
University of Nigeria Activity of... · 2015-08-28 · University of Nigeria Research Publications...
105
University of Nigeria Research Publications Author NJOKU, Ugochi Olivia PG/M.Sc/03/34258 Title Antioxidant Activity of Seed Extract and Fractions of Monodora Tenuifolia (Annonaceae) Faculty Medicine Department Pharmacology and Toxicology Date February, 2007 Signature
Transcript of University of Nigeria Activity of... · 2015-08-28 · University of Nigeria Research Publications...

University of Nigeria Research Publications
Aut
hor
NJOKU, Ugochi Olivia
PG/M.Sc/03/34258
Title
Antioxidant Activity of Seed Extract and Fractions of Monodora Tenuifolia
(Annonaceae)
Facu
lty
Medicine
Dep
artm
ent
Pharmacology and Toxicology
Dat
e
February, 2007
Sign
atur
e

ANTIOXIEANT ACTIVITY OF SEED EXTRACT AND FRACTIONS OF Monodora tsnuifolia
(Annonaceae)
NJOKU, UGOCHI OLlVlA PG/M.Sc/03/34258
A PROJECT SUBMITTED TO THE DEPARTMENT OF PHARMACOLOGY~~OXlCOLOGY, UNIVERSITY OF NIGERIA, NSUKKA IN PARTIAL FULFILLMENT OF
THE REQUIREMENT FOR THE AWARD OF MASTERS OF SCIENCE (M.Sc) DEGREE.

TITLE PAGE
ANTIOXIDANT ACTIVITY OF SEED EXTRACT AND FRACTIONS OF Monodora tenuifolia
(Rnnonaceae)
A PROJECT SUBMITTED TO THE DEPARTMENT OF PHARMACOLOGY/TOXlCOLOGY, UNIVERSITY OF NIGERIA, NSUKKA IN PARTIAL FULFILLMENT OF
THE REQUIREMENT FOR THE AWARD OF MASTERS OF SCIENCE (M.Sc.) DEGREE
NJOKU, UGOCHI OLlVlA DEPARTMENT OF PHARMACOLOGY/TOXlCOLOGY
UNIVERSITY OF NIGERIA, NSUKKA
SUPERVISOR: Prof. P. A. AKAH DEPARTMENT OF PHARMACOLOGY/TOXlCOLOGY UNIVERSITY OF NIGERIA, NSUKKA

CERTIFICATION
NJOKU, Ugochi Olivia, a postgraduate student in the Department
of PharmacologyiToxicology and with registration number
PG/M.Sc/03/32458 has satisfactorily completed the requirement of
research work for the degree of Masters of Science (M.Sc) in
PharmacologyiToxicology.
This work embodied in this project is original and has not been
submitted in part or full for any other diploma or degree of this or any
other University. 5 F t i ,. .
roject Supervisor s Head of Department

iii
DEDICATION
To my beloved husband, great and wonderful kids: Onyii, Obi. Jnr. &
Chioma, parents and loved ones.

ACKNOWLEDGEMENT
My most profound gratitude goes to my supervisor, Prof. P.A.
Akah for his warmly guidance and supervision. His unreserved kindness
and understanding not only inspired me but also encouraged me
throughout the trying moments of this work.
1 also wish to express my indebtedness to Dr. Charles Okoli of the
Department of Pharmacology and Toxicology who by close supervision
and assistance contributed in no small measure to the completion of this
work.
My gratitude goes to Okoye and Ajaba for their unrelentful
assistance throughout my laboratory work.
To my husband, Prof. 0. U. Njoku, my kids: Onyinye, Obi Jnr.,
and Chioma, I really appreciate your moral and financial support.
My'thanks also goes to my caring colleagues, friends who have
contributed inestimably throughout the course of this work. In this
respect, I am particularly grateful to Ada Onyirioha, Thelma Ihedioha,
Ada Ibiam, Uju Udeogaranya, Prof. Mrs. E. Okeke, and a host of others.
I thank my parents: Elder and Elder (Mrs.) Ibe, my parents' in-law:
Sir and Lady Rufus Njoku, my siblings: Paul, Uloma, Nwajiulo, Ngozi,
Jude, Chizoba, and Tochukwu who have been an everlasting source of
love, strength and encouragement.
Finally, above all, to my gracious heavenly father for the grace,
strength and ability, for perfecting all that concerneth me to him be all the
glory thanksgiving and honour.

TABLE OF CONTENTS Page
List of Figures .................................................................................... viii
................................................................................................ Abstract X
CHAPTER ONE
1 . 1 Antioxidant: An Ovef-View ................................................... 1
1 . 1 . 1 Natural Antioxidant of low and high molecular weight 2
1 . 1 . 2 Mechanism of action of antioxidants .............................. 5
1.2 Lipid Pef-oxidation ................................................................. 6
1.3 Mechanisms of lipid peroxidation induction .................. 7
........................................................................... 1 ~ 3 - 1 Autoxidation
1.3.2 Photo-oxidation .....................................................................
1 a3.3 Enzymatic peroxidation ......................................................
1.4 General antioxidant actions ...............................................
1.4.1 Free f-adicals ..........................................................................
1 a4.2 Antioxidant mokcules .........................................................
1.5 Oxidative stress .................................................................... 1.6 Diseases associated with free radicals ..........................

Flavonoids ..............................................................................
Glycosides ..............................................................................
Cardiac glycosides ...............................................................
Steroidal aglycone ................................................................
Saponins .................................................................................
Tannins ....................................................................................
Carbohydrates ....................................................................... Literature survey of Monodora tenuifolia plant ............
Characteristics/Morphology of Monodora tenuifolia ...
HabitaVEcology of Monodora tenuifolia .........................
Local names ........................................................................... Pharmacological uses of Monodora tenuifolia .............
Aim of research ..................................................................... CHAPTER TWO
Materials and Methods ....................................................................
2.1 Materials .................................................................................. 2.1 . 1 Phnt materials .......................................................................
2.1 2 Animals ....................................................................................
2- 1 a3 Chemicals ............................................... : ...............................
2.1 -4 EquipmenVApparatus .........................................................
2.1.5 Preparation of reagents for Phytochemical Analysis ..
2-2 Methodology .......................................................................... 2.2. Extraction of oil from Monodora tenuifolia seeds .am.....
2.3 Thin-layer chromatographic technique ..........................
2.3.1 Determination of lipid components .................................. 2.4 Determination of vitamin in Monodora tenuifolia .........
2.5 Phytochemical analysis of Monodora tenuifolia seed
2.6 c o h l n chromatographic technique ............................... 2.6.1 Elution of the column ...........................................................
2.7 Nitric oxidellipid peroxidation scavenging assay ........
2-7-1 Scavenging of nitric oxide test ..........................................

vii
2.7.2 Carbon tetrachloride induced lipid peroxidation test
2.8 Statistical analysis ................................................................
CHAPTER THREE
Results ....................-......................~~....--.......-....-.-~~.....-.-....-.--....*..-..... 3-1 Percentage yield of extract .--*.-.......-.----....----....-.--.**.----....-
3.2 The result of the phytochemical analysis of Monodora tenuifolia seed ..................................................
3.3 The result of acute toxicity test: Acute toxicity (LDS0) of the extract ..........................................................................
3.4 Nitric oxidellipid peroxidation scavenging ...---.-...----.-..--
3.4.1 Effect of pet-ether extract and fractions F1 and F p on Carbon tetrachloride-induced lipid peroxidation in rat Liver homogenate .................................................................
3.4.2 Effect of nitric oxide scavenging activity of pet-ether extract and the fractions ....----.....---.-..-.---. * .----...----..*.---......-
3.5 Result of the lipid compounds present in the seed extract ......................................................................................
CHAPTER FOUR

viii
LIST OF FIGURES
Figure 1 : Shows a balance between Oxidative Stress and defense system ........................................................................ 2
Figure 3: Effect of extract and fraction on carbon-tetrachloride- Induced lipid peroxidation in rat liver Homogenate ....... 75
Figure 4: Effect of Extract and fractions on nitric oxide Scavenging activity ................................................................. 78

LIST OF TABLES
Table 1: Shows the Radical Reaction Potentials ........................... 20
Table 3.1 Results of Phytochemical Test ............................................ 70
Table 3.3 Effect of Crude Extract and Fractions on CCI4-lnduced Lipid Peroxidation ........................................ 74
Table 3.4 Effect of Nitric Oxide Scavenging activity of Crude Extract and Fractions ............................................................. 77
Table 3.5 Result of Vitamin Assay of the Extract ......................... 79
Table 3.6 Result of the Lipid Components present in the seed extract 80

ABSTRACT
The antioxidant activity of the seed extract and fractions of
Monodora tenuifolia (Fam. Annonaceae) was evaluated. The Monodora
tenuifolia seed was extracted with pet ether 40-60°C to produce the
crude extract. Fractionation of the extract by column chromatography
using pet ether 60-80°C and diethyl-ether produced 2 fractions (F1) and
(F2). Phytochemical analysis of Monodora tenuifolia seed extract
showed the presence of some plant secondary metabolites, viz:
alkaloids, flavonoids, proteins, carbohydrates, saponins, glycosides,
cyanogenic glycosides, cardiac glycosides, tannins, steroidal aglycon
while, 0 and C glycosides, anthracene glycosides and reducing sugar
were absent. The 3 fractions showed the presence of vitamin A and
vitamin El The pet-ether extract and the fractions (F1 and F P ) reduced
CCle-induced lipid peroxidation in rat liver homogenate. They also
exhibited significant antioxidant activity in nitric oxide induced lipid
peroxidation. The crude extract and diethylether fraction (F2) produced
dose-dependent protective effect against lipid peroxidaiton and free
radical generation in liver homogenate. The acute toxicity study with the
crude extract showed no signs of obvious toxicity up to a dose level of
5000 mglkg. These results suggest that Monodora tenuifolia seed
extract possessed significant antioxidant properties and could be used
for the treatment of diseases associated with free radical generation.

CHAPTER ONE
1 .I Antioxidant: An overview
Antioxidants are a group of substances, which when present at
low concentrations, in relation to oxidizable substrates, significantly
inhibit or dctlay oxidative processes, while often being oxidized
themselves (Kanner et al., 1999).
The application of antioxidants are widespread, in industries they
are used in preventing polymer from oxidative degradation, rubber and
plastic from losing strength, gasoline from autooxidation, synthetic and
natural pigments from discolouration and as additives to cosmetics, food
(especially food with high fat content) beverages and baking products
(Kanner et a/, 1999).
In recent years there has been an increase in the application of
antioxidant in medicine as information is constantly gathered linking the
development of human diseases to oxidative stress (Halliwell et a/.,
1999). The generally accepted hypothesis in any biological system is
that, an important balance must be maintained between the formation of
reactive oxygen and nitrogen species (ROS and RNS, respectively). The
reactive species such as superoxide (02)) hydrogen peroxide (H202),
hydroxyl radical (OH), nitrogen oxide (NO), and hypochlorous acid
(HOCI), are all products of normal metabolic pathways of human organs,
but under certain condition, when in excess they can exert harmful
effects.

To maintain an oxido/redox balance, organs protect themselves
from the toxicity of excess ROSlRNS in different ways, including the use
of endogenous and exogenous antioxidants.
OXIDATIVE STRESS DEFENSE SYSTEM I I
- Mitochondria - Radiation - ~araoxonase ' - Vitamin C ' - Peroxisomes - Ozone - Glutathione- - Flavonoids
* -Inflammatory Cells - Xenobiotics peroxidase - Glutathione -Superoxide
dimutase
Fig. 1: Shows a balance between oxidative stress and defense system
. 11.1 Natural antioxidant of low and high molecular weight
Naturally occurring antioxidants of high or low molecular weight,
can differ in their mechanism and site of action (Sahart, 2001). They can
be divided into the following categories: -
(a) Enzymes
(b) High molecular weight proteins
(c) Low molecular weight antioxidants

a) Enzymes: The best studied cellular antioxidants are the enzymes,
superoxide dimutase (SOD), catalase and glutathione peroxidase (GPx).
These attenuate the generation of reactive oxygen species by removing
potential oxidants or by transforming ROSIRNS into relatively stable
compounds. SOD, which was discovered in the late 60s, catalyzes the
transformation of the superoxide radical into hydrogen peroxide, which
can then be further transformed by the enzyme catalase into water and
molecular oxygen (Sahart, 2001). Glutathione peroxidase (GPx)
reduces lipid peroxides (ROOH), formed by the oxidation of poly-
unsaturated fatty acid (PUFA) to a stable, non-toxic molecule hydroxyl
fatty acid (ROH) (Sahart, 2001). Less well studied (but probably just as
important) enzymatic antioxidants are the peroxiredoxins and the
recently discovered sulfiredoxin. Other enzymes that have antioxidant
properties (though this is not their primary role) include Paraxonase,
Glutathione - strainsferases, and aldehyde, dehydrogenases (Current
Medicinal Chemistry, 2005).
' b) High molecular weight proteins: These preventive antioxidants
hinder the formation of new ROS. These antioxidants are protein that
binds ROS to protect essential proteins. The group includes albumin,
metallothonine, transferring, ceruloplasmin, myoglobin, happtoglobin and
ferritin (Current Medicinal Chemistry, 2005).
These are all present in plasma and bind to redox active metals
and limit the production of metal - catalyzed free radicals (Current

Medicinal Chemistry, 2005). Metals such as iron, copper, chromium,
vanadium and cobalt are capable of redox cycling in which a single
electron may be accepted or donated by the metal (Current Medicinal
Chemistry, 2005). Albumin and ceruloplasmin can bind copper, ions, and
transferin binds free iron. Haptoglobin binds heme-containing protein
and can thus clear them from the circulations (Current Medicinal
Chemistry, 2005). Both free and heme associated protein have pro-
"oxidant properties due to their reaction with H202 to form ferry1 species,
which can easily initiate lipid peroxidation (Current Medicinal Chemistry,
2005).
c) Low molecular weight antioxidants: These are subdivided into
lipid-soluble antioxidants (Tocopherol, carotenoids, quinines, bilirubin
and some poly-phenols) and water-soluble antioxidant (ascorbic acid,
uric acid and some polyphenols) (Niki, 1987).
a-Tocopherol (vitamin E) and p-carotene have considerable
support as lipid-soluble antioxidants; tocopherol might act synergistically
with ascorbate. Vitamin C in living organisms regenerates vitamin E by
reducing the tocopherol radical that is produced when vitamin E
scavenges a peroxyl radical (Niki, 1987). Uric acid is another antioxidant
in primates as their blood has a higher concentration than that of other
mammals; uric acid might serve to scavenge reactive free radicals (Re)
and therefore account for the prolonged life span of humans (Ames et
al., 1981). Some carotenoids, including p-carotene, quench highly

reactive singlet oxygen under certain conditions and can block free
radical-mediated reactions (Bendich, 1989).
1 .I .2 Mechanism of action of antioxidants
Two principle mechanisms of action have been proposed for
antioxidant. The first is chain-breaking mechanism, by which primary
antioxidants donate an electron to the free radical present in the system.
The second mechanism involves removal of ROSIRNS initiators
(secondary antioxidants) by eliminating chain-initiating catalyst (Murray,
Electron donation
Primary antioxidants are compounds that are able to donate
hydrogen atom rapidly to a lipid radical forming a new radical, more
stable than the initial one (Murray et a/., 1990). Biological organs
contain many polyunsaturated fatty acids (PUFA), such as linoleic,
linolenic and arachidonic acid, mainly in the form of ester with
cholesterol. These PUFA can undergo lipid peroxidation that can be
interrupted by the primary antioxidant by the donation of electrons.
The whole process can be depicted as follows.
RH + O2 (Singlet oxygen) - - - - -ROOH
ROOH + ~ e ' ~ - - - - - RO. + HO- + ~ e ' ~
ROOH + ~ e ' ~ - - - - ROO. + H' + ~ e ' ~
ROO.+a-TO. - - - - Non radical products.
RH = Polyunsaturated fatty acid (PUFA)

ROOH = PUFA hydroperoxide
RO. = Alkoxyl radical
ROO. = Peroxyl radical
a - TO. = Tocopheryl radical
Metal Chelation
Secondary antioxidant can retard the effect of ROSIRNS radical
initiated action by means of initiator removal or elimination. This can be
accomplished by deactivation of high-energy species, absorption of UV
light, scavenging of oxygen and thus reducing its concentration (Omenn
et a/, 1996). Chelation of metal catalyzes free radical reaction or inhibits
peroxidase, such as NADPH oxidase, xanthine oxidase, dopamine-p-
hydroxylase or lipoxygenases (Omenn et al., 1996).
The ability of antioxidant to chelate transition metal ions can be
followed spectroscopically. High molecular weight proteins bind directly
or indirectly to redox active metals and thus inhibit the production of
metal-catalyzed free radicals. Some low molecular weight compounds,
such as polyphenols, in addition to their ability to donate hydrogen atom
and thus act as chain-breaking antioxidant, can also chelate transition
metal ions and hence inhibit free radical formation (Omenn et a/., 1996).
1.2 Lipid peroxidation
Peroxidation (autooxidation) of lipids exposed to oxygen is
responsible not only for deterioration of food (rancidity) but also for
damage to tissues in vivo, where it may be a cause of cancer,

inflammatory diseases, atherosclerosis, aging, etc (Murray et al., 1990).
The deleterious effects are initiated by free radicals (ROO., RO., OH.)
produced during peroxide formation from fatty acids containing
methylene interrupted double bonds, i.e., those found in the naturally
occurring polyunsaturated fatty acid (PUFA) (Murray et a/., 1990).
Lipid peroxidation is a chain reaction providing a continuous
supply of free radicals that initiate further peroxidation. Since the
molecular precursor for the initiation process is generally the
hydroperoxide product (ROOH), lipid peroxidation is a branching
reaction with potentially devastating effects. To control and reduce lipid
peroxidaiton both humans and nature involves the use of antioxidants.
1.3 Mechanisms of lipid peroxidation induction
Three different mechanisms are able to induce lipid peroxidation.
1 .3.l Autoxidation
This is a radical - chain process involving three sequences:
(a) Initiation: In a peroxide-free lipid system, the initiation of a
peroxidation sequence refers to the attack of a l iOS (with sufficient
reactivity) able to abstract a hydrogen atom from a methylene group
(-CH2-); these hydrogen having very high mobility (Morel, 1997). This
attack generates easily free radical from polyunsaturated fatty acids: OH
is the most efficient ROS to do that attack, whereas 02. is sufficiently
reactive.

The carbon radical tends to be stabilized by a molecular
rearrangement to form a conjugated diene. Under aerobic conditions
conjugated dienes are able to combine with O2 to give a peroxyl (or
peroxyl) radical, ROO'.
(b) Propagation: A peroxyl radical is able to abstract hydrogen from
another lipid molecule (adjacent fatty acid), especially in the presence of
metals such as copper or iron, thus causing an autocalalytic chain
reaction. The peroxyl radical combines with hydrogen to give a lipid
hydroperoxide. This reaction characterizes the propagation stage
(Morel, 1997).
(c) Termination: Termination (formation of a hydroperoxide is most
often achieved by reaction of a peroxyl radical with a-tocopherol, which
is the main lipophilic "chain-breaking molecule" in the cell membranes.
Furthermore, any kind of alkyl radical (lipid free radical) can react with a
lipid peroxide to give non-initiating and non-propagating species such as
the relatively stable dimers (Morel, 1997).
1.3.2 Photo-Oxidation
As singlet oxygen ('02.) is highly electrophilic, it can react rapidly
with unsaturated lipid but by a different mechanism than free radical
autoxidation. In the presence of sensitizers (chlorophyll, porphyrins,
myoglobin, riboflavin, methylene blue, etc.) a double bond interacts with
singlet oxygen produced from O2 by light. Oxygen is added at either

ends of carbon of a double bond, which takes the trans configuration.
Thus, one possible reaction of singlet 0 2 with a double bond between
C12 and Cl3 of one fatty acid is to produce 12- and 13-hydroperoxide.
The inhibition of photosensitized oxidation is efficiently inhibited by
an antioxidant present and rich in green leafy carotenoids, vegetables
and many coloured fruits (Khachik et a/., 1986). The inhibitory
mechanism is thought to be through an interference with the formation of
' singlet oxygen from the oxygen molecule. In contrast tocopherols
inhibits its oxidation by quenching the previously formed singlet oxygen,
and this forms stable additional products (Morel, 1997).
1.3.3 Enzymatic Peroxidation
Lipoxygenase enzymes (from plants and animals) catalyze
reaction between O2 and polyunsaturated fatty acid, such as arachidonic
acid (20 : 4, - 6), containing methylene interrupted double bonds. When
20: 4,-6 is the substrate, these hydroperoxides are known as HpETES,
which can be transformed into hydroxyl products (HETEs). These
HETEs are also formed directly via cytochrome P450 induced reactions.
Cycloxygenase enzymes (in plants and animals) catalyzed the
addition of molecular oxygen to various polyunsaturated fatty acid, they
are thus converted into biologically active molecule called endoperoxide
(PGG, PGH), intermediates in the transformation of fatty acid to
prostaglandin (Morel, 1997).

1.4 General antioxidant actions
1.4.1 Free radicals
Every cell has chemical reactions involving the oxidation and
reduction of molecules. These reaction or redox pathways can lead to
the production of free radicals.
A free radical is any chemical species capable of independent
existence possessing one or more unpaired electrons. Biological free
radicals are thus highly unstable molecules that have electrons available
to react with various organic substrates (Sahart, 2001).
Many free radicals are generated from naturally occurring
processes such as oxygen metabolism and inflammatory processes.
For example, when cells use oxygen to generate energy, free radicals
are created as a consequence of ATP production by the mitochondria
(Sahart, 2001). Exercise can increase the levels of free radicals as can
environmental stimuli such as ionizing radiation (from industry, sun
exposure, cosmic rays, and medical x-rays), environmental toxins,
altered atmospheric conditions (e.g. hypoxia and hyperoxia), ozone and
nitrous oxide (primarily from automobile exhaust). Lifestyle stressors
such as cigarette smoking and excessive alcohol consumption are also
known to affect levels of free radicals (Omenn et a/., 1996).
It has been noted that certain organ systems are predisposed to
greater levels of oxidative or nitrosative stress. Those organ systems
most susceptible to damage are the pulmonary system (exposed to high

levels of oxygen), the brain (exhibits intense metabolic activity yet has
lower levels of endogenous antioxidants), the eye (constantly exposed to
damaging UV light), circulatory system (victim to fluctuating oxygen and
nitric oxide levels) and reproductive systems (at risk from the intense
metabolic activity of sperm cells). Nearly every organ system can be
found to have an Oxidative or Nitrosative "Achilles heel" (Omenn, et a/.,
1996).
Reactive Oxygen Species (ROS)
ROS is a term collectively describing radicals and other non-
radical reactive oxygen derivatives. These intermediates may participate
in reactions giving rise to free radicals or that are damaging to organic
substrates. ROS in living organisms include the following:
Radicals Non-Radicals
Hydroxyl OH* Peroxynitrite ONOO-
Superoxide 02*- Hypochloric acid HOCl
Nitric oxide NO* Hydrogen peroxide H202
Thyl RS* Singlet oxygen A&-' 02
Peroxyl ROzO Ozxone 0 3
Lipid peroxyl LOO* Lipid peroxide LOOH
Reactive Nitrogen Species (RNS)
RNS are nitrogen-based molecules that can act to facilitate
nitrosylation reactions. Reactive nitrogen species (RNS) include:

Nitrous oxide NO*
Peroxynitrite OONO-
Peroxynitrous acid ONOOH
Nitroxyl anion NO'
Nitryl chloride N02CI
Nitrosyl cation NO'
Nitrogen dioxide NO2*
Dinitrogen trioxide N203
Nitrous acid HN02
The most reactive and damaging free radicals are the OH* and OONO-
(Sahart, 2001). Many other radical species can be formed by biological
reactions, for example: phenolic and other aromatic species are often
formed during xenobiotic metabolisms as part of natural detoxification
mechanisms (Sahart, 2001). Most of the free radicals are produced by
mitochondria and most of the free radical damage is to mitochondria
membranes and mitochondria1 DNA (Wei and Lee, 2002). Between one
and five percent of the oxygen used by mitochondria to generate energy
results in the formation of superoxide radicals (Wei and Lee, 2002).
Although mitochondria are the major source of free radicals, there
are numerous other sources. A green peroxidase of phagocytic cells (as
neutrophils and monocytes) are another source of free radicals. They
(neutrophils and monocytes) assist in bacteriocidal activity by catalyzing

the oxidation of ionic halogen to free halogen (Buetner and Jurkiewicz,
1996).
Myeloperoxidase enzyme, which is a peroxidase found in the
lysosomal granules of myeloid cells, particularly macrophages and
neutrophils, responsible for generating potent bacteriocidal activity by the
hydrolysis of hydrogen peroxide (produced in the metabolic burst) in the
presence of halide ions (Buettner and Jurkiewicz, 1996). Free radicals
are generated by eicosanoids from arachidonic acid during Ischemia-
reperfusion injuries. During reperfusion the endothelial enzyme xanthine
oxidase converts oxygen to superoxide, which can react with nitric oxide
to produce peroxynitrite (Buettner and Jurkiewicz, 1996). Free radicals
from tobacco smoke and air pollution can cause oxidative damage to
* lungs, blood vessels and other body tissue (Bendich and Olson, 1989).
Reactive free radicals ( R - ) ~ appear to have a role in the general
process of aging and in tissue damage that results from radiation,
reactive oxygen metabolite and carcinogen metabolism (Rose and Bode,
1993):
Details of underlying chemistry of ascorbate (Levine and Morita,
1985) and free radical generation (Grisham and McCord, 1986) are
available. Although many or most R' that are generated in the body are
metabolized to non-reactive species (Fig. 2), cellular damage is initiated
under some conditions.

Animals have evolved intricate and interrelated processes for
protecting against the effects of R'. The enzymatic reactions of
superoxide dimutase (SOD), catalase, glutathione peroxidase are not
100% effective in eliminating the formation of all free radicals. For
example, the very reactive hydroxyl free radical, HO', is not eliminated
by these mechanisms (Rose and Bode, 1993).
At the body's non-enzymatic protective mechanisms is a
scavenging reaction in which some endogenous compounds with the
inherent trait of entering into redox reactions contributes an electron to
fill the outer shell of R' and thereby neutralize it to a non-reactive
species. In principle, many chemicals could serve this purpose because
the high reactivity of R* results in it extracting an electron from almost
any available molecule. A few of the compounds shown to have this
property are: Mannitol (Caughey and Watkins (1 985) heamoglobin
(Giulivi and Davies, 1990), estrogens (Niki and Nakon, 1990), bile acids
and derivatives (Stocker et a/., 1990) and serotonin (Jovanovic et a/.,
1990).
For a substance to function biologically, it must do more than
simply react with R*. The present emphasis is on water-soluble
compounds that might have been useful throughout the long
evolutionary development from microbes to mammals. Particular
emphasis is on primates, as they are subjected to threat from R' over
long lifespan.

It must be considered that the source of R* changed over the last
10 years, with the threat from reactive oxygen species increasing (due to
plant generation of 02) and the threat from solar radiation diminishing
(due to emergence of the earth's stratospheric ozone layer) (Rose and
Bode, 1993). Carcinogen metabolites tend to be electron-deficient or
electrophilic (Cavalieri and Rogan, 1984).
Properties of an ideal free radical scavenger
Protection is thought to be available in the form of endogenous
compounds that react with and thereby "scavenge" the R*. Because
many R* are reactive forms of oxygen, an effective scavenger is often
referred to as an antioxidant.
To be an effective antioxidant physiologically, a substance must
have certain chemical and biological properties.
(a) It must be present in adequate amounts in the body
In that most R* have a brief half-life in the body and diffuse only
over short distances, the probability that they react with any given
antioxidant is proportional to the antioxidant's concentration in the
immediate environment where the R* is generated. Most potential
scavengers are present in the mammalian body at a low concentration.
Some have appeared only recently in evolution.
For instance, the introduction of hemoglobin coincided with the
appearance of animals having a closed circulatory system; most earlier
forms of aerobic life depended on cutaneous respiration and did not

have the possibility of respiratory pigments protecting them from free
radical threats (Rose and Bode, 1993).
(b) It must be versatile
The ideal scavenger should combine with a wide variety of free
radicals, i.e., it must be readily oxidized. One limitation of superoxide
dismutase (SOD) in eliminating free radicals is its lack of versatility; it
has but one substrate (Rose and Bode, 1993).
(c) It must be suitable to be compartmentalized
The antioxidant must be suitable for the body to translocate it
between tissues and must accumulate within compartments where a
need for protection exists at the time. A frequent cellular mechanism for
directing substrates to specific sites of the body is membrane transport
, e.g. through polarized cells of the gastrointestinal tract, renal tubule,
liver, placenta etc. (Rose and Bode, 1993). Molecualr size of the
antioxidant is important. Small molecules may be so permanent that
even if they were recognized by a transport mechanism, they would
. readily diffuse out of any membrane-bound compartment. Very large
compounds may not be transported across cellular membranes at rates
great enough to be useful (Rose and Bode, 1993).
(d) It must have tolerable toxicity
Ideally, the antioxidant would be non-toxic, both before and after it
performs the scavenging reaction. If toxicity is a possibility, careful

management of the toxic form must be accomplished under normal
conditions (Rose and Bode, 1993).
(e) It must be available
If the compound is to be accessible to all animal species, it should
either be synthesized de novo or acquired in the diet. A particular
antioxidant might be produced by some animal species or acquired in
the diet by others. If some organisms became devoid of synthetic
capability (e.g. primates, in the case of ascorbic acid) the compound
must be suitable to be ingested as food. Therefore, it must exist in plant
products and be stable for periods of days or weeks after harvest. It
must also be suitable for the normal processes of ingestion, digestion,
and intestinal absorption.
- (f) It might be suitable for regeneration
The process of neutralizing a R' results in the scavenger
becoming oxidized to a form that has less capacity to react with
additional Re. Thus a scavenger would be particularly useful if it is
' recycled so that dietary acquisition does not become prohibitively
expensive. The compound must have a biologically convenient reducing
mechanism, which could be either a specific enzyme or a direct chemical
reaction.
(g) It must be conserved by the kidneys
If the compound is filtered in the glomerular of the kidney, it must
be suitable for reabsorption. Because renal clearance of small

compounds that are filtered but not reabsorbed is high in most animals
(with the half-life of plasma disappearance < 1 hr), large urinary losses
would occur in the absence of active reabsorption (Rose and Bode,
1993).
1.4.2 Antioxidant molecules
We can accept that many substances interact with free radicals or
at least with the most reactive of them. This is not surprising, as some
R' (such as HOD) are so electrophylic that they strip an electron or
hydrogen atom from almost any compound with which they come in
contact. Some of these reactions immediately result in products that are
stable, thus terminating the free radical activity. Many of those
compounds, however, have few of the properties listed above. For
instance, mannitol is present in plants but it is not synthesized in
animals. It is not recognized by mammalian membrane receptors or
transporters; thus it is not absorbed in the gastrointestinal tract or
directed to specific sites of the body. Mannitol does not enter most
animal cells, as evidenced by its use in research as an extracellular
space marker (Rose and Bode, 1993).
a-Tocopherol (Vitamin E) and p-carotene have considerable
support as lipid-soluble antioxidants; tocopherol might act synergistically
with ascorbate. Vitamin C in living organisms regenerates vitamin E by
reducing the tocopherol radical that is produced when vitamin E
scavenges a peroxyl radical (Niki, 1987).

ASCH- + 'TOC TOC + 'ASC-
ASCH- -ASC-
Fig. 2: Ascorbate free Radical Formation
R H
This interaction is consistent with the results of a 1 3 ~ - ~ ~ ~ study,
which showed that the phenolic head group of a-tocopherol in
unilamellar vesicles is located very close to the lipid-water interface
(Perly et a/. , 1985).
Urate is another likely candidate for an antioxidant role in
primates, as their blood has a higher concentration than that of other
mammals. Urate might serve to scavenge R' and thereby account for
the prolonged life span of humans (Ames eta/., 1981).
Free radicals can be listed by one-electron reduction potentials in
millivolts (mV) at pH 7.0. The reduced form of each radical is capable of
neutralizing (reducing) free radicals having a higher potential. As can be
seen from the Table 1, the hydroxyl radical (,OH) has the highest
potential and is the most destructive (reactive) of biological free radicals.

20
Table I: Radical Reaction Potentials
Radical
'OH (hydroxyl)
'LO (alkoxyl)
LOO'
'GS (glutathione)
'HU (Urate)
'TOC (Tocopherol)
'ASC (Ascorbate)
I ~ e ~ ' - EDTA
Vitamin C (Ascorbate ASCH-)
Vitamin C can donate a hydrogen atom to a free radical molecule
(R') thereby neutralizing the free radical while becoming an ascrobate
radical itself. It accumulates in many tissues, both in animal species that
produce it and in those that absorb it as a vitamin. Considering the levels
of ascorbate in humans compared with the plasma, it is highly
concentrated in leukocytes, adrenal, pituitary and compartments of the
eye (Evans et a/, 1982).
Table 1 shows that the ascorbate radicallascorbate
thermodynamic couple is low compared with the reduction potential of
the a-tocopherol free radical, the glutathione radical, the aliphatic alkoxyl
and alkyl peroxyl radicals, and the hydroxyl free radical. Because of this,
ascorbate will act as an antioxidant in each system, and also with

superoxide (Nishikimi, 1975), the urate free radical and other radicals not
prevalent in the body such as nitroxides (Melhorn, 1991). The biological
damaging reactive oxidative species come from a variety of sources,
including ionizing radiation, oxygen metabolism and carcinogen
metabolism. The ease with which ascorbate is oxidized has resulted in
significant commercial utility; ascorbate, or its stereoisomers form, D-
Isoascorbate, is effective in preventing (or reversing) oxidation in a wide
variety of food products. The evidence is also strong that ascorbate has
high reactivity with R' in body fluids (Buettner and Jurkiewicz, 1993).
Ascorbate's efficacy as a scavenger depends on the reactivity of
the ascorbyl free radical (AFR). If AFR were highly reactive with other
substances at the biological pH, temperature, electrolyte composition,
, etc. the chain of free radical reactions would be propagated to
completion as with other intermediate forms of R* in the cell.
In addition to being well suited for an antioxidant role in biology,
ascorbate has also been shown to have a pro-oxidant role in vitro (Borg
and Schaich, 1989).
Chelation of ~ e ~ ' with EDTA actually enhances the reactivity of
iron toward superoxide, thus favouring the Haber-Weiss Reaction
(Buettner and Jurkiewicz, 1996). F~~'-EDTA chelate can catalyze the
Fenton Reaction to generate hydroxyl ion without reduction of Fe3' to
~ e * ' on the other hand, ~ e ~ ' and F~~'-EDTA can be reduced to Fe2' by
ascorbate (ASCH-) to generate the ascorbate radical ('ASC-). The

reduced Iron can then generate a hydroxyl radical by the Fenton
Reaction. Copper ion (cu*') is 80 times more efficient at reacting with
ascorbate than ~ e ~ ' (Buettner and Jurkiewicz, 1996). Thus vitamin C
can be a powerful antioxidant as long as metal ions are not present, but
small amounts of vitamin C in the presence of metal ions can make
vitamin C a dangerous pro-oxidant. Large amounts of vitamin C can
restore the antioxidant function. (Vitamin C has been called an
'. "oxymoron antioxidant") (Buettner and Jurkiewicz, 1996).
Ascorbate is present in many plants, microorganisms, and
animals; it therefore appears to have been present throughout animal
evolution. Evidence that early forms of life use ascorbate comes from
the finding of ascorbate oxidase isoenzymes in tea leaves (Chen and
Asada, 1989).
There are three (3) principal reasons for suggesting that
ascorbate serves an important role as scavenger of free radicals in the
human body:
(a) It is chemically suited to react with oxidizing free radicals;
(b) It is present in the body at sufficiently high concentrations to
be effective;
(c) It fits into the physiology of cellular transport and metabolism.
This combination of properties is well suited for this antioxidant
molecule to contribute to the extended life span potential of
humans (Cutler, 1984).

The most effective singlet oxygen quenchers are carotenoids,
phytochemicals, which plants produce to protect themselves from singlet
oxygen produced by ultraviolet light (Cutler, 1984). rruur" rc-*-
Carotenoids F L * . ~ .b -. ' -
Of 600 carotenoids from natural sources that have been
characterized, fewer than 10% serve as precursors of vitamin A. Many
dietary carotenoids, both with and without provitamin A activity, are
formed in the blood and tissues of humans (Bendich and Olson, 1989).
p-carotene, the most nutritionally active carotenoid, comprises
15-30% of total serum carotenoids (Bendich and Olson, 1989).
Green leafy vegetables and many coloured fruits are rich in
carotenoids and polyenes, (Khachik et a/., 1986). In animal models,
carotenoids have been implicated as chemo-protective or chemo-
preventive agents in several kinds of cancer (Peto et a/., 1981),
particularly skin cancer. Epstein (1977) first showed that injected P-
carotene slowed the growth of skin tumors in hairless mice exposed to
ultraviolet light (UV-A, UV-B). Similarly, feeding either p-carotene,
canthaxanthin, or phytoene to hairless mice exposed to UV-B irradiation
delayed the appearance of skin tumors and reduced their number
(Mathews-Roth, 1982).
Carotenoids may protect cells from oxidative stress by quenching
free radicals capable of causing cellular damage. Unsaturated lipids in
cell membranes are prime targets for free radical reactions. A free

radical-mediated attack on lipid membranes can initiate a chain reaction
that results first in lipid peroxidation and ultimately in functionally
significant damage to membranes, enzyrnes and nucleic acids
(Benedich and Olson, 1989).
Both in vivo and in vitro, p-carotene has been shown to protect
isolated lipid membranes from peroxidation, LDL-containing lipids from
oxidation, and liver lipids from oxidation induced by carbon tetrachloride-
induced free radicals.
In chemical studies, the possible basis for the protective actions of
carotenoids have been examined. Although p-carotene primarily has
been studied, theoretically all carotenoids with a similar conjugated
double bond system should act similarly (Krinsky and Deneke, 1982). In
purely chemical studies, p-carotene interacts with peroxyl radicals
irreversibly to form a carbon-centered carotenoid radical (Burton, 1989).
It is difficult to extrapolate directly from chemical and biological
systems. For example, although antioxidant effects in a chemical
system were noted at a carotenoid concentration of 50 pM, maximal
inhibition of peroxidation was observed at 0.5 mM (Burton and Ingold,
1984). On the other hand, as singlet oxygen quenchers, low
concentrations of carotenoids with nine or more conjugated double
bonds can inhibit the peroxidation of linolenate (Burton, 1989).
At high oxygen tensions, a-tocopherol is the most effective
antioxidant (Burton, 1989).

More than 20 epidemiological studies, both prospective and
retrospective types, have shown that the risk of developing or dying from
certain types of cancer (usually in both men and women) is inversely
associated with the intake of carotenoid-containing fruits and vegetables
and with higher levels of serum p-carotene concentration (Ziegler, 1989).
In the case of lung cancer, which has been most closely associated with
intake of fruits and vegetables, individuals with the lowest carotenoid
intake or serum p-carotene concentrations were at a two- to sevenfold
higher risk of developing neoplasms than those in the highest intake and
highest serum level groups (Ziegler, 1989).
1.5 Oxidative stress
Oxidative stress is a medical term for damage to animal or plant
cells (and thereby the organs and tissues composed of those cells) that
occurs in normal metabolic processes through the production of "free
radicals" caused by reactive oxygen species (Current Medicinal
Chemistry, 2005). In the quest to find a "mate" and become stable, free
radicals interact with the nearest molecule, targeting proteins, fats, or
even DNA. These actions can be so violent that they create bursts of
light within the body. If not neutralized rapidly, they may crate more free
radicals or cause damage to vessel and cell walls, lipids, proteins and
even the nucleus (DNA) of the cell, processes which can lead to cell
death (apoptosis) by induction of mitochondria1 membrane permeability

transition and release of apotogenic factors such as cytochrome C (Wei
and Lee, 2002).
Oxidative stress could be defined as an imbalance between pro-
oxidants prevailing (Current Medicinal Chemistry, 2005). Superoxide is
produced deleteriously by I-electron transfers in the mitochondria1
electron transfer chain. Other enzymes capable of producing superoxide
are xanthine oxidase, NADPH oxidases and cytochrome P450(s).
Hydrogen peroxide is produced by a wide variety of enzymes including
monoxygenases and oxidases (Current Medicinal Chemistry, 2005).
Meals such as Iron, Copper, Chromium, Vanadium and Cobalt are
capable of redox cycling in which a single electron may be accepted or
donated by the metal. This action catalyzes reactions that produce
reactive radicals and can produce reactive oxygen species such as
hydroxyl radical in reactions like Fenton's reaction. The hydroxyl radical
then can lead to modifications of amino acids (e.g. meta-tyrosine and
ortho-tyrosine formation from phenyl-alanine) carbohydrates, initiate lipid
peroxidation (Current Medicinal Chemistry, 2002).
Most enzymes that produce reactive oxygen species contain one
of these metals. The presence of such metals in biological systems in
an unsequestered form (not in an enzyme or other protein) can
significantly increase the level of oxidative stress (Current Medicinal
Chemistry, 2002). Under normal circumstances, there is a delicate
balance between the production of oxidants and antioxidants. However,

during periods of increase pro-oxidant production, antioxidants can
become depleted in the process of detoxification, leaving the body with
little or no defense against the next attack of free radicals (Ray, 2006).
Oxidative stress is a combination of hereditary, environment and
lifestyle factors. While it is possible to change lifestyle and some
environmental factors through diet, physical activity and antioxidant
supplementation, we cannot change our genes (Ray, 2006).
Intense exercise can actually increase oxidative stress in humans,
as the body struggles to detoxify free-radicals while muscles use oxygen
at 100 - 200 times their normal rate (Stauth ef a/., 2002). In this type of
intense exercise for example exercise by ultramarathon runners, who
have competed in races of 50 kilometers, or more than 30 miles, it is
found that here is increased oxidative stress and depleted levels of
vitamin E. When running or doing other intense exercise, there is a 10-
20 fold increase in whole body oxygen consumption; this can produce
reactive oxygen species, which can be harmful, at rates that exceed the
body's natural capacity to detoxify them (Stauth et a/, 2002).
a-ketoglutarate dehydrogenase (a-KGDH), a key enzyme in the
Kreb's cycle is a crucial early target of oxidative stress (Tretter and
Adam-Vizi, 2000). The study on the generation of ROS in the reaction
catalyzed by a-ketoglutarate dehydrogluase demonstrates that a-KGDH
is able to generate H202 and thus, could also be a source of ROS in
mitochondria. Isolated a-KGDH with coenzyme A (HS-CoA) and

thiamine pyrophosphate started to produce H202 after addition of a-
ketoglutarate in the absence of nicotinamide adenine dinucleotide-
oxidized (NAD'). NAD', which proved to be a powerful inhibitor of a-
KGDH-mediated H202 formation, switched the H202 form mode of the
enzyme to the catalytic [nicotinamide adenine dinucleotide-reduced
(NADH) forming] mode (Treter and Adam-Vizi, 2000). In contrast, NADH
stimulated H202 formation by a-KGDH, and for this, neither a-
ketoglutarate nor HS-CoA were required. When all of the substrates and
cofactors of the enzyme were present, the NADHINAD' ratio determined
the rate of H202 production. The higher the NADHINAD' ratio the higher
the rate of H202 production. H202 production as well as the catalytic
function of the enzyme was activated by ca2'. In synaptosomes, using
a-keto-glutarate as respiratory substrate, the rate of H202 production
increased by 2.5-fold, and a conitase actikity decreased, indicating that
a-KGDH can generate H202 in in siu mitochondria. Thus a-KGDH is not
only a target of ROS but could significantly contribute to generation of
oxidative stress in the mitochondria (Tretter and Adam-Vizi, 2000).
1.6 Diseases associated with free radicals
Certain organ systems are predisposed to greater levels of
oxidative or nitrosative stress. Those organ systems most susceptible to
damage are the pulmonary system (exposed to high levels of oxygen),
the brain (exhibits intense metabolic activity yet has lower levels of

endogenous antioxidants), the eye (constantly exposed to damaging UV
light), circulatory system (victim to fluctuating oxygen and nitric oxide
levels) and reproductive systems (at risk from the intense metabolic
activity of sperm cells) (Wee and Lee, 2002).
Oxidative stress definitely contributes to tissue injury following
irradiation and hyperoxia and is suspected (though not proven) to be a
cause of neuro degenerative diseases including Lou Gehrigls disease
(aka MND or ALS), Parkinson's disease, Alzheimer's disease and
Huntington's disease. Oxidative stress is thought to be linked to certain
cardiovascular disease, since oxidation of LDL in the endothelium is a
precursor to plague formation. Oxidative stress also contributes to many
diseases including inflammation, cancer, autoimmune diseases (Current
* Medicinal Chemistry, 2005).
These diseases associated with free radicals are: -
1.6.1 Neurodegenerative diseases
a) Parkinson's disease
Parkinson's disease is a degenerative disorder of the central
nervous system that affects the control of muscles, and so may affect
movement, speech and posture. Parkinson's disease belongs to a
group of conditions called movement disorders, characterized by muscle
rigidity, tremor, a slowing of physical movement (bradykinesia), and in
extreme cases, a loss of physical movement (akinesia) (Parkinson,
2002).

Current concepts of the pathogensis of Parkinson's disease (PD)
center on the formation of reactive oxygen species and the onset of
oxidative stress leading to oxidative damage to substantia nigra pars
compacta (Jenner and Olanow, 1996). Extensive postmortem studies
have provided evidence to support the involvement of oxidative stress in
the pathogenesis of Parkinson's disease; in particular, these include
alterations in brain iron content, impaired mitochondria1 function,
alteration in the antioxidant protective systems (most notably superoxide
dismutase [SODJ and reduced glutathione [GSH]), and evidence of
oxidative damage to lipids, proteins and DNA (Jenner and Olanow,
1996).
Iron can induce oxidative stress, and intranigral injections have
been shown to induce a model of progressive Parkinsonism (Klein and
Ackerman, 2003). A loss of GSH is associated with incidental lewy body
disease and may represent the earliest biochemical marker of nigral cell
loss. GSH depletion alone may not result in damage to nigral neurons
but may increase susceptibility to subsequent toxic or free radical
exposure (Jenner and Olanow, 1996). The nature of the free radical
species responsible for cell death in Parkinson's disease remains
unknown, but there is evidence of involvement of hydroxyl radical (OH.),
peroxynitrite, and nitric oxide. Indeed, OH. and peroxynitrite formation
may be critically dependent on nitric oxide formation (Jenner and
Olanow, 1996).

Central to many of the processes involved in oxidative stress and
oxidative damage in Parkinson's disease are the actions of monoamine
oxidase-B (MAO-B). MAO-B is essential for the activation of l-methyl-4-
phenyl-I ,2,3,6-tetrahydropyridine to 1-methyl-4-phenylpyridinium ion, for
a component of the enzymatic conversion of dopamine to hydrogen
peroxide (H202), and for activation of other potential toxins such as
isoquinolines and beta-carbolines (Nakamura et a/., 2000).
"b) Huntington's Disease (HD)
Huntington's disease (HD), also known as Huntington disease and
previously as Huntington's chorea and chorea major, is a rare inherited
neurological disorder affecting up to 8 people per 100,000 (Vessie,
1932).
Huntington's disease is caused by a rinucleotide repeat expansion
in the Huntingtin (tt) gene and is one of several polyglutamine (or polyQ)
diseases (Vessie, 1932). This produces and extended form of protein,
mutant Huntingtin (mHtt), which causes cell death in selective areas of
the brain. HD's most obvious symptoms are abnormal body movements
called chorea, but also affects a number of mental abilities (Graham et
a/. , 2006).
The gene's coding involved in HD, called the HD gene, is used to
produce a 348 KDa cytoplasmic protein called Huntingtin (Htt). Htt has a
characteristic sequence of fewer than 40 glutamine amino acid residues

in the normal form, more than this and a mutated form of Htt that causes
the disease, mHtt, is produced (Ridley et a/., 1988).
The severity of the disease is generally proportional to the number
of extra residues. The continuous aggregation of the mHt molecules in
neuronal cells causes them to die off in selected regions of the brain.
Although the full function of Htt is unknown, it acts as a transcription
factor in upregulating the expression of Brain-derived neurotrophic factor
(BDNF). With mHtt, there is suppression of this transcription regulatory
function of Htt and hence underexpression of BDNF (Ridley et a/., 1998).
It is suspected that the cross-linking of Htt results in aggregates, which
are toxic, and can lead to dysfunction of the proteasome system. This
mitochondria1 dysfunction can lead to excitotoxicity and oxidative stress
(Graham et a/., 2006). The neurodegeneration caused by mHtt 8is
related to the caspase-6 enzyme clearing the Htt protein (Graham et a/.,
2006).
c) Autism
Autism is a complex neurological disorder and oxidative
imbalance is one feature of the autistic syndrome (Pratico, 2006).
Autism, an early onset neurological disorder, is characterized by
impaired social interactions, limited verbal and non-verbal
communication and repetitive and restricted behavioural patterns
(Pratico, 2006). Children with autism shows signs of abnormal blood-
vessels function and damaging levels of oxidative stress compared to

healthy children. Previous studies on Autism have shown that autistic
patients have reduced cerebral blood flow, presumably due to
constricted blood vessels in the brain, versus healthy controls. Urinary
samples of autistic children who were similar in age and healthy controls
were provided by the Pfeiffer Treatment Center where patients were
diagnosed with autism disorder and evaluated. Patients were excluded
from analysis if they had ever received anti-oxidant treatments or
medicine with any known anti-oxidant effect; if they suffered from chronic
illness, such as depression, psychosis, or inflammatory disorders; andlor
if they were sick at the time of the sample collection.
Isoprostane, a biomarker for oxidative stress; thromboxane, an
index of platelets activation; and prostacyclin, a measure of blood vessel
activation in the samples were measured. It was observed that the rates
thromboxane and prosacyclin synthesis re both not significantly
increased in autism, but are closely related with the rate of oxidative
stress. Compared with controls, children with autism had significantly
higher urinary levels of isoprostane, thromboxane and prostacyclin
(Pratico, 2006).
In patients with autism, the levels of isoprostane (the chemical
byproduct of free radicals attacking fat cells) are nearly double the level
of oxidative stress than the levels in healthy controls (Pratico, 2006).
In autistic patients, there is a biochemical imbalance in the
patients' blood vessels, resulting in high levels of thromboxane (an

indicator of platelet activity) and prostacyclin (an indicator of constricting
endothelial cells). During normal function, thromboxane and prostacyclin
work together to maintain the integrity of vessels. In response to
different kinds of stress, platelets release thomboxane, which causes
vessels to contract. The endothelium responds to elevated levels of
thromboxane by releasing prostacyclin. This event counterbalances the
effect on vessels, inducing dilation of the vessels and, in turn, more
'blood flow (Pratico, 2006). Oxidative imbalance may also play a role in
this disease: autism is characterized by an impaired antioxidant defense
system, higher free-radical production, and improvement of behavioural
symptoms after taking anti-oxidants (Pratico, 2006).
1.6.2 Cardiovascular disease
Cardiovascular disease refers to the class of diseases that involve
the heart and/or blood vessels (arteries and veins). While the term
technically refers to any disease that affects the cardiovascular system,
it is usually used to refer to those related to artherosclerosis (arterial
disease) (Andraws et a/., 2005).
Cardiovascular disease usually occurs as a result of arterial
damage. In coronary heart disease, atherosclerotic plaques (inflamed
fatty deposits in the blood vessel wall) obstruct the coronary arteries
(blood vessels supplying the heart). Narrowing of arteries is called
arterial stenosis. When the blockages become severe enough, the
blood flow to the heart is restricted (cardiac ischemia), especially during

increased demand (i.e. during exertion or emotion). This disease occurs
when one of the plaques ruptures, forming thrombus (blood clot) that
acutely occludes the whole artery. The portion of the heart muscle
supplied by that artery dies; this is known as a myocardial infarction (or a
heart attack in lay parlance). This may result in the death of the patient if
the affected area is large enough. If the patient survives, congestive
heart failure may result. (Andraws et a/., 2005).
Similarly, inflammation and blood clots may obstruct the cerebral
arteries (those supplying the brain). As the disease progresses, an
artery may be transiently blocked, causing cerebral ischemia. These
results in a transient ischemic attack (TIA) (Leaf et a/., 2003) called a
mini-stroke in lay parlance. If the obstruction is severe, a
cerebrovascular accident or stroke may result, due to the death of brain
tissue supplied by the artery (Leaf et a/., 2003).
In peripheral artery disease, obstruction occurs in the arteries of
the arms or legs. This results initially in pain, during temporary
obstruction, and finally in tissue death and gangrene if not treated
(Andraws et a/., 2005). There are many specific illnesses that may occur
in association with these and other cardiovascular disease. In addition
to the ones mentioned above, these include hypertension (high blood
pressure), arterial aneurysms (arterial enlargement and weakening),
cardiomegaly (abnormal enlargement of the heart), tachycardia1
braqdycardialarrhythmia (fast/slowlirregular heart rates), cardiac arrest

(heart stoppage), cardiomyopathy (heart muscle weakness), heart valve
regurgitation (leakage), and heart valve stenosis (narrowing) (Andraws
et a/., 2005).
There are many risk factors, which predispose to various forms of
cardiovascular disease. These include the following: -
- Age
- Absence of key nutritional elements, such as polyphenol
antioxidants
- Diabetes mellitus
- Hypercholesterolemia (elevated cholesterol levels) and less than
excellent lipoprotein particle profile (cholesterol subtypes)
- Tobacco smoking
- Obesity, especially central or male-type obesity; apart from being
linked to diabetes, this form of obesity independently increases
cardiovascular risk, presumably by inducing an inflammatory and
procoagulant state.
- Genetic factors/family history of cardiovascular disease
- High blood pressure (Leaf et a/., 2003).
1.6.3 Aging
Human cells rely on ATP for growth, differentiation, and response
to physiological stimuli and environmental changes. It has been
established that mitochondria make ATP by the coupling of respiration-
generated proton gradient with the proton-driven phosphorylation of ADP

by F,, F1 ATPase (Wei and Lee, 2002). Mitochondria do not only
produce less ATP, but they also increase the production of reactive
oxygen species (ROS) as byproducts of aerobic metabolism in the aging
tissues of the human and animals (Wei and Leo, 2002). It is now
generally accepted that aging-associated respiratory function decline
can result in enhanced production of ROS in mitochondria. Moreover,
the activities of free radical-scavenging enzymes are altered in the aging
.. process. The concurrent age-related changes of these two systems
result in the elevati.on of oxidative stress in aging tissues. Within a
certain concentration range, ROS may induce stress response of the
cells by altering expression of respiratory genes to uphold the energy
metabolism to rescue the cell. However, beyond the threshold, ROS
. may cause a wide spectrum of oxidative damage to various cellular
components to result in cell death or elicit apoptosis by induction of
mitochondrial membrane permeability transition and release of
apoptogenic factors such as cytochrome C. (Wei and lee, 2002).
Moreover, oxidative damage and large-scale deletion and
duplication of mitochondrial DNA (MTDNA) have been found to increase
with age in various tissues of the human. Mitochondria act like a
biosensor of oxidative stress and they enable cell to undergo changes in
aging and age-related diseases. On the other hand, it has recently been
demonstrated that impairment in mitochondrial respiration and oxidative
phosphorylation elicits an increase in oxidative stress and causes a host

of mtDNA rearrangements and deletions (Wei and Lee, 2002). This
aging of the skin is an outward manifestation of "Oxidative stress" which
is occurring within every cell in our body (Ray, 2006).
1.6.4 Cancer
Cancer is caused in all or almost all instances by mutation of
cellular genes that control cell growth and cell mitosis. The mutated
genes are called oncogenes (Antoniades and Owen, 1982). Usually two
or more different oncogenes must occur in a cell before the cell will
become cancerous. Mutation can occur as a result of exposure to
certain chemical, physical, or biological factors (Hall, 1984). Some of
these are the following:
(1) It is well known that ionizing radiation such as x-rays, gamma
rays, and particle radiations from radioactive substances and
even ultraviolet light, can predispose to cancer (Antoniades and
Owen, 1982). Ions formed in tissue cells under the influence of
such radiation are highly reactive and can rupture DNA strands,
thus causing many mutations (Hall, 1984).
(2) Chemical substances of certain types also have a high propensity
for causing mutations (Hall, 1984). Historically, it was long ago
discovered that various aniline dye derivatives are very likely to
cause cancer so that workers in chemical plants producing such
substances, if unprotected, have a special predisposition to
cancer (Hall, 1984). Chemical substances that can cause

mutation are called carcinogens. The carcinogens that cause by
far the greatest number of deaths in our present day society are
those in cigarette smoke. These can cause about one quarter of
all cancer deaths (Ziegler, 1989).
(3) Physical irritants can also lead to cancer, such as continued
abrasion of the linings of the intestinal tract by some types of food
(Gey et a/., ,1987). The damage to the tissue, leads to rapid
mitotic replacement of the cells. The more rapid the mitosis, the
greater the chance for mutation (Lerman, 1984).
1.7 Pharmacological options
Parkinson's disease is a chronic disorder that requires broad-
based management including patient and family education, support
group services, general wellness maintenance, exercise and nutrition
(de Lau et a/., 2004). The most widely used form of treatment is L-dopa
in various forms. L-dopa is transformed into dopamine in the
dopaminergic neurons by L-aromatic amino acid decarboxylase (often
known by its former name dopa-decarboxylase) (Jenner and Olanow,
1996). However, only 1-5% of L-DOPA enters the dopaminergic
neurons (de Lau et a/., 2004).
Carbidopa and benserazide are dopa decarboxylase inhibitors.
They help to prevent the metabolism of L-dopa before it reaches the
dopaminergic neurons and are generally given as combination
preparations of carbidopa/levodopa (co-careldopa) (e.g. sinemet

(http;//www.sinemet.com/parcopa) and benserazide/levodopa (co-
beneldopa) (e.g. Madopar) (de Lau et al., 2004).
There are also controlled release versions of Sinemet and
Madopar that spread out the effect of the L-dopa. Duodopa is a
combination of Levodopa and Carbidopa, dispersed as a viscous gel.
Using a patient-operated portable pump, the drug is continuously
delivered via a tube directly into the upper small intestine, where it is
rapidly absorbed (Marras et al., 2005). Talcopone inhibits the COMT
enzyme, thereby prolonging the effects of L-dopa, and so has been used
to complement L-dopa. Stalevo another drug containing Levodopa,
Carbidopa and Entacopone Stalevo (http://www.stalevo.com/info./sp).
Mucuna prurious, is a natural source of therapeut6ic quantities of L-dopa
, (de Lau et al., 2004).
The Dopamine-agonists bromocriptine (parlodel), pergolide
(permax), pramipexole (Mirapex), ropinirole (Requip), cabergoline
(Cabazer), apomorphine (Apokyn), and Lisuride (Revanil), are
. moderately effective (Mona, 2000).
Dopamine agonists can be useful for patients experiencing on-off
fluctuations and dyskinesian as a result of high doses of L-dopa.
Apomorphine can be administered via subcutaneous injection using a
small pump, which is carried by the patient. A low dose is automatically
administered throughout the day, reducing the fluctuations of motor
symptoms by providing a steady dose of dopaminergic stimulation

(Michael, 2005). Apomorphine is also available in a more acute dose as
an autoinjector pen for emergency doses such as after a fall or first thing
in the morning (Marras eta/., 2005).
Selegiline (Eldepryl) and rasogiline (Azilect) reduce the symptoms
of Parkinson's disease by inhibiting monoamino oxidaze-B (MAO-B),
which inhibits the breakdown od dopamine secreted by the dopaminergic
neurons (Jenner and Olanow, 1996).
MAO-B is essential for the activation of l-methyl-4-phenyl-I ,2,3,6-
tetrahydropyridine to 1-methyl-4-phenylpyridinium ion, for a component
of the enzymatic conversion of dopamine to hydrogen peroxide (H202),
and for the activation of other potential toxins such as isoquinolines and
beta-carbolines (Jeriner and Olanow, 1996).
Thus the inhibition of MAO-6 by drugs such as selegiline may
protect against activation of some toxins and fee radicals formed from
the MAO-B oxidation of dopamine. In addition, selegiline may act
through a mechanism unrelated to MAO-6 to increase neurotrophic
factor activity and upregulate molecules such as glutathione, superoxide
dimutase (SOD), catalase, and BCL-2 protein, which protect against
oxidant stress and apostosis. Consequently, selegiline may be
advantageous in the long-term treatment of Parkinson's disease (Jenner
and Olanow, 1996).
There are standard treatments to alleviate emotional symptoms of
Huntington's disease. These include the use of antidepressants and

sedatives with antipsychotics (in low doses) for psychotic symptoms
(Graham et a/., 2006). Nutrition is an important part of treatment, most
Huntington's disease sufferers need two-three times the calories than
the average person to maintain body weight. Note, an average calorie
intake is between 2000 (women) to 2500 (children and men) (Graham et
a/., 2006).
To aid swallowing, thickener can be added to drinks when
* swallowing becomes hazardous the option of using a stomach PEG for
intake of nutrients is often chosen, this reduces the chances of
pneumonia due to aspiration of food and increases the amount of
nutrients that can be given (Sirna therapeutics, 2006). Other agents and
measures that have shown promise in initial experiments include
. dopamine receptor blockers, creatine, CoQ10, the antibiotic Minocycline,
Trehalose, exercise, antioxidant-containing foods and nutrients (Sirna
Therapeutics, 2006).
Treatment of cardiovascular disease depends on the specific form
of the disease in each patient, but effective treatment always includes
preventive lifestyle changes, which take the form of modifying risk
factors (Leaf, et a/., 2003). Some, such as gender (male or female), age,
and family history cannot be modified. Smoking cessation (or
abstinence) is one of the most effective and easily modifiable changes
(Andraws et a/., 2005). Regular cardiovascular exercise (aerobic
exercise) complements the healthful eating habits. Medications, such as

blood pressure reducing medications, aspirin and the statin cholesterol-
lowering drugs may be helpful (Leaf et a/., 2003).
Sometimes, the combination of diet and exercise will improve
lipoprotein (cholesterol) levels. Aspirin has been shown to decrease the
clot formation that may lead to myocardial infarctions and strokes; it is
routinely prescribed for patients with one or more cardiovascular risk
factors (Andraws et a/., 2005). Eating oily fish at least twice a week may
help reduce the risk of sudden death and arrhythmias (Leaf et a/., 2003).
Studies of individual heart cells showed that the fatty acids blocked
excessive sodium and calcium currents in the heart, which could
otherwise cause dangerous, unpredictable changes in its rhythm (Leaf et
a/., 2003).
Epidemiological evidence suggests that vitamin E
supplementation decreases the incidence of ALS (bou Gehrig's disease)
and Alzheimer's (Andraws, et a/., 2005).
In epidemiological studies, the intake of caroteroid-rich, fruits and
vegetables has been correlated with protection from some forms of
cancer, particularly lung cancer (Bendich and Olson, 1989). In the
treatment of cancer, some classes of chemotherapeutic agents are used
such agents like Alkylating agents (e.g. Nitrogen Mustards,
Ethylenimines and Methylmelamines, Folic acid analogs),
Antimetabolites, (e.g. pyrimidine analogs, purine analogs and related
inhibitors) Natural products, (e.g. Antibiotics, Enzymes, biological

response modifiers, Anthracenedione etc.) and hormones and
antagonists (e.g. Estrogens, Antiestrogen) androgens, antiandrogen,
Gonadotropin-Releasing hormone analog) (Chang et a/., 1993).
1.8 General review of phytochemistry
1.8.1 Alkaloids
They are a group of basic secondary plant substance, which
usually possesses an n-containing heterocyte. Alkaloids exist in plants
as salts, amine or n-oxides. Dicotyledonous plants are the real
producers of alkaloids (Evans, 1989).
They appear in large members and in many variation in these
plants. They are bitter to taste, so when present in plants, insects and
predators tend to move away from such plants. They also protect the
plant from the effect of singlet oxygen (Bonner and Varner, 1965).
Alkaloids at high concentration, produces a variety of toxic effects on
animals.
Their pharmaceutical and medicinal importance can be seen to
act on the cardiovascular system and some have been resorted to be
antihypertensive. Alkaloids also contribute to liver disease and
hepatocellular tumor (Antoniodes and Owen, 1982). Alkaloids of
Catharanthus roseus are used in cancer chemotherapy.
1.8.2 Flavonoids
The origin of the names is from a Latin word "FLAVUS" meaning
yellow. They are a series of related water soluble phenolic glycosides

having in common a basic structural unit. The CI5 skeleton of flavones.
The flavones are sap-soluble (Bonner and Varner, 1965). The phonetic
compound contributes to the colour of soft fruits, which are scarlet,
crimson and purple anthocyamins e.g. cyamidin-3-rutinoside. They are
widely distributed in nature but are more common in the higher plants
and in young tissues, where they occur in the cell sap. Flavonoids
contribute to the taste and flavour of foodstuffs (Bonner and Varner,
.. 1965). Flavonoids when consumed in certain quantity could lead to
serious disorder in the system.
1.8.3 Glycosides
These are the products obtained after condensation of sugar with
different types of organic hydroxyl compounds. These are referred to as
the cardiac-active or cardio-tonic glycosides examples include amygdalin
(Stryer, 1975). In small doses, glycosides promote mild gastric irritation
causing a reflux from the bronchioles. This can be attributed to its wide
usage but in larger dose, they lead to vomiting (Evans, 1989). A larger
number of glycosides and their aglycone have antimicrobial activities.
1.8.4 Cardiac glycosides
This is a derivative of glycosides. It is normally seen in urine,
fruits. It is made up of a non-sugar moiety called aglyone and this non-
sugar moiety, determines the pharmacological effectiveness. The sugar
causes the compounds to be soluble and also power glycosides fixation
to the cardiovascular muscle (Evans, 1989).

I .8.5 Steroidal aglycone
These are non-sugar moiety compounds. They are important
because of their relationship with compounds like the sex hormones,
cortisone, vitamin D and cardiac glycosides (Evans, 1989).
These compounds possess both toxic and play active roles when
they are in the forms of steroids; they have drastic effect on animals.
I .8.6 Saponins
Saponins are useful in the production of soft drinks, beers,
confectioneries, shampoos, soaps, fibre extinguishers and beverages
and this is attributed to its foaming ability (Liener, 1972). They are quite
toxic when injected into the bloodstream and are harmless when taken
by mouth since the sarsaparilla is rich in saponins but is used in the
preparation of non-alcoholic beverages (Evans, 1989).
The highest sapogenin concentration occurs in the reproductive
parts of the plants, the seeds containing 18% trigonenin (Bonner and
Varner, 1965).
Saponin have some medicinal properties, since it has been
reported to have anti-inflammatory, anti-fungal, antimycolic,
bacteriostatic and other biological activities. When ground in a
powdering form, causes violent sneezing.
I .8.7 Tannins
The word "tannin" signifies substances present in plant extracts,
which are able to combine with protein of animal hides, prevent their

putrefaction and the conversion to leather (Evans, 1989). Those tannins
are responsible for the taste qualities of wines, tea and coffee. They are
astrigent and styptic (i.e. the dry sensation felt in the mouth). Tannins
due to their antiseptic properties prevent fungal attacks (Bonner and
Varner, 1965; Evans, 1989). They also have tumorigenic and
carcinogenic effects.
1.8.8 Carbohydrates
Carbohydrates being the most abundant biomolecule with starch
as its precursor is considered of very high pharmaceutical importance. It
is found most abundantly in roots, rhizomes, fruits and seeds. It is also
located in the chlorophyll containing tissues of plants (Evans, 1989).
Monosaccharide, one of the three major size classes of starch are
simple sugars and consist of a single polyhydroxy aldelyde or ketone
unit. It can be oxidized by relatively mild oxidizing agents such as ferric
(I=e3+) or cupric (cu2*) ion. Glucose and other sugars capable of reducing
ferric (Fe3') or Cupric (cu2') ion are termed "Reducing Sugars". This
property is the basis of Fehling's reaction, a qualitative test for the
presence of reducing sugar.
1.9 Literature Survey of Monodora tenuifolia Plant
Taxonomical Classification - Monodora tenuifolia belongs to the:
Kingdom - Plantae
Phylum/Division - Spermatophytae
Class - Dicotyledonae

Subclass - Angrospermal
Order - Archichlamydae
Family - Annonaceae
Gems - Monodora
Specie - Tenuifolia
1.9.1 Characteristics/Morphology of Monodora tenuifolia
Monodora tenuifolia is a widespread and attractive small tree with
very decorative flowers appearing just before the leaves. The fruit is
suspended on a long green stalk with numerous seeds embedded in
whitish sweet smelling pulp. The seed is oblong and pale brown when
fresh with a thin seed coat and hard kernel (Dalziel, 1948). The seed
production is seasonal occurring between April to June. The fruits are
globular and ovoid; 3-4 inch long and about 3-5 inch diameter. The
wood is hard. The seeds are contained in a hard shell and have a very
strong aroma (Neilson, 1965). This plant is commonly called Orchid
flower tree in English, Ehuru Ofia in lgbo (Okafor, 1989) (www.Terv-uren
Xylariumwood Database.com). Monodora tenuifolia is a specie of
calabash nutmeg, the edible seeds yield a nutmeg-flavoured oil which is
used in West Africa for cooking (Eggeling, 1952).
1.9.2 HabitatlEcology of Monodora tenuifolia
Monodora tenuifolia is tropically distributed. It is cultivated in East
India, Malaysia, Sri Lanka, West lndies and Africa. It could be
propagated by stem culturing and budding (Okafor, 1989). The

Monodora species are also found in West Africa and are cultivated in the
southern parts of Nigeria. The trees are very common in Anambra, Abia
and Enugu States.
I .9.3 Local Names
The plant is usually called Orchid flower and is also referred and
called:
Ehuru Ofia - lgbo name
Ehinawosin - lkale name
Lakosin - Yoruba name
Uyenghen - Edo name
Keay, 1989 (www.TervurenXylariumwoodDatabse.com)
I .9.4 Pharmacological uses of Monodora tenuifolia
The plant is widely used in ethnomedicine, especially to relieve
toothache as well as in the treatment of dysentery (Nielson, 1979). When
roasted and ground, the seeds are rubbed on the skin for (unspecified)
skin diseases (Irvine, 1961). This suggests that the seeds of M.
tenuifolia plant could be germicidal or antiseptic.
The roasted ground seeds are chewed, then spat into the hand
and then rubbed across the forehead to relieve headache. The seeds
are also crushed and used as insecticide, while the root relieves
toothache when crushed (Oguntinein et a/, 1989).
Monodora tenuifolia seeds are also used for the treatment of
constipation and as a stimulant (Irvine, 1961). The essential oil from

Monodora tenuifolia seed is used in pharmaceutical and dental
preparation (Talalaji, 1965).
1 . I0 Aim of research
The aim and objective of this research is to:
(1) Investigate the antioxidant activity profile of Monodora tenuifolia
seed extract; and possibly determine the antioxidant principles.

CHAPTER TWO
MATERIALS AND METHODS
2.1 MATERIALS
2.1 .I Plant material
The fresh seeds of the Monodora tenuifolia were collected within
the surroundings of the University of Nigeria, Nsukka Campus, Enugu
State in the month of April to June and the seeds were identified and
authenticated by Mr. A. 0 . Ozioko of Bioresources Development and
Conservation Programme (BDCP), Nsukka.
2.1.2 Animals
Twenty-seven mice (100-120 g) of either sex and 8 Swiss albino
rats (170 - 200 g) of either sex were used for the LD5(-), nitric oxide
scavenging activity and CC14 lipid peroxidation scavenging activity.
These mice and rats were bought from the Department of
Pharmacology & Toxicology Animal House, University of Nigeria,
Nsukka. Animals were kept in cages at 25OC + Z°C. All the animals were
acclimatized to the laboratory conditions for a week before use. They
were fed with standard animal feed and water ad libitum.
2.1.3 Chemicals
All the chemical used in this work were all of analytical grade
commercially available.
The chemicals include:
Absolute ethanol
Acetic acid
BDH
Vicker

Acetone
Ammonia
Ammonium Moybdate
Ammonium solution
Ascorbic acid
Carbon-tetrachloride
Chloroform
Diethyl-ether (40-60°C)
Ethyl acetate
Ferric Chloride
Ferrous Sulphate
Hydrochloric acid (Conc.)
Lead acetate
Methanol
Naphthylethylenediamine dihydrochloride
n-Hexane
Nitric Oxide
Petroleum ether (40-60°C, 60-80°C)
Phosphoric acid
Potassium chloride
Potassium hydroxide
Potassium Iodide
Silica gel
Sodium hydroxide
Sodium nitroprusside
Sulphanilamide
Sulphuric acid (Conc.)
Thiobarbituric acid
Trichloroacetic acid
Tween 80
Vicker
BDH
Merck
Sigma
BDH
Numex
May and Baker
Merck
BDH
Vicker Lab. Ltd.
BDH
Merck
Sigma
Fluka
BDH
Riedel-de Haen
Merck
Frutaran
BDH
BDH
BDH
Merck
Merck
MAY AND BAKER
BDH
Sigma
BDH
Sigma
Sigma
Sigma

2.1.4 EquipmentlApparatus
The equipment used for this includes:
Column
Glass wares (test tubes, beakers etc.)
Mortar and Pestle
Refrigerator
Rotary evaporator
Spatula
Separating funnel
Soxhlet apparatuss
Spectrophotometer
Spreader & TLC applicator
TLC plates
Weighing balance
Water bath
2.1.5 Reagents for phytochemical analysis
Aluminium chloride solution (0.5%)
Ammonium solution
Dilute sulphuric acid solution
Dragendorff's reagent
Ethanol (45%)
Ferric chloride solution (5%)
Lead sub-acetate solution (1 5%)
Mayer's reagent
Pyrex
Pyrex
Life Scan Company
Thermocool
Mecitherm (PFV)
Pyrex
Giant Instrument
Mecitherm (PFV)
Novospec (LKB)
Merck
Merck
Meter HAS 6
Mecitherm (PFV)
Wagner's reagent

2.1.6 Reagents for nitric oxidellipid peroxidation scavenging assay
Ferrous sulphate (1 5 mM)
Naphthylene diamine dihydrochloride (0.1 %)
Phosphoric acid (2%)
Potassium chloride (0.15 m)
Potassium sulphate buffer (pH 7.4)
Sodium nitroprusside (5 pm)
Standard phosphate buffer (0.025 m, pH 7.4)
Thiobarbituric acid (0.67%)
Thiobarbituric acid (0.67%) in acetic acid (50%)
Trichloroacetic acid (1 0%)
. 2.1.7 Preparation of reagents for thin-layer chromatographic separation of lipid components
Preparation of chromatoplates
The clean thin-layer chromatographic plates were cleaned with
acetone and placed on an applicator. The silica gel was spread on the
plates using a spreader.
The plates were allowed to dry in air and stored in a dust free
desiccator. Before usage, the plates were activated in an oven at I 10°C
for an hour and left to cool before usage.

55
Solvents for chromatographic studies
Petroleum ether, diethylether and acetic acid solvent mixture
(1 80:20:2 vlvlv)
n-hexane, diethylether and acetic acid solvent mixture (80:20:2
vlvlv).
n-hexane and benzene (85:l5 vlv) solvent mixture
Chloroform, methanol and water (95:20:2.5 vlvlv) solvent mixture
Chloroform, .acetone, methanol and acetic acid (50:32O: 10: 10.5
v/v/v/v) solvent mixture.
n-hexane, diethylether and acetic acid (69:29:2 vlvlv) solvent
mixture.
Methodology
' 2.2.1 Extraction of oil from Monodora tenuifolia Seeds
The freshly collected seeds of the Monodora tenuifolia were sun
dried and powdered using a pistle and mortar. The powder was defatted
with petroleum ether (40 - 60°C) using soxhlet apparatus.
The whole filtrate was allowed to evaporate at room temperature
leaving the oil.
% yield of oil = Weight of Oil 100 -X-
Weight of Sample I
2.3 Thin-Layer chromatographic technique
The extracted oil of Monodora tenuifolia was prepared with
chloroform (1:5) (0.1 ml of oil was dissolved in 0.5 ml chloroform). This

56
solution was spotted 2 cm from the base of the chromatographic plates.
Before developing in the appropriate solvent system, they were allowed
to dry. Micropipetes were used for spotting of the sample.
2.3.1 Determination of lipid components
A chromatographic tank is used for the thin layer chromtography
and the solvent sy'stem (pet-ether, diethylether and acetic acid (180:20:2
V/V/V) was poured inside with vigorous shaking so that it will systemize.
This solvent was allowed to equilibrate for about 1 hr. The spotted
plates were placed inside the tank containing the solvent system and
allowed to develop for 45 minutes.
The spotted plates were left for a while to allow for separation
shown by relative distance moved by different components of the oil.
Development of the plate was done in an iodine tank with iodine vapour.
On observation, it was discovered that there were seven distinct
components. Their relative distance was measured and the solvent front
was determined. From the measurements, specific Rf values were
calculated using the formular:
Rf = Distance moved by each component Distance moved by solvent front
On using different chemical agents, series of thin-layer
chromatography were conducted to determine the presence of
aminolipids, glycolipids, free fatty acids and cholesterol.

Determination of aminolipids
The spotted plates were placed in the chromatotank containing
the solvent system to separate. The plates were then sprayed with
ninhydrin reagent (0.2 g of ninhydrin dissolved in 100 mi of ethanol).
The sprayed plates were heated at 120°C for 5 minutes in a humid
atmosphere. A red violet colour indicates the presence of aminolipids.
Determination of free fatty acids
After spotting the chromatographic plates with the lipid sample, it
was placed in the chomatotank containing the solvent system to
separate. The plate was then sprayed with ferric chloride solution (1.0 g
of ferric chloride in 100 ml of H20). A Rose-Violet spot on the plate
shows the presence of free fatty acid.
Determination of glycolipids
The plate having been spotted with the oil dissolved in chloroform
was developed in the solvent system. After air-drying the plates, was
sprayed with a-naphthol reagent (0.5 g of a-naphthol dissolved in a
solution containing equal volumes of 25 ml methanol and H20) and
subsequently heated in an oven at 120°C for 5 minutes. A purple blue
spot indicates the presence of glycolipids.
Determination of c.holesterol
The plate from the chromatotank was air-dried and sprayed with
acidic ferric chloride solution. The sprayed plate was heated for 2 - 3

minutes, in an oven at 100°C. Red violet colour/spots indicate the
presence of cholesterol.
2.4 Determination of vitamin in Monodora tenuifolia
Preparation of plant extract
In preparation of plant extract, a quantity of 0.1 g of the sample
were each dissolved in 10 ml of chloroform and 10 ml of ethanol in 2
different test tubes. The mixture was shaken for 30 minutes, filtered with
filter paper and the filtrate was used for determination of the presence of
Vitamins A and E.
Vitamin A assay
Two drops of the extracts were added to 5 drops of chloroform
using a pipette with the addition of 2 drops of concentrated H2SO4. A
' brown colour indicates the presence of Vitamin A.
Vitamin E assay
In assaying for vitamin E, a quantity of 0.1 g of the extracts from
the sample was added to 1 ml of ethanol and 10 drops of concentrated
HN03. The presence of red colour indicates the presence of Vitamin E.
2.5 Phytochemical analysis of Monodora tenuifolia Seed
Test for alkaloids
Some quantity of the Monodora tenuifolia sample was ground in a
mortar and some quantity of about 0.2 g of the macerated sample was
boiled with 5 ml of 2% HCI on a steam bath. This was filtered and 1 ml
portion of the filtrate was treated with 2 drops of the following reagent:

Dragendorff's Reagent (Bismuth potassium-Iodide Solution: A
red precipitate was observed indicating the presence of
alkaloids.
Mayer's Reagent (Potassium Mercuric Iodide Solution): A
creamy white coloured precipitate indicates the presence of
alkaloids.
Wagner's Reagent (Iodine in Potassium Iodide Solution): A
reddish brown precipitate indicates the presence of alkaloids.
Picric Acid (1%): A yellow precipitate indicates the presence
of alkaloids. The following colours were observed using the
reagents: Dragendorffs reagent (red precipitate), Mayer's
reagent (white precipitate), 1 % picric acid (yellow precipitate
and Wagner's reagent (brown precipitates).
' Test for the presence of flavonoids
To test for the presence of flavonoids, 0.2 g of the macerated
sample was heated with 10 ml of ethylacetate in a boiling water for 3
minutes. The mixture was filtered and the filtrate used for the following
tests:
(i) 4 ml of the filtrate was shaken with 1 ml of 1% aluminium
chloride solution and observed for light yellow coloured
ethylacetate layer. A yellow coloration observed in the
ethylacetate layer indicates the presence of Flavonoids.
(ii) 4 ml of the filtrate was shaken with 1 ml of dilute ammonia.
The layers were allowed to separate. A yellow coloration

Dragendorffs Reagent (Bismuth potassium-Iodide Solution: A
red precipitate was observed indicating the presence of
alkaloids.
Mayer's Reagent (Potassium Mercuric Iodide Solution): A
creamy white coloured precipitate indicates the presence of
alkaloids.
Wagner's Reagent (Iodine in Potassium Iodide Solution): A
reddish brown precipitate indicates the presence of alkaloids.
Picric Acid (1%): A yellow precipitate indicates the presence
of alkaloids. The following colours were observed using the
reagents: Dragendorff's reagent (red precipitate), Mayer's
reagent (white precipitate), 1% picric acid (yellow precipitate
and Wagner's reagent (brown precipitates).
* Test for the presence of flavonoids
To test for the presence of flavonoids, 0.2 g of the macerated
sample was heated with 10 ml of ethylacetate in a boiling water for 3
minutes. The mixture was filtered and the filtrate used for the following
tests:
(i) 4 ml of the filtrate was shaken with 1 ml of 1% aluminium
chloride solution and observed for light yellow coloured
ethytacetate layer. A yellow coloration observed in the
ethylacetate layer indicates the presence of Flavonoids.
(ii) 4 ml of the filtrate was shaken with 1 ml of dilute ammonia.
The layers were allowed to separate. A yellow coloration

observed at the ammonia layer indicates the presence of
Flavonoids.
Test for the presence of glycosides
To test for the presence of glycosides, 2.0 g of the ground sample
was added to 10 ml of water, the solution was heated on a water bath for
5 minutes, filtered and used for the following tests:
(i) To 5 ml of the filtrate was added 0.2 ml of Fehlings solution A
and Fehlings solution B until it turned alkaline (tested with
litmus paper) and heated on a water bath for 2 minutes. A
brick red colour was observed.
(ii) 15 ml of dilute H2SO4 instead of water was used, the above
process was repeated and the quantity of the precipitate
formed was compared with that of the former experiment.
Test for the presence of cardiac glycosides
To test for the presence of cardiac glycosides, 0.1 g of the sample
was warmed with 5 ml of chloroform in a water bath and decanted. The
decanted solution was evaporated to dryness on a water bath and the
residue dissolved in 3 ml glacial acetic acid containing a drop of ferric
chloride. The concentrated sulphuric acid was poured carefully by the
side of the tube. A brown ring was observed at the interface.
Test for presence of cyanogenic glycosides
In the test for the presence of cyanogenic glycosides, 0.1 g of the
ground sample was placed in a conical flask and enough water was

added to cover the sample. A piece of sodium picrate paper was
suspended in the flask placed on a water bath for one hour. A colour
change from yellow to white was observed.
Test for presence of anthracene glycosides
The test for the presence of anthracene glycosides, 5 ml dilute
sulphuric acid and 5 ml ferric chloride solution was added to 0.1 g of the
ground sample of Monodora tenuifolia. The resultant mixture was boiled
for 5 minutes, cooled and filtered into a 50 ml separating funnel. The
filtrate was shaken with equal volume of carbon tetrachloride. The lower
organic layer was carefully separated into a test tube and 5 ml of dilute
ammonia solution was added to it and was gently shaken. A brown
coloration in the ammonium layer was observed.
Test for presence of 0- and C-glycosides
To test for the presence of 0- and C-glycosides, 0.1 g of the
ground sample was heated with 5 ml of water in a boiling water bath for
15 minutes, cooled and filtered. The filtrate was treated with 5 ml of 25%
HCI and the solution heated once more for 15 minutes, cooled with 10 ml
of diethylether in separating funnel. Both phases were checked for
purple red colour in the aqueous layer.
The combined ether extract was shaken with 5 ml of 3.5% dilute
ammonia solution and observed. The acid phase was titrated with 0.5 g
of ferric chloride and then heated for about 30 minutes in a water bath
and cooled. The solution was extracted with 10 ml of chloroform. The
layer was washed with 5 ml of water and then 3 ml of 3.5% dilute

ammonia solution was added. If a dark red precipitate is observed, then
it shows a positive result.
Test for presence of tannins
In the test of tannins, 20 g of the ground sample was boiled with 5
ml of 45% ethanol for 5 minutes, cooled and filtered, the filtrate was used
for the following tests:
(i) Lead Sub-Acetate:
In the lead sub-acetate test, 0.1 ml of the filtrate was
added a few drops of lead sub-acetate solution and observed
for a gelatinous white precipitate.
(ii) Bromine Water:
For the bromine water test, 1.0 ml of the filtrate was
added 0.5 ml bromine water and a pale yellow precipitate was
observed.
(iii) Ferric Chloride:
For the ferric chloride test, 0.1 ml of filtrate was diluted
with distilled water and a few drops of Fed3 solution was
added. A thick yellow colour was observed, which later
turned black.
Test for presence of saponins
The test for the presence of saponins, 0.1 g of the ground sample
was boiled with 5 ml of distilled water for 5 minutes and decanted while
still hot. The filtrate was used for the following tests:

(i) Frothing Test:
In the frothing test, 1 ml of the filtrate was diluted with
4 ml of distilled water, shaken vigorously. A stable froth was
observed on standing the test tube.
(ii) Emulsion Test:
For the emulsion test, 1 ml of the filtrate was added to
2 drops of olive oil. The mixture was shaken and there was
formation of an emulsion.
Test for presence of reducing Sugar
To test for reducing sugars, 1.0 g of the ground sample was
shaken vigorously with 5 ml of distilled water and filtered. About 1 ml
portion of the filtrate was added to equal volume of Fehlings solutions A
and B and shaken vigorously. A brick red precipitate was observed.
Test for carbohydrate (Molish Test)
The test for carbohydrates, 0.1 g of the ground sample was
shaken vigorously with water and filtered. To the aqueous filtrate was
added few drops of Molish reagent followed by vigorous shaking.
Then 1 ml of concentrated sulphuric acid was carefully added to
form a layer below the aqueous solution. A brown ring was observed at
the interface.
Test for steroidal aglycon
To test for the presence of steroidal aglycon, 10 ml of lead acetate
solution (9% wlv) and 20 ml aqueous ethanol were added to about 1 gm
of the ground material in a 100 ml conical flask. This was placed in a

boiling water bath for about 2 minutes, then cooled and filtered. The
filtrate was extracted twice with 15 ml chloroform. The lower chloroform
layer was retained while 5 ml of the chloroform extract was evaporated
to dryness on a water bath. To the residue, 2 ml of 3,5-diintrobenzoic
acid solution (2% on ethanol) and 1 ml NaOH solution were added and
then observed for pinkish coloration.
2.6 Column chromatographic technique
The column was set up by clamping in a vertical position, a small
quantity of glass wool was placed in the bottom of the column. These
retain solid materials in the column while allowing liquid to percolate
freely through them. Air trapped in the glass wool was removed by
pushing it with a glass rod.
The medium i.e. dry silica gel was poured in, such that it allowed
uniform packing of the column and a free flow of solvent through it in
order to attain optimum results. The silica gel was poured into the tube a
little at a time and allowed to separate under gravity until a column of the
correct height was attained. A little of the solvent was poured into the
column to a depth of a few inches.
Finally, the tap at the bottom of the column was opened to allow
the liquid to run out until it just covers the top of the silica gel.
2.6.1 Elution of the column
The extract to be separated is mixed with some quantity of the
silica gel until a fine mixture is gotten. Thus mixture is introduced into
the packed column to a depth of a few inches.

Some of the silica gel is further introduced to a depth of few
inches covering the extract - silica gel mixture. The first eluting solvent
petroleum ether is introduced and allowed to flow steadily through the
medium adjusting the clips to give the required rate. The flow is
continued until the mixture is completely separated into its components.
After the lSt elution, using petroleum-ether (60-80°C), which
readily must have eluted some proportion of the component in the
'mixture further elution is carried out with a second solvent diethyl-ether.
Here, the separated compounds emerge in the eluate in different
fractions. This results in different fractions of eluant viz: petroleum ether
(60-80°C) fraction (F1) and Diethyl ether Fraction (F2).
2.7 Nitric oxidellipid peroxidation scavenging assay
The crude extract as well as fractions F1 and F2 were evaporated
to dryness to remove the solvents and suspended in 20% (vlv) of Tween
80 and then used for the nitric oxidellipid peroxidation scavenging tests.
2.7.1 Scavenging o f nitric oxide
Sodium nitroprusside 1 ml of 15 pglml was incubated with
different concentrations (100, 200, 400, 800 pglml respectively) of F1
dissolved in standard phosphate buffer (0.025 m, pH 7.4) and the tubes
were incubated at 25OC for 5 hours. After 5 hrs, 0.5 ml of incubation
solution was removed and diluted with 0.5 ml of Griess reagent
(prepared by mixing equal volume of 1% suphanilamide in 2%
phosphoric acid and 0.1% naphthylene diamine dihydrochloride in

water). The chromophore formed was read at 546 nm in a
spectrophotometer. The control experiment was also carried out in
similar manner, using distilled water in the place of F1. The experiment
was repeated in same manner using F p and crude pet-ether extract. The
tests with each of the extracts - F1, Fp and crude extract were carried out
in triplicates and % scavenging activity was calculated using the
following formular:
Abs T = Mean Absorbance of the Extracts (Fl, F2 and crude
extract)
Abs C = Mean Absorbance of the control (Distilled water).
The activity of each fraction and extract was compared with
ascorbic acid of different concentrations (100, 200, 400 and 800 pglmi),
which was used as a standard antioxidant.
2.7.2 Carbon tetrachloride induced lipid peroxidation test
The method of (Comporti, 1989) was used in the CCL4 induced
lipid peroxidation assay. Eight adult Swiss albino rats weighing 170 -
200 g were anaesthetized in chloroform and dissected to remove the
liver and their livers were carefully removed.
The total liver from the rats weighing 8 g was homogenized with
20 ml of ice-cold potassium chloride. Aliquots of 0.5 ml of homogenates
were taken in different test tubes. These test tubes with different
concentrations of Fi (100, 200, 400 and 800 pg/ml) were incubated at

37OC in a constant shaker water bath for 45 minutes with 1.5 ml of
potassium sulphate (pH 7.4), 2 ml of potassium chloride and finally 10 ml
of carbon tetrachloride (CC14) was added.
The reaction was stopped by the addition of 2 ml of 10% (wlv)
trichloroacetic acid and after incubation, the contents of each test tubes
were centrifuged at 4000 rpm for 10 minutes and 2 ml of the clear
supernatant was transferred to another test tubes and finally 2 ml of
0.67% (wlv) of thiobarbituric acid was added to the test tubes and
heated in a boiling water bath for 15 minutes. Each of the test tubes was
cooled and the mixture adjusted to pH 12.5 with potassium hydroxide.
The chromophore was measured at 543 nm in a Novaspec
Spectrophotometer. The control test was carried out in same manner
but distilled water was used in place of F1. The test was also carried
with different concentrations (100, 200, 400, 800 pglml) of the other
extracts (F2 ands crude extract) all in triplicate and the %
scavenginglinhibition was calculated using the following:
where:
Abs T = Mean Absorbance of the Extracts (F1, F2 and crude
extract)
Abs C = Mean Absorbance of the control (Distilled water).

Again activities of each extracts (F1, F2 and crude extract) were
compared with that of ascorbic acid of different concentration (100, 200,
400, 800 pglml), which was used as a standard antioxidant drug.
2.8 Statistical analysis
The results were analyzed with one-way ANOVA expressed as
Mean + SEM. The FISCHER LSD Posthoc test was used to test the
differences between mean of treated and control groups and were
regarded significant at pc 0.05.

CHAPTER THREE
RESULTS
3.1 Percentage yield of extract
In the present study, the % yield of the crude extract was found to
be 18.9%.
3.2 The result of the phytochemical analysis of Monodora tenuifolia seed
The result shows that the Monodora tenuifolia seed contains
alkaloids, carbohydrates, cardiac glycosides, cyanogenic glycosides,
flavonoids, glycosides, proteins, saponins, steroidal aglycon and tannins
while anthracene glycosides, 0 and C glycosides and reducing sugar
were absent (Table 3.1).

Table 3.1: Results of phytochemical tests
COMPONENTS
Alkaloids
Anthracene glycosides
Carbohydrate
Cardiac glycosides
Cyanogenic glycosides
Flavonoids
Glycosides
0 and C glycosides
Protein
Reducing sugar
Saponins
Steroidal aglycones
Tannins
RESULTS -
Present
Absent
Present
Present
Present
Present
Present
Absent
Present
Absent
Present
Present
Present

3.3 The result of acute toxicity test: (LDS0) of the extract
In the acute toxicity test, administration of 10, 100 and 1000
mglkg body weight of the extract caused no death in the first phase of
the test. In the second phase of the test, no death was recorded even
up to 5000 mglkg body weight.
The LD50 was therefore greater than 5000 mglkg (Table 3.2).

Table 3.2: Result of acute toxicity test
1 ST PHASE
2ND PHASE
CRUDE EXTRACT
Crude extract
Crude extract
DOSE (mglkg)
10
MORTALITY GROUP
013

3.4 Nitric oxidellipid peroxidation scavenging
3.4.1 Effect of pet-ether extract and fractions fl and f2 on carbon tetrachloride-induced lipid peroxidation in rat liver homogenate
Lipid peroxide formation by CC14 was inhibited by the pet-
ether(40-60°C) extract, the pet-ether fraction (60-80°C) (F1) and
diethylether fraction (F2) at all tested doses (100 - 800 pglml). The
percentage inhibition of peroxide formation was increased in a,dose
dependent manner for the pet-ether (40-60°C) extract and diethylether
fraction (F2). While for the pet-ether fraction (F1) the percentage
inhibition showed a result independent of the dose (Table 3.3)..

Table 3.3: Effect of extract and fractions on CCL4 induced lipid peroxidation
EXTRACT
Pet-ether fraction (F,)
Diethlether fraction (F2)
Pet-ether Extract (crude)
Vitamin C (standard)
BLANK
CONC. pglml
100
200
400
800
100
200
400
800
100
200
400
800
100
200
400
800
ABSORBANCE (Mean f SE)
0.49 + 0.03*
0.54 + 0.06*
0.48 + 0.04*
0.45 + 0.01*
0.64 + 0.01*
0.59 + O.OO*
0.45 + O.OO*
0.43 + 0.01*
0.64 + 0-OO*
0.49 + 0.01*
0.45 + O.OO*
0.40 f 0.00*
0.41 + 0.04"
0.34 + 0.02*
0.24 + O.OO*
0.12 _+ o.oo*
1.30 + 0-00* $ BLANK
INHIBITION ( O W
62.07
57.93
63.33
65.40
51.03
54.33
64.40
66.67
50.80
62.30
65.67
69.50
68.47
73.87
81.50
90.53

0 100 200 300 4 00 500 GO0 700 000 900
Concentration (uglnil)
Fig. 3: Effect of,extract and fractions on carbon tetrachloride-induced lipid peroxidation in the rat liver homogenate
--+a- Pel Elher Fraction (FI ) -0- Oielhylelher fraclion (F2)
-A- Oil (Pel elhcr exlracl) - -n- - Vilamin C (Slandard)

3.4.2 Effect of nitric oxide scavenging activity of pet-ether extract and the fractions
The Monodora tenuifolia pet ether (40-60°C) seed extract and the
fractions; pet-ether (60-80°C) fraction (Fl), and diethylether fraction (F2)
showed significant free radical, scavenging action against nitric oxide
(NO) induced release of free radicals. Diethylether fraction and the pet-
ether (40-60°C) extract showed an increased percentage inhibition,
which was dose dependent while the pet-ether (60-80°C) fraction (F1)
showed a percentage inhibition, which was independent of the dose.
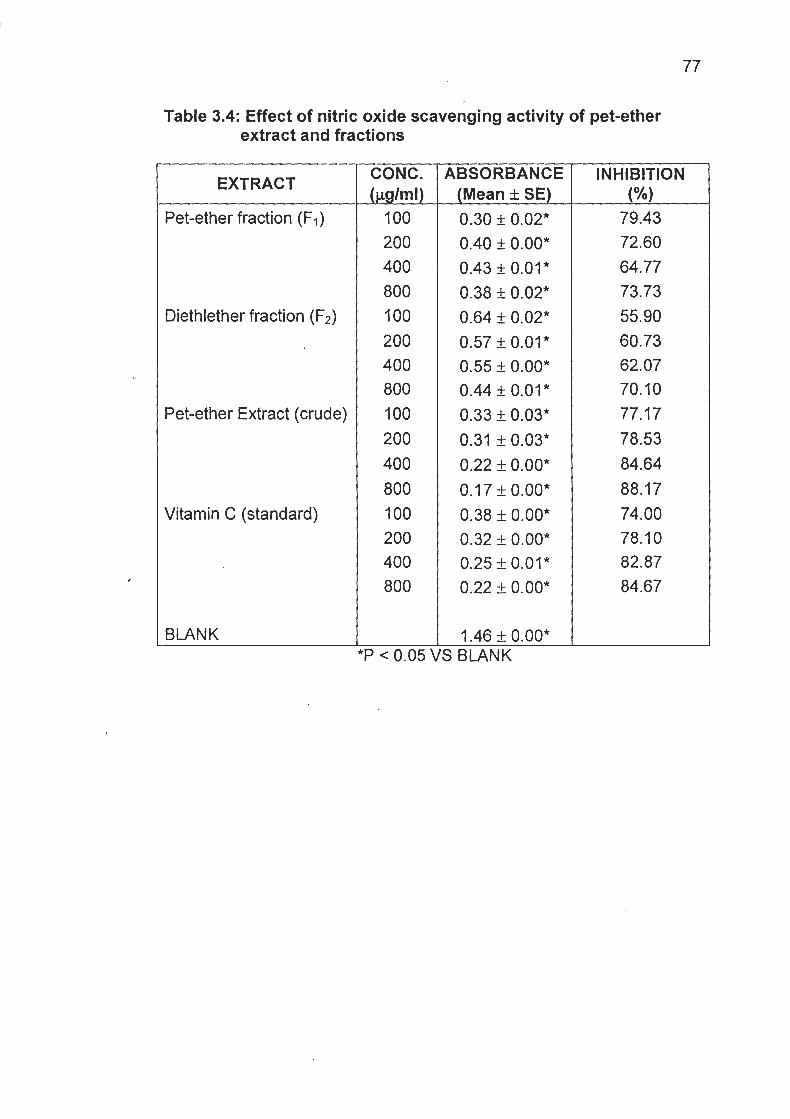
Table 3.4: Effect of nitric oxide scavenging activity of pet-ether extract and fractions
EXTRACT
Pet-ether fraction (F1)
Diethlether fraction (F2)
Pet-ether Extract (crude)
Vitamin C (standard)
BLANK
CONC. ( pglml)
100 200 400 800 100 200 400 800 100 200
400
800 100 200 400 800
P < 0.05 \ S BLANK
ABSORBANCE (Mean f SE)
0.30 + 0.02" 0.40 + O.OO* 0.43 + 0.01* 0.38 + 0.02* 0.64 + 0.02* 0.57 + 0.01* 0.55 + O.OO* 0.44 + 0.01* 0.33 + 0.03* 0.31 + 0.03*
0.22 + O.OO* 0.17 + 0.00" 0.38 f 0.00* 0.32 + O.OO* 0.25 + 0.01* 0.22 + O.OO*
INHIBITION (%)
79.43 72.60 64.77 73.73 55.90 60.73 62.07 70.10 77.17
78.53 84.64
88.17 74.00 78.10 82.87 84.67

0 100 200 300 100 ' 500 600 700 000 900
Concenlralibn (uglm)
Fig. 4: Effect of extract and fractions on nitric oxide scavenging activity
--a- Pel Elher Fraction ( F I ) --I+-- Dielhylether fraction (F2)
-a- Oil (Pel elhcr cxlract) . W . . Vilarn~n C (Standard)
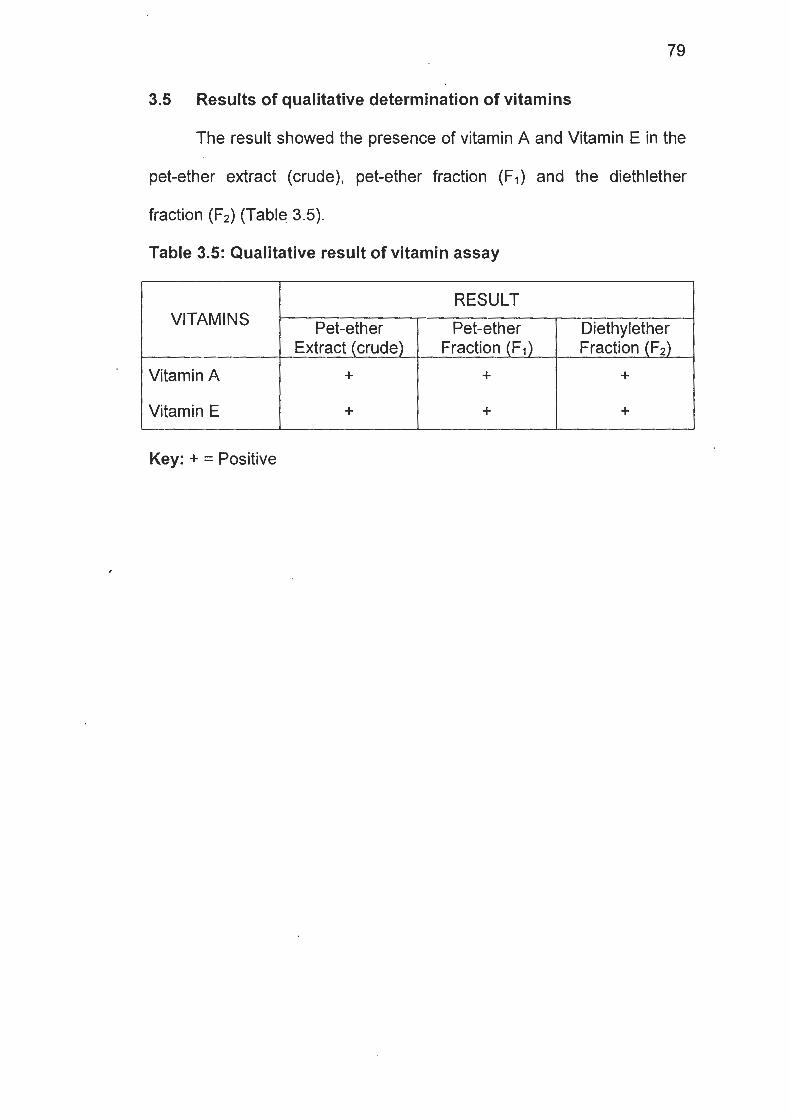
3.5 Results of qualitative determination of vitamins
The result showed the presence of vitamin A and Vitamin E in the
pet-ether extract (crude), pet-ether fraction (F,) and the diethlether
fraction (F2) (Table 3.5).
Table 3.5: Qualitative result of vitamin assay
Vitamin A I + I + I + I
VITAMINS
Key: + = Positive
RESULT
Vitamin E
Pet-ether Extract (crude)
+
Pet-ether Fraction (F,)
Diethylether Fraction (F2)
+ +

3.6 Lipid Compounds Present inthe Seed Extract
The result shows the different classes of lipids present in
Monodora tenuifolia seed. The result shows the presence of
aminolipids, free fatty acids, glycolipids, phospholipids and sterols. This
is shown in Table 3.6.
Table 3.6: Result of lipid components present in the seed extract
"
Key: +ve = Positive
Free fatty acids Glycolipids Phospholipids Sterols
LIPID COMPONENT Aminolipids
+ve +ve +ve +ve
RESULTS +ve

CHAPTER FOUR
DISCUSSION
In traditional system of medicine, certain herbs are claimed to
have medicinal properties ranging from the relief of pains to curing of
diabetes etc. The claimed therapeutic effects have to be verified in a
scientific manner. In the present study, one such drug Monodora
tenuifolia seed extract was studied for its possible antioxidant
properties..
Monodora tenuifolia is a tropical plant found in forest regions and
they appear in colonies (Dalziel, 1948) and is mostly used as spices or
condiments in soup as well as in pharmaceutical preparations as a
dental cure (Talalaji, 1965). From the study, it is evident that the extract
of seeds of Monodora tenuifolia has promising antioxidant activity.
In the present study, the percentage (%) yield of the extract was
found to be 18.9%, which is relatively low when compared to a previous
study on the plant. In a previous study (Esuoso et al, 2000) reported
that Monodora tenuifolia seed extract had a yield of 34.7-68.8%.
The possible difference in the O h yield could be as a result of
geographical and climatic factors, which has been found to affect plant
constituents, or time of collection of the seed, method of storage, the
variety of the parent plant and the nature of the soil on which it is planted
(Alam et all 1982).

In the acute toxicity test, administration of up to 5000 mg/kg
caused no death, suggesting that the extract could be safe even at very
high doses. Monodora tenuifolia seed was found to contain a lot of
secondary plant metabolites namely: alkaloids, carbohydrates, cardiac
glycosides, cynogenic glycosides, flavonoids, glycosides, proteins,
saponins, steroidal aglycones and tannins. Alkaloids at high
concentration, has been found to produce a variety of toxic effects on
" animals. They also protect the plant from the effect of singlet oxygen
(Bonner and Varner, 1965). These plant constituents are known to be
biological active, eliciting a variety of pharmacological actions such as
antioxidant effects (Bauer et a/., 1996). It can also be concluded that the
antioxidant activity of the extract could be attributed to flavonoids, which
. are fourid in antioxidant plant such as Aspidium cacutarium (Ghoghari et
all 2006), Phyllanthus debilis klein ex Willd (Kumaran and Karunakaran,
2006) as well in Tephrosia purpurea (Jain ef al., 2006).
In the Nitric oxide scavenging test, systemic administration of the
pet-ether (40-60°C) extract, pet-ether (60-80°C) fraction and
diethylether fraction showed significant free radical scavenging action
against nitric oxide (NO) induced release of free radicals.
Sodium nitroprusside serves as a chief source of the free radicals.
The absorbance of the chromophore formed during diazotization of the
nitrite with sulphanilamide and subsequent coupling with napthyethylene
diamine is used as the marker of NO scavenging activity (Mukherjee,

1989). The chromophore formation was not complete in the presence of
Monodora tenuifolia seed extract (pet-ether (40-60°C) extract),
diethyether fraction (F2) and pet-ether (60-80°C) fraction, which
scavenged the NO formed from the sodium nitroprusside. The
absorbance for the pet-ether extract and the diethylether fraction (F2)
decreases as their concentration increases in a dose-dependent
manner.
In terms of the vitamin content, Monodora tenuifolia seed extract
contained vitamin A and vitamin E (Table 3.6). These vitamins offer
protective measures from oxidative stress by quenching free radicals
capable of causing cellular damage. Vitamin A in the form of
carotenoids can exhibit maximum inhibition of peroxidation of linolenate
. at low concentration (Krinsky and Deneke, 1982). At high O2 tensions,
a-tocopherol is the most effective antioxidant (Burton, 1989).
Many antioxidant agents modify oxidative degradation in the body
by accelerating the destruction or antagonizing the action of mediators of
oxidative processes e.g. the reactive species superoxide (02-), hydrogen
peroxide (H202), hydroxyl radical (HO.), nitrogen oxide (NO.) and
hypochlorous acids (HOCI) (Akah et al., 1997). These free radicals play
an important role in the genesis of various diseases such as
inflammation, arthrosclerosis, arthritis, rheumatism, cirrhosis of liver,
lung damage, cataract, aging and ischemia (Halliwal, 1994). There are
various pharmacological options in the treatment of these diseases,

which are geared towards protecting the human body from damage due
to free radicals (Osawa et a/., 1990).
Lipid peroxidation has been implicated in the apthogenesis of
various diseases including arthritis. It is well established that
bioenzymes are very much susceptible to lipid peroxidation, which is
considered to be the starting point of many toxic as well as degradative
processes. Thus the presence of flavonoids and alkaloids, as well as
vitamins A and E may be responsible for the antioxidant effects
observed with Monodora tenuifolia seed extract in the presence study.
The lipids present in this study are important viz., aminolipids, free
fatty acids, glycolipids, phospholipids and sterols. They equally play
important roles in both cellular functions and in the free radical
- reactions.The results of present study with Monodora tenuifolia seed
extract and fractions have good correlation with the therapeutic use of
Monodora tenuifolia in the possible treatment of oxidative stress
conditions. Plants that belong to Annonaceae family are rich in
flavonoids and bioflavonoids and are known to have antioxidant activity.
Monodora tenuifolia seed extract contains important pharmacological
compounds, alkaloids, flavonoids, and vitamins A and E as well as many
important lipids; arhinolipids, free fatty acids, glycolipids, phospholipids
and sterols.

REFERENCES
Akah, P. A,, Gamaniel, K. S., Wambebe, C. O., (1997). Evaluation of Nigerian Traditional Medicine: Effects of Gkani, a herbal anti- asthmatic drugs. Journal of Ethnopharmacology 55: 87-92.
Alam, M. N., Farug, M. O., Hossain, M. E. and Alam, M. J. (1982). Investigation of Rubber seed oil Bangladesh. J. Sci. Ind. Res. xvii, (3-4), 200-204.
Ames, B. N., ~a thcar t , R., Schwiers, E., and Hochstein, P. (1981). Uric acid provides an antioxidant defense in humans against oxidant and radical caused aging and cancer; a hypothesis. Proc. Natl. Acad. Sci. USA. 78: 6858-6862.
Andraws, R., Berger, J. S. and Brown, D. L. (2005). Effects of antibiotic therapy on outcomes of patients with coronary artery disease. JAMA. 293: 2641-7.
Antoniades, H. N. and Owen, A. J. (1982). Growth factors and regulation of cell growth. Ann. Rev. Med., 33: 445.
Bauer, R., Probosthe, A., Latter, H. Wasgherredecker, W., Matheissen, U. (1996). Phytomedicine, J. Pharm. Col. 4: 305-308.
Bendich A. and Olson, J. A. (1989). Biological actions of Carotenoids. FASEB J. 3: 1927-1932.
Bendich, A. (1989). Carotenoids and Immune Response. J. Nutr. 119: 112-115.
Bonner, J. and Varner, J. E. (1965). Plant Biochemistry Academic Press London, pp. 252-703.
Boogerd, W., Vander Sande, J. J. and Van Zandwijk, N. (1999). Teniposide sometimes effective in brain metastases from non- small cell lung cancer. J. Neurooncol. 41 : 285-289.
Borg, D. C. and Schaich, K. (1989). Pro-oxidant action of antioxidants. CRC Handbook of Free Radicals and ~ntioxidants in Biomedicine. (Miquel, J., Quintanilha, A. T., and Weber, H., eds) CRC Press, Boca Raton, Florida. Vol. I. pp. 12-45.
Borg, D. M., (1993). Oxygen free radicals and tissue injury; a reference in oxygen free Radical in Tissue Damage (Tarr, M. and Sampson, eds.) C. V. Press, Mosby, by St. Louis, pp. 10-32.

Buettner, G. R., and Jurkiewicz, B. A. (1993). Ascorbate free radical as a marker of oxidative stress: an EPR study. Free Rad. Biol. Med. 14; 49-55.
Buettner, G. R., and Jurkiewicz, B. A. (1996). Catalytic Metals, Ascorbate and Free Radicals: Combination to Avoid. Radiation Res. 145: 532-541.
Burton, G. W. (1989). Antioxidant action of carotenoids. J. Nutr. 119: 109-111. .
Burton, G. W., and Ingold, K. U. (1984). Beta-Carotene: an unusual type of lipid antioxidant . Science 224; 569-573.
Caughey, W. S., and Watkins, J. A. (1985). Oxy-radical and Peroxide formation by haemoglobin and myoglobin. In CRC Handbook of Methods for oxygen Radical Research (Greenwald R. A., ed): pp. 95-1 04.
Cavalieri, E. L. and Rogan, E. G. (1984). One-electron and two-electron oxidation in aromatic hydrocarbon carcinogenesis. In Free Radicals in Biology (Pryor, W. A., ed.) I : 323-370.
Chang, T. K., Weber, G. F., Crespi, C. L., and Waxman, D. J. (1993). Differential activation of cyclophosphamide and ifophamide by cytochromes p-450 23 and 3A in human liver microsomes. Cancer Res. 53: 5629-5637.
Chen, G. X., and Asada, K. (1989). Ascorbate peroxidase in tea leaves: Occurrence of two lsozymes and the differences in their enzymatic and molecular properties. Plant Cell Physiol. 30: 987- 998.
Comporti, M. (1989). Three models of free radical induced cell injury, Chem. Biol. Interact, 72: 1-56.
Current Medicinal Chemistry (2005). Metals, toxicity and oxidative stress. Free Radical heory of aging. 12(10): 1 I 61 -1 208.
Cutler, R. G. (1984). Antioxidants, aging, and longevity. In Free Radicals in Biology, Academic, Orlando, Florida. Vol. I. pp. 371-428.
Dalziel, F. M. (1948). The useful plant of West Tropical Africa, being an appendix to the flora of West Tropical Africa by Hutchinson. J. and Dalziel, J. M. The Crown Agent for the Colonies, London. S.W. T. pp. 5-9.

De Lau, L., Giesbergen, P., Rijk, M., Hofman, A., Koudstaal, P. and Breteler, M. (2004). Incidence of Parkinsonism and Parkinson disease in a general population: the Rockenstein study. Neurology 63(7): 1240-4.
Eggeling, W. J. (1952). The Indigenous Trees of the Uganda protectorate (Revised and enlarged by Ivan R. Dale). Government printer, Entebbe Uganda, Crown Agents for the Colonies London. pp. xiii and 491.
Epstein, J. H. (1977). Effects of Beta-Carotene on Ultraviolet induced cancer formation in the hairless mouse skin. Photochem. Photobiol. 25: 21 1-21 3.
',Esuoso, K. O., Lutz, H., Bayer, E., Kutubuddin, M. (2000). Unsaponifiable lipid constituents of some underutilized tropical seed oil. J. Agric. Fd.Chem. 48(2): 231-234.
Evans, R. M., Currie, L., and Campbell, A. (1982). The distribution of ascorbic acid between various cellular components of blood, in normal individuals, and its relation to the plasma concentration. Br. J. Nature 47: 473-482.
Evans, R. W. (1989). Pharmacognosy (13'~ ed); Baillere Tindall, London. pp. 527-665.
Gey, K. F., Braubacher, G. B. and Stahelin, H. B. (1987). Plasma levels of antioxidant vitamins in relation to ischemic heart disease and cancer. Am. J. Clin. Nutri., 45: 1368-1 377.
Ghoghari, A. M., Bagul, M.S., Anandjiwala, S., Chauhan, M. G. and Rajani, M. (2006). Free radical scavenging activity of Aspidium cicutarium rhizome. J. Natural Remedies, 6(2): 1 31 -1 34.
Graham, R. K., Deng, Y., Slow, E. J., Haigh, B., Bissada, N., Lu, G., Pearson J., Shehadh, J., Bertram, L. and Murphy, Z. (2006). Cleavage at the caspase-6 site is required for neuronal dysfunction and degeneration due to mutant Huntingtin. Cell., 125: 1179-1191.
Grisham, M. B. and McCord, J. M. (1986). Chemistry and cytotoxicity of reactive oxygen metabolites. In: Physiology of Oxygen Radicals (Taylor, A. E., Matalon, S. and Ward, P., eds.). Am. Physiological Society. Bethesda, Md. Pp. 248-261.
Guilivi, C. and Davies, K. J. A. (1990). A novel antioxidant role for haemoglobin. The com-proportionation of ferrylhaemoglobin with oxyhaemoglobin. J. Biol. Chem. 265: 19453-1 9460.

Hall, A. (1 984). Oncogenes-implications for human cancer. A review. J. Res. Soc. Med., 77: 41 0.
Halliwal, H. (1 994). Lancet. 344: 721-724.
Halliwell, B., Gutteridge, J. M. C. and Holbrook, N. J. (1999). Free radical in Biology and Medicine, Oxford University Press. London, p. 625.
Irvine, F. R. (1961). Woody Plants of Ghana with special reference to their uses. Oxford University Press, London pp. 13-23.
Jain, A., Singhai, A. K. and Dixit, V. K. (2006). In vitro evaluation of Tephrosia purpurea pers for antioxidant activity. J. Natural remedies. 6(1): 162-1 64.
~enner, P. and Olanow, C. W. (1996). Oxidative stress and the pathogenesis of Parkinson's disease. Neurology. 47: 161-1 70.
Jovanovic, S. V., Steenken, S. and Simic, M. G. (1990). One electron reduction potentials of 5-lndoxyl radicals. A pulse radiolysis and laser photolysis study. J. Phys. Chem. 94: 3583-4588.
Kanner, P. H., Guyton, K. E., and Gorospe, M. (1999). Oxidative stress and the molecular biology of antioxidant defense (Scandolios, J. G. ed) Cold Spring Harbor Laboratory Press, New York, pp. 247- 272.
Khachik, F., Beecher, G. R., and Whittaker, N. F. (1986). Separation, Identification and quantification of the major Carotenoid and Chlorophyll Constituents in extracts of several green vegetables by liquid chromatography. J. Agr. Food Chem. 34: 603-616.
Klein, J. a. and Ackerman, S. L. (2003). Oxidative stress, cell cycle and neurodegeneration. J. Clin. Invest; 11 l(6): 785-793.
Krinsky, N. I. and Deneke, S. M. (1982). Interaction of Oxygen and Oxyradicals with Carotenoids. J. Natl. Cancer lnst. 69: 206-210.
Kumaran, A. and Karunakaran, J. (2006). Antioxidant activity of polyphenols from phyllantus debilis klein ex Willd. J. Natural Remedies. 6(2): 141 -146.
Leaf, a., Kang, J. X., Xiao, Y. F. and Billman, G. E. (2003). Clinical prevention of sudden cardiac death by n-3 polyunsaturated acids and mechanism of prevention of arrhythmias by n-3 fish oils. Circulation: 107: 2646-2652.

Lerman, L.S. (1 984). Sequence determined DNA separations. Ann. Rev. Biophys. Bioeng. 13: 399.
Levine, M.. and Morita, K. (1985). Ascorbic acid in endocrine systems. Vitamin. Horm:42
Liener, I. E. (1972). Toxic Constituents of Plant Food Stuff, Academic Press, London, pp. 143-1 55.
Marras, C., McDermott, M., Rochon, P., Tanner, C., Naglie, G., Rudolph, A., and Lang, A. (2005). Survival in Parkinson disease: thirteen- year follow-up of the DATATOP Cohort. Neurology, 64(1): 87-93.
Mathews-Roth, M. M. (1982). Antitumor activity of beta-carotene, Canthaxanthin and Phytoene. Oncology, 39: 33-37.
Melhorn, R. J. (1991). Ascorbate and dehydroascorbic acid mediated reduction of free radicals in the human erythrocylc. J. Biol. Chem. 266: 2724-273 1.
Michael, J. F. (2005). Dynamic dopamine modulation in the basal ganglia: A neurocomputational account of cognitive deficits in medicated and non-medicated Parkinsonism. J. Cognitive Neuroscience; 1 7: 5 1 -73.
. Mona, T. (2000). The Nigrostriatal Dopaminergic system as a preferential target of repeated exposures to combined paraqual and manels: implications for Parkinson's disease. J. Neuroscience 20(24): 9207-921 4.
Morel, C. 0 . (1997). Mechanisms of Fatty Acid Oxidation. http:llwww.atozofhealth.com. Downloaded on the 1 lth ~ u n e , 2005.
Mukherjee, K. 1. (1 989). Medical laboratory technology. Tata McGraw Hill Publishing Company Limited, New Delhi, 1" edition, pp. 1123- 1124.
Murray, P. E., Vaya, J. and Aviran, M. (1990). Nutritional Antioxidant: Mechanisms . of Action, Analyses of Activities and Modern Application. Bentham Science Publishers Ltd. New York, pp. 99- 110.
Nakamura, K., Bindokas, V. P., Marks, J. D., Wright, D. A., Frim, D. M., Miller, R. J. and Kang, U. J. (2000). The selective toxicity of 1- methyl-4-phenylpyridinium to dopaminergic neurons: The role of mitochondria1 complex I and reactive oxygen species revisted. Mol. Pharmacol. 58(2): 271 -278.

Neilson, M. (1965). Introduction to flowering plants of West Africa, University of London Press Ltd, London. pp. 90-105.
Niki, E. (1987). Interaction of ascorbate and a-toco-pherol. In: Third Conference on Vitamin C. (Burres, J. J., Rivers, J. M. and Machlin, L. J., eds). Ann. N. Y. Acad. Sci. 498: 186-199.
Niki, E. and Nakon, M. (1990). Estrogens as antioxidants. Methods Enzymol. 186: 330-333.
Nishikimi, M. (1975). Oxidation of Ascorbic acid with superoxide amion generated by the Xanthine-Xanthine Oxidase System. Biochem. Biophys. Res. Commun. 63: 463-468.
Oduyemi, 0 . (1993). Insecticidal properties of certain indigenous plant oils against Sitophlius zeamais mots. Appl. Entomol. and Phytopathol., 1 : 1 9-27.
Oguntimein, B., Ekundayo, O., Laasko, I. and Hitunen, R. (1989). Constituents of the essential oil of Monodora tenuifolia. Flav. and Fragr. J., 4: 193-1 95.
Okafor, J. C. (1989). Some useful Tropical Plans in Health Care Delivery, Forestry Commission HQ, Enugu. pp. 3-9.
Dmenn, G. S., Goodman, G. E., Thornquist, M. D., and Cullen, M. R. (1996). Free radicals and Antioxidants. Bio. Med., 334: 11 50- 11 55.
Osawa, T., Kavakishi, S. and Namiki, M. (1990). In: Kuroda, Shankal D. M., waters MD. (Eds). Antimutagenesis and anticarcinogenesis mechanisms II. New York, pp. 139.
Parkinson, J. (2002). As essay on the shaking palsy. J. Neuropsychiatry Clin. Neuroscience; l4(2): 223-236.
Perly, B., Smith, I.C.P. and Hughes, L. (1 985). Estimation of the Location of natural alpha-tocopherol in lipid bilayers by ' 3 ~ - ~ ~ ~ spectroscopy. Biochem. Biophys. Acta, 81 9: 131 -1 35.
Peto, R., Doll, R., Buckley, J. D. and Sporn, M. B. (1981). Can dietary beta-carotene materially reduce cancer rates? Nature, London. 290: 201-208.
Pratico, D. (2006). Link between autism and abnormal blood-vessel function, oxidative stress. http://www.med.upenn.edu/ Downloaded on the 2dth sept., 2006.

Ray, D. S. (2006). Oxidative stress. www.bionutrition.org. downloaded on the 24" Sept., 2006.
Ridley, R. M., Frith, C. D., Crow, T. J. and Conneally, P. M. (1988). Anticipation in Huntington's disease is inherited through the male line but may originate in the female. J. Medical Genetics. 25: 589- 595.
Robbelen, G., Downey, R. K. and Ashri, A. (1989). Oil Crops of the World, McGraw-Hill Publishing Company, New York, pp. 107-1 08.
Rose, R. C. and Bode, A. M. (1993). Biology of free radical scavengers: An evaluation of Ascorbate. FASEB J. 7: 11 35-1 141.
"
Sahart, M. H. (2001). Free radicals and Antioxidant. Biol. Med., 28: 1685-1 696.
Sawa, T., Kavakishi, S. and Namiki, M. (19900. In: Kuroda, Shankal D.M., Waters, M. D. (Eds.) Antimutagenesis and anticarcinogenesis mechanisms I!. New York; p. 139.
Sirna Therapeutics (2006). Huntington's disease overview. http://www.Sirna.com/wt/page/neurology. downloaded on the 24th Sept., 2006.
Stauth, D., Traber, M. and Mastaloudis, A. (2002). Marathon runners deplete vitamins, raise oxidative stress. Free Radical Biol. & Medicine. 54: 737-797.
Stocker, R., McDonagh, A F., Glazer, A. N. and Ames, B. N. (1990). Antioxidant activities of bile pigments: bilverdin and bilirubin. Methods Enzymol. 186: 301-309.
Stryer, L. (1988). Biochemistry 3rd ed. W. A. Freeman and Company, New York pp. 331 and 470.
Talalaji, S. J. (1 965). Essential oil from Monodora myristica grown in Ghana. West African Pharmacist, 4: 64-65.
Tretter, L. and Adam-Vizi, V. (2000). Generation of reactive oxygen species in the raction catalyzed by a-ketoglutarate dehydrogenase. http:www.neurosci.orcr/cai/contentlfu11/24/36/777l
Vessie, P. R. (1932). On the transmission of Huntington's chorea for 300 years - the Bures Family Group. J. Nen/ous and Mental Disease. 76: 553-573.

Wei, Y. N., and Lee, H. C. (2002). Oxidative Stress and Mitochondria1 DNA mutations in Human Aging. Proc. Soc. Exp. Biol. Med. 227: 671 -682.
Ziegler, R. G. (1989). A review of epidemiologic evidence that carotenoids reduce the risk of cancer. J. Nufr. 119: 116-122.


















