum levels of adhesion
molecules and thrombomodulin as indicators of vascular injury in severe
Plasmodium falciparum malaria. Clin. Investig. 72, 598-603.
162
University of Ghana http://ugspace.ug.edu.gh

Boufenzer. A., Sennoun. N., Bouazza, Y., Derive, M., BoUaert, P., Gibot, S .• 2012.
Role of TREM-I in endothelial dysfunction during experimental sepsis. Crit.
Care 16, P49.
Bouharoun-Tayoun, H., Attanath, P., Sabchareon, A .• Chongsuphajaisiddhi, T.,
Druilhe, P., 1990. Antibodies that protect humans against Plasmodium
falciparum blood stages do not on their own inhibit parasite groMil and invasion
in vitro, but act in cooperation with monocytes. J. Exp. Med. 172. 1633-1641.
Bouveyron, C., Girard, S .. Schmid, C., 2007. High-dimensional data clustering.
Comput. Stat. Data Anal. 52, 502-519.
Bouyou-Akotet, M.K., Offouga, C.L., Mawili-Mboumba. D.P.. Essola, L..
Madoungou, B .• Kombila, M., 2014. Falciparum Malaria as an Emerging Cause
of Fever in Adults Living in Gabon, Central Africa [WWW Document).
BioMed Res. Int. URL https:llwww.hindawi.com/joumals/bmril2014/3512811
(accessed 2.28.18).
Boyle. MJ., Jagannathan, P., Bowen, K., McIntyre, T.I., Vance, H.M., Farrington.
L.A., Greenhouse, B., Nankya, F., Rek, J., Katureebe, A, Arinaitwe, E., Dorsey,
G., Kamya. M.R., Feeney, M.E., 2015a. Effector Phenotype of Plasmodium
falciparum-Specific CD4+ T Cells Is Influenced by Both Age and Transmission
Intensity in Naturally Exposed Populations. J. Infect. Dis. 212,416-425.
Boyle, M.J., Reiling, L., Feng, G., Langer, C., Osier, F.H., Aspeling-Jones, H., Cheng,
Y.S., Stubbs, J., Tetteh, K.K.A, Conway, D.J., McCarthy, J.S., Muller, I..
Marsh, K., Anders, R.F., Beeson, J.G., 2015b. Human antibodies fix
complement to inhibit Plasmodium falciparum invasion of erythrocytes and are
associated with protection against malaria. Immunity 42, 580--590.
Braga, E.M., Scopel, K.K.G., Komatsu, N.T., daSilva-Nunes, M., Ferreira, M.U., 2005.
Polymorphism of the Fey receptor IIA and malaria morbidity. J. Mol. Genet.
Med. Int. J. Biomed. Res. 1,5-10.
Bryceson, AD., Fleming, A.F., Edington, G.M., 1976. Splenomegaly in Northern
Nigeria. Acta Trop. 33, 185-214.
Budczies, J., Klauschen, F., Sinn, B.V., Gy6rffy, B., Schmitt, W.O., Darb-Esfahani, S.,
Denkert, C., 2012. Cutoff Finder: A Comprehensive and Straightforward Web
Application Enabling Rapid Biomarker Cutoff Optimization. PLoS ONE 7,
e51862.
163
University of Ghana http://ugspace.ug.edu.gh

BujiJa, I., Schwarzer, E., Skorokhod, 0., Weidner, J.M., Troye-Blomberg, M., Ostlund
Farrants, A.-K., 2016. Malaria-derived hemozoin exerts early modulatory
effects on the phenotype and maturation of human dendritic cells. Cell.
Microbiol. 18,413-423.
Burda. P.-C., CaldeJari, R., HeussJer, V.T., 2017. Manipulation of the Host Cell
Membrane during Plasmodium Liver Stage Egress. mBio 8.
Campos, F.M.F., Franklin, B.S., Teixeira-Carvalho, A., Filho, A.L.S., de Paula, S.C.O.,
Fontes, C.J., Brito, C.F., Carvalho, L.H., 2010. Augmented plasma
microparticles during acute Plasmodium vivax infection. Malar. J. 9, 327.
Canavese, M., Spaccapelo, R., 2014. Protective or pathogenic effects of vascular
endothelial growth factor (VEGF) as potential biomarker in cerebral malaria.
Pathog. Glob. Health 108,67-75.
Cappadoro, M., Giribaldi, G., O'Brien, E., Turrini, F., Mannu, F., Ulliers, D., Simula,
G., Luzzatto, L., Arese, P., 1998. Early Phagocytosis of Glucose-6-Phosphate
Dehydrogenase (G6PD)-Deficient Erythrocytes Parasitized by Plasmodium
falciparwn May Explain Malaria Protection in G6PD Deficiency. Blood 92,
2527-2534.
Carneiro. l.. Roca-Feltrer, A., Griffin, J.T., Smith, L., Tanner, M., Schellenberg, J.A .•
Greenwood, B., Schellenberg, D., 2010. Age-Patterns of Malaria Vary with
Severity, Transmission Intensity and Seasonality in Sub-Saharan Africa: A
Systematic Review and Pooled Analysis. PLOS ONE 5, e8988.
Carter, R., Mendis, K.N., 2002. Evolutionary and historical aspects of the burden of
malaria. Clin. Microbiol. Rev. 15,564-594.
Ceesay, S.J., Casals-Pascual, C., Erskine, J., Anya, S.E., Duah, N.O., Fulford. A.J.,
Sesay, S.S., Abubakar, I., Dunyo, S., Sey, 0., Palmer, A., Fofana, M., Corrah,
T., Bojang, K.A., Whittle, H.C., Greenwood, B.M., Conway, OJ., 2008.
Changes in malaria indices between 1999 and 2007 in The Gambia: a
retrospective analysis. The Lancet 372, 1545-1554.
Chamnanchanunt, S., Fucharoen, S., Umemura, T., 2017. Circulating microRNAs in
malaria infection: bench to bedside. Malar. J. 16.
Cheesbrough, M., 1984. Medical laboratory manual for tropical countries Vol. II,
Vol.lI,. M. Cheesbrough, Doddington (14 Bevills Close, Doddington, Cambs.
PEl5 OTT).
164
University of Ghana http://ugspace.ug.edu.gh

Chen, L., Zhang, Z.·H., Sendo, F., 2000. Neutrophils playa critical role in the
pathogenesis of experimental cerebral malaria. Clin. Exp. Immunol. 120. 125-
133.
Cherif, M .• Amoako·Sakyi. D., Dolo, A., Pearson, J.·O., Gyan, B., Obiri· Yeboah, D.,
Nebie, I., Sirima, S.B., Doumbo, 0., Troye·Blomberg, M., Bakary, M., 2016.
Distribution of FcyR gene polymorphisms among two sympatric populations in
Mali: differing allele frequencies, associations with malariometric indices and
implications for genetic susceptibility to malaria. Malar. J. IS.
Cholera. R., Brittain, N.J., Gillrie, M.R., Lopera.Mesa, T.M., Diakite, S.AS., Arie, T.,
Krause, M.A., Guindo, A., Tubman, A., Flijioka, H., Diallo, D.A, Doumbo,
O.K., Ho, M., Wellems, T.E., Fairhurst, R.M., 2008. Impaired cytoadherence of
Plasmodium falciparum-infected erythrocytes containing sickle hemoglobin.
Proc. Nat!. Acad. Sci. U. S. A 105,991-996.
Clark, l.A., Alleva, L.M., 2009. Is human malarial coma caused, or merely deepened,
by sequestration? Trends Parasitol. 25, 314-318.
Clark, I.A, Budd, A.C., Alleva, L.M., Cowden, W.B., 2006. Human malarial disease:
a consequence of inflammatory cytokine release. Malar. J. 5, 85.
Clark. I.A, Rockett, K.A., 1994. The cytokine theory of human cerebral malaria.
Parasitol. Today Pers. Ed 10,410-412.
Clark, T.G., Diakite, M., Auburn, S., Campino, S., Fry, A.E., Green, A., Richardson,
A., Small, K., Teo, Y.Y., Wilson, 1., Jallow, M., Sisay·Joof, F., Pinder, M.,
Griffiths, M., Peshu, N., Williams, T.N., Marsh, K., Molyneux, M.E., Taylor,
T.E., Kirk A., R., Kwiatkoski, D.P., 2009. Tumor necrosis factor and
Iymphotoxin-a polymorphisms and severe malaria in African populations. J.
Infect. Dis. 199, 569-575.
Coban, C., Ishii, K.J., Kawai, T., Hemmi, H., Sato, S., Uematsu, S., Yamamoto, M.,
Takeuchi, 0., Itagaki, S., Kumar, N., Horii, T., Akira, S., 2005. Toll-like
receptor 9 mediates innate immune activation by the malaria pigment hemozoin.
J. Exp. Med. 201, 19-25.
Cohen, S., McGregor, l.A, Carrington, S., 1961. Gamma-globulin and acquired
immunity to human malaria Nature 192,733-737.
165
University of Ghana http://ugspace.ug.edu.gh

Combes. V., EI-Assaad, F., Faille, D., Jambou, R., Hunt, N.H .• Grau, G.E.R., 2010.
Microvesiculation and cell interactions at the brain-endothelial interface in
cerebral malaria pathogenesis. Prog. Neurobiol. 91, 140-151.
Conroy, A.L., Glover, S.J., Hawkes, M., Erdman, L.K., Seydel, K.B., Taylor, T.E.,
Molyneux, M.E .. Kain, K.C., 2012. Angiopoietin-2 levels are associated with
retinopathy and predict mortality in Malawian children with cerebral malaria: a
retrospective case-control study·. Crit. Care Med. 40, 952-959.
Conroy, A.L., Lafferty, E.I., Lovegrove, F.E.. Krudsood, S., Tangpukdee, N., Liles,
W.C., Kain, K.C., 2009. Whole blood angiopoietin-I and -2 levels discriminate
cerebral and severe (non-cerebral) malaria from uncomplicated malaria. Malar.
J. 8,295.
Conroy, A.L., Phiri. H., Hawkes. M., Glover, S., Mallewa, M .• Seydel, K.B., Taylor,
T.E., Molyneux, M.E., Kain, K.C., 2010. Endothelium-based biomarkers are
associated with cerebral malaria in Malawian children: a retrospective case
control study. PloS One 5, e15291.
Cooper. D.N., 2010. Functional intronic polymorphisms: Buried treasure awaiting
discovery within our genes. Hum. Genomics 4, 284-288.
Coppi, A., Tewari, R., Bishop, J.R., Bennett, B.L., Lawrence, R., Esko, J.D., Billker,
0., Sinnis, P., 2007. Heparan Sulfate Proteoglycans Provide a Signal to
Plasmodium Sporozoites to Stop Migrating and Productively Invade Cells. Cell
Host Microbe 2, 316-327.
Cannier, L.A., 2016. The Ten-Thousand Year Fever: Rethinking Human and Wild
Primate Malarias. Routledge.
Cowman. A.F., Healer, J., Marapana, D., Marsh, K., 2016. Malaria: Biology and
Disease. Cell 167,610-624.
Cox, F.E., 2010. History of the discovery of the malaria parasites and their vectors.
Parasit. Vectors 3, 5.
Crispe,I.N., 2009. The liver as a lymphoid organ. Annu. Rev. Immuno!. 27,147-163.
Crompton, P.O., Moebius, J., Portugal, S., Waisberg, M., Hart, G., Garver, L.S., Miller,
L.H., Barillas, C., Pierce, S.K., 2014. Malaria immunity in man and mosquito:
insights into unsolved mysteries of a deadly infectious disease. Annu. Rev.
Immuno!. 32, 157-187.
166
University of Ghana http://ugspace.ug.edu.gh

Cserti-Gazdewich, C.M., Dzik, W.H., Erdman. L., Ssewanyana, I., Dhabangi, A.,
Musoke, c., Kain, K.C., 2010. Combined measurement of soluble and cellular
ICAM-I among children with Plasmodium falciparum malaria in Uganda.
Malar. J. 9,233.
Cserti-Gazdewich, C.M., Mayr. W.R., Dzik. W.H., 2011. Plasmodium falciparum
malaria and the immunogenetics of ABO, HLA. and CD36 (platelet
glycoprotein IV). Vox Sang. 100,99-111.
Cunnington, A.1., Riley, E.M., Walther, M., 2013. Stuck in a rut? Reconsidering the
role of parasite sequestration in severe malaria syndromes. Trends Paras ito!' 29,
585-592.
Dalaba, M.A., Akweongo, P., Aborigo, R., Awine, T., Azongo, D.K., Asaana, P.,
Atuguba. F., Oduro, A.. 2014. Does the national health insurance scheme in
Ghana reduce household cost of treating malaria in the Kassena-Nankana
districts? Glob. Health Action 7, 23848.
Daou, M., Kituma, E., Kavishe, R., Chilongola. J., Mosha, F .• van der Ven, A., Kouriba,
B., Bousema, T., Sauerwein, R., Doumbo, 0., 2015. a-Thalassaemia trait is
associated with Antibody prevalence against Malaria Antigens AMA-I and
MSP-1. J. Trop. Pediatr. 61, 139-142.
Des. S .• Hertrich, N., Perrin, A.1., Withers-Martinez, C., Collins, C.R., Jones, M.L.,
Watermeyer, J.M., Fobes, E.T., Martin, S.R., Saibil, H.R., Wright, G.J., Treeck,
M .• Epp, C., Blackman, M.J., 2015. Processing of Plasmodium falciparum
Merozoite Surface Protein MSPI Activates a Spectrin-Binding Function
Enabling Parasite Egress from RBCs. Cell Host Microbe 18. 433-444.
Davies. D.H., Duffy, P., Bodmer, J.-L., FeIgner, P.L., Doolan, D.L., 2015. Large screen
approaches to identify novel malaria vaccine candidates. Vaccine 33, 7496-
7505.
Dayeh, I.A., Olsson, A.H., Volkov, P., Almgren, P., ROnn, T., Ling, C., 2013a.
Identification of CpG-SNPs associated with type 2 diabetes and differential
DNA methylation in human pancreatic islets. Diabetologia 56, 1036-1046.
Dayeh, I.A., Olsson, A.H., Volkov, P., Almgren, P., ROnn, T., Ling, C., 2013b.
Identification of CpG-SNPs associated with type 2 diabetes and differential
DNA methylation in human pancreatic islets. Diabetologia 56, 1036-1046.
167
University of Ghana http://ugspace.ug.edu.gh

DeCoster. J., Gallucci, M., Iselin. A.-M., n.d. Best Practices for Using Median Splits,
Artificial Categorization, and their Continuous Alternatives. J. Exp.
Psychopathol. 2, 197-209.
Deng, Y., Chang, C., Ido, M.S., Long, Q., 2016. Multiple Imputation for General
Missing Data Patterns in the Presence of High-dimensional Data. Sci. Rep. 6,
21689.
Derbyshire, E.R., Prudencio, M., Mota, M.M., Clardy, J., 2012. Liver-stage malaria
parasites vulnerable to diverse chemical scaffolds. Proc. Natl. Acad. Sci. U. S.
A. 109,8511-8516.
Desruisseaux, M.S., Machado, F.S., Weiss, L.M., Tanowitz, H.B., Golightly, L.M.,
2010. Cerebral Malaria. Am. J. Pathol. 176, 1075-1078.
Dhangadamajhi, G., Kar, A., Rout, R., Dhangadamajhi, P., Dhangadamajhi, G., Kar,
A., Rout, R., Dhangadamajhi, P., 2017. A meta-analysis of TLR4 and TLR9
SNPs implicated in severe malaria. Rev. Soc. Bras. Med. Trop. 50, 153-160.
Dickinson-Copeland, C.M., Wilson, N.O., Liu, M., Driss, A., Salifu, H., Adjei, A.A.,
Wilson, M., Gyan, B., Oduro, D., Badu, K., Botchway, F., Anderson, W., Bond,
V., Bacanamwo, M., Singh, S., Stiles, J.K., 2015. Heme-Mediated Induction of
CXCLlO and Depletion of CD34+ Progenitor Cells Is Toll-Like Receptor 4
Dependent. PLOS ONE 10, e0142328.
Dodoo, D., Theander, T.G., Kurtzhals, J.A.L., Koram, K., Riley, E., Akanmori, B.D.,
Nkrumah, F.K., Hviid, L., 1999. Levels of Antibody to Conserved Parts of
Plasmodium falciparum Merozoite Surface Protein 1 in Ghanaian Children Are
Not Associated with Protection from Clinical Malaria. Infect. Immun. 67,
2131-2137.
Dondorp. A.M., Desakorn, V., Pongtavornpinyo, W., Sahassananda, D., Silamut, K.,
Chotivanich, K., Newton, P.N., Pitisuttithum, P., Smithyman, A.M., White.
N.J., Day, N.P.J., 2005. Estimation of the Total Parasite Biomass in Acute
Falciparum Malaria from Plasma PfHRP2. PLOS Med. 2, e204.
Dorovini-Zis, K., Schmidt, K., Huynh, H., Fu, W., Whitten, R.O., Milner, D., Kamiza,
S., Molyneux, M., Taylor, T.E., 2011. The neuropathology of fatal cerebral
malaria in maJawian children. Am. J. Pathol. 178,2146-2158.
Dunst. J., Kamena, F., Matuschewski, K., 2017. Cytokines and Chemokines in Cerebral
Malaria Pathogenesis. Front. Cell. Infect. Microbiol. 7.
168
University of Ghana http://ugspace.ug.edu.gh

Dunst, J.. Pigorsch, S., H!nsgen, G., Hintner, I., Lautenschillger, C., Becker, A., 1999.
Low hemoglobin is associated with increased serum levels of vascular
endothelial growth factor (VEGF) in cancer patients. Does anemia stimulate
angiogenesis? Strahlenther. Onkol. Organ Otsch. Rontgengesellschaft AI 175,
93-96.
Dzau, V.J., Gnecchi, M., Pachori, A.S., Morello, F., Melo, L.G., 2005. Therapeutic
Potential of Endothelial Progenitor Cells in Cardiovascular Diseases.
Hypertension 46,7-18.
Edstein, M.D., Kotecka, B.M., Anderson, K.L., Pombo, OJ., Kyle, D.E., Rieckmann,
K.H., Good, M.F .• 2005. Lengthy antimalarial activity ofatovaquone in human
plasma following atovaquone-proguanil administration. Antimicrob. Agents
Chemother. 49, 4421~422.
Edwards, D.R.V., Romero, R., Kusanovic, J.P., Hassan, S.S., Mazaki-Tovi, S.,
Vaisbuch, E., Kim. C.J., Erez, 0., Chaiworapongsa, T., Pearce, B.D., Bartlett,
J .. Friel, L.A., Salisbury, B.A., Anant, M.K., Vovis, G.F., Lee, M.S., Gomez,
R.. Behnke, E., Oyarzun, E., Tromp, G., Menon, R., Williams, S.M., 2011.
Polymorphisms in maternal and fetal genes encoding for proteins involved in
extracellular matrix metabolism alter the risk for small-for-gestational-age. J.
Matern.-Fetal Neonatal Med. Off. J. Eur. Assoc. Perinat. Med. Fed. Asia Ocean.
Perinat. Soc. Int. Soc. Perinat. Obstet. 24, 362-3S0.
li,.,de. T.P., Larsen, D., Steketee, R.W., 2010. Protective efficacy of interventions for
preventing malaria mortality in children in Plasmodium falciparum endemic
areas. Int. J. Epidemiol. 39 Suppl 1, iSS-lO 1.
Erdman, L.K., Dhabangi, A., Musoke, C., Conroy, A.L., Hawkes, M., Higgins, S.,
Rajwans, N., Wolofsky, K.T., Streiner, D.L., Liles, W.C., Cserti-Gazdewich,
C.M., Kain, K.C., 2011. Combinations of host biomarkers predict mortality
among Ugandan children with severe malaria: a retrospective case-control
study. PloS One 6, e 17440.
Esser, C., Bachmann, A., Kuhn, D., Schuldt, K., FOrster, B., Thiel, M., May, J., Koch
Nolte, F., Yanez-Mo, M., Sanchez-Madrid, F., Schinkel, A.H., Jalkanen, S.,
Craig, A.G., Bruchhaus, I., Horstmann, R.D., 2014. Evidence of promiscuous
endothelial binding by Plasmodium falciparum-infected erythrocytes. Cell.
Microbiol. 16, 70 I-70S.
169
University of Ghana http://ugspace.ug.edu.gh

Evans, S.S., Repasky, E.A., Fisher, D.T., 2015. Fever and the thennal regulation of
immunity: the immune system feels the heat. Nat. Rev. Immunol. IS, 335-349.
Farner!, A., Yman, V., Vafa Homann, M., Wandell, G., Mhoja, L., Johansson, M.,
Jesaja, S., Sandlund, J., Tanabe, K., Hammar, U., Bottai, M., Premji, Z.G.,
BjOrkman, A., Rooth, I., 2014. Epidemiology of malaria in a village in the Rufiji
River Delta, Tanzania: declining transmission over 25 years revealed by
different parasitological metrics. Malar. J. 13,459.
Faruque, M.U., Chen, G., Doumatey, A., Huang, H., Zhou. J., Dunston, G.M .• Rotimi,
C.N., Adeyemo, A.A., 2011. Association of ATPIBI, RGS5 and SELE
polymorphisms with hypertension and blood pressure in African-Americans. J.
Hypertens.29,1906-1912.
Fleming, A.F., Storey, J., Molineaux, L., Iroko, E.A., Attai, E.D., 1979. Abnonnal
haemoglobins in the Sudan savanna of Nigeria. I. Prevalence of haemoglobins
and relationships between sickle cell trait, malaria and survival. Ann. Trop.
Med. Parasitol. 73.161-172.
Fowkes, F.J.I.. Richards, J.S., Simpson, J.A., Beeson, J.G., 2010. The relationship
between anti-merozoite antibodies and incidence of Plasmodium falciparum
malaria: A systematic review and meta-analysis. PLoS Med. 7, e I 000218.
Furini. A.A. da c., Cassiano, G.C., Petrolini Capobianco, M., dos Santos, S.E.B.,
Dantas Machado, R.L., 2016. Frequency of TNFA, INFG, and ILlO Gene
Polymorphisms and Their Association with Malaria Vivax and Genomic
Ancestry. Mediators Inflamm. 2016.
Gallup, J.L., Sachs, J.D., 2001. The economic burden of malaria. Am. J. Trop. Med.
Hyg. 64, 85-96.
Gao, X .. Gunalan, K., Yap, S.S.L., Preiser, P.R., 2013. Triggers of key calcium signals
during erythrocyte invasion by Plasmodium falciparum. Nat. Commun. 4, 2862.
Gardner, M.1., Hall, N., Fung, E., White, 0., Berriman, M., Hyman, R.W., Carlton,
J.M., Pain, A., Nelson, K.E., Bowman, S., Paulsen, l.T., James, K., Eisen, J.A.,
Rutherford, K., Salzberg, S.L., Craig, A., Kyes, S., Chan, M.-S., Nene, V.,
Shallom, S.1., Sub, B., Peterson, J., Angiuoli, S., Pertea, M., Allen, J., Selengut.
J., Haft, D., Mather, M.W., Vaidya, A.B., Martin, D.M.A., Fairlamb, A.H.,
Fraunholz. M.1., Roos, D.S., Ralph, S.A., McFadden, G.I., Cummings, L.M.,
Subramanian, G.M., Mungall. C., Venter, J.C., Carucci, D.J., Hoffman, S.L .•
170
University of Ghana http://ugspace.ug.edu.gh

Newbold, C., Davis, R. W., Fraser, C.M., Barrell, B., 2002a. Genome sequence
of the human malaria parasite Plasmodium falciparum. Nature 4 I 9, 498-511.
Gardner. M.J., Hall, N., Fung, E., White, 0., Beniman, M., Hyman, R. W., Carlton,
J.M., Pain, A., Nelson, K.E., Bowman, S., Paulsen, I.T., James, K., Eisen. J.A.,
Rutherford, K., Salzberg, S.L., Craig, A., Kyes, S., Chan. M.-S., Nene, V.,
Shallom, S.J., Suh, B., Peterson, J., Angiuoli, S., Pertea, M., Allen, J., Selengut.
J., Haft, D., Mather, M.W., Vaidya, AB., Martin, D.M.A. Fairlamb. A.H .•
Fraunholz, M.J., Roos, D.S., Ralph, S.A, McFadden, G.I., Cummings, L.M.,
Subramanian, G.M., Mungall, C., Venter. J.C., Carucci, DJ., Hoffman. S.L .•
Newbold, C., Davis, R. W., Fraser. C.M., Barrell, B., 2002b. Genome sequence
of the human malaria parasite Plasmodium falciparum. Nature 419, 498-511.
Ghazanfari, N., Mueller. S.N .• Heath, W.R., 2018. Cerebral Malaria in Mouse and Man.
Front. Immunol. 9.
Giha, H.A .• A-Elbasit, I.E., A-Elgadir. T.M.E., Adam. I., Berzins. K., Elghazali, G.,
Elbashir, M.I .• 2005. Cerebral malaria is frequently associated with latent
parasitemia among the semi-immune population of eastern Sudan. Microbes
Infect. 7,1196-1203.
Goeij, D., C.m. M .• van Diepen, M., Jager, KJ., Tripepi. G., Zoccali, C., Dekker, F. W.,
2013. Multiple imputation: dealing with missing data. Nephrol. Dial.
Transplant. 28, 2415-2420.
Graewe, S., Rankin, K.E., Lehmann, C., Deschermeier, c., Hecht, L., Froehlke, U.,
Stanway, R.R., Heussler, V., 2011. Hostile takeover by Plasmodium:
reorganization of parasite and host cell membranes during liver stage egress.
PLoS Pathog. 7, el002224.
Graham. S.M .• Chen, J., Chung, D.W., Barker. K.R., Conroy, AL., Hawkes, M.T.,
Namasopo, S., Kain, K.C., L6pez, J.A., Liles, W.C .• 20]6. Endothelial
activation, haemostasis and thrombosis biomarkers in Ugandan children with
severe malaria participating in a clinical trial. Malar. J. ]5,56.
Greene, J.A., Sam-Agudu, N., John, C.C., Opoka, R.O., Zimmerman, P.A., Kazura,
J.W., 2012. Toll-like receptor polymorphisms and cerebral malaria: TLR2 ~2
polymorphism is associated with protection from cerebral malaria in a case
control study. Malar. J. 11.47.
171
University of Ghana http://ugspace.ug.edu.gh

Greenwood. B .• Marsh, K., Snow, R., 1991. Why do some African children develop
severe malaria? Parasitol. Today Pers. Ed 7, 277-281.
Guillemin, J., 2002. Choosing Scientific Patrimony: Sir Ronald Ross, Alphonse
Laveran, and the Mosquito-Vector Hypothesis for Malaria. J. Hist. Med. Allied
Sci. 57,385-409.
Gupta, H., Chaudhari, S., Rai, A., Bhat, S., Sahu, P.K.. Hande, M.H.. D'Souza. S.C .•
Shashikiran, U., Satyamoorthy, K., 2017. Genetic and epigenetic changes in
host ABCB 1 influences malaria susceptibility to Plasmodium falciparum. PLoS
ONE 12.
Gupta, S .• Seydel. K .• Miranda-Roman. M.A., Feintuch, C.M., Saidi, A., Kim, R.S.,
Birbeck. G.L.. Taylor. T.E., Daily, J.P .• 2017. Extensive alterations of blood
metabolites in pediatric cerebral malaria. PLOS ONE 12, e0175686.
Gyan, B., Goka, B.Q .• Adjei, G.O., Tetteh, J.K.A., Kusi, K.A., Aikins, A., Dodoo. D.,
Lesser, M.L.. Sison, C.P .• Das, S., Howard, M.E .• Milbank, E., Fischer, K.,
Rafli, S., Jin, D .• Golightly, L.M .. 2009a. Cerebral malaria is associated with
low levels of circulating endothelial progenitor cells in African children. Am. J.
Trop. Med. Hyg. 80,541-546.
Gyan. 8.. Goka, B.Q., Adjei, G.O., Tetteh, J.K.A., Kusi, K.A., Aikins, A., Dodoo, D .•
Lesser, M.L., Sison, C.P .• Das, S., Howard, M.E., Milbank, E., Fischer, K.,
Rafii, S .• Jin, D., Golightly, L.M., 2009b. Cerebral Malaria Is Associated with
Low Levels of Circulating Endothelial Progenitor Cells in African Children.
Am. J. Trop. Med. Hyg. 80, 541-546.
Gyan, B.A., Goka, B., Cvetkovic, J.T., Kurtzhals, J.L., Adabayeri, V., Perl mann, H.,
Lefvert, A.-K., Akanmori. B.D., Troye-Blomberg, M., 2004. Allelic
polymorphisms in the repeat and promoter regions of the interleukin-4 gene and
malaria severity in Ghanaian children. Clin. Exp. Immunol. 138, 145-150.
Hadley, T.J., Miller, L.H., 1988. Invasion of Erythrocytes by Malaria Parasites:
Erythrocyte Ligands and Parasite Receptors. Malar. Immunol. 41. 49-71.
Hafalla, J.C.R., Cockburn, LA., Zavala, F., 2006. Protective and pathogenic roles of
CD8+ T cells during malaria infection. Parasite Immunol. 28, 15-24.
Hamik, A., Wang, B., Jain, M.K., 2006a. Transcriptional Regulators of Angiogenesis.
Arterioscler. Thromb. Vase. BioI. 26,1936-1947.
172
University of Ghana http://ugspace.ug.edu.gh

Hamik. A., Wang. B., Jain, M.K., 2006b. Transcriptional Regulators of Angiogenesis.
Arterioscler. Thromb. Vase. BioI. 26, 1936-1947.
Handel. T.M., Horuk, R., 2010. Duffy antigen inhibitors: useful therapeutics for
malaria? Trends Parasitol. 26,329-333.
Hansen, D.S., Bernard, N.J., Nie, C.Q., Schofield, L., 2007. NK Cells Stimulate
Recruitment of CXCR3+ T Cells to the Brain during Plasmodium berghei
Mediated Cerebral Malaria. J. Immunol. 178,5779-5788.
Hanson, J., Dondorp, A.M., Day, N.P., White, N.J., 2013. Reply to Cunnington et aI. J.
Infect. Dis. 207, 370-371.
Haq. I., Chappell, S .. Johnson, S.R., Lotya, J., Daly, L., Morgan, K., Guetta-Baranes,
T., Roca, J., Rabinovich, R., Millar, A.B., Donnelly, S.C., Keatings, V.,
MacNee, W., Stolk, J., Hiemstra, P.S., Miniati, M., Monti, S., O'Connor, C.M.,
Kalsheker, N., 2010. Association of MMP-2 polymorphisms with severe and
very severe COPD: a case control study of MMPs-I, 9 and 12 in a European
population. BMC Med. Genet. 11,7.
Hemingway, J., Shrelta, R., Wells, T.N.C., Bell, D .• Djimde, A.A., Achee, N., Qi, G.,
2016. Tools and Strategies for Malaria Control and Elimination: What Do We
Need to Achieve a Grand Convergence in Malaria? PLOS BioI. 14, el002380.
Hendriksen, I.C.E., Ferro, J., Montoya, P., Chhaganlal, K.D., Seni, A., Gomes, E.,
Silamut, K., Lee, S.J., Lucas, M., Chotivanich, K., Fanello, C.I., Day, N.P.J.,
White, N.J., von Seidlein, L., Dondorp, A.M., 2012. Diagnosis, Clinical
Presentation, and In-Hospital Mortality of Severe Malaria in HIV -Coinfected
Children and Adults in Mozambique. Clin. Infect. Dis. Off. Publ. Infect. Dis.
Soc. Am. 55, 1144-1153.
Herrera, R., Anderson, C., Kumar, K., Molina-Cruz, A., Nguyen, V., Burkhardt, M.,
Reiter, K., Shimp, R., Howard, R.F., Srinivasan, P., Nold, M.J., Ragheb, D.,
Shi, L., DeCotiis, M., Aebig, J., Lambert, L., Rausch, K.M., Muratova, 0., Jin,
A., Reed, S.G., Sinnis, P., Barillas-Mury, C., Duffy, P.E., MacDonald, N.J.,
Narum, D.L., 2015. Reversible Conformational Change in the Plasmodium
falciparum Circumsporozoite Protein Masks Its Adhesion Domains. Infect.
Immun. 83,3771-3780.
Hill, A.V., 1999. The Immunogenetics of Resistance to Malaria. Proc. Assoc. Am.
Physicians I 11,272-277.
173
University of Ghana http://ugspace.ug.edu.gh

Hill, A.V., Jepson, A., Plebanski, M., Gilbert, S.C., 1997. Genetic analysis of host
parasite coevolution in human malaria. Philos. Trans. R. Soc. Lond. B. BioI.
Sci. 352,1317-1325.
Hirunpetcharat, C., Vukovic, P., Liu, X.Q., Kaslow, D.C., Miller, L.H., Good, M.F.,
1999. Absolute requirement for an active immune response involving B cells
and Th cells in immunity to Plasmodium yoelii passively acquired with
antibodies to the 19-kDa carboxyl-terminal fragment of merozoite surface
protein-I. J.lmrnunol. Baltim. Md 1950 162,7309-7314.
Hoffman. S.1., Cole, C.B., 2018. Defining the global health system and systematically
mapping its network of actors. Glob. Health 14.
Holder, A.A., 1994. Proteins on the surface of the malaria parasite and cell invasion.
Parasitology 108 Suppl, S5-18.
Hollestelle, M.1., Donkor, C., Mantey, E.A., Chakravorty, S.J., Craig, A., Akoto, A.O.,
O'Donnell, J., van Mourik, J.A., Bunn, J., 2006. von Wille brand factor
propeptide in malaria: evidence of acute endothelial cell activation. Br. J.
Haematol. 133, 562-569.
Honda. T., Miyachi, Y., Kabashima, K., 2011. Regulatory T cells in cutaneous immune
responses. J. Dermatol. Sci. 63, 75-82.
Hong. E.P., Park, J.W., 2012. Sample Size and Statistical Power Calculation in Genetic
Association Studies. Genomics Inform. 10, 117-122.
Hristov, M., Erl, W., Weber, P.C., 2003. Endothelial Progenitor Cells Mobilization,
Differentiation, and Homing. Arterioscler. Thromb. Vasco BioI. 23, 1185-1189.
Hsieh, Y.-C., Hsieh, F.-I., Chen, Y.-R., Hu, c.-J., Jeng, J.-S., Tang, S.-C., Chi, N.-F.,
Lin, H.-J., Lien, L.-M., Peng, G.-S., Chiou, H.-Y., Formosa Stroke Genetic
Consortium (FSGC), 2017. Associations of estradiol levels and genetic
polymorphisms of inflammatory genes with the risk of ischemic stroke. J.
Biomed. Sci. 24, 25.
Hu, Z., Bruno, A.E., 2011. The Influence of 3 'UTRs on MicroRNA Function Inferred
from Human SNP Data. Compo Funct. Genomics 2011, 910769.
Hviid, L., Jensen, A.T.R., 2015. PfEMPl - A Parasite Protein Family of Key
Importance in Plasmodium falciparum Malaria Immunity and Pathogenesis.
Adv. Parasitol. 88,51-84.
174
University of Ghana http://ugspace.ug.edu.gh

Iacobucci, D., Posavac, S.S., Kardes. F.R .. Schneider, M.J., Popovich. D.L., 2015. The
median split: Robust, refined, and revived. J. Consum. Psychol. 25, 690-704.
Idro, R., Jenkins, N.E., Newton, C.RJ.C., 2005. Pathogenesis, clinical features, and
neurological outcome of cerebral malaria. Lancet Neurol. 4, 827-840.
Idro, R., Marsh, K., John, C.C., Newton, C.R., 2010. Cerebral Malaria; Mechanisms Of
Brain Injury And Strategies For Improved Neuro-Cognitive Outcome. Pediatr.
Res. 68, 267-274.
Imai, T., Ishida, H .. Suzue, K .• Hirai, M., Taniguchi, T., Okada, H .• Suzuki, T.,
Shimokawa, C., Hisaeda, H., 2013. CD8(+) T cell activation by murine
erythroblasts infected with malaria parasites. Sci. Rep. 3, 1572.
imai, T., Shen, J., Chou, B., Duan, X., Tu, L., Tetsutani, K., Moriya, C., Ishida, H.,
Hamano, S., Shimokawa, C., Hisaeda, H., Himeno, K., 2010. Involvement of
CD8+ T cells in protective immunity against murine blood-stage infection with
Plasmodium yoelii 17XL strain. Eur. J. Immunol. 40, 1053-1061.
Ishino, T., Yano, K., Chinzei, Y., Yuda, M., 2004. Cell-passage activity is required for
the malarial parasite to cross the liver sinusoidal cell layer. PLoS BioI. 2. E4.
Jagannathan, P., Eccles-James, I., Bowen, K., Nankya, F., Auma, A., Wamala, S.,
Ebusu, C., Muhindo, M.K., Arinaitwe, E., Briggs, J., Greenhouse, B., Tappero,
J.W., Kamya, M.R., Dorsey, G., Feeney, M.E., 2014. IFN'YIIL-IO co-producing
cells dominate the CD4 response to malaria in highly exposed children. PLoS
Pathog. 10, e1003864.
Jain, V., Lucchi, N.W., Wilson, N.O., Blackstock, A.J., Nagpal, A.C., Joel, P.K., Singh,
M.P., Udhayakumar, V., Stiles, J.K., Singh, N., 2011. Plasma levels of
angiopoietin-l and -2 predict cerebral malaria outcome in Central India. Malar.
J. 10,383.
Jones. C.O.H., Williams, H.A., 2004. The Social Burden of Malaria: What Are We
Measuring? American Society of Tropical Medicine and Hygiene.
Kafsack, B.F.C., Rovira-Graells, N., Clark, T.G., Bancells, C., Crowley, V.M.,
Campino, S.G., Williams, A.E., Drought, L.G., Kwiatkowski, D.P., Baker,
D.A., Cortes, A., Lliruis, M., 2014. A transcriptional switch underlies
commitment to sexual development in human malaria parasites. Nature 507.
248-252.
175
University of Ghana http://ugspace.ug.edu.gh

Kaushansky. A., Douglass. A.N., Arang, N., Vigdorovich, V., Dambrauskas, N., Kain,
H.S., Austin, L.S., Sather, D.N., Kappe, S.H.I., 2015. Malaria parasites target
the hepatocyte receptor EphA2 for successful host infection. Science 350,
1089-1092.
Kennedy, A.T., Schmidt, C.Q., Thompson, J.K., Weiss, G.E., Taechalertpaisam, T.,
Gilson, P.R., Barlow, P.N., Crabb, B.S., Cowman, A.F., Tham, W.-H., 2016.
Recruitment of Factor H as a Novel Complement Evasion Strategy for Blood
Stage Plasmodium falciparum Infection. J. Immunol. Baltim. Md 1950 196,
1239-1248.
Kidson, c., Lamont, G., Saul, A., Nurse, G.T., 1981. Ovalocytic erythrocytes from
Melanesians are resistant to invasion by malaria parasites in culture. Proc. Natl.
Acad.Sci. 78,5829-5832.
Kikuchi, M., Looareesuwan, S., Ubalee, R., Tasanor, 0., Suzuki, F., Wattanagoon, Y.,
Na-Bangchang, K., Kimura, A., Aikawa, M., Hirayama. K., 2001. Association
of adhesion molecule PECAM-I/CD31 polymorphism with susceptibility to
cerebral malaria in Thais. Parasitol. Int. 50,235-239.
Kim, H.S., Bae, S.-C., Kim, T.-H., Kim, S.-Y., 2013. Endothelial nitric oxide synthase
gene polymorphisms and the risk of osteonecrosis of the femoral head in
systemic lupus erythematosus. Int. Orthop. 37, 2289-2296.
King, A.F.A., 1883. Insects and Disease - Mosquitoes and Malaria. In: Popular Science
Monthly Volume 23 September 1883.
King, C.L., Adams, J.H., Xianli, J., Grimberg, B.T., McHenry, A.M., Greenberg, L.J .•
Siddiqui, A., Howes, R.E., Silva-Nunes, M. da, Ferreira, M.U., Zimmennan,
P .A., 2011. FyalFyb antigen polymorphism in human erythrocyte Duffy antigen
affects susceptibility to Plasmodium vivax malaria. Proc. Natl. Acad. Sci. 108,
20113-20118.
Koita, O.A., Doumbo, O.K., Ouattara, A., Tall, L.K., Konare, A., Diakite, M., Diallo,
M., Sagara, I., Masinde, G.L., Doumbo, S.N., Dolo, A., Tounkara, A., Trao~,
I., Krogstad, D.J., 2012. False-Negative Rapid Diagnostic Tests for Malaria and
Deletion of the Histidine-Rich Repeat Region of the hrp2 Genet. Am. J. Trop.
Med. Hyg. 86, 194-198.
176
University of Ghana http://ugspace.ug.edu.gh

Koplan. J.P .• Bond, T.e.. Merson, M.H., Reddy, K.S., Rodriguez, M.H., Sewankambo,
N.K., Wasserheit, J.N., 2009. Towards a common definition of global health.
The Lancet 373,1993-1995.
Kordes, M., Matuschewski, K., Hafalla, J.C.R., 2011. Caspasc-l activation of
interleukin-l p (IL-l P) and IL-I8 is dispensable for induction of experimental
cerebral malaria. Infect. Immun. 79, 3633-3641.
Kraemer, S.M., Smith, J.D., 2003. Evidence for the importance of genetic structuring
to the structural and functional specialization of the Plasmodium falciparum Vat
gene family. Mol. Microbiol. 50, 1527-1538.
Krief, S., Escalante, A.A., Pacheco, M.A., Mugisha, L., Andre, C., Halbwax, M.,
Fischer, A., Krief, J.-M., Kasenene, J.M., Crandfield, M., Cornejo, O.E.,
Chavatte, J.-M., Lin, C., Letoumeur, F., GrOner, A.C., McCutchan, T.F., Renia,
L., Snounou, G., 2010. On the Diversity of Malaria Parasites in African Apes
and the Origin of Plasmodium falciparum from Bonobos. PLOS Pathog. 6,
el000765.
Krishnegowda. G .• Hajjar, A.M., Zhu, J., Douglass, E.J., Uematsu, S., Akira, S.,
Woods, A.S., Gowda, D.C., 2005. Induction of proinflammatory responses in
macrophages by the glycosylphosphatidylinositols of Plasmodium falciparurn:
cell signaling receptors, glycosylphosphatidylinositol (GPI) structural
requirement, and regulation ofGPI activity. J. BioI. Chern. 280, 8606-8616.
Krotoski, W.A., Collins, W.E., Bray, R.S., Garnham, P.C., Cogswell, F.B., Gwadz,
R.W., Killick-Kendrick, R., Wolf, R., Sinden, R., Koontz, L.C., Stanfill, P.S.,
1982. Demonstration of hypnozoites in sporozoite-transmitted Plasmodium
vivax infection. Am. J. Trop. Med. Hyg. 31, 1291-1293.
Kusi, K.A., Gyan, B.A., Goka, B.Q., Dodoo, D., Obeng-Adjei, G., Troye-Blomberg.
M., Akanmori, B.D., Adjimani, J.P., 2008. Levels of soluble CD163 and
severity of malaria in children in Ghana. Clin. Vaccine Immunol. CVI 15, 1456-
1460.
Kwiatkowski, D., Cannon, J.G., Manogue, K.R., Cerami, A., Dinarello, C.A.,
Greenwood, B.M., 1989. Tumour necrosis factor production in Falciparurn
malaria and its association with schizont rupture. Clin. Exp. Immunol. 77, 361-
366.
177
University of Ghana http://ugspace.ug.edu.gh

Kwiatkowski. D .• Hill. A.V., Sambou. I., Twumasi, P .• Castracane, J., Manogue, K.R.,
Cerami, A .• Brewster, D.R., Greenwood, B.M., 1990. TNF concentration in
fatal cerebral, non-fatal cerebral, and uncomplicated Plasmodium falciparum
malaria. Lancet Lond. Eng!. 336, 1201-1204.
Langhorne. J., Meding, S.J .• Eichmann, K., Gillard, S.S., 1989. The response ofCD4+
T cells to Plasmodium chabaudi chabaudi. Immunol. Rev. 112.71-94.
Lawrie. E., 1898. ON THE FLAGELLATED FORM OF THE MALARIA PARASITE.
The Lancet 151,432--434.
Lee, W.-C., 2003. Searching for disease-susceptibility loci by testing for Hardy
Weinberg disequilibrium in a gene bank of affected individuals. Am. J.
Epidemiol. 158, 397--400.
Leisch. F., 2004. FlexMix: A General Framework for Finite Mixture Models and Latent
Class Regression in R I Leisch I Journal of Statistical Software.
Lepenies, B., Cramer, J.P., Burchard, G.D., Wagner, H., Kirschning, C.J., Jacobs, T.,
2008. Induction of experimental cerebral malaria is independent of TLRl/4/9.
Med. Microbiol. Immunol. (Berl.) 197,39--44.
Lerner, H., Berg, C., 2017. A Comparison of Three Holistic Approaches to Health: One
Health. EcoHealth, and Planetary Health. Front. Vet. Sci. 4,163.
Liehl. P .• Mota, M.M., 2012. Innate recognition of malarial parasites by mammalian
hosts. Int. J. Parasito!. 42, 557-566.
Liehl. P., Zuzarte-Luis, V., Chan, J., Zillinger, T., Baptista, F., Carapau, D., Koner!,
M., Hanson, K.K., Carret, C., Lassnig, C., MUlier, M., Kalinke, U., Saeed, M.,
Chora, A.F., Golenbock, D.T., Strobl, B., Prudencio, M., Coelho, L.P., Kappe,
S.H., Superti-Furga, G., Pichlmair, A., Vigario, A.M., Rice, C.M., Fitzgerald,
K.A., Barchet, W., Mota, M.M., 2014. Host-cell sensors for Plasmodium
activate innate immunity against liver-stage infection. Nat. Med. 20, 47-53.
Liu. M., Solomon, W., Cespedes, J.C., Wilson, N.O., Ford, B., Stiles, J.K., 2018.
Neuregulin-l attenuates experimental cerebral malaria (ECM) pathogenesis by
regUlating ErbB4/AKT/STAT3 signaling. J. Neuroinflammation 15. 104.
Liu, W., Li, Y., Learn, G.H., Rudicell, R.S., Robertson, J.D., Keele, B.F., Ndjango, J.
B.N., Sanz, C.M., Morgan, D.B., Locatelli, S., Gonder, M.K., Kranzusch. PJ.,
Walsh, P.O., Delaporte, E., Mpoudi-Ngole, E., Georgiev, A.V., Muller, M.N.,
Shaw, G.M., Peeters, M., Sharp, P.M., Rayner, J.C .• Hahn, B.H., 2010. Origin
178
University of Ghana http://ugspace.ug.edu.gh

of the human malaria parasite Plasmodium falciparwn in gorillas. Nature 467,
420-425.
Lopansri, B.K., Anstey, N.M., Weinberg, J.8., Stoddard. 0.1., Hobbs, M.R., Levesque,
M.C., Mwaikambo, E.D.. Granger, D.L., 2003. Low plasma arginine
concentrations in children with cerebral malaria and decreased nitric oxide
production. Lancet Lond. Engl. 361, 676-678.
Lovegrove, F.E., Tangpukdee, N., Opoka, R.O., Lafferty, E.!.. Rajwans. N., Hawkes,
M., Krudsood, S., Looareesuwan, S., John, C.C., Liles, W.C., Kain, K.C., 2009.
Serum angiopoietin-l and -2 levels discriminate cerebral malaria from
uncomplicated malaria and predict clinical outcome in African children. PloS
One 4, e4912.
MacClellan, L.R., Howard, T.D., Cole, J.W., Stine, O.C., Giles, W.H., O'Connell, J.R.,
Wozniak. M.A., Stem, B.J., Mitchell, B.D., Kittner, S.J., 2009. Relation of
candidate genes that encode for endothelial function to migraine and stroke: the
Stroke Prevention in Young Women study., Relation of Candidate Oenes that
Encode for Endothelial Function to Migraine and Stroke: The Stroke Prevention
in Young Women Study. Stroke Stroke J. Cereb. Circ. 40,40, e550, e550-7.
Mac-Daniel, L., Buckwalter. M.R., Berthet, M., Virk, Y., Yui, K., Albert, M.L.,
Gueirard, P., Menard, R., 2014. Local immune response to injection of
Plasmodium sporozoites into the skin. J. Immunol. Baltim. Md 1950 193, 1246-
1257.
Machado, F.S., Desruisseaux, M.S., Nagajyothi, null, Kennan, R.P., Hetherington,
H.P., Wittner, M., Weiss, L.M., Lee, S.C., Scherer, P.E., Tsuji, M., Tanowitz.
H.B., 2006. Endothelin in a murine model of cerebral malaria. Exp. BioI. Med.
Maywood NJ 231,1176-1181.
MacPherson, G.G., Warrell, M.J., White, N.J., Looareesuwan, S., Warrell, D.A., 1985.
Human cerebral malaria. A quantitative ultrastructural analysis of parasitized
erythrocyte sequestration. Am. J. Pathol. 119,385-401.
Maestre, A., Muskus, C., Duque, V., Agudelo, 0., Liu, P., Takagi, A., Ntumngia, F.B.,
Adams, J.H., Sim, K.L., Hoffman, S.L., Corradin, G., Velez, 1.0., Wang, R.,
2010. Acquired Antibody Responses against Plasmodium vivax Infection Vary
with Host Genotype for Duffy Antigen Receptor for Chemokines (DARC).
PLOS ONE 5, e11437.
1'70
University of Ghana http://ugspace.ug.edu.gh

Maier, A.G., Duraisingh, M.T., Reeder, J.C .• Patel, S.S., K.azura, J.W .• Zimmerman,
P.A.. Cowman, A.F., 2003. Plasmodium falciparum erythrocyte invasion
through glycophorin C and selection for Gerbich negativity in human
populations. Nat. Med. 9, 87-92.
Malaria diagnosis: Memorandum from a WHO Meeting. 1988 .. Bull. World Health
Organ. 66,575-594.
Malaria Genomic Epidemiology Network. 2015. A novel locus of resistance to severe
malaria in a region of ancient balancing selection. Nature 526, 253.
Maneerat, Y., Pongponratn, E., Viriyavejakul, P., Punpoowong, B., Looareesuwan, S.,
Udomsangpetch. R., 1999. Cytokines associated with pathology in the brain
tissue of fatal malaria. Southeast Asian J. Trop. Med. Public Health 30, 643-
649.
Manjurano, A., Sepulveda. N., Nadjm, B., Mtove, G., Wangai, H., Maxwell, c., Olomi,
R., Reyburn, H., Drakeley, C.J., Riley, E.M., Clark, T.G., 2015. USP38,
FREM3, SOC I, DOC, and LOC727982 Gene Polymorphisms and Differential
Susceptibility to Severe Malaria in Tanzania. J. Infect. Dis. 212. 1129-1139.
Manke, T., Heinig, M., Vingron, M., 2010. Quantifying the effect of sequence variation
on regulatory interactions. Hum. Mutat. 31,477-483.
Manning, L., Rosanas-Urgell, A., Laman, M., Edoni, H., McLean, C., Mueller, I., Siba,
P., Davis, T.M.E., 2012. A histopathologic study of fatal paediatric cerebral
malaria caused by mixed Plasmodium faiciparumlPlasmodium vi vax infections.
Malar. J. II, 107.
Manson, P., 1898. The Mosquito And The Malaria Parasite. Br. Med. J. 2, 849-853.
Mantel, P.-Y., Hjelmqvist, D., Walch, M., Kharoubi-Hess, S., Nilsson, S., Ravel, D.,
Ribeiro, M., GrUring, c., Ma, S., Padmanabhan, P., Trachtenberg, A.,
Ankarklev, J., Brancucci, N.M., Huttenhower, C., Duraisingh, M.T., Ghiran, I.,
Kuo, W.P., Filgueira, L., Martinelli, R., Marti, M., 2016. Infected erythrocyte
derived extracellular vesicles alter vascular function via regulatory Ag02-
miRNA complexes in malaria. Nat. Commun. 7.
Marsh, K., 1992. Malaria--a neglected disease? Parasitology 104 Suppl. S53-69.
May, J., Evans, J.A., Timmann, C., Ehmen, C., Busch, W., Thye, T., Agbenyega, T.,
Horstmann, R.D., 2007. Hemoglobin variants and disease manifestations in
severe falciparum malaria. JAMA 297,2220-2226.
180
University of Ghana http://ugspace.ug.edu.gh

Mazier, D., Nitcheu. J.. Idrissa-Boubou. M., 2000. Cerebral malaria and
immunogenetics. Parasite Immuno!. 22, 613-623.
McCall, M.B.B., Sauerwein, R.W., 2010. Interferon-y--central mediator of protective
immune responses against the pre-erythrocytic and blood stage of malaria. J.
Leukoc. BioI. 88, 1131-1143.
McGuire, W., Hill, A.V.S., Allsopp, C.E.M., Greenwood. B.M., Kwiatkowski, D.,
1994. Variation in the TNF-a promoter region associated with susceptibility to
cerebral malaria. Nature 371, 508-511.
Meding, SJ., Langhorne, J., 1991. CD4+ T cells and B cells are necessary for the
transfer of protective immunity to Plasmodium chabaudi chabaudi. Eur. J.
Immuno!. 21, 1433-1438.
Mendon~a. D., R, V.R., Goncalves, M.S., Bacral-Netto, M., 2012. The Host Genetic
Diversity in Malaria Infection [WWW Document]. J. Trop. Med. URL
https:llwww.hindawLcomljoumals/jtml20 12/9406161 (accessed 7.24.18).
Meyer, H.A., Astor, R.A., 2002. Child and Parent Perspectives on Routes to and from
School in High Crime Neighborhoods. J. Sch. Violence 1, 101-128.
Miller, J.L.. Sack, B.K., Baldwin, M., Vaughan, A.M., Kappe, S.H.I., 2014. Interferon
mediated innate immune responses against malaria parasite liver stages. Cell
Rep. 7,436-447.
Miller, L.H., Baruch, 0.1., Marsh, K., Doumbo, O.K., 2002. The pathogenic basis of
malaria. Nature 415,673-679.
Miyashita, H., Kanemura, M., Yamazaki, T., Abe, M., Sato, Y., 2004. Vascular
endothelial zinc finger I is involved in the regulation of angiogenesis: possible
contribution of stathrninlOP 18 as a downstream target gene. Arterioscler.
Thromb. Vasco BioI. 24, 878-884.
Montasser, M.E., Shimmin, L.C., Gu, D., Chen, J., Gu, c., Kelly, T.N., Jaquish, C.E.,
Rice, T., Rao, D.C., Cao, J., Chen, J., Liu, D.P., Whelton, P., He, J., Hixson,
J.E., 2010. Blood pressure response to potassium supplementation is associated
with genetic variation in endothelin 1 and interactions with E selectin in rural
Chinese. J. Hypertens. 28, 748-755.
Moszyilska. A., Gebert, M., Collawn, J.F., Bartoszewski, R., 2017. SNPs in microRNA
target sites and their potential role in human disease. Open BioI. 7.
181
University of Ghana http://ugspace.ug.edu.gh

Mota. M.M., Brown, K.N., Holder, A.A., Jarra, W., 1998. Acute Plasmodium chabaudi
chabaudi malaria infection induces antibodies which bind to the surfaces of
parasitized erythrocytes and promote their phagocytosis by macrophages in
vitro. Infect. Immun. 66, 4080-4086.
Mullins, R.F., Skeie, J.M., Folk, J.C., Solivan-Timpe, F.M., Oetting, T.A., Huang, J.,
Wang, K., Stone, E.M., Fingert, J.H., 2011. Evaluation of variants in the selectin
genes in age-related macular degeneration. BMC Med. Genet. 12, 58.
Munde. E.O., Okeyo, W.A., Anyona, S.B., Raballab, E., Konab, S., Okumu, W.,
Ogonda, L., Vulule, J., Ouma, c., 2012. Polymorphisms in the Fc Gamma
Receptor IlIA and Toll-Like Receptor 9 Are Associated with Protection against
Severe Malarial Anemia and Changes in Circulating Gamma Interferon Levels.
Infect. Immun. 80,4435-4443.
Munde, E.O., Okeyo, W.A., Raballab, E., Anyona, S.B., Were, T., Ong'echa, J.M.,
Perkins, D.J., Ouma, C., 2017. Association between Fcy receptor IIA, IlIA and
I1IB genetic polymorphisms and susceptibility to severe malaria anemia in
children in western Kenya. BMC Infect. Dis. 17,289.
Nagel, R.L., Raventos-Suarez, C., Fabry, M.E., Tanowitz, H., 1981. Impainnent of the
Growth of Plasmodium Falciparum in HbEE Erythrocytes. J. Clin. Invest. 68,
303-305.
Naka. I., Nishida, N., Patarapotikul, J., Nuchnoi, P., Tokunaga, K., Hananantachai, H.,
Tsuchiya, N., Ohashi, J., 2009. Identification of a haplotype block in the 5q31
cytokine gene cluster associated with the susceptibility to severe malaria. Malar.
J. 8, 232.
Nakashima, K., Hirota, T., Obara, K., Shimizu, M., Doi, S., Fujita. K., Shirakawa, T.,
Enomoto, T., Yoshihara, S., Ebisawa, M., Matsumoto, K., Saito, H., Suzuki, Y.,
Nakamura, Y., Tamari, M., 2006. A functional polymorphism in MMP-9 is
associated with childhood atopic asthma. Biochem. Biophys. Res. Commun.
344,300-307.
Nantakomol, D., Dondorp, A.M., Krudsood, S., Udomsangpetch, R., Pattanapanyasat,
K., Combes. V .• Grall, G.E .• White, N.J .• Viriyavejakul. P., Day, N.P.J.,
Chotivanich, K., 2011. Circulating red cell-derived microparticles in human
malaria. J. Infect. Dis. 203, 700-706.
182
University of Ghana http://ugspace.ug.edu.gh

Nasibullin. T.R .• Timasheva. Y.R, Sadikova. R.I., Tuktarova. I.A., Erdman, V.V .•
Nikolaeva. I.E., Sabot J., Kruzliak, P., Mustafina. O.E., 2016. Genotype/allelic
combinations as potential predictors of myocardial infarction. Mol. Bioi. Rep.
43, 11-16.
N'Dilimabaka. N., Taoufiq, Z., Zoll8b6d6, S., Bonnefoy, S., Lorthiois, A., Couraud,
P.O., Rebollo, A., Snounou, G., Mazier, D., Moreno Sabater, A., 2014. P.
falciparum Isolate-Specific Distinct Patterns of Induced Apoptosis in
Pulmonary and Brain Endothelial Cells. PLoS ONE 9, e90692.
Ndungu, F.M., Cadman, E.T., Coulcher, J., Nduati, E., Couper, E., Macdonald, D.W.,
Ng, D., Langhorne, J., 2009. Functional memory B cells and long-lived plasma
cells are generated after a single Plasmodium chabaudi infection in mice. PLoS
Pathog. 5, el000690.
Ndungu, F.M., Olotu, A., Mwacharo, J., Nyonda. M., Apfeld, J., Mramba, L.K., Fegan,
G.W., Bejon, P., Marsh, K., 2012. Memory B cells are a more reliable archive
for historical antimalarial responses than plasma antibodies in no-longer
exposed children. Proc. Natl. Acad. Sci. U. S. A. 109,8247-8252.
Newton, C.R. Hien, T.T., White, N., 2000. Cerebral malaria. J. Neurol. Neurosurg.
Psychiatry 69, 433-441.
Nitcheu, J., Bonduelle, 0., Combadiere, C., Tefit, M., Seilhean, D., Mazier, D .•
Combadiere, B., 2003. Perf orin-Dependent Brain-Infiltrating Cytotoxic CD8+
T Lymphocytes Mediate Experimental Cerebral Malaria Pathogenesis. J.
Immunol. 170, 2221-2228.
Nkumama, I.N., O'Meara, W.P., Osier, F.H.A., 2017. Changes in Malaria
Epidemiology in Africa and New Challenges for Elimination. Trends Parasitol.
33, 128-140.
Nonvignon, J., Aryeetey, G.C., MaIm, K.L., Agyemang, S.A., Aubyn, V.N.A., Peprah,
N.Y., Bart-Plange, C.N., Aikins, M., 2016. Economic burden of malaria on
businesses in Ghana: a case for private sector investment in malaria control.
Malar. J. 15,454.
Oakley, M.S., Gerald, N., McCutchan, T.F., Aravind, L., Kumar, S., 2011. Clinical and
molecular aspects of malaria fever. Trends Parasitol. 27, 442-449.
Obermeier, B., Daneman, R, Ransohoff, RM., 2013. Development, maintenance and
disruption of the blood-brain barrier. Nat. Med. 19,1584-1596.
183
University of Ghana http://ugspace.ug.edu.gh

Oduro. D .• 2015. THE ROLE OF CIRCULATING ENDOTHEUAL PROGENITOR
CELLS (CEPCs) AND OTHER BIOMARKERS IN THE PATHOGENESIS
OF CEREBRAL MALARIA. University of Ghana.
Offeddu. V., Thathy, V., Marsh, K., Matuschewski, K.., 2012. Naturally acquired
immune responses against Plasmodium falciparum sporozoites and liver
infection. Int. J. Parasitol. 42, 535-548.
Olivier, M., Van Den Ham. K .• Shio, M.T., Kassa, F.A., Fougeray, S., 2014. Malarial
pigment hemozoin and the innate inflammatory response. Front. Immunol. 5,
25.
Omar, A.H., Yasunami, M., Yamazaki, A., Shibata, H., Ofori, M.F., Akanmori, B.D.,
Shuaibu, M.N., Kikuchi. M., Hirayama, K., 2012. Toll-like receptor 9 (TLR9)
polymorphism associated with symptomatic malaria: a cohort study. Malar. J.
11,168.
O'Meara. W.P., Bejon, P., Mwangi, T.W., Okiro, E.A., Peshu, N., Snow, R.W.,
Newton, C.R., Marsh, K., 2008. Effect of a fall in malaria transmission on
morbidity and mortality in Kilifi, Kenya. The Lancet 372, 1555-1562.
Para, S., Mungkalasut, P., Chanda, M., Nuchprayoon, I., Krudsood, S., Cheepsunthorn,
C.L.. 2018. An Observational Study of the Effect of Hemoglobinopathy. Alpha
Thalassemia and Hemoglobin E on P. Vivax Parasitemia. Mediterr. J. Hematol.
Infect. Dis. 10.
Parroche, P., Lauw, F.N., Goutagny, N., Latz, E., Monks, B.G., Visintin, A., Halmen.
K.A.. Lamphier, M., Olivier, M., Bartholomeu, D.C., Gazzinelli, R.T.,
Golenbock. D.T., 2007. Malaria hemozoin is immunologically inert but
radically enhances innate responses by presenting malaria DNA to Toll-like
receptor 9. Proc. Natl. Acad. Sci. 104, 1919-1924.
Pedersen, A.B., Mikkelsen, E.M., Cronin-Fenton, D., Kristensen. N.R., Pham, T.M.,
Pedersen, L., Petersen, I., 2017. Missing data and multiple imputation in clinical
epidemiological research. Clin. Epidemiol. 9, 157-166.
Pemberton-Ross, P., Smith, T.A., Hodel, E.M., Kay, K., Penny, M.A., 2015. Age
shifting in malaria incidence as a result of induced immunological deficit: a
simulation study. Malar. J. 14,287.
Perez-Mazliah, D., Langhorne, J., 2015. CD4 T-Cell Subsets in Malaria: THlffH2
Revisited. Front. Immunol. S.
184
University of Ghana http://ugspace.ug.edu.gh

Pluess, B., Tanser, F.C., Lengeler, C., Sharp, B.L., 2010. Indoor residual spraying for
preventing malaria. Cochrane Database Syst. Rev. CD0066S7.
Podoba, J .E., Stevenson, M.M., 1991. CD4+ and CD8+ T lymphocytes both contribute
to acquired immunity to blood-stage Plasmodium chabaudi AS. Infect. Immun.
59,51-58.
Poggesi, A., Pasi, M., Pescini, F., Pantoni, L., Inzitari, D., 2016a. Circulating biologic
markers of endothelial dysfunction in cerebral small vessel disease: A review.
J. Cereb. Blood Flow Metab. 36, 72-94.
Poggesi, A., Pasi, M., Pescini, F., Pantoni, L., Inzitari, D., 2016b. Circulating biologic
markers of endothelial dysfunction in cerebral small vessel disease: A review.
J. Cereb. Blood Flow Metab. 36, 72-94.
Pombo, OJ., Lawrence, G., Hirunpetcharat, C., Rzepczyk, C., Bryden, M., Cloonan,
N., Anderson, K., Mahakunkijcharoen, Y., Martin, L.B., Wilson, D., Elliott,
Salenna, Elliott, Suzanne, Eisen, D.P., Weinberg, J.B., Saul, A., Good, M.F.,
2002. Immunity to malaria after administration of ultra-low doses of red cells
infected with Plasmodium falciparum. Lancet Lond. Engl. 360, 610~17.
Ponsford, M.J., Medana, I.M., Prapansilp, P., Hien, T.T., Lee, SJ., Dondorp, A.M.,
Esiri, M.M., Day, N.P.J., White, N.J., Tumer, G.D.H., 2012. Sequestration and
microvascular congestion are associated with coma in human cerebral malaria.
J. Infect. Dis. 205, 663~71.
Postels, D.G., Birbeck, G.L., 2013. Cerebral malaria. Handb. Clin. Neurol. 114, 91-
102.
Prapansilp, P., Medana,I., Mai, N.T.H., Day, N.P.J., Phu, N.H., Yeo, T.W., Hien, T.T.,
White, NJ., Anstey, N.M., Tumer, G.D.H., 2013. A clinicopathological
correlation of the expression of the angiopoietin-Tie-2 receptor pathway in the
brain of adults with Plasmodium falciparum malaria. Malar. J. 12,50.
Prudencio, M., Rodriguez, A., Mota, M.M., 2006. The silent path to thousands of
merozoites: the Plasmodium liver stage. Nat. Rev. Microbiol. 4, 849-856.
Pruim, RJ., Welch, R.P., Sanna, S., Teslovich, T.M., Chines, P.S., Gliedt, T.P.,
Boehnke, M., Abecasis, G.R., Willer, CJ., 2010. LocusZoom: regional
visualization of genome-wide association scan results. Bioinfonna. Oxf. Eng!.
26,2336-2337.
185
University of Ghana http://ugspace.ug.edu.gh

Rafii. S .• 2000. Circulating endothelial precursors: mystery, reality, and promise. J.
Clin. Invest. 105, 17-19.
Ravenhall, M., Campino, S., Sepulveda, N., Manjurano, A., Nadjm, B., Mtove, G.,
Wangai, H., Maxwell, c., Olomi, R., Reyburn, H. t Orakeley, C.J., Riley, E.M.,
Clark, T.G., MalariaGEN, I.C.W., 2018. Novel genetic polymorphisms
associated with severe malaria and under selective pressure in North-eastern
Tanzania. PLOS Genet. 14, eI007172.
Renia, L., Howland, S.W., Claser, C., Charlotte Gruner, A., Suwanarusk, R., Hui Teo,
T., Russell, B., Ng, L.F.P., 2012. Cerebral malaria: mysteries at the blood-brain
bamer. Virulence 3, 193-201.
Resch. T., Pircher, A., Kllhler, C.M., Pratschke, J., Hilbe, W., 2012. Endothelial
Progenitor Cells: Current Issues on Characterization and Challenging Clinical
Applications. Stem Cell Rev. Rep. 8,926-939.
Ribatti, D., 2007. The discovery of endothelial progenitor cells. An historical review.
Leuk. Res. 31, 439-444.
Rich, S.M., Leendertz, F.H., Xu, G., LeBreton, M., Djoko, C.F., Aminake, M.N.,
Takang, E.E., Diffo, J.L., Pike, B.L., Rosenthal, B.M., others, 2009. The origin
of malignant malaria. Proc. Natl. Acad. Sci. 106, 14902-14907.
Riggle, B.A., Miller, L.H., Pierce, S.K., 2017. Do we know enough to find an adjunctive
therapy for cerebral malaria in African children? FI OOOResearch 6.
Riglar, D.T., Richard, D., Wilson, D.W., Boyle, M.J., Dekiwadia, C., Turnbull, L.,
Angrisano, F., Marapana, D.S., Rogers, K.L., Whitchurch, C.B., Beeson, J.G.,
Cowman, A.F., Ralph, S.A., Baum, J., 2011. Super-Resolution Dissection of
Coordinated Events during Malaria Parasite Invasion of the Human
Erythrocyte. Cell Host Microbe 9, 9-20.
Riley, E.M., Wagner, G.E., Ofori, M.F., Wheeler, J.G., Akanmori, B.D., Tetteh, K.,
McGuinness, D., Bennett, S., Nkrumah, F.K., Anders, R.F., Koram, K.A., 2000.
Lack of Association between Maternal Antibody and Protection of African
Infants from Malaria Infection. Infect. Immun. 68, 5856-5863.
RiSCO-Castillo, V., Top~u, S., Marinach, C., Manzoni, G., Bigorgne, A.E., Briquet, S.,
Baudin, X., Lebrun, M., Dubremetz, J.-F., Silvie, 0., 2015. Malaria Sporozoites
Traverse Host Cells within Transient Vacuoles. Cell Host Microbe 18, 593-
603.
186
University of Ghana http://ugspace.ug.edu.gh

Robertson, K.D., 2005. DNA methylation and human disease. Nat. Rev. Genet. 6, 597-
610.
Roestenberg, M., McCall, M., Hopman, J., Wiersma, J., Luty, A.J.F., van Gernert. G.J.,
van de Vegte-Bolmer, M., van Schaijk, B., Teelen, K., Arens, T., Spaannan, L.,
de Mast, Q., Roeffen, W., Snounou, G., Renia. L., van der Ven, A., Hennsen,
C.C., SaueIWein, R., 2009. Protection against a malaria challenge by sporozoite
inoculation. N. Eng!. J. Med. 361,468-477.
Rosa, T.F.A., Flammersfeld, A., Ngwa, C.J., Kiesow, M., Fischer, R., Zipfel, P.F.,
Skerka, C., Pradel. G., 2016. The Plasmodium falciparum blood stages acquire
factor H family proteins to evade destruction by human complement. Cell.
Microbiol. 18, 573-590.
Rowe, J.A., Claessens, A., Corrigan, R.A., Arman, M., 2009. Adhesion of Plasmodium
falciparum-infected erythrocytes to human cells: molecular mechanisms and
therapeutic implications. Expert Rev. Mol. Med. 11.
Rucker, D.O., McShane, B.B., Preacher, K.J., 2015. A researcher's guide to regression,
discretization, and median splits of continuous variables. J. Consum. Psychol.
25, 666-678.
Ruwende, C., Khoo. S.C., Snow, R.W., Yates, S.N., Kwiatkowski, D., Gupta. S., Warn,
P., Allsopp, C.E., Gilbert, S.C., Peschu, N., 1995. Natural selection of hemi
and heterozygotes for G6PD deficiency in Africa by resistance to severe
malaria. Nature 376, 246-249.
Sabchareon, A., Burnouf, T., Ouattara, D., Attanath, P., Bouharoun-Tayoun, H.,
Chantavanich, P., Foucault, C., Chongsuphajaisiddhi. T., Druilhe, P., 1991.
Parasitologic and clinical human response to immunoglobulin administration in
falciparum malaria. Am. J. Trop. Med. Hyg. 45, 297-308.
Safeukui, I., Gomez, N.D., Adelani, A.A., Burte, F., Afolabi, N.K., Akondy, R.,
Velazquez, P., Holder, A., Tewari, R., Buffet, P., Brown, B.1., Shokunbi, W.A.,
Olaleye, D., Sodeinde, 0., Kazura, J., Ahmed, R., Mohandas, N., Fernandez
Reyes, D., Haldar, K., 2015. Malaria induces anemia through CD8+ T cell
dependent parasite clearance and erythrocyte removal in the spleen. mBio 6.
Sahu, U., Sahoo, P.K., Kar, S.K., Mohapatra, B.N., Ranjit, M., 2013. Association of
TNF level with production of circulating cellular microparticles during clinical
manifestation of human cerebral malaria. Hum. Immunol. 74,713-721.
187
University of Ghana http://ugspace.ug.edu.gh

Salanti. G., Amountza, G., Ntzani, E.E., Ioannidis, J.P.A., 2005. Hardy-Weinberg
equilibrium in genetic association studies: an empirical evaluation of reporting,
deviations, and power. Eur. J. Hum. Genet. EJHG 13,840-848.
Santos, J.C.D., Cruz, M.S., Bortolin, R.H., Oliveira, K.M. de, Aralijo, J.N.G. de,
Duarte. V.H.R., Silva, A.M.G. da, Santos, I.C.C.D., Dantas, J.M. de 0., Paiva,
M.S.M. de 0., Rezende, A.A., Hirata, M.H., Hirata, R.D.C., luchessi, A.D.,
Silbiger, V.N., 2018. Relationship between circulating VCAM-I, ICAM-I, E
selectin and MMP9 and the extent of coronary lesions. Clin. Sao Paulo Braz.
73, e203.
Schofield, L., Hackett, F., 1993. Signal transduction in host cells by a
glycosylphosphatidylinositol toxin of malaria parasites. J. Exp. Med. 177, 145-
153.
SchrOder, N.W., Schumann, R.R., 2005. Single nucleotide polymorphisms of Toll-like
receptors and susceptibility to infectious disease. lancet Infect. Dis. 5, 156-
164.
Schubert, C., Pryds, A., Zeng, S., Xie, Y., Freund, K.B., Spaide, R.F., Merriam. J.C.,
Barbazetto, I., Slakter, J.S., Chang, S., Munch, I.C., Drack, A.V., Hernandez,
J., Yzer, S., Merriam. J.E., Linneberg, A., Larsen, M., Yannuzzi, L.A., Mullins.
R.F., AIlikmets, R., 2014. Cadherin 5 is Regulated by Corticosteroids and
Associated with Central Serous Chorioretinopathy. Hum. Mutat. 35, 859-867.
Schulman, S., Roth, E.F., Cheng, B., Rybicki, A.C., Sussman, I.I., Wong, M., Wang.
W., Ranney, H.M., Nagel. R.L., Schwartz, R.S., 1990. Growth of Plasmodium
falciparum in human erythrocytes containing abnormal membrane proteins.
Proc. Natl. Acad. Sci. 87, 7339-7343.
Schumak, B., Klocke, K., Kuepper, J.M., Biswas, A .• Djie-Maletz, A., Limmer, A., van
Rooijen, N., Mack, M., Hoerauf, A., Dunay, I.R., 2015. Specific Depletion of
ly6Chi Inflammatory Monocytes Prevents Immunopathology in Experimental
Cerebral Malaria. PloS ONE 10.
Shabani, E., Ouma, B.J., Idro, R., Bangirana, P., Opoka, R.O., Park, G.S., Conroy, A.L..
John, C.C., 2017. Elevated cerebrospinal fluid tumor necrosis factor is
associated with acute and long-term neurocognitive impairment in cerebral
malaria. Parasite Immunol. 39.
188
University of Ghana http://ugspace.ug.edu.gh

Shames, D.S., Minna, J.D., Gazdar, A.F., 2007. DNA Methylation in Health, Disease,
and Cancer. Curro Mol. Med. 7, 85-102.
Shaw, G.M., Lu. W., Zhu. H., Yang, W., Briggs, F.B., Carmichael, S.L., Barcellos,
L.F., Lammer, EJ., Finnell, R.H., 2009. 118 SNPs of folate-related genes and
risks of spina bifida and conotruncal heart defects., 118 SNPs of folate-related
genes and risks of spina bifida and conotruncal heart defects. BMC Med. Genet.
BMC Med. Genet. 10, 10,49,49-49.
Shortt, H.E., Garnham, P.C.C., 1948. Pre-erythrocytic stage in mammalian malaria
parasites. Nature 161, 126.
Singh, S., Alam, M.M., Pal-Bhowmick, I., Brzostowski, J.A.. Chitnis, C.E., 2010.
Distinct external signals trigger sequential release of apical organelles during
erythrocyte invasion by malaria parasites. PLoS Pathog. 6, e 1 000746.
Smith, R.C., Vega-Rodriguez. J., Jacobs-Lorena, M., 2014. The Plasmodium
bottleneck: malaria parasite losses in the mosquito vector. Mem. Inst. Oswaldo
Cruz 109,644--061.
Sole, X., Guin6. E .• Valls, J., lniesta, R., Moreno. V .• 2006. SNPStats: a web tool for
the analysis of association studies. Bioinformatics 22.1928-1929.
Solomon. W .• Wilson, N.O., Anderson, L., Pitts, S., Patrickson, J., Liu, M., Ford, B.D.,
Stiles, J.K., 2014. Neuregulin-l attenuates mortality associated with
experimental cerebral malaria. J. Neuroinflammation 11,9.
Sol us, J.F., Chung, C.P., Oeser. A., Li, c., Rho, Y.H., Bradley, K.M., Kawai, V.K.,
Smith, J.R., Stein, C.M., 2015. Genetics of serum concentration of IL-6 and
TNFa in systemic lupus erythematosus and rheumatoid arthritis: a candidate
gene analysis. Clin. Rheumatol. 34, 1375-1382.
Souza, M.C., Silva, J.D., Padua, T.A., Torres, N.D., Antunes, M.A., Xisto, D.G., Abreu,
T.P., Capelozzi, V.L., Morales, M.M., Sa Pinheiro, A.A., Caruso-Neves, C.,
Henriques, M.G., Rocco, P.R., 2015. Mesenchymal stromal cell therapy
attenuated lung and kidney injury but not brain damage in experimental cerebral
malaria. Stem Cell Res. Ther. 6.
Starke, R.D., Ferraro, F., Paschalald, K.E., Dryden, N.H .• McKinnon, T.A.J., Sutton,
R.E., Payne, E.M., Haskard, D.O., Hughes, A.D., Cutler, D.F., Laffan, M.A.,
Randi, A.M., 2011. Endothelial von Willebrand factor regulates angiogenesis.
Blood 117, 1071-1080.
189
University of Ghana http://ugspace.ug.edu.gh

Storm. J., Craig, A.G., 2014. Pathogenesis of cerebral malaria-inflammation and
cytoaciherence. Front. Cell. Infect. Microbiol. 4.
Siiss, G., Eichmann. K., Kury, E., Linke, A., Langhorne, J., 1988. Roles ofCD4- and
CDS-bearing T lymphocytes in the immune response to the erythrocytic stages
of Plasmodium chabaudi. Infect. Immun. 56, 30S1-30S8.
Swanson, P.A., Hart, G.T., Russo, M.V., Nayak, D .• Yazew, T .• Pefia, M., Khan, S.M.,
Janse, C.J., Pierce. S.K .• McGavern, D.B., 2016. CDS+ T Cells Induce Fatal
Brainstem Pathology during Cerebral Malaria via Luminal Antigen-Specific
Engagement of Brain Vasculature. PLoS Pathog. 12.
Tachado, S.D., Gerold. P., McConville, M.J., Baldwin, T., Quilici, D., Schwarz, R.T.,
Schofield, L.. 1996. Glycosylphosphatidylinositol toxin of Plasmodium induces
nitric oxide synthase expression in macrophages and vascular endothelial cells
by a protein tyrosine kinase-dependent and protein kinase C-dependent
signaling pathway. J. Immunol. Baltim. Md 1950 156, IS97-1907.
Taguchi, A., Matsuyama. T., Nakagomi, T., Shimizu, Y., Fukunaga, R., Tatsumi, Y.,
Yoshikawa, H., Kikuchi-Taura, A., Soma, T., Moriwaki, H., Nagatsuka, K.,
Stern, D.M., Naritomi, H., 200S. Circulating CD34-Positive Cells Provide a
Marker of Vascular Risk Associated with Cognitive Impairment. J. Cereb.
Blood Flow Metab. 28, 445-449.
Tan, J., Pieper, K., Piccoli, L., Abdi, A., Perez, M.F., Geiger, R., Tully, C.M., Jarrossay,
D., Maina Ndungu, F., Wambua, J., Bejon, P., Fregni, C.S., Fernandez
Rodriguez, B., Barbieri, S., Bianchi, S., Marsh, K., Thathy, V., Corti, D.,
Sallusto, F .• Bull, P., Lanzavecchia, A., 2016. A LAIRI insertion generates
broadly reactive antibodies against malaria variant antigens. Nature 529, 105-
109.
Tang, S., Feng, D., Wang, R., Ghose, B., Hu, T., Ji, L., Wu, T., Fu. H., Huang. Y., Feng,
Z., 2017. Economic burden of malaria inpatients during National Malaria
Elimination Programme: estimation of hospitalization cost and its inter
province variation. Malar. 1. 16,291.
Tangpukdee, N., Krudsood, S., Kano, S., Wilairatana, P., 2012. Falciparum malaria
parasitemia index for predicting severe malaria. Int. J. Lab. Hematol. 34, 320-
327.
190
University of Ghana http://ugspace.ug.edu.gh

Tarazona-Santos, E., Castilho, L., Amaral, D.R. T., Costa, D.C., Furlani, N.G.,
Zuccherato, L.W., Machado, M., Reid, M.E., Zalis, M.G., Rossit, A.R., Santos,
S.E.B., Machado, R.L., Lustigman, S., 2011. Population Genetics ofGVPB and
Association Study between GYPB·S/s Polymorphism and Susceptibility to P.
falciparum Infection in the Brazilian Amazon. PLOS ONE 6, e16123.
Tavares, J., Formaglio, P., Thiberge, S., Mordelet. E., Van Rooijen. N., Medvinsky, A .•
Menard, R., Amino, R., 2013. Role of host cell traversal by the malaria
sporozoite during liver infection. J. Exp. Med. 210,905-915.
Tawiah, T., Asante, K.P., Dwommoh, R.A., Kwarteng. A., Gyaase, S., Mahama, E.,
Abokyi, L., Amenga-Etego. S., Hansen, K., Akweongo, P., Owusu-Agyei, S.,
2016. Economic costs of fever to households in the middle belt of Ghana. Malar.
1. 15,68.
Taylor, S.M., Parobek, C.M., Fairhurst, R.M., 2012. Impact ofhaemoglobinopathies on
the clinical epidemiology of malaria: a systematic review and meta-analysis.
Lancet Infect. Dis. 12, 457-468.
Taylor, T.E., Fu, W.J., Carr, R.A., Whitten, R.O., Mueller, J.S., Fosiko, N.G., Lewallen,
S., Liomba, N.G., Molyneux, M.E., Mueller, J.G., 2004. Differentiating the
pathologies of cerebral malaria by postmortem parasite counts. Nat. Med. 10,
143-145.
Tetteh, H., 2014. Endothelial Biomarkers and the Pathogenesis of Cerebral Malaria
(Thesis). University of Ghana.
Tham, W.-H., Healer, J., Cowman, A.F., 2012. Erythrocyte and reticulocyte binding
like proteins of Plasmodium fa\Ciparum. Trends Parasitol. 28, 23-30.
Timasheva, Y.R., Nasibullin, T.R., Imaeva, E.B., Erdman, V.V., Kruzliak, P ..
Tuktarova, I.A., Nikolaeva, I.E., Mustafina, O.E., 2015. Polymorphisms of
inflammatory markers and risk of essential hypertension in Tatars from Russia.
Clin. Exp. Hypertens. N. Y. N 199337,398-403.
Togbe, D., Schofield, L., Grau, G.E., Schnyder, B., Boissay, V., Charron, S., Rose, S.,
Beutler, B., Quesniaux, V.F.J., Ryffel, B., 2007. Murine cerebral malaria
development is independent of toll-like receptor signaling. Am. J. Pathol. 170,
1640-1648.
ToreHa, D., Ellison, G.M., Torella, M., Vicinanza, C., Aquila, I., Iaconetti, C., Scalise,
M., Marino, F., Henning. B.J., Lewis, F.C., Gareri, C., Lascar, N., Cud&, G.,
191
University of Ghana http://ugspace.ug.edu.gh

Salvatore, T., Nappi, G., Indolfi, C., Torella, R., Cozzolino, D., Sasso, F.C.,
2014. Carbonic Anhydrase Activation Is Associated With Worsened
Pathological Remodeling in Human Ischemic Diabetic Cardiomyopathy. J. Am.
Heart Assoc. Cardiovasc. Cerebrovasc. Dis. 3.
TRANG, N.V., CHOISY, M., NAKAGOMI, T., CHINH, N.T.M .• DOAN. Y.H ••
YAMASHIRO, T., BRYANT, J.E., NAKAGOMI, 0., ANH, D.O., 2015.
Detennination of cut-off cycle threshold values in routine RT -PCR assays to
assist differential diagnosis of norovirus in children hospitalized for acute
gastroenteritis. Epidemiol. Infect. 143,3292-3299.
Turner, G.D., Ly, V.C., Nguyen, T.H., Tran, T.H., Nguyen, H.P., Bethell, D., Wyllie.
S., Louwrier, K., Fox, S.B., Gatter, K.C., Day, N.P., Tran. T.H., White, N.J.,
Berendt, A.R., 1998. Systemic endothelial activation occurs in both mild and
severe malaria. Correlating dennal microvascular endothelial cell phenotype
and soluble cell adhesion molecules with disease severity. Am. J. Pathol. 152,
1477-1487.
Turner, L., Lavstsen. T., Berger, S.S., Wang, C.W., Petersen, J.E.V., Avril, M., Brazier,
A.J., Freeth, J., Jespersen, J.S., Nielsen, M.A., Magistrado, P., Lusingu, J.,
Smith, J.D., Higgins, M.K., Theander, T.G., 2013. Severe malaria is associated
with parasite binding to endothelial protein C receptor. Nature 498, 502-505.
Urbich, C., Dimmeler, S., 2004. Endothelial progenitor cells: characterization and role
in vascular biology. Circ. Res. 95, 343-353.
Valletta, J.J., Recker, M., 2017. Identification of immune signatures predictive of
clinical protection from malaria. PLOS Comput. BioI. 13, e1005812.
van Loon, W., Gai, P.P., Hamann, L.. Bedu-Addo, G., Mockenhaupt, F.P., 2019.
MiRNA-146a polymorphism increases the odds of malaria in pregnancy. Malar.
J.18.
Verma, I., Syngle, A., Krishan, P., Garg, N., 2017. Endothelial Progenitor Cells as a
Marker of Endothelial Dysfunction and Atherosclerosis in Ankylosing
Spondylitis: A Cross-Sectional Study. Int. J. Angiol. Off. Publ. Int. Coli.
Angiol. Inc 26, 3&-42.
Wagner, J.R., Busche, S., Ge, B., Kwan, T., Pastinen, T., Blanchette, M., 2014. The
relationship between DNA methylation, genetic and expression inter-individual
variation in untransfonned human fibroblasts. Genome BioI. IS. R37.
192
University of Ghana http://ugspace.ug.edu.gh

Wahlgren, M., Goel, S., Akhouri, R.R, 2017a. Variant surface antigens of Plasmodium
falciparum and their roles in severe malaria. Nat. Rev. Microbiol. 15, 479-491.
Wahlgren, M., Goel, S., Akhouri, R.R, 2017b. Variant surface antigens of Plasmodium
falciparum and their roles in severe malaria. Nat. Rev. Microbiol. 15,479-491.
Wang. Q., Liu, H., Xiong, H., Lillo Z., Wang, L.-E., Qian, J., Muddasani, R, Lu, V.,
Tan, D., Ajani, J.A., Wei, Q., 2013. Polymorphisms at the microRNA binding
site of the stem cell marker gene CD 133 modifY susceptibility to and survival
of gastric cancer. Mol. Carcinog.
Wassmer, S.C., Grau, G.E.R., 2017. Severe malaria: what's new on the pathogenesis
front? Int. J. Parasitol. 47,145-152.
Wassmer, S.C., Taylor, T.E., Rathod, P.K., Mishra, S.K., Mohanty, S., Arevalo
Herrera, M., Duraisingh, M.T., Smith, J.D., 2015. Investigating the
Pathogenesis of Severe Malaria: A Multidisciplinary and Cross-Geographical
Approach. Am. J. Trop. Med. Hyg. 93,42-56.
Waters, A.P., 2016. Epigenetic Roulette in Blood Stream Plasmodium: Gambling on
Sex. PLoS Pathog. 12.
Wattavidanage, J., Carter, R., Perera, K.L.RL., Munasingha, A., Bandara, S.,
Mcguinness, D., Wickramasinghe, A.R, Alles, H.K., Mendis, K.N.,
Premawansa, S., 2001. TNFa·2 marks high risk of severe disease during
Plasmodium falciparum malaria and other infections in Sri Lankans. Clin. Exp.
Immunol. 115,350-355.
Weiss, G.E., Gilson, P.R., Taechalertpaisarn, T., Tham, W.-H., de Jong, N.W.M.,
Harvey, K.L., Fowkes, F.J.l., Barlow, P.N., Rayner, J.C., Wright, G.J.,
Cowman, A.F., Crabb, B.S., 2015. Revealing the sequence and resulting cellular
morphology of receptor-ligand interactions during Plasmodium falciparum
invasion of erythrocytes. PLoS Pathog. II, e 1 004670.
Werner, N., Kosiol, S., Schiegl, T., Ahlers, P., Walenta, K., Link, A., Bijhm, M.,
Nickenig, G., 2005. Circulating endothelial progenitor cells and cardiovascular
outcomes. N. Eng!. J. Med. 353, 999-1007.
White, N.J., 2004. Antimalarial drug resistance. J. Clin. Invest. 113, 1084-1092.
White, N.J., Turner, G.D.H., Day, N.P.J., Dondorp, A.M., 2013. Lethal malaria:
Marchiafava and Bignami were right. J. Infect. Dis. 208, 192-198.
193
University of Ghana http://ugspace.ug.edu.gh

WHO, 2000. Severe falciparum malaria. World Health Organization, Communicable
Diseases Cluster. Trans. R. Soc. Trap. Med. Hyg. 94 Suppl I, S 1-90.
Widmer, R.J., Lerman, A., 2014. Endothelial dysfunction and cardiovascular disease.
Glob. Cardiol. Sci. Pract. 2014, 291-308.
Wilairatana, P., Tangpukdee, N., Krudsood, S., 2013. Definition of hyperparasitemia
in severe falciparum malaria should be updated. Asian Pac. J. Trap. Biomed. 3,
586.
Williams, T.N., Mwangi, T.W., Wambua, S., Alexander, N.D., Kortok, M., Snow,
R.W., Marsh, K., 2005. Sickle Cell Trait and the Risk of Plasmodium
falciparum Malaria and Other Childhood Diseases. J. Infect. Dis. 192, 178-186.
Wilson. N.O., Jain, V., Roberts, C.E., Lucchi, N., Joel, P.K., Singh, M.P., Nagpal, A.C.,
Dash, AP., Udhayakumar, V., Singh, N., Stiles, lK., 2011. CXCL4 and
CXCLI 0 predict risk offatal cerebral malaria. Dis. Markers 30, 39-49.
World Health Organisation, 2015. Guidelines for the Treatment of Malaria. Third
Edition. World Health Organization.
World Health Organisation, 2018. World malaria report 2018.
World Health Organization (Ed.), 2010. Basic malaria microscopy, 2nd ed. ed. WHO.
Geneva.
World Health Organization, 2016a. World malaria report 2016. Geneva WHO
Embargoed Until 13.
World Health Organization, 2016b. World Malaria Report 2015. World Health
Organization.
World Health Organization (Ed.), 2016c. Malaria microscopy quality assurance
manual, Version 2. ed. World Health Organization, Geneva.
World Health Organization, Global Malaria Programme, 2017. World malaria report
2017.
Wu, J.M., Visco, AG., Grass, E.A., Craig, D.M., Fulton, R.G., Haynes, C., Weidner,
A.C., Shah, S.H., 2012. Matrix Metalloproteinase-9 Genetic Polymorphisms
and the Risk for Advanced Pelvic Organ Prolapse. Obstet. Gynecol. 120,587-
593.
Wu, S., Hsu, L.-A, Teng, M.-S., Lin, J.-F., Chang, H.-H., Sun, Y.-C., Chen, H.-P., Ko,
Y.-L., 2012. Association ofSELE genotypeslhaplotypes with sE-selectin levels
194
University of Ghana http://ugspace.ug.edu.gh

in Taiwanese individuals: interactive effect ofMMP91evel. BMC Med. Genet.
13, 115.
Wu, X., Gowda, N.M., Gowda, D.C., 2015. Phagosomal Acidification Prevents
Macrophage Inflammatory Cytokine Production to Malaria, and Dendritic Cells
Are the Major Source at the Early Stages of Infection: IMPLICATION FOR
MALARIA PROTECTIVE IMMUNITY DEVELOPMENT. J. Bioi. Chern.
290,23135-23]47.
Wykes. M.N., Stephens. R., Cockburn, I.A., 20] 7. Adaptive Immunity to Plasmodium
Blood Stages. In: Malaria. Springer, Cham, pp. 47-66.
Wykes, M.N., Zhou, Y.-H .• Liu. X.Q., Good, M.F., 2005. Plasmodium yoelii can ablate
vaccine-induced long-term protection in mice. J. Immunol. Baltim. Md 1950
]75.2510-2516.
Yeo. T.W., Lampah, D.A., Gitawati, R., Tjitra, E., Kenangalem, E., McNeil, Y.R.,
Darcy, C.J., Granger. D.L., Weinberg, J.B., Lopansri, B.K., Price. R.N., Duffull.
S.B.. Celermajer, D.S., Anstey, N.M., 2007. Impaired nitric oxide
bioavailability and L-arginine reversible endothelial dysfunction in adults with
falciparum malaria. J. Exp. Med. 204, 2693-2704.
Yeo. T.W., Lampah. D.A., Gitawati, R., Tjitra, E., Kenangalem, E., Piera, K., Price,
R.N., Duffull. S.B .• Celermajer, D.S .• Anstey, N.M., 2008. Angiopoietin-2 is
associated with decreased endothelial nitric oxide and poor clinical outcome in
severe falciparum malaria. Proc. Natl. Acad. Sci. U. S. A. 105,17097-17]02.
Yeo, T.W., Lampah. D.A., Tjitra, E .• Piera, K .• Gitawati, R., Kenangalem, E .• Price.
R.N., Anstey, N.M., 2010. Greater endothelial activation, Weibel-Palade body
release and host inflammatory response to Plasmodium vivax, compared with
Plasmodium falciparum: a prospective study in Papua, Indonesia. J. Infect. Dis.
202, 109-112.
Zaina, S., Perez-Luque, E.L., Lund, G., 2010. Genetics Talks to Epigenetics? The
Interplay Between Sequence Variants and Chromatin Structure. Curro Genomics
11,359-367.
lakeri, S., Pirahrnadi, S., Mehrizi, A.A., Djadid, N.D., 20] 1. Genetic variation ofTLR-
4, TLR-9 and TIRAP genes in Iranian malaria patients. Malar. J. 10, 77.
Zeng, L., Cui, J., Wu, H., Lu, Q., 2014. The emerging role ofcircuiating rnicroRNAs
as biomarkers in autoimmune diseases. Autoimmunity 1-11.
195
University of Ghana http://ugspace.ug.edu.gh

Zhu, J .• Krishnegowda, G., Li, G., Gowda, D.C., 2011. Proinflammatory responses by
glycosylphosphatidylinositols (GPls) of Plasmodium falciparum are mainly
mediated through the recognition ofTLR2IfLRl. Exp. Parasitol. 128, 205-211.
Zbu, J., Wu, X., Goel, S., Gowda, N.M., Kumar, S., Krishnegowda, G., Mishra, G.,
Weinberg, R., Li, G., Gaestel, M., Muta. T., Gowda, D.C., 2009. MAPK
activated protein kinase 2 differentially regulates plasmodium falciparum
glycosylphospbatidylinositol-induced production of tumor necrosis factor
{alpha} and interleukin-12 in macrophages. J. BioI. Chem. 284, 15750-15761.
Zymo Research Corporation, 2014. Quick-gDNATM MidiPrep.
196
University of Ghana http://ugspace.ug.edu.gh

APPENDICES
Appendix I: Ethical Approval
NOGUCHI MEMORIAL INsnroTE FOR MEDICAL RESEARCH
EsUlbllsh«l , 919 A CoItsr/rutnr of rile CoIIc/IC of HeIItrh Sdcnccs University of Chana
Ph_ +233-302418431 (Ohc:t) +233-218-622574
Fix: +233-21-6021821513202 E-mail: ~."*..com.orv
My Refe'enc:e: OF 22
DaoieI Amoako-Sakyi, PbD Ua.iVVlity of Cape COlIC, Dept ofMicrobiololY IIId Imm\lllololY
NMlMR .... P. O. Box LO 511 LMaD,Accn 0IIInI
R£: OUr Study 1032/14-15 RESEARCH-IRS
At NOGUCHI MEMORIAl INSTITUTE FOR MEDICAl
Our DIllie! Amouo-Sakyi, PhD:
.... ung 01 .. : 11M014 RESEARCH-IRB ProIoc:oI TIUe:
At: NOGUCHI MEMORIAL INSTITUTE FOR MEDICAL
Genelc and Epigenetic baSil 01 EndoIIeUII Damage and Repair In cerebral Malllla In 0hanafM dllldren
ThIs lito aClvile you IIIat !lie IIbovI ref8renc:ecI study has been presenled 10 l1li InsUlullonal Review BoarcI, and tile following ICIIon lakin IUIIjtct 10 the condlUons and txptanaUon provldld below.
In .. mall: New AppI Expiration Date: 111412015 On Agenda For: InIIIaI SubmlUion Reason 1: Reason 2: OIacrlpllon: Dale RtceIvtcI- 1012112014 IRS ACTION: Con8ngtnt AIJprovaI Condition 1: Action ExpIanaUOR: The protOCOl WIll epprovtd IUIIjed 10 ICICIrestMIg tile comment below. AI the aIIbreYta1ions used In Ihelludy should be expllined upfIont
Yours SIncerely.
NMIMR-IRB IRB Adn*IIstrator
197
University of Ghana http://ugspace.ug.edu.gh

Appendix II: Scientific and Technical Committee Approval
NOGUCHI MEMORIAL INSTITUTE FOR MEDICAL RESEARCH
Administration (MeMO)
FROM STC COORDINATOR
TO MR. DANIEL AMOAKO-SAKYI
cc DIRECTOR, PROF. BEN aYAN
SUBJECT STCOUTCOME
DATE 13'" OCTOBER, 2014
", Its meeting held on 2nd October, 2014, the Scientific and TechnIc:aI Committee (STC)
reYleWed your protoalI entitled "Genetic and epigenetic basis d endothelial damage
and repair inc:erebnll malana In Ghanaian childrenw (STC] Paper 1(2)2014-15
and reconvnended that you make the following amendments to your proposal:
• The school c:hiIdren should be IIIected from sdIooIs dose to the hospiIaIs within
the catchment area.
• The sample sue calwlaticn should be provided.
• The study is a PhD work. Therefore, "Co-PIs- should be replaced with .. student supervisorsw.
• A consent fann should be provided.
I&rJJkm The proposal was approved subject to addressing the above concerns. The revIIa:I
version should be forwarded to the SfC Coordinator.
198
University of Ghana http://ugspace.ug.edu.gh

Appendix III: Inform Consent
Title: CircubtiaaJ eadolbelW celli :as die padIogeneSIS ot m.aIana
PriDap!llnvrstJgatoc Baa Gym, PlIO
Addreta: Dqw1maIt of ImmllDOlogy. NMIMR. BOll LG 511. Lqoo
1ll1o .... dD.: (1'0 be read or 1raDsbted to paJl'IllJ/ljUaJ'diDI ill tbeiJ own mother 1UDp)
o.ar Yotvnt.r,
lbJJ ~ fIxm coaWn. iD1bmWi0ll about the reSClJ"Ch cntrtled Cl1'cuialllt8 .1!doIM/Jal
c.1b tDtd Illtl ptllIJog.1IIIS1S olmaJiula III order to be 11ft that you are mformed about beang III
Ibis rese.arcb. "'" are ulciq you to read (or ha\'e rud to you) IhiI Coraent Form. You W1Il allO
be ukecI to AID it (or make your mart ia Jtout of. wtlJlen). We will give you. copy oflhil
form. Tlu. cOllSelll fIxm IIIiIbt COIItliD. some WOI'dI that _ un1'.uDiliar to you. PIeue ask \IS to
expbm anythmg you may DOt UIIdentaDd.
Why Ihls .ReIy Is pluned
Your child II bema .asked to participate iIIlbe above study ill order to ftad out fatton ill die
blood !bat may be of risk 10 wvere ~.an.a.. Mmria II QUJed by • germ that it pUled hID
ODe periOD 10 lilt olber by die bite of a mosquito !bat cames the maJ.aria germ. Mabna II •
very senoUI heUh problem ill GIIaria, as it is illlIWIY At1can COUDtria. We do DOt IaJow wily
some c:biIdtea become 1e1'eRly ill tom ~ari.a or wtay lOme of diose cbildrCII die hID
malan.a. To 1IIIlkntmd this problem we Deed to study childrm who come 10 tile IloIpital with
Ie\'ere malaria IIId compare !hem to c:hildrcn who have leu wvere ma1aria, IIId to other
cJuldren who are feeliDl well. The purpoae of Ibe study iI to tiDd out wII3I !acton they already
199
University of Ghana http://ugspace.ug.edu.gh

,
llave ill dIeir blood IIW may auIre III8a ~ &idt WIIaIIbcy IIIw IN\atIi. If_ caD II1II
die IIIIWU ID dIis qucstlOQ, 11ft: bope 10 be IbIe 10 IUQCIt _ lQ)'I 01 controUiD& NCta KWR
lidIDeua ill malaria.
c...aJ liii0 ..... 110. ud )'lIIlf put .. die oily
for a d1i1c11o quaW'y 10 be pan ofllus stud)' dial cluJd sbotIld be belwealdle .. 0I11IId 12
yean. If your cluldl\vard qrea 10 be ill die 1tIIdy, we "''ill collect veooUI blood .-pIe tbr
!aboralOf)' c1iapoaillll4 2 all [teatpOOII1\II) for our r_:IIth • the um. of admiIIIOII, 1 daya
....s 14 dIyI after rKOvay. 1f}'IIU a .. a i acbecIuJed follow-up viais (7 dIyI aad 14 days aftu
recovay), 1ft..., CIIIIW:t you aI bam. by pilon" or ill penoa 10 sc:Mdu.le moIber VIlA oIIId 10
tee If you IliII WIIIt 10 tab part ill the resean:h. WbeD lII1a COIIUct 11 made ),OU wjJJ DOt be
iMaIified as belDl illlIus rae.udl
1'bcre are 110 cbrect beDdlb 10 yout child &om tbia 1IUdy. However, billber p:amclp3!JOG may
IIrIp UI develop better malana trntment. He;she will DOt be paid for parucipabon m litis ItIIdy
but you will be relDlbuoed willi m amOUlll of fifteen Gbma cedis for your rune IIId nvel
dwiDa die follow up \lIIa.
'onllllllllsks
'lbll1IIIICIIIIII of blood coUeckd II 1wmIeu, altboup tIIa"e miY be allipt paiD IIId 1mIiIIDI
• die blcedmg lite. All IUbJCct! will receive approprim IftabDcIIl as DCCessuy. SIaiJe
! IleduliqIlClIIId diJpolable, siap-ule equipllleDt WIll be uaed II all lim ..
L_
200
University of Ghana http://ugspace.ug.edu.gh

Whlldnwat froa ...,.
We would like 10 _ dlallIuI study .. IInctIy ¥IIIuatary. Should tit cIIiId decide ... ID
PII1iciP*: it will 111,. DO Coaoequmces1br1lilMler. Sbauldlllno!unte .... lfaaypoialdtlliq
tile 1IDdy. decldc dial betsb. do !lOt willi 10 J*1icJpue lDy ftIo1ber. )'ClOI ate ltft 10 IaII1iDMt
die panicipatioa. dll!ctive iawcdulely. Ally __ decuioo will be RIpICted wiIbout l1li)'
ftIo1ber ~ y_ dcdIiaD wiIJ DOl aJIKt tbc bcaltb care you would DmIIIIIJy RCCi\'e.
Visits
If lilt cbiId _ • scbeduJed YIIiI. ... may CIIIIIact you II ~ by pIIaac. or ill ~ 10
IdIeduIe lIlotbcr Wit aDd 10 tee ilyou IIiII WaD! to tile. part illlIIe martb. Wbcn IIIiI COIIIII:\
is made you wilIlIOl be KIaIblied u beiDa illlIIiI martb.
CoddeadaJlly
AU iIII'IlnuIIiclG pIbcftd would be lruted iD IIrict caafidcaIiaIity. We wllJ protect UlformalJoo
about yoar clIiI4l1ki11a pan ill !lui racarcb 10 IIIc belt of our ability. The cbiId will DOt be
lIIIDed iD l1li)' reporta. line, ..... tile lid' of (lilt a111l0llpl dial may ICCm lilt rese.cb
records) may IOIIIeIimes look at bJs,ber retCilfcb ruordl. If you bve my qu<.b .... please
feel fttc 10 Ilk die pllyslcilll .. dIaq. Someooc 6'om die IllB or Edu~ CommiIIee miIbt
W1IIIIO ask you qllClUOUS about beiDllD IIIc rescarcb, but you do IlOl have 10 __ them. A
_ 0(0 (ould order medical records sbowD 10 OIlIer people. but lllal islllllllcely.
COAIaca: If you ever ba\'e l1li)' qucstiaal about lilt raan:b INdy or lIIIdyoftiaed problllllS,
you may CODtaClDI'. M2ame Yu Nyllko at PriDe. Mme l.ouiK HotplIaJ (Tel: 0244 OIlSS.)
or Dr.Bm Gym of !be Nopcbi Memorial Institute for MedicallW .... cII (0244 726016) II
201
University of Ghana http://ugspace.ug.edu.gh

aylilllC. for quatiou about the edIicaI upectI oflbis INdy or your nIhIIu a \'Oluzuecr,
you l1l.I)' COIDCt Dr. SaDtl A~ CIwnuaa, IuciIuliaaaJ IteYIew Bon,
NMIMR. Uniwnity of Gbaaa (021 50117119) or CIIalnIw! of die GIIua HaI1b s.w:c EIIUcal Coau!udee (Tel 021 611109)
V,ar rtph as a parddpul
1bit research bas beaI re"eMd aad apprO\'" by die NMIMR IRB aad Gballa Heal1ll Service
Etbical CommiIIee. AIIIRB or Etbical Committee is a committee Ibm reviews RUarch ItUdieI
ia order to help protect patIicipaII. If you have .y quatiou aboul yoar riPlI II I research
paniapallt you may CODtact [Dr. Samuel Ayete-Nyampoaa, Tel 21·501·1781179 or CIIainDaD
oldie Gbana HaI1b Service E1IIiI:al CaauJUIIee (Tel 021 681109)
VOLUNTEER AGRIDlENT
The abovt documrnl delcnbllli die baIefits, riW aDd procedura for the research tit1e
CImtkltIlflIlldotMlI4I ellis fWI 1M ptJIltopl/lS/s ofWIQIar/Q lw beaI read aad explaiaed to
me. I have been P\'COIII oppclIIUDIty to have lIlY queItiou about the resean:b "''''Creel to
II)' salisfactioo. lqree my child/ware! to particiJllle as a vo1uateer.
Date
II volaa ..... '. Pll'lllrlGaardlu ClllDot rtad the 10,. th_seh'n, a wilaUI mast lip
ben:
I was present while the beadits, rUb II1II pruc;edum were read to Ibc \'OIuateer. All questioIu
were IIISwered md Ibc VOluotcer'l Guardtm'Pareat bas qreed to take part ill Ibc research.
202
University of Ghana http://ugspace.ug.edu.gh

I ~ lballbe IIII1n ., PIII)lOIe. die poccatIaJ beae1Its, IIId poaiblc nib ulOdated willi
panicipItIq Ia dtiI.-.:h IIIw beai apWaed to the above iDdIviduli (TdIdI, 2014)
Siptw't PetIOlI who obtained COOlellt
203
University of Ghana http://ugspace.ug.edu.gh

uIIs IIIId tltI pathor-b~""" Ia Older til be IIIR .. )'011 are iDIbnDed abouI beiq ill
Ibu raardI, we are IIkiDI yoa til read (or llave read til yov) dIiI ~ F-. Y0II1riII aIIo
be liked tlllIlD it (or mae your IUrIi: ill ht of a willlen). We \diM }'VII a copy oflllil
fbrm. nw cCllJCDt tbnn might COOtailllOlllC wordI that are ~ to you. PIMIe alii: us til
explailllll)1llilll you may DOC UDdentaad.
WIt,. dIU HIldy b pluald
Your child II beiDa asked to panicipete m Ibe Ibovc ltUdy ill order til find out factDn ill Ibe
bIIIod dial l1li)' be otrilk tIIlCwre IDllari&. Mmria II CIIIICd by a Jam_ il pwcd hili
CIlIa paIOIItII Ibe other by tbe bite of. lIlOIquito that carrie. tile IIIIIIria ImIL MaJana II •
\'eIY smOUl beaIIh problem ill GIwII, as it is ill _y Alhcaa cOUDlriet. We do DOt bow wily
lome cIIIIdrcu become severely ill li'om malaria or wtIy lome or those c:Iu1dm1 die rtom
malaria. To IIIIdcntaDd tIIis problem we Deed to study cIIildreo who come to tile bOlpital With
severe malaria and compare them to t:IIiIdreD who bave las severe mabria, and to other
cbildreD who are t'eeIiDg well The purpose ottlle study Is to ftDd out wbat tmon they alre.,.
ba\'e ill !lleir blood thai may make them severe1y lick \\1Iea they have ma1aria. IIwe CID ftDd
!lie __ to !Ius questioo, we hope to be able to SlIgest Dew ways ofCODInI11iDa such m"ere
liclaltua ill malaria
CfttnilDlormalloD ud Y08r pan ill die mad)'
For a child to ~ to be part oftllis ltUdy thai child IbouI4 be bctweeo the aaes or I and 12
)'arI. II r- c:IliIcIIwwd asrees to be ill the study. we will colla:l VerJO\ll blood Nlllple tor
laboratory diagnOSIS aod :I all (taspooDfW) for our NMarth irIitiaIJy and 7 day. and 14 days
later. II you mias a scbeduJed follow-up visitI (7 days 1114 14 days) ill your 1Cboo1. we may
COlltact}'VII at lIome by pboaa. or m paIOII to Icbedule lDother VIIi! 1114 to _ it you JtilI_t
204
University of Ghana http://ugspace.ug.edu.gh

to take part ill 1M researdL WIleD thai coatact il made you will IlOl be 1deaIified &I bciq In
dlilmardL
Possible Beadh
'I'1In IRlIII direct baIdiII to y_cbiJ4 a- tbu 1tUdy. Howna' ...... pllltJclpatlOa auy
bclp UI develop bcner JDaIaN treatmenl Helshe WIll nOl be paid tar partlcJpatiOll ill thai ltudy
but you WIll be reimbuned wiIIIm lIII0II1II of fifteen Gbaaa cedis tar your lime II1II IrInl
clurmg die follow up VW!l
Possible lUaks
The amollDt ot blood c:oIIec&ed is banDIcA. aJIbousb dIcrc may be a IIiIbt paIIl OIIId bruiliq at
the bleedirll lite. AU lUbJecti will recein appropnate trutment &I aeceuary. StmIe
tecbDJques mel dlspoqble.liDgIe-UK equlpmeDI will be used at all blDcs.
"'ltbdrl.11 fro. ,"dy We would like to ... lllat dIiI IIUdy .. strictly vaIuaIary. Sbouid the cbild decide !lilt to
pirtle 'p*; it WIll !law DO consequcm:ea Col JWn,1u:r. Should 1be voluateer. at my poiat cIunI:Ii
dae 11Udy. decide 1ba helshe do not wish to participate III)' 1IIr1ba. you 1ft he to tCl1l1llllle
the partJClpatson, dfedive imIIIediardy. All.y such decision WIll be rapectm without III}'
ftJnbu diKusaiOll. Your deci.ioa WIll DOl a1I'ect the beaJth care you would normally receive.
\'blll
20S
University of Ghana http://ugspace.ug.edu.gh

(fllle dulclllUSlt'S a sdIeduIed V1IiI. WI..." COIIIICt yw .lIome II)' pboDc. or ill pcncIIIlD
ICbeduIe IIIIIIber viIilllld to I« if you IIiIlwaal.,,,, part ID IIIe re-aa. ..... IblICOIIIICt
IS made you "ill DO( be idmtified u beiaa ill 1bia-.dL
CealldeatbllJly
AU illformatioa pdIemI woaId \Ie lUatN ID IMrict COIIIIdaatiaIit· We wiD pI'GCIct iafbnDatioa
about r-- child '*iDa pelt ill duJ rnadllD IIIe \lett of our ability. TIle child wiD DO( \Ie
DDed m lIlY repIIItI. Howevcr,Ibe lWI'ot[lllt Illlf011p1tbat may _e.. die mcarchftCClfCbj
may _ts loot IlIIiI/IIIr ...-cIa ncords. (fyou bave lIlY queatioa.s. pleue Cede-to
uk Ibe physlciIII ill cbarp Someone tam IIIe 1RB or Ethical COIIUDJIIee IIIiPt WlDt to uk
you questlClIII about bciq mdle racardI, bile you do DOl bave to IIIIMr tb_. A court otlaw
could order medial _cis sbowa to otber people, but !bat II uaIikeIy.
Coautts: ltyou C\'a" ba\"e lIlY qucstioaa about Ibe researell study or 1II!dy«lated problema,
you may ccaact Dr. M-. Yu Nyllto. PriDce Marie Louiac HOlpiIaJ (Tel: 0244 018888)
or DrBea GYID oflbe Nop:lu MaaoriaJ lDstJtUte for Medial Jtesearch (0244 720016) at Illy
tuDe. For quutioas about die etJucal upccu of tb.iJ study or your nabtt u a volualeer, you
may CODtact Dr. Samuel Ayete-Ny1lJlPOlll, CIwnDaa. IJllIiluIioGa1 Review BoaId. NMIMJt..
Umvlllily of Gbma (021 50117119) or CbainDaa oldie GbaIIa Healtb SeMce Etlucal
C4lnllllJtt~ (Tel. 021 681109)
V ... r ....... as I partldpu.
Tbia _cb bu bcea reviewed mel approved II)' die NMIMJt 1RB IIId Gbaaa HRIIh Service
EdIicaI Commil!ft. Au IRB or EtbicaI Committee II a commillee !bat r • .,.\eWI reautcb .tudies
in order 10 bcIp protect partlapaDII.ltyou bavc any questiCIIII abollt r-- rilbll u a resean:ll
206
University of Ghana http://ugspace.ug.edu.gh

~_you..,cOIIIaCI[Dr.s-JA~Tdll.S01·1711179orCllaJnJuo
GIlle CiII.aoa HeaJtlI Sen"ittElllit:alc-illel (Tel 021611109)
VOLUNTEER ACR!:DlDO"
Tbe above documeaI duaibiDc !be beaefiI.I, rIIU aDd proce4ura I'« die reMIIdI title
CIn:ulDlIIIr I1ItItItJwIItII alb fftI 1M pt1IJtGptwts ~".."", .... been IUd IIId cap1aiDed 10
me. I have been SWeD an oppor\Ufti1y to .\'C DY quntiaDI abouI!be reulld!lM\\'end 10 my
satut.cta0lL I apee lIlY cbildIwIrd 10 pIIticipUe U I \·olwlleer.
U ftJurMr'1 P ..... lICu.rdJaD .... er nad tIM r. .. $ .......... wtnaaa .ut lip
11 ... :
1_ pmaII wbiIe die beaeftta, rilb ami procedum were read 10 !be \'OIlIIIteer AU questlODJ
MR aaswae4111d !be VOIullllMr'I GuudlmlP:armtlw qreed 10 take JII'lID die rese¥Cb.
I ~ tbat die DaIUre aDd IJUI'POK. die poIaItlII baellb, aDd possible rIIU UIOCiaIed mill
PIltlClpalllll in tbiI raean:II have bem expIaiDed III die above iadividuaJ (TeIIeb, 2014)
207
University of Ghana http://ugspace.ug.edu.gh

Appendix IV: Primer for iPLEX Reaction
ExIaIded Pr1I1ICI Forwn Primer 10 Forw.d Primer Sequcnc;e Rcvcnc Primer ID Reverse Primer Sequence ID ElltcDdcd PriJDcr Scqucncc
1 .u7209I_WI_F
2 n2ai416_WI_'
3 ISIOI9994o_WI_F
• 1S2304S27 _ WI1
, n943012_WI_F
6 1II1'm22_WI_F
7 ",UI900'85_WI_F
a n207IS59_WI_F
9 n3911211_WI1
10 1I2l747S'_ WI_F
11 nQ524054_WI]
12 ",'T.M22262_WI1
Il n.591155740_ WI_F
14 ",16I31S32_ WI_F
IS rsllOO783_WI_F
16 rs56213 104_ WI_F
17 nQ070744_WI1
18 rs606630J_ WIJ
19 nI331l099_WI_F
20 1'110489181 WI F
21 .. 7604879_WI_F
22 n42J6Ol!.1 WI_F
B n3918256_WI_F
ACGlTGGATGAOOATAGTCCTAGACA(JCAC n37209I_ WI_R
ACGlTGGAroccACA1T1'CCACCACTATCC n2236416_ WUt
ACGlTGGATGTC'1OOro1'CACTGC 1S10000000_WI_R
ACGTTGGATGGGAAGACCCTCTTT ACCTTG B23CMS27 _ W 1_ R
ACGlTGGATGGGTGACAACACTGAGG'CTG n943OI2_WI_R
ACGTTGGATGAGAGACCTCMTGTCCACAG 111176722_ WI_R
ACGTTGGA1'G11CAACTCTGCTATACAC ISI000000S_WI_R
ACGTTGGATGATCAGAAAACGCACTTGCCC n207IS59_WI_R
ACGTTGGATGAGACCTAOOroCAGGACA TC n3911211_WI_R
ACGTTGGATGTTCGACGATGACGAGTTGTG rs2274US_ WI_R
ACGTTGGATGMCACAACCTGACTTTTACG nQ524054_WUt
ACGTTGGATGGACTGAGGT MAAGGACTGC 1573422262_ WI_R
ACGTTGGATGMCATGGTGAAACCCCCTCT n.59IISS740_WI_R
ACGTTGGATGGGTCATAGTTAAAGAGACCG nl683IS32_WI_R
ACGTTGGATGAGTCATCCTTGGTCATGCAC nl800783 WI_R
ACGTTGGATGTCCAGAGTGGGCTCCTT AC rs5623JI04_WI_R
ACGTTGGATGACCAGGGCA TCMGCTCTTC rs2070744 _ WI_ R
ACGTTGGATGCAAGGGCAGCACT A TCTTGA rs6066303 _ W I_R
ACGTTGGATGT AGMCGAT AAGGAGGGTGC nl3JI 3099_WI_R
ACGTTGGATGCATGMTGCMGGGTTTGTC nICNI9IBI_WI_R
ACGTTlrtiATGTGC AGC AAACGGAAACT MC .. 7604179 WI .R
ACGTTGGATGGGAGGTTTGAAAGTTGCCAG 1'14236014 WI R
ACGTTGGATGTTGACAGCGACMGMGTGG nJ918256_ WI_R
ACGlTGGATGGACCAA TGAGCAA TGGCT AC rs37209I_ WI E GTCCCAGCAGCAT
ACGTTGGATGGACACCAGACCAAGGAAGAG n2236416_ WI_E MAGGCCGAACCT
ACGTTGGATGTGCAGTCCAGCACAGMAAG .. 10199940_ WI_E GAGCCTGGGAGTCT
ACGlTGGATGATAGACMGGCGATroccTC n2J04S27 _ WI_E 1TGCCTCI1TCAGCT
ACGTTGGATGCCTGAAATTTCTCAGCCAGC n'M3OII2 _WI _ E GTGGCAGAGTGAGCC
ACGTTGGATGCACCACCATTGGGTTAACTG "'176722_ WI_E TGTCCACAGTCACTCG
ACGTTGGATGATTGCACTCCAGCCTGCCT rsI09OO585_WI_E MGGAGTCTCACTCTT
ACGTTGGATGCTAGGCAGGTCACTTCAAAC rs2071S59 _ WI_E GGAAATAGCGGGMTG
ACGTTGGATGGTCTGCAGGACGTTGGTTG .u911211_ WI_E aTTGTCTGCGGCGATGT
ACGTTGGATGGGGAGAGMTGMGGGM TC 1S22747SS_ WI_E TGGGCAAGOOCGTCGGT
ACGTTGGATGTATGGGMGCTCT ACCACAC rs2524054 WI_E TGACTTTT ACGATCATCA
ACGTTGGATGATTGCATGCCTM TAGGAAC rs73422262_WI_E GCCTMTAGGMCCA TCCA
ACGTTGGATGTATGATCTTGGCTCACTGCG rsS90SS740 _ WI_ E IIIjIGATT ACAGGCCACCA
ACGTTGGATGCTTTCAGATAGACAAAGTG rsl6831S32_WI_E GACAAAGTGAAAACAAAA T
ACGTTGGATGATCCAGCCCCTACTTTTCAG nl800783 _ WI_E ggcgGGAGGAGACAACAGA
ACGTTGGATGTCCCGAGTTCTGGGCA TTTC rsS62J3104_WI_E cas-AGCACCTTGCTCTGCAT
ACGTTGGATGCTGTCA TTCAGTGACGCACG rsl070744 _ WI E ctnCTGAGGCAGGGTCAGCC
ACGTTGGATGAAACCGGAGGCTATTTGTG rs606630J_ WI_E GGCTATTTGTGTAAATGT AM
ACGTTGGATGGACTCGT AACTTAC ACCCTC 1513313099_ WI_E aGATTTT AGA TCACCTTAACTC
ACGTTGGATGAAGATAAATGCCAAGGGGAC nl0489181_WI_E AAAGTACAGGAGATA TT A TGGG
ACGTTGGATGCTCTGTGTACCTCAGA TTGG ",7604879_ WI_E AA TT A TTTGTTCTCAT MGTTTG
ACGTTGGATGTGCTGCTTTCACTCTCACCC 1'14236084. WI.E agallTCACTCTCACCCCCGCCCC
ACGTTGGATGACAAACTGT ATCCTGGAGGG n3918256_WI_E cce<:GCCCCGGAGCCGCGGGACCA
.~ _L_
University of Ghana http://ugspace.ug.edu.gh

24 rs3917419_WI_' ACCil'TOOATOCiC'TCATCTC'TCACCTTTAAC rs3917419_WI_R ACGTTOGATGAAAOOGGTAGAATTACAGTC rs3917419_WI_E ... TTTAACNJATAAGAACACTG
2' ral07'73II_III_F AC'GTTOOATGAQATCGATCTGATGACAAa: .. 1077311_WI_R ACGTTGGATGTTCACACTATCa:CAGGATG .. 1077311_ WI_E -.croTGAAA TGGGAA TGATA
:Ii6 .. 1466S41_WI_F ACGTTGGATGAAGAGT AAGGGGT AGGGAAC .. 1466S48_WI_R ACGTTGGATGTCCAGTAATATCAATGGAOG .. 1466548 WI_E apaGGAGGATATA TGTGTGCA T CTTAAGTTCTGCTTTTAAAAATAT
27 11614951 WI F ACGTTGGATGTOGTTGAGCTTAAGTTCTGC n6lI49S1 WI R ACGTTOGATGGTAAGTGACCTAAACCTTGC n6U9S1 WI E A
University of Ghana http://ugspace.ug.edu.gh

Appendix V: iPLEX protocol
Table I: Multiplexed peR cocktail, without DNA (Same multiplexed assays, different
DNA)
.'" volume Reagent Concentration In 5 J&l (1 rxi!1 .
Water (HPlC grade) NA 1.8 JIl 10x PeR Butrer with 20 mM MgCI2 1x (2 mM MgClz) 0.5 JIl MgCI2 (25 mM) .. 2mM OA JIl dNTP mix (25 mM each) ••• 500 pM 0.1 III
Primer mix (500 nM each) 100nM 1.0 III
peR Enzyme (5 UlIll) 1.0 Ulrxn 0.2 III
Total VOlume: 4.0"l
The SAP mix should be prepared as below and can be scaled up to encompass more
samples. Table2: SAP enzyme solution
'" ... '." ,
Reagent Volume (1rxn)·
Water (HPlC grade) 1.53 J&l SAP Buffer (1Ox) O.17~
SAP enzyme (1.7 U/JlL) O.30~
Total Volume 2.00 J&l
RNgII'It Cone. In 9 III Volume (1rxn,' WIII1It (HPLC grade) NA 0.G19 III
FlEX BuIll Plus (10x) 0222)( 0.200 III IPlEX Tennmlion mix 1x 0.200 III Pm. mil: (' pM: 10 pM: 15 pM)" 0.52 pM:1.04 I'M: 1.57 I'M 0.940 III
IPlEX 8ftQme 1x 0.041 III
Total VoIl.IIle: 2.000 III
Add 2 ul of iPlex-SBE master to each well of the SAP treated plate (total vol. 9 ul).
Vortex and centrifuge
210
University of Ghana http://ugspace.ug.edu.gh


















