
UNIVERSIDADE ESTADUAL DE CAMPINAS FACULDADE DE...
84
UNIVER FAC J O CONSUMO DE FRU IMPLICAÇÕES PA PRODUÇÃO DE M LIGHT PHASE RE IMPLICATIONS ON H PRODU RSIDADE ESTADUAL DE CAMPINAS CULDADE DE CIÊNCIAS MÉDICAS JULIANA DE ALMEIDA FARIA UTOSE RESTRITO A FASE CLARA DO CIC ARA A ATIVAÇÃO DA AMPK HIPOTALÂ MELATONINA E O METABOLISMO GLIC ESTRICTED FRUCTOSE CONSUMPTION A HYPOTHALAMIC AMPK ACTIVATION, ME UCTION AND GLUCOSE METABOLISM CAMPINAS 2016 ICLO E SUAS ÂMICA, A CÍDICO AND ITS ELATONIN
Transcript of UNIVERSIDADE ESTADUAL DE CAMPINAS FACULDADE DE...

UNIVERSIDADE ESTADUAL DE CAMPINAS
FACULDADE DE CIÊNCIAS MÉDICAS
JULIANA DE ALMEIDA FARIA
O CONSUMO DE FRUTOSE RESTRITO A FASE CLARA DO CICLO E SUAS IMPLICAÇÕES PARA A ATIVAÇÃO DA AMPK HIPOTALÂMICA, A
PRODUÇÃO DE MELATONINA E O METABOLISMO
LIGHT PHASE RESTRICTED FRUCTOSE CONSUMPTION AND ITS IMPLICATIONS ON HYPOTHALAMIC AMPK ACTIVATION, MELATONIN
PRODUCTION AND GLUCOSE METABOLISM
UNIVERSIDADE ESTADUAL DE CAMPINAS
FACULDADE DE CIÊNCIAS MÉDICAS
JULIANA DE ALMEIDA FARIA
O CONSUMO DE FRUTOSE RESTRITO A FASE CLARA DO CICLO E SUAS IMPLICAÇÕES PARA A ATIVAÇÃO DA AMPK HIPOTALÂMICA, A
PRODUÇÃO DE MELATONINA E O METABOLISMO GLICÍDICO
LIGHT PHASE RESTRICTED FRUCTOSE CONSUMPTION AND ITS IMPLICATIONS ON HYPOTHALAMIC AMPK ACTIVATION, MELATONIN
PRODUCTION AND GLUCOSE METABOLISM
CAMPINAS
2016
O CONSUMO DE FRUTOSE RESTRITO A FASE CLARA DO CICLO E SUAS IMPLICAÇÕES PARA A ATIVAÇÃO DA AMPK HIPOTALÂMICA, A
GLICÍDICO
LIGHT PHASE RESTRICTED FRUCTOSE CONSUMPTION AND ITS IMPLICATIONS ON HYPOTHALAMIC AMPK ACTIVATION, MELATONIN

JULIANA DE ALMEIDA FARIA
O CONSUMO DE FRUTOSE RESTRITO A FASE CLARA DO CICLO E SUAS IMPLICAÇÕES PARA A ATIVAÇÃO DA AMPK HIPOTALÂMICA, A
PRODUÇÃO DE MELATONINA E O METABOLISMO GLICÍDICO
LIGHT PHASE RESTRICTED FRUCTOSE CONSUMPTION AND ITS IMPLICATIONS ON HYPOTHALAMIC AMPK ACTIVATION, MELATONIN
PRODUCTION AND GLUCOSE METABOLISM
Tese apresentada à Faculdade de Ciências Médicas da Universidade Estadual de Campinas como parte dos requisitos exigidos para a obtenção do título de Doutora em Farmacologia.
Doctoral Thesis presented to the Faculty of Medical Sciences, State University of Campinas - UNICAMP to obtain the title of Doctor of Pharmacology.
Orientador: Professor Dr. Gabriel Forato Anhê Tutor: Professor Dr. Gabriel Forato Anhê ESTE EXEMPLAR CORRESPONDE À VERSÃO FINAL DA TESE DEFENDIDA PELA ALUNA JULIANA DE ALMEIDA FARIA E ORIENTADA PELO PROF. DR. GABRIEL FORATO ANHÊ.
CAMPINAS
2016

Agência(s) de fomento e nº(s) de processo(s): FAPESP, 2012/11409-4
Ficha catalográfica
Universidade Estadual de Campinas
Biblioteca da Faculdade de Ciências Médicas
Maristella Soares dos Santos - CRB 8/8402
Faria, Juliana de Almeida, 1986-
F225c FarO consumo de frutose restrito a fase clara do ciclo e suas implicações para
a ativação da AMPK hipotalâmica, a produção de melatonina e o metabolismo
glicídico / Juliana de Almeida Faria. – Campinas, SP : [s.n.], 2016.
FarOrientador: Gabriel Forato Anhê.
FarTese (doutorado) – Universidade Estadual de Campinas, Faculdade de
Ciências Médicas.
Far1. Frutose. 2. Ingestão fora de fase. 3. Hipotálamo. 4. Proteína quinase
ativada por AMP. 5. Corticosterona. 6. Melatonina. I. Anhê, Gabriel
Forato,1980-. II. Universidade Estadual de Campinas. Faculdade de Ciências
Médicas. III. Título.
Informações para Biblioteca Digital
Título em outro idioma: Light phase restricted fructose consumption and its implications on
hypothalamic AMPK activation, melatonin production and glucose metabolism
Palavras-chave em inglês:Fructose
Out-of-phase feeding
Hypothalamus
AMP-Activated protein kinase
Corticosterone
Melatonin
Área de concentração: Farmacologia
Titulação: Doutora em Farmacologia
Banca examinadora:Gabriel Forato Anhê [Orientador]
Andrea Maculano Esteves
Gabriela Freitas Pereira de Souza
Thomas Prates Ong
José Donato Júnior
Data de defesa: 29-07-2016
Programa de Pós-Graduação: Farmacologia

BANCA EXAMINADORA DA DEFESA DE DOUTORADO
JULIANA DE ALMEIDA FARIA
ORIENTADOR: PROF. DR. GABRIEL FORATO ANHÊ
MEMBROS:
1. PROF. DR. GABRIEL FORATO ANHÊ
2. PROF. DRa. ANDREA MACULANO ESTEVES
3. PROF. DRa. GABRIELA FREITAS PEREIRA DE SOUZA
4. PROF. DR. THOMAS PRATES ONG
5. PROF. DR. JOSÉ DONATO JÚNIOR
Programa de Pós-Graduação em Farmacologia da Faculdade de Ciências Médicas da Universidade Estadual de Campinas.
A ata de defesa com as respectivas assinaturas dos membros da banca examinadora encontra-se no processo de vida acadêmica do aluno.
Data: 29/07/2016

"I do not know what I may appear to the world,
but to myself I seem to have been only like a boy
playing on the sea-shore, and diverting myself in now and
then finding a smoother pebble or a prettier shell than ordinary,
whilst the great ocean of truth lay all undiscovered before me."
Sir Isaac Newton

AGRADECIMENTOS
A Deus, minha “...humilde admiração do espírito superior e ilimitado que se revela nos menores
detalhes que podemos perceber em nossos espíritos frágeis e incertos... Essa convicção,
profundamente emocional na presença de um poder racionalmente superior, que se revela no
incompreensível universo...”.
Aos meus pais, Mauro e Marilza, pelo amor. Pela persistência e dedicação em me proporcionar
o que não tiveram. Pela educação e pelos valores transmitidos, baseados no exemplo e não
simplesmente em palavras vãs.
Ao meu irmão e aos meus avós, tios e primos pelo carinho, pela torcida e confiança no caminho
escolhido pra seguir.
Ao professor Gabriel Anhê, pela convivência diária, pela valiosa e consistente orientação, pela
inquestionável reverência e estima a ciência, pelas diversas oportunidades proporcionadas e pela
confiança em me integrar ao seu “time científico”. Agradeço pelo tempo dedicado a minha
formação profissional e intelectual. Serei eternamente grata.
Aos amigos, Letícia M. Ignácio de Souza e Thiago M. Ferreira de Araújo, presenças constantes
dentro e fora do laboratório. Pela sinceridade. Por desafiarem minhas convicções e permitirem o
questionamento e ampliação do meu limitado universo de ideias e teorias. Com vocês, entendi
que a amizade pode (e deve) ser catalisadora de transformações.
Ao Lucas pelo amor e companheirismo. Pela paciência. Por acreditar em mim mais do que eu
mesma. Por dividir (e somar) os planos e os sonhos dessa breve vida. Por achar graça dos meus
devaneios. Por trazer calma onde a ansiedade e a agitação predominam.
Ao querido Senhor Miguel pelo empenho no cuidado dos animais experimentais. Pelo seu
fantástico senso de humor.
Agradeço aos colegas e amigos do Laboratório de Farmacologia e Fisiopatologia do Diabetes
Mellitus e do Laboratório de Sinalização Celular pelas discussões sobre ciência e sobre a vida
(quantas vezes elas se confundem).

Pelo universo desconhecido que nos desafia constantemente. Pela curiosidade, alavanca das
descobertas e propulsora da inovação. Pelos que, acometidos pela doença, necessitam de
esperança em terapias mais eficientes.
Agradeço ainda a Thiago M. Ferreira de Araújo, Letícia M. Ignácio de Souza, Daniela Razolli,
Dailson Souza e ao professor Gabriel Anhê, os quais se dedicaram diretamente a realização deste
trabalho. Por questionarem sempre. Pelos experimentos diurnos e/ou noturnos (os ZT’s não serão
esquecidos). Pela paciência quando o meu cansaço já substituía o entusiasmo.
À Universidade Federal dos Vales do Jequitinhonha e Mucuri por minha formação básica.
Exemplo de que, mesmo diante de tantas limitações, é possível capacitar seus graduandos.
À Universidade Estadual de Campinas pela inspiração e orgulho. Nem nos planos mais otimistas,
me imaginei “filha” dessa grandiosa Instituição.
À Universidade de Cambridge, ao Instituto de Ciências Metabólicas e a professora Susan Ozanne
pela calorosa acolhida durante parte do meu Doutorado. Por permitir que as “janelas” do
conhecimento fossem ampliadas.
Aos animais experimentais, sem os quais seria impossível desenhar e desenvolver esse projeto.
Pela vida doada à ciência.
À Fundação de Amparo à Pesquisa do Estado de São Paulo, pelo auxílio financeiro. Pela
concessão da bolsa de estudo no país e da bolsa de estágio em pesquisa no exterior.

RESUMO
DE ALMEIDA FARIA, J. O consumo de frutose restrito a fase clara do ciclo e suas
implicações para a ativação da AMPK hipotalâmica, a produção de melatonina e o
metabolismo glicídico. 2016. 84 f. Tese (Doutorado) – Faculdade de Ciências Médicas,
Universidade Estadual de Campinas, São Paulo, 2016.
O consumo de dietas ricas em frutose exerce uma conhecida modulação a qual se caracteriza por
resistência periférica à ação da insulina, aumento de gliconeogênese e esteatose hepáticas sobre o
metabolismo glicêmico. Os mecanismos exatos pelos quais a frutose realiza estas ações estão em
contínua elucidação, no entanto já se sabe que tanto o aumento da gliconeogênese quanto o
estímulo à hiperfagia resultam da ativação da proteína AMPK no hipotálamo. Dentre as diversas
modulações endócrinas que a frutose exerce, foi recentemente descrita a redução na produção
noturna de melatonina. A melatonina, um hormônio produzido de maneira circadiana pela
glândula pineal, tem a capacidade de sincronizar o metabolismo energético aumentando a
sensibilidade à insulina e reduzindo a gliconeogênese. Assim como a produção de melatonina,
sabe-se que a atividade hipotalâmica da AMPK apresenta um ritmo circadiano com valores
aumentados no período noturno. Foi demonstrado recentemente que o aumento da atividade
hipotalâmica da AMPK com ativadores farmacológicos reduz a produção noturna de melatonina.
Interessantemente, a frutose ingerida exclusivamente durante a fase clara do ciclo claro/escuro
resulta em modificações metabólicas similares às descritas acima, enquanto que a frutose
ingerida durante a fase escura parece exercer efeitos metabólicos mais brandos. O presente
estudo traz os resultados oriundos da investigação das alterações metabólicas resultantes da
ingestão de frutose em diferentes fases do ciclo claro/escuro; e nesse contexto, avalia se tais
alterações mostram-se dependentes de modificações nos padrões de atividade da AMPK no
hipotálamo e da produção de corticosterona.
Palavras-chave: Frutose, Ingestão fora de fase, Hipotálamo, AMPK, Corticosterona,
Melatonina.

ABSTRACT
DE ALMEIDA FARIA, J. The light phase restricted fructose consumption and its
implications on hypothalamic AMPK activation, melatonin production and glucose
metabolism. 2016. 84 f. Thesis (Doctoral) – Faculty of Medical Sciences, University of
Campinas, Sao Paulo, 2016.
A high fructose diet exerts a known modulation which is characterized by peripheral resistance
to the insulin action, increased gluconeogenesis and hepatic steatosis on glucose metabolism.
The exact mechanisms by which fructose triggers such actions are in continuous elucidation
however it is known that the increased gluconeogenesis and the hyperphagia stimulation both
result from activation of hypothalamic AMPK protein. Among several endocrine modulations,
fructose promotes a reduction on nocturnal melatonin production. Melatonin, a hormone
produced by pineal gland in a circadian manner, has the ability to synchronize the energetic
metabolism by increasing insulin sensitivity and reducing gluconeogenesis. As melatonin
production, it is known that hypothalamic AMPK activity has a circadian rhythm with increased
values during the night phase. It has been shown recently that increased hypothalamic AMPK
activity through pharmacologic activators is able to reduce nocturnal melatonin production.
Interestingly, fructose consumption only during the light phase of the light / dark cycle results in
metabolic changes similar to those described above, while fructose intake exclusively during the
dark phase appears to exert milder metabolic effects. This study presents the results arising from
the research of metabolic changes resulting from fructose intake at different phases of light / dark
cycle; and in this context, we have assessed whether such changes are dependent on
hypothalamic AMPK activity patterns and corticosterone production.
Key words: Fructose, Out-of-phase feeding, Hypothalamus, AMPK, Corticosterone, Melatonin.

LISTA DE ABREVIATURAS E SIGLAS
AMP
AMPK
Adenosina monofosfato
Proteína quinase ativada por AMP
ANOVA
AUC
Análise de variância
Area under curve
ATP Trifosfato de adenosina
CRH Hormônio liberador de corticotrofina
DMSO Dimetilsulfóxido
DTT Ditiotreitol
EDTA Ácido etileno diamino tetracético
ELISA
EPM
FoxO1
G-6-Pase
Ensaio imunoenzimático
Erro padrão da média
Forkhead box protein O1
Glicose-6-fosfatase
GLUT4 Transportador de glicose tipo 4
GTT
HFCS
HPA
icv
Teste de tolerância à glicose
High fructose corn syrup
Hipotálamo-pituitária-adrenal
Intracerebrovenricular
ITT
NaCl
NES
Teste de tolerância à insulina
Cloreto de sódio
Night eating syndrome

PEPCK
PHG
PTT
Fosfoenolpiruvato carboxiquinase
Produção hepática de glicose
Teste de tolerância ao piruvato
SDS-PAGE Eletroforese em gel de poliacrilamida com dodecil sulfato de sódio
Tris
ZT
Tri(hidroximetil)-aminometano
Zeitgeber time

SUMÁRIO
INTRODUÇÃO
14
OBJETIVOS 22
Objetivo Geral 22
Objetivos Específicos
METODOLOGIA
22
23
CAPÍTULO 1 – Artigo referente à tese 29
Abstract 30
Introduction 31
Methods 32
Results 36
Discussion 42
References 47
Figure Legends
Figures
DISCUSSÃO
CONCLUSÃO
53
56
60
65

CAPÍTULO 2 – Co-autoria/autoria de artigos publicados na área durante o período 66
REFERÊNCIAS 68
APÊNDICE 79
ANEXOS 84

14
INTRODUÇÃO
O açúcar e seu consumo através dos diferentes momentos históricos
Vários registros históricos demonstram que o consumo de açúcar, tal como
conhecemos, teve início apenas no último século. De forma breve, destaco aqui os
principais momentos históricos que permitiram o gradativo aumento do consumo de
açúcar até os padrões atuais.
Durante o período Paleolítico (conhecido ainda como Idade da Pedra Lascada),
as proteínas constituíam a base alimentar do Homem e eram oriundas de atividades
como pesca e caça. Os carboidratos eram obtidos exclusivamente de vegetais
encontrados casualmente e coletados da natureza, uma vez que a agricultura ainda não
havia sido desenvolvida. Nesse período, portanto, os açúcares, presentes naturalmente e
em quantidades modestas, eram consumidos de forma totalmente dependente da estação
do ano principalmente em países localizados em média e alta latitudes onde as estações
são marcadamente definidas.
No período Neolítico ou Idade da Pedra Polida, indivíduos, em diferentes
posições geográficas e de forma independente, perceberam que os grãos coletados
poderiam ser enterrados e, após algum tempo, dariam origem a novas plantas e,
consequentemente, a novos frutos. Nesse momento, entende-se que é possível cultivar
espécies vegetais e, portanto, controlar, pelo menos em parte, a disponibilidade de
alimentos.
Acredita-se que a escassez e a palatabilidade do açúcar foram responsáveis pelo
interesse recorrente de diversos povos em desenvolver técnicas que ampliassem a
obtenção de tal nutriente (Tappy & Lê, 2010).

15
Nessa linha cronológica, os próximos eventos históricos permitiram o contato de
países localizados na Europa com civilizações fixadas no Oriente Médio e em regiões
que hoje pertencem à Índia e à China. As Cruzadas e as Grandes Navegações levaram
ao estreitamento das relações comerciais entre o continente europeu e o asiático
viabilizando a substituição do mel, até então o principal agente edulcorante usado na
Europa, pelo açúcar extraído da cana e de outros vegetais. O açúcar oriundo de países
orientais foi incorporado rapidamente à cultura alimentar europeia e, mais tarde, seu
consumo, na forma de sacarose, foi difundido para os outros continentes.
Porém, a Revolução Industrial constituiu o período no qual aconteceram as mais
profundas transformações nos padrões alimentares. Durante esse período, o açúcar
passou a ser o principal componente da dieta ocidental (Licht, 2005). O
desenvolvimento e aperfeiçoamento de métodos de extração pela indústria alimentícia
permitiram maior eficiência na obtenção de frutose através do processo de isomerização
enzimática (Marshall, 1957). Com isso, na América do Norte, o xarope de milho rico
em frutose (originalmente nomeado como high fructose corn syrup ou simplesmente
compilado na sigla HFCS) passou a ser largamente produzido (White, 2008). O alto
poder edulcorante, a facilidade na extração e o baixo custo de produção foram fatores
que favoreceram a implementação e expansão do consumo da frutose à época e
explicam o interesse na manutenção desse monossacarídeo como um dos principais
componentes de produtos industrializados até os dias atuais (Tappy & Lê, 2010).
Efeitos metabólicos da frutose e a dependência do horário de ingestão
Dentre os diversos fatores ambientais reconhecidos como determinantes para o
ganho de peso e o estabelecimento de quadros que favorecem a resistência à insulina, a
composição da dieta é aquele que recebe maior atenção da comunidade científica.

16
Estudos realizados em humanos e em roedores mostram que, de maneira geral, a
ingestão de dietas ricas em açúcares e/ou em gorduras saturadas tem capacidade de
induzir resistência à insulina e aumento de adiposidade (Lê et al., 2009; Ragheb et al.,
2008; Rajasekar et al., 2007).
Antes de 1950, a frutose era encontrada em baixas quantidades na dieta humana,
a qual dispunha de alimentos que apresentavam naturalmente tal monossacarídeo, entre
os quais, cereais, vegetais e mel. O interesse em implementar e difundir o uso da frutose
residiu, em um primeiro momento, no fato de que esse nutriente apresenta poder
edulcorante maior que a glicose pura. Assim sendo, a frutose passou a ser
comercializada e ingerida na forma de sacarose (dissacarídeo frutose/glicose) ou ainda,
e principalmente, em sua forma pura, presente no xarope de milho rico em frutose; a
forma pura tem sido amplamente utilizada na dieta norte-americana até os dias atuais
conforme já mencionado anteriormente (Tappy & Lê, 2010). Porém, a rápida inserção e
difusão do consumo deste nutriente no ocidente e sua associação com o aumento no
número de casos de obesidade fizeram com que a frutose se tornasse alvo de vários
estudos buscando sua relação com o aumento de adiposidade corporal.
Foi demonstrado que a metabolização da frutose não depende da ação da
insulina. Além disso, após o consumo desse monossacarídeo, ocorrem flutuações
glicêmicas discretas, ao contrário do que ocorre após a ingestão de glicose. Por essas
razões, a frutose foi, em um primeiro momento, utilizada como alternativa à sacarose e
à glicose por pacientes diabéticos (Havel, 2005). No entanto, diversos estudos em
roedores mostraram uma consistente relação entre consumo crônico de frutose e
resistência à insulina em órgãos metabolicamente importantes, obesidade, Diabetes
Mellitus tipo II e hipertensão arterial (revisado por Tappy & Lê, 2010). Apesar de ainda
não determinados com exatidão, muitos mecanismos foram propostos para explicar o

17
possível efeito pró-resistência à insulina causada pela frutose. As primeiras abordagens
mostraram que o metabolismo da frutose pelo fígado leva a formação de fosfo-triose a
qual pode, à jusante da via, ser convertida à glicose. Desta maneira, mais de 50% da
frutose absorvida pelo trato gastrointestinal é convertida à glicose pelo fígado,
contribuindo para o aumento da glicemia de jejum (Bode et al., 1981).
Além disso, o próprio metabolismo hepático da frutose predispõe a síntese de
ácidos graxos e acúmulo de gordura em hepatócitos. Diversos relatos experimentais
descrevem um quadro de esteatose hepática em animais submetidos a dietas ricas em
frutose por um longo período de tempo (Mayes, 1993; Ragheb et al., 2008). Acredita-se
que o acúmulo de gordura no fígado leva um aumento da expressão de citocinas pró-
inflamatórias pelo hepatócito que, por sua vez, diminui a transmissão do sinal
intracelular da insulina. A resistência hepática a ação da insulina é marcada por um
drástico aumento da gliconeogênese e aumento da glicemia de jejum (Samuel, 2011).
Além dos eventos acima mencionados, a oxidação da frutose pela
cetohexocinase gera uma baixa quantidade de ATP e leva a uma diminuição da razão
ATP/AMP intracelular, facilitando deste modo a ativação da proteína kinase dependente
de AMP (AMPK). Recentemente, experimentos de nosso grupo mostraram que a
frutose pode aumentar a gliconeogênese em decorrência de ações extra-hepáticas.
Nossos estudos mostram que a frutose tem a capacidade de agir diretamente no
hipotálamo e estimular a ativação da AMPK. A ativação hipotalâmica da AMPK resulta
em disparo de uma resposta contra-reguladora da ação da insulina marcada por aumento
da produção de corticosterona, um clássico estimulador da gliconeogênese (Kinote et al,
2012). A ativação hipotalâmica da AMPK pela frutose também apresenta outras
relevâncias, entre as quais, a estimulação direta da ingestão alimentar (Cha et al., 2008).

18
Um estudo recente mostra que também o horário da ingestão de frutose parece
determinar a intensidade de seus efeitos metabólicos descritos acima. Ratos expostos à
frutose durante a fase clara do ciclo claro/escuro apresentaram um aumento de
adiposidade e resistência à insulina ao passo que estas adaptações metabólicas não
foram observadas em animais expostos à frutose durante a fase escura do ciclo
claro/escuro (Morris et al., 2012). Apesar deste estudo não descrever o mecanismo que
explica o efeito horário-dependente da frutose, pode se levar em conta uma possível
interferência no ritmo circadiano da atividade hipotalâmica da AMPK. Neste sentido, já
está relativamente bem estabelecido que as flutuações nas atividades da AMPK no
hipotálamo apresentam valores máximos durante a noite e mínimos durante o dia em
roedores (Um et al., 2011).
Dependendo de sua intensidade e frequência, a ingestão de alimentos fora de
fase por seres humanos pode ser classificada como a night eating syndrome (NES)
(Stunkard et al., 1955), traduzida livremente como síndrome da alimentação noturna. A
prevalência de NES é relativamente baixa na população em geral, mas varia entre 8,9 e
27% em subgrupos obesos (Rand et al., 1957). Os estudos de coorte mostraram que a
NES está positivamente correlacionada ao diagnóstico de síndrome metabólica,
aumento de triglicerídios e da circunferência da cintura (Gallant et al., 2014). Entre as
várias adaptações, o perfil endócrino circadiano dos pacientes com NES é caracterizado
por níveis de melatonina reduzidos durante a noite e aumento das concentrações de
cortisol pela manhã (Birketvedt et al., 1999).
Inter-relações entre frutose, melatonina e adaptações metabólicas
A exposição à frutose tem como resultado, além das alterações metabólicas
descritas acima, uma complexa modulação em várias secreções endócrinas. Por

19
exemplo, em humanos a exposição à frutose resulta em diminuição nas concentrações
dos hormônios leptina e grelina (Teff et al., 2004). Em ratos, a frutose promove uma
redução do pico noturno da produção de melatonina pela glândula pineal (Leibowitz et
al., 2008) e aumento da produção de corticosterona (Kinote et al., 2012). A redução da
secreção de melatonina, por sua vez, parece não ser uma simples coincidência posto que
a reposição da melatonina em animais que recebem frutose resulta numa reversão de
alguns distúrbios metabólicos causados por este monossacarídeo (Kitagawa et al.,
2012).
A descrição das ações reguladoras do metabolismo desencadeadas pela
melatonina data, porém, do início da década de 90. Em trabalho publicado em 1994
(Lima et al., 1994), foram relatados os primeiros resultados de experimentos com
adipócitos isolados e incubados na presença de melatonina. Este trabalho mostrou que a
incubação de adipócitos com melatonina, aumenta a captação de glicose estimulada pela
insulina. Em continuidade, outros estudos do mesmo grupo demonstraram que a
remoção da glândula pineal (pinealectomia) gera um quadro de intolerância à glicose e
resistência à insulina (Lima et al, 1998).
Em seguida, foi demonstrado que ratos submetidos a uma remoção da glândula
produtora de melatonina (pineal) apresentavam resistência à insulina e diminuição da
expressão do GLUT4 no tecido adiposo e na musculatura esquelética (Zanquetta et al.,
2003). É importante ressaltar que, ao contrário de outros parâmetros metabólicos, a
captação de glicose por adipócitos em animais sem a glândula pineal está reduzida em
diferentes momentos da fase do ciclo claro/escuro (Alonso-Vale et al., 2004). Mais
interessante, foi a constatação de que a incubação intermitente in vitro de adipócitos
com melatonina (12h de presença e 12h de ausência de melatonina in vitro) é um
potencializador da insulina mais eficiente que a exposição continua à melatonina (24h)

20
(Alonso-Vale et al., 2006). Após estes achados, verificou-se que a melatonina leva ao
aumento também da captação de glicose em células musculares in vitro em paralelo a
um aumento da sinalização da insulina (Ha et al., 2006) e que a administração de
melatonina in vivo para animais obesos atenua a resistência à insulina (Sartori et al.,
2009).
De maneira geral, todos estes achados sugerem que a ritmicidade de produção da
melatonina favorece a sensibilidade à insulina de maneira sincronizada com a
capacidade secretora do pâncreas endócrino, de modo a evitar uma intolerância à
glicose. A insulina secretada, em situações fisiológicas, exerce uma potente ação sobre
o fígado que resulta na supressão da produção hepática de glicose (PHG). O mecanismo
proposto para esta ação envolve a inibição do fator de transcrição FoxO1 (Forkhead
box-Other 1), proteína que quando ativa, estimula a expressão das enzimas
Fosfoenolpiruvato-Carboxicinase (PEPCK) e glicose-6-fosfatase (G-6-Pase) (Zhang et
al., 2006). O mecanismo pelo qual a insulina inibe a atividade transcricional do FoxO1
envolve sua fosforilação em resíduos de serina e treonina pela AKT (Brunet et al.,
1999). Nosso grupo publicou recentemente que animais sem a glândula pineal
apresentam, no final do período noturno, aumento da expressão do fator de transcrição
FoxO1 e da proteína PEPCK no território hepático; classicamente, a PEPCK é descrita
como uma enzima chave da gliconeogênese por converter oxalacetato em
fosfoenolpiruvato (Nogueira et al., 2011).
No que diz respeito ao mecanismo pelo qual a ritmicidade da produção de
melatonina leva a melhora da homeostasia glicêmica, a literatura atual aponta duas
tendências não excludentes. A melatonina diretamente pode ativar a via de sinalização
da insulina em diversos tecidos (Anhê et al., 2004; Ha et al., 2006) e assim estimular,
por exemplo, a captação de glicose por adipócitos e fibras musculares (Alonso-Vale et

21
al., 2004; Ha et al., 2006). Outro possível mecanismo pelo qual a melatonina facilitaria
a ação da insulina seria secundário à supressão da secreção de cortisol ou corticosterona.
Uma série de estudos já demonstrou que a melatonina consegue agir através de
receptores específicos na adrenal de primatas e de roedores resultando na supressão da
produção de glicocorticóides (Torres-Farfan et al., 2004; Torres-Farfan et al., 2011).

22
OBJETIVOS
Objetivo geral
Investigar o papel do consumo de frutose em diferentes fases do ciclo claro/escuro sobre
a modulação do metabolismo energético.
Objetivos específicos
• Investigar o efeito do consumo de frutose sobre a ingestão alimentar e ganho de
peso para os diferentes grupos experimentais.
• Investigar possíveis alterações na sensibilidade sistêmica à insulina, tolerância à
glicose e gliconeogênese.
• Investigar possíveis alterações na ativação da proteína AMPK no tecido
hipotalâmico resultantes do consumo de frutose em diferentes fases do ciclo
claro/escuro.
• Avaliar se a inibição farmacológica da proteína AMPK no sistema nervoso
central pode interferir nas ações metabólicas da frutose oferecida durante a fase clara do
ciclo claro/escuro.
• Avaliar a produção de melatonina em animais expostos à frutose durante a fase
clara do ciclo claro/escuro.
• Avaliar se a reposição de melatonina exclusivamente durante a fase escura
suprime os prejuízos metabólicos da frutose ingerida exclusivamente durante a fase
clara do ciclo claro/escuro.

23
METODOLOGIA
Animais e tratamentos
Os procedimentos experimentais foram aprovados pelo Comitê de Ética em
Experimentação Animal da Universidade Estadual de Campinas (protocolo n ° 2798-1)
e foram conduzidos de acordo com as orientações do Colégio Brasileiro de
Experimentação Animal (COBEA).
Ratos machos da linhagem Sprague-Dawley apresentando três semanas de idade
foram fornecidos pelo Centro de Melhoramento Animal da Universidade Estadual de
Campinas (CEMIB, Campinas, São Paulo, Brasil) e foram mantidos sob temperatura
(22 ± 2 ° C) e fotoperíodo de 12 horas. Os animais tiveram livre acesso a ração padrão e
água durante 5 semanas. Na oitava semana de vida, os animais foram atribuídos aos
respectivos grupos experimentais e tratados durante as próximas 8 semanas com frutose
e/ou melatonina. As medidas de ingestão alimentar e de massa corporal foram feitas
duas e uma vez, por semana, respectivamente, durante todo o período de tratamento.
Quando especificado na seção de resultados (Capítulo 1), duas estratégias experimentais
foram combinadas ao tratamento com frutose durante a fase clara do ciclo: (i) a
disponibilidade de ração padrão foi restrita à fase escura (grupo Chow-R) durante as 8
semanas de tratamento, ou (ii) O Composto C foi administrado durante a última semana
de tratamento através de uma cânula implantada no ventrículo lateral.
A frutose foi dissolvida em água potável para produzir uma solução estoque
70% (m/v). A solução estoque de frutose foi posteriormente diluída a 10% (m/v)
novamente com água potável imediatamente antes do tratamento. A frutose foi
disponibilizada aos animais exclusivamente durante a fase clara ou durante a fase escura

24
(representados pelas siglas LPF e DPF, respectivamente). Garrafas contendo água foram
oferecidas aos ratos apenas quando frutose estava ausente.
A melatonina (Cat. A9525, Sigma-Aldrich, St. Louis, MO, EUA) foi
inicialmente diluída em etanol absoluto para produzir uma solução estoque de
concentração 100 mg/ml a qual foi mantida ao abrigo da luz em freezer -20 ° C durante,
no máximo, os próximos 7 dias. A solução estoque de melatonina foi posteriormente
diluída (1:50) com água destilada a cada 3 dias para gerar uma solução de concentração
2 mg/ml. Volumes variáveis (levando em consideração a massa corporal do respectivo
animal) desta última solução de melatonina foram adicionados à garrafa de água
potável, para se obter uma solução a 0,5 mg de melatonina/kg de animal. A garrafa
contendo solução de melatonina foi colocada na respectiva gaiola 30 minutos antes do
apagar das luzes e removida 30 minutos após a o acender. Os cálculos individuais para
os volumes necessários de solução de melatonina foram baseados na massa corporal
(avaliada semanalmente) e na ingestão noturna de água (avaliada diariamente). Tais
cálculos foram ajustados diariamente.
Procedimento cirúrgico e tratamento intracerebroventricular
Sete semanas após o início do tratamento com frutose, os ratos foram
anestesiados através da administração de uma mistura de diazepam e cetamina (2 e 50
mg/kg, respectivamente) e posicionados em aparelho estereotáxico para inserir uma
cânula de aço inoxidável no ventrículo lateral do hipotálamo. As coordenadas
estereotáxicas foram de 0,8 mm (anteroposterior), 1,5 mm (lateral), e 4,0 mm
(profundidade) (Lin et al, 2013). A localização da cânula foi testada avaliando-se a
resposta dipsogênica individual após a injeção intracerebroventricular (icv) de

25
angiotensina II (5 ng/uL de solução salina; Sigma, St Louis, MO, EUA) uma semana
após o procedimento cirúrgico (7 semanas de tratamento com frutose). Apenas os
animais que apresentaram uma resposta positiva neste teste foram utilizados nos
próximos experimentos. Para a injeção icv foi utilizada seringa Hamilton (50 µL)
acoplada, por meio de uma cânula plástica a uma agulha 30G, de modo que esta
ultrapasse o comprimento da cânula metálica (0,1 – 0,2 mm) garantindo a entrada do
líquido no ventrículo lateral.
O Composto C (catálogo n ° 171260; EMD4 Biosciences, Gibbstown, NJ, EUA)
foi diluído em 5% de dimetilsulfóxido (DMSO) a uma concentração final de 200 mM e
foi injetado diariamente através da cânula, durante os cinco dias finais da última semana
de tratamento com frutose. As injeções (2 ul) foram realizadas uma hora após o
acendimento das luzes (ZT 1) do biotério e volumes iguais de veículo foram injetados
nos animais do grupo controle. Os experimentos foram realizados duas horas antes de
as luzes serem apagadas no quinto dia de injeção icv utilizando composto C.
Previamente às análises, os animais foram submetidos ao jejum (imediatamente após a
última injeção icv). As amostras foram coletadas duas horas antes do apagar das luzes
(ZT 10) do quinto dia de injeção icv de composto C.
Produção de glicose após desafio intraperitoneal com piruvato
O teste de tolerância intraperitoneal ao piruvato (PTT) foi realizado com a
finalidade de estimar a taxa de gliconeogênese em cada animal submetido a um
diferente tratamento (Yao & Nyomba, 2008). Os ratos receberam uma injeção
intraperitoneal de piruvato (2 g/kg) dissolvido em uma solução de NaCl 0,9% (m/v). A
glicemia foi medida anteriormente a injeção de piruvato e, após a mesma, nos tempos

26
15, 30, 60, 90 e 120 minutos a partir do sangue total colhido de uma pequena incisão na
cauda do animal.
Teste de tolerância à glicose
Para se avaliar a tolerância à glicose, os animais foram submetidos ao teste de
tolerância à glicose (GTT). Neste teste os animais receberam uma injeção
intraperitoneal de glicose (2g/kg) e a glicemia foi medida antes e 5, 10, 15, 30, 60 e 120
minutos após a injeção. A área sob a curva (glicemia vs. tempo) foi calculada como
parâmetro de tolerância à glicose.
Teste de tolerância à insulina
Os animais foram submetidos ao teste de tolerância à insulina (ITT) para se
avaliar a sensibilidade sistêmica à insulina. Neste teste os animais receberam uma
injeção intraperitoneal de insulina (2 UI/kg) e a glicemia foi aferida antes e 5, 10, 15,
20, 25 e 30 minutos após a injeção. Os valores da glicemia foram convertidos para
escala logarítimica na base e. Em seguida a inclinação da reta (resultante do logarítimo
da glicemia vs. tempo) foi calculada. Assumiu-se este valor em módulo multiplicado
por 100 como a constante de decaimento de glicose após o ITT (KITT).
Extração de proteínas e immunoblotting
Os animais anestesiados foram decapitados e o hipotálamo foi removido. O
hipotálamo foi em seguida homogeneizado com um Polytron (Kinematica. Suíça) em
0.8 mL de tampão de extração (SDS 1%, Tris (pH 7,4) 100mM, pirofosfato de sódio
100mM, fluoreto de sódio 100mM, EDTA 10mM, ortovanadato de sódio 100mM) e
incubadas à 96ºC por 10 min. Em seguida, as amostras foram centrifugadas para a

27
remoção do material insolúvel. Após centrifugação, parte do sobrenadante das amostras
foi utilizado para determinação do conteúdo protéico por espectrofotometria com
reagente Bradford (Biorad, CA, USA) e o restante foi acrescido de tampão Laemmli 5X
com DTT e incubado à 96ºC em banho-maria por 10 min. A mesma quantidade de
proteínas totais de cada amostra tratada com Laemmli foi fracionada em SDS-PAGE em
aparelho para mini-gel (Mini-Protean, Bio-Rad). Após separação eletroforética, as
proteínas foram transferidas para uma membrana de nitrocelulose (Bio-Rad, CA, USA).
As extrações ocorreram duas horas antes do desligamento das lâmpadas do
biotério. Os anticorpos primários utilizados foram: anti-pAMPK alfa (T172) ( Cell
Signaling Technology, Cat #2531S, Danvers, MA, EUA) e anti-GAPDH ( Cell
Signaling, Cat #2118S, Danvers, MA, EUA). Os anticorpos secundários conjugados
com peroxidase (Bio-Rad Laboratories, Hercules, CA, EUA) foram utilizados, seguidos
de detecção quimioluminescente das bandas sobre filmes de raios-X. A densitometria
óptica dos filmes foi realizada utilizando o software de análise Scion Image (Scion
Corp., Frederick, MD, EUA).
Dosagens hormonais (direta e indireta)
Sangue do tronco foi colhido duas horas antes do desligamento das luzes (ZT
10) e o plasma foi extraído com heparina e armazenado a -80 ° C para determinação dos
níveis de corticosterona utilizando kit comercial de ELISA (Cat # 402810;. Neogen,
Lexington, KY, EUA). Para colher a urina, os ratos foram colocados numa gaiola
metabólica durante a fase escura do ciclo. A urina foi coletada em tubo de vidro
posicionado sob as gaiolas após seu acúmulo durante todo o período da noite. As
amostras de urina foram usadas para dosagem da 6-sulfatoxi-melatonina (6-S-Mel)

28
utilizando um kit de ELISA comercialmente disponível (Cat # RE54031; IBL
Internacional, Hamburgo, Alemanha).
Análise estatística
Os resultados foram expressos como média ± erro padrão da média (EPM) e
analisados estatisticamente por análise de variância (ANOVA de uma via com pós-teste
de Tukey-Kramer). Em todos os resultados adotou-se 5% como limite de significância
estatística (p < 0,05).

29
CAPÍTULO 1
______________________________________________________________________Regular Manuscript Metabolic impact of light phase-restricted fructose consumption is linked to
changes in hypothalamic AMPK phosphorylation and melatonin production
Juliana de Almeida Faria1, Thiago Matos F. de Araújo1, Letícia Martins I. de Souza2,
Silvana Bordin3, Gabriel Forato Anhê1*
Author affiliation: 1-Department of Pharmacology, Faculty of Medical Sciences, State
University of Campinas, Campinas, Brazil; 2-Faculty of Nutrition, Federal University of
Mato Grosso, Cuiabá, Brazil; 3-Department of Physiology and Biophysics, Institute of
Biomedical Sciences, University of Sao Paulo, Sao Paulo, Brazil
Running title: Fructose intake and melatonin production
Key words: Fructose, out-of-phase feeding, AMPK, corticosterone, melatonin
*Corresponding author: Please address correspondence to Dr. Gabriel Forato Anhê,
Department of Pharmacology, Faculty of Medical Sciences, State University of
Campinas, #105 Alexander Fleming St., Campinas, SP, Brazil. Zip Code: 13084-971.
Phone: 55-19-35219527. Fax: 55-19-32892968. E-mail: [email protected]

30
Abstract
Excessive fructose consumption can facilitate the development of glucose intolerance in
rodents and in humans. Recent studies have revealed that the metabolic outcomes
caused by fructose in mice are dependent on the phase of its consumption along with the
light/dark cycle. It is still unclear, however, if this time-of-the-day dependence is linked
to perturbations in parameters that display circadian rhythmicity. In the present study,
we investigated the effects of fructose consumption either during the light (LPF) or the
dark (DPF) phases of the light dark cycle over 8 weeks. We also investigated how these
changes were dependent on modulations of feeding behavior, hypothalamic AMPK
phosphorylation and melatonin and corticosterone production. LPF rats exhibited
increased AMPK phosphorylation in the hypothalamus and more pronounced glucose
intolerance and whole-body conversion of pyruvate into glucose when compared with
DPF rats. LPF rats, but not DPF rats, exhibited increased chow ingestion during the
light phase that was blunted by pharmacological inhibition of AMPK in the central
nervous system. LPF rats subjected to dark phase-restricted feeding still exhibited
increased hypothalamic AMPK phosphorylation but failed to develop the endocrine and
metabolic outcomes. Moreover, melatonin administration to LPF rats reduced
corticosterone levels and prevented glucose intolerance. Altogether, the present data
suggests that consumption of fructose during the light phase, rather than during the dark
phase, results in out-of-phase feeding due to increased hypothalamic AMPK
phosphorylation. This shift in spontaneous chow ingestion is responsible for the
reduction of melatonin production and the deleterious metabolic outcomes in LPF rats.

31
Introduction
It is well recognized that excessive fructose consumption can facilitate the development
of glucose intolerance in rodents and in humans. Sprague-Dawley rats fed a fructose-
enriched diet develop glucose intolerance and whole body insulin resistance [1]. In
mice, fructose-enriched diets were found to cause glucose intolerance with concomitant
hepatic triglyceride accumulation and insulin resistance [2]. Interventional experiments
with humans have also demonstrated that overweight subjects display glucose
intolerance after a 10 week interval of consumption of fructose sweetened beverages
[3]. Among the myriad of endocrine changes that putatively underlie these metabolic
effects, the consumption of a fructose-enriched diet was shown to reduce nocturnal
melatonin production in rats [4].
The relationship between melatonin and the control of energy metabolism has
been supported by several studies using distinct experimental approaches. Surgical
ablation of the pineal gland was reported to result in glucose intolerance and insulin
resistance with increased nocturnal levels of glycemia and gluconeogenesis [5,6,7].
Accordingly, the aging-related reduction of melatonin levels was shown to mediate the
increase of adiposity in middle-aged rats [8]. In turn, exogenous melatonin
administration is able to improve metabolic control in rodents rendered glucose
intolerant either by high-fat diets or fructose administration [9,10,11].
Recent studies have revealed that the metabolic outcomes caused by fructose
and high-fat diet intake are influenced by their out-of-phase consumption. Mice allowed
to consume a high-fat diet exclusively during the dark-phase fail to develop abrupt body
weight gain and glucose intolerance compared with those subjected to ad libitum or
light phase-restricted consumption [12,13]. In addition, fructose consumption by mice

32
exclusively during the light phase, but not during the dark phase, resulted in increased
body weight, adiposity and insulin levels [14].
Depending on its intensity and frequency, out-of-phase food intake by humans
can be classified as the night eating syndrome (NES) [15]. The prevalence of NES is
relatively low in the general population but ranges between 8.9 and 27% in obese
subgroups [16]. Cohort studies have shown that NES positively correlates with the
diagnosis of metabolic syndrome, increased triglycerides and waist circumference [17].
Among several adaptations, the circadian endocrine profile of NES patients is
characterized by reduced melatonin levels during the night and increased morning
cortisol concentrations [18].
Given the above mentioned observations, the present study was conducted to
investigate whether the metabolic impact resulting from fructose consumption during
different phases of the light/dark cycle is dependent on changes in the circadian pattern
of food intake in rats. We also collected results showing that disruption of melatonin
production is a key event in the mechanism linking the light phase-restricted fructose
consumption and its metabolic outcomes.
Materials and Methods
Animals, Surgical Procedure and Treatments
The experimental procedures were approved by the State University of Campinas
Committee for Ethics in Animal Experimentation (protocol n° 2798-1) and were
conducted in accordance with the guidelines of the Brazilian College for Animal
Experimentation.
Three-week-old male Sprague-Dawley rats were obtained from the Animal
Breeding Center at the University of Campinas (CEMIB, Campinas, Sao Paulo, Brazil)

33
and were housed at 22±2°C under a 12:12 h light:dark cycle (lights on at 7:00 a.m.)
with free access to food and water for 5 weeks. At 8 weeks of age, rats were assigned to
the experimental groups for an additional 8 weeks of treatment with fructose and/or
melatonin. Measurements of chow consumption and body mass were made twice and
once a week during the period of treatment, respectively. When specified in the results
section, two experimental strategies were combined to the fructose treatment: (i) chow
availability was restricted to the dark phase (Chow-R rats) during the 8 weeks of
treatment, or (ii) Compound C was administered during the last week of treatment
through a cannula placed in the lateral ventricle.
Fructose was dissolved in regular water to produce a 70% stock solution (w/v).
The fructose stock solution was further diluted to 10% (w/v) with regular water
immediately before treatment. Fructose was made available exclusively during the light
or the dark phases (respectively LPF and DPF rats). Bottles containing just water were
offered to the rats only when fructose was absent.
Melatonin (Cat. A9525; Sigma-Aldrich, St. Louis, MO, USA) was initially
diluted in 100% ethanol to generate a 100 mg/ml stock solution that was kept protected
from the light in -20°C for no longer than 7 days. The melatonin stock solution was
further diluted (1:50) with distilled water every 3 days of treatment to generate a 2
mg/ml solution. Variable volumes of this solution were added to the drinking water
bottle to yield a 0.5 mg/kg ingestion of melatonin. Bottles with melatonin were placed
on the cages 30 minutes before "lights off" and removed 30 min after "lights on". The
individual calculations of the required volumes of melatonin solution were based on the
body weight (weekly assessed) and nocturnal water intake (daily assessed) and were
adjusted daily.

34
Surgical Procedure and icv treatment
Six weeks after the beginning of fructose treatment, rats were anesthetized with
diazepam and ketamine (2 and 50 mg/kg, respectively) and placed in a stereotaxic
apparatus to insert a stainless-steel cannula into the lateral ventricle. Stereotaxic
coordinates were 0.8 mm (anteroposterior), 1.5 mm (lateral), and 4.0 mm (depth) [19].
The localization of the cannula was tested by evaluating the dipsogenic response to an
intracerebroventricular (icv) angiotensin II injection (15 ng/3 µl saline; Sigma, St Louis,
MO, USA) 1 week after the surgical procedure (7th week of fructose treatment). Only
animals that presented a positive response in these tests were used for further
experimentation.
Compound C (catalog no. 171260; EMD4 Biosciences, Gibbstown, NJ, USA)
was diluted in 5% dimethylsulfoxide (DMSO) to a final concentration of 200 mM and
was injected daily through the cannula for five days during the last week of fructose
treatment. Injections (2 µl) were performed 1 hour after "lights on" and equal volumes
of vehicle were injected in the controls. Experiments with these rats were carried out
two hours before "lights off" on the day of the last icv injection. Fasting prior to the
analyses and sample collection started immediately after the last injection.
Intraperitoneal pyruvate tolerance test (PTT)
Rats were fasted for 10 h, and a sodium pyruvate solution (250 mg/ml) was injected i.p.
at a dosage of 2 g/kg. Pyruvate injections were made two hours before "lights off".
Glucose concentration was determined in blood extracted from the tail before (0 min)
and 15, 30, 60, 90, and 120 min after pyruvate injection. The area under the curve
(AUC) of glycemia vs. time was calculated using each individual baseline (basal
glycemia) to estimate glucose clearance after pyruvate injection.

35
Intraperitoneal glucose tolerance test (GTT)
Rats were fasted for 10 h prior to i.p. glucose injection (2 g/kg of a 25% solution of D-
glucose) two hours before "lights off". The blood samples were collected from the tail at
0, 10, 15, 30, 60 and 120 min to determine the blood glucose concentration. The area
under the curve (AUC) of glycemia vs. time was calculated from each individual
baseline (basal glycemia) to estimate glucose tolerance.
Intraperitoneal insulin tolerance test (ITT)
Rats were fasted for 10 h prior to i.p. insulin injection (2 IU/kg) two hours before "lights
off". Blood glucose was measured before and 5, 10, 15, 20, 25 and 30 min after insulin
injection to determine the sensitivity of insulin-responsive tissues. Blood glucose values
were converted to a logarithmic scale, and the slope of the curve was calculated. This
value multiplied by 100 was assumed to be the glucose decay constant (KITT).
Protein extraction and immunoblotting
Anesthetized rats were decapitated, and the hypothalamus was removed and processed
for Western blotting as previously described [20]. The extractions occurred two hours
before "lights off". The primary antibodies used were as follows: anti-pAMPK alpha
(T172) from Cell Signaling Technology (Cat #2531S, Danvers, MA, USA) and anti-
GAPDH from Cell Signaling (Cat #2118S, Danvers, MA, USA). Secondary antibodies
conjugated with horseradish peroxidase (Bio-Rad Laboratories, Hercules, CA, USA)
were used, followed by chemiluminescent detection of the bands on x-ray-sensitive
films. Optical densitometry of the films was performed using the Scion Image analysis
software (Scion Corp., Frederick, MD, USA).

36
Hormone measurements
Trunk blood was collected two hours before "lights off", and plasma was extracted with
heparin and stored at -80°C for corticosterone determination with a commercially
available ELISA kit (Cat. #402810; Neogen, Lexington, KY, USA). For urine
collection, rats were placed in a metabolic cage during the dark phase. Urine was
allowed to accumulate overnight in a glass tube placed under the cages. Urine samples
were used for 6-sulfatoxi-melatonin (6-S-Mel) determination using a commercially
available ELISA kit (Cat. #RE54031; IBL International, Hamburg, Germany).
Statistical analysis
The results are presented as the means ± SE. Comparisons were performed using one-
way ANOVA, followed by Tukey-Kramer post hoc testing (INStat; GraphPad Software,
Inc., San Diego, CA, USA). Values of P˂0.05 indicate a significant difference.
Results
Metabolic and endocrine changes in rats exposed to fructose consumption during the
light or the dark phases
Body weights of CTL, LPF and DPF rats were similar at baseline, prior to the beginning
of treatments. Body weight gains after the 8 week treatments did not differ among the
groups so that final body weights were similar among rats assigned to the CTL, LPF
and DPF groups (Fig. 1A). Food intake was also assessed at the eighth week after the
beginning of treatments. The rats exhibited an expected nocturnal eating pattern so that,
for every group, food intake during the dark phase was higher than that during the light
phase (P<0.001). Consumption of chow during the light phase, however, was increased
in LPF rats (58% higher than CTL values; P<0.001), but reduced in DPF rats (44%

37
lower than CTL values; P<0.01). Chow consumption during the dark phase was
similarly reduced in LPF and DPF rats (23 and 31% lower than CTL, respectively;
P<0.01) (Fig. 1B). Glucose intolerance was found in LFP rats, but not in DFP rats, as
evidenced by increased AUC values obtained from the GTTs (120% higher than CTL;
P<0.05) (Fig. 1C). Conversely, the AUC values obtained from the curve glycemia vs.
time after a pyruvate load were increased in both LPF and DPF rats (respectively 114
and 42% higher than CTL; P<0.05). The values of DPF were, however, 44% lower than
those of LFP (P<0.001) (Fig. 1E). Apart from these changes in glucose tolerance, our
data revealed that neither LPF nor DPF rats exhibited insulin resistance as shown by
similar KITT values (Fig. 1G). Endocrine changes in LPF rats were hallmarked by
increased levels of corticosterone (91% higher than CTL; P<0.05) and reduced urinary
levels of 6-S-Mel (63% lower than CTL; P<0.05) (Figs. 1D and 1F, respectively). These
changes were not observed in DPF rats. The levels of hypothalamic AMPK
phosphorylation were increased in LPF rats but not in DPF rats (102% higher than CTL;
P<0.05) (Fig. 1H).
Increased food intake during the light phase is involved in the glucose intolerance
observed in LPF rats
To investigate if the increase in food intake during the light phase was involved in the
metabolic and endocrine changes observed in LPF animals, we designed an
experimental protocol in which the animals were assigned to four different groups: CTL
(rats that consumed chow ad libitum and did not receive fructose), LPF (rats that
consumed chow ad libitum and received fructose during the light phase), Chow-R (rats
that consumed chow exclusively during the dark phase and did not receive fructose) and

38
LPF/Chow-R (rats that consumed chow exclusively during the dark phase and received
fructose during the light phase).
The body weights of the animals assigned to the four groups were similar at the
beginning of the treatments. The four groups of animals exhibited a similar increase in
body weight so that absolute body weights at the end of the treatments were also similar
among the groups (Fig. 2A). The increase in food intake during the light phase exhibited
by LPF rats was replicated in this set of experiments (40% higher than food intake of
CTL during the light phase; P<0.05). The food intake during the dark phase was
increased in Chow-R compared with CTL rats (26% higher; P<0.05). Apart from that,
consumption of fructose during the light phase resulted in reduced food intake during
the dark phase irrespective of food restriction to this phase. Thus, food intake during the
dark phase was reduced in LPF/Chow-R (14% lower than in Chow-R; P<0.05) and in
LPF (12% lower than in CTL; P<0.05) (Fig. 2B).
LPF animals exhibited consistent glucose intolerance as evidenced by the AUC
values obtained from the GTT (98% higher than CTL; P<0.05). The tolerance to
exogenous glucose was not modulated in Chow-R compared with CTL animals.
Interestingly, glucose intolerance observed in LPF rats was not replicated in LPF/Chow-
R rats (Fig. 2C). With regard to the endocrine profile, Chow-R exhibited corticosterone
and urinary 6-S-Mel concentrations similar to those of CTL rats. The increase in
circulating corticosterone levels (77%; P<0.05) and the reduction in the urinary
concentration of 6-S-Mel (74%; P<0.05) observed in LPF rats when compared with
CTL rats were not detected in LPF/Chow-R (respectively, Figs. 2D and 2E). The
hypothalamic AMPK phosphorylation levels observed in CTL and in LPF rats were not
affected by restricting food availability to the dark phase. Thus, the amounts of

39
phosphorylated AMPK were increased in LPF compared with CTL rats (72%; P<0.05)
and in LPF/Chow-R compared with Chow-R rats (41%; P<0.05) (Fig. 2F).
Increased hypothalamic AMPK phosphorylation in LPF phase advances food intake
resulting in metabolic and endocrine changes
The previous data indicated that the increase of chow ingestion during the light phase
in LPF rats, although indispensable for the endocrine/metabolic changes, was not
causing the increase in hypothalamic AMPK phosphorylation. We assessed the
relevance of this increase in hypothalamic AMPK phosphorylation for LPF rats in a set
of experiments in which they received icv injections with Compound C, a
pharmacological AMPK inhibitor. The rats were assigned to four different groups in
these experiments, as follows: CTL (rats that did not receive fructose and were treated
with vehicle icv), LPF (rats that received fructose during the light phase and were
treated with vehicle icv), CC (rats that did not receive fructose and were treated with
Compound C icv) and LPF/CC (rats that received fructose during the light phase and
were treated with Compound C icv). Icv treatments lasted for 5 days during the eighth
week of fructose treatment.
Icv treatments did not interfere with the final body weight so that similar values
were obtained among the groups at the end of the eighth week of treatment. For the four
groups, the final body weight reached values higher than in the beginning of treatments
(P<0.05) (Fig. 3A). Before the beginning of icv treatments, food intake during the light
phase was increased in animals assigned to both LPF and LPF/CC groups (139 and
119% higher than CTL, respectively; P<0.05). Thus, surgical implantation of the
cannula in the lateral ventricle alone did not affect the response to light phase fructose
described in the previous experiments. After icv treatments, the LPF animals treated

40
with vehicle, but not those treated with Compound C, maintained increased values of
food intake during the light phase (200% higher than CTL after treatment with vehicle;
P<0.05) (Fig. 3B).
Food intake during the dark phase was similarly not affected by cannula
implantation per se so that before icv treatments, the amount of chow ingested by LPF
and LPF/CC groups during the period of "lights off" was lower than that ingested by
rats assigned to the CTL group (25 and 28% lower than CTL before icv treatment;
P<0,05). After icv treatments, food intake during the dark phase remained reduced in
both LPF and LPF/CC groups (32 and 34% lower than CTL after icv treatment with
vehicle; P<0.05) (Fig. 3C).
Glucose tolerance tests performed after icv treatments showed that icv injections
with vehicle did not interfere with the metabolic effect of fructose consumption during
the light phase. In these experiments, as in those described above, LPF animals treated
with vehicle became glucose intolerant as evidenced by the AUC values of the GTTs
(114% higher than in CTL after icv treatment with vehicle; P<0.05). When compared
with CTL rats, neither the CC nor the LPF/CC rats exhibited glucose intolerance after
icv treatments (Fig. 3D).
Corticosterone levels were increased in LPF rats treated with vehicle via icv
(150% higher than in CTL treated with vehicle; P<0.05). Corticosterone levels of
LPF/CC and CC rats remained similar to those of CTL rats after icv treatments (Fig.
3E). The urinary 6-S-Mel concentrations in LPF rats, but not in LPF/CC and CC rats,
were reduced after icv treatments (65% lower than CTL after icv treatment with vehicle;
P<0.05) (Fig. 3F). The phosphorylation levels of hypothalamic AMPK were increased
in LPF rats after treatment with vehicle (460% higher than in CTL treated with vehicle;

41
P<0.05). Treatment with Compound C icv blunted this response so that hypothalamic
AMPK phosphorylation in LPF/CC rats was similar to that of CTL rats (Fig. 3G).
Reduced melatonin production in LPF rats leads to increased corticosterone levels and
glucose intolerance
To determine the metabolic relevance for reduced melatonin production in LPF rats, we
designed an experimental protocol in which the animals were assigned to the following
four different groups: CTL (rats that did not receive either fructose or melatonin), LPF
(rats that received fructose during the light phase), Mel (rats that received melatonin
exclusively during the dark phase) and LPF/Mel (rats that received fructose during the
light phase and melatonin exclusively during the dark phase).
When compared to their initial body weights, the rats belonging to the four
experimental groups exhibited increased body weight at the end of the treatment
(P<0.05). Treatment with melatonin did not affect the changes in body weights
throughout the experimental period so that final body weights of the rats belonging to
the CTL, LPF, Mel and LPF/Mel groups were similar (Fig. 4A). The changes in the
food intake profile observed in LPF animals, hallmarked by increased values during the
light phase and reduced values during the dark phase, were not altered by melatonin
treatment. Additionally, melatonin treatment per se did not affect food intake. Thus,
food intake by both LPF and LPF/Mel rats was increased during the light phase
(approximately 50% higher than CTL; P<0.05) and reduced during the dark phase
(approximately 17% lower than CTL; P<0.05) (Fig. 4B).
Treatment with melatonin was able to reduce glucose intolerance induced by
fructose consumption during the light phase. The AUC values obtained from the GTT
were increased in LPF (138% higher than CTL; P<0.05) but not in LPF/Mel rats. In

42
turn, treatment with melatonin in fructose-naive rats did not alter glucose tolerance (Fig.
4C). As example of what was observed for glucose intolerance, corticosterone levels
were increased in LPF (249% higher than CTL; P<0.05) but not in LPF/Mel animals.
Melatonin treatment alone did not alter corticosterone concentrations (Fig. 4D).
In this set of experiments, we also found that LPF rats had reduced urinary 6-S-
Mel concentrations (80% lower than CTL; P<0.05). Fructose consumption during the
light phase, however, failed to reduce urinary 6-S-Mel concentrations in rats consuming
melatonin (urinary 6-S-Mel concentration are similar between Mel and LPF/Mel
groups). The urinary 6-S-Mel concentration was found to be similarly increased in both
Mel and LPF/Mel groups (187 and 254% higher than CTL, respectively; P<0.05) (Fig.
4E).
Melatonin treatment alone did not interfere with hypothalamic AMPK
phosphorylation so that the levels in rats belonging to the Mel group were similar to
those of the CTL group. Melatonin treatment was also unable to modulate the increase
in hypothalamic AMPK phosphorylation induced by the consumption of fructose during
the light phase (LPF and LPF/Mel were, respectively, 302 and 325% higher than CTL;
P<0.05) (Fig. 4F).
Discussion
The data presented herein show that rats receiving fructose during the light phase
exhibited glucose intolerance and increased whole-body conversion of pyruvate to
glucose simultaneously with increased chow ingestion during the light phase.
Importantly, these combined changes were not observed in rats receiving fructose
exclusively during the dark phase. Our data supports the conclusion that this shift in
food intake is of pivotal relevance for metabolic outcomes because fructose ingestion

43
during the light phase with simultaneous chow restriction to the dark phase fails to
induce glucose intolerance. This finding is in accordance with recent publications
showing that rats that increase their food intake during the light phase (either by forced
activity protocols during the light phase or by simple restriction of food availability)
become glucose intolerant [21]. Out-of-phase feeding seems also to be relevant for
human metabolism as subgroups of diabetic patients who display night eating behavior
also have impaired glycemic control based on increased glycated hemoglobin levels
[22].
The present data also allow us to conclude hypothalamic 5’-AMP-activated
protein kinase (AMPK) activation is a key event induced by fructose ingestion during
the light phase that increases out-of-phase feeding. In turn, out-of-phase feeding is
likely to be ultimately responsible for glucose intolerance. This sequential cause/effect
relationship is supported by our data showing: 1-fructose consumption during the light
phase is still able to induce hypothalamic AMPK phosphorylation in rats for which food
availability has been restricted to the dark phase; and 2-pharmacological inhibition of
AMPK with Compound C abrogates the shift in food intake and glucose intolerance
induced by fructose consumption during the light phase. Accordingly, previous studies
have already shown that fructose metabolism in the hypothalamus results in an increase
in the AMP/ATP ratio that activates AMPK and stimulates food intake [23].
Previous publications have also shown that hypothalamic AMPK activation with
pharmacological approaches triggers a counter-regulatory response hallmarked by
increased Endogenous Glucose Production (EGP). The mechanisms for this response
are not completely understood. However, it was demonstrated that hypothalamic AMPK
activation is able to spread peripheral signals that lead to the secretion of
glucocorticoids, glucagon and catecholamines [24,25,26]. These hormones are

44
classically known to act in the liver by increasing gluconeogenesis and glycogenolysis,
therefore stimulating EGP. In this context, we have previously demonstrated that intra-
cerebro ventricular injections with fructose during the light phase lead to an acute
activation of hypothalamic AMPK in the central nervous system and consequently
increases corticosterone levels that raise whole-body gluconeogenesis [20]. Increased
corticosterone levels as an acute response to an oral fructose load have been formerly
demonstrated by other groups [27,28].
The present data add further information to this field by revealing that chronic
fructose consumption can also increase both hypothalamic AMPK phosphorylation and
corticosterone levels when it occurs exclusively during the resting light phase. As
example of glucose intolerance, increased corticosterone levels were also prevented in
LPF rats by restricting chow availability to the night or by pharmacologically inhibiting
AMPK in the central nervous system. Based on our interpretation, the ability of chronic
fructose consumption during the light phase to increase corticosterone levels relies on
the chronically light phase-shifted chow ingestion induced by hypothalamic AMPK
activation.
Having established that hypothalamic AMPK activation and increased food
intake during the light phase are important for the increase in corticosterone levels
observed in LPF rats, we next explored how changes in the central nervous system
result in peripheral endocrine modulations. It was previously demonstrated that
exposing Sprague-Dawley rats to a 60% fructose-enriched diet ad libitum resulted in a
reduction of the levels of urinary 6-sulfatoxy melatonin [4]. The day-time consumption
of carbohydrates by rodents, however, seems to be particularly relevant to yield
reductions in nocturnal melatonin production. Selmaoui et al. have demonstrated that
the reduced amplitude of nocturnal melatonin levels is caused by offering a combination

45
of a carbohydrate-enriched diet and standard chow during the light-resting phase.
Similar protocols with standard chow alone failed to modulate melatonin production
[29]. Accordingly, the present data reveals that the consumption of fructose during the
light phase needs to co-exist with increased chow ingestion during this period of the
light/dark cycle to result in reduced 6-S-Mel production and increased corticosterone
levels. This was concluded because restriction of chow availability to the dark phase
prevented the reduction in 6-S-Mel observed in LPF rats. Our data from the experiments
with Compound C further corroborates this hypothesis because the pharmacological
inhibition of AMPK in the central nervous system abrogated both the shift in food
intake to the light phase observed in LPF rats and the reduction in 6-S-Mel.
To date, the negative modulation of melatonin secretion secondary to
hypothalamic AMPK activation has already been shown in other species. Menassol et
al. demonstrated that acute icv injection with AICAR (a pharmacological AMPK
activator) in ewes can actually reduce the amplitude of the nocturnal melatonin surge.
This modulation occurred irrespective of changes in the rhythm of melatonin production
[30]. As our data suggest, the ability of hypothalamic AMPK activation induced by
fructose ingestion during the light phase to reduce melatonin production relies on
changes in the rhythm of feeding behavior of the rat. Whether this applies to different
species remains to be determined.
The causal relationship between the reduced melatonin and increased
corticosterone was further examined in our experiments in which LPF rats were treated
with melatonin. We collected evidence to support the proposition that reduced
melatonin in rats consuming fructose during the light phase is a key event for the
increase in corticosterone levels and glucose intolerance as these adaptations were not
observed in LPF rats receiving melatonin. Accordingly, melatonin has already been

46
demonstrated to blunt insulin resistance induced by ad libitum consumption of a 60%
fructose enriched diet in Wistar rats [10].
The increased corticosterone levels observed in experimental conditions
characterized by reduced melatonin production can be explained by the suppressive
action that the pineal hormone exert on the Hypothalamus-Pituitary-Adrenal (HPA)
axis. It has already been shown that melatonin acts through MT1 receptors to suppress
ACTH-induced cortisol production in cultured adrenal glands isolated from primates
[31]. A similar response was found in cultured adrenal glands from rats [32]. It is
important to note that the suppressive action of melatonin over the HPA axis might not
be restricted to direct action on the adrenal glands because rats treated with melatonin
were also shown to have reduced CRH and ACTH levels after a stress stimulus [33].
In summary, the present study demonstrates that fructose consumption during
the light phase, but not the dark phase, results in glucose intolerance and increased
corticosterone levels. The effects of day-time consumption of fructose are secondary to
hypothalamic AMPK activation and to upregulation of food intake during the light
phase. We also show that the reduction of melatonin production due to hypothalamic
AMPK activation is a key event that mediates the increase in corticosterone and glucose
intolerance induced by the consumption of fructose during the light phase.
Acknowledgements
The authors would like to thank Mr. Miguel Borges da Silva, Mr. Antonio Vilson dos
Santo and Mr. Agnaldo Fernando de Azevedo for their technical assistance (State
University of Campinas). This study was supported by the Research Foundation of the
State of Sao Paulo (FAPESP) and the National Council of Research (CNPq).

47
Author Contributions
Juliana de Almeida Faria designed and performed the experiments, acquired the data,
contributed to data analysis/interpretation, prepared and approved the manuscript.
Thiago Matos F. de Araújo performed the experiments and approved the article. Letícia
Martins I. de Souza performed the experiments and approved the manuscript. Silvana
Bordin prepared and approved the manuscript. Gabriel Forato Anhê designed the
experiments, contributed to the data analysis/interpretation, prepared and approved the
manuscript.
References
1. ORON-HERMAN M, KAMARI Y, GROSSMAN E, et al. Metabolic syndrome:
comparison of the two commonly used animal models. Am J Hypertens 2008;
21:1018-1022
2. CHAN SM, SUN RQ, ZENG XY, et al. Activation of PPARα ameliorates
hepatic insulin resistance and steatosis in high fructose-fed mice despite
increased endoplasmic reticulum stress. Diabetes 2013; 62:2095-2105.
3. STANHOPE KL, SCHWARZ JM, KEIM NL, et al. Consuming fructose-
sweetened, not glucose-sweetened, beverages increases visceral adiposity and
lipids and decreases insulin sensitivity in overweight/obese humans. J Clin
Invest 2009; 119:1322-1334.
4. LEIBOWITZ A, PELEG E, SHARABI Y, et al. The role of melatonin in the
pathogenesis of hypertension in rats with metabolic syndrome. Am J Hypertens
2008; 21:348-351.

48
5. LA FLEUR SE, KALSBEEK A, WORTEL J, et al. Role for the pineal and
melatonin in glucose homeostasis: pinealectomy increases night-time glucose
concentrations. J Neuroendocrinol 2001; 13:1025-1032.
6. LIMA FB, MACHADO UF, BARTOL I, et al. Pinealectomy causes glucose
intolerance and decreases adipose cell responsiveness to insulin in rats. Am J
Physiol 1998; 275:E934-941.
7. NOGUEIRA TC, LELLIS-SANTOS C, JESUS DS, et al. Absence of melatonin
induces night-time hepatic insulin resistance and increased gluconeogenesis due
to stimulation of nocturnal unfolded protein response. Endocrinology 2011;
152:1253-1263.
8. WOLDEN-HANSON T, MITTON DR, MCCANTS RL, et al. Daily melatonin
administration to middle-aged male rats suppresses body weight, intraabdominal
adiposity, and plasma leptin and insulin independent of food intake and total
body fat. Endocrinology 2000; 141:487-497.
9. SHIEH JM, WU HT, CHENG KC, et al. Melatonin ameliorates high fat diet-
induced diabetes and stimulates glycogen synthesis via a PKCzeta-Akt-
GSK3beta pathway in hepatic cells. J Pineal Res 2009; 47:339-344.
10. KITAGAWA A, OHTA Y, OHASHI K. Melatonin improves metabolic
syndrome induced by high fructose intake in rats. J Pineal Res 2012; 52:403-
413.
11. CANO BARQUILLA P, PAGANO ES, JIMÉNEZ-ORTEGA V, et al.
Melatonin normalizes clinical and biochemical parameters of mild inflammation
in diet-induced metabolic syndrome in rats. J Pineal Res 2014; 57:280-290.

49
12. ARBLE DM, BASS J, LAPOSKY AD, et al. Circadian timing of food intake
contributes to weight gain. Obesity (Silver Spring) 2009; 17:2100-2102.
13. HATORI M, VOLLMERS C, ZARRINPAR A, et al. Time-restricted feeding
without reducing caloric intake prevents metabolic diseases in mice fed a high-
fat diet. Cell Metab 2012; 15:848-860.
14. MORRIS M, ARAUJO IC, POHLMAN RL, et al. Timing of fructose intake: an
important regulator of adiposity. Clin Exp Pharmacol Physiol 2012; 39:57-62.
15. STUNKARD AJ, GRACE WJ, WOLFF HG. The night-eating syndrome: a
pattern of food intake among certain obese patients. Am J Med 1955; 19:78-86.
16. RAND CS, MACGREGOR AM, STUNKARD AJ. The night eating syndrome
in the general population and among postoperative obesity surgery patients. Int J
Eat Disord 1997; 22:65-69.
17. GALLANT A, DRAPEAU V, ALLISON KC, et al. Night eating behavior and
metabolic heath in mothers and fathers enrolled in the QUALITY cohort study.
Eat Behav 2014; 15:186-191.
18. BIRKETVEDT GS, FLORHOLMEN J, SUNDSFJORD J, et al. Behavioral and
neuroendocrine characteristics of the night-eating syndrome. JAMA 1999;
282:657-663.
19. LIN QM, ZHAO S, ZHOU LL, et al. Mesenchymal stem cells transplantation
suppresses inflammatory responses in global cerebral ischemia: contribution of
TNF-α-induced protein 6. Acta Pharmacol Sin 2013; 34:784-792.

50
20. KINOTE A, FARIA JA, ROMAN EA, et al. Fructose-induced hypothalamic
AMPK activation stimulates hepatic PEPCK and gluconeogenesis due to
increased corticosterone levels. Endocrinology 2012; 153:3633-3645.
21. SALGADO-DELGADO RC, SADERI N, BASUALDO MDEL C, et al. Shift
work or food intake during the rest phase promotes metabolic disruption and
desynchrony of liver genes in male rats. PLoS One 2013; 8:e60052.
22. HOOD MM, REUTRAKUL S, CROWLEY SJ. Night eating in patients with
type 2 diabetes. Associations with glycemic control, eating patterns, sleep, and
mood. Appetite 2014; 79:91-96.
23. CHA SH, WOLFGANG M, TOKUTAKE Y, et al. Differential effects of central
fructose and glucose on hypothalamic malonyl-CoA and food intake. Proc Natl
Acad Sci USA 2008; 105:16871-16875.
24. MCCRIMMON RJ, SHAW M, FAN X, et al. Key role for AMP-activated
protein kinase in the ventromedial hypothalamus in regulating counterregulatory
hormone responses to acute hypoglycemia. Diabetes 2008; 57:444-450.
25. HAN SM, NAMKOONG C, JANG PG, et al. Hypothalamic AMP-activated
protein kinase mediates counter-regulatory responses to hypoglycaemia in rats.
Diabetologia 2005 48:2170-2178.
26. ALQUIER T, KAWASHIMA J, TSUJI Y, et al. Role of hypothalamic adenosine
5'-monophosphate-activated protein kinase in the impaired counterregulatory
response induced by repetitive neuroglucopenia. Endocrinology 2007; 148:1367-
1375.

51
27. BRINDLEY DN, COOLING J, GLENNY HP, et al. Effects of chronic
modification of dietary fat and carbohydrate on the insulin, corticosterone and
metabolic responses of rats fed acutely with glucose, fructose or ethanol.
Biochem J 1981; 200:275-283.
28. BRINDLEY DN, SAXTON J, SHAHIDULLAH H, et al. Possible relationships
between changes in body weight set-point and stress metabolism after treating
rats chronically with D-fenfluramine. Effects of feeding rats acutely with
fructose on the metabolism of corticosterone, glucose, fatty acids, glycerol and
triacylglycerol. Biochem Pharmacol 1985; 34:1265-1271
29. SELMAOUI B, OGUINE A, THIBAULT L. Food access schedule and diet
composition alter rhythmicity of serum melatonin and pineal NAT activity.
Physiol Behav 2001; 74:449-455.
30. MENASSOL JB, TAUTOU C, COLLET A, et al. The effect of an
intracerebroventricular injection of metformin or AICAR on the plasma
concentrations of melatonin in the ewe: potential involvement of AMPK? BMC
Neurosci 2011; 12:76.
31. TORRES-FARFAN C, RICHTER HG, ROJAS-GARCÍA P, et al. mt1
Melatonin receptor in the primate adrenal gland: inhibition of
adrenocorticotropin-stimulated cortisol production by melatonin. J Clin
Endocrinol Metab 2003; 88:450-458.
32. RICHTER HG, TORRES-FARFAN C, GARCIA-SESNICH J, et al. Rhythmic
expression of functional MT1 melatonin receptors in the rat adrenal gland.
Endocrinology 2008; 149:995-1003.

52
33. KONAKCHIEVA R, MITEV Y, ALMEIDA OF, et al. Chronic melatonin
treatment and the hypothalamo-pituitary-adrenal axis in the rat: attenuation of
the secretory response to stress and effects on hypothalamic neuropeptide
content and release. Biol Cell 1997; 89:587-596.

53
Figure Legends
Figure 1: Metabolic and endocrine changes in rats exposed to fructose
consumption during the light or the dark phases. Rats assigned to the groups CTL
(no fructose), LPF (fructose during the light phase) and DPF (fructose during the dark
phase) had their body weights assessed before and after (8 weeks) treatments (A). Food
intake during the light and the dark phases were also assessed at the end of the
treatments (B). After these measurements, the rats were subjected to glucose tolerance
tests (C), pyruvate tolerance tests (D) and insulin tolerance tests (E). Tests were
performed two hours before "lights off". Euthanasia was performed two hours before
"lights off" and the hypothalamus and plasma were collected. Plasma samples were
used for the determination of corticosterone levels (F). Urine was collected overnight
before euthanasia for determination of 6-S-Mel concentration (G). Hypothalamus
samples were used for Western blot detection of pAMPK and normalization GAPDH
(H). The results are presented as the means ± SE. *P<0.05 vs. same group before
treatment; **P<0.05 vs. CTL at the same phase of the light/dark cycle; #P<0.05 vs. LPF
at the same phase of the light/dark cycle.
Figure 2: Night-restricted feeding in LPF rats. Rats were assigned to the groups CTL
(rats that consumed chow ad libitum and did not receive fructose), LPF (rats that
consumed chow ad libitum and received fructose during the light phase), Chow-R (rats
that consumed chow exclusively during the dark phase and did not receive fructose) and
LPF/Chow-R (rats that consumed chow exclusively during the dark phase and received
fructose during the light phase). Body weights were assessed before and after (8 weeks)
treatments (A). Food intake during the light and the dark phases were also assessed at
the end of the treatments (B). After these measurements, the rats were subjected to

54
glucose tolerance tests (C). Tests were performed two hours before "lights off".
Euthanasia was performed two hours before "lights off" and the hypothalamus and
plasma were collected. Plasma samples were used for the determination of
corticosterone levels (D). Urine was collected overnight before euthanasia for
determination of the 6-S-Mel concentration (E). Hypothalamus samples were used for
Western blot detection of pAMPK and normalization GAPDH (F). The results are
presented as the means ± SE.*P<0.05 vs. same group before treatment; **P<0.05 vs.
CTL at the same phase of the light/dark cycle; #P<0.05 vs. Chow-R at the same phase
of the light/dark cycle.
Figure 3: Pharmacological inhibition of AMPK in the central nervous system of
LPF rats. Rats were assigned to the groups CTL (rats that did not receive fructose and
were treated with vehicle icv), LPF (rats that received fructose during the light phase
and were treated with vehicle icv), CC (rats that did not receive fructose were treated
with Compound C icv) and LPF/CC (rats that received fructose during the light phase
and were treated with Compound C icv). Cannula implantation and icv treatments (5
days) occurred during the sixth and eighth weeks of fructose treatment, respectively.
Body weights were assessed before and after (8 weeks) fructose treatment (A). Food
intake was assessed before and after icv injections during the last week of fructose
treatment. Data were acquired separately during the light (B) and the dark (C) phases.
After these measurements, the rats were subjected to glucose tolerance tests (D). Tests
were performed two hours before "lights off". Euthanasia was performed two hours
before lights off, and the hypothalamus and plasma were collected. Plasma samples
were used for corticosterone determinations (E). Urine was collected overnight before
euthanasia for determination of 6-S-Mel concentration (F). Hypothalamus samples were

55
used for Western blot detection of pAMPK and normalization GAPDH (G). The results
are presented as the means ± SE.*P<0.05 vs. same group before fructose treatment;
**P<0.05 vs. CTL at the same moment of icv treatment.
Figure 4: Nocturnal melatonin administration in LPF rats. Rats were assigned to the
groups CTL (rats that did not receive either fructose or melatonin), LPF (rats that
received fructose during the light phase), Mel (rats that received melatonin exclusively
during the dark phase) and LPF/Mel (rats that received fructose during the light phase
and melatonin exclusively during the dark phase). Body weights were assessed before
and after (8 weeks) treatments (A). Food intake during the light and the dark phases
were also assessed at the end of the treatments (B). After these measurements, the rats
were subjected to glucose tolerance tests (C). Tests were performed two hours before
"lights off". Euthanasia was performed two hours before "lights off" and the
hypothalamus and plasma were collected. Plasma samples were used for the
determination of corticosterone levels (D). Urine was collected overnight before
euthanasia for determination of 6-S-Mel concentration (E). Hypothalamus samples were
used for Western blot detection of pAMPK and normalization GAPDH (F). The results
are presented as the means ± SE.*P<0.05 vs. same group before treatment; **P<0.05 vs.
CTL at the same phase of the light/dark cycle.

56
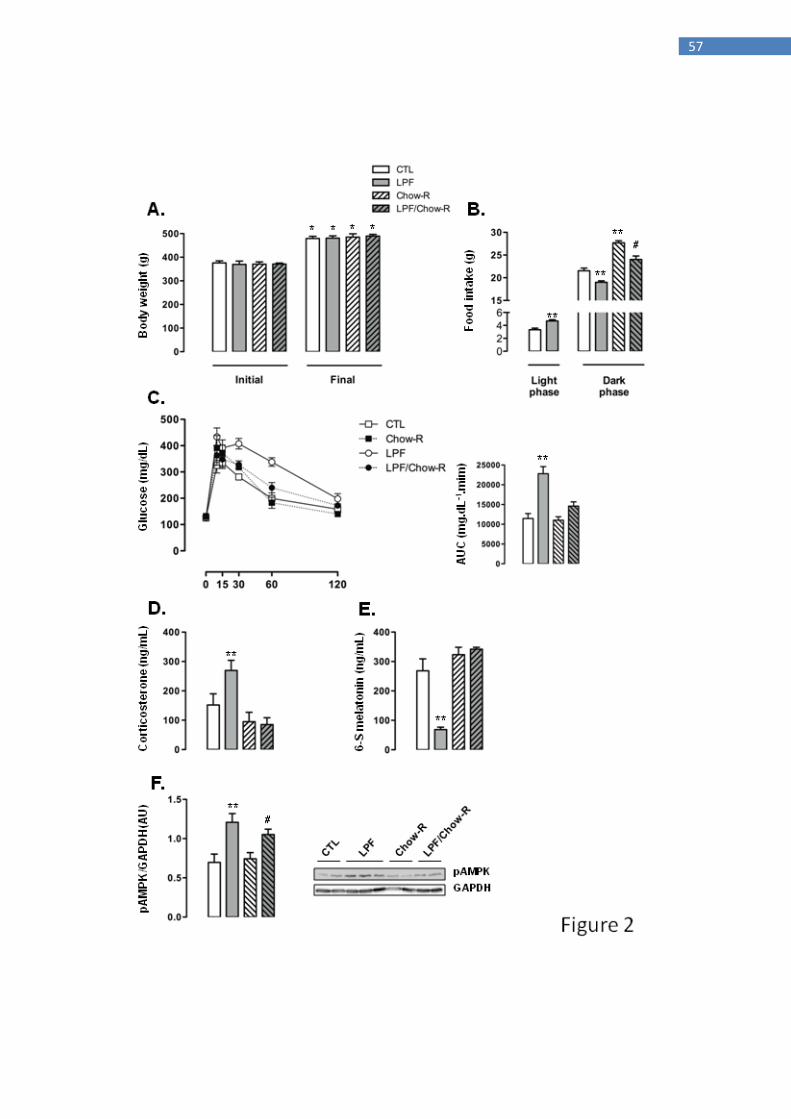
57
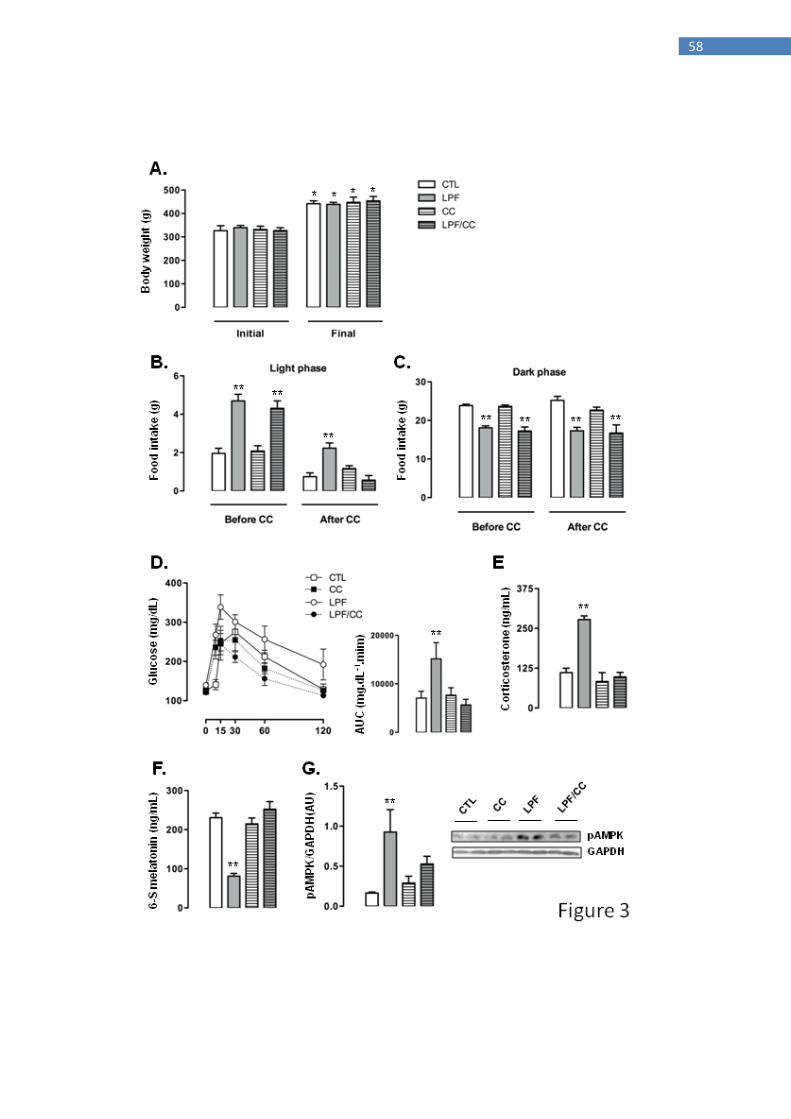
58

59

60
DISCUSSÃO
Os dados aqui apresentados mostram que os animais que receberam frutose
durante a fase clara do ciclo claro/escuro exibiram intolerância à glucose e aumento da
conversão sistêmica de piruvato em glicose simultaneamente ao aumento da ingestão
alimentar durante o mesmo o período do ciclo. É importante ressaltar que as alterações
citadas não foram observadas em ratos que receberam frutose exclusivamente durante a
fase escura do ciclo. Nossos dados permitem afirmar que o aumento da ingestão total de
calorias durante a fase clara do ciclo é de relevância fundamental para os resultados
metabólicos observados, uma vez que a ingestão de frutose durante a fase clara
associada simultaneamente à restrição de ração nessa mesma fase (a ração, nesse caso, é
oferecida exclusivamente durante a fase escura) falha em induzir intolerância à glicose.
Este resultado está de acordo com as publicações recentes que demonstram que os ratos
que aumentam sua ingestão de alimentos durante a fase clara (ou por protocolos de
atividade forçada durante essa fase ou pela simples restrição da disponibilidade de
alimentos durante a fase escura) tornam-se intolerantes à glicose (Salgado-Delgado,
2013). A ingestão alimentar fora de fase (em geral, relacionada ao período de menor
atividade da espécie em questão) também parece ser relevante no que tange o
metabolismo humano como já descrito para subgrupos de pacientes diabéticos os quais
apresentam controle glicêmico prejudicado (com base no aumento dos níveis de
hemoglobina glicada) quando ocorre aumento no comportamento alimentar noturno
desses indivíduos (Hood, 2014).
Os presentes dados também permitem afirmar que a ativação hipotalâmica da
proteína quinase 5'-AMP (AMPK), induzida pela ingestão de frutose durante a fase
clara, constitui um evento chave levando, por sua vez, ao aumento da alimentação fora
de fase. Em última análise, a alimentação fora de fase é responsável pela intolerância à

61
glicose. A proposição da existência desta relação de causa/efeito sequencial é suportada
pelo nosso conjunto de dados mostrando: 1- O consumo de frutose durante a fase clara é
capaz de induzir a fosforilação da AMPK hipotalâmica em ratos mesmo quando a
disponibilidade de alimentos é restrita à fase escura do ciclo; 2- A inibição
farmacológica da AMPK com Composto C atenua o aumento da ingestão calórica fora
de fase e a intolerância à glicose induzidos pelo consumo de frutose exclusivamente
durante a fase clara do ciclo. Em consonância com os dados apresentados, estudos
anteriores já demonstraram que o metabolismo da frutose gera no hipotálamo uma
resposta caracterizada pelo aumento da razão AMP/ATP a qual estimula a ativação da
proteína AMPK e, em consequência, o aumento da ingestão de alimentos (Cha, 2008).
Publicações anteriores também demonstraram que a ativação da AMPK
hipotalâmica através do uso de agentes farmacológicos desencadeia uma resposta contra
regulatória caracterizada pelo aumento da produção endógena de glicose. Os
mecanismos para esta resposta não foram completamente elucidados. No entanto, foi
demonstrado que a ativação da AMPK hipotalâmica é capaz de desencadear sinais
periféricos que levam à secreção de glicocorticóides, glucagon e catecolaminas
(McCrimmom, 2008; Han, 2005; Alquier, 2007). Esses hormônios são classicamente
conhecidos por atuar no fígado ocasionando aumentados níveis de gliconeogênese e
glicogenólise e, portanto, estimulando a produção endógena de glicose. Neste contexto,
demonstramos previamente que as injeções intracerebroventriculares com frutose
durante a fase clara levam a ativação aguda da AMPK hipotalâmica e,
consequentemente, aumentos dos níveis de corticosterona que elevam a gliconeogênese
sistêmica (Kinote, 2012). O aumento dos níveis de corticosterona como uma resposta
aguda a uma carga de frutose por via oral foi anteriormente demonstrado por outros
grupos (Brindley, 1981; Brindley, 1985). Os dados aqui apresentados adicionam mais

62
informações a este campo ao revelar que o consumo crônico de frutose também reflete
em aumentados níveis de fosforilação da AMPK hipotalâmica e de corticosterona
quando ocorre o consumo desse nutriente exclusivamente durante a fase de menor
atividade do roedor, ou seja, a fase clara do ciclo. No que diz respeito à intolerância à
glicose, o aumento do nível de corticosterona foi prevenido quando o grupo LPF teve a
disponibilidade de alimento restrita ao período escuro do ciclo ou ainda quando a
proteína AMPK foi farmacologicamente inibida no sistema nervoso central. Com base
em nossa interpretação, a habilidade de o consumo crônico de frutose durante a fase
clara levar ao aumento dos níveis de corticosterona, baseia-se no deslocamento parcial
da ingestão alimentar (ingestão fora de fase) induzida pela ativação da AMPK
hipotalâmica.
Tendo estabelecido que a ativação da AMPK hipotalâmica com consequente
aumento da ingestão calórica durante a fase clara representa o gatilho metabólico para o
aumento dos níveis de corticosterona, o próximo objetivo foi explorar o papel das
modificações no sistema nervoso central sobre possíveis modulações endócrinas
periféricas. Interessantemente, foi demonstrado que a exposição de ratos Sprague-
Dawley machos a uma dieta rica em frutose (60% m/m) ad libitum resultou numa
redução considerável dos níveis urinários de 6-sulfatoxi-melatonina (Leibowitz, 2008),
principal produto do metabolismo hepático da melatonina. O consumo de carboidratos
durante a fase clara do ciclo por roedores, no entanto, parece ser particularmente
relevante para produzir reduções na produção de melatonina noturna. Selmaoui e
colaboradores demonstraram que a associação de uma dieta rica em carboidrato e uma
dieta balanceada durante a fase de menor atividade mostrou-se capaz de reduzir a
amplitude dos níveis noturnos de produção da melatonina. Protocolos semelhantes, com
uso exclusivo de dieta padrão, falharam em modular a produção de melatonina

63
(Selmaoui, 2001). Por conseguinte, os presentes dados revelam que o consumo de
frutose durante a fase clara precisa coexistir com o aumento da ingestão alimentar
durante o mesmo período para resultar na redução da produção de 6-S-Mel e aumento
dos níveis de corticosterona. Este desfecho foi confirmado uma vez que a restrição da
ingestão alimentar ao período escuro do ciclo preveniu a redução da 6-S-Mel observada
em animais do grupo LPF. Adicionalmente, os dados experimentais obtidos com a
administração do inibidor Composto C corroboram esta hipótese já que a inibição
farmacológica da AMPK no sistema nervoso central revoga tanto o aumento da ingestão
fora de fase quanto a citada redução da 6-S-Mel em animais do grupo LPF.
A modulação negativa da secreção de melatonina secundária à ativação da
AMPK hipotalâmica já foi demonstrada em outras espécies experimentadas. Menassol e
colaboradores demonstraram que a injeção icv aguda com AICAR (um clássico ativador
farmacológico da proteína AMPK) em ovelhas leva a efetiva redução na amplitude do
pico noturno de produção da melatonina. Esta modulação ocorreu independentemente
de mudanças no ritmo da produção de melatonina (Menassol, 2011). Como os dados
aqui mostrados sugerem, a capacidade da ativação da AMPK hipotalâmica induzida
pela ingestão diurna da frutose em reduzir a produção noturna de melatonina, depende
de alterações no comportamento alimentar. Ainda não foi completamente esclarecido se
tal fato aplica-se a diferentes espécies.
A relação causal entre a redução nos níveis de melatonina e aumento da
corticosterona foi ainda investigada em nossos experimentos onde o grupo que recebeu
frutose exclusivamente durante a fase clara foi tratado com melatonina (suplementada
em água potável durante a fase escura). Verificou-se que os animais que ingeriram
frutose durante a fase clara e concomitantemente melatonina durante a fase escura do
ciclo tiveram níveis aumentados de corticosterona e intolerância à glicose, prevenidos.

64
Por conseguinte, demonstrou-se previamente que a melatonina atenua a resistência à
insulina induzida pelo consumo de uma dieta composta por 60% de frutose ad libitum
em ratos Wistar (Kitagawa, 2012).
O aumento dos níveis de corticosterona observado em condições experimentais,
caracterizado por produção reduzida de melatonina, pode ser explicado pela ação
supressora que o hormônio produzido pela pineal exerce sobre o eixo hipotálamo-
pituitária-adrenal (HPA). Já foi demonstrado que a melatonina atua através de
receptores MT1 para suprimir a produção de cortisol induzida pelo hormônio ACTH em
glândulas supra-renais em cultura isoladas de primatas (Torres-Farfan, 2003). Uma
resposta similar foi encontrada nas glândulas supra-renais em cultura de ratos (Richter,
2008). É importante destacar que a ação supressora da melatonina ao longo do eixo
HPA não pode ser restrita a ação direta sobre as glândulas supra-renais uma vez que
demonstrou-se também que os ratos tratados com melatonina apresentaram reduzidos
níveis de CRH e ACTH depois de um estímulo de stress (Konakchieva, 1997).

65
CONCLUSÃO
Em resumo, a presente tese demonstra que o consumo de frutose durante a fase
clara, mas não durante a fase escura do ciclo claro/escuro, resulta em intolerância à
glucose e aumento dos níveis de corticosterona. Os prejuízos associados ao consumo de
frutose restrito à fase clara são secundários à ativação da AMPK hipotalâmica e à
regulação positiva da ingestão alimentar durante a mesma fase. Também demonstramos
que a redução da produção de melatonina devido à ativação da AMPK hipotalâmica
constitui um evento chave que medeia o aumento da corticosterona e a intolerância à
glucose induzidos pelo consumo de frutose exclusivamente durante o período claro do
ciclo claro/escuro.

66
CAPÍTULO 2
____________________________________________________________________________
Co-autoria/autoria de artigos publicados durante o período
1. De Almeida Faria, J., De Araújo, T.M.F., Isler Mancuso, R., Meulman, J.,
Ferreira, D.S., Batista, T.M., Vetorazzi, J., Silva, P.M.R., Rodrigues, S.C.,
Kinote, A.P. B.M., Carneiro, E.M., Bordin, S., Anhê, G.F. (2016) Day-restricted
feeding during pregnancy and lactation programs glucose intolerance and
impaired insulin secretion in male rat offspring. Acta Physiol 217:240-53.
Pag. 80
2. Ramos-Filho, A.C., Moscoso, J.A., Calmasini, F., Faria, J. de A., Anhê,
G.F., Mónica, F.Z., Antunes, E. (2014) Blockade of renin-angiotensin system
prevents micturition dysfunction in renovascular hypertensive rats. Eur J
Pharmacol 738:285-292.
Pag. 81
3. Faria, J. A., Kinote, A., Ignacio-Souza, L.M., de Araujo, T.M., Razolli, D.S.,
Doneda, D.L., Paschoal, L.B., Lellis-Santos, C., Bertolini, G.L., Velloso, L.A.,
Bordin, S., Anhe, G. F. (2013) Melatonin acts through MT1/MT2 receptors to
activate hypothalamic Akt and suppress hepatic gluconeogenesis in rats. Am J
Physiol Endocrinol Metab. 305(2):E230-42
Pag.82
4. Ramos-Filho, A.C., A. Faria, J., Calmasini, F.B., Teixeira, S.A., Mónica,
F.Z., Muscará, M.N., Gontijo, J.A., Anhê, G.F., Zanesco, A., Antunes, E. (2013)
The renin-angiotensin system plays a major role in voiding dysfunction of
ovariectomized rats. Life Sci. 93(22):820-9.
Pag. 83

67
5. Kinote, A., Faria, J. A.; Roman, E. A., Solon, C., Razolli, D.S., Ignacio-Souza,
L.M., Sollon, C.S., Nascimento, L.F., de Araujo, T.M., Barbosa, A.P.L., Lellis-
Santos, C., Velloso, L.A., Bordin, S., Anhe, G.F. (2012) Fructose-Induced
Hypothalamic AMPK Activation Stimulates Hepatic PEPCK and
Gluconeogenesis due to Increased Corticosterone Levels. Endocrinology
153(8):3633-45.
Pag. 84

68
REFERÊNCIAS BIBLIOGRÁFICAS
Alonso-Vale, M.I.; Andreotti, S.; Borges-Silva, C.N.; Mukai, P.Y.; Cipolla-Neto, J.;
Lima, F.B. (2006). “Intermittent and rhythmic exposure to melatonin in primary
cultured adipocytes enhances the insulin and dexamethasone effects on leptin
expression.” J Pineal Res 41: 28-34.
Alonso-vale, M.I.; Borges-Silva, C.N.; Anhê, G.F.; Andreotti, S.; Machado, M.A.;
Cipolla-Neto, J.; Lima, F.B. (2004) “Light/dark cycle-dependent metabolic changes in
adipose tissue of pinealectomized rats.” Horm Metab Res 36: 474-479.
Alquier, T., Kawashima, J., Tsuji, Y., Kahn, B.B. (2007). “Role of hypothalamic
adenosine 5'-monophosphate-activated protein kinase in the impaired counterregulatory
response induced by repetitive neuroglucopenia”. Endocrinology 148: 1367-1375.
Anhê, G.F., Caperuto, L.C., Pereira-da-Silva, M., Souza, L.C., Hirata, A.E., Velloso,
L.A., Cipolla-Neto, J., Carvalho, C.R. (2004) “In vivo activation of insulin receptor
tyrosine kinase by melatonin in the rat hypothalamus.” J Neurochem 90: 559-566.
Arble, D.M., Bass, J., Laposky, A.D., Vitaterna, M.H., Turek, F.W. (2009) “Circadian
timing of food intake contributes to weight gain.” Obesity 17: 2100-2102.
Birketvedt, G.S., Florholmen, J., Sundsfjord, J., Osterud, B., Dinges, D., Bilker, W.,
Stunkard, A. (1999) Behavioral and neuroendocrine characteristics of the night-eating
syndrome. JAMA 282: 657-663.

69
Bode, C., Dürr, H.K., Bode, J.C. (1981) “Effect of fructose feeding on the activity of
enzymes of glycolysis, gluconeogenesis, and the pentose phosphate shunt in the liver
and jejunal mucosa of rats.” Horm Metab Res 13: 379-383.
Brindley, D.N., Cooling, J., Glenny, H.P., Burditt, S.L., McKechnie, I.S. (1981)
“Effects of chronic modification of dietary fat and carbohydrate on the insulin,
corticosterone and metabolic responses of rats fed acutely with glucose, fructose or
ethanol.” Biochem J 200: 275-283.
Brindley, D.N., Saxton, J., Shahidullah, H., Armstrong, M. (1985) “Possible
relationships between changes in body weight set-point and stress metabolism after
treating rats chronically with D-fenfluramine. Effects of feeding rats acutely with
fructose on the metabolism of corticosterone, glucose, fatty acids, glycerol and
triacylglycerol.” Biochem Pharmacol 34: 1265-1271.
Brunet, A., Bonni, A., Zigmond, M.J., Lin, M.Z., Juo, P., HU, L.S., Anderson,
M.J., Arden, K.C., Blenis, J., Greenberg, M.E. (1999) “Akt promotes cell survival by
phosphorylating and inhibiting a Forkhead transcription factor.” Cell 96: 857-868.
Cano Barquilla, P., Pagano, E.S., Jiménez-Ortega, V,. Fernández-Mateos, P., Esquifino,
A.I., Cardinali, D.P. (2014) “Melatonin normalizes clinical and biochemical parameters
of mild inflammation in diet-induced metabolic syndrome in rats.” J Pineal Res 57: 280-
290.

70
Cha, S.H., Wolfgang, M., Tokutake, Y., Chohnan, S., Lane, M.D. (2008) “Differential
effects of central fructose and glucose on hypothalamic malonyl-CoA and food intake.”
Proc Natl Acad Sci U S A. 105: 16871-16875.
Chan, S.M., Sun, R.Q., Zeng, X.Y., Hoong, Z., Wang, H., Watt, M.J., Ye, J.M. (2013)
“Activation of PPARα ameliorates hepatic insulin resistance and steatosis in high
fructose-fed mice despite increased endoplasmic reticulum stress.” Diabetes 62: 2095-
2105.
Gallant, A., Drapeau, V., Allison, K.C., Tremblay, A., Lambert, M., O'Loughlin,
J., Lundgren, J.D. (2014) “Night eating behavior and metabolic heath in mothers and
fathers enrolled in the QUALITY cohort study.” Eat Behav 15: 186-191.
Ha, E., Yim, S.V., Chung, J.H., Yoon, K.S., Kang, I., Cho, Y.H., Baik, H.H. (2006)
“Melatonin stimulates glucose transport via insulin receptor substrate-
1/phosphatidylinositol 3-kinase pathway in C2C12 murine skeletal muscle cells.” J
Pineal Res 41: 67-72.
Han, S.M., Namkoong, C., Jang, P.G., Park, .IS., Hong, S.W., Katakami, H., Chun,
S., Kim, S.W., Park, J.Y., Lee, K.U., Kim, M.S. (2005) Hypothalamic AMP-activated
protein kinase mediates counter-regulatory responses to hypoglycaemia in rats.
Diabetologia 48:2170-2178.

71
Hatori, M., Vollmers, C., Zarrinpar, A., DiTacchio, L., Bushong, E.A., Gill, S., Leblanc,
M., Chaix, A., Joens, M., Fitzpatrick, J.A., Ellisman, M.H., Panda, S. (2012) “Time-
restricted feeding without reducing caloric intake prevents metabolic diseases in mice
fed a high-fat diet.” Cell Metab 15: 848-860.
Havel, P.J. (2005) “Dietary fructose: implications for dysregulation of energy
homeostasis and lipid/carbohydrate metabolism.” Nutr Rev 63: 133–157.
Hood, M.M., Reutrakul, S., Crowley, S.J. (2014) “Night eating in patients with type 2
diabetes. Associations with glycemic control, eating patterns, sleep, and mood.”
Appetite 79: 91-96.
Kinote, A., Faria, J.A., Roman, E.A., Solon, C., Razolli, D.S., Ignacio-Souza,
L.M., Sollon, C.S., Nascimento, L.F., de Araújo, T.M., Barbosa, A.P., Lellis-Santos,
C., Velloso, L.A., Bordin, S., Anhê, G.F. (2012) “Fructose-Induced Hypothalamic
AMPK Activation Stimulates Hepatic PEPCK and Gluconeogenesis due to Increased
Corticosterone Levels.” Endocrinology 153(8): 3633-45.
Kitagawa, A., Ohta, Y., Ohashi, K. (2012) “Melatonin improves metabolic syndrome
induced by high fructose intake in rats.” J Pineal Res 52: 403-413.
Konakchieva, R., Mitev, Y., Almeida, O.F., Patchev, V.K. (1997) “Chronic melatonin
treatment and the hypothalamo-pituitary-adrenal axis in the rat: attenuation of the

72
secretory response to stress and effects on hypothalamic neuropeptide content and
release.” Biol Cell 89: 587-596.
La Fleur, S.E., Kalsbeek, A., Wortel, J., Van Der Vliet, J., Buijs, R.M. (2001) Role for
the pineal and melatonin in glucose homeostasis: pinealectomy increases night-time
glucose concentrations. J Neuroendocrinol 13: 1025-1032.
Lê, K., Ith, M., Kreis R., Faeh, D., Bortolotti, M., Tran, C., Boesch, C., Tappy, L.
(2009) “Fructose overconsumption causes dyslipidemia and ectopic lipid deposition in
healthy subjects with and without a family history of type 2 diabetes.” Am J Clin Nutr
89: 1760-1765.
Leibowitz, A., Peleg, E., Sharabi, Y., Shabtai, Z., Shamiss, A., Grossman, E. (2008)
“The role of melatonin in the pathogenesis of hypertension in rats with metabolic
syndrome.” Am J Hypertens. 21: 348-351.
Licht, F.O. International Sugar Economic Yearbook and Directory. (2005) Turnbridge
Wells, UK: Agna Informa.
Lima, F.B., Matsushita, D.H., Hell, N.S., Dolnikoff, M.S.; Okamoto, M.M., Cipolla-
Neto J. (1994) “The regulation of insulin action in isolated adipocytes. Role of
periodicity of food intake, time of the day and melatonin.” Braz.J.Med.Biol.Res 27:
995-1000.

73
Lima, F.B., Machado, U.F., Bartol, I., Seraphim, P.M., Sumida, D.H., Moraes, S.M.,
Hell, N.S., Okamoto, M.M., Saad, M.J., Carvalho, C.R., Cipolla-Neto, J. (1998)
“Pinealectomy causes glucose intolerance and decreases adipose cell responsiveness to
insulin.” Am J Physiol 275: E934-941.
Lin, Q.M., Zhao, S., Zhou, L.L., Fang, X.S., Fu, Y., Huang, Z.T. (2013) “Mesenchymal
stem cells transplantation suppresses inflammatory responses in global cerebral
ischemia: contribution of TNF-α-induced protein 6.” Acta Pharmacol Sin 34: 784-792.
Marshall, R.O., Kooi, E.R. (1957) “Enzymatic conversion of D-glucose to D-fructose.”
Science.125(3249): 648-9.
Mayes, P.A. (1993) “Intermediary metabolism of fructose.” Am J Clin Nutr 58: 754S-
765S.
McCrimmon, R.J., Shaw, M., Fan, X., Cheng, H., Ding, Y., Vella, M.C., Zhou,
L., McNay, E.C., Sherwin, R.S. (2008) “Key role for AMP-activated protein kinase in
the ventromedial hypothalamus in regulating counterregulatory hormone responses to
acute hypoglycemia.” Diabetes 57: 444-450.
Menassol, J.B., Tautou, C., Collet, A., Chesneau, D., Lomet, D., Dupont, J., Malpaux,
B., Scaramuzzi, R.J. (2011) “The effect of an intracerebroventricular injection of

74
metformin or AICAR on the plasma concentrations of melatonin in the ewe: potential
involvement of AMPK?” BMC Neurosci 12: 76.
Morris, M., Araujo, I.C., Pohlman, R.L., Marques, M.C., Rodwan, N.S., Farah, V.M.A.
(2012) Timing of fructose intake: an important regulator of adiposity. Clinical and
Experimental Pharmacology and Physiology 39: 57-62
Nogueira, T.C., Lellis-Santos, C., Jesus, D.S., Taneda, M., Rodrigues, S.C., Amaral,
F.G., Lopes, A.M., Cipolla-Neto, J., Bordin, S., Anhê, G.F. (2011) “Absence of
melatonin induces night-time hepatic insulin resistance and increased gluconeogenesis
due to stimulation of nocturnal unfolded protein response.” Endocrinology 152: 1253-
1263.
Oron-Herman, M., Kamari, Y., Grossman, E., Yeger, G., Peleg, E., Shabtay,
Z., Shamiss, A., Sharabi, Y. (2008) “Metabolic syndrome: comparison of the two
commonly used animal models.” Am J Hypertens 21:1018-1022
Paxinos, G. and Watson, C. (1986) “The Rat Brain in Stereotaxic Coordinates.” 2nd
edn. Academic Press Inc San Diego, California.
Ragheb, R., Medhat, A.M., Shanab, G.M., Seoudi, D.M., Fantus, I.G. (2008) “Links
between enhanced fatty acid flux, protein kinase C and NFkappaB activation, and apoB-
lipoprotein production in the fructose-fed hamster model of insulin resistance.”
Biochem Biophys Res Commun 370: 134-139.

75
Rajasekar, P., Anuradha, C.V. (2007) “Fructose-induced hepatic gluconeogenesis: effect
of L-carnitine.” Life Sci 80: 1176-1183.
Rand, C.S., Macgregor, A.M., Stunkard, A.J. (1997) The night eating syndrome in the
general population and among postoperative obesity surgery patients. Int J Eat Disord
22: 65-69.
Richter, H.G., Torres-Farfan, C., Garcia-Sesnich, J., Abarzua-Catalan, L., Henriquez,
M.G., Alvarez-Felmer, M., Gaete, F., Rehren, G.E., Seron-Ferre, M. (2008) “Rhythmic
expression of functional MT1 melatonin receptors in the rat adrenal gland.”
Endocrinology 149: 995-1003.
Salgado-Delgado, R.C., Saderi, N., Basualdo Mdel C., Guerrero-Vargas, N.N., Escobar,
C., Buijs, R.M. (2013) “Shift work or food intake during the rest phase promotes
metabolic disruption and desynchrony of liver genes in male rats.” PLoS One 8:e60052.
Samuel, V.T. (2011) “Fructose induced lipogenesis: from sugar to fat to insulin
resistance.” Trends Endocrinol Metab 22: 60-65.
Sartori, C., Dessen. P., Mathieu, C., Monney, A., Bloch, J., Nicod, P., Scherrer, U.,
Duplain, H. (2009) “Melatonin improves glucose homeostasis and endothelial vascular
function in high-fat diet-fed insulin-resistant mice.” Endocrinology 150: 5311-5317.

76
Selmaoui, B., Oguine, A., Thibault, L. (2001) “Food access schedule and diet
composition alter rhythmicity of serum melatonin and pineal NAT activity.” Physiol
Behav 74: 449-455.
Shieh, J.M., Wu, H.T., Cheng, K.C., Cheng, J.T. (2009) “Melatonin ameliorates high fat
diet-induced diabetes and stimulates glycogen synthesis via a PKCzeta-Akt-GSK3beta
pathway in hepatic cells.” J Pineal Res 47: 339-344.
Stanhope, K.L., Schwarz, J.M., Keim, N.L., Griffen, S.C., Bremer, A.A., Graham,
J.L., Hatcher, B., Cox, C.L., Dyachenko, A., Zhang, W., McGahan, J.P., Seibert,
A., Krauss, R.M., Chiu, S., Schaefer, E.J., Ai, M., Otokozawa, S., Nakajima,
K., Nakano, T., Beysen, C., Hellerstein, M.K., Berglund, L., Havel, P.J. (2009)
“Consuming fructose-sweetened, not glucose-sweetened, beverages increases visceral
adiposity and lipids and decreases insulin sensitivity in overweight/obese humans.” J
Clin Invest 119: 1322-1334.
Stunkard, A.J., Grace, W.J., Wolff, H.G. (1955) “The night-eating syndrome: a pattern
of food intake among certain obese patients.” Am J Med 19: 78-86.
Tappy, L., Lê, K.A. (2010) “Metabolic effects of fructose and the worldwide increase in
obesity.” Physiol Rev 90: 23-46. Review.

77
Teff, K.L., Elliott, S.S., Tschöp, M., Kieffer, T.J., Rader, D., Heiman, M., Townsend,
R.R., Keim, N.L., D'alessio, D., Havel, P.J. (2004) “Dietary fructose reduces circulating
insulin and leptin, attenuates postprandial suppression of ghrelin, and increases
triglycerides in women.” J Clin Endocrinol Metab 89: 2963-2972.
Torres-Farfan, C., Richter, H.G., Rojas-García, P., Vergara, M., Forcelledo,
M.L., Valladares, L.E., Torrealba, F., Valenzuela, G.J., Serón-Ferré, M. (2003) “mt1
Melatonin receptor in the primate adrenal gland: inhibition of adrenocorticotropin-
stimulated cortisol production by melatonin.” J Clin Endocrinol Metab 88: 450-458.
Torres-Farfan, C., Richter, H.G., Germain, A.M., Valenzuela, G.J., Campino, C., Rojas-
García, P., Forcelledo, M.L., Torrealba, F., Serón-Ferré, M. (2004) “Maternal melatonin
selectively inhibits cortisol production in the primate fetal adrenal gland.” J Physiol
554: 841-856.
Torres-Farfan, C., Mendez, N., Abarzua-Catalan, L., Vilches, N., Valenzuela, G.J.,
Seron-Ferre, M. (2011) “A circadian clock entrained by melatonin is ticking in the rat
fetal adrenal.” Endocrinology 152: 1891-1900.
Um, J.H., Pendergast, J.S., Springer, D.A., Foretz, M., Viollet, B., Brown, A., Kim,
M.K., Yamazaki, S., Chung, J.H. (2011) “AMPK regulates circadian rhythms in a
tissue- and isoform-specific manner.” PLoS One 6: e18450.
White, J.S. (2008) “Straight talk about high-fructose corn syrup: what it is and what it
ain't.” Am J Clin Nutr 88(6): 1716S-1721S.

78
Wolden-Hanson, T., Mitton, D.R., McCants, R.L., Yellon, S.M., Wilkinson,
C.W., Matsumoto, A.M., Rasmussen, D.D. (2000) Daily melatonin administration to
middle-aged male rats suppresses body weight, intraabdominal adiposity, and plasma
leptin and insulin independent of food intake and total body fat. Endocrinology 141:
487-497.
Yao, X.H., Nyomba, B.L. (2008) “Hepatic insulin resistance induced by prenatal
alcohol exposure is associated with reduced PTEN and TRB3 acetylation in adult rat
offspring.” Am J Physiol Regul Integr Comp Physiol 294(6): R1797-806.
Zanquetta, M.M., Seraphim, P.M., Sumida, D.H., Cipolla-Neto, J., Machado, U.F.
(2003) “Calorie restriction reduces pinealectomy-induced insulin resistance by
improving GLUT4 gene expression and its translocation to the plasma membrane.” J
Pineal Res 35: 141-148.
Zhang, W., Patil, S., Chauhan, B., Guo, S., Powell, D.R., Le, J., Klotsas, A., Matika,
R., Xiao, X., Franks, R., Heidenreich, K.A., Sajan, M.P., Farese, R.V., Stolz, D.B., Tso,
P., Koo, S.H., Montminy, M., Unterman, T.G. (2006) “FoxO1 regulates multiple
metabolic pathways in the liver: effects on gluconeogenic, glycolytic, and lipogenic
gene expression.” J Biol Chem. 281: 10105-10117.

79
APÊNDICE

80

81

82

83

84
ANEXO


















