UNIVERSIDAD POLITÉCNICA DE MADRID ESCUELA TÉCNICA …oa.upm.es/32621/1/Jelena_Barbir.pdf ·...
145
UNIVERSIDAD POLITÉCNICA DE MADRID ESCUELA TÉCNICA SUPERIOR DE INGENIEROS AGRÓNOMOS HABITAT MANAGEMENT FOR CONSERVATION OF POLLINATORS IN AGRO-ECOSYSTEMS OF CENTRAL SPAIN TESIS DOCTORAL JELENA BARBIR BSc in Biology Madrid, 2014
Transcript of UNIVERSIDAD POLITÉCNICA DE MADRID ESCUELA TÉCNICA …oa.upm.es/32621/1/Jelena_Barbir.pdf ·...

UNIVERSIDAD POLITÉCNICA DE MADRID
ESCUELA TÉCNICA SUPERIOR DE INGENIEROS AGRÓNOMOS
HABITAT MANAGEMENT FOR CONSERVATION OF
POLLINATORS IN AGRO-ECOSYSTEMS OF CENTRAL SPAIN
TESIS DOCTORAL
JELENA BARBIR
BSc in Biology
Madrid, 2014

UNIVERSIDAD POLITÉCNICA DE MADRID
ESCUELA TÉCNICA SUPERIOR DE INGENIEROS AGRÓNOMOS
Habitat management for conservation of pollinators in
agro-ecosystems of Central Spain
TESIS DOCTORAL
Jelena Barbir
BSc in Biology
Madrid, 2014

Departamento de Producción Vegetal: Botánica y Protección Vegetal
Escuela Técnica Superior de Ingenieros Agrónomos
Habitat management for conservation of pollinators in
agro-ecosystems of Central Spain
PhD candidate: Jelena Barbir
BSc in Biology
Supervisors: José Dorado
PhD in Agronomy (ICA, CSIC, Madrid)
Francisco Rubén Badenes Pérez
PhD in Entomology (ICA, CSIC, Madrid)
Madrid, 2014

Tribunal nombrado por el Magfco. Y Excmo. Sr. Rector de la Universidad Politécnica
de Madrid, el día de de 2014.
Presidente D./Da
Vocal D./D.a
Vocal D./D.a
Vocal D./D.a
Secretario D./D.a
Suplente D./D.a
Suplente D./D.a
Realizada la lectura y defensa de la tesis el día de de 2014 en Madrid, en la
Escuela Técnica Superior de Ingenieros Agrónomos.
Calificación:
El Presidente Los Vocales
El Secretario

To the planet Earth ...
... with hope that this work will contribute its beauty and magnificence;
... and to my family.

Acknowledgments
First of all, I would like to use this opportunity to express gratitude to my supervisors,
José Dorado and Francisco Rubén Badenes Pérez.
José, thank you for being like a father to me, for helping me to adapt easily to the new
environment and group. Thank you for sharing with me your knowledge in the field and your
passion for science and agriculture. Thank you for your patience, for all your time spent helping
me out with my decisions, experimental designs and doubts. Also, thank you for being always
available when I needed someone to talk about anything (work, stress, happy moments,
vacations, health or even about missing my family). It was a real pleasure to work with you
during those 4 years.
Ruben, thank you for sharing with me all your knowledge and passion for science and
entomology. It was a unique experience to work with you and I am happy that I had an
opportunity to learn so much from you about science and life in general. I would like to thank
you for encouraging me to participate in several international conferences and for financing all
the inscriptions and trips. Thank you for being always there for me when I needed an advice
regarding my work, and also thank you for always being available to prioritize my deadlines.
Although not my supervisor, I feel like he was one of them. César, thank you for
recognizing my passion for science and for giving me an opportunity to be part of your research
team where I felt as another member of the family. Also, I want to thank you for being always
so nice to me, and always finding time for me with a big smile on your face. Thank you for your
positive energy!
In addition, I would like to thank to all members of the group “Weed Ecology”: to David
and José Manuel, who were always there for me whenever I needed help in the field, lab or to
share a tortilla in a bar; to Carolina, for her smiles and amazing trip to Japan; to Bea, for her
positive energy, empathy, smiles and help in the field where we shared some great moments
counting Plutellas; also special thanks to my friend, colleague and flatmate Dioni, for
motivating me to finish my thesis as soon as possible and have life again:).
I would also like to use this chance to thank Elisa Viñuela, my tutor at Polytechnic
University of Madrid, who helped me to identify some of the first pollinators in my study area
and for always being so positive and energetic. Also, I would like to use this chance to thank all
members of her group for being supportive and nice to me. Especially, I would like to thank to
my friend Agustin for sharing with me all those happy and frustrating moments during the PhD
thesis “process”, and to Celeste, who helped me in the field during that hot summer of 2012,
patiently counting bees and keeping aromatic plants alive.

My thanks go to Cedo Maksimovic (Imperial College London, UK) and Tijana Blanusa
(University of Reading, UK) who warmly received me during my 3-month research visit and
helped me with the experiments I was conducting there. Also, thanks to Rebeka Siling who was
helping me with my experiments in Reading and to Simon Potts and Robert Stuart for finding
time to receive me in Reading and help me sort out some of my major doubts regarding
experimental designs.
I would like to express my gratitude to Luis Oscar Aguado Martín and Mike Edwards.
Those two entomologists made me understand bees. Their passion reflects through my work,
my experiments and love for what I was doing those 4 years. Without this passion, all this work
would be just boring. So, Oscar and Mike, thank you for enabling me to have fun during those 4
years of my PhD, and thanks to your wives for being so friendly and cooking such a lovely
dishes while I was visiting you.
Probably none of my field experiments would turn out right without help of Pedro
Hernaiz and his “crew”, who solved all my problems in the experimental farm “La Poveda”.
Pedro, gracias por tu paciencia y todo el esfuerzo que hiciste para que mis experimentos salgan
bien. Muchas gracias a José, Tasio, Jesús, Paulino y especialmente quiero dar las gracias a
Carlos que venía incluso durante sus vacaciones a regar mis plantas en el campo.
I would like to thank to the Spanish National Research Council (CSIC) that awarded me
with the JAEpredoc scholarship, to the head of the Institute of Agricultural Sciences - María
Teresa García González, and to the CSIC staff: Cristina, Maria Cruz, Belen, Raimundo and
Chelo for helping me out with all the paperwork and documentation I needed to deal with
during those 4 years; to Clara for assuring me to get any weird or super old article I needed to
include in my thesis; to Laura Barrios for her help with statistic analysis and experimental
designs; to Antonio for having the cars always ready for me; to David and Victor for constantly
repairing my office light; and last but not least, I would like to thank to my “polish” friend
Casimiro for keeping my plants happy and in a good condition and to Elisa and Antonio who
were always so helpful and nice to me, maintaining my greenhouse experiments running. Also, I
would like to thank to all techs, master, PhD and postdoc students I have met in CSIC: to Clara,
Sergio, Raul, Lilia, Mikel, Pablo, MJ, Vanesa, Irene, Francesca, Leti, Carlos, Carlos, Dani,
Mario, Miguel, Celia, Anita, Rocco, Mariana... thanks for your friendship, for sharing with me
Spanish food, Madrid and “out of Madrid” excursions, parties ...
Thanks to all those amazing people I have met in Madrid: José, Paula, Fahmi, Ruth (for
being so patient correcting my English), Gorjana, Alicia, Ana Cobo ..., and of course to my
“precious” friend Clemence, for being always there for me whenever I needed a nice word or a
hug. To all my great international friends for being there for me: to Sergio, Paul, Andy, Marta,

Manal, Jojo, Jasna, Mike, Rob, Quentin, Hara, Jovanka, Meera, Gabi, Catalan group ... Many
thanks to my fantastic Serbian friends: Jelena, Duska, Ivana, Marija, Sanja NS, Elvira, Vesna,
Sanela, Ranko, Alpi, Zeljko... guys thank you for all those years of friendship and time we have
spent together. Especially, I would like to thank to Sanja and Jovanka for always, always,
always being there for me weather I am feeling good or bad, happy or sad, being annoying or
super-fun. Thanks girls for always holding my back!
The most important people in my life, I saved for the end. To my parents, Ljubisa and
Dragana, who passed through the war and fights, but they never stopped believing in life and
people. I want to thank them, for transmitting that kind of thinking to me and for making me the
person I am. Thank you for all fantastic moments, endless chats, trips, advices, hugs and
positive energy... but above all thank you for believing in me. Mama, tata, ne zaboravite, vi ste
najbolji roditelji koje jedno dete moze pozeleti. Ponosna sam na vas i volim vas do neba! I
would like to thank to my sister, Natasa, for her endless love, for her happy and funny spirit that
always cheers me up, for her “lousy” karaoke singing, for helping me out in the field and for
never stopping to believe in me. Also, I would like to thank to my super-grandmas, Milica and
Zlata, for being my inspiration and support. Hvala vam na vasoj neizmernoj ljubavi, na svakom
telefonskom razgovoru, savetu, poljupcu i zagrljaju. Takodjer, hvala vam na svakom obroku,
torti i soku pripremljenom sa ogromnom kolicinom ljubavi. Also, I would like to thank to the
rest of my family for always being there for me: Vedo, Tiki, Ivana, Bojana, tetka and Saco, teco,
strikan and Vesna, tetka Anica, Bojan, Bojan, Bilja, cika Mijo, and of course thanks to the
youngest crew of our family: Stefku, Daci and Dori! Also, I thank to the newest members of our
family, Alonso and Maite, for their love, support and for being part of my life.
Finally, I would like to thank to the love of my life and the best life partner one can
imagine - Xavieru. Your love, support, indefinite patience and care for me keep me alive and
joyful. I am grateful for every moment I have spent with you and I am looking forward to spend
with you the rest of my life.

Table of Content
Abstract .................................................................................................................... i
Resumen .................................................................................................................. iv
I. General introduction ................................................................................... 1
1. Agriculture and pollination ..................................................................... 1
2. Plant-pollinator interactions .................................................................... 3
3. Important groups of pollinators .............................................................. 4
4. Managing semi-natural habitats of pollinators within agro-ecosystems10
II. Research question and objectives ............................................................. 15
III. Functionality of aromatic (Lamiaceae) floral mixture in conservation of
pollinators in Central Spain ........................................................................................ 18
1. Introduction ........................................................................................... 18
2. Materials and methods .......................................................................... 20
3. Results ................................................................................................... 24
4. Discussion ............................................................................................. 33
IV. The attractiveness of flowering herbaceous plants to bees
(Hymenoptera: Apoidea) and hoverflies (Diptera: Syrphidae) in agro-ecosystems of
Central Spain .............................................................................................................. 38
1. Introduction ........................................................................................... 38
2. Materials and methods .......................................................................... 39
3. Results ................................................................................................... 44
4. Discussion ............................................................................................. 52

V. Wild rocket - effect of water deficit on growth, flowering and
attractiveness to pollinators ........................................................................................ 57
1. Introduction ........................................................................................... 57
2. Material and methods ............................................................................ 58
3. Results ................................................................................................... 63
4. Discussion ............................................................................................. 77
VI. Can proximity of floral field margins improve pollination and seed
production of coriander crop, Coriandrum sativum L. (Apiaceae)? .......................... 81
1. Introduction ........................................................................................... 81
2. Material and methods ............................................................................ 83
3. Results ................................................................................................... 87
4. Discussion ............................................................................................. 93
VII. General discussion ................................................................................ 96
VIII. Conclusions ......................................................................................... 102
IX. Conclusiones ....................................................................................... 104
X. References .............................................................................................. 106
XI. Appendices .......................................................................................... 121
1. List of abbreviations ........................................................................... 121
2. List of figures ...................................................................................... 123
3. List of tables ........................................................................................ 126

Abstract
i
Abstract
Habitat management, aimed to conserve pollinators in agro-ecosystems, requires
selection of the most suitable plant species in terms of their attractiveness to pollinators
and simplicity of agronomic management. However, since all flowers are not equally
attractive to pollinators and many plant species can be weedy or invasive in the
particular habitat, it is important to test which plant species are the most appropriate to
be implemented in specific agro-ecosystems. For that reason, this PhD dissertation has
been focused on determination of the most appropriate aromatic and herbaceous plants
for conservation of pollinators in agro-ecosystems of Central Spain.
Therefore, in a first approximation, spatial expansion (i.e. potential weediness)
and attractiveness to pollinators of six aromatic perennial plants from the Lamiaceae
family, native and frequent in the Mediterranean region, were evaluated. Preliminary
plant selection was based on designing a functional mixed margins consisting of
plants attractive to pollinators and with different blooming periods, in order to extend
the availability of floral resources in the field. After a year of vegetative growth, the
next two years the plant species were studied in a randomized block design
experiment in order to estimate their attractiveness to pollinators in Central Spain and
to investigate whether floral morphology and density affect attractiveness to pollinators.
The final aim of the study was to evaluate how their phenology and attractiveness to
pollinators can affect the functionality of a flowering mixture of these plants. In
addition, the spatial expansion, i.e. potential weediness, of the selected plant species
was estimated under field conditions, as the final purpose of the studied plants is to be
implemented within agro-ecosystems. The results of the experiment showed that floral
morphology did not affect the attractiveness of plants to pollinators, but floral density
did, as plant species with higher floral density (i.e. Nepeta tuberosa and Hyssopus
officinalis) showed significantly higher attractiveness to pollinators. In addition, of six
plant species, two summer species (Melissa officinalis and Thymbra capitata) did not
efficiently contribute to the attractiveness of the mixture to pollinators, which reduced
its attractiveness during the summer period. Finally, as none of the plants showed
weedy behaviour under field conditions, the attractive plant species, i.e. N. tuberosa, H.
officinalis and the early spring flowering Salvia verbenaca, showed good potential to
conserve the pollinators.

Abstract
ii
Similarly, in a second approximation, the attractiveness to pollinators and
agronomic behaviour of twelve herbaceous plants blooming in spring were studied.
This experiment was conducted over two years in a randomized block design in order
to evaluate attractiveness of preselected plant species to pollinators, as well as their
attractiveness efficiency (a combination of duration of flowering and insect visitation),
their response to two different agronomic management practices (growing in mixed vs.
mono-specific plots; tillage vs. no-tillage), and their potential weediness. The results of
this experiment showed that the flowers of Borago officinalis, Echium plantagineum,
Phacelia tanacetifolia and Diplotaxis tenuifolia were attractive to bees, while Calendula
arvensis, Coriandrum sativum, D. tenuifolia and Lobularia maritima were attractive to
hoverflies. In addition, floral mixture resulted in lower attractiveness efficiency to
pollinators than mono-specific D. tenuifolia, but higher than most of the mono-specific
stands. On the other hand, although some of the most attractive plant species (e.g. P.
tanacetifolia and C. arvensis) showed potential weediness, their self-seeding was
reduced by tillage. After comparing attractiveness efficiency of various herbaceous
species to pollinators and their potential weediness, the results indicated that D.
tenuifolia showed the highest attractiveness efficiency to pollinators and efficient self-
reproduction, making it highly recommended to attract bees and hoverflies in agro-
ecosystems of Central Spain. In addition, this plant, commonly known as wild rocket,
has a supplementary economic value as a commercialized crop.
The implementation of a new floral margin in agro-ecosystems means increased
production costs, especially in regions with frequent and long droughts (as it is Central
and South-East area of Iberian Peninsula), where the principal agricultural cost is
irrigation. Therefore, before recommending D. tenuifolia for sustainable habitat
management within agro-ecosystems, it is necessary to study the effect of drought
stress and moderate and severe deficit irrigation on its growth, flower development
and attractiveness to pollinators. The results of this experiment showed that in
greenhouse conditions, potted D. tenuifolia could be without irrigation for 4 days
without affecting its growth, flowering and attractiveness to pollinators. However, lack
of irrigation for 8 days or longer significantly reduced the vegetative growth, number of
open flowers, total floral area, flower diameter, corolla tube diameter and corolla tube
length of D. tenuifolia. This study showed that regulated deficit irrigation can improve
water use efficiency, and depending on the purpose of growing D. tenuifolia, as a crop

Abstract
iii
or as a beneficial plant to attract pollinators, it can reduce water consumption by 40% to
70% without affecting its vegetative and floral development and without reducing its
attractiveness to pollinators.
Finally, the following experiment was developed in order to understand how
habitat management can influence on the agricultural production. For this purpose, it
was evaluated if the vicinity of mixed and mono-specific field margins, preselected
to conserve pollinators within agro-ecosystems, can improve seed production in
coriander (C. sativum). The selection of this plant species for the experiment was
based on its necessity for insect pollination for production of seeds (even though some
pollen can be transmitted from one flower to another by wind) and the fact that under
semi-controlled field conditions established in the field it is possible to estimate its total
seed production. Since D. tenuifolia is attractive for both bees and hoverflies in Central
Spain, the main objective of this experiment was to estimate the impact of two different
types of field margins, i.e. mono-specific margin with D. tenuifolia and mixed margin
with six herbaceous species, on the seed production of potted coriander. For that reason,
it was tested: i) if open pollination (control without proximate field margin and
treatments with nearby mono-specific and mixed margin) increases the seed production
of coriander when compared with no-pollination (covered inflorescences of coriander)
under field conditions; ii) if frequency of pollinator visitation to the flowers of coriander
was higher in the presence of field margins than in the control without field margin; and
iii) if seed production was higher in the presence of field margins than in control plants
of coriander without field margin. The results showed that the proximity of both types
of floral margins (mixed and mono-specific) improved the seed quality of coriander
plants, as seed weight and germination rate were higher than in control plants without
field margin. Furthermore, the number of seeds produced was significantly higher in
coriander plants grown near mixed margins than near mono-specific margin, probably
due to an increase in pollinator visits. Since the experiment was conducted under semi-
controlled field conditions, it can be concluded that pollinator visits was the main factor
that biased the results, and that presence of both mixed or mono-specific (D. tenuifolia)
margins can improve the production of coriander for more than 200% in small-scale
gardens and, in addition, conserve the local pollinators.

Resumen
iv
Resumen
La gestión de hábitat orientada a la conservación de polinizadores en los agro-
ecosistemas requiere una selección de especies vegetales atendiendo fundamentalmente
a dos criterios: i) el potencial atractivo de sus flores a los polinizadores; y ii) la
simplicidad en su manejo agronómico. Además de estas premisas, es necesario
considerar la capacidad invasora de estas especies vegetales, debido a que algunas de las
más atractivas pueden resultar invasoras en determinados agro-ecosistemas. Por lo
tanto, es preciso determinar qué especies vegetales son las más indicadas para ser
implementadas en cada agro-ecosistema. En la presente tesis doctoral se plantea la
búsqueda de las especies vegetales adecuadas para atraer polinizadores en los agro-
ecosistemas del centro de España.
En una primera aproximación, se ha evaluado la atracción y expansión espacial
(potencial invasivo) de seis plantas perennes de la familia Lamiaceae (aromáticas),
elegidas por ser nativas de la región mediterránea. La elección de las especies vegetales
se ha llevado a cabo con el fin de crear márgenes funcionales basados en la mezcla de
especies vegetales con distintos periodos de floración, de modo que prolonguen la
disponibilidad de recursos florales en el tiempo. Tras un primer año dedicado al
establecimiento de las especies aromáticas, en los dos años siguientes se ha estudiado la
atracción individual y combinada de las especies vegetales sobre los polinizadores, y
como ésta se ve afectada por la densidad y la morfología floral, utilizando para ello un
diseño experimental en bloques al azar. Los resultados de este estudio han puesto de
manifiesto que la morfología floral no tuvo influencia sobre la atracción de las especies
vegetales, pero si la densidad floral, puesto que las especies vegetales con mayor
densidad de flores (Nepeta tuberosa e Hyssopus officinalis) han mostrado mayor
atracción a polinizadores. Cabe destacar que de las seis especies consideradas, dos
especies de verano (Melissa officinalis y Thymbra capitata) no han contribuido de
forma efectiva a la atracción de la mezcla hacia los polinizadores, mostrando una
reducción significativa de este parámetro respecto a las otras especies aromáticas a lo
largo del verano. Se ha observado que ninguna de las especies aromáticas evaluadas ha
mostrado tendencia invasora a lo largo del estudio. En base a estos resultados, se puede
concluir que entre las especies aromáticas estudiadas, N. tuberosa, H. officinalis y
Salvia verbenaca son las que ofrecen mayor potencial para ser utilizadas en la
conservación de polinizadores.

Resumen
v
De forma similar al caso de las plantas aromáticas, se ha llevado a cabo una
segunda experimentación que incluía doce plantas anuales con floración de
primavera, en la que se evaluó la atracción a polinizadores y su comportamiento
agronómico. Este estudio con especies herbáceas se ha prolongado durante dos años,
utilizando un diseño experimental de bloques aleatorios. Las variables analizadas
fueron: el atractivo de las distintas especies vegetales a los polinizadores, su eficiencia
de atracción (calculada como una combinación de la duración de la floración y las
visitas de insectos), su respuesta a dos tipos de manejo agronómico (cultivo en mezcla
frente a monocultivo; laboreo frente a no-laboreo) y su potencial invasivo. Los
resultados de esta segunda experimentación han mostrado que las flores de Borago
officinalis, Echium plantagineum, Phacelia tanacetifolia y Diplotaxis tenuifolia son
atractivas a las abejas, mientras que las flores de Calendula arvensis, Coriandrum
sativum, D. tenuifolia y Lobularia maritima son atractivas a los sírfidos. Con
independencia del tipo de polinizadores atraídos por cada especie vegetal, se ha
observado una mayor eficiencia de atracción en parcelas con monocultivo de D.
tenuifolia respecto a las parcelas donde se cultivó una mezcla de especies herbáceas, si
bien en estas últimas se observó mayor eficiencia de atracción que en la mayoría de
parcelas mono-específicas. Respecto al potencial invasivo de las especies herbáceas, a
pesar de que algunas de las más atractivas a polinizadores (P. tanacetifolia and C.
arvensis) mostraron tendencia a un comportamiento invasor, su capacidad de auto-
reproducción se vio reducida con el laboreo. En resumen, D. tenuifolia es la única
especie que presentó una alta eficiencia de atracción a distintos tipos de polinizadores,
conjuntamente con una alta capacidad de auto-reproducción pero sin mostrar carácter
invasor. Comparando el atractivo de las especies vegetales utilizadas en este estudio
sobre los polinizadores, D. tenuifolia es la especie más recomendable para su cultivo
orientado a la atracción de polinizadores en agro-ecosistemas en el centro de España.
Esta especie herbácea, conocida como rúcula, tiene la ventaja añadida de ser una
especie comercializada para el consumo humano.
Además de su atractivo a polinizadores, deben considerarse otros aspectos
relacionados con la fisiología y el comportamiento de esta especie vegetal en los agro-
ecosistemas antes de recomendar su cultivo. Dado que el cultivo en un campo agrícola
de una nueva especie vegetal implica unos costes de producción, por ejemplo debidos a
la utilización de agua de riego, es necesario evaluar el incremento en dichos costes en

Resumen
vi
función de demanda hídrica específica de esa especie vegetal. Esta variable es
especialmente importante en zonas dónde se presentan sequías recurrentes como es el
caso del centro y sur-este de la península Ibérica. Este razonamiento ha motivado un
estudio sobre los efectos del estrés hídrico por sequía y el estrés por déficit
moderado y severo de riego sobre el crecimiento y floración de la especie D.
tenuifolia, así como sobre la atracción a polinizadores. Los resultados muestran que
tanto el crecimiento y floración de D. tenuifolia como su atracción a polinizadores no se
ven afectados si la falta de riego se produce durante un máximo de 4 días. Sin embargo,
si la falta de riego se extiende a lo largo de 8 días o más, se observa una reducción
significativa en el crecimiento vegetativo, el número de flores abiertas, el área total y el
diámetro de dichas flores, así como en el diámetro y longitud del tubo de la corola. Por
otro lado, el estudio pone de manifiesto que un déficit hídrico regulado permite una
gestión eficiente del agua, la cual, dependiendo del objetivo final del cultivo de D.
tenuifolia (para consumo o solo para atracción de polinizadores), puede reducir su
consumo entre un 40 y un 70% sin afectar al crecimiento vegetativo y desarrollo floral,
y sin reducir significativamente el atractivo a los polinizadores.
Finalmente, esta tesis aborda un estudio para determinar cómo afecta el manejo de
hábitat a la producción de los cultivos. En concreto, se ha planteado una
experimentación que incluye márgenes mono-específicos y márgenes con una mezcla
de especies atractivas a polinizadores, con el fin de determinar su efecto sobre la
producción del cultivo de cilantro (C. sativum). La elección del cultivo de cilantro se
debe a que requiere la polinización de insectos para su reproducción (aunque, en menor
medida, puede polinizarse también por el viento), además de la facilidad para estimar su
producción en condiciones semi-controladas de campo. El diseño experimental consistía
en la siembra de márgenes mono-específicos de D. tenuifolia y márgenes con mezcla de
seis especies anuales situados junto al cultivo de cilantro. Estos cultivos con márgenes
florales fueron comparados con controles sin margen floral. Además, un segundo grupo
de plantas de cilantro situadas junto a todos los tratamientos, cuyas flores fueron
cubiertas para evitar su polinización, sirvió como control para evaluar la influencia de
los polinizadores en la producción del cultivo. Los resultados muestran que la presencia
de cualquiera de los dos tipos de margen floral mejora el peso y el porcentaje de
germinación de las semillas de cilantro frente al control sin margen. Si se comparan los
dos tipos de margen, se ha observado un mayor número de semillas de cilantro junto al

Resumen
vii
margen con mezcla de especies florales respecto al margen mono-específico,
probablemente debido al mayor número visitas de polinizadores. Puesto que el
experimento se realizó en condiciones de campo semi-controladas, esto sugiere que las
visitas de polinizadores fueron el factor determinante en los resultados. Por otro lado,
los resultados apuntan a que la presencia de un margen floral (ya sea mono-especifico o
de mezcla) en cultivos de pequeña escala puede aumentar la producción de cilantro en
más de un 200%, al tiempo que contribuyen a la conservación de los polinizadores.

Chapter I
1
I. General introduction
1. Agriculture and pollination
Agriculture, defined as cultivation of plants, animals and other forms of life for
food, biofuel, medicinal and other products used to sustain and enhance human life
(ILC, 2000), is one of the oldest and greatest modifications of the ecosystems. Through
the history, a constant increase in human population have caused a gradual switching
from small-scale farming to an application of intensive agricultural practices where
usage of modern technologies, i.e. tillage, pesticides and fertilizers, have resulted in
massive biodiversity simplification and a reduction of ecosystem services (Altieri, 1999;
Benton et al., 2003; Watling and Donnelly, 2006; Gill et al., 2012). Nowadays,
agricultural land occupies around 45% of total Earth land area (New, 2005), irregularly
distributed worldwide (Fig. I-1) and therefore it should be managed in the most
sustainable way.
Fig. I-1. Percentage of agricultural land relative to the total land area across the Earth Source: World Bank, 2013

Chapter I
2
Pollination, i.e. transfer of pollen from anther to stigma, is one of the most
important ecosystem services in agriculture as it enables reproduction of plants
(Bastian, 2013). There are two types of pollination, abiotic and biotic. Abiotic
pollination takes place without the involvement of living organisms, for example, where
pollen is transported by wind (anemophily), while biotic pollination is the result of the
movement of pollen by living organisms and it is the most common form of pollination.
The most diverse and abundant group of pollinators are insects (Steffan-Dewenter et al.,
2005), hence in the following text the term “pollinators” will be used for insect
pollinators restrictively.
Focusing on insects, in one natural and balanced ecosystem, it is assumed that all
insects contribute to maintaining the system and therefore no species can be considered
to have positive or negative impact on the ecosystem. On the other side, in agriculture,
where the goal is to raise selected crops, insects that hinder production are classified as
pests, while insects that assist production are considered beneficial. Along with natural
enemies of pests, pollinators are the most important beneficial insects for agricultural
production (Carvalheiro et al., 2012) and wild plant reproduction (Biesmeijer et al.,
2006), as approximately 75 % of crop species of global significance for food production
and 65 % of plant species rely on pollinators (Klein et al., 2007).
Recently, interactions between pollinators and agriculture have generated
considerable research interest, stimulated by rising recognition of the significance of
insect pollination in maintaining yields of some crops (Santos et al., 2008; Carvalheiro
et al., 2012; Bartomeus et al., 2014) and coupled with the concern that many species of
pollinating insects have declined substantially (Biesmeijer et al., 2006). In the face of
global decline in biodiversity, the species interactions deserve particular attention. As
pollinators are strongly dependent on floral diversity due to mutual specialization,
declines in plant diversity (caused by land use intensification), may be associated with
extinction of pollinators (Frund et al., 2010). Although intensive agriculture is mainly
responsible for pollinator decline (Tscharntke et al., 2005; Rundlof et al., 2008; Le Feon
et al., 2010), there are other factors that negatively affect diversity and abundance of
pollinators, as the potential spread of diseases (Vanbergen and Insect Pollinators
Initiative, 2013) and changes in climate (Memmott et al., 2010; Rader et al., 2013).
Furthermore, various scientific proofs have been published recently showing the

Chapter I
3
negative effect of neonicotinoid insecticides on development and health of pollinators
(Gill et al., 2012; Stokstad, 2013; van der Sluijs et al., 2013).
Consequently, the decline in abundance and diversity of pollinators has become a
subject of public and political interest. Therefore, to promote farming in a way that it
supports biodiversity, many regions have introduced agri-environment schemes (AES),
including Australia, the European Union, USA and Switzerland (Morris and Potter,
1995; Albrecht et al., 2007; Batary et al., 2011; Elts and Lohmus, 2012). In that way,
conservation of semi-natural habitats has been supported economically by AES,
compensating the farmers’ income losses associated with employing environmentally
sustainable land (Dobbs and Pretty, 2004). In addition, in 2013 the European Union
decided to suspend the use of a number of neonicotinoids, which has boosted further
research on pollinator exposure to insecticides and the potential for sublethal effects
(Sandrock et al., 2014).
2. Plant-pollinator interactions
Recently, parallel diversity decline of bees and insect-pollinated plants has been
reported in Europe (Biesmeijer et al., 2006), suggesting a functional coupling and
mutual dependence between both guilds. This diversity correlation may be driven by an
influence of pollinator diversity on the seed set of plants (Klein et al., 2003a; Fontaine
et al., 2006) and in turn, as pollinators are strongly dependent on floral diversity due to
mutual specialization, declines in plant diversity, e.g. caused by land use intensification,
may be associated with linked extinctions of pollinators (Frund et al., 2010).
Although the general extent of pollinator specialization is still poorly known
many findings highlight the complexity of plant-pollinator interactions and confirm the
importance of flower diversity for bee and hoverfly communities (Ambrosino et al.,
2006; Bosch et al., 2009). Since floral density and diversity are one of the most
important factors for conservation of pollinators (Scriven et al., 2013), importance of a
positive correlation between richness of flower species and pollinators has been
reported from several countries and habitats (Steffan-Dewenter and Tscharntke, 2000;
Frund et al., 2010; Nicholls and Altieri, 2013). For that reason, exposure of diverse
flowers to pollinators is a way of maintaining and promoting genetic diversity in crops
and other plants, and studies focusing on the behaviour of floral visitors are an

Chapter I
4
important tool to identify patterns of ecological interactions between visitors and
flowers. The majority of such ecological interactions are mutualistic, whereby plant
species offer floral resources and obtain pollination benefits. However, some relations
are unilateral, where plants can be used for other purposes such as for prey capture,
mating, and resting (Comba et al., 1999). One of interesting ecological relations are
those that involve the pillage of plant resources by floral visitors, i.e. the use of
resources by illegitimate visitation incapable of promote pollination (Inouye, 1980). For
those reasons it is important to know which insects are attracted to which floral species,
and also which insects are efficient in pollination.
3. Important groups of pollinators
The most recognized pollinators are various species of bees (Hymenoptera:
Apoidea), with their body structure plainly adapted to pollinate (Michener, 2007). Other
groups of insects important in pollination are: hoverflies (Diptera: Syrphidae); ants and
wasps (Hymenoptera); moths and butterflies (Lepidoptera) and some beetles
(Coleoptera).
This study has been mainly focused only on bees and hoverflies as those two
groups of pollinators were the most abundant and diverse in the study area. In addition,
several species of beetles were included in the study as they were highly abundant
visitors to some of the studied plant species. For these reasons, taxonomy and biology
of these three groups will be described, while other groups of pollinators will be
briefly introduced.
3.1. Bees (Hymenoptera: Apoidea)
Bees (Hymenoptera: Apoidea) are the most important pollinating insects as they
depend on flower nectar and pollen. Nectar is the main energy source for adults, while
pollen is a protein source, collected by adult females primarily to feed their larvae
(Michener, 2007). However, they are important pollinators because some of the
collected pollen that they inevitably lose in going from flower to flower falls on the
pistils (reproductive structures) of other flowers of the same species, resulting in cross-
pollination.

Chapter I
5
The bees have been classified by different authors following different criteria, but
in this research the classification proposed by Michener (2007) has been used, where
seven families of bees were set: Andrenidae, Colletidae, Halictidae, Melittidae,
Stenotritidae, Megachilidae and Apidae. Two of those families (Melittidae,
Stenotritidae) are small families, mainly distributed in Africa and Australia, while other
five families (Apidae, Halictidae, Colletidae, Andrenidae and Megachilidae) are
cosmopolitan families and numerous species of those families are present in Spain (Fig.
I-2).
Fig. I-2. Different groups of pollinators [1. Honey bee (Apidae: Apis sp.); 2. Bumblebee
(Apidae: Bombus sp.); 3. Carpenter bee (Apidae: Xylocopa sp.); 4. Digger bee (Apidae:
Amegilla sp.); 5. Leafcutter bee (Megachilidae: Megachile sp.); 6. Mining bee
(Andrenidae: Andrena sp.; 7. Sweat bee (Halictidae: Halictus sp.; 8. Hoverfly
(Syrphidae: Sphaerophoria scripta.); 9. Beetle (Coleoptera: Heliotaurus ruficollis)]
Nevertheless, in pollination ecology it is frequent to group bees in accordance
with their morpho-physiological features to function as pollinators, such as body size

Chapter I
6
and method of pollen collection. Therefore, honey bees, bumblebees and wild bees are
in general analyzed separately (Carrie et al., 2012; Miñarro and Prida, 2013; Bartomeus
et al., 2014) and sometimes, depending on the scope of the study, wild bees can also be
divided into different groups (Barbir et al., 2014a) easily distinguished in the field. In
continuation, bee groups that are frequent in Central Spain will be briefly described.
Honey bees
Honey bees (Family: Apidae) represent only a small fraction of the approximately
20,000 known species of bees and are a subset in the genus Apis, primarily
distinguished from other species by the production of honey and the construction of
nests out of wax. Currently, there are only seven recognized species of honey bee with a
total of 44 subspecies (Engel, 1999).
Honey bees (as well as bumble bees) are social insects, with a firmly specialized
division of labour (Johnson, 2010). Besides the important role in producing honey,
honey bees have the important ecological and economical role of pollination, for which
their hind legs are modified into a structure called corbicula, widely known as the
"pollen basket" (Fig. I-2.1). Economically, for all USA agriculture, the marginal
increase in the value attributable to honey bees—that is, the value of the increased yield
and quality achieved through pollination by honey bees alone—was $9.3 billion in 1989
and $14.6 billion in 2000 (Morse and Calderone, 2000).
Bumblebees
Another important genus from the family Apidae in pollination of plants is
Bombus (bumblebees). There are over 250 known species, existing primarily in the
Northern Hemisphere. Bumblebees are, like honey bees, social insects and are mostly
characterized by black and yellow body (Fig. I-2.2) while in some species it might be
orange, red or entirely black (Michener, 2007).
In Spain there are just twelve species of bumblebees of which the most abundant
is Bombus terrestris L. (Rasmont and Iserbyt, 2010). However, in other regions, e.g.
UK or USA, bumblebees are rather diverse and abundant, but this diversity is declining
rapidly and therefore conservation of bumblebees have received a lot of attention
© P
©

Chapter I
7
recently (Carvell et al., 2007; Heard et al., 2007). In general, bumblebees are good
pollinators due to their hairy body, but some species of bumblebee also exhibit what is
known as "nectar robbing", i.e. instead of inserting the mouthparts into the flower, they
bite directly through the base of the corolla to extract nectar, avoiding pollen transfer
(Maloof, 2001).
Solitary bees
Solitary bees (frequently called wild bees) are important pollinators, because of
their advanced types of pollen-carrying structures, i.e. the scopa (made up of thick,
plumose setae), on the lower abdomen (Fam. Megachilidae) or on the hind legs (other
families of wild bees).
Solitary bees are often oligoleges (or simply specialist pollinators), gathering
pollen from one or a few species/genera of plants (unlike honey bees and bumblebees
which are generalists). Due to their high specialization, some solitary bee species are
endangered, simultaneously with the plant species they are associated with (Biesmeijer
et al., 2006). In addition, solitary bees create nests in hollow reeds, holes in wood, or
most commonly, in tunnels in the soil, which are frequently destroyed by agricultural
tillage (Nicholls and Altieri, 2013).
Although solitary bees are important pollinators, a very few species of solitary
bees are reared for commercial pollination, e.g. Osmia ribifloris Cockerell, because of
their difficult management (providing nest sites). Instead, especially in organic
gardening, it became widely popular to provide nest boxes for solitary bees and in that
way improve the conservation of the native solitary bee species. In the landscape level,
conservation of pollinators mainly takes place within agro-ecosystems by managing the
habitat and offering additional floral sources to forage, with final aim to maintain
pollinators close to agricultural fields (New, 2005).
Among the solitary bees different groups can be found, such as:
Carpenter bees (Fam. Apidae: Xylocopini) are large bees which make their nests
by tunnelling into wood. These bees are important pollinators of open-faced flowers,
although some species are known to rob nectar by slitting the sides of flowers with deep

Chapter I
8
corollas (Zhang et al., 2007). One of the most abundant species of this group in Central
Spain is Xylocopa violacea L. (see Xylocopa sp. In Fig. I-2.3).
Digger bees (Fam. Apidae: Anthophorini) are very buzzy solitary bees which dig
their nests in the soil. This group consists of over 750 species with different shapes and
sizes, but the majority of bees are large (up to 3 cm), hairy, fast-flying and robust
(Michener, 2007). The most abundant genera in Spain are: Amegilla spp. (Fig. I-2.4),
Antophora spp. and Habropoda spp.
Leafcutter bees (Fam. Megachilidae) (Fig. I-2.5) are a cosmopolitan family of
solitary bees whose pollen-carrying structure is restricted to the ventral surface of the
abdomen and because of this they are easy to recognize in the field (Michener, 2007).
They also produce characteristic buzzy sound when flying. As they nest in the wood, it
is easier to commercially produce some of the species and to use them for pollination in
greenhouses (O. ribifloris). Some of the frequent species in Central Spain are:
Chelostoma florisomnis, Chalicodoma pyrenaica, Osmia spp., Anthidium spp.
Mining bees (Fam. Andrenidae) is a large non-parasitic bee family, with most of
the diversity in temperate and arid areas (warm temperate xeric). Therefore, this group
of bees is very diverse and abundant in the Mediterranean region. They are typically
small to moderate-sized bees and often have scopa on the basal segments of their legs in
addition to the tibiae. They can be separated from other bee families by the presence of
two subantennal sutures on the face (Michener, 2007). The most frequent genus in
Central Spain is Andrena with many different species (Fig. I-2.6).
Sweat bees (Fam. Halictidae) is a cosmopolitan family consisting mainly of
small to midsize bees (up to 10 mm), although few species can be bigger. Their hives
are usually found inside the hollow trunk or wood or underground. Several species are
completely or partly green and a few are red; a number of them have yellow markings,
especially the males, which commonly possess yellow faces, a pattern widespread
among the various families of bees (Engel, 2000). Two genera are the most abundant in
Spain: Halictus spp. (Fig. I-2.7) and Lasioglossum spp.
© J l

Chapter I
9
3.2. Hoverflies (Diptera: Syrphidae)
Hoverflies, also known as syrphid flies, are classified in the family Syrphidae
(Diptera), which consists of about 6,000 species. Hoverflies are an extremely variable
family of flies that range from the large, bulky and hairy to the small, slender, and shiny
adults which are easily recognized by their ability to hold a seemingly motionless
position in the air (Stubbs and Falk, 2002). The genera Sphaerophoria (Fig. I-2.8),
Eristalis, Episyrphus, Syrphus and Eupeodes are some of the most abundant genus of
the family Syrphidae in Central Spain (Pineda and Marcos-Garcia, 2008).
Many adult hoverflies survive on nectar and pollen, but the larvae eat a much
more varied diet that may include other insects or decaying plants and animals. For that
reason, some species of hoverfly are highly prized as biological control agents because
the larval forms of those flies feed on aphids (Chambers and Adams, 1986; Ambrosino
et al., 2006; Smith and Chaney, 2007). Therefore, it has been recommended to plant
flowers that produce pollen and nectar that adult hoverflies use as the energy to produce
large numbers of eggs all of which turn into aphid-munching larvae (Stubbs and Falk,
2002).
3.3. Beetles
Beetles (Coleoptera) are generally viewed as poor pollinators (Bartomeus et al.,
2008), although in Mediterranean ecosystems they are very abundant floral visitors
(Bernhardt, 2000) (Fig. I-2.9). On the other side, beetles spend long periods of time on
each flower they visit, and therefore they visit fewer flowers than bees (Bosch, 1992). In
addition, some beetles can occupy or destroy flowers, not allowing other pollinators to
feed on the flowers (Barbir et al., 2014a). In this study, only the most abundant flower
visitors have been included.
3.4. Other pollinator groups
Butterflies and moths are the second largest order of insects (Lepidoptera)
however, they are not major pollinators. Butterflies and moths tend to sit at the edges of
flowers, and extend their long tongues (probosces) to reach nectar (Subba Reddi and
Meera Bai, 1984). They rarely come in contact with the pollen at the centres of the
©Jelena

Chapter I
10
flowers. Nevertheless, some moths are effective pollinators of deep-throated flowers
that require the insects to crawl inside to reach the nectar.
In addition, ants and wasps (Hymenoptera) are also insects that frequently visit
flowers and contribute to their pollination, but they are not as efficient pollinators as
bees (Philpott et al., 2006).
4. Managing semi-natural habitats of pollinators within agro-
ecosystems
Since agricultural practices can damage biodiversity through various pathways,
agriculture is frequently considered anathema to conservation. However, appropriate
management can ameliorate many of the negative impacts of agriculture, while largely
maintaining provisioning ecosystem services (Power, 2010). Although honey bee has
high economic importance in honey production and it is the most abundant
cosmopolitan pollinator, it has been discovered that there are various crops for which
honey bees are poor pollinators compared to wild bees (Michener, 2007). For that
reason, national and international organizations are publicizing the need to conserve
native pollinators (Michener, 2007).
Although wild pollinators are better conserved within protected areas and natural
parks, where floral diversity and shelters are available, for agricultural production it is
more convenient to conserve pollinators within agricultural fields. Conventional farmers
routinely manage their fields for greater provisioning services by using inputs and
practices to increase yields, but management practices can also enhance other ecosystem
services, such as pollination, biological control, soil fertility and structure, and support
biodiversity (Power, 2010). Therefore, if introduced properly, habitat management
within the agro-ecosystem can provide resources needed for conservation of pollinators
and natural enemies (Tscharntke et al., 2005).
Large monocultures of bee-pollinated crops, i.e. almond, melon, or canola,
normally provide abundant food sources for pollinators, but only for a few weeks
period. Therefore, lack of within field or adjacent wild plants, blooming before and after
the main crops bloom, can result in a decline of healthy pollinators abundance
(Goulson, 2003). Recently, implementation of floral margins in agricultural landscapes

Chapter I
11
has been shown to increase the abundance of pollinators in agro-ecosystems (Kells et
al., 2001; Marshall, 2005). Naturally regenerated uncropped margins were primarily
designed to conserve rare local arable flora and their associated fauna (Kells et al.,
2001). However, sowing an uncropped margin with wildflower seed mixture can offer
better food sources to pollinators and improve their conservation (Pywell et al., 2011).
Therefore, in order to maintain native plant species in each region for pollinator
conservation, there is a need for pollination biologists to integrate their findings with
those of plant restoration ecologists and to ensure sustainable restoration of pollinator
habitats within agro-ecosystems by using native plants (Menz et al., 2011). For that
reason, special attention should be focused on the selection of beneficial insectary plants
and their management in the agro-ecosystems (Nicholls and Altieri, 2013).
4. 1. Beneficial insectary plants
Beneficial insectary plants are plants that attract beneficial insects (pollinators and
natural enemies of pests) by producing nectar and pollen which are their main food
sources (Colley and Luna, 2000). Although many pollinators are generalist, e.g. honey
bee and the majority of hoverfly species (Branquart and Hemptinne, 2000; Frund et al.,
2010), some pollinators have preferences when choosing the plants to visit (Frund et al.,
2010).
Pigmentation in flowers seems to play a major role in pollination success (Miller
et al., 2011) and can significantly influence the insects’ preference (Campbell et al.,
2012b). In addition, colour can help beneficial insects with plant species selection,
flower location and ripeness, and nectar and pollen location within the flower. Besides,
shape and size of the flower (Gomez et al., 2008; Schiestl and Johnson, 2013), nectar
availability and chemical composition of the plant species (Baker and Baker, 1983;
Stout and Goulson, 2002) also contribute to the preferences of pollinators.
Some plant families have been widely known as highly attractive to beneficial
insects, for instance Boraginaceae, Brassicaceae and Apiaceae, and many plant species
from those families have been introduced within agricultural fields worldwide. Plants as
buckwheat (Fagopyrum esculentum Moench) (Fig. I-3.1), phacelia (Phacelia
tanacetifolia Benth.) (Fig. I-3.2), alyssum (Lobularia maritima L. Desv.) (Fig. I-3.3)

Chapter I
12
and coriander (Coriandrum sativum L.) (Fig. I-3.4), have been used in beneficial insect
conservation practices in the last 10 years (Landis et al., 2000; Pfiffner and Wyss,
2004), where their attractiveness to beneficial insects have been highly ranked on the
basis of feeding visit frequencies (Ambrosino et al., 2006).
Fig. I-3. Beneficial insectary plants [1. Buckwheat (Fagopyrum esculentum Moench);
2. Phacelia (Phacelia tanacetifolia Benth.); 3. Alyssum (Lobularia maritima L. Desv.);
4. Coriander (Coriandrum sativum L.)]
While it is important to consider all the factors mentioned above when choosing
plants to attract beneficial insects, it is also necessary to consider the ecology,
physiology, floral phenology, and potential weediness of the plants prior introducing
them in agro-ecosystems (Nicholls and Altieri, 2013), with the final aim to apply the
most appropriate and sustainable habitat management for conservation of pollinators.
© Aldo De Bastiani © Jelena Barbir
© Jelena Barbir © H. Zell
1 2
3 4

Chapter I
13
4. 2. Management of beneficial insectary plants within agro-ecosystems
Although some plants can be widely attractive to cosmopolitan pollinators, other
plants have flowers that are more attractive to specific groups of pollinators, e.g. open
flowers are attractive to hoverflies and plants from Boraginaceae family are very
attractive to bumblebees. Therefore, it is important to have fundamental knowledge
about the abundance and diversity of groups of pollinators in the region before choosing
plant species for efficient management of pollinator habitats within the landscapes.
In addition, to manage beneficial insectary plants within agro-ecosystems it is
essential to consider the climate conditions of the region (Memmott et al., 2010). In the
Mediterranean region, where precipitation and irrigation water are scarce, usage of plant
species that can cope with water stress are preferred, while in other regions, i.e. tropical,
Central and Northern Europe, irrigation is not a limiting factor. Under different
environmental conditions, beneficial insectary plants easily managed in one region can
show weediness or invasiveness in another region. Therefore, it is recommendable to
test under field conditions the agricultural behaviour and the attractiveness to pollinators
of introduced or native beneficial plants (Comba et al., 1999; Corbet et al., 2001) before
recommending its usage in specific agro-ecosystems.
Furthermore, beneficial insectary plants can be managed in different ways within
agro-ecosystems depending on the scale of the field, climate and agricultural practices
of specific region (Altieri, 1991). One of the most common approaches to implement
beneficial plants within large agricultural fields is to sow field margins. However, some
plant species are more efficient if sown as mono-specific margins (Fig. I-4.1), while
others are more efficient if mixed with other plant species (Fig. I-4.2) (Pontin et al.,
2006; Barbir et al., 2014a).

Chapter I
14
Fig. I-4. Sowed field margins: 1. Mono-specific field margin (Phacelia tanacetifolia
Benth.), 2. Mixed field margin
In addition, sustainable and organic agriculture have recently received a lot of
attention as strategies that will stop or at least reduce the damage to ecosystems
(Andersson et al., 2012; Crowder et al., 2012) and improve conservation of beneficial
insects (Power and Stout, 2011; Gill et al., 2012). In organic farming, beneficial
insectary plants are widely used for their positive impact in conservation of natural
enemies of pests and pollinators (Power and Stout, 2011). Since the commercialization
of organic products is becoming increasingly widespread, this implies an
unquestionable opportunity for organic production on a small-scale, where the use of
beneficial insectary plants should be a key part of the production system (Fig. I-5)
(Markussen et al., 2014).
Fig. I-5: Use of beneficial insectary plants in small-scale gardening
©OrganicGarden ©The Climate Compliannce Conference

Chapter II
15
II. Research question and objectives
There is a need to improve habitat management to promote the conservation of
pollinators in agro-ecosystems from Central Spain. Considering the specific
characteristics in term of climate, native flora, agricultural practices, pollinator species
and their habitats within different regions worldwide, it is important to find the most
appropriate beneficial plants in a specific region and to manage them in the most
suitable way. For this, the following research question should be posed:
How to manage the habitat of pollinators in order to conserve them
within agro-ecosystems of Central Spain?
In order to answer the research question and to conduct the experiments
systematically, several specific objectives have been proposed:
Objective 1
to improve the functionality of a mixture of aromatic plants (Fam. Lamiaceae) in
order to conserve pollinators (Chapter III)
Objective 2
to assess the preferences of pollinators to different species of annual herbaceous
plants in order to use them for their conservation in agro-ecosystems of Central
Spain (Chapter IV)
Objective 3
to evaluate the agronomic behaviour, i.e. potential weediness and self-seeding,
of different aromatic and herbaceous plant species based on their biological,
physiological and morphological characteristics (Chapter III and IV)

Chapter II
16
Objective 4
to study the effects of drought and regulated deficit irrigation (RDI) on growth
and potential attractiveness of the plant selected as the most appropriate to
conserve pollinators in Central Spain (Chapter V)
Objective 5
to test if the proximity of mono-specific or mixed field margins can improve
pollination in the neighbouring crop (Chapter VI)

CHAPTER III
Submitted to: Insect Conservation and Diversity Journal (Submitted: 06/06/2014)
Barbir, J., Azpiazu, C., Badenes-Peréz, F.R., Fernández-Quintanilla, C., Dorado,
J. (submitted). Functionality of aromatic (Lamiaceae) floral mixture in conservation of
pollinators in Central Spain, Insect Conservation and Diversity

Chapter III
18
III. Functionality of aromatic (Lamiaceae) floral mixture in
conservation of pollinators in Central Spain
1. Introduction
In the Mediterranean region, medicinal and aromatic plants (in further text
referred only to family Lamiaceae) are broadly distributed in agro-ecosystems as both
cultivated and spontaneous (Carrubba and Scalenghe, 2012). Besides being
commercially used by food, perfume and pharmaceutical industries (Hussain et al.,
2011; Zihlif et al., 2013), those plants have good potential as a forage source for
pollinators and other beneficial insects (Song et al., 2010; Macukanovic-Jocic et al.,
2011). As such, native aromatic species could play a significant role in the enhancement
of multi-functionality of agriculture (Carrubba and Scalenghe, 2012), since
environmentally friendly farming practices are mandatory in the European Union.
Furthermore, use of aromatic plants could improve the conservation of beneficial insects
and their natural habitat within agro-ecosystems.
Agricultural landscapes, dominated by large monocultures, have caused alarming
declines in farmland biodiversity, threatening the optimal functionality of agro-
ecosystems (Krebs et al., 1999; Benton et al., 2003). As reported by Landis et al. (2000)
the presence of wild plants in crop ecosystems enhances the survivorship of beneficial
insects responsible for ecosystem services such as pollination and biological control of
pests in both agricultural and natural habitats (Klein et al., 2007). Thus, the recent report
of parallel decline of pollinators and insect-pollinated plants (Biesmeijer et al., 2006)
encouraged further investigations in plant-pollinator interactions (Campbell et al.,
2012a; Carrie et al., 2012; Wratten et al., 2012) and habitat management (Cranmer et
al., 2012; Holzschuh et al., 2012; Wratten et al., 2012) to enhance the presence of
beneficial insects within agro-ecosystems.
In the intensively managed agro-ecosystems, insect species tend to suffer from
deficit of nectar and pollen sources, shelter and nesting sites (Cane, 2008; Pywell et al.,
2011; Carvalheiro et al., 2012). Therefore, some recent studies have focused on
improving the habitat of beneficial insects within agricultural landscapes by sowing
field margins with a mixture of wild flowers (Marshall, 2005; Carvell et al., 2007;

Chapter III
19
Olson and Wackers, 2007). For that purpose, some studies recommend plant species
that are attractive to pollinators (Pontin et al., 2006), while others suggest the usage of
native or naturalized plants in the area (Tuell et al., 2008; Morandin and Kremen, 2013).
In addition, the importance of adjusting the floral phenologies of the plants used in the
field margins has been emphasized, by combining their different bloom periods and
providing pollinators with constant floral resources available from early spring to early
autumn (Memmott et al., 2010). Although the attractiveness to pollinators of many
aromatic species has been combined with their floral phenology and morphology
(Herrera, 2001; Keasar et al., 2006), only few studies have simultaneously compared the
attractiveness of various aromatic plant species under the same field conditions
(Petanidou and Vokou, 1993; Macukanovic-Jocic et al., 2011). Some studies have
emphasized that the beneficial plants should not be invasive towards neighbouring crop
fields (Stout and Morales, 2009), but in the Mediterranean region little is known about
the behaviour of beneficial plants in floral mixtures as well as their competitiveness
towards neighbouring crops.
Considering that many native aromatic species are naturally present in the
landscapes, hedgerows and margins of crop fields, there is a need to compare their
attractiveness to pollinators and to study their role in floral mixtures in order to choose
the most efficient plants for functional field margins. Therefore, it was hypothesized
that mixture of plants attractive to pollinators and with different floral phonologies, can
create a functional, long-flowering floral margin which will offer food for pollinators
during their active season. Consequently, six plant species from the Lamiaceae family,
with different floral phenologies, frequent and native in the Mediterranean region, were
selected for this study by following three main objectives: i) to compare their
attractiveness to pollinators, ii) to investigate whether floral morphology affects the
attractiveness; and iii) to evaluate the functionality of the aromatic plants grown in
mixture by taking into account different phenologies and attractiveness to pollinator.
Finally, as the purpose of the studied plants is to be implemented within agro-
ecosystems, their spatial expansion, i.e. potential weediness, was estimated under field
conditions.

Chapter III
20
2. Materials and methods
2.1. Study area and experimental design
This study has been conducted over three years at the experimental farm La
Poveda (40º 19' N and 3º 29' W, elevation 536 MASL), Arganda del Rey, Madrid,
Spain. The study area is characterized by a continental Mediterranean climate with cold
winters and hot summers and scant precipitation (about 400 mm per year). The
meteorological data for both years of study was obtained from the meteorological
station located at a distance of 1 km from the experiment, available online at Agro-
climatic Information System for Irrigation (SIAR, 2014).
In February 2011, three-month-old seedlings of six aromatic species (Lamiaceae)
were organised in randomized block design experiment (24.0 m × 15.2 m) within a
barley field, with the longer side orientated NE-SW. The experiment was divided into
three blocks (repetitions) each consisting of eight plots (2.4 m × 2.4 m) including six
mono-specific plots and two plots (M1 and M2) with a mixture of these six species
equally proportioned and randomly distributed within a plot (Fig. III-1).
Fig. III-1. Experimental design (randomized block design - each number represents one
Lamiaceae plant species (mono-specific plot), while M1 and M2 represent mixed plots)
1 6
5 4
3 M1
M2 2
M2 4
6 1
M1 5
3 2
1 5
3 M1
M2 4
2 6

Chapter III
21
Each plot consisted of 64 plants organized in 8 lines and 8 columns, with a
separation of 0.3 m between them. Finally, in order to avoid the influence of the
neighbouring species, a separation between plots was 1.2 m and between blocks 2.0 m.
2.2. Groups of pollinators
Insects were grouped in accordance with their morpho-physiological features to
function as pollinators, such as body size and method of pollen collection (Barbir et al.,
2014a). The study was focused on seven groups of insects easily distinguished in the
field: i) honey bees (Apis mellifera L.), ii) leafcutter bees (Fam. Megachilidae), iii)
digger bees (Fam. Apidae, tribe Anthophorini), iv) small solitary bees (Fam. Halictidae
and Andrenidae, with body size ≤ 1cm), v) big solitary bees (Fam. Halictida e and
Andrenidae, with body size > 1cm), vi) hoverflies, and vii) beetles. Bumblebees,
Bombus spp. (Apidae) and carpenter bees, Xylocopa violacea L. (Apidae) were not
included in the final analysis as fewer than ten visits were recorded during the whole
study.
2.3. Attractiveness of the plants to pollinators
In order to estimate the attractiveness of aromatic flowers to local pollinators,
their visitation rate, i.e. the mean number of insect visits per minute (Campbell et al.,
2012a), was compared between plant species and their mixtures during 2012 and 2013.
Insect visits to the flowers (direct contact with the reproductive structures of the flower,
i.e. stamens and pistils) were counted once per week (9 min/plot) within three marked
areas (0.25 m × 0.25 m) randomly scattered within each plot. The observations were
conducted by three observers throughout the flowering period of each species, always in
the morning (9.00 h – 13.00 h local time) when the weather conditions were preferable
for the insects to fly and visit flowers (sunny days without wind). Switching plots, each
observer counted visits of insects for three minutes in each plot.
2.4. Floral assessment
As one of the objectives of the study was to create a functional long flowering
mixture of aromatic species, floral phenology was one of the main criteria for plant
selection. For that purpose, six aromatic species (Fam. Lamiaceae), native and frequent

Chapter III
22
in ecosystems of Central Spain, were selected for this study (Table III-1) in order to
assess their potential contribution in floral mixtures.
Table III-1. Aromatic plant species used (taken from Castroviejo (1993))
Plant species Plant height (cm)
Flowering period Habitat Use
Salvia verbenaca L. 5 – 60 april -may
grasslands, cultivated fields,
roadsides, wastelands
medical
Salvia officinalis L. < 60 may -june cultivated or sub-spontaneous food and medical
Nepeta tuberosa L. 30 – 100 may - june
juniper and oak woods, thickets,
roadsides, meadows, grasslands
ornamental and medical
Melissa officinalis L. 45 – 83 june - july nitrified, shady and cool places
food, medical and perfume industry
Thymbra capitata (L.) Cav. 10 – 40 june - july rocky slopes,
limestone, marl or clay soils
food, medical and perfume industry
Hyssopus officinalis L. 15 – 52 july - octobre thickets, roadsides medical
In order to understand why some plant species can be more attractive to
pollinators than others, morphological characteristics of flowers were assessed.
Morphological characteristics of flowers were measured at the full bloom phenological
stage in ten randomly chosen flowers, using a Vernier caliper. Immediately after cutting
off the flower, certain morphological characteristics (flower length, flower height,
corolla tube length, corolla height and calyx length) were measured, and the mean value
of those measurements was used in further analysis.
The capacity of plants to produce flowers was assessed. For this, the number of
flowers was counted in three frames of 0.25 m × 0.25 m within each plot once per week.
In addition, each observer estimated the percentage of flower coverage in each plot, and
the mean value of the three readings was calculated. Both parameters were used to
estimate floral density as well as floral phenology of each plant species under the field
conditions, with the final aim to estimate their influence on the attractiveness of the
plants (Stout et al., 1998). Any plot was considered in bloom when it had more than five
open flowers per observed area.

Chapter III
23
2.5. Spatial expansion of the plant species
In the early spring of 2014, the spatial expansion from the original area (i.e. the
percentage of increased area) of each plant species, i.e. the three replicates of each
mono-specific plots (2.40 m × 2.40 m) and the proportional part (i.e. one-sixth) of all
the mixed plots, was calculated considering the area occupied the following year by
using real time kinematic differential GPS (RTK-GPS). The percentage increase in area
occupied by each plant species relative to their original area (i.e. potential weediness)
was calculated using the software ArcGIS® 9.3 (ESRI, Redlands, California, USA).
2.6. Data analysis
The data from 2012 and 2013 were independently analysed with SPSS®
Statistics19 software (IBM, Chicago, IL, USA). A simple general linear model (GLM)
was used in order to assess the ability of different insects to discriminate between
flower species. The mean value of insect visits per minute within the observation area
(0.25 m × 0.25 m) was used as response variable for these analyses. Prior to ANOVA
analysis, the assumptions of normal distribution (Kolmogorov Smirnov test) and
homogeneity of variances (Levene’s test) were tested. Differences among means were
explored by Tukey’s B post hoc test (P ≤ 0.05). In addition, univariate ANOVA was
carried out to assess if there were significant differences between measured
morphological characteristics of the flowers.
Discriminate Canonical Analysis (DCA) was performed in order to test if any of
morphological characteristics influence on the discrimination between selected plants,
and number of dimensions necessary to express this relationship. Finally, Categorical
Partial Component Analysis (CATPCA) was used to assess the relationship between
morphological characteristics and number of insect visits per minute. A bivariate two-
tailed Pearson analysis was used to assess the correlation between floral density and
visits of pollinators per minute.

Chapter III
24
3. Results
3.1. Attractiveness of the selected aromatic plants to pollinators
The attractiveness of the plants to pollinators was estimated by insect visitation
rate (Table III-2). Nepeta tuberosa and Hyssopus officinalis, which had the highest
visitation rate of pollinators in both years of the study, were characterized as the most
attractive plant species. On the other hand, Salvia verbenaca, with no insect visits in
2012, and few beetles, digger bees and hoverflies visits in 2013, was characterized as
the least attractive plant species (Table III-2).

Chapter III
25
Tab
le II
I-2.
Mea
n (±
SE)
inse
ct v
isits
per
min
ute
to fl
ower
s of s
elec
ted
arom
atic
pla
nt sp
ecie
s in
2012
and
201
3
Mea
ns w
ith th
e sa
me
lette
r in
each
row
are
not
sign
ifica
ntly
diff
eren
t at P
≤ 0
.05
as d
eter
min
ed b
y Tu
key’
s B m
ultip
le c
ompa
rison
test
.
Yea
rS.
ver
bena
caS.
off
icin
alis
N. t
uber
osa
M. o
ffic
inal
isT.
cap
itata
H. o
ffic
inal
isM
1M
2
Hon
ey b
ees
2012
0±
0c0.
05±
0.02
abc
0.07
±0.
02ab
0±
0c0.
01±
0.01
c0.
14±
0.05
a0.
02±
0.01
bc0.
04±
0.01
abc
2013
0±
0b0
±0b
0.04
±0.
02a
0±
0b0.
01±
0.01
b0.
03±
0.01
b0.
01±
0.01
b0.
01±
0.01
b
Dig
ger
bees
2012
0±
0c0.
01±
0.01
c0.
09±
0.02
a0.
02±
0.01
c0
±0c
0.04
±0.
01b
0±
0c0.
01±
0.01
c
2013
0.02
±0.
02b
0.02
±0.
01b
0.09
±0.
03a
0.01
±0.
01b
0±
0b0.
05±
0.02
ab0.
03±
0.01
ab0.
02±
0.01
b
Leaf
cutt
er b
ees
2012
0±
0b0.
02±
0.01
b0.
12±
0.03
a0.
04±
0.01
ab0.
03±
0.02
ab0.
12±
0.03
a0.
03±
0.01
b0.
04±
0.01
b
2013
0±
0b0
±0b
0.09
±0.
03ab
0.03
±0.
02b
0.01
±0.
01b
0.20
±0.
04a
0.02
±0.
01b
0.03
±0.
01b
Smal
l sol
itary
bee
s20
120
±0c
0.28
±0.
05a
0.14
±0.
02ab
0.15
±0.
04ab
c0.
29±
0.07
a0.
37±
0.09
a0.
05±
0.01
c0.
08±
0.02
bc
2013
0±
0d0.
01±
0.01
d0.
06±
0.02
bcd
0.22
±0.
07b
0.55
±0.
10a
0.18
±0.
03bc
0.05
±0.
01d
0.06
±0.
01bc
d
Big
solit
ary
bees
2012
0±
0c0.
12±
0.03
b0.
44±
0.06
a0.
02±
0.01
c0.
06±
0.02
bc0.
04±
0.02
bc0.
09±
0.02
bc0.
12±
0.03
bc
2013
0±
0a0.
03±
0.03
a0.
06±
0.02
a0
±0a
0.05
±0.
02a
0.08
±0.
02a
0.03
±0 .
01a
0.02
±0.
001a
Hov
erfli
es20
120
±0b
0±
0b0.
03±
0.01
a0.
01±
0.01
ab0.
01±
0.00
ab0.
04±
0.02
a0.
01±
0.00
ab0.
01±
0.01
ab
2013
0.07
±0.
04ab
0.10
±0.
03ab
0.17
±0.
04a
0.01
±0.
010.
08±
0.03
bc0.
10±
0.03
ab0.
05±
0.01
bc0.
07±
0.01
bc
Beet
les
2012
0±
0b
0.14
±0.
05a
0±
0b
0±
0b0
±0b
0±
0b0.
01±
0.01
b0.
01±0
.01b
2013
0.06
±0.
04b
0±
0b0.
29±
0.11
a0
±0b
0±
0b0.
01±
0.01
b0.
03±
0.01
b0.
03±
0.01
b
Polli
nato
rs (t
otal
)20
120
±0c
0.61
±0.
11a
0.93
±0.
11a
0.24
±0.
05b
0.39
±0.
09b
0.74
±0.
12a
0.20
±0.
03b
0.31
±0.
4b
2013
0.15
±0.
04c
0.15
±0.
04b
0.83
±0.
14a
0.27
±0.
07b
0.69
±0.
14a
0.64
±0.
08a
0.23
±0.
03b
0.23
±0.
03b

Chapter III
26
In 2012, Salvia officinalis was highly attractive to small solitary bees (F6,363 =
8.98, P ≤ 0.01) and beetles (F6,363 = 11.56, P ≤ 0.01), while in 2013 was attractive only
to hoverflies (F7,231 = 7.40, P ≤ 0.01). The flowers of N. tuberosa showed high
attractiveness to digger bees (2012: F6,363 = 15.73, P ≤ 0.01; 2013: F7,231 = 3.79, P ≤
0.01), leafcutter bees (2012: F6,363 = 5.75, P ≤ 0.01; 2013: F7,231 = 10.17, P ≤ 0.01) and
hoverflies (2012: F6,363 = 3.25, P ≤ 0.05; 2013: F7,231 = 7.40, P ≤ 0.01) in both years of
the study. In 2012, N. tuberosa was attractive to small solitary bees (F6,363 = 8.98, P ≤
0.01) and big solitary bees (F6,363 = 18.23, P ≤ 0.01), and in 2013 was the most
attractive plant species to honey bees (F7,231 = 3.06, P ≤ 0.05) and beetles (F7,231 = 3.79,
P ≤ 0.01). In general, Melissa officinalis was not attractive to pollinators in both years
of the study. The most frequent visitors to M. officinalis flowers were leafcutter bees
(F6,363 = 5.75, P ≤ 0.01) and small solitary bees (F6,363 = 8.98, P ≤ 0.01), but only in
2012. Small solitary bees were the most frequent visitors in the plots of Thymbra
capitata in both years, 2012 (F6,363 = 8.98, P ≤ 0.01) and 2013 (F7,231 = 16.75, P ≤ 0.01).
In addition, H. officinalis was highly attractive to leafcutter bees (2012: F6,363 = 5.75, P
≤ 0.01; 2013: F7,231 = 10.17, P ≤ 0.01) in both years of the study, while in 2012 was
frequently visited by honey bees (F6,363 = 5.22, P ≤ 0.01), small solitary bees (F6,363 =
8.98, P ≤ 0.01) and hoverflies (F6,363 = 3.25, P ≤ 0.05), and in 2013 by digger bees
(F7,231 = 3.79, P ≤ 0.01). On the other hand, when compared with the mono-specific
aromatic plots, both mixed plots (M1 and M2) showed low attractiveness to all groups
of pollinators, although the diversity of insects was high (Table III-2).
3.2. Relationship between morphological characteristics of the flowers and
insect visitation
Although the results showed statistically significant differences among plant
species for each of the measured morphological flower characteristics (Table III-3), the
results of DCA showed that only two of the studied morphological characteristics were
significant for plant discrimination (Table III-4).
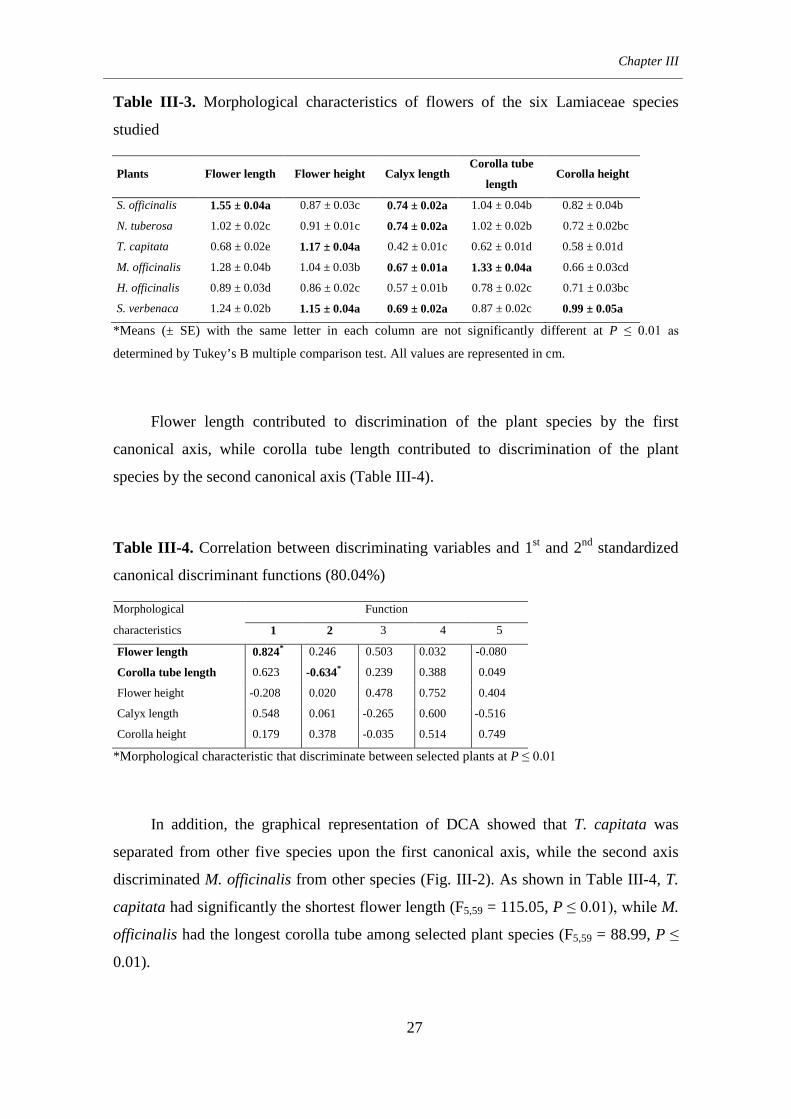
Chapter III
27
Table III-3. Morphological characteristics of flowers of the six Lamiaceae species
studied
Plants Flower length Flower height Calyx length Corolla tube
length Corolla height
S. officinalis 1.55 ± 0.04a 0.87 ± 0.03c 0.74 ± 0.02a 1.04 ± 0.04b 0.82 ± 0.04b
N. tuberosa 1.02 ± 0.02c 0.91 ± 0.01c 0.74 ± 0.02a 1.02 ± 0.02b 0.72 ± 0.02bc
T. capitata 0.68 ± 0.02e 1.17 ± 0.04a 0.42 ± 0.01c 0.62 ± 0.01d 0.58 ± 0.01d
M. officinalis 1.28 ± 0.04b 1.04 ± 0.03b 0.67 ± 0.01a 1.33 ± 0.04a 0.66 ± 0.03cd
H. officinalis 0.89 ± 0.03d 0.86 ± 0.02c 0.57 ± 0.01b 0.78 ± 0.02c 0.71 ± 0.03bc
S. verbenaca 1.24 ± 0.02b 1.15 ± 0.04a 0.69 ± 0.02a 0.87 ± 0.02c 0.99 ± 0.05a
*Means (± SE) with the same letter in each column are not significantly different at P ≤ 0.01 as
determined by Tukey’s B multiple comparison test. All values are represented in cm.
Flower length contributed to discrimination of the plant species by the first
canonical axis, while corolla tube length contributed to discrimination of the plant
species by the second canonical axis (Table III-4).
Table III-4. Correlation between discriminating variables and 1st and 2nd standardized
canonical discriminant functions (80.04%)
Morphological Function
characteristics 1 2 3 4 5
Flower length 0.824* 0.246 0.503 0.032 -0.080
Corolla tube length 0.623 -0.634* 0.239 0.388 0.049
Flower height -0.208 0.020 0.478 0.752 0.404
Calyx length 0.548 0.061 -0.265 0.600 -0.516
Corolla height 0.179 0.378 -0.035 0.514 0.749
*Morphological characteristic that discriminate between selected plants at P ≤ 0.01
In addition, the graphical representation of DCA showed that T. capitata was
separated from other five species upon the first canonical axis, while the second axis
discriminated M. officinalis from other species (Fig. III-2). As shown in Table III-4, T.
capitata had significantly the shortest flower length (F5,59 = 115.05, P ≤ 0.01), while M.
officinalis had the longest corolla tube among selected plant species (F5,59 = 88.99, P ≤
0.01).
Chapter III
28
Fig. III-2. Discrimination among plant species upon the first and second canonical axis
Figure III-3 shows the two-dimensional scatterplot of CATPCA analysis, relating
the morphological features of flowers and the visits of the different groups of
pollinators.

Chapter III
29
Fig. III-3. Relationship between morphological characteristics and number of insect
visits in 2012 (A) and in 2013 (B)

Chapter III
30
The figure shows that beetles and hoverflies are closest to the centroid and distant
from the two principal components, meaning that their contribution to the analysis is
low. Although DCA showed that some floral characteristics discriminate between plant
species, CATPCA showed that there is no correlation between insect visitation and
morphological characteristics of the flowers. Therefore, the fact of no clustering
between floral characteristics and insect visits emphasized that those two traits are not
related (Fig. III-3).
3.3. Relationship between floral phenology and insect visitation
In early spring 2012 mean daily temperatures were below 15 ºC and no pollinators
visited the flowers of S. verbenaca (Fig. III-4).

Chapter III
31
Fig. III-4. Relationship between floral phenology and insect visitation

Chapter III
32
In May, when the mean temperatures increased, S. officinalis and N. tuberosa
started to bloom, and the peaks of floral density and insect visits overlapped at the end
of May (S. officinalis) and beginning of June (N. tuberosa), respectively. The peaks of
floral density and the insect visits overlapped in mid-June for M. officinalis, and at the
beginning of July 2012 for T. capitata. Finally, the peak of floral density in H.
officinalis occurred at the end of September, while the insect visits had two peaks, the
first at the end of August and the second at the end of September. Both mixtures (M1
and M2) started to bloom at the beginning of May, reaching their peaks of floral density
and insect visits at the beginning of June. In the summer period (July-August) both traits
decreased until September, when they reached their second peaks (Fig. III-4).
In early spring 2013 mean daily temperatures were higher than the previous year
and pollinators visited the flowers of S. verbenaca. However, May 2013 was rainy and
cool (with the mean temperatures around 15 oC). This cool spring favoured the high
floral density of S. officinalis and N. tuberosa, but it led to low frequency of insect visits
(Fig. III-4). At the beginning of June the mean temperature started to increase and
stayed between 20 and 30 oC until September, when it started to decrease again. The
curves of floral density and insect visits for M. officinalis and T. capitata in 2013
showed similar tendencies as in 2012. However, in 2013 both the insect visits and the
floral density of H. officinalis reached one peak in mid-September, but the abundance of
insects was lower than the previous year. Similarly to 2012, both mixtures (M1 and M2)
started to bloom in May 2013, reaching the peaks of floral density and insect visits at
the beginning of June. In the summer period (July-August), both traits reduced their
values approximately to zero, and at the end of September reached their second peak
(Fig. III-4). In both years of study, the results showed a strong positive correlation
between floral density and number of insect visits (year 2012: r = 0.748, n = 608, P ≤
0.01; year 2013: r = 0.661, n = 648, P ≤ 0.01).
3.4. Spatial expansion of the plant species
The results of spatial expansion from 2014 showed that of the six plant species
included in the experiment, only two, S. verbenaca and N. tuberosa, expanded from
their original plots planted in 2011 (Table III-5).

Chapter III
33
Table III-5. Spatial increase (%) of the area occupied by plant species one year after
planting
Plant species Initial area in
2011 (m2)
Area in 2014
(m2)
Spatial increase of
initial area (%)
S. verbenaca 23.04 32.17 40
S. officinalis 23.04 23.04 0
N. tuberosa 23.04 31.39 36
M. officinalis 23.04 17.27 -25
T. capitata 23.04 23.04 0
H. officinalis 23.04 23.04 0
In contrast, the area occupied by M. officinalis was reduced by 25% in
comparison with its initial area. Other three plant species (S. officinalis, T. capitata and
H. officinalis) maintained their area within the initial plots during 3 years of the study
(Table III-5). None of the plant species intruded in the neighbouring barley fields, thus
showing no weediness or danger to invade the crop.
4. Discussion
The selected aromatic plant species attracted a great diversity of bees, while
hoverflies were in general less frequent visitors. Nevertheless, hoverflies are described
as less effective pollinators of Lamiaceae species than bees (Garcia, 2000). However,
results of both years of study showed clear differences in the floral attractiveness to
pollinators among aromatic flower species, where N. tuberosa and H. officinalis were
more attractive to local pollinators than S. verbenaca, M. officinalis and the mixtures
(M1 and M2). In addition, similarly to previous studies (Pontin et al., 2006),
preferences of the different groups of pollinators towards specific plants were detected.
In both years of study, T. capitata was mostly visited by small solitary bees. Nepeta
tuberosa and H. officinalis were mainly visited by leafcutter bees and hoverflies, while
digger bees preferred the flowers of N. tuberosa. Although honey bees are polylectic
and have been reported as the most frequent visitors and the most efficient pollinators of
H. officinalis, S. officinalis and M. officinalis (Macukanovic-Jocic et al., 2011), in this
study, honey bees were in general scarce, visiting only the flowers of H. officinalis and
N. tuberosa. Nevertheless, since the main purpose of field margins is the conservation

Chapter III
34
of beneficial insects, plants that attract mayor diversity of pollinators, like N. tuberosa
are preferable.
Although many studies have stated that the morphological characteristics of
flowers influence the plant’s attractiveness to insect visitors (Buide, 2006; Reith et al.,
2006; Reith et al., 2007), few studies have compared the morphology and the
attractiveness to pollinators of various aromatic species under the same conditions
(Petanidou and Vokou, 1993; Macukanovic-Jocic et al., 2011). This study shows that
only two morphological characteristics discriminated among plant species in terms of
their attractiveness to pollinators, i.e. flower length and corolla tube length. Thus, T.
capitata, visited mainly by short-tongued small solitary bees, was discriminated from
other plant species due to its short flowers. Conversely, despite the longest corolla tube
of M. officinalis, this species was rarely visited by the long-tongued bees and was
mainly visited by small solitary bees. Although this finding is in accordance with Engel
and Dingemansbakels (1980), the CATPCA analysis showed that morphological
characteristics and visits of pollinators were unrelated, suggesting that flower size does
not have a crucial influence on the attraction to pollinators. However, although
morphological characteristics of flowers did not influence the insect preference toward
different plant species, the results revealed a strong positive correlation between insect
visits and floral density, showing its important role in plant attractiveness (Dauber et al.,
2010; Scriven et al., 2013).
Low mean temperatures limited the presence of pollinators in the field and
consequently their visits to the flowers of S. verbenaca in April 2012, while April 2013
was warm and pollinator’s visits were observed. Although S. verbenaca was not
frequently visited by pollinators, it blooms in early spring when flowers in landscapes
are scarce, offering food source for pollinators before the crops flower (Nicholls and
Altieri, 2013). As such, the contribution of S. verbenaca may be important in a floral
mixture (Petanidou and Vokou, 1993). The flowering period of N. tuberosa and S.
officinalis coincided with high temperatures and high abundance of pollinators in the
field in 2012. Nevertheless, in 2013, May and June were cool and rainy, and the plants
in full bloom with high floral density were rarely visited by pollinators. Therefore, this
study emphasizes that weather conditions can limit the presence of pollinators in the
field, overriding the influence of floral phenology (Martin Gonzalez et al., 2009;
Abrahamczyk et al., 2011). However, as weather conditions can change from one year

Chapter III
35
to another, it is important to maintain floral sources available to pollinators from early
spring until late autumn.
Field margins containing a large floral diversity have become the focus of
numerous conservation efforts (Kells et al., 2001). In this study, although plant mixtures
allowed a longer blooming period than any of the individual stands, maximum flower
densities and insect visits were attained with some specific plant species. Several
reasons may explain this fact. Although mono-specific plots of S. officinalis and N.
tuberosa flower profusely and attract numerous pollinators in early spring, N. tuberosa
was significantly more attractive to all functional groups of pollinators than S.
officinalis. This probably was reflected on the lower relative attractiveness of the
mixture in the period of their flowering (May-June). During early summer (June-July),
flowering of M. officinalis and T. capitata in both mixed plots was considerably reduced
compared with their respective mono-specific plots. Although relatively high floral
density and insect visitation was observed in mono-specific plots of T. capitata, due to
its low height (20-30 cm), flowers were not available to pollinators when mixed with
taller plants, limiting its contribution to the mixture. During late-summer and early-
autumn, H. officinalis was the only plant species in the mixture with high floral density,
which contributed to the high attractiveness of the mixed plots. Even so, it is likely that
the growth and flowering capacity of this late growing plant was somewhat reduced by
competition from the other early growing species. In addition, it has to be considered
that floral density in the mixed plots has been lower than in mono-specific plots, due to
the mismatch in flowering periods of different plant species.
The plants assessed in this study are native and frequent in the study area, and
none of them showed a weedy behaviour nor intruded in the neighbouring crop. Most of
them maintained their original stands during three years, with the exception of the
reduction in the case of M. officinalis. Considering that perennial plants are
recommended as floral sources to pollinators in agro-ecosystems due to their low
maintenance (Tuell et al., 2008), the fact that the initial area of M. officinalis was
reduced may represent a limitation for its usage in floral margins. Therefore, a correct
plant selection is essential for the efficient functioning of plant mixtures (Nicholls and
Altieri, 2013).

Chapter III
36
The plants in this study were pre-selected to obtain continuous flowering of their
mixture. However, the results showed that under field conditions, other factors such as
weather and competition between plants can influence their availability to pollinators in
the mixtures. Therefore, the results provide some interesting hints for the design of
improved species mixtures, where mixing individual species with different flowering
periods and avoiding competition between them may be critical. Thus, other possible
option could be planting a mosaic of mono-specific stands and trying to optimize their
management. The explanation for lower number of pollinator visits in mixed plots was
probably due to the lower floral density in these plots (Essenberg, 2012; Hudewenz et
al., 2012), as within the mixed plots it never occurred that all plants were flowering at
the same time. Therefore, there is a need to exclude the plants that were not efficient in
the floral mixture (i.e. M. officinalis and T.capitata) in order to increase the floral
density of the mixture, and offer constant food sources to pollinators.

CHAPTER IV
Published online: Agricultural and Forest Entomology (Accepted: 15/05/2014)
Barbir, J., Badenes-Peréz, F.R., Fernández-Quintanilla, C., Dorado, J. (2014). The
attractiveness of flowering herbaceous plants to bees (Hymenoptera: Apoidea) and
hoverflies (Diptera: Syrphidae) in agro-ecosystems of Central Spain. Agricultural and
Forest Entomology (DOI: 10.1111/afe.12076)

Chapter IV
38
IV. The attractiveness of flowering herbaceous plants to bees
(Hymenoptera: Apoidea) and hoverflies (Diptera: Syrphidae)
in agro-ecosystems of Central Spain
1. Introduction
The global biodiversity decline is affecting the functioning of ecosystems given
that it is threatening the stability of the ecosystem services (Dobson et al., 2006;
Wratten et al., 2012; Vanbergen and Insect Pollinators Initiative, 2013). Under these
circumstances, agro-biodiversity can play a fundamental role in providing ecosystem
services such as pollination and pest control in agro-ecosystems (Altieri, 1999; Gurr et
al., 2003; Scheper et al., 2013). Bees are generally the most efficient pollinators of wild
flowers and commercial crops (LaSalle and Gauld, 1993; Jauker et al., 2012; Woodcock
et al., 2013), while hoverflies are effective pollinators of open flowers and many species
are also natural enemies of aphid pests (Ambrosino et al., 2006; Fontaine et al., 2006).
Many studies have reported an overall decline in the abundance and diversity of
wild pollinators (Altieri, 1991; Potts et al., 2010; Carvalheiro et al., 2013). Some studies
have linked this decline to the potential spread of diseases (Vanbergen and Insect
Pollinators Initiative, 2013) or competition (Goulson, 2003; Walther-Hellwig et al.,
2006) from managed honey bees. On the other hand, changes in climate (Memmott et
al., 2010; Rader et al., 2013), a reduction and a fragmentation of habitats (Pfiffner and
Wyss, 2004), an expansion of intensive agriculture (Benton et al., 2003; Watling and
Donnelly, 2006) and high inputs of pesticides (Brittain and Potts, 2011; Gill et al.,
2012) are also important factors that are associated with the decline in pollinators.
Nevertheless, semi-natural habitats can offer diverse forage plants and shelter for
beneficial insects within the agricultural fields, which has been recognized as essential
for both biodiversity conservation (Billeter et al., 2008) and for the maintenance of
ecosystem services in agro-systems (Klein et al., 2003b; Carvalheiro et al., 2012).
Therefore, as the parallel diversity decline of pollinators and insect-pollinated plants has
been reported (Biesmeijer et al., 2006), suggesting a functional coupling and mutual
dependence between both guilds (Frund et al., 2010), several studies have focused on

Chapter IV
39
improving the habitat for beneficial insects within agro-ecosystems in order to conserve
them. Some of those studies have suggested using diverse field margins to provide
nectar and pollen resources (Carvell et al., 2007; Heard et al., 2007) or nesting habitat
(Goulson et al., 2010) to pollinators in agro-ecosystems, while others have investigated
the differences in attractiveness of flowers to pollinators (Pontin et al., 2006; Hogg et
al., 2011). Although plant attractiveness is considered an important driver when
choosing the plants for beneficial insects’ conservation practices (Campbell et al.,
2012a; Carrie et al., 2012), it is essential to combine it with the factor of flowering
duration of the different plant species (Nicholls and Altieri, 2013).
Due to the lack of practical information on attractiveness and management of
beneficial plants in Spain (Miñarro and Prida, 2013), the implementation of known
beneficial plants within Spanish agro-ecosystems is still limited. Therefore, the main
objective of this study was to find the most appropriate plant species to improve the
habitat for pollinators in agro-ecosystems of Central Spain, where a preliminary
selection of plants prioritized native or naturalized (Tuell et al., 2008; Morandin and
Kremen, 2013) and not invasive (Stout and Morales, 2009) plants. Consequently, this
study addressed several specific objectives: i) to evaluate both attractiveness (insect
visitation) and attractiveness efficiency (a combination of duration of flowering and
insect visitation) of various plant species from diverse botanical families; ii) to evaluate
how the selected plants respond to two different agronomic managements (growing in
mixture vs. mono-specific stands; tillage vs. no-tillage at the end of the season to check
their ability of self-seeding); and iii) to estimate the spatial expansion, i.e. potential
weediness, of the different plant species, because it may threaten the functionality of
agro-ecosystems (Marshall and Moonen, 2002).
2. Materials and methods
2.1. Study area and experimental design
This study has been conducted over two years at the experimental farm La Poveda
(40º 19' N and 3º 29' W, elevation 536 m), Arganda del Rey, Madrid, Spain. The study
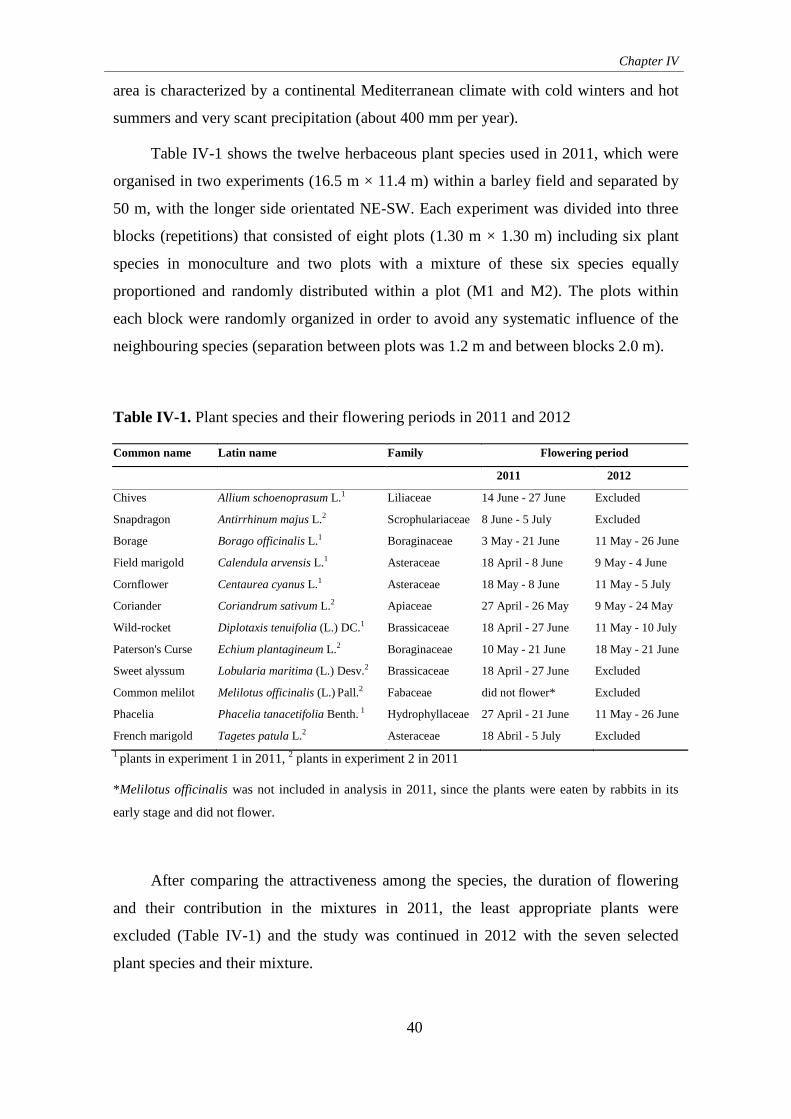
Chapter IV
40
area is characterized by a continental Mediterranean climate with cold winters and hot
summers and very scant precipitation (about 400 mm per year).
Table IV-1 shows the twelve herbaceous plant species used in 2011, which were
organised in two experiments (16.5 m × 11.4 m) within a barley field and separated by
50 m, with the longer side orientated NE-SW. Each experiment was divided into three
blocks (repetitions) that consisted of eight plots (1.30 m × 1.30 m) including six plant
species in monoculture and two plots with a mixture of these six species equally
proportioned and randomly distributed within a plot (M1 and M2). The plots within
each block were randomly organized in order to avoid any systematic influence of the
neighbouring species (separation between plots was 1.2 m and between blocks 2.0 m).
Table IV-1. Plant species and their flowering periods in 2011 and 2012
Common name Latin name Family Flowering period
2011 2012
Chives Allium schoenoprasum L.1 Liliaceae 14 June - 27 June Excluded
Snapdragon Antirrhinum majus L.2 Scrophulariaceae 8 June - 5 July Excluded
Borage Borago officinalis L.1 Boraginaceae 3 May - 21 June 11 May - 26 June
Field marigold Calendula arvensis L.1 Asteraceae 18 April - 8 June 9 May - 4 June
Cornflower Centaurea cyanus L.1 Asteraceae 18 May - 8 June 11 May - 5 July
Coriander Coriandrum sativum L.2 Apiaceae 27 April - 26 May 9 May - 24 May
Wild-rocket Diplotaxis tenuifolia (L.) DC.1 Brassicaceae 18 April - 27 June 11 May - 10 July
Paterson's Curse Echium plantagineum L.2 Boraginaceae 10 May - 21 June 18 May - 21 June
Sweet alyssum Lobularia maritima (L.) Desv.2 Brassicaceae 18 April - 27 June Excluded
Common melilot Melilotus officinalis (L.) Pall.2 Fabaceae did not flower* Excluded
Phacelia Phacelia tanacetifolia Benth. 1 Hydrophyllaceae 27 April - 21 June 11 May - 26 June
French marigold Tagetes patula L.2 Asteraceae 18 Abril - 5 July Excluded 1 plants in experiment 1 in 2011, 2 plants in experiment 2 in 2011
*Melilotus officinalis was not included in analysis in 2011, since the plants were eaten by rabbits in its
early stage and did not flower.
After comparing the attractiveness among the species, the duration of flowering
and their contribution in the mixtures in 2011, the least appropriate plants were
excluded (Table IV-1) and the study was continued in 2012 with the seven selected
plant species and their mixture.

Chapter IV
41
2.2. Insect visitation assessment
Grouping of pollinators. The local insects were grouped in accordance with their
morpho-physiological features to function as pollinators, such as body size and method
of pollen collection. The analysed groups were: honey bees, Apis mellifera L. (Apidae),
leafcutter bees (Megachilidae), small solitary bees (Halictidae and Andrenidae, with
body size ≤ 1cm), big solitary bees (Apidae, Halictidae and Andrenidae, with body size
> 1cm) and hoverflies. Bumblebees, Bombus spp. (Apidae) and carpenter bees,
Xylocopa violacea L. (Apidae) were not included in the final analysis as fewer than ten
visits were recorded during the whole experiment.
Although the main focus of this study was on the flower visitation of bees and
hoverflies, some beetles were studied as they occupied the same feeding niche as bees
and hoverflies and in that way indirectly influenced the visits of the main pollinators.
Attractiveness of the plants. In order to estimate the attractiveness of flowers to
local pollinators, the mean number of visits of bees and hoverflies per minute
(Campbell et al., 2012) was compared between plant species and their mixtures during
the spring of 2011 and 2012. Insect visits to the flowers were counted twice per week (9
min/plot) in a marked area of 0.5 m × 0.5 m, located in the centre of each plot. Visits
were counted only when the insect had a direct contact with the reproductive structures
of the flower (i.e., stamens and pistils). The observations were conducted by three
observers, always in the morning (9.00 h – 13.00 h local time, Greenwich Mean Time +
2:00) when the weather conditions were preferable for the insects to fly and visit
flowers (sunny days without wind and mean temperatures between 22oC and 32oC).
Switching plots, each observer counted visits of insects for three minutes in each plot.
In order to identify the different insect species, insect captures were made twice per
month using a butterfly net and an aspirator. Insect were always caught in the morning,
a day after the experimental observations thus not biasing the results.
Attractiveness efficiency of the plants. Considering that the attractiveness of a
plant species to pollinators cannot be efficient in agro-ecosystems if its flowering
duration is short, the term attractiveness efficiency has been introduced in this study,
visualizing a relation between the total pollinator visits to the flower/minute and the
duration of flowering (the total number of observation days).
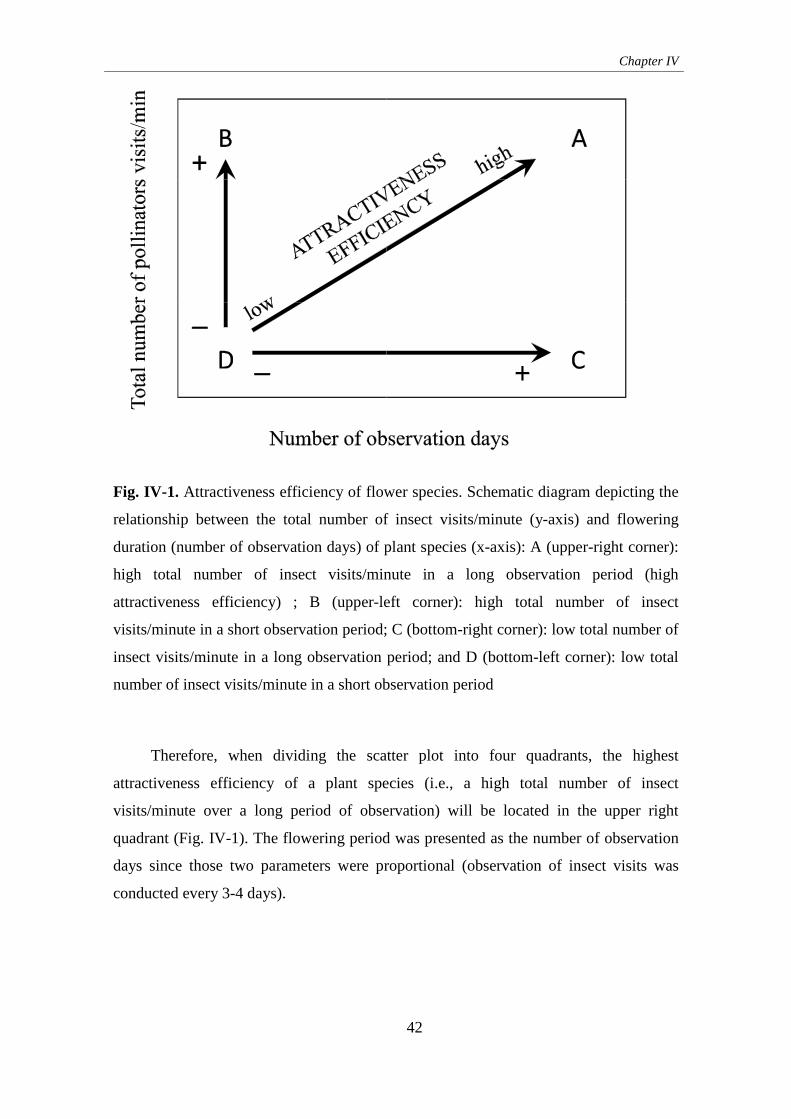
Chapter IV
42
Fig. IV-1. Attractiveness efficiency of flower species. Schematic diagram depicting the
relationship between the total number of insect visits/minute (y-axis) and flowering
duration (number of observation days) of plant species (x-axis): A (upper-right corner):
high total number of insect visits/minute in a long observation period (high
attractiveness efficiency) ; B (upper-left corner): high total number of insect
visits/minute in a short observation period; C (bottom-right corner): low total number of
insect visits/minute in a long observation period; and D (bottom-left corner): low total
number of insect visits/minute in a short observation period
Therefore, when dividing the scatter plot into four quadrants, the highest
attractiveness efficiency of a plant species (i.e., a high total number of insect
visits/minute over a long period of observation) will be located in the upper right
quadrant (Fig. IV-1). The flowering period was presented as the number of observation
days since those two parameters were proportional (observation of insect visits was
conducted every 3-4 days).

Chapter IV
43
2.3. Floral density
The capacity of plants to produce flowers was assessed. For this, the number of
flowers was counted on each observation day within the marked area (0.5 m × 0.5 m),
while the percentage of floral coverage per plot was assessed once per week. Each
observer estimated the percentage of flower coverage per m2 in each plot, and the mean
value of the three readings was calculated. Those parameters were used in order to
estimate the floral density within the studied plots and to investigate its influence on the
attractiveness of the plants (Stout et al., 1998). Each capitulum of species belonging to
the Asteraceae family (i.e., T. patula, C. cyanus and C. arvensis) was considered as an
individual flower. Any plot was considered in bloom when it had more than five open
flowers per observed area.
2.4. Effect of tillage and no-tillage on the self-seeding of plant species
At the end of each floral season (July), how the two different soil management
practices (tillage and no-tillage) affected self-seeding (the ability of the plant to
reproduce itself) in the studied plant species was compared. In order for this to take
place, half of each plot was shallow tillage while in the other half no-tillage was
applied. At the beginning of the following season (January), when the majority of
seedlings emerged, the number of seedlings under each treatment was counted three
times using a 25 cm × 25 cm frame. At this time, the density of seedlings was used to
estimate which agronomical treatment was more appropriate for the successful
maintenance of the plant species in successive years.
2.5. Potential weediness of plant species
In spring (April-May), the spatial expansion from the original area [Δ Area (%)]
of each plant species (the sum of the three mono-specific plots (1.30 m × 1.30 m) and
one sixth of all the mixed plots), was calculated considering the area occupied the
following year by using real time kinematic differential GPS (RTK-GPS). The
percentage increase in area occupied by each plant species relative to their original area
(i.e. potential weediness) was calculated using the software ArcGIS® 9.3 (ESRI,
Redlands, California, USA).

Chapter IV
44
2.6. Data analysis
The data from 2011 and 2012 were independently analysed with SPSS®
Statistics19 software (IBM, Chicago, IL, USA). A simple general linear model (GLM)
was used in order to assess the ability of different insects to discriminate between
flower species. The mean value of insect visits per minute within the observation area
(0.5 m × 0.5 m) was used as response variable for these analyses. The data of hoverflies
visitation in 2012 was square root transformed prior to analysis in order to meet the
assumption of normally distributed residuals and homogeneity of variance. Differences
among means were explored by Tukey’s B post hoc test (P ≤ 0.05). Univariate ANOVA
was also used to test whether there were significant differences between tillage and no-
tillage treatments. Finally, a bivariate two-tailed Pearson analysis was used to assess the
correlation between floral density and visits of pollinators/minute.
3. Results
3.1. Diversity of pollinators in the studied area
Bees were the most diverse group of pollinators (Table IV-2) with 41 different
species from five families (Apidae, Andrenidae, Colletidae, Halictidae and
Megachilidae). Due to the semiarid conditions of the experimental area, only 7 species
of hoverflies were observed (Table IV-2). Even though weather conditions in the spring
2011 were favourable for hoverflies (mean temperature 17oC and mean daily
precipitation 3 mm), a drier and colder spring in 2012 (mean temperature 15oC and
mean daily precipitation 1 mm) reduced their abundance and diversity in 2012 (SIAR,
2014). In addition, four species of beetles (Table IV-2) were found occupying the same
feeding niche as the insects of interest.

Chapter IV
45
Table IV-2. List of insects visiting the studied plant species Orden Family Species Size (mm) Groups of insects *
Coleoptera Dasytidae Psilothrix viridicoerulea (Geoffroy, 1785) 7 beetles
Coccinelidae Coccinela septempunctata (Linnaeus, 1758) 7 beetles
Tenebrioidae Heliotaurus ruficollis (Fabricius, 1781) 10 beetles
Scarabaeidae Tropinota squalida (Scopoli, 1783) 14 beetles
Hymenoptera Apidae Apis mellifera (Linnaeus, 1758) 11-16 honey bee
Bombus terrestris (Linnaeus, 1758) 12-21 bumblebee
Bombus muscorum (Linnaeus, 1758) 10-20 bumblebee
Xylocopa violacea (Linnaeus, 1758) 22-23 carpenter bee
Amegilla quatrifasciata (Villers, 1789) 12-15 solitary big
Anthophora agama (Radoszkowski, 1869) 15-20 solitary big
Anthophora atroalba (Lepeletier, 1841) 12-16 solitary big
Anthophora fulvitarsis (Brullé, 1832) 16-18 solitary big
Habropoda zonatula (Smith 1854) 14-17 solitary big
Eucera caspica (Morawitz, 1873) 11-12 solitary big
Eucera clypeata (Erichson, 1835) 11-12 solitary big
Eucera elongatula (Vachal, 1907) 9-11 solitary big
Andrenidae Andrena albopunctata ssp. melona (Rossi, 1792) 13-16 solitary big
Andrena bicolorata (Rossi, 1792) 11-15 solitary big
Andrena bimaculata (Kirby 1802) 10-13 solitary big
Andrena florea (Fabricius, 1781) 10-15 solitary big
Andrena gelriae ssp. gredana (van der Vecht, 1927) 9 - 13 solitary big
Andrena haemorrhoa (Fabricius, 1781) 9-11,5 solitary big
Andrena hispania (Warncke, 1967) 13-16 solitary big
Andrena morio (Brullé, 1832) 12-19 solitary big
Andrena nigroaenea (Kirby 1802) 10-14 solitary big
Andrena ovatula (Kirby 1802) 10-13 solitary big
Andrena similis (Smith, 1849) 7-12 solitary big
Andrena thoracica (Fabricius, 1775) 11-15 solitary big
Andrena truncatilabris ssp. espanola (Morawitz, 1877) 9-12 solitary big
Andrena variabilis (Smith 1853) 10-16 solitary big
Andrena vetula (Lepeletier, 1841) 10-12,5 solitary big
Andrena sp. 2-7 solitary small
Andrena sp. 1-5 solitary small
Panurgus canescens (Latreille, 1811) 4 -6 solitary small
Colletidae Colletes cunicularius (Linnaeus, 1761) 12-13,5 solitary big
Halictidae Halictus scabiosae (Rossi, 1790) 14-17 solitary big
Lasioglossum callizonius (Perez, 1895) 6-8 solitary small
Lasioglossum sp. 2-5 solitary small
Lasioglossum sp. 9 solitary small
Lasioglossum sp. 2-8 solitary small

Chapter IV
46
Family Species Size (mm) Group of insect
Megachilidae Anthidium oblongatum (Illiger, 1806) 7-8,5 leafcutter bee
Chelostoma emarginatum (Nylander, 1856) 9 leafcutter bee
Megachile sp. 1 9 leafcutter bee
Megachile sp. 2 11 leafcutter bee
Osmia tricornis (Latreille, 1811) 10-14 leafcutter bee
Diptera Syrphidae Episyrphus balteatus (De Geer, 1776) 9-12 hoverfly
Eupeodes corollae (Fabricius, 1794) 6-11 hoverfly
Sphaerophoria scripta (Linnaeus, 1758) 8 -11 hoverfly
Sphaerophoria rueppellii (Weidemann, 1820) 8-11 hoverfly
Eristalis tenax (Linnaeus, 1758) 10-12 hoverfly
Melanostoma sp. 9-11 hoverfly
Scaeva sp. 8-10 hoverfly
* Insects were classified in eight groups (beetles, honey bees, bumblebees, carpenter bees, solitary big,
solitary small, leafcutter bees and hoverflies). All solitary bees, except leafcutter bees, were classified in
two groups using the body length [solitary small (≤ 10 mm) and solitary big (> 10mm)] as main criteria
3.2. Attractiveness of different plants and their mixtures to pollinators
In general, the visitation frequency of bees was significantly higher in 2012 than
in 2011, while the mean number of visits per minute of hoverflies and beetles was much
higher in 2011 than in 2012 (Fig. IV-2 and IV-3).
In 2011, bees were most attracted to the flowers of B. officinalis, P. tanacetifolia
and D. tenuifolia in experiment 1 (F6,17 = 6.78, P≤ 0.01), while in experiment 2 the
flowers of E. plantagineum and M2 were most attractive to the bees (F5,15 = 24.08, P≤
0.01). In 2012, flowers of C. sativum, E. plantagineum and P. tanacetifolia were
significantly more visited by the bees than the flowers of B. officinalis and C. arvensis
(F7,16 = 7.20, P≤ 0.01) (Fig. IV-2). In general, the most frequent visitors were solitary
bees, although the results of both years point to the preference of honey bees for B.
officinalis (year 2011: F6,17 = 16.80, P≤ 0.01; year 2012: F7,16 = 6.23, P≤ 0.01) as well
as leafcutter bees for the flowers of C. cyanus (year 2011: F6,17 = 2.35, P= 0.07; year
2012: F7,16 = 9.79, P≤ 0.01).
In 2011, hoverflies visited significantly more frequently the flowers of C.
arvensis, D. tenuifolia and M1 (F6,17 = 16.69, P≤ 0.01) in experiment 1, while in
experiment 2 the flowers of L. maritima were the most attractive followed by C.
sativum (F5,15 = 49.08, P ≤ 0.01). The most regular hoverflies to visit the plants in 2011
were Sphaerophoria scripta and Sphaerophoria rueppellii, two species grouped as

Chapter IV
47
Sphaerophoria spp. (Fig. IV-2). In 2012, few hoverflies were observed in the study
area, preferably visiting the same plant species as in 2011, i.e. D. tenuifolia, C. sativum
and C. arvensis (F7,16 = 4.48, P≤ 0.01), except L. maritima that was not used in 2012.
Fig. IV-2. Visits of bees, hoverflies and beetles to the flowers of the studied plant
species. Means with same letter in each graph are not significantly different at P ≤ 0.05,
as determined by Tukey’s B multiple comparison test.
Abbreviations: 2011 (1): Bor= Borago officinalis; Cal= Calendula arvensis; Cen= Centaurea cyanus;
Dpl= Diplotaxis tenuifolia; Pha= Phacelia tanacetifolia; Ali= Alium schoenoprasum; M1= mixture of
those species. 2011 (2): Cor= Coriandrum sativum; Ech= Echium plantagineum; Lob= Lobularia
maritima; Tag= Tagetes patula; Ant= Antirrhinum majus; M2= mixture of those species. 2012: M=
mixture of the species used in 2012.

Chapter IV
48
According to the results, the flowers of E. plantagineum (F5,15 = 9.62, P≤ 0.01)
and P. tanacetifolia (F6,17 = 32.61, P≤ 0.01) were the most attractive to beetles in 2011,
while in 2012 beetles were more abundant on the flowers of C. sativum (F7,16 = 9.01, P≤
0.01) (Fig. IV-2). The most frequent visitor to all flowers was Heliotaurus ruficolis,
except C. arvensis and C. cyanus which mainly attracted Psilothrix viridicoerulea and
Coccinella septempunctata.
Furthermore, the results showed a strong positive correlation between floral
density and the number of visits by bees in both years of the study (year 2011: r= 0.119,
n= 810, P≤ 0.01; year 2012: r= 0.413, n= 407, P≤ 0.01), while visits of hoverflies were
significantly positively correlated with the floral density in 2011 (r= 0.634, n= 810, P≤
0.01), but not in 2012, probably because of the low abundance of hoverflies in the field
area that year.
3.3. Attractiveness efficiency of different plants and their mixtures to
pollinators
Figure IV-3 illustrates the scatter plot showing the relationship between the total
number of beneficial insect visits and the duration of the flowering period of each plant
species (i.e., the attractiveness efficiency). The plant species with the highest
attractiveness efficiency (i.e., the upper-right square of the scatter plot) for bees were B.
officinalis, D. tenuifolia, P. tanacetifolia and the mixtures in 2011, and P. tanacetifolia,
E. plantagineum, C. cyanus, D. tenuifolia and the mixture in 2012. Regarding
hoverflies, the highest attractiveness efficiency was observed in L. maritima in 2011
and D. tenuifolia in 2012. The lowest attractiveness efficiency (i.e., bottom-left square
of the scatter plot) for both bees and hoverflies was found in A. majus, A.
schoenoprasum and C. cyanus in 2011 and in C. sativum and C. arvensis in 2012 (Fig.
IV-3).

Chapter IV
49
Fig. IV - 3. Attractiveness efficiency of the studied plant species for bees and hoverflies
in 2011 and 2012 (abbreviations of plant species as in Fig. IV-2.).
Scatter-plots depict the relationship between the total number of insect visits/minute (y-axis) and
flowering duration (number of observation days) of plant species (x-axis). Bars represent the standard
deviation from 3 replicated plots.
After consulting the results of attractiveness efficiency for 2011 it was decided to
exclude some plant species which were not attractive to bees (L. maritima), not
attractive to hoverflies (T. patula and A. schoenoprasum) and that which was not
attractive to neither bees nor hoverflies (A. majus). Additionally, A. schoenoprasum, L.
maritima and T. patula were also excluded from the experiment as their height
(maximum 20 cm) was limiting their contribution when mixed with taller plant species
(60-100 cm) used in the experiment (J. Barbir, personal observation). Considering that
the flowering period of C. cyanus was significantly reduced in 2011 due to aphid attack,
it was decided to test again its attractiveness to pollinators in 2012.

Chapter IV
50
3.4. Effect of tillage and no-tillage on the self-seeding of plant species
The results of the first year (2011) showed that seedling emergence was
significantly reduced by tillage in C. arvensis (F1,16 = 6.57, P= 0.02) and P. tanacetifolia
(F1,16 = 4.79, P= 0.04) with respect to no-tillage treatment. In contrast, C. sativum
showed higher seedling emergence after tillage (F1,16 = 12.99, P= 0.02). Some plant
species such as C. cyanus and D. tenuifolia did not emerge in 2011 (Table IV-3).
Table IV-3. Differences in seedling emergence (Nº plants/m2) after applying two
different managements at the end of plants cycle: shallow tillage and no-tillage
Plant species Treatment No. plants m–2 ± SD
2011 2012
B. officinalis tillage 26.67 ± 30.98 1.78 ± 5.33
no-tillage 26.67 ± 17.88 19.56 ± 31.78
C. arvensis tillage 2816.89 ± 1103.66 231.11 ± 119.23
no-tillage 4522.67 ± 1664.15 * 1223.11 ± 78.02 *
C. cyanus tillage 0 58.67 ± 32.98
no-tillage 0 190.22 ± 97.47 *
C. sativum tillage 313.56 ± 246.96 * 1.78 ± 5.33
no-tillage 16.19 ± 16.54 0
D. tenuifolia tillage 0 5.33 ± 8.00
no-tillage 0 152.89 ± 78.43 *
E. plantagineum tillage 234.61 ± 60.1 1.78 ± 5.33
no-tillage 230.26 ± 135.35 236.44 ± 127.94 *
P. tanacetifolia tillage 226.12 ± 96.79 1.78 ± 5.33
no-tillage 318.13 ± 80.83 * 115.56 ± 160.75 *
*significant difference between the treatments at P≤ 0.05
Similar results to those of 2011, only with a lower seedling emergence after
tillage, were obtained in 2012 (Table IV-3) for C. arvensis (F1,16 = 436.24, P≤ 0.01) and
P. tanacetifolia (F1,16 = 4.50, P= 0.05). Although there was no significant difference
between tillage and no-tillage treatments in C. sativum in 2012, there was a tendency
that tillage positively affected the seedling emergence of this plant. Both, C. cyanus and
D. tenuifolia emerged in 2012, and the number of seedlings was significantly reduced
by tillage for both species (F1,16 = 14.71, P≤ 0.01 and F1,16 = 31.53, P≤ 0.01,
respectively).

Chapter IV
51
3.5. Potential weediness of plant species
The results of both years demonstrated the potential weediness of C. arvensis
since its initial area increased by 1632% (2011) and 1300% (2012) within one year
(Table IV-4). Furthermore, in 2011 the initial area of P. tanacetifolia increased by 611%
and in 2012 by 124%. Although the initial area of E. plantagineum increased by 275%
in 2011, in 2012 it increased by only 12% (Table IV-4).
Table IV-4. Spatial increase (%) of the area occupied by plant species one year after
planting
Plant species Initial
area (m2)
Area in the year after
planting (m2) Δ Area (%)
2011 2012 2011 2012 2011 2012
B. officinalis 7.29 6.38 0.17 4.19 -98 -34
C. arvensis 7.29 6.38 126.26 89.35 1632 1300
D. tenuifolia 7.29 6.38 3.58 8.05 -51 26
C. cyanus 7.29 6.38 1.25 7.84 -83 23
P. tanacetifolia 7.29 6.38 51.8 14.26 611 124
A. schoenoprasum 7.29 - 0 - -100 -
E. plantagineum 7.29 6.38 27.31 7.15 275 12
T. patula 7.29 - 0 - -100 -
L. maritima 7.29 - 0 - -100 -
M. officinalis 7.29 - 0 - -100 -
A. majus 7.29 - 1.3 - -82 -
C. sativum 7.29 6.38 0 0 -100 -100
External weeds 0 0 171.66 67.89 17166 6789
In both years of study the plants of B. officinalis, D. tenuifolia and C. cyanus
appeared the following years but did not show weediness and maintained their area
within the initial plots. On the other hand, seedlings from A. schoenoprasum, T. patula,
L. maritima and M. officinalis did not emerge the following year.

Chapter IV
52
4. Discussion
In accordance with some previous studies (Colley and Luna, 2000; Frund et al.,
2010) the results of this study showed differences in attractiveness among the plant
species. In both years of study, the most attractive plants for bees were P. tanacetifolia
and E. plantagineum, followed by D. tenuifolia and mixed plots. However, no
consistent results were found for the other plant species from one year to another.
Therefore, in 2011, B. officinalis was more frequently visited by honey bees than in
2012, while the flowers of C. cyanus and C. sativum had more bee visits in 2012 than in
2011. Some plants selected for this study, such as P. tanacetifolia and B. officinalis,
have been previously studied in other regions, i.e. USA and UK, and are frequently
cited as being attractive to bees (Carreck and Williams, 2002; Campbell et al., 2012a).
In this study, the results confirmed that P. tanacetifolia was highly attractive to bees in
Central Spain, while B. officinalis was the most attractive plant species to honey bees
although being less visited in 2012 than in 2011.
On the other hand, the most attractive plant to hoverflies in 2011 was L. maritima,
followed by C. sativum, C. arvensis, D. tenuifolia and mixed plots. In 2012 there were
very few hoverflies in the study area, probably because of the cool and dry spring
(Amoros-Jimenez et al., 2012). However, the flowers of D. tenuifolia were the most
attractive to the few hoverfly visitors recorded. The results from 2011 confirmed the
high attractiveness of L. maritima and C. sativum to hoverflies reported in other regions
and under different field conditions (Ambrosino et al., 2006; Campbell et al., 2012a).
In addition to the attractiveness of plants to pollinators, a long flowering period is
considered important (Adhikari and Adhikari, 2010). For that reason, the analysis of
attractiveness efficiency of the plant species acquired important significance as it
combined the total number of insect visits/minute and flowering duration, facilitating
the choice of plants which are beneficial for insect conservation. Among the other plant
species assessed, D. tenuifolia and E. plantagineum were attractive to bees for a long
flowering period, reflecting their high attractiveness efficiency in both years of study.
Similarly, the analysis of the attractiveness efficiency to hoverflies showed that L.
maritima and D. tenuifolia maintained their attractiveness constantly during their long
flowering period, while the short biological cycle of C. sativum reduced its

Chapter IV
53
attractiveness efficiency in the field. Therefore, although attractive to hoverflies, a short
flowering period limited C. sativum as a food source for hoverflies in agro-ecosystems.
In contrast, D. tenuifolia which had a long flowering period and showed to be attractive
for both groups of beneficial insects (bees and hoverflies) in both years of study could
have an important potential in the conservation of pollinators within agro-ecosystems.
In addition, D. tenuifolia could have an added economic value as it is widely used as
rocket salad (D'Antuono et al., 2009; Hall et al., 2012).
The study pointed out that some inter-specific interactions between pollinators
and non-pollinator insects can affect the attractiveness of plants through long flower
occupation (Bosch, 1992) or damage (J. Barbir, personal observation). For example, H.
ruficolis occupied the flowers of P. tanacetifolia and E. plantagineum, while P.
viridicoerulea, described as a poor pollinator (Bartomeus et al., 2008), occupied the
flowers of C. arvensis, reducing the approachability of the flowers to bees and
hoverflies. Additionally, Tropinota squalida, described as a pest in agro-systems
(Khater et al., 2003), negatively influenced the attractiveness of the studied flowers to
beneficial insects by destroying the bottom part of corollas when reaching for the nectar
of E. plantagineum (J. Barbir, personal observation). Nevertheless, in 2012 the number
of beetle visits was significantly lower than in 2011, coinciding with an increase in the
number of bee visits in P. tanacetifolia and E. plantagineum. As these inter-specific
insect interactions can influence the final estimation of the attractiveness of the plants to
bees and hoverflies, it would be advisable to be considered in further studies on the
conservation of pollinators.
Regarding the management of plant species in a mixture, the hypothesis was that
mixtures could combine the advantages of all species in terms of a longer flowering
period and offering complementary resources relative to single species. However, the
results of this study have revealed that, in general, mixed plots showed lower
attractiveness efficiency than mono-specific plots of D. tenuifolia for both groups of
pollinators and in both years of study. The lower number of visits of pollinators in
mixed plots might be due to the lower floral density in these plots (Essenberg, 2012;
Hudewenz et al., 2012). In contrast, although mixed plots were less visited by
pollinators than some mono-specific plots, i.e. P. tanacetifolia, E. plantagineum or L.
maritima, they showed a longer flowering period and higher diversity of floral traits.
Furthermore, some plant species did not contribute efficiently to floral mixtures because

Chapter IV
54
of their low attractiveness to pollinators (A. majus), low height (L. maritima) or both (A.
schoenoprasum and T. patula). Therefore, plants such as L. maritima, which is
attractive to hoverflies (Colley and Luna, 2000) and improves biological control in
vineyards (Begum et al., 2006), has a greater potential as a beneficial plant, if planted as
a mono-specific stand.
As the study area was relatively small, the results regarding the weediness of
plants and their behaviour under field conditions should be considered as preliminary.
Although tillage reduced the seedling emergence of C. arvensis within the plots, it was
not effective in stopping its wide spatial expansion. This finding is in accordance with
Saramago et al. (2012) who also described C. arvensis as weedy. On the other hand,
seedlings of P. tanacetifolia also expanded beyond their original plots in the following
years, but as the seedling emergence was significantly reduced after tillage treatment,
this management practice may be appropriate as a control against its potential
weediness. Indeed, previous studies (Rasmussen, 1991) suggested the positive weed-
killing effect of harrowing on P. tanacetifolia. On the other hand, E. plantagineum
expanded beyond its original plots only in 2011 and its self-seeding was successfully
reduced by tillage. Although E. plantagineum has not been reported as weedy in the
Mediterranean region, in other regions such as Australia, it has been described as
invasive and weedy (Grigulis et al., 2001).
In conclusion, of twelve herbaceous plants included in this study D. tenuifolia can
be implemented in agro-ecosystems of Central Spain as mono-specific stands, due to its
high attractiveness efficiency to both bees and hoverflies and its efficient self-
reproduction within the plots where it was planted. However, it can also be included in
mixtures with other plant species recommended in this study, i.e. B. officinalis and C.
sativum, as attractive to pollinators, not weedy, but with shorter flowering period than
D. tenuifolia. On the other hand, C. arvensis, despite its attractiveness to hoverflies, is
not recommendable as a beneficial plant in agro-ecosystems because of its potential
weediness. Conversely, when considering the interactions among the studied insects, P.
tanacetifolia and E. plantagineum, besides being attractive to bees, proved to be
attractive to beetles that occupy the same feeding niche as local pollinators. In addition,
those two plants also require tillage treatment to control their high self-seeding which
can lead to potential weedines. Therefore, it is necessary to expand this study to a larger

Chapter IV
55
scale in order to fully recommend the implementation of those beneficial plant species
in agro-ecosystems of Central Spain.

CHAPTER V
Published online: Acta Agriculturae Scandinavica, Section B – Soil & Plant Science
(Accepted: 13/05/2014)
Jelena Barbir, José Dorado, César Fernández-Quintanilla, Tijana Blanusa, Cedo
Maksimovic & Francisco R. Badenes-Pérez (2014). Wild rocket – effect of water
deficit on growth, flowering and attractiveness to pollinators. Acta Agriculturae
Scandinavica, Section B – Soil & Plant Science (DOI:10.1080/09064710.2014.925575)

Chapter V
57
V. Wild rocket - effect of water deficit on growth, flowering and
attractiveness to pollinators
1. Introduction
Although fresh water is one of the most important natural resources in the world,
it is not equally distributed, and in many regions is very limited and scarce. Considering
that the largest part of available water is used for irrigation in agriculture, it is important
to apply irrigation treatments that reduce water consumption and maximize water use
efficiency (Howell, 2001; Christian-Smith et al., 2012). Regulated deficit irrigation
(RDI), i.e. the deliberate water deficit applied to a crop at certain times (English, 1990),
has been shown to save water while maintaining plant growth of various crops in areas
with limited water sources (Chartzoulakis et al., 2002; Fereres and Soriano, 2007;
Perez-Pastor et al., 2014; Perez-Perez et al., 2014). On the other hand, exposure of
plants to water deficit can negatively affect plant development (Hsiao, 1973; Gutbrodt
et al., 2011; Garcia-Tejero et al., 2012) and for this reason, while applying strategies
that optimise water use in crop production, it is essential to mitigate the negative effects
of water stress on plant development.
As it is closely related to crop production, the effect of water deficit on plant
attraction to insect herbivores has been well-studied (Huberty and Denno, 2004).
Preference for water-stressed over well-watered plants has been shown in some studies
with certain insect herbivores (Showler and Moran, 2003), but other studies have shown
the opposite (Hoffman and Hogg, 1992) or no effect (Badenes-Perez et al., 2005) of
water deficit on herbivore preference. Considering that pollinators’ richness and
abundance have been decreasing with a decrease in plant diversity (Biesmeijer et al.,
2006), plants attractive to pollinators have taken an important role in their conservation
(Batary et al., 2010; Carrie et al., 2012). Nevertheless, the effect of water deficit on the
attraction of flowering plants to insect pollinators has not been studied yet.
Wild rocket, Diplotaxis tenuifolia (L.) DC. (Family: Brassicaceae), is one of the
three cultivated rocket species whose demand and production has been increasing in
Mediterranean countries (Bennett et al., 2007; Lamy et al., 2008; Znidarcic et al., 2011;

Chapter V
58
Hall et al., 2012). Besides being used as a crop, it can be used to attract bees and
hoverflies over its long flowering season (Barbir et al., 2014a). Although D. tenuifolia is
a C3 – C4 intermediate plant, with more efficient photosynthetic performance at high
temperatures than other cruciferous plants with C3 photosynthesis (Schuster and
Monson, 1990; Ueno et al., 2006), little is known about its ecology and physiology
under water deficit. As pollinators are mainly nectar feeders and the effect of water
deficit in insect preference is mediated by changes in plant physiology and plant
chemistry (Huberty and Denno, 2004; Gutbrodt et al., 2011), it is important to test if
water deficit could affect flower morphology and nectar composition in D. tenuifolia.
Considering the dry Mediterranean climate and its scarcity of water for irrigation,
it is important to study the effect of water deficit on the growth and the attractiveness to
pollinators of D. tenuifolia in order to evaluate its capability to cope with water scarcity
without negative consequences for its use as a crop and a beneficial plant to pollinators.
For this reason, the main objective of this study was to test different irrigation
treatments to find the most efficient irrigation regime for D. tenuifolia, both as a crop
and as a plant attractive to pollinators. In addition, two other objectives were to test the
effect of different drought duration on growth, flower development and attractiveness to
pollinators in D. tenuifolia; and to assess the effect of moderate deficit irrigation (MDI)
and severe deficit irrigation (SDI) on growth, flower development and attractiveness to
pollinators of D. tenuifolia.
2. Material and methods
The study was divided in two experiments. Experiment 1 was set up to test the
effect of different drought treatments on growth and floral development of D. tenuifolia,
and was conducted from May to August 2012 in Madrid, Spain. Experiment 2 was set
up to test the effect of different RDI treatments on growth and floral development of D.
tenuifolia, and it was conducted from August to October 2012 in Reading, UK. Both
experiments also tested the effect of the different water deficit treatments on
attractiveness of D. tenuifolia to bees and hoverflies.

Chapter V
59
2.1. Experiment 1 – Response of D. tenuifolia to drought
Seedlings of D. tenuifolia were transplanted to pots (10 cm × 13 cm) filled with
520 g of a mixture of peat-based compost and vermiculite (3:1) by volume, irrigated to
container capacity and moved to the greenhouse. The minimal (night-time) temperature
in the greenhouses during the experiment was 18-20 oC and the maximal (day-time) was
25-29 oC. The relative humidity varied from 50% to 80%.
Before starting the experiment, a preliminary study was conducted in order to
estimate the duration of the drought treatments (D) and the number of replicates
necessary to show significant differences in plant transpiration. For that purpose, plant
transpiration was measured every 2-3 days in five plants that were not being irrigated
(data not shown). All plants died after 16 days, so it was decided to apply drought
treatments over a period of 14 days, in order to test if plants could recover by re-
watering. Based on the preliminary differences in plant transpiration, it was decided to
use 5 different drought treatments and a replication of six plants per treatment.
In the greenhouse, plants were randomly organized, over four tables and the
drought treatments were applied after the plants of D. tenuifolia developed their first
flowers. In the control treatment, each plant was under 100% potential
evapotranspiration (ETp), i.e. adding all the water lost by evapotranspiration in the
previous 24 h, while the other treatments did not receive any irrigation for 4, 8, 11 or 14
days (D-4, D-8, D-11 and D-14), respectively. Therefore, the D-14 treatment started
first, followed by the D-11 treatment (three days later), the D-8 treatment (six days
later) and the D-4 treatment (ten days later).
At the end of the experiment, three plants of each treatment were chosen for
destructive analysis and other three plants were re-watered for 19 days to study if they
could recover after drought treatments applied (R19). The measured parameters were
plant height, biomass, number of leaves and open flowers, flower area (area/flower),
total floral area (total area of all open flowers/plant, on the day of the measurements),
flower diameter, corolla tube diameter, corolla tube length, and sugar concentrations in
floral nectar. These parameters estimated the effect of water deficit on plant survival
and development, while the effect of water deficit on the attractiveness of D. tenuifolia
to pollinators was measured directly under field conditions.

Chapter V
60
2.2. Experiment 2 – Response of D. tenuifolia to RDI
Seedlings of D. tenuifolia were transplanted to individual pots (10 cm × 10 cm)
filled with 400 g of peat-based compost, irrigated to container capacity and randomly
organized over tables in a greenhouse with a minimal (night-time) temperature of 12-16 oC and a maximal (day-time) temperature 20-33 oC. The relative humidity varied from
50% to 80%.
Plants were exposed to three irrigation treatments (six plants per each) based on
ETp. Control plants were grown under 100% ETp (all plants were weighed daily and
watered back to the container capacity). Plants under moderate deficit irrigation (MDI)
were grown under 60% ETp (plants received 60% of the 100% ETp values) while
severe deficit irrigation (SDI) plants were under 30% ETp (plants received 30% of the
100% ETp values). Plants within this experiment were divided into two groups. In the
first group, RDI was applied for a total of 30 days, 16 days before flowering (RDI-BF)
and 14 days during flowering (RDI-DF); while in the second group, RDI started when
all plants had developed their first three flowers (RDI-DF).
2.3. Water consumption
In order to estimate the quantity of water used for each irrigation treatment, total
water consumption/plant for control and water deficit treatments (D-4, D-8, D-11, D-14,
MDI-BF, SDI-BF, MDI-DF and SDI-DF) was calculated using following equation:
Total water consumption/plant = (CWD × No. daysWD) + (C100% ETp × No. days100% ETp)
Where CWD is the water consumption/plant under water deficit; No. daysWD is the
number of days under water deficit; C100% ETp is the water consumption/plant under
100% ETp; and No. days100% ETp is the number of days under 100% ETp. Sum of the
number of days under water deficit and the number of days under 100% ETp irrigation
was 30 days for all treatments.

Chapter V
61
2.4. Plant transpiration and growth
As plants close their stomata as a result of water stress (Chaves et al., 2003), an
AP4 Leaf Porometer (Delta-T Devices Ltd., Cambridge, UK) was used to measure leaf
stomatal conductance (gs). Stomatal conductance measurements were made between
11:00 and 13:00 h every 3–5 days in two leaves per plant, randomly chosen among
medium size leaves (6-10 cm length). Stomatal conductance was also used to assess
apparent plant survival (and 0 mmolm-2s-1 indicating the cessation of transpiration by a
plant).
The assessments of vegetative parameters were mainly focused on leaf number
and plant biomass, because of their importance for commercial production of wild
rocket (Znidarcic et al., 2011). Plant height and fresh and dry biomass of shoots (leaves,
stems and flowers) were measured at the end of each experiment. Total number of dead
and viable (living) leaves per plant was counted and the total leaf area (of living leaves)
was measured using a Leaf Area Meter (Delta – T Devices Ltd., Cambridge, UK). In
addition, leaves shorter than 10 cm (small leaves) and leaves longer than 10 cm (big
leaves) were counted separately in order to estimate if the impact of water deficit varied
depending on the size of the leaves.
2.5. Floral characteristics
In order to estimate total production of flowers per plant, open and senescent
flowers were counted separately. At the end of each experiment, three open flowers per
plant (the top youngest, the bottom oldest and one intermediate flower) of the highest
stem were used to measure the diameter of the corolla and the corolla tube, and the
corolla tube length using a Vernier calliper.
Total floral area per plant was measured using an Area Meter (Delta – T Devices
Ltd., Cambridge, UK) by placing all open flowers from a single plant with petals facing
the measurement pane. The average area per flower was calculated by dividing the total
floral area with the number of flowers per plant.

Chapter V
62
2.6. Nectar analysis
As the effect of water deficit on insect preference is mediated by changes in plant
chemistry (Gutbrodt et al., 2011), it was tested if water deficit could affect nectar
composition. For that purpose, the nectar was extracted by using a modification of the
rinsing method described in Petit et al. (2011). This method was preferred as it is not
destructive, which facilitated the use of flowers for further measurements. Three flowers
were used per plant, and each flower was washed twice with 2 μl of miliQ H2O. The
content was transferred into an erlenmeyer and diluted to 5 ml with miliQ H2O. High-
Performance Liquid Chromatography (Dionex, 4500i) was used to measure the content
of glucose, fructose and sucrose in each sample. Separation was done on an ionic
column (RCX-10, 250 × 4.6 mm, 7μm) using 5% sodium hydroxide 1N and 95% miliQ
H2O as solvents with a flow rate of 2 ml/minute (injection volume 50 μl).
2.7. Pollinator visits
In order to estimate if water deficit can affect the attractiveness of D. tenuifolia to
pollinators, their visits to the flowers of plants grown under different water deficit
treatments were monitored under field conditions. In both experiments, each insect visit
was counted only when the insect had a direct contact with the reproductive structures
of the flower (i.e., stamens and pistils). The observations were conducted on the last day
of each experiment, in the morning (9.00 h – 14.00 h local time), when the weather
conditions were preferable for the insects to fly and visit flowers (sunny days without wind
and temperatures between 22oC and 32 oC). In each observation set (group of three plants)
all opened flowers and insect visits per minute were counted (nine repetitions over a
three-minute period for each observation set). Some insect species were identified in
situ, while others were captured using a butterfly net for further identification.
In experiment 1, floral attractiveness of well-watered and drought-stressed plants
to pollinators (bees and hoverflies) was estimated under field conditions in two sites: the
CSIC experimental farm “La Poveda” (Arganda del Rey, Madrid, Spain) and the
vegetable garden of the Center for Ecology and Education “Puente del Perdón”
(Rascafría, Madrid, Spain). Data collection in the first site was conducted on the 16th of
May 2012, while on the second site data collection was conducted on the 17th of July

Chapter V
63
2012. Plants were transported from the greenhouse to the field sites by van (driving
distance < 1h). The experimental unit consisted of four observation sets (control, D-4,
D-8, D-11) organized in a circle (Ø3 m). The experiment consisted of three
experimental units separated 15 m from each other. Plants in the D-14 treatment were
excluded from the experiment because none of them had open flowers.
In experiment 2, floral attractiveness of well-watered and RDI water stressed
plants to pollinators was estimated under field conditions in the experimental garden of
the University of Reading, UK. The experimental unit consisted of five observation sets
(control, MDI-BF, SDI-BF, MDI-DF and SDI-DF) distributed in a circle (Ø3 m). The
experiment consisted of two experimental units separated 15m from each other.
2.8. Statistical analysis
Data were analysed with univariate analysis of variance (ANOVA) using SPSS®
Statistics19 software (IBM, Chicago, IL, USA). Prior to the ANOVA, the assumptions
of normal distribution and homogeneity of variances were tested by Kolmogorov
Smirnov and Levene’s tests, respectively. Differences among means were analyzed by
Tukey’s B post hoc test (P ≤ 0.05). A bivariate two-tailed Pearson analysis was used to
assess the correlation between floral parameters and visits of pollinators per minute.
3. Results
3.1. Response of D. tenuifolia to drought
Effect of drought on plant growth
Although gs decreased in all drought treatments (Fig. V-1A), significant
differences were found only for the plants that were drought-stressed for 11 and 14 days
(F4,85=6.96, P ≤ 0.01). After six days of recovery, the gs of D-14 plants continued to be
significantly lower than in other plants (F4,40=3.29, P = 0.02), while after the recovery
period of 16 and 19 days, gs reached values similar to the control treatment (Fig. V-1B).
After finishing the drought treatment, plants with gs zero (no transpiration)
appeared dead. However, all plants of control, D-4 and D-8 treatments, as well as 83%

Chapter V
64
of D-11 maintained their gs values above zero, while in 17% of D-11 plants and 100%
of D-14 leave transpiration was reduced to zero. After 19 days of recovery, gs values in
some plants recovered and the plants started to flower again (plant survival becoming
100% for D-11 and 33% for D-14 treatment at the end of experiment 1).
Fig. V-1. Mean stomatal conductance of D. tenuifolia under different drought
treatments (no irrigation for 4 [D-4], 8 [D-8], 11 [D-11] or 14 days [D-14]) (A) and
during a recovery period (well-irrigated) of 19 days (B).
For each measurement (day), means with different letter are significantly different at P ≤ 0.05, as
determined by the Tukey’s B multiple comparison test.
Furthermore, 14 days without irrigation (D-14 treatment) caused significant
decrease in all vegetative parameters: plant height (F4,25=7.84, P ≤ 0.01), fresh biomass
(F4,10=23.77, P ≤ 0.01), dry biomass (F4,10=5.01, P = 0.02), total number of leaves
(F4,25=10.09, P ≤ 0.01), number of leaves ≤10 cm length (F 4,25=7.79, P ≤ 0.01), number
of leaves >10 cm length (F4,25=7.75, P ≤ 0.01) while number of dead leaves significantly
increased (F4,25=17.03, P ≤ 0.01). Eleven days without irrigation (D-11 treatment)
caused a significant decrease of fresh biomass (F4,10=23.77, P ≤ 0.01), dry biomass

Chapter V
65
(F4,10=5.01, P = 0.02), total number of leaves (F4,25=7.75, P ≤ 0.01), number of leaves
>10 cm length (F4,25=7.75, P ≤ 0.01), while number of dead leaves significantly
increased (F4,25=17.03, P ≤ 0.01). However, plant growth was not significantly
influenced by 8 and 4 days without irrigation (Table V-1).
After 19 days of recovery, only fresh biomass weight was significantly lower in
D-14 plants compared to the control plants (F4,10=3.42, P = 0.05), while all other
parameters were comparable to control levels (Table V-1).
Effect of drought on floral characteristics
After 14 days without irrigation, D. tenuifolia plants did not have any open
flowers, so only senescent flowers and number of stems were counted. Significantly
lower values for total floral area (F4,24=15.15, P ≤ 0.01) and number of open flowers
(F4,25=15.93, P ≤ 0.01) were found in D-8 and D-11 (Table V-2). The values of flower
area (F4,24=66.59, P ≤ 0.01), flower diameter (F4,25=37.69, P ≤ 0.01), corolla tube
diameter (F4,25=37.67, P ≤ 0.01), corolla tube length (F4,25=36.67, P ≤ 0.01) and number
of stems were significantly lower in D-11 than in control, D-4 and D-8 plants
(F4,25=3.28, P = 0.05). After 19 days of recovery, no significant differences between
treatments were found for any of the measured floral parameters in any of the treatments
(Table V-2).

Chapter V
66
Tab
le V
-1.
Effe
cts o
f diff
eren
t dro
ught
trea
tmen
ts (n
o irr
igat
ion
for 4
[D-4
], 8
[D-8
], 11
[D-1
1] o
r 14
days
[D-1
4]) a
nd o
f re-
wat
erin
g ov
er 1
9 da
ys o
n m
orph
olog
ical
cha
ract
eris
tics o
f D. t
enui
folia
pla
nts.
Mea
ns w
ithin
a ro
w w
ith d
iffer
ent l
ette
r are
sign
ifica
ntly
diff
eren
t by
Tuke
y’s B
mul
tiple
com
paris
on te
st (*
P ≤
0.0
5; *
* P
≤ 0.
01)
At t
he en
d of
dro
ught
trea
tmen
tsA
t the
end
of 1
9 da
ys re
cove
ry
Cont
rol
D-4
D-8
D-1
1D
-14
Con
trol
D-4
D-8
D-1
1D
-14
Plan
t hei
ght (
cm)
68.2
7a67
.58a
68.0
0a57
.83a
b47
.7b
**-
Biom
ass f
resh
(g)
32.0
7a28
.79a
23.6
0a12
.38b
6.91
b**
22.3
2a14
.30a
b16
.24a
b13
.80a
b7.
18b
*
Biom
ass d
ry (g
)7.
73a
6.34
ab5.
68ab
5.05
b4.
65b
*7.
53a
5.70
a5.
71a
4.40
a4.
57a
No.
of l
eave
s21
.50a
20.1
7ab
18.5
0ab
10.5
bc1.
00c
**28
.33a
28.5
0a26
.33a
24.0
0a28
.00a
No.
of l
eave
s (≤10cm)
14.3
3a14
.00a
12.3
3a7.
50ab
0.00
b**
26.0
0a17
.00a
25.3
3a22
.00a
9.33
a
No.
of l
eave
s (>1
0cm
)7.
17a
6.17
ab6.
17ab
3.00
bc1.
00c
**2.
33a
2.00
a1.
00a
2.00
a0.
00a
No.
of l
eave
s (de
ad)
3.83
c4.
00c
3.33
c13
.67b
27.0
0a**
18.0
0a11
.00a
18.3
3a11
.33a
23.0
0a

Chapter V
67
Tab
le V
-2. E
ffec
t of
diff
eren
t dro
ught
trea
tmen
ts (
no ir
rigat
ion
for
4 [D
-4],
8 [D
-8],
11 [
D-1
1] o
r 14
day
s [D
-14]
) an
d of
re-
wat
erin
g ov
er 1
9 da
ys o
n th
e flo
ral c
hara
cter
istic
s of D
. ten
uifo
lia.
Mea
ns w
ithin
a r
ow w
ith d
iffer
ent l
ette
r ar
e si
gnifi
cant
ly d
iffer
ent b
y Tu
key’
s B
mul
tiple
com
paris
on te
st (
* P
≤ 0.
05; *
* P
≤ 0.
01).
Plan
ts o
f D. t
enui
folia
und
er th
e D
-14
drou
ght t
reat
men
t did
not
hav
e an
y op
en
Att
heen
dof
drou
ghtt
reat
men
tsA
t the
end
of 1
9 da
ys re
cove
ry
Con
trol
D-4
D-8
D-1
1D
-14
Con
trol
D-4
D-8
D-1
1D
-14
Tota
l flo
ral a
rea (
cm2 )
50.2
3a36
.93a
b25
.54b
c15
.75c
d-
**10
.01a
9.08
a4.
82a
9.56
a1.
96a
Flow
er a
rea
(cm
2 )1.
55a
1.47
a1.
46a
1.08
b-
**1.
20a
1.16
a1.
24a
1.48
a0.
84a
Flow
er d
iam
eter
(cm
)2.
22a
2.16
a2.
12a
1.84
b-
**19
.39a
19.0
0a19
.78a
21.6
7a16
.34a
Coro
lla tu
be d
iam
eter (
cm)
0.41
a0.
40a
0.40
a0.
32b
-**
3.44
a3.
22a
3.44
a3.
83a
2.67
a
Coro
lla tu
be le
ngth
(cm
)0.
41a
0.41
a0.
40a
0.32
b-
**4.
00a
4.11
a4.
33a
4.44
a3.
33a
No.
stem
s9.
17a
6.00
ab6.
17ab
5.20
b5.
83ab
*12
.33a
7.67
a8.
33a
6.67
a12
.00a
No.
flow
ers (
sene
scen
t)69
.17a
51.8
3a66
.83a
66.0
0a62
.83a
267.
00a
124.
67a
196.
00a
150.
33a
75.6
7a
No.
flow
ers (
open
)32
.17a
25.1
7ab
17.3
3b15
.00b
-**
8.00
a7.
33a
3.67
a6.
33a
7.00
a

Chapter V
68
The concentration of sugars (glucose, fructose and sucrose) in nectar was
measured directly after the drought treatments as well as after the recovery period.
Although there is an apparent tendency for concentration of glucose and fructose to
decrease in the plants exposed to longer period without irrigation, no significant
differences were found between the treatments (Fig. V-2A). However, this tendency did
not remain after re-watering the plants for 19 days (Fig. V-2B).
Fig. V-2. Concentration of sugars in nectar after different drought treatments (no
irrigation for 4 [D-4], 8 [D-8], 11 [D-11] or 14 days [D-14]) (A) and after re-watering of
19 days (B).
No significant differences were found between drought treatments. The vertical bars represent standard
errors. Concentrations of sugars in D-14 after re-watering are measured in only one plant that recovered
after drought stress.

Chapter V
69
Effect of drought on pollinator visits
Due to the different environmental conditions between two sites (“La Poveda”
and “Puente del Perdón”), there was some variation in the species of pollinators visiting
the flowers of D. tenuifolia. In “La Poveda”, flowers of D. tenuifolia were mainly
visited by bees (Anthophora spp., Andrena spp., and Lassioglossum spp.) and hoverflies
(Sphaerophoria rueppellii and/or S. scripta, Scaeva sp. and Syrphus sp.). In “Puente del
Padrón” both groups, hoverflies (Sphaerophoria rueppellii and/or S. scripta, Eupeodes
corollae, Eristalis tenax, Melanostoma sp. and Episyrphus balteatus) and bees (Apis
melifera, Megachile spp., Anthophora spp., Andrena spp., and Lassioglossum spp.)
were more diverse.
At the site “La Poveda”, the mean number of bees visiting the flowers of D.
tenuifolia was significantly higher in control plants and D-4 plants than in plants that
were under longer drought (F3,4=16.94, P ≤ 0.01). However, the overall number of
hoverflies was low and no significant differences were found between treatments (Fig.
V-3A). Also, there was a strong positive correlation between number of flowers and
number of visits by bees/min in the first experimental site (r=0.92, n=8, P ≤ 0.01), while
there was no significant correlation between number of flowers and number of
hoverflies visits (r=0.67, n=8, P = 0.07).

Chapter V
70
Fig. V-3. Number of pollinators visiting flowers of D. tenuifolia after different drought
treatments (no irrigation for 4 [D-4], 8 [D-8] and 11 days [D-11]) in two sites of Central
Spain: La Poveda (A) and La Sierra (B).
For each treatment, means that are significantly different at P ≤ 0.05 are marked with different letter
(comparison within insect groups), as determined by the Tukey’s B multiple comparison test. The vertical
bars represent standard errors.
At the site “Puente del Perdón”, the mean number of bees visiting the flowers of
D. tenuifolia did not significantly differ between drought treatments (Fig. V-3B).
Nevertheless, hoverflies apparently distinguished between different plants, visiting less
the plants under drought (F3,8=7.29, P ≤ 0.01). There was no significant correlation

Chapter V
71
between number of flowers and number of visits by bees (r=0.39, n=12, P = 0.21) and
hoverflies (r=0.55, n=12, P = 0.07).
3.2. Response of D. tenuifolia to RDI
Effect of RDI on plant growth
Regulated deficit irrigation (RDI) applied before and during flowering
significantly reduced gs in the D. tenuifolia plants (Fig. V-4). The gs of control plants
averaged 330 and 300 mmolm-2s-1 before and during flowering, respectively, while gs of
the plants under the MDI and SDI were constantly lower than 150 mmolm-2s-1. On the
first day of the experiment, there were no significant differences between treatments,
but 4 and 6 days after RDI was applied, values of gs in MDI and SDI were significantly
lower than in the control plants. In general, no significant differences in gs were found
between MDI and SDI.

Chapter V
72
Fig. V-4. Mean stomatal conductance of D. tenuifolia under moderate deficit irrigation
(MDI) and severe deficit irrigation (SDI). Deficit irrigation applied before and during
flowering (A) and deficit irrigation applied only during flowering (B).
For each measurement, means with different letter are significantly different at P ≤ 0.05, as determined by
the Tukey’s B multiple comparison test.
Applied before flowering, SDI significantly reduced height (F2,21=6.18, P ≤ 0.01),
total leaf area (F2,17=43.63, P ≤ 0.01), fresh biomass (F2,17=23.36, P ≤ 0.01) and dry
biomass (F2,17=6.98, P ≤ 0.01) of the plants, while the number of dead leaves increased
after 30 days of SDI (F2,21=3.54, P = 0.12). However, no significant differences were
found between control and MDI plants (Table V-3).

Chapter V
73
Table V-3. Effect of different regulated deficit irrigation (RDI) treatments (moderate
deficit irrigation before flowering (MDI-BF) and during flowering (MDI-DF); and
severe deficit irrigation before flowering (SDI-BF) and during flowering (SDI-DF) on
the vegetative characteristics of D. tenuifolia. Measured after the treatments were done.
RDI (16 days BF + 14 days DF) RDI (14 days DF)
Control MDI-BF SDI-BF Control MDI-DF SDI-DF
Plant height (cm) 43.50a 46.00a 33.40b ** 43.50a 42.17a 36.83a
Biomass fresh (g) 22.81a 27.35a 8.14b ** 18.33a 20.67a 13.80b **
Biomass dry (g) 2.62a 2.59a 1.67b ** 2.62a 3.14a 2.60a
No. leaves (tot) 35.25a 32.63a 26.88a 31.83ab 37.67a 25.70b **
No. leaves (≤10cm) 28.75a 26.25a 21.88a 24.83a 33.50a 24.67a
No. leaves (>10cm) 6.50a 6.38a 5.00a 7.00a 4.83a 1.00b **
No. leaves (dead) 2.00b 4.25ab 4.88a * 2.67c 10.70b 17.70a **
Total leaf area (cm2) 219.22a 206.44a 140.11b ** 219.22a 214.05a 103.60b **
Means within a row with different letter are significantly different by Tukey’s B multiple comparison test
(* P ≤ 0.05; ** P ≤ 0.01)
Applied during flowering, SDI significantly reduced fresh biomass (F2,15=13.32, P
≤ 0.01), number of big leaves (F2,15=11.41, P ≤ 0.01), number of dry leaves
(F2,15=27.26, P ≤ 0.01) and total number of leaves (F2,15=6.08, P ≤ 0.01), as well as total
leaf area (F2,15=9.56, P ≤ 0.01). In contrast, MDI significantly increased the number of
dry leaves when applied during flowering (Table V-3).
Effect of RDI on floral characteristics
While SDI applied before flowering influenced significantly growth and
development of the vegetative parts of D. tenuifolia, it affected only the total floral area
(F2,17=4.78, P = 0.02) (Table V-4).

Chapter V
74
Table V-4. Effect of different regulated deficit irrigation (RDI) regimes (moderate
deficit irrigation before flowering (MDI-BF) and during flowering (MDI-DF); and
severe deficit irrigation before flowering (SDI-BF) and during flowering (SDI-DF) on
the floral characteristics of D. tenuifolia. Measured after the treatments were done.
RDI (16 days BF + 14 days DF) RDI (14 days DF)
Control MDI-BF SDI-BF Control MDI-DF SDI-DF
Total floral area (cm2) 18.13ab 20.17a 9.06b * 18.13a 12.18ab 8.00b *
Flower area (cm2) 1.78a 1.66a 1.42a 1.78a 1.66a 1.14b **
Flower diameter (cm) 2.19a 2.07a 1.88a 2.21a 2.13ab 1.90b *
Corolla tube diameter (cm) 0.38a 0.39a 0.34a 0.38a 0.36a 0.34a
Corolla tube length (cm) 0.41a 0.34a 0.36a 0.41a 0.36a 0.30b **
No. stems 3.88a 4.25a 2.50a 3.33a 2.67a 2.50a
No. flowers (senescent) 25.13a 28.13a 18.63a 20.17a 13.67a 17.17a
No. flowers (open) 10.00a 12.29a 6.43a 10.00a 7.50a 6.67a
Means within a row with different letter are significantly different by Tukey’s B multiple comparison test
(* P ≤ 0.05; ** P ≤ 0.01)
However, when applied during flowering, SDI caused significant reduction of
total floral area (F2,15=3.50, P ≤ 0.05), flower area (F2,15=8.40, P ≤ 0.01), flower
diameter (F2,15=5.53, P = 0.02), and corolla tube length (F2,15=10.32, P ≤ 0.01). Instead,
MDI (when applied before and during flowering) did not have an effect on the number
and characteristics of D. tenuifolia flowers compared to control plants (Table V-4).
Effect of RDI on pollinator visits
At the University of Reading (UK) site, both groups of pollinators (bees and
hoverflies) were detected. Of all bees, two species of bumblebees were the most
frequent visitors to the flowers of D. tenuifolia (Bombus terrestris and Bombus
pascuorum). Conversely, the group of hoverflies was more diverse including four
genera (Sphaerophoria sp., Eupeodes sp., Episyrphus sp. and Eristalis sp.).

Chapter V
75
Fig. V-5. Number of pollinators visiting flowers of D. tenuifolia after moderate deficit
irrigation (MDI) and severe deficit irrigation (SDI). Deficit irrigation applied before and
during flowering (A) and deficit irrigation applied only during flowering (B).
For each treatment, means with different letter are significantly different at P ≤ 0.05 (comparison within
insect groups), as determined by Tukey’s B multiple comparison test. The vertical bars represent standard
errors.
Figure V-5 illustrates the mean number of visits of pollinators/min to the flowers
of D. tenuifolia under different deficit irrigation treatments. Irrespective of whether
MDI or SDI, were applied before or during flowering, there was no significant effect on
the attractiveness of the plants to bees and hoverflies (Fig. V-5). However, no
significant correlations between number of flowers and visits of bees (r=0.06, n=30, P =
0.74) and hoverflies (r=-0.07, n=30, P = 0.71) were found.

Chapter V
76
3.3. Water consumption
In order to assess the most suitable irrigation treatment for D. tenuifolia, the total
water consumption was calculated for each treatment for 30 days (Table V-5).
Table V-5. Total water consumption per plant for irrigation of potted D. tenuifolia
during 30 days experiment after drought treatments (D) of 4 [D-4], 8 [D-8], 11 [D-11]
or 14 days [D-14]) and regulated deficit irrigation (RDI) treatments (moderate deficit
irrigation before flowering [MDI-BF] and during flowering [MDI-DF]; and severe
deficit irrigation before flowering [SDI-BF] and during flowering [SDI-DF]
Treatment
Water deficit No. of days
under 100% ETp
irrigationa
Total water
consumption/plant
(g)
Water
saved/plant
(%)
No.
days
Water
(g/day)
D-control - - 30 1467.6 0
D-4 4 - 26 1271.9 13
D-8 8 - 22 1076.2 27
D-11 11 - 21 929.5 37
D-14 14 - 16 782.7 47
RDI-control - - 30 1129.6 0
MDI-BF 30 22.6 - 677.7 40
SDI-BF 30 11.2 - 336.9 70
MDI-DF 11 22.6 19 963.8 15
SDI-DF 11 11.2 19 838.9 26 a 100% ETp irrigation (replacing all the water lost by evapotranspiration in the previous 24 h) in drought
treatments was 48.92 g/day; while 100% ETp in RDI treatments was 37.65 g/day
The maximal water savings were measured for MDI applied before flowering,
while the minimal water savings were measured in plants that were without irrigation
for 4 days (D-4 treatment) and MDI applied during flowering (Table V-5).

Chapter V
77
4. Discussion
This study shows that drought periods of 11 and 14 days can reduce the gs to 0
mmolm-2s-1 in 33% and 100% of D. tenuifolia plants, respectively. However, after 19
days of re-watering most plants had gs values increased to the control plants’ values,
except in 67% of D-14 plants that died. In plants that survived, drought stress of 11 and
14 days negatively influenced biomass and number of leaves, while no significant
differences were found between control, D-4 and D-8 plants. Regardless of whether
MDI was applied before or during flowering, it did not affect plant growth. In contrast,
SDI applied before flowering significantly reduced biomass, total leaf area and plant
height, which are all important parameters for commercial production of D. tenuifolia
(Znidarcic et al., 2011). When applied during flowering, SDI significantly reduced
biomass, total leaf area, total number of leaves, and caused a significant decrease in
number of big leaves. Indeed, the negative effect of SDI on plant growth was less
evident when applied before flowering. If produced as a crop under greenhouse
conditions, D. tenuifolia can be under drought stress for no more than 8 days, or MDI
can be applied without further negative effects on its commercial production. However,
if produced under field conditions, additional experiments would be necessary as
selection of an irrigation regime depends on the local climate (McDonald and Girvetz,
2013).
On the other hand, if the main purpose of D. tenuifolia is to attract pollinators,
water deficit should not affect quantity and quality of its flowers. The results of this
study show that all floral parameters, except sugar concentration in nectar, were
negatively affected by drought of 11 or more days. These findings agree with Carroll et
al. (2001), who also reported that drought stress of 12 days did not affect the
concentration of sugars in the nectar of Epilobium angustifolium L. and with Cerekovic
et al. (2013), reporting that drought stress of 12 days negatively affect development of
leaves and flowers of blackcurrant (Ribes nigrum L.). In addition, this research shows
that flower development in D. tenuifolia is more sensitive to drought stress than
vegetative development, as drought of 8 days did not affect any of the measured
vegetative parameters, while it reduced the number of open flowers and the total floral
area. On the other hand, MDI and SDI did not influence floral traits when applied

Chapter V
78
before flowering. However, when applied during flowering, SDI reduced floral area. As
in other studies conducted with other plants (Mingeau et al., 2001; Cakir, 2004; Alvarez
et al., 2013), these results show that D. tenuifolia responds differently to regulated
deficit irrigation and water stress depending on the phenological stage of the plant.
Since pollinator visits are correlated with the number of open flowers and their
total floral area (Conner and Rush, 1996), our results suggest that, under the
temperature and relative humidity conditions of this experiment, D. tenuifolia could
tolerate drought for 4 days without affecting its attractiveness to pollinators.
Additionally, direct observations of pollinators’ visits to water-stressed plants were
included in this study, confirming that the number of flowers can affect flower visitation
by pollinators (Eckhart, 1992; Conner and Rush, 1996; Caraballo-Ortiz et al., 2011) as
bees and hoverflies preferred to visit plants that had more opened flowers. This could be
the main reason why there was no significant difference in the frequency of visits by
pollinators to flowers of D. tenuifolia after RDI treatments, which did not affect the
number of open flowers per plant. Nevertheless, the positive correlation between
number of pollinator visits/min and number of flowers was not significant when the
abundance of bees and hoverflies was low.
The simultaneous assessment of vegetative and floral parameters under different
regimes of RDI and drought stress showed that RDI treatments caused less of a
disturbance to the plant than 11 or 14 days of drought stress. Although not many studies
have compared those two water deficit treatments, similar findings were found by Luo
et al. (2007) and Cuevas et al. (2007) when measuring the effect of both, drought stress
and RDI, on the growth and flowering of Eriobotrya japonica Lindl. Additionally,
Cuevas et al. (2007) showed that MDI applied before flowering positively influenced
the flowering of E. japonica, while if applied during flowering, it restricted the normal
flower development.
Application of MDI before flowering was the most efficient water-saving
technique for commercial production of D. tenuifolia, as it did not reduce plant growth
and saved up to 40% of irrigation water. The longest drought period tested (D-14)
reduced up to 50% the consumption of irrigation water, but resulted in death of most of
the plants even after irrigation was reinstated. Although reducing vegetative growth,

Chapter V
79
SDI before flowering could save water consumption by 70% without affecting
flowering and plant attractiveness of D. tenuifolia to pollinators.
In conclusion, in the environmental conditions of this study, potted D. tenuifolia
can cope with drought of 4 days without having any consequences on its growth,
flowering and attractiveness to pollinators. However, a longer drought period can
negatively affect development of its flowers and reduce its vegetative growth.
Additionally, this study shows that RDI can improve water use efficiency. Depending
on the purpose of growing D. tenuifolia, water use can be reduced by 40% (if grown as
a crop) or by 70% (if grown to attract pollinators).

CHAPTER VI
Submitted to: Agricultural and Forest Entomology (Submitted: 23/07/2014)
Barbir, J., Badenes-Pérez, F.R., Fernández-Quintanilla, C., Dorado, J. (submitted).
Can proximity of floral field margins improve pollination and seed production in
Coriandrum sativum L. (Apiaceae)?

Chapter VI
81
VI. Can proximity of floral field margins improve pollination and
seed production of coriander crop, Coriandrum sativum L.
(Apiaceae)?
1. Introduction
Coriander, Coriandrum sativum L. (Apiaceae), is a native plant in Mediterranean
region, where it is mainly grown in a small-scale private gardens or fields (Carrubba et
al., 2006). However, its cultivation has been widespread in a lot of regions all over the
world (Western Europe, South and North America, India) and its production has
become large-scaled (Maroufi et al., 2010). Coriander has been cultivated for its fruits
(hereinafter referred to as “ seeds”), leaves and oils (Maroufi et al., 2010; Nadeem et al.,
2013), characterized by high antioxidant activity (Wangensteen et al., 2004) desirable in
healthy human diet.
Insect pollination is necessary for production of seeds in coriander, even though
some pollen can be transmitted from one flower to another by wind (Koul et al., 1989;
Bendifallah et al., 2013). As flowers of coriander are characterized for their exposed
nectaries, abundant pollen production and zygomorphic flowers (Koul et al., 1989;
Sinacori et al., 2014), they are attractive to flies (mainly hoverflies), bees and beetles
(Colley and Luna, 2000; Bendifallah et al., 2013; Barbir et al., 2014a). However,
foraging of pollinators in C. sativum flowers is limited due to its short bloom period (20
to 40 days, depending on the region). Therefore, there is a need to complement the
agricultural landscapes with additional floral sources where pollinators can forage
before and after C. sativum blooming.
Intensification of agriculture has reduced the availability of forage sources for
pollinators (Tscharntke et al., 2005; Winfree et al., 2009; Le Feon et al., 2010), leading
to the decline in abundance and diversity of pollinators in agro-ecosystems (Biesmeijer
et al., 2006). This lack of pollinators in agricultural fields can cause poor pollination of
crops, as well as a significant reduction in crop yields (Richards, 2001; Garibaldi et al.,
2009). Therefore, in order to buffer this impact of intensive agriculture and to conserve
wild pollinators within their natural habitats, it has been recommended to introduce

Chapter VI
82
floral margins, as additional forage and shelter sources to pollinators, within mono-
culture landscapes (Landis et al., 2000; Marshall, 2005; Pywell et al., 2011; Rands and
Whitney, 2011).
Many studies evaluated the importance of field margins in conservation of the
beneficial insects in general (Landis et al., 2005; Olson and Wackers, 2007; Haaland et
al., 2011) or natural enemies (Tinkeu et al., 1996; Steingrover et al., 2010) and
pollinators (Pywell et al., 2011; Nicholls and Altieri, 2013; Scriven et al., 2013) in
particular. In addition, some studies directly estimated how implementation of field
margins reflected on the crop production by comparing yield/ha in the fields with and
without floral margins (Carvalheiro et al., 2012). However, when assessing the effect of
field margins on crop production in large-scale studies, the possibility for additional
external factors (i.e. rainfall and temperature, physical and chemical properties of soil or
plant development) biasing the results, is high. Therefore, the assessment of the
relationship between yield, presence of field margins, and visits of pollinators to flowers
in margins and crop should be preformed, excluding major external factors that can
influence plant development and yield production. These factors can be minimized by
potting plants of coriander (Ghazoul, 2006) and reducing the scale of the study, and in
that way studying only a direct relation between pollinators visits and seed production.
The selection of coriander as a target crop for this experiment was based on its
requirement for insect pollination to produce seeds (even though some pollen can be
transmitted from one flower to another by wind) as well as the easy way to estimate its
total seed production under semi-field conditions.
In the previous study it has been shown how Diplotaxis tenuifolia L. DC. is
equally attractive for two groups of the most abundant pollinators in Central Spain, i.e.
bees and hoverflies, (Barbir et al., 2014a). Accordingly, it has been hypothesized that a
margin with this floral species can attract as much pollinators and improve seed
production in coriander to the same extend as a mixed margin including various plant
species. Hence, the main objective of this study was to estimate the impact of field
margins on seed production of coriander, where two different types of field margins
were tested: mono-specific margin (D. tenuifolia) and mixed margin (six annual
herbaceous species). For that reason, it was tested: i) if open pollination, i.e. control
(without field margins), vicinity of mono-specific margins and mixed margins,
increases the seed production of coriander when compared with no-pollination, i.e.

Chapter VI
83
covered inflorescences of coriander, under field conditions; ii) if frequency of pollinator
visitation to the flowers of coriander was higher in the presence of field margins than in
control; and iii) if the crop seed production was higher in presence of field margins than
in control without floral margins.
2. Material and methods
2.1. Study area and experimental design
This study was conducted in the spring of 2013 at the experimental farm La
Poveda (40º 19' N and 3º 29' W, elevation 536 m) sited in Arganda del Rey (Madrid,
Spain). The study area is characterized by a continental Mediterranean climate with cold
winters and hot summers and very scant precipitation (about 400 mm per year). In
general, floral areas are scarce within the experimental farm, as in the last four years
maize, barley and poplars were mainly produced. The experiments were randomly
distributed within those fields.
In order to test how the proximity of different field margins affects visits of
pollinators and seed production in coriander, three different treatments were applied: the
vicinity of mixed margin (mixture of six herbaceous species), mono-specific margin (D.
tenuifolia) and control (absence of floral margin). Mixed margin (10 m × 1.5 m),
consisting of six herbaceous species attractive to bees and hoverflies in Central Spain
(Phacelia tanacetifolia Benth., Echium plantagineum L., D. tenuifolia, Centaurea
cyanus L., Calendula arvensis L. and Borago officinalis L.), was sown in fall 2012.
This floral mixture has been found as efficient and attractive to pollinators in the region
(Barbir et al., 2014a).The mono-specific margins of D. tenuifolia were planted in
February 2013, where 18 plants were planted in a row, with the separation of 0.5 m
between them. From the previous experience with D. tenuifolia this distribution of the
plants forms 10 m × 1.5 m margin (J. Barbir, unpublished results). Each experiment
with a different type of margin was separated from others at least 200 m, while
minimum separation between three repetitions within each treatment was 20 m.
In February 2012, seedlings of coriander were transplanted to pots (10 cm × 13
cm) filled with 520 g of a mixture of peat-based compost and vermiculite (3:1 by

Chapter VI
84
volume), irrigated to container capacity and distributed over tables in the greenhouse.
The minimum temperature in the greenhouse was 18-20 oC and the maximum
temperature was 25-29 oC. The relative humidity varied from 50% to 80%. In April, one
month before blooming, plants of coriander were transported to the field. Fifteen plants
of coriander were organized in a row (separation between plants was 0.5 m) at 1.5 m
distance from the margins and were buried in the ground together with their pots. The
plants were potted in order to avoid the differences in soil properties which could affect
crop development and seed production (Moore and Lawrence, 2013). During the
experiment, each potted plant was irrigated 4 times/week with 0.5 l of water and each
margin was irrigated 4 times/week with 10 litres of water each time.
2.2. Visitation of pollinators
In order to estimate the attractiveness of the margins to pollinators and its effect
on the pollination of coriander, the mean number of visits of bees and hoverflies per
minute (Campbell et al., 2012a) was counted in the flowers of the margins and
coriander plants. Insect visits to the flowers of the margins were counted twice per week
(9 min/margin) in three marked areas of 0.5 m × 0.5 m (3min/marked area), located in
the centre and both ends of each margin. Insect visits to the flowers of coriander were
also counted twice per week in four randomly chosen plants/row (3 min/plant). Visits
were counted only when the pollinator had a direct contact with the reproductive
structures of the flower (i.e., stamens and pistils). The observations were conducted
always in the morning (9.00 h – 13.00 h local time, Greenwich Mean Time + 2:00)
when the weather conditions were preferable for the insects to fly and visit flowers
(sunny days without wind).
2.3. Floral assessment
The flower density (number of flowers) was counted every observation day within
the marked area (0.5 m × 0.5 m) of all margins in order to investigate its influence on
the attractiveness of the plants (Stout et al., 1998). Each capitulum of species belonging
to the Asteraceae family (i.e., C. cyanus and C. arvensis) was considered as an
individual flower. In addition, the total number of flowers/plant in coriander was

Chapter VI
85
estimated by multiplying the mean number of flowers in 10 inflorescences randomly
chosen and the total number of inflorescences found in the plants selected for
observation of insect visits.
2.4. Seed production
In order to study how the presence of different field margins can affect pollination
and seed production in coriander, potted plants of coriander were managed, i.e.
regularly irrigated, under field conditions until all seeds were formed. At harvest time,
all 135 plants were unburied from the ground and transported to the laboratory where
the seeds were counted and weighted. Differently than in other studies which measured
the number of seeds/plant to estimate the efficiency of insect pollination in coriander
(Al-Fattah, 2001; Chaudhary and Singh, 2007), this study used number of
seeds/inflorescence because the number of stems and inflorescences can vary from one
plant to another. Therefore, for each plant, number of inflorescences, number of formed
seeds/inflorescence (actual SN/inf) and number of dry flowers/inflorescence (flowers
that did not develop its fruit) were counted in order to estimate the percentage of actual
seed production in coriander. Sum of dry flowers/inflorescence and actual SN/inf
represented potential of one inflorescence to produce seeds (potential SN/inf).
Seed production in coriander with and without open pollination
In order to estimate the grade of auto pollination in coriander and to study how
different treatments of open pollination, i.e. control (without margins), vicinity of mixed
margins and mono-specific margins, and no pollination can affect its seed production, a
minimum of three inflorescences per plant were covered to prevent insect pollination.
Therefore, three inflorescences per plant were covered 3-5 days before flowering while
other three inflorescences (at the same plant and its stage of development) were marked
with a blue string as not-covered. In total 220 inflorescences were covered (no
pollination) and 220 were not-covered (open pollination).

Chapter VI
86
Seed quality
Seed weight (SW). Seeds from each plant were collected, counted and weighted
using a precision balance. The mean weight per seed was calculated for each treatment.
In addition, the seed weight/plant (SW/plant) was used as a parameter to evaluate
whether the presence of floral margins improves the pollination in coriander, by
calculating the percentage increase compared to control without margins.
Germination rate of coriander seeds. This parameter has been used to estimate
the seed quality (Putievsky, 1980). Since there were no more than 30 seeds in some
replications of covered inflorescences, the sample size in germination tests including
covered inflorescences was limited to 10 seeds per petri dish in order to perform three
repetitions per treatment. In contrast, the germination tests that did not include covered
inflorescences (i.e., the comparison between controls without margin and crops with
margins) did not have this limitation and therefore a greater number of seeds (e.g., four
replications with 25 seeds per petri dish) were used.
2.5. Statistical analysis
The data were analysed with SPSS® Statistics19 software (IBM, Chicago, IL,
USA). A simple general linear model (GLM) was used in order to assess the ability of
different insects to discriminate between flower species. The mean value of insect visits
per minute within the observation area was used as response variable for these analyses.
Prior to an ANOVA analysis, the assumptions of normal distribution (Kolmogorov
Smirnov test) and homogeneity of variances (Levene’s test) were tested. Differences
among means were explored by Tukey’s B post hoc test. Univariate ANOVA was also
used to test whether there were significant differences in seed production and seed
germination rate between the treatments. Finally, a bivariate two-tailed Pearson analysis
was used to assess the correlation between floral density and visits of
pollinators/minute.

Chapter VI
87
3. Results
3.1. Visits of pollinators to the flowers
Visits of pollinators to the flowers of margins
The results showed no significant differences in the mean number of opened
flowers per marked area between mono-specific and mixed margins (Table VI-1).
However, frequency of pollinator visits was significantly higher in mono-specific than
in mixed margins for all groups of pollinators. Indeed, the results showed that visits of
bees (F1,34 = 10.54, P ≤ 0.01), hoverflies (F1,34 = 5.48, P ≤ 0.01) and beetles (F1,34 =
12.09, P ≤ 0.001) were more frequent in mono-specific margins than in mixed margins.
Consequently, the mean number of visits/minute of total pollinators was significantly
higher (F1,34 = 12.45, P ≤ 0.001) in the mono-specific margins than in mixed margins
(Table VI-1).
Table VI-1. Mean (± S.E.) flower density and mean (± S.E.) number of visits of
pollinators to the flowers of mono-specific and mixed floral margins
Mono-specific Mixed
No. flowers/marked area 38.44 ± 2.73a 41.60 ± 9.63a
No. beetles/min*** 0.68 ± 0.17a 0.08 ± 0.02b
No. bees/min*** 0.51 ± 0.17a 0.13 ± 0.05b
No. hoverflies/min** 0.36 ± 0.17a 0.16 ± 0.05b
Total No. pollinators/min*** 1.55 ± 0.17a 0.37 ± 0.08b
***significant differences between the treatments at P≤ 0.001; the same letter in a row represents no
significant differences between treatments
Visits of pollinators to the flowers of coriander
The results showed that number of inflorescences/plant and number of
flowers/plant of coriander did not significantly differ between treatments (Table VI-2).

Chapter VI
88
Table VI-2. Mean (± S.E.) flower density and mean (± S.E.) number of visits of
pollinators to the flowers of coriander without proximate floral margins (control) and in
vicinity of mono-specific and mixed margins
Control Mono-specific Mixed
No. inflorescence/plant 33.10 ± 4.42a 35.07 ± 3.14a 36.50 ± 3.66a
No. flowers/plant 546.15 ± 72.86a 578.60 ± 51.85a 602.25 ± 60.46a
No. beetles/min*** 0.07 ± 0.02b 0.52 ± 0.14a 0.87 ± 0.25a
No. bees/min 0.11 ± 0.04a 0.12 ± 0.05a 0.30 ± 0.12a
No. hoverflies/min*** 0.06 ± 0.03b 0.58 ± 0.12a 0.46 ± 0.15a
Total No. pollinators/min*** 0.24 ± 0.06b 1.22 ± 0.19a 1.63 ± 0.36a
***significant differences between the treatments at P≤ 0.001; the same letter in a row represents no
significant differences between treatments
However, number of beetles/min, number of hoverflies/min and number of total
pollinators/min was significantly higher in mono-specific and mixed margins treatments
(F2,42 = 5.82, P ≤ 0.01; F2,42 = 5.97, P ≤ 0.01; F2,42 = 9.06, P ≤ 0.001) than in control
without margins.
Correlations between number of flowers and number of pollinators in coriander
plants and margins
Significant correlations between number of flowers and number of visits of
pollinators/min were found for mono-specific and mixed margins treatments (Table VI-
3). In contrast, no significant correlations were found for coriander plants in the control
treatment.
The number of flowers in mixed margins was positively correlated with the
number of pollinators visiting flowers of both, mixed margins and coriander in the
vicinity of mixed margins. Therefore, these results suggest an increase in the number of
pollinators visiting flowers of mixed margins with the increase in number of flowers of
coriander and vice versa. On the other hand, a strong positive correlation was found
between number of flowers in mono-specific margins and number of pollinators visiting
them.

Chapter VI
89
Table VI-3. Correlations between number of flowers and number of pollinators'
visits/min
No. pollinators
No. flowers
Coriander Margins
Control Mono-specific Mixed Mono-specific Mixed
Coriander
Control 0.291 - - - -
Mono-specific - 0.145 - - 0.571* -
Mixed - - 0.376 - 0.522*
Margins Mono-specific - 0.219 - 0.695** -
Mixed - - 0.528* - 0.552*
*significant correlation at P≤ 0.05; **significant correlation at P≤ 0.01; (-) no correlation
Although no significant correlation was found between number of flowers in
mono-specific margins and visits of pollinators to the adjacent coriander flowers, a
negative correlation between number of coriander flowers and number of pollinators
visiting flowers of mono-specific margins was detected (Table IV-3).
3.2. Seed production in coriander
Seed production in coriander with and without pollination
The results in Table VI-4 showed that the number of seeds/inflorescence formed
in plants in the control was higher than in plants in the mono-specific and mixed margin
treatments (F2,217 = 15.43, P ≤ 0.001).
Table VI-4. Potential (± S.E.), actual (± S.E.) and percentage (± S.E.) of actual seed
number (SN) per inflorescence in covered inflorescences of coriander under control,
mono-specific and mixed margin treatments
Control Mono-specific Mixed
Potential SN/inf.*** 15.68 ± 0.46a 12.54 ± 0.31c 14.38 ± 0.34b
Actual SN/inf.*** 1.28 ± 0.23a 0.20 ± 0.06b 0.42 ± 0.11b
Actual SN/inf. (%)*** 7.99 ± 1.45a 1.59 ± 0.52b 2.85 ± 0.72b
***significant differences between the treatments at P≤ 0.001; the same letter in a row represents no
significant differences between treatments
In addition, the total potential of inflorescences to produce seeds was significantly
higher in the control plants of coriander than in coriander plants in vicinity of the mixed

Chapter VI
90
and the mono-specific margins (F2,217 = 18.73, P ≤ 0.001). As these inflorescences of
coriander were covered and insects did not get in contact with the flowers, consequently
actual percentage of coriander seed number per inflorescence was higher in control than
in the other two treatments (F2,217 = 12.81, P ≤ 0.001) (Table VI-4).
Considering the effect of insect pollination, the results showed a significantly
lower number of seeds produced in the covered inflorescences of coriander than in
inflorescences exposed to open pollination, e.g. control, mono-specific and mixed
margin treatments (Table VI-5). Significant differences were also found between open
pollination treatments, where coriander plants under control and mono-specific margins
treatments produced significantly less seeds/inflorescence than the plants located next to
mixed margins (F2,205 = 4.85, P ≤ 0.01).
Table VI-5. Potential (± S.E.), actual (± S.E.) and percentage (± S.E.) of actual seed
number (SN) per inflorescence of coriander in not-covered (control, mono-specific and
mixed margin treatments) vs. covered inflorescences
Control Mono-specific Mixed Covered inf.
Potential SN/inf.*** 16.13 ± 0.31a 13.55 ± 0.31c 14.78 ± 0.34b 14.10 ± 0.23bc
Actual SN/inf.** 2.57 ± 0.28b 2.83 ± 0.28b 4.00 ± 0.38a 0.60 ± 0.09c
Actual SN/inf. (%)** 15.86 ± 1.85b 20.15 ± 1.85b 25.68 ± 2.24a 3.99 ± 0.57c
**significant differences between the treatments at P≤ 0.01; ***significant differences between the
treatments at P≤ 0.001; the same letter in a row represents no significant differences between treatments
In contrast, the highest potential seed production was found in the control plants,
while the lowest was found in mono-specific margin treatment (F2,205 = 13.47, P ≤
0.001). Considering the relative value of actual production of seeds, the highest
percentage of actual seed number was found in coriander plants next to the mixed
margins (F2,205 = 5.32, P ≤ 0.01), while less than 4% of potential SN reached maturity in
treatments with covered inflorescences (Table VI-5).
Seed quality
After counting the potential and actual number of seeds per inflorescence in all
plants included in the experiment (4,426 inflorescences in total), significant differences

Chapter VI
91
in number of seeds produced per inflorescence were found between control, mono-
specific and mixed margin treatments (F2,4426 = 26.17, P ≤ 0.001) (Table VI-6).
Table VI-6. Potential (± S.E.), actual (± S.E.) and percentage (± S.E.) of actual seed
number (SN) per inflorescence of coriander, and seed weight (SW) per seed (± S.E.),
SW per plant (± S.E.) and percentage of SW per plant (± S.E.) under control, mono-
specific and mixed margin treatments
Control Mono-specific Mixed
Potential SN/inf.*** 16.68 ± 0.11a 14.89 ± 0.08b 14.96 ± 0.08b
Actual SN/inf.*** 4.06 ± 0.10c 4.60 ± 0.09b 5.04 ± 0.09a
Actual SN/inf. (%) *** 23.25 ± 0.55c 28.97 ± 0.48b 31.82 ± 0.49a
SW/seed (mg)*** 7.8 ± 0.9b 12.8 ± 0.8a 13.3 ± 1.0a
SW/plant (g)** 0.96b 2.12a 2.45a
SW/plant (%) 100.00 220.83 255.21
**significant differences between the treatments at P≤ 0.01; ***significant differences between the
treatments at P≤ 0.001; the same letter in a row represents no significant differences between treatments
The highest potential SN per inflorescence was found in control plants of
coriander (F2,4426 = 115.04, P ≤ 0.001), while no differences were found in coriander
plants under mono-specific and mixed margin treatments (Table VI-6). However, this
potential production did not translate into actual production, with significantly lower
values of percentage of SN per inflorescence in control plants than in coriander plants
close to mixed margins (the highest percentages) and those under mono-specific
treatment (F2,4426 = 65.83, P ≤ 0.001).
Seed weight. The results showed that the average weight of each seed was
significantly higher in coriander plants next to margins treatment than in control plants
(F2,4426 = 11.66, P ≤ 0.001), indicating better seed quality. Seed weight per plant was
also significantly higher in mono-specific and mixed margin treatments than in control
plants of coriander (F2,15 = 6.36, P ≤ 0.01), increasing the seed production for more than
220 % and 255 %, respectively (Table VI-6).
Germination rate of coriander seeds. Seed quality was higher in plants next to
floral margins since the results of germination tests showed higher values in the

Chapter VI
92
germination rate of these treatments relative to control plants (F2,21 = 10.76, P ≤ 0.001)
(Figure VI-1A).
Fig. VI-1. Germination rate of coriander seeds. Covered inflorescences of coriander
(covered) and not-covered inflorescences of coriander in vicinity of: mono-specific
margins (NO - D. tenuifolia), mixed margin (NO - mixed) and without margins (NO -
control). Means with same letter in each graph are not significantly different at P ≤ 0.05,
as determined by Tukey’s B multiple comparison test. Seed germination in: (A) sample
of 25 seeds x 4 repetitions and (B) sample of 10 seeds x 3 repetitions.

Chapter VI
93
In the second batch of seed germination tests including seeds from covered
inflorescences, the results showed similar values for seeds from plants next to the field
margins (mixed and mono-specific) (F3,20 = 9.59, P ≤ 0.001), while seed from covered
inflorescences, i.e. not exposed to open pollination, showed lower quality similar to the
control plants (Fig. VI-1B).
4. Discussion
Although many studies confirmed that number of pollinator visits increases with
higher density of flowers (Conner and Rush, 1996; Dauber et al., 2010; Essenberg,
2012; Barbir et al., 2014b), in this study no significant differences in flower density
were found between mono-specific and mixed margins. Still abundance of pollinator
visitors (bees, hoverflies and beetles) was significantly higher in mono-specific margins
than in mixed margins, which could be explained by the relatively higher attractiveness
of D. tenuifolia flowers to local pollinators compared to the floral mixture (Barbir et al.,
2014a). The attractive effect of floral margins to pollinators has also affected nearby
plants of coriander. Indeed, when comparing the open pollination treatments, i.e.
control, mono-specific and mixed margin, number of coriander flowers per plant did not
significantly differ, but the frequency of hoverfly and beetle visits to their flowers was
significantly higher in the vicinity of mono-specific and mixed margins than in control
plants without margins. However, no significant differences in bee visits/min were
found between the different treatments of coriander.
The possible explanation could be the different biology of these insect groups.
The most frequent beetles (Heliotaurus ruficolis L. and Psilothrix viridicoerulea L.) are
highly attracted to the species used in the margins (Barbir et al., 2014a) and prefer
diverse floral areas over bare soil (Woodcock et al., 2012). Hoverflies, unlike bees, do
not collect pollen and nectar for their offspring and disperse into landscapes in linear
manner, whereas bees return to their brood cells after foraging (Kleijn and van
Langevelde, 2006). On the other hand, most hoverflies, show an extremely specialized
selection of microhabitat sites for larval development, usually not found in intensively
managed agro-ecosystems (Jauker et al., 2009), which can explain higher abundance of
hoverflies in vicinity of floral margins than in control plants of coriander without

Chapter VI
94
proximate field margins and surrounded by bare soil. In addition, majority of bee
species in the study area are solitary-mining bees (Barbir et al., 2014a), which prefer to
nest in bare soil (Cane, 1997) and in vicinity of their host plants (Julier and Roulston,
2009). Since control plants of coriander were mainly surrounded by bare soil, it
probably contributed to the nesting of bees in surrounding area, which resulted in higher
abundance of bees in the control plants of coriander compared with other treatments.
Recently, many studies directly correlated the abundance and diversity of
pollinators with crop production under controlled conditions, i.e. cages or greenhouses
(Jauker and Wolters, 2008; Jauker et al., 2012) or in open fields by estimating yield/ha
(Carvalheiro et al., 2012). However, this study mixed those two methodologies, as it
was conducted under semi-controlled field conditions (potted plants and regulated
irrigation) in order to directly correlate insect visits with seed production by eliminating
the influence of some environmental factors, as for example the soil properties.
Therefore, a strong positive correlation between number of flowers in mixed margins
and pollinator visits to nearby plants of coriander has been found, confirming that visits
of pollinators were more frequent in the flowers of crop located next to the mixed
margins than in control without floral margins (Ghazoul, 2006).
On the other hand, the highly attractive mono-specific margins of D. tenuifolia did
not contribute to the spillover of insects from the margin to the crop flowers. However,
the negative correlation between number of coriander flowers and visits of pollinators to
the margin showed that when blooming of coriander ends, i.e. number of flower
decreases, pollinators switch flower-foraging from coriander flowers to D. tenuifolia
flowers. Therefore, although not contributing directly to the attraction of the crop to
pollinators, D. tenuifolia showed good potential in their conservation in the study area
(Barbir et al., 2014a).
In accordance with Jacobs et al. (2009), number of seeds/inflorescence and
percentage of seeds produced/inflorescence were significantly lower in covered
inflorescences than in open pollinated ones, thus demonstrating the essential role of
insect pollination in this crop. However, when comparing the number of seed produced
in the different treatments of open pollination, the highest number of coriander seeds per
inflorescence was found near mixed margins, while the lowest number of
seeds/inflorescence was achieved in control plants without margins, despite their

Chapter VI
95
significantly higher potential for seed production. Indeed, crop plants in the vicinity of
mixed margins produced 31% of their potential seed number, crop plants close to the D.
tenuifolia margins produced 29%, while control plants without margins produced only
23% of their potential seed number.
Average weight per seed was one of the measures for estimation of the seed
quality, which resulted significantly higher in both floral margin treatments than in the
control plants of coriander. In addition, when comparing seed production, i.e. seed
weight per plant, the results showed that when floral margins were present in the field,
the seed production of coriander increased by more than 200%. Although the number of
seeds/inflorescence was slightly lower in mono-specific treatment than in the mixed
margin treatment, it did not reflect on the differences in final seed production/plant as
the quality of seed was equally good. Another parameter for seed quality estimation was
germination rate of produced seeds. The seeds of coriander plants in the vicinity of
floral margins showed significantly higher germination rate than seeds produced in the
control treatment.
Finally, it can be concluded that the proximity of both floral margins (mixed and
mono-specific) improved the seed quality of coriander plants, as seed weight and
germination rate were significantly higher than in control plants. However, number of
seeds produced was significantly higher in the crop grown near mixed margins than
near mono-specific margin, probably due to stronger positive correlations in pollinator
visits to coriander flowers and number of margins’ flowers. As the experiment was
conducted under semi-controlled conditions, it can be concluded that pollinator visits
was the main factor that biased the results, and that presence of both mixed or mono-
specific (D. tenuifolia) margins improve the crop production for more than 200% in
small-scaled gardens and, in addition, conserve the local pollinators.

Chapter VII
96
VII. General discussion
Agricultural landscapes, dominated by large monocultures, have caused alarming
declines in farmland biodiversity, threatening the optimal functionality of agro-
ecosystems (Krebs et al., 1999; Benton et al., 2003) and leaving beneficial insects to
suffer from deficit of food sources, shelter and nesting sites (Cane, 2008; Pywell et al.,
2011; Carvalheiro et al., 2012). In addition, the recent report of parallel decline of
pollinators and insect-pollinated plants (Biesmeijer et al., 2006) encouraged further
investigations in plant-pollinator interactions (Campbell et al., 2012a; Carrie et al.,
2012; Wratten et al., 2012) and habitat management (Cranmer et al., 2012; Holzschuh et
al., 2012; Wratten et al., 2012) to enhance the presence of beneficial insectary plants
within agro-ecosystems and therefore attract beneficial insects.
In order to promote the conservation of beneficial insects in a specific region,
there is a need to ensure sustainable restoration of pollinator habitats within agro-
ecosystems by using native beneficial insectary plants that can offer better food sources
and shelter to pollinators (Menz et al., 2011). However, to manage beneficial insectary
plants within specific agro-ecosystems, it is essential to reflect on its climate conditions
(Memmott et al., 2010), as beneficial insectary plants of one region can behave as
weedy or invasive under different environmental conditions of another region. On the
other hand, it is also important to be familiar with the abundance and diversity of
different pollinator groups of the specific region, mainly because the habitat
management for conservation of pollinators frequently vary among different groups. For
example, in Central Spain where diversity and abundance of solitary bees (especially
mining bees) is relatively high in comparison with other groups, the habitat
management applied should differ from the UK landscape management where the most
abundant pollinators are bumblebees and hoverflies. Therefore, it is recommendable to
study the agricultural behaviour and the attractiveness to pollinators of introduced or
native beneficial plants under the field conditions of a particular region (Comba et al.,
1999; Corbet et al., 2001), before recommending their use.
In the Mediterranean region, use and availability of aromatic (Lamiaceae) plants
is very common. As many species of the aromatic plants are native, highly attractive to
pollinators and well adapted to the semi-arid conditions of the Mediterranean region

Chapter VII
97
(Petanidou and Vokou, 1993; Garcia, 2000), their use for conservation of pollinators in
agro-ecosystems should be considered. However, before implementing these plants in
the agro-ecosystems, it is required to be familiar with the most appropriate ways to
manage them within agricultural fields. For this reason, six aromatic plant species, with
different bloom periods, were pre-selected for this study, as their good adaptation to
semi-arid conditions and ability to survive in the field during hot summer months
(Petanidou and Vokou, 1993) showed good potential for creation of a functional long-
flowering floral mixture (from early spring to mid-fall).
The results revealed a strong positive correlation between insect visits and floral
density (Dauber et al., 2010; Scriven et al., 2013). Since, floral morphology did not
affect the attractiveness of different plant species to pollinators, high floral density of
Nepeta tuberosa and Hyssopus officinalis was probably responsible for the high
attractiveness of these plants to pollinators, mainly visited by leafcutter bees and
hoverflies. On the other hand, the lower floral density in mixed plots was probably the
explanation for reduced number of pollinator visits in these plots (Essenberg, 2012;
Hudewenz et al., 2012), mainly because within the mixed plots never occurred that all
plant species were flowering at the same time. In addition, some plant species included
in the mixture were less attractive and consequently not efficient in the floral mixture
(Melissa officinalis and Thymbra capitata). Therefore, these plant species should be
replaced with more efficient ones, in order to increase the floral density of the mixture,
and optimize the supply of food for pollinators.
In addition, the results showed that under field conditions, besides floral density,
other factors such are weather and plant competition, can negatively influence the
efficiency of plants in the mixture to attract pollinators. Therefore, this study provides
some interesting hints for the design of floral margins, such is to avoid the usage of low-
growing plant species in floral mixtures, since pollinators cannot reach the flowers. For
example, the reduced pollinator visitation of T. capitata in the floral mixture was
detected because pollinators were unable to approach the flowers. On the other hand, in
mono-specific plots this plant was highly attractive to small solitary bees and for that
reason its use in mono-specific stands is preferred over mixed. Following this example,
this study emphasizes the importance to find the most appropriate management for each
plant species, in order to achieve the greatest benefit from its implementation within
agro-ecosystems.

Chapter VII
98
With a similar aim, twelve spontaneous native or naturalized annual herbaceous
plants were studied under the field conditions of Central Spain. As all pre-selected
herbaceous plants are spring flowering species, their attractiveness to pollinators was
simultaneously compared together with their floral phenology (Adhikari and Adhikari,
2010), using the attractiveness efficiency, i.e. combination of the total number of insect
visits/minute and flowering duration, as a parameter for comparison. This parameter
goes one step further in the evaluation of beneficial insectary plants for conservation of
pollinators, showing that some plant species as Borago officinalis or Coriandrum
sativum, frequently cited as highly attractive and efficient beneficial plants (Ambrosino
et al., 2006; Campbell et al., 2012a), are less efficient in conservation of pollinators due
to their short flowering period. In contrast, Diplotaxis tenuifolia, which had a long
flowering period and showed to be attractive for bees and hoverflies in both years of
study, showed to be highly efficient in both, mono-specific stands (due to its high
attractiveness efficiency) and mixed stands (due to its high contribution to the
functionality of the mixture).
The study also pointed out that some inter-specific interactions between studied
pollinators and other insects can affect the attractiveness of plants through flower
occupation (Bosch, 1992) or damage (Barbir et al., 2014a). For example, Heliotaurus
ruficollis occupied the flowers of Phacelia tanacetifolia and Echium plantagineum,
while Psilothrix viridicoerulea, described as a poor pollinator (Bartomeus et al., 2008),
occupied the flowers of Calendula arvensis, reducing the approachability of the flowers
to bees and hoverflies. Additionally, Tropinota squalida, described as a pest in agro-
ecosystems (Khater et al., 2003), negatively influenced the attractiveness of the studied
flowers to beneficial insects by destroying the bottom part of corollas when reaching for
the nectar of E. plantagineum (J. Barbir, personal observation). As these inter-specific
insect interactions can influence the final estimation of the attractiveness of the plants to
bees and hoverflies, it would be advisable to be considered in further studies on the
conservation of pollinators.
Although the plants assessed in these two studies are native and frequent in the
study area, some of them showed a weedy behaviour as they intruded in the
neighbouring crop. Most of aromatic plant species maintained their original stands
during three years, with the exception of the reduction in the case of M. officinalis.
Considering that perennial plants are recommendable as floral sources to pollinators in

Chapter VII
99
agro-ecosystems due to their low maintenance (Tuell et al., 2008), the fact that the
initial area of M. officinalis was reduced may represent a limitation for its usage in floral
margins. On the other hand, some herbaceous plants, i.e. C. arvensis, P. tanacetifolia
and E. plantagineum, showed wide spatial expansion, intruding the neighbouring crop.
Although shallow tillage significantly reduced their self seeding, it did not stop weedy
behaviour of C. arvensis (Saramago et al., 2012). Hence, the usage of this plant as a
beneficial insectary plant in agro-ecosystems of Central Spain is not recommendable.
After analyzing the potential attractiveness of all pre-selected plants (perennial
aromatics and annual herbaceous) to pollinators and their agricultural management in
agro-ecosystems it was necessary to deepen the recommendations regarding the habitat
management practices. Therefore, the scale of further experiments was expanded, the
number of plant species included (using only the most efficient plants) was reduced, and
the experiments were focused on the crops of interests. In this way, besides conserving
pollinators in agro-ecosystems, it is also possible to improve the pollination of targeted
crops.
Therefore, as D. tenuifolia showed high attractiveness efficiency, good self-
seeding and no weediness under field conditions, it has been considered as a good
beneficial plant for conservation of local pollinators. However, before recommending it
to local farmers, it was important to estimate its capacity to cope with water deficit as
well as to recommend the most appropriate regulated deficit irrigation (RDI) regime due
to the semi-arid conditions of Central Spain. Since the study was conducted under
controlled green house conditions, the exact RDI regime under field conditions could
not be recommended, as it highly depends of precipitation and soil type (McDonald and
Girvetz, 2013). However, it was evaluated how different RDI regimes can influence the
growth and the attractiveness of D. tenuifolia to pollinators. The study showed how the
development of its flowers is more sensitive to drought stress than vegetative
development, as drought of 8 days did not affect any of the measured vegetative
parameters, since it reduced the number of open flowers and the total floral area.
Considering that pollinator visits are correlated with the number of open flowers and
their total floral area (Conner and Rush, 1996), the results suggest that, under the
temperature and relative humidity conditions of this experiment, D. tenuifolia could
tolerate drought for 4 days without affecting its attractiveness to pollinators. However,
those results should be considered with precaution, as the experiment was conducted

Chapter VII
100
under controlled conditions and it is recommendable to estimate the effect of drought
under field conditions.
In agreement with previous researches conducted with other plants (Mingeau et
al., 2001; Cakir, 2004; Cuevas et al., 2007; Alvarez et al., 2013), this study showed that
D. tenuifolia responds differently to RDI and water stress depending on the
phenological stage of the plant. Although moderate deficit irrigation (MDI) did not
affect plant growth or floral attractiveness to pollinators in any phenological stage,
severe deficit irrigation (SDI) applied before flowering significantly reduced some
vegetative parameters, but not floral; whereas when applied after flowering SDI
negatively affected all vegetative parameters and surface area of individual flowers. The
study revealed that the negative effect of drought was reflected mainly on floral
characteristics, while the negative effect of RDI was primarily expressed on the
vegetative parameters of the plant. Application of MDI before flowering was the most
efficient water-saving technique for commercial production of D. tenuifolia, as it did
not reduce plant growth and saved up to 40% of irrigation water. On the other hand, the
longest drought period tested (14 days) reduced up to 50% the consumption of irrigation
water, but as it resulted in death of most of the plants, even after irrigation was
reinstated, its application was inadvisable. Although reducing vegetative growth, SDI
before flowering could save water consumption by 70% without affecting flowering and
plant attractiveness of D. tenuifolia to pollinators. Therefore, this irrigation regime
could be used as sustainable and beneficial when managing the pollinator habitats in
agro-ecosystems.
Once estimating the efficiency in attracting pollinators of all studied plants
included in the study, choosing the most appropriate plant for efficient mono-specific
stands, and optimizing the attractiveness of herbaceous and aromatic mixture, the effect
of floral margins (mono-specific or mixed) on pollination and crop yield production was
estimated under field conditions. The mixture consisted of the most attractive
herbaceous plants chosen in the previous study. This spring blooming mixture was
considered appropriate for the experiment as coriander is also a spring blooming crop.
The experiment showed that the proximity of both types of floral margins improved
quality (i.e. seed weight and seed germination rate) and quantity of coriander seeds. On
the other hand, number of seeds produced was significantly higher in the crop growing
near mixed margins, than near mono-specific margin, probably due to stronger

Chapter VII
101
correlations in pollinator visits to coriander flowers and number of flowers in mixed
margins (Ghazoul, 2006). As the experiment was conducted under semi-controlled
conditions (potted plants of coriander), it can be concluded that pollinator visits was the
main factor that biased the results and that presence of both mixed or mono-specific (D.
tenuifolia) margins can improve the crop production for more than 200% and, in
addition, conserve the local pollinators in agro-ecosystems. Nevertheless, it is
recommendable to link the understanding of insect conservation practices with the
existing information on agronomic production of insect-pollinated crops, in order to
develop more sustainable agricultural practices. Therefore, further studies should be
expanded to a larger scale and carried out with different insect-pollinated crops.
Finally, after evaluating potential of various aromatic and herbaceous plants for
conservation of pollinators in agro-ecosystems of Central Spain, it has been found that
some plants have higher value as beneficial insectary plants than others, and that some
plants are more recommendable to be used for conservation of pollinators than others.
However, it is important to apply the appropriate management when introducing the
selected beneficial insectary plants in agro-ecosystems in order to provide benefits to
both local pollinators and local farmers (by improving the agricultural production in the
field through increased rate of pollination).

Chapter VIII
102
VIII. Conclusions
Conclusions on Objective 1:
Although floral morphology, e.g. flower length, flower height, corolla tube
length, corolla height and calyx length, did not affect the attractiveness of six
aromatic (Lamiaceae) plant species to pollinators, the floral density did, as plant
species with higher floral density, i.e. Nepeta tuberosa and Hyssopus officinalis,
were characterized as the most attractive plant species.
Of six aromatic (Lamiaceae) plant species, two summer species (Melissa
officinalis and Thymbra capitata) did not efficiently contribute to the
attractiveness of the floral mixture to pollinators, reducing its attractiveness
during the summer period.
Conclusions on Objective 2:
Among twelve herbaceous annual plant species, the flowers of Borago
officinalis, Echium plantagineum, Phacelia tanacetifolia and Diplotaxis
tenuifolia were the most attractive to bees; while the flowers of Calendula
arvensis, Coriandrum sativum, D. tenuifolia and Lobularia maritima were
attractive to hoverflies; therefore, D. tenuifolia was the only species with high
attractiveness efficiency to both bees and hoverflies.
The floral mixture resulted in lower attractiveness efficiency to pollinators than
mono-specific stand of D. tenuifolia, but higher than most of the other mono-
specific stands.
Conclusion on Objective 3:
Of six aromatic plant species studied, none showed weedy behaviour under field
conditions. In contrast, some of the most attractive herbaceous plant species,
such as P. tanacetifolia and C. arvensis, showed potential weediness under field
conditions, but their self-seeding was significantly reduced by tillage which
could be recommended as a suitable weed management.

Chapter VIII
103
Conclusions on Objective 4:
In controlled greenhouse conditions, plants of D. tenuifolia could be without
irrigation for 4 days without affecting its growth, flowering and attractiveness to
pollinators. However, lack of irrigation for 8 days or longer significantly
reduced their vegetative growth and floral morphology, i.e. number of open
flowers, total floral area, flower diameter, corolla tube diameter and corolla tube
length.
Regulated deficit irrigation can improve water use efficiency, and depending on
the purpose of growing D. tenuifolia, as a crop or as a beneficial plant to attract
pollinators, it can reduce water consumption by 40% to 70% without affecting
its vegetative and floral development and without reducing its attractiveness to
pollinators.
Conclusions on Objective 5:
The proximity of floral margins improved the seed quality (e.g., seed weight and
germination rate) of coriander crop plants with regard to the controls without
floral margin. In addition, coriander crop production, i.e. number of seeds, was
significantly higher in plants grown near floral mix margins than near mono-
specific margins.
Since the experiments were conducted under semi-controlled field conditions, it
can be concluded that pollinator visits was the main factor that biased the
results, and that presence of both mixed or mono-specific (D. tenuifolia) margins
can improve the coriander production for more than 200% in small-scale
gardens and, in addition, conserve the local pollinators within agricultural fields.

Chapter IX
104
IX. Conclusiones
Conclusiones del Objetivo 1:
Aparentemente, las características morfológicas florales, e.g., diámetro y altura
de la flor, diámetro y longitud del tubo de la corola, longitud del cáliz, no
afectan significativamente a la atracción de las seis especies aromáticas
(Lamiaceae) sobre los polinizadores. Por el contrario, si ha tenido un efecto
significativo la densidad floral, resultando las especies con mayor densidad de
flores (Nepeta tuberosa e Hyssopus officinalis) las especies más atractivas a
polinizadores dentro de este estudio.
De las seis especies perennes aromáticas consideradas, las dos especies con
fenología de verano (Melissa officinalis y Thymbra capitata) no han contribuido
eficientemente al atractivo de la mezcla floral sobre los polinizadores, debido a
la reducción de la atracción total durante el periodo estival.
Conclusiones del Objetivo 2:
De entre las doce especies herbáceas anuales consideradas en este estudio, las
flores de Borago officinalis, Echium plantagineum, Phacelia tanacetifolia y
Diplotaxis tenuifolia han resultado ser las más atractivas para las abejas,
mientras que las flores de Calendula arvensis, Coriandrum sativum, D.
tenuifolia y Lobularia maritima han demostrado ser las más atractivas para los
sírfidos. Por lo tanto, D. tenuifolia es la única de las especies consideradas con
elevada atracción a ambos grupos de polinizadores, abejas y sírfidos.
La mezcla floral ha tenido menor eficiencia de atracción a polinizadores que las
parcelas mono-específicas de D. tenuifolia, aunque mayor que el resto de
parcelas con cultivos mono-específicos.
Conclusiones del Objetivo 3:
Ninguna de las seis especies aromáticas perennes consideradas en este estudio
ha mostrado tendencia invasora. Por el contrario, algunas de las especies
herbáceas anuales con mayor atractivo a polinizadores (P. tanacetifolia and C.

Chapter IX
105
arvensis), se han comportado como invasoras en condiciones de campo. Sin
embargo, este comportamiento como malas hierbas se redujo significativamente
con el laboreo, una labor agronómica habitual que puede recomendarse para
prevenir la expansión de estas especies.
Conclusiones del Objetivo 4:
En condiciones controladas de invernadero, se ha comprobado que D. tenuifolia
puede mantenerse sin riego durante 4 días sin que ello afecte a su potencial
atractivo a los polinizadores. Por otro lado, la falta de riego durante 8 días o
más, reduce significativamente su crecimiento vegetativo y sus características
morfológicas florales, e.g. número de flores, diámetro y área de la flor, diámetro
y longitud del tubo de la corola.
Un Déficit de Riego Regulado puede mejorar la eficiencia del uso del agua, el
cual, dependiendo del uso final de la especie D. tenuifolia, como cultivo para el
consumo o como planta atractiva a polinizadores, puede reducir el consumo de
agua entre un 40 y un 70 % sin que afecte su desarrollo vegetativo y floral, y sin
reducir su atracción a polinizadores.
Conclusiones del Objetivo 5:
La proximidad de un margen floral junto al cultivo de cilantro mejora la calidad
de sus semillas (peso y tasa de germinación) respecto a las plantas sin margen
floral. Por otro lado, los márgenes compuestos por una mezcla floral
incrementan el rendimiento del cultivo (numero de semillas) respecto a las
parcelas junto a márgenes mono-específicos.
Puesto que los experimentos han sido desarrollados en condiciones de campo
semi-controladas, se puede concluir que las visitas de polinizadores han sido el
factor determinante en los resultados, y que la presencia de un margen floral (ya
sea mono-especifico o de mezcla) puede aumentar la producción a pequeña
escala del cultivo de cilantro en más de un 200%, ayudando además a la
conservación de los polinizadores en los agro-ecosistemas.

Chapter X
106
X. References
Abrahamczyk, S., Kluge, J., Gareca, Y., Reichle, S. and Kessler, M. (2011) The influence of climatic seasonality on the diversity of different tropical pollinator groups. Plos One, 6, 9.
Adhikari, S. and Adhikari, M. K. (2010) Floral phenology and pollination ecology of Punica granatum L. in Kathmandu, Nepal. Nepal Journal of Science and Technology, 11, 115-124.
Al-Fattah, M. A. (2001) The ecological behaviour of honeybee foragers on flowers of certain aromatic crops and their role in seed production.The 37th International Apicultural Congress, Durban, South Africa
Albrecht, M., Duelli, P., Mueller, C., Kleijn, D. and Schmid, B. (2007) The Swiss agri-environment scheme enhances pollinator diversity and plant reproductive success in nearby intensively managed farmland. Journal of Applied Ecology, 44, 813-822.
Altieri, M. A. (1991) How best we can use biodiversity in agroecosystems. Outlook on Agriculture, 20, 15-23.
Altieri, M. A. (1999) The ecological role of biodiversity in agroecosystems. Agriculture Ecosystems & Environment, 74, 19-31.
Alvarez, S., Banon, S. and Sanchez-Blanco, M. J. (2013) Regulated deficit irrigation in different phenological stages of potted geranium plants: water consumption, water relations and ornamental quality. Acta Physiologiae Plantarum, 35, 1257-1267.
Ambrosino, M. D., Luna, J. M., Jepson, P. C. and Wratten, S. D. (2006) Relative frequencies of visits to selected insectary plants by predatory hoverflies (Diptera: Syrphidae), other beneficial insects, and herbivores. Environmental Entomology, 35, 394-400.
Amoros-Jimenez, R., Pineda, A., Fereres, A. and Marcos-Garcia, M. A. (2012) Prey availability and abiotic requirements of immature stages of the aphid predator Sphaerophoria rueppellii. Biological Control, 63, 17-24.
Andersson, G. K. S., Rundlof, M. and Smith, H. G. (2012) Organic farming improves pollination success in strawberries. PLoS One, 7.
Badenes-Perez, F. R., Nault, B. A. and Shelton, A. M. (2005) Manipulating the attractiveness and suitability of hosts for diamondback moth (Lepidoptera : Plutellidae). Journal of Economic Entomology, 98, 836-844.
Baker, H. G. and Baker, I. (1983) In Handbook of experimental pollen biology.(Ed, Jones, C. E. L. R. J.), pp. 117-141.
Barbir, J., Badenes-Perez, F. R., Fernandez-Quintanilla, C. and Dorado, J. (2014a) The attractiveness of flowering herbaceous plants to bees (Hymenoptera: Apoidea) and hoverflies (Diptera: Syrphidae) in agro-ecosystems of Central Spain. Agricultural and Forest Entomology.
Barbir, J., Dorado, J., Fernández-Quintanilla, C., Blanusa, T., Maksimovic, C. and

Chapter X
107
Badenes-Pérez, F. R. (2014b) Wild rocket – effect of water deficit on growth, flowering and attractiveness to pollinators. Acta Agriculturae Scandinavica, Section B – Soil & Plant Science.
Bartomeus, I., Bosch, J. and Vila, M. (2008) High invasive pollen transfer, yet low deposition on native stigmas in a Carpobrotus-invaded community. Annals of Botany, 102, 417-424.
Bartomeus, I., Potts, S. G., Steffan-Dewenter, I., Vaissiere, B. E., Woyciechowski, M., Krewenka, K. M., Tscheulin, T., Roberts, S. P. M., Szentgyorgyi, H., Westphal, C. and Bommarco, R. (2014) Contribution of insect pollinators to crop yield and quality varies with agricultural intensification. PeerJ, 2, e328.
Bastian, O. (2013) The role of biodiversity in supporting ecosystem services in Natura 2000 sites. Ecological Indicators, 24, 12-22.
Batary, P., Andras, B., Kleijn, D. and Tscharntke, T. (2011) Landscape-moderated biodiversity effects of agri-environmental management: a meta-analysis. Proceedings of the Royal Society B-Biological Sciences, 278, 1894-1902.
Batary, P., Baldi, A., Saropataki, M., Kohler, F., Verhulst, J., Knop, E., Herzog, F. and Kleijn, D. (2010) Effect of conservation management on bees and insect-pollinated grassland plant communities in three European countries. Agriculture Ecosystems & Environment, 136, 35-39.
Begum, M., Gurr, G. M., Wratten, S. D., Hedberg, P. R. and Nicol, H. I. (2006) Using selective food plants to maximize biological control of vineyard pests. Journal of Applied Ecology, 43, 547-554.
Bendifallah, L., Louadi, K. and Doumandji, S. (2013) Bee fauna potential visitors of coriander flowers Coriandrum sativum L. (Apiaceae) in the Mitidja area (Algeria). Journal of Apicultural Science, 57, 59-70.
Bennett, R. N., Carvalho, R., Mellon, F. A., Eagles, J. and Rosa, E. A. S. (2007) Identification and quantification of glucosinolates in sprouts derived from seeds of wild Eruca sativa L. (salad rocket) and Diplotaxis tenuifolia L. (wild rocket) from diverse geographical locations. Journal of Agricultural and Food Chemistry, 55, 67-74.
Benton, T. G., Vickery, J. A. and Wilson, J. D. (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends in Ecology & Evolution, 18, 182-188.
Bernhardt, P. (2000) Convergent evolution and adaptive radiation of beetle-pollinated angiosperms. Plant Systematics and Evolution, 222, 293-320.
Biesmeijer, J. C., Roberts, S. P. M., Reemer, M., Ohlemuller, R., Edwards, M., Peeters, T., Schaffers, A. P., Potts, S. G., Kleukers, R., Thomas, C. D., Settele, J. and Kunin, W. E. (2006) Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science, 313, 351-354.
Billeter, R., Liira, J., Bailey, D., Bugter, R., Arens, P., Augenstein, I., Aviron, S., Baudry, J., Bukacek, R., Burel, F., Cerny, M., De Blust, G., De Cock, R., Diekotter, T., Dietz, H., Dirksen, J., Dormann, C., Durka, W., Frenzel, M., Hamersky, R., Hendrickx, F., Herzog, F., Klotz, S., Koolstra, B., Lausch, A., Le Coeur, D., Maelfait, J. P., Opdam, P., Roubalova, M., Schermann, A., Schermann, N., Schmidt, T., Schweiger, O., Smulders, M. J. M., Speelmans, M.,

Chapter X
108
Simova, P., Verboom, J., van Wingerden, W., Zobel, M. and Edwards, P. J. (2008) Indicators for biodiversity in agricultural landscapes: a pan-European study. Journal of Applied Ecology, 45, 141-150.
Bosch, J. (1992) Floral biology and pollinators of three co-occurring Cistus species (Cistaceae). Botanical Journal of the Linnean Society, 109, 39-55.
Bosch, J., Gonzalez, A. M. M., Rodrigo, A. and Navarro, D. (2009) Plant-pollinator networks: adding the pollinator's perspective. Ecology Letters, 12, 409-419.
Branquart, E. and Hemptinne, J. L. (2000) Selectivity in the exploitation of floral resources by hoverflies (Diptera : Syrphinae). Ecography, 23, 732-742.
Brittain, C. and Potts, S. G. (2011) The potential impacts of insecticides on the life-history traits of bees and the consequences for pollination. Basic and Applied Ecology, 12, 321-331.
Buide, M. L. (2006) Pollination ecology of Silene acutifolia (Caryophyllaceae): Floral traits variation and pollinator attraction. Annals of Botany, 97, 289-297.
Cakir, R. (2004) Effect of water stress at different development stages on vegetative and reproductive growth of corn. Field Crops Research, 89, 1-16.
Campbell, A. J., Biesmeijer, J. C., Varma, V. and Wackers, F. L. (2012a) Realising multiple ecosystem services based on the response of three beneficial insect groups to floral traits and trait diversity. Basic and Applied Ecology, 13, 363-370.
Campbell, D. R., Bischoff, M., Lord, J. M. and Robertson, A. W. (2012b) Where have all the blue flowers gone: pollinator responses and selection on flower colour in New Zealand Wahlenbergia albomarginata. Journal of Evolutionary Biology, 25, 352-364.
Cane, J. H. (1997) Ground-nesting bees: the neglected pollinator resource for agriculture. Acta Horticulturae (Wageningen), 437, 309-324.
Cane, J. H. (2008) A native ground-nesting bee (Nomia melanderi) sustainably managed to pollinate alfalfa across an intensively agricultural landscape. Apidologie, 39, 315-323.
Caraballo-Ortiz, M. A., Santiago-Valentin, E. and Carlo, T. A. (2011) Flower number and distance to neighbours affect the fecundity of Goetzea elegans (Solanaceae). Journal of Tropical Ecology, 27, 521-528.
Carreck, N. L. and Williams, I. H. (2002) Food for insect pollinators on farmland: insect visits to flowers of annual seed mixtures. Journal of Insect Conservation, 6, 13-23.
Carrie, R. J. G., George, D. R. and Wackers, F. L. (2012) Selection of floral resources to optimise conservation of agriculturally-functional insect groups. Journal of Insect Conservation, 16, 635-640.
Carroll, A. B., Pallardy, S. G. and Galen, C. (2001) Drought stress, plant water status, and floral trait expression in fireweed, Epilobium angustifolium (Onagraceae). American Journal of Botany, 88, 438-446.
Carrubba, A., la Torre, R., Saiano, F. and Alonzo, G. (2006) Effect of sowing time on coriander performance in a semiarid Mediterranean environment. Crop Science,

Chapter X
109
46, 437-447.
Carrubba, A. and Scalenghe, R. (2012) The scent of Mare Nostrum: medicinal and aromatic plants in Mediterranean soils. Journal of the Science of Food and Agriculture, 92, 1150-1170.
Carvalheiro, L. G., Kunin, W. E., Keil, P., Aguirre-Gutierrez, J., Ellis, W. N., Fox, R., Groom, Q., Hennekens, S., Van Landuyt, W., Maes, D., Van de Meutter, F., Michez, D., Rasmont, P., Ode, B., Potts, S. G., Reemer, M., Roberts, S. P. M., Schaminee, J., WallisDeVries, M. F. and Biesmeijer, J. C. (2013) Species richness declines and biotic homogenisation have slowed down for NW-European pollinators and plants. Ecology Letters, 16, 870-878.
Carvalheiro, L. G., Seymour, C. L., Nicolson, S. W. and Veldtman, R. (2012) Creating patches of native flowers facilitates crop pollination in large agricultural fields: mango as a case study. Journal of Applied Ecology, 49, 1373-1383.
Carvell, C., Meek, W. R., Pywell, R. F., Goulson, D. and Nowakowski, M. (2007) Comparing the efficacy of agri-environment schemes to enhance bumble bee abundance and diversity on arable field margins. Journal of Applied Ecology, 44, 29-40.
Castroviejo, S., Aedo, C., Cirujano, S., Laínz, M., Montserrat, P., Morales, R., Muñoz Garmendia, F., Navarro, C., Paiva, J., Soriano, C. (1993) Flora iberica 12, Real Jardín Botánico, CSIC, Madrid.
Cerekovic, N., Pagter, M., Kristensen, H. L., Pedersen, H. L., Brennan, R. and Petersen, K. K. (2013) Effects of drought stress during flowering of two pot-grown blackcurrant (Ribes nigrum L.) cultivars. Scientia Horticulturae, 162, 365-373.
Chambers, R. J. and Adams, T. H. L. (1986) Quantification of the impact of hoverflies (Diptera, Syrphidae) on cereal aphids in winter-wheat - an analysis of field populations. Journal of Applied Ecology, 23, 895-904.
Chartzoulakis, K., Patakas, A., Kofidis, G., Bosabalidis, A. and Nastou, A. (2002) Water stress affects leaf anatomy, gas exchange, water relations and growth of two avocado cultivars. Scientia Horticulturae, 95, 39-50.
Chaudhary, O. P. and Singh, J. (2007) Diversity, temporal abundance, foraging behaviour of floral visitors and effect of different modes of pollination on coriander (Coriandrum sativum L.). Journal of Spices and Aromatic Crops, 16, 8–14.
Chaves, M. M., Maroco, J. P. and Pereira, J. S. (2003) Understanding plant responses to drought - from genes to the whole plant. Functional Plant Biology, 30, 239-264.
Christian-Smith, J., Cooley, H. and Gleick, P. H. (2012) Potential water savings associated with agricultural water efficiency improvements: a case study of California, USA. Water Policy, 14, 194-213.
Colley, M. R. and Luna, J. M. (2000) Relative attractiveness of potential beneficial insectary plants to aphidophagous hoverflies (Diptera : Syrphidae). Environmental Entomology, 29, 1054-1059.
Comba, L., Corbet, S. A., Hunt, L. and Warren, B. (1999) Flowers, nectar and insect visits: Evaluating British plant species for pollinator-friendly gardens. Annals of Botany, 83, 369-383.

Chapter X
110
Conner, J. K. and Rush, S. (1996) Effects of flower size and number on pollinator visitation to wild radish, Raphanus raphanistrum. Oecologia, 105, 509-516.
Corbet, S. A., Bee, J., Dasmahapatra, K., Gale, S., Gorringe, E., La Ferla, B., Moorhouse, T., Trevail, A., Van Bergen, Y. and Vorontsova, M. (2001) Native or exotic? Double or single? Evaluating plants for pollinator-friendly gardens. Annals of Botany, 87, 219-232.
Cranmer, L., McCollin, D. and Ollerton, J. (2012) Landscape structure influences pollinator movements and directly affects plant reproductive success. Oikos, 121, 562-568.
Crowder, D. W., Northfield, T. D., Gomulkiewicz, R. and Snyder, W. E. (2012) Conserving and promoting evenness: organic farming and fire-based wildland management as case studies. Ecology, 93, 2001-2007.
Cuevas, J., Canete, M. L., Pinillos, V., Zapata, A. J., Fernandez, M. D., Gonzalez, M. and Hueso, J. J. (2007) Optimal dates for regulated deficit irrigation in 'Algerie' loquat (Eriobotrya japonica Lindl.) cultivated in Southeast Spain. Agricultural Water Management, 89, 131-136.
D'Antuono, L. F., Elementi, S. and Neri, R. (2009) Exploring new potential health-promoting vegetables: glucosinolates and sensory attributes of rocket salads and related Diplotaxis and Eruca species. Journal of the Science of Food and Agriculture, 89, 713-722.
Dauber, J., Biesmeijer, J. C., Gabriel, D., Kunin, W. E., Lamborn, E., Meyer, B., Nielsen, A., Potts, S. G., Roberts, S. P. M., Sober, V., Settele, J., Steffan-Dewenter, I., Stout, J. C., Teder, T., Tscheulin, T., Vivarelli, D. and Petanidou, T. (2010) Effects of patch size and density on flower visitation and seed set of wild plants: a pan-European approach. Journal of Ecology, 98, 188-196.
Dobbs, T. L. and Pretty, J. N. (2004) Agri-environmental stewardship schemes and "multifunctionality". Review of Agricultural Economics, 26, 220-237.
Dobson, A., Lodge, D., Alder, J., Cumming, G. S., Keymer, J., McGlade, J., Mooney, H., Rusak, J. A., Sala, O., Wolters, V., Wall, D., Winfree, R. and Xenopoulos, M. A. (2006) Habitat loss, trophic collapse, and the decline of ecosystem services. Ecology, 87, 1915-1924.
Eckhart, V. M. (1992) Spatiotemporal variation in abundance and variation in foraging behavior of the pollinators of gynodioecious Phacelia linearis (Hydrophyllaceae). Oikos, 64, 573-586.
Elts, J. and Lohmus, A. (2012) What do we lack in agri-environment schemes? The case of farmland birds in Estonia. Agriculture Ecosystems & Environment, 156, 89-93.
Engel, M. S. (1999) The taxonomy of recent and fossil honey bees (Hymenoptera: Apidae; Apis). Journal of Hymenoptera Research, 8, 165-196.
Engel, M. S. (2000) Classification of the bee tribe Augochlorini (Hymenoptera: Halictidae). Bulletin of the American Museum of Natural History, 250, 1-89.
Engel, M. S. and Dingemansbakels, F. (1980) Nectar and pollen resources for stingless bees (Meliponinae, Hymenoptera) in Surinam (South-America). Apidologie, 11, 341-350.

Chapter X
111
English, M. J., Musich, J. T., Murty, V. V. N. (1990) In Management of Farm Irrigation Systems(Ed, Hoffman, G. J., Howell, T. A., Solomon, K. H.) American Society of Agricultural Engineers, St. Joseph, MI, USA, pp. 631-663.
Essenberg, C. J. (2012) Explaining variation in the effect of floral density on pollinator visitation. American Naturalist, 180, 153-166.
Fereres, E. and Soriano, M. A. (2007) Deficit irrigation for reducing agricultural water use. Journal of Experimental Botany, 58, 147-159.
Fontaine, C., Dajoz, I., Meriguet, J. and Loreau, M. (2006) Functional diversity of plant-pollinator interaction webs enhances the persistence of plant communities. Plos Biology, 4, 129-135.
Frund, J., Linsenmair, K. E. and Bluthgen, N. (2010) Pollinator diversity and specialization in relation to flower diversity. Oikos, 119, 1581-1590.
Garcia-Tejero, I., Duran-Zuazo, V. H., Arriaga-Sevilla, J. and Muriel-Fernandez, J. L. (2012) Impact of water stress on citrus yield. Agronomy for Sustainable Development, 32, 651-659.
Garcia, J. D. D. (2000) Patterns of insect flower visitation in Lavandula bucchii Webb (Lamiaceae), an endemic shrub of Tenerife (Canary Islands). Journal of Natural History, 34, 2145-2155.
Garibaldi, L. A., Aizen, M. A., Cunningham, S. and Klein, A. M. (2009) Pollinator shortage and global crop yield: looking at the whole spectrum of pollinator dependency. Communicative & Integrative Biology, 2, 37-39.
Ghazoul, J. (2006) Floral diversity and the facilitation of pollination. Journal of Ecology, 94, 295-304.
Gill, R. J., Ramos-Rodriguez, O. and Raine, N. E. (2012) Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature, 491, 105-8.
Gomez, J. M., Bosch, J., Perfectti, F., Fernandez, J. D., Abdelaziz, M. and Camacho, J. P. M. (2008) Spatial variation in selection on corolla shape in a generalist plant is promoted by the preference patterns of its local pollinators. Proceedings of the Royal Society B-Biological Sciences, 275, 2241-2249.
Goulson, D. (2003) Effects of introduced bees on native ecosystems. Annual Review of Ecology Evolution and Systematics, 34, 1-26.
Goulson, D., Lepais, O., O'Connor, S., Osborne, J. L., Sanderson, R. A., Cussans, J., Goffe, L. and Darvill, B. (2010) Effects of land use at a landscape scale on bumblebee nest density and survival. Journal of Applied Ecology, 47, 1207-1215.
Grigulis, K., Sheppard, A. W., Ash, J. E. and Groves, R. H. (2001) The comparative demography of the pasture weed Echium plantagineum between its native and invaded ranges. Journal of Applied Ecology, 38, 281-290.
Gurr, G. M., Wratten, S. D. and Luna, J. M. (2003) Multi-function agricultural biodiversity: pest management and other benefits. Basic and Applied Ecology, 4, 107-116.
Gutbrodt, B., Mody, K. and Dorn, S. (2011) Drought changes plant chemistry and causes contrasting responses in lepidopteran herbivores. Oikos, 120, 1732-1740.

Chapter X
112
Haaland, C., Naisbit, R. E. and Bersier, L.-F. (2011) Sown wildflower strips for insect conservation: a review. Insect Conservation and Diversity, 4, 60-80.
Hall, M. K. D., Jobling, J. J. and Rogers, G. S. (2012) Some perspectives on rocket as a vegetable crop: a review. Vegetable Crops Research Bulletin, 76, 21-41.
Heard, M. S., Carvell, C., Carreck, N. L., Rothery, P., Osborne, J. L. and Bourke, A. F. G. (2007) Landscape context not patch size determines bumble-bee density on flower mixtures sown for agri-environment schemes. Biology Letters, 3, 638-641.
Herrera, C. M. (2001) Deconstructing a floral phenotype: do pollinators select for corolla integration in Lavandula latifolia? Journal of Evolutionary Biology, 14, 574-584.
Hoffman, G. D. and Hogg, D. B. (1992) Effect of alfalfa water-stress on potato leafhopper (Homoptera, Cicadellidae) plant preference and oviposition rate. Annals of the Entomological Society of America, 85, 506-516.
Hogg, B. N., Bugg, R. L. and Daane, K. M. (2011) Attractiveness of common insectary and harvestable floral resources to beneficial insects. Biological Control, 56, 76-84.
Holzschuh, A., Dudenhoffer, J. H. and Tscharntke, T. (2012) Landscapes with wild bee habitats enhance pollination, fruit set and yield of sweet cherry. Biological Conservation, 153, 101-107.
Howell, T. A. (2001) Enhancing water use efficiency in irrigated agriculture. Agronomy Journal, 93, 281-289.
Hsiao, T. C. (1973) Plant responses to water stress. Annual Review of Plant Physiology and Plant Molecular Biology, 24, 519-570.
Huberty, A. F. and Denno, R. F. (2004) Plant water stress and its consequences for herbivorous insects: A new synthesis. Ecology, 85, 1383-1398.
Hudewenz, A., Klein, A. M., Scherber, C., Stanke, L., Tscharntke, T., Vogel, A., Weigelt, A., Weisser, W. W. and Ebeling, A. (2012) Herbivore and pollinator responses to grassland management intensity along experimental changes in plant species richness. Biological Conservation, 150, 42-52.
Hussain, A. I., Anwar, F., Nigam, P. S., Sarker, S. D., Moore, J. E., Rao, J. R. and Mazumdar, A. (2011) Antibacterial activity of some Lamiaceae essential oils using resazurin as an indicator of cell growth. Lwt-Food Science and Technology, 44, 1199-1206.
ILC (2000) Safety and health in agriculture.International Labour Conference, Geneva
Inouye, D. W. (1980) The terminology of floral larceny. Ecology, 61, 1251-1253.
Jacobs, J. H., Clark, S. J., Denholm, I., Goulson, D., Stoate, C. and Osborne, J. L. (2009) Pollination biology of fruit-bearing hedgerow plants and the role of flower-visiting insects in fruit-set. Annals of Botany, 104, 1397-1404.
Jauker, F., Bondarenko, B., Becker, H. C. and Steffan-Dewenter, I. (2012) Pollination efficiency of wild bees and hoverflies provided to oilseed rape. Agricultural and Forest Entomology, 14, 81-87.
Jauker, F., Diekotter, T., Schwarzbach, F. and Wolters, V. (2009) Pollinator dispersal in

Chapter X
113
an agricultural matrix: opposing responses of wild bees and hoverflies to landscape structure and distance from main habitat. Landscape Ecology, 24, 547-555.
Jauker, F. and Wolters, V. (2008) Hover flies are efficient pollinators of oilseed rape. Oecologia, 156, 819-823.
Johnson, B. R. (2010) Division of labor in honey bees: form, function and proximate mechanisms. Behavioral Ecology and Sociobiology, 64, 305-316.
Julier, H. E. and Roulston, T. H. (2009) Wild bee abundance and pollination service in cultivated pumpkins: Farm management, nesting behavior and landscape effects. Journal of Economic Entomology, 102, 563-573.
Keasar, T., Pollak, G., Arnon, R., Cohen, D. and Shmida, A. (2006) Honesty of signaling and pollinator attraction: The case of flag-like bracts. Israel Journal of Plant Sciences, 54, 119-128.
Kells, A. R., Holland, J. M. and Goulson, D. (2001) The value of uncropped field margins for foraging bumblebees. Journal of Insect Conservation, 5, 283-291.
Khater, A. M., El-Zakardy, K. A. H. and Ebadah, I. M. A. (2003) The efficiency of honeybees and other insect pollinators in pollination of faba bean (Vicia faba L.). Bulletin of Faculty of Agriculture Cairo University, 54, 465-481.
Kleijn, D. and van Langevelde, F. (2006) Interacting effects of landscape context and habitat quality on flower visiting insects in agricultural landscapes. Basic and Applied Ecology, 7, 201-214.
Klein, A.-M., Vaissiere, B. E., Cane, J. H., Steffan-Dewenter, I., Cunningham, S. A., Kremen, C. and Tscharntke, T. (2007) Importance of pollinators in changing landscapes for world crops. Proceedings of the Royal Society Biological Sciences Series B, 274, 303-313.
Klein, A. M., Steffan-Dewenter, I. and Tscharntke, T. (2003a) Fruit set of highland coffee increases with the diversity of pollinating bees. Proceedings of the Royal Society B-Biological Sciences, 270, 955-961.
Klein, A. M., Steffan-Dewenter, I. and Tscharntke, T. (2003b) Pollination of Coffea canephora in relation to local and regional agroforestry management. Journal of Applied Ecology, 40, 837-845.
Koul, A. K., Hamal, I. A. and Gupta, S. K. (1989) Pollination mechanism in Coriandrum sativum L. (Apiaceae). Proceedings of the Indian Academy of Sciences-Plant Sciences, 99, 509-515.
Krebs, J. R., Wilson, J. D., Bradbury, R. B. and Siriwardena, G. M. (1999) The second silent spring? Nature, 400, 611-612.
Lamy, E., Schroder, J., Paulus, S., Brenk, P., Stahl, T. and Mersch-Sundermann, V. (2008) Antigenotoxic properties of Eruca sativa (rocket plant), erucin and erysolin in human hepatoma (HepG2) cells towards benzo(a)pyrene and their mode of action. Food and Chemical Toxicology, 46, 2415-2421.
Landis, D. A., Menalled, F. D., Costamagna, A. C. and Wilkinson, T. K. (2005) Manipulating plant resources to enhance beneficial arthropods in agricultural landscapes. Weed Science, 53, 902-908.

Chapter X
114
Landis, D. A., Wratten, S. D. and Gurr, G. M. (2000) Habitat management to conserve natural enemies of arthropod pests in agriculture. Annual Review of Entomology, 45, 175-201.
LaSalle, J. and Gauld, I. D. (1993) Hymenoptera: their diversity, and their impact on the diversity of other organisms, CAB International.
Le Feon, V., Schermann-Legionnet, A., Delettre, Y., Aviron, S., Billeter, R., Bugter, R., Hendrickx, F. and Burel, F. (2010) Intensification of agriculture, landscape composition and wild bee communities: A large scale study in four European countries. Agriculture Ecosystems & Environment, 137, 143-150.
Luo, H. J., Zheng, Z. B., Luo, S., Pan, Y. S. and Liu, X. H. (2007) In Proceedings of the Second International Symposium on Loquat(Eds, Huang, X. M. and Janick, J.) International Society Horticultural Science, Leuven 1, pp. 417-422.
Macukanovic-Jocic, M., Stevanovic, Z. D., Mladenovic, M. and Jocic, G. (2011) Flower morphophysiology of selected Lamiaceae species in relation to pollinator attraction. Journal of Apicultural Research, 50, 89-101.
Maloof, J. E. (2001) The effects of a bumble bee nectar robber on plant reproductive success and pollinator behavior. American Journal of Botany, 88, 1960-1965.
Markussen, M. V., Kulak, M., Smith, L. G., Nemecek, T. and Ostergard, H. (2014) Evaluating the sustainability of a small-scale low-input organic vegetable supply system in the United Kingdom. Sustainability, 6, 1913-1945.
Maroufi, K., Farahani, H. A. and Darvishi, H. H. (2010) Importance of coriander (Coriandrum sativum L.) between the medicinal and aromatic plants. Advances in Environmental Biology, 4, 433-436.
Marshall, E. J. P. (2005) Field margins in northern Europe: Integrating agricultural, environmental and biodiversity functions, Canadian Weed Science Soc, Soc Canadienne Malherbologie, Quebec.
Marshall, E. J. P. and Moonen, A. C. (2002) Field margins in northern Europe: their functions and interactions with agriculture. Agriculture Ecosystems & Environment, 89, 5-21.
Martin Gonzalez, A. M., Dalsgaard, B., Ollerton, J., Timmermann, A., Olesen, J. M., Andersen, L. and Tossas, A. G. (2009) Effects of climate on pollination networks in the West Indies. Journal of Tropical Ecology, 25, 493-506.
McDonald, R. I. and Girvetz, E. H. (2013) Two challenges for u. S. Irrigation due to climate change: Increasing irrigated area in wet states and increasing irrigation rates in dry states. Plos One, 8, 10.
Memmott, J., Carvell, C., Pywell, R. F. and Craze, P. G. (2010) The potential impact of global warming on the efficacy of field margins sown for the conservation of bumble-bees. Philosophical Transactions of the Royal Society B, 365, 2071-2079.
Menz, M. H. M., Phillips, R. D., Winfree, R., Kremen, C., Aizen, M. A., Johnson, S. D. and Dixon, K. W. (2011) Reconnecting plants and pollinators: challenges in the restoration of pollination mutualisms. Trends in Plant Science, 16, 4-12.
Michener, C. D. (2007) The bees of the world, Johns Hopkins University Press.

Chapter X
115
Miller, R., Owens, S. J. and Rorslett, B. (2011) Plants and colour: Flowers and pollination. Optics and Laser Technology, 43, 282-294.
Mingeau, M., Perrier, C. and Ameglio, T. (2001) Evidence of drought-sensitive periods from flowering to maturity on highbush blueberry. Scientia Horticulturae, 89, 23-40.
Miñarro, M. and Prida, E. (2013) Hedgerows surrounding organic apple orchards in north-west Spain: potential to conserve beneficial insects. Agricultural and Forest Entomology, 15, 382-390.
Moore, S. R. and Lawrence, K. S. (2013) The effect of soil texture and irrigation on Rotylenchulus reniformis and cotton. Journal of Nematology, 45, 99-105.
Morandin, L. A. and Kremen, C. (2013) Bee preference for native versus exotic plants in restored agricultural hedgerows. Restoration Ecology, 21, 26-32.
Morris, C. and Potter, C. (1995) Recruiting the new conservationists - farmers adoption of agri-environmental schemes in the uk. Journal of Rural Studies, 11, 51-63.
Morse, R. A. and Calderone, N. W. (2000) In Bee Culture Magazine Vol. 128, pp. 1-14.
Nadeem, M., Anjum, F. M., Khan, M. I., Tehseen, S., El-Ghorab, A. and Sultan, J. I. (2013) Nutritional and medicinal aspects of coriander (Coriandrum sativum L.) A review. British Food Journal, 115, 743-755.
New, T. R. (2005) Invertebrate conservation and agricultural ecosystems, Cambridge University Press.
Nicholls, C. I. and Altieri, M. A. (2013) Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agronomy for Sustainable Development, 33, 257-274.
Olson, D. M. and Wackers, F. L. (2007) Management of field margins to maximize multiple ecological services. Journal of Applied Ecology, 44, 13-21.
Perez-Pastor, A., Ruiz-Sanchez, M. C. and Domingo, R. (2014) Effects of timing and intensity of deficit irrigation on vegetative and fruit growth of apricot trees. Agricultural Water Management, 134, 110-118.
Perez-Perez, J. G., Robles, J. M. and Botia, P. (2014) Effects of deficit irrigation in different fruit growth stages on 'Star Ruby' grapefruit trees in semi-arid conditions. Agricultural Water Management, 133, 44-54.
Petanidou, T. and Vokou, D. (1993) Pollination ecology of Labiatae in a phryganic (East Mediterranean) ecosystem. American Journal of Botany, 80, 892-899.
Petit, S., Rubbo, N. and Schumann, R. (2011) Nectar collected with microcapillary tubes is less concentrated than total nectar in flowers with small nectar volumes. Australian Journal of Botany, 59, 593-599.
Pfiffner, L. and Wyss, E. (2004) Use of sown wildflower strips to enhance natural enemies of agricultural pests, CSIRO Publishing.
Philpott, S. M., Uno, S. and Maldonado, J. (2006) The importance of ants and high-shade management to coffee pollination and fruit weight in Chiapas, Mexico. Biodiversity and Conservation, 15, 487-501.
Pineda, A. and Marcos-Garcia, M. A. (2008) Seasonal abundance of aphidophagous

Chapter X
116
hoverflies (Diptera : Syrphidae) and their population levels in and outside Mediterranean sweet pepper greenhouses. Annals of the Entomological Society of America, 101, 384-391.
Pontin, D. R., Wade, M. R., Kehrli, P. and Wratten, S. D. (2006) Attractiveness of single and multiple species flower patches to beneficial insects in agroecosystems. Annals of Applied Biology, 148, 39-47.
Potts, S. G., Biesmeijer, J. C., Kremen, C., Neumann, P., Schweiger, O. and Kunin, W. E. (2010) Global pollinator declines: trends, impacts and drivers. Trends in Ecology & Evolution, 25, 345-353.
Power, A. G. (2010) Ecosystem services and agriculture: tradeoffs and synergies. Philosophical Transactions of the Royal Society B-Biological Sciences, 365, 2959-2971.
Power, E. F. and Stout, J. C. (2011) Organic dairy farming: impacts on insect-flower interaction networks and pollination. Journal of Applied Ecology, 48, 561-569.
Putievsky, E. (1980) Germination studies with seed of caraway, coriander and dill. Seed Science and Technology, 8, 245-254.
Pywell, R. F., Meek, W. R., Hulmes, L., Hulmes, S., James, K. L., Nowakowski, M. and Carvell, C. (2011) Management to enhance pollen and nectar resources for bumblebees and butterflies within intensively farmed landscapes. Journal of Insect Conservation, 15, 853-864.
Rader, R., Reilly, J., Bartomeus, I. and Winfree, R. (2013) Native bees buffer the negative impact of climate warming on honey bee pollination of watermelon crops. Global Change Biology, 19, 3103-3110.
Rands, S. A. and Whitney, H. M. (2011) Field margins, foraging distances and their impacts on nesting pollinator success. Plos One, 6, e25971, 1-8.
Rasmont, P. and Iserbyt, I. (2010) In Atlas HymenopteraMons, Gembloux.
Rasmussen, J. (1991) A model for prediction of yield response in weed harrowing. Weed Research, 31, 401-408.
Reith, M., Baumann, G., Classen-Bockhoff, R. and Speck, T. (2007) New insights into the functional morphology of the lever mechanism of Salvia pratensis (Lamiaceae). Annals of Botany, 100, 393-400.
Reith, M., Classen-Bockhoff, R. and Speck, T. (2006) Biomechanics of salvia flowers: The role of lever and flower tube in specialization on pollinators, Crc Press-Taylor & Francis Group, Boca Raton.
Richards, A. J. (2001) Does low biodiversity resulting from modern agricultural practice affect crop pollination and yield? Annals of Botany, 88, 165-172.
Rundlof, M., Nilsson, H. and Smith, H. G. (2008) Interacting effects of farming practice and landscape context on bumblebees. Biological Conservation, 141, 417-426.
Sandrock, C., Tanadini, L. G., Pettis, J. S., Biesmeijer, J. C., Potts, S. G. and Neumann, P. (2014) Sublethal neonicotinoid insecticide exposure reduces solitary bee reproductive success. Agricultural and Forest Entomology, 16, 119-128.
Santos, S. A. B. d., Roselino, A. C. and Bego, L. R. (2008) Pollination of cucumber, Cucumis sativus L. (Cucurbitales: Cucurbitaceae), by the stingless bees

Chapter X
117
Scaptotrigona aff. depilis moure and Nannotrigona testaceicornis Lepeletier (Hymenoptera: Meliponini) in greenhouses. Neotropical Entomology, 37, 506-12.
Saramago, I. S., Regato, M. A., Guerreiro, I. M. and Regato, J. E. (2012) In Xxviii International Horticultural Congress on Science and Horticulture for People, Vol. 924 (Eds, Tous, J., Gucci, R. and Fevereiro, P.) Int Soc Horticultural Science, Leuven 1, pp. 255-259.
Scriven, L. A., Sweet, M. J. and Port, G. R. (2013) Flower density is more important than habitat type for increasing flower visiting insect diversity. International Journal of Ecology, 2013, 1-12.
Scheper, J., Holzschuh, A., Kuussaari, M., Potts, S. G., Rundlof, M., Smith, H. G. and Kleijn, D. (2013) Environmental factors driving the effectiveness of European agri-environmental measures in mitigating pollinator loss - a meta-analysis. Ecology Letters, 16, 912-20.
Schiestl, F. P. and Johnson, S. D. (2013) Pollinator-mediated evolution of floral signals. Trends in Ecology & Evolution, 28, 307-315.
Schuster, W. S. and Monson, R. K. (1990) An examination of the advantages of C3-C4 intermediate photosynthesis in warm environmnets. Plant Cell and Environment, 13, 903-912.
Showler, A. and Moran, P. (2003) Effects of drought stressed cotton, Gossypium hirsutum L., on beet armyworm, Spodoptera exigua (Hubner), oviposition, and larval feeding prefernces and growth. Journal of Chemical Ecology, 29, 1997-2011.
SIAR (2014) Ministerio de agricultura, alimentación y madio ambiente, http://eportal.magrama.gob.es/websiar/SeleccionParametrosMap.aspx?dst=1.
Sinacori, A., Carrubba, A., Oliveri, E. and Sinacori, M. (2014) Api e pronubi selvatici nella produzione di Coriandrum sativum L. (Famiglia Apiaceae) in Sicilia occidentale. Avenue media; IT.
Smith, H. A. and Chaney, W. E. (2007) A survey of syrphid predators of Nasonovia ribisnigri in organic lettuce on the Central Coast of California. Journal of Economic Entomology, 100, 39-48.
Song, B. Z., Wu, H. Y., Kong, Y., Zhang, J., Du, Y. L., Hu, J. H. and Yao, Y. C. (2010) Effects of intercropping with aromatic plants on the diversity and structure of an arthropod community in a pear orchard. Biocontrol, 55, 741-751.
Steffan-Dewenter, I., Potts, S. G. and Packer, L. (2005) Pollinator diversity and crop pollination services are at risk. Trends in Ecology & Evolution, 20, 651-652.
Steffan-Dewenter, I. and Tscharntke, T. (2000) Resource overlap and possible competition between honey bees and wild bees in central Europe. Oecologia, 122, 288-296.
Steingrover, E. G., Geertsema, W. and van Wingerden, W. (2010) Designing agricultural landscapes for natural pest control: a transdisciplinary approach in the Hoeksche Waard (The Netherlands). Landscape Ecology, 25, 825-838.
Stokstad, E. (2013) Pesticides under fire for risks to pollinators. Science, 340, 674-676.

Chapter X
118
Stout, J. C., Allen, J. A. and Goulson, D. (1998) The influence of relative plant density and floral morphological complexity on behaviour of bumblebees. Oecologia, 117, 543-550.
Stout, J. C. and Goulson, D. (2002) The influence of nectar secretion rates on the responses of bumblebees (Bombus spp.) to previously visited flowers. Behavioral Ecology and Sociobiology, 52, 239-246.
Stout, J. C. and Morales, C. L. (2009) Ecological impacts of invasive alien species on bees. Apidologie, 40, 388-409.
Stubbs, A. E. and Falk, S. (2002) British hoverflies: an illustrated identification guide. Second edition, British Entomological and Natural History Society.
Subba Reddi, C. and Meera Bai, G. (1984) Butterflies and pollination biology. Proceedings of the Indian Academy of Sciences Animal Sciences, 93, 391-396.
Tinkeu, L. S. N., VanImpe, G., Hance, T. and Univ, G. (1996) In 48th International Symposium on Crop Protection, Pts I-Iv, Vol. 61, pp. 917-925.
Tscharntke, T., Klein, A. M., Kruess, A., Steffan-Dewenter, I. and Thies, C. (2005) Landscape perspectives on agricultural intensification and biodiversity - ecosystem service management. Ecology Letters, 8, 857-874.
Tuell, J. K., Fiedler, A. K., Landis, D. and Isaacs, R. (2008) Visitation by wild and managed bees (Hymenoptera : Apoidea) to eastern US native plants for use in conservation programs. Environmental Entomology, 37, 707-718.
Ueno, O., Wada, Y., Wakai, M. and Bang, S. W. (2006) Evidence from photosynthetic characteristics for the hybrid origin of Diplotaxis muralis from a C-3-C-4 intermediate and a C-3 species. Plant Biology, 8, 253-259.
Van der Sluijs, J. P., Simon-Delso, N., Goulson, D., Maxim, L., Bonmatin, J. M. and Belzunces, L. P. (2013) Neonicotinoids, bee disorders and the sustainability of pollinator services. Current Opinion in Environmental Sustainability, 5, 293-305.
Vanbergen, A. J. and Insect Pollinators Initiative (2013) Threats to an ecosystem service: pressures on pollinators. Frontiers in Ecology and the Environment, 11, 251-259.
Walther-Hellwig, K., Fokul, G., Frankl, R., Buchler, R., Ekschmitt, K. and Wolters, V. (2006) Increased density of honeybee colonies affects foraging bumblebees. Apidologie, 37, 517-532.
Wangensteen, H., Samuelsen, A. B. and Malterud, K. E. (2004) Antioxidant activity in extracts from coriander. Food Chemistry, 88, 293-297.
Watling, J. I. and Donnelly, M. A. (2006) Fragments as islands: a synthesis of faunal responses to habitat patchiness. Conservation Biology, 20, 1016-1025.
Winfree, R., Aguilar, R., Vazquez, D. P., LeBuhn, G. and Aizen, M. A. (2009) A meta-analysis of bees' responses to anthropogenic disturbance. Ecology, 90, 2068-2076.
Woodcock, B. A., Bullock, J. M., Nowakowski, M., Orr, R., Tallowin, J. R. B. and Pywell, R. F. (2012) Enhancing floral diversity to increase the robustness of grassland beetle assemblages to environmental change. Conservation Letters, 5,

Chapter X
119
459-469.
Woodcock, B. A., Edwards, M., Redhead, J., Meek, W. R., Nuttall, P., Falk, S., Nowakowski, M. and Pywell, R. F. (2013) Crop flower visitation by honeybees, bumblebees and solitary bees: Behavioural differences and diversity responses to landscape. Agriculture Ecosystems & Environment, 171, 1-8.
Wratten, S. D., Gillespie, M., Decourtye, A., Mader, E. and Desneux, N. (2012) Pollinator habitat enhancement: Benefits to other ecosystem services. Agriculture Ecosystems & Environment, 159, 112-122.
Zhang, Y. W., Robert, G. W., Wang, Y. and Guo, Y. H. (2007) Nectar robbing of a carpenter bee and its effects on the reproductive fitness of Glechoma longituba (Lamiaceae). Plant Ecology, 193, 1-13.
Zihlif, M., Afifi, F., Abu-Dahab, R., Majid, A., Somrain, H., Saleh, M. M., Nassar, Z. D. and Naffa, R. (2013) The antiangiogenic activities of ethanolic crude extracts of four Salvia species. Bmc Complementary and Alternative Medicine, 13.
Znidarcic, D., Ban, D. A. and Sircelj, H. (2011) Carotenoid and chlorophyll composition of commonly consumed leafy vegetables in Mediterranean countries. Food Chemistry, 129, 1164-1168.


Chapter XI
121
XI. Appendices
1. List of abbreviations
AES – Agri-environmental Schemes
Ali – Alium schoenoprasum
Ant – Antirrhinum majus
ArcGIS – Arc Geographic Information System
Bor – Borago officinalis
Cal – Calendula arvensis
CATPCA – Categorical Partial Component Analysis
Cen – Centaurea cyanus
Cor – Coriandrum sativum
CSIC – Consejo Superior de Investigaciones Científicas
D – Drought
D-11 – Drought of 11 days
D-14 – Drought of 14 days
D-4 – Drought of 4 days
D-8 – Drought of 8 days
DCA – Discriminate Canonical Analysis
Dpl – Diplotaxis tenuifolia
Ech – Echium plantagineum
ETp – Potential Evapotranspiration
GLM – General Linear Model
gs – Stomatal Conductance
Lob – Lobularia maritima

Chapter XI
122
M – Mixture of floral species
MASL – Meters above Sea Level
MDI – Moderate Deficit Irrigation
MDI-BF – Moderate Deficit Irrigation – before flowering
MDI-DF – Moderate Deficit Irrigation – during flowering
NE-SW – North-East – South-West
Pha – Phacelia tanacetifolia
R19 – recover of 19 days after drought treatments applied
RDI – Regulated Deficit Irrigation
RDI-BF – Regulated Deficit Irrigation – before flowering
RDI-DF – Regulated Deficit Irrigation – during flowering
RTK-GPS – Real Time Kinematic-the Global Positioning System
SDI – Severe Deficit Irrigation
SDI-BF – Severe Deficit Irrigation – before flowering
SDI-DF – Severe Deficit Irrigation – during flowering
SE – Standard Error
SN/inf – Seed Number per Inflorescence
sp. – species
spp. – species (plural)
SPSS – Statistical Package for the Social Sciences
SW – Seed Weight
Tag – Tagetes patula
UK – The United Kingdom
USA – The United States of America

Chapter XI
123
2. List of figures
I General introduction
Fig. I-1. Percentage of agricultural land relative to the total land area across the Earth
Fig. I-2. Different groups of pollinators [1. Honey bee (Apidae: Apis sp.); 2. Bumblebee
(Apidae: Bombus sp.); 3. Carpenter bee (Apidae: Xylocopa sp.); 4. Digger bee (Apidae:
Amegilla sp.); 5. Leafcutter bee (Megachilidae: Megachile sp.); 6. Mining bee
(Andrenidae: Andrena sp.; 7. Sweat bee (Halictidae: Halictus sp.; 8. Hoverfly
(Syrphidae: Sphaerophoria scripta.); 9. Beetle (Coleoptera: Heliotaurus ruficollis)]
Fig. I-3. Beneficial insectary plants [1. Buckwheat (Fagopyrum esculentum Moench);
2. Phacelia (Phacelia tanacetifolia Benth.); 3. Alyssum (Lobularia maritima L. Desv.);
4. Coriander (Coriandrum sativum L.)]
Fig. I-4. Sowed field margins: 1. Mono-specific field margin (Phacelia tanacetifolia
Benth.), 2. Mixed field margin
Fig. I-5: Use of beneficial insectary plants in small-scale gardening
III Functionality of aromatic (Lamiaceae) floral mixture in conservation of
pollinators in Central Spain
Fig. III-1. Experimental design (randomized block design - each number represents one
Lamiaceae plant species (mono-specific plot), while M1 and M2 represent mixed plots)
Fig. III-2. Discrimination among plant species upon the first and second canonical axis
Fig. III-3. Relationship between morphological characteristics and number of insect
visits in 2012 (A) and in 2013 (B)
Fig. III-4. Relationship between floral phenology and insect visitation
IV The attractiveness of flowering herbaceous plants to bees (Hymenoptera:
Apoidea) and hoverflies (Diptera: Syrphidae) in agro-ecosystems of Central Spain
Fig. IV-1. Attractiveness efficiency of flower species. Schematic diagram depicting the
relationship between the total number of insect visits/minute (y-axis) and flowering

Chapter XI
124
duration (number of observation days) of plant species (x-axis): A (upper-right corner):
high total number of insect visits/minute in a long observation period (high
attractiveness efficiency); B (upper-left corner): high total number of insect
visits/minute in a short observation period; C (bottom-right corner): low total number of
insect visits/minute in a long observation period; and D (bottom-left corner): low total
number of insect visits/minute in a short observation period
Fig. IV-2. Visits of bees, hoverflies and beetles to the flowers of the studied plant
species. Means with same letter in each graph are not significantly different at P ≤ 0.05,
as determined by Tukey’s B multiple comparison test. Abbreviations: 2011 (1): Bor=
Borago officinalis; Cal= Calendula arvensis; Cen= Centaurea cyanus; Dpl= Diplotaxis
tenuifolia; Pha= Phacelia tanacetifolia; Ali= Alium schoenoprasum; M1= mixture of
those species. 2011 (2): Cor= Coriandrum sativum; Ech= Echium plantagineum; Lob=
Lobularia maritima; Tag= Tagetes patula; Ant= Antirrhinum majus; M2= mixture of
those species. 2012: M= mixture of the species used in 2012
Fig. IV - 3. Attractiveness efficiency of the studied plant species for bees and hoverflies
in 2011 and 2012 (abbreviations of plant species as in Fig. IV-2.). Scatter-plots depict
the relationship between the total number of insect visits/minute (y-axis) and flowering
duration (number of observation days) of plant species (x-axis). Bars represent the
standard deviation from 3 replicated plots
V Wild rocket - effect of water deficit on growth, flowering and attractiveness to
pollinators
Fig. V-1. Mean stomatal conductance of D. tenuifolia under different drought
treatments (no irrigation for 4 [D-4], 8 [D-8], 11 [D-11] or 14 days [D-14]) (A) and
during a recovery period (well-irrigated) of 19 days (B). For each measurement (day),
means with different letter are significantly different at P ≤ 0.05, as determined by the
Tukey’s B multiple comparison test.
Fig. V-2. Concentration of sugars in nectar after different drought treatments (no
irrigation for 4 [D-4], 8 [D-8], 11 [D-11] or 14 days [D-14]) (A) and after re-watering of
19 days (B). No significant differences were found between drought treatments. The

Chapter XI
125
vertical bars represent standard errors. Concentrations of sugars in D-14 after re-
watering are measured in only one plant that recovered after drought stress.
Fig. V-3. Number of pollinators visiting flowers of D. tenuifolia after different drought
treatments (no irrigation for 4 [D-4], 8 [D-8] and 11 days [D-11]) in two sites of Central
Spain: La Poveda (A) and La Sierra (B). For each treatment, means that are significantly
different at P ≤ 0.05 are marked with different letter (comparison within insect groups),
as determined by the Tukey’s B multiple comparison test. The vertical bars represent
standard errors.
Fig. V-4. Mean stomatal conductance of D. tenuifolia under moderate deficit irrigation
(MDI) and severe deficit irrigation (SDI). Deficit irrigation applied before and during
flowering (A) and deficit irrigation applied only during flowering (B). For each
measurement, means with different letter are significantly different at P ≤ 0.05, as
determined by the Tukey’s B multiple comparison test.
Fig. V-5. Number of pollinators visiting flowers of D. tenuifolia after moderate deficit
irrigation (MDI) and severe deficit irrigation (SDI). Deficit irrigation applied before and
during flowering (A) and deficit irrigation applied only during flowering (B). For each
treatment, means with different letter are significantly different at P ≤ 0.05 (comparison
within insect groups), as determined by Tukey’s B multiple comparison test. The
vertical bars represent standard errors.
VI Can proximity of floral field margins improve pollination and seed production
in coriander, Coriandrum sativum L. (Apiaceae)?
Fig. VI-1. Germination rate of coriander seeds. Covered inflorescences of coriander
(covered) and not-covered inflorescences of coriander in vicinity of: mono-specific
margins (NO - D. tenuifolia), mixed margin (NO - mixed) and without margins (NO -
control). Means with same letter in each graph are not significantly different at P ≤ 0.05,
as determined by Tukey’s B multiple comparison test. Seed germination in: (A) sample
of 25 seeds x 4 repetitions and (B) sample of 10 seeds x 3 repetitions.

Chapter XI
126
3. List of tables
III Functionality of aromatic (Lamiaceae) floral mixture in conservation of
pollinators in Central Spain
Table III-1. Aromatic plant species used
Table III-2. Mean (± SE) insect visits per minute to flowers of selected aromatic plant
species in 2012 and 2013
Table III-3. Morphological characteristics of flowers of the six Lamiaceae species
studied
Table III-4. Correlation between discriminating variables and 1st and 2nd standardized
canonical discriminant functions (80.04%)
Table III-5. Spatial increase (%) of the area occupied by plant species one year after
planting
IV The attractiveness of flowering herbaceous plants to bees (Hymenoptera:
Apoidea) and hoverflies (Diptera: Syrphidae) in agro-ecosystems of Central Spain
Table IV-1. Plant species and their flowering periods in 2011 and 2012
Table IV-2. List of insects visiting the studied plant species
Table IV-3. Differences in seedling emergence (Nº plants/m2) after applying two
different managements at the end of plants cycle: shallow tillage and no-tillage
Table IV-4. Spatial increase (%) of the area occupied by plant species one year after
planting
V Wild rocket - effect of water deficit on growth, flowering and attractiveness to
pollinators
Table V-1. Effects of different drought treatments (no irrigation for 4 [D-4], 8 [D-8],
11 [D-11] or 14 days [D-14]) and of re-watering over 19 days on morphological
characteristics of D. tenuifolia plants

Chapter XI
127
Table V-2. Effect of different drought treatments (no irrigation for 4 [D-4], 8 [D-8], 11
[D-11] or 14 days [D-14]) and of re-watering over 19 days on the floral characteristics
of D. tenuifolia
Table V-3. Effect of different regulated deficit irrigation (RDI) treatments (moderate
deficit irrigation before flowering (MDI-BF) and during flowering (MDI-DF); and
severe deficit irrigation before flowering (SDI-BF) and during flowering (SDI-DF) on
the vegetative characteristics of D. tenuifolia. Measured after the treatments were done.
Table V-4. Effect of different regulated deficit irrigation (RDI) regimes (moderate
deficit irrigation before flowering (MDI-BF) and during flowering (MDI-DF); and
severe deficit irrigation before flowering (SDI-BF) and during flowering (SDI-DF) on
the floral characteristics of D. tenuifolia. Measured after the treatments were done.
Table V-5. Total water consumption per plant for irrigation of potted D. tenuifolia
during 30 days experiment after drought treatments (D) of 4 [D-4], 8 [D-8], 11 [D-11]
or 14 days [D-14]) and regulated deficit irrigation (RDI) treatments (moderate deficit
irrigation before flowering [MDI-BF] and during flowering [MDI-DF]; and severe
deficit irrigation before flowering [SDI-BF] and during flowering [SDI-DF]
VI Can proximity of floral field margins improve pollination and seed production
in coriander, Coriandrum sativum L. (Apiaceae)?
Table VI-1: Mean (± S.E.) flower density and mean (± S.E.) number of visits of
pollinators to the flowers of mono-specific and mixed floral margins
Table VI-2. Mean (± S.E.) flower density and mean (± S.E.) number of visits of
pollinators to the flowers of coriander without proximate floral margins (control) and in
vicinity of mono-specific and mixed margins
Table VI-3. Correlations between number of flowers and number of pollinators'
visits/min
Table VI-4. Potential (± S.E.), actual (± S.E.) and percentage (± S.E.) of actual seed
number (SN) per inflorescence in covered inflorescences of coriander under control,
mono-specific and mixed margin treatments

Chapter XI
128
Table VI-5. Potential (± S.E.), actual (± S.E.) and percentage (± S.E.) of actual seed
number (SN) per inflorescence of coriander in not-covered (control, mono-specific and
mixed margin treatments) vs. covered inflorescences
Table VI-6. Potential (± S.E.), actual (± S.E.) and percentage (± S.E.) of actual seed
number (SN) per inflorescence of coriander, and seed weight (SW) per seed (± S.E.),
SW per plant (± S.E.) and percentage of SW per plant (± S.E.) under control, mono-
specific and mixed margin treatments