
UNCOMFORTABLE PROTECTION: FÖRST. SHELTERS FORMICA FUSCA...
10
INTRODUCTION Formica (Serviformica) fusca L. is biotically re- lated both to F. (Formica) polyctena Först. and F. (Raptiformica) sanguinea Latr. in a special way. It is, at least in Central Europe, the most frequent host species for temporary social parasitism of F. polycte- na at nest foundation (and in the case of other wood ants of the subgenus Formica s. str.; see e.g., Cze- chowski and Pisarski 1994, Pisarski and Czechowski 1994) and the most frequent slave species of F. sangu- inea [and Polyergus rufescens (Latr.); see e.g., Kut- ter 1968, Buschinger et al. 1980]; for definitions of so- cial parasitism and its particular forms in ants see e.g. Buschinger (1986, 1990) and Hölldobler and Wilson (1990). Both territorial species thus affect negatively F. fusca, though not in the same way. Consequently these three species are closely interrelated ecological- ly, besides being common co-members of ant assem- blages in boreal and temperate forest habitats. The multi-species ant assemblages are organized hierar- chically, and are dominated by territorial species, i.e. species whose colonies defend their foraging areas (Pisarski 1973, 1982, Gallé 1975, 1978, Vepsäläinen and Pisarski 1982, Reznikova 1983, Savolainen and Vep- säläinen 1988). Nests of such colonies play the role of spatial organizing centres of local ant communities (Savolainen and Vepsäläinen 1988, 1989, Savolainen et UNCOMFORTABLE PROTECTION: FORMICA POLYCTENA FÖRST. SHELTERS FORMICA FUSCA L. FROM FORMICA SANGUINEA LATR. (HYMENOPTERA: FORMICIDAE) ANNALES ZOOLOGICI (Warszawa), 2006, 56(3): 539-548 WOJCIECH CZECHOWSKI 1 and BÁLINT MARKÓ 2 1 Laboratory of Social and Myrmecophilous Insects, Museum and Institute of Zoology, Polish Academy of Sciences, Wilcza 64, 00-679 Warszawa, Poland e-mail: [email protected] 2 Department of Taxonomy and Ecology, Babeş-Bolyai University, Clinicilor 5-7 3400 Cluj-Napoca, Romania; e-mail: [email protected] Abstract.— The wood ant Formica polyctena Först. is a territorial species, a regular top dominant of ant communities in forests. Its colonies defend their whole foraging areas (territories) against other territorial ants, including F. sanguinea Latr., a common facul- tative slave-maker. The most frequent ‘victim’ of F. sanguinea is F. fusca L., a ubiquitous submissive ant species. On the basis of some earlier observations, the presumption was made that F. polyctena, when defending its own territories, would indirectly protect F. fusca colonies, which nest within these territories, from F. sanguinea raids. It was expected that F. fusca should be more abundant in F. polyctena territories, than in F. sanguinea territories, while other subordinate ants, which are not potential slaves of F. sanguinea, should not show such difference. This hypothesis was supported by the results of the baiting experiments carried out in the Białowieża Forest, NE Poland. The findings are discussed in the context of interspecific competition hierarchy in ants. Key words.— Ants, Formicidae, Formica polyctena, Formica fusca, Formica san- guinea, Polyergus rufescens, territoriality, interspecific competition, competition hierar- chy, foraging, slavery.
Transcript of UNCOMFORTABLE PROTECTION: FÖRST. SHELTERS FORMICA FUSCA...

INTRODUCTION
Formica (Serviformica) fusca L. is biotically re-lated both to F. (Formica) polyctena Först. and F. (Raptiformica) sanguinea Latr. in a special way.It is, at least in Central Europe, the most frequent hostspecies for temporary social parasitism of F. polycte-na at nest foundation (and in the case of other woodants of the subgenus Formica s. str.; see e.g., Cze-chowski and Pisarski 1994, Pisarski and Czechowski1994) and the most frequent slave species of F. sangu-inea [and Polyergus rufescens (Latr.); see e.g., Kut-ter 1968, Buschinger et al. 1980]; for definitions of so-cial parasitism and its particular forms in ants see e.g.
Buschinger (1986, 1990) and Hölldobler and Wilson(1990). Both territorial species thus affect negatively F. fusca, though not in the same way. Consequentlythese three species are closely interrelated ecological-ly, besides being common co-members of ant assem-blages in boreal and temperate forest habitats. Themulti-species ant assemblages are organized hierar-chically, and are dominated by territorial species, i.e.species whose colonies defend their foraging areas(Pisarski 1973, 1982, Gallé 1975, 1978, Vepsäläinen andPisarski 1982, Reznikova 1983, Savolainen and Vep-säläinen 1988). Nests of such colonies play the role ofspatial organizing centres of local ant communities(Savolainen and Vepsäläinen 1988, 1989, Savolainen et
UNCOMFORTABLE PROTECTION: FFOORRMMIICCAA PPOOLLYYCCTTEENNAAFÖRST. SHELTERS FFOORRMMIICCAA FFUUSSCCAA L. FROM FFOORRMMIICCAA
SSAANNGGUUIINNEEAA LATR. (HYMENOPTERA: FORMICIDAE)
AA NN NN AA LL EE SS ZZ OO OO LL OO GG II CC II ((WWaarrsszzaawwaa)),, 22000066,, 5566((33)):: 553399--554488
WOJCIECH CZECHOWSKI1 and BÁLINT MARKÓ2
1Laboratory of Social and Myrmecophilous Insects, Museum and Institute of Zoology, Polish Academy of Sciences, Wilcza 64, 00-679 Warszawa, Poland
e-mail: [email protected] of Taxonomy and Ecology, Babeş-Bolyai University, Clinicilor 5-7
3400 Cluj-Napoca, Romania; e-mail: [email protected]
Abstract.— The wood ant Formica polyctena Först. is a territorial species, a regular topdominant of ant communities in forests. Its colonies defend their whole foraging areas(territories) against other territorial ants, including F. sanguinea Latr., a common facul-tative slave-maker. The most frequent ‘victim’ of F. sanguinea is F. fusca L., a ubiquitoussubmissive ant species. On the basis of some earlier observations, the presumption wasmade that F. polyctena, when defending its own territories, would indirectly protect F. fusca colonies, which nest within these territories, from F. sanguinea raids. It wasexpected that F. fusca should be more abundant in F. polyctena territories, than in F. sanguinea territories, while other subordinate ants, which are not potential slaves of F. sanguinea, should not show such difference. This hypothesis was supported by theresults of the baiting experiments carried out in the Białowieża Forest, NE Poland. Thefindings are discussed in the context of interspecific competition hierarchy in ants.
Key words.— Ants, Formicidae, Formica polyctena, Formica fusca, Formica san-guinea, Polyergus rufescens, territoriality, interspecific competition, competition hierar-chy, foraging, slavery.

al. 1989). Non-territorial (subordinate) ants may nestonly at a certain distance from the dominant’s nest andsuffer other restrictions regarding e.g. foraging activi-ty (Punttila et al. 1991, Pisarski and Vepsäläinen 1989;see also Reznikova 1999). Relations between wood ant F. polyctena as the top dominant and F. fusca asa typical subordinate, submissive species perfectlyillustrate this phenomenon (see Savolainen 1990,1991). Usually subordinate species are more abundantoutside, than within territories of dominant species insimilar habitats.
The matter is not so explicit in the case of thosesubordinate species, which, alike F. fusca, are poten-tial slave species for the common facultative slave-maker, F. sanguinea, if the latter is present in the specific area. In respect of its hierarchical status, F. sanguinea is a territorial species subordinatedeven to moderately sized colonies of wood ants(Czechowski and Vepsäläinen 2001). During its raidingperiods, however, it does not respect the boundaries offoreign territories (Czechowski 2000), including thoseof wood ants. Competition between wood ants and F.sanguinea indirectly influences the distribution andabundance of potential slave species of F. sanguinea,including F. fusca (Punttila et al. 1996). Behaviouralmechanisms of this influence were observed and de-scribed specifically in Lasius fuliginosus (Latr.) andthe wood ant Formica rufa L. as the top dominant,territorial species on one hand, and Formica cinereaMayr and F. fusca as F. sanguinea’s slave species onthe other (see Czechowski 1999, 2000). The effect ofrelationships between F. rufa and F. sanguinea on thedistribution of F. fusca was estimated quantitatively intwo field arrangements of monodomous F. rufa coloniesand one versus several linearly situated colonies of F.sanguinea (Czechowski and Vepsäläinen 2001). Theabundance of F. fusca turned out to be higher within ter-ritories of the dominant species, than outside them, if F.sanguinea was present in the neighbourhood.
Would the above phenomena still occur so eloquent-ly if the competitive pressure of the territorial wood ant species was stronger? A polydomous system of a territorial species could have greater negative impacton subordinate species, than a monodomous colony.The aim of the present study is to verify and possiblygeneralize this principle, testing the possible protectiverole of the wood ant species with a different socialorganization, namely polydomous F. polyctena, inmore complicated field arrangement of the coloniesand territories of competitive species, and under differ-ent habitat conditions. On the basis of the formerresults, the following hypothesis can be formulated:there should be a clear-cut difference between the territories of F. polyctena and F. sanguinea, which border each other, as regards the presence of F. fusca. F. fusca should be more abundant within the
F. polyctena territory, than within that of F. san-guinea, while other subordinate species, which are notpotential slaves of F. sanguinea, should not show suchdifference.
STUDY AREA
The investigations were carried out in the managedpart of the Białowieża Forest, NE Poland, in July 2001.The study area was a fairly homogenous part of a neg-lected forest plantation stretching as a ca. 15-m widestrip between a mature mixed coniferous-deciduous(pine, spruce, oak, hornbeam, birch) forest and a neg-lected thick, mainly deciduous, young coppice (Figs 1, 2). The top dominant of the local ant community inrespect of the competitive hierarchy was an extentpolydomous F. polyctena colony consisting mainly ofsmall nests situated closely to each other at old sprucestumps (Fig. 3). The vast territory of F. polyctenaspread throughout the forest and an adjacent openarea including the above-mentioned strip of the for-est plantation. Here and there, territories of single F. sanguinea colonies, the local subdominant territo-rial species, cut into the F. polyctena’s territory on theplantation area and at the forest margin. The territo-ries of F. polyctena and F. sanguinea bordered eachother (Fig. 4), and both were settled and searched by F. fusca. In the forest interior, the F. polyctena’s territory was partly cut by the territory of a Lasiusfuliginosus colony.
The study area comprised twelve colonies (= nests)of F. polyctena (FP-I–XII). Colony FP-I – with the big-gest (Ø 1 m, h 30 cm) and typically formed nest mound– was probably the maternal colony for the whole polydomous system. There were also two colonies of F. sanguinea (FS-I and FS-II), both with F. fuscaslaves. A part of the investigated F. polyctena territo-ry lay within the range of raids of the obligatory slave-maker, Polyergus rufescens (Fig. 4), 55 m away fromits nest (see Czechowski 2002, 2007), which makes theresult even more interesting. Due to a high density ofsmall, but densely-populated F. polyctena nests thedynamic density of wood ant workers within the terri-tory of F. polyctena under discussion was very high,visibly higher than those within the territories of theformerly studied monodomous F. rufa colonies (seeCzechowski and Vepsäläinen 2001).
MATERIAL AND METHODS
Baiting observations
Baiting is widely used for studying interactionsamong different species of ants. It is also appropriate
540 W. CZECHOWSKI and B. MARKÓ

UNCOMFORTABLE PROTECTION: FORMICA POLYCTENA SHELTERS F. FUSCA FROM F. SANGUINEA 541
Figure 1. Study area seen along the transect from the Formica polyctena territory (photo W. Czechowski).
Figure 4. Map of the study site, the location of the bait transect in the territories of Formica polyctena (nests FP-I–XII) and F. san-guinea (nests FS-I and FS-II); CV – the nest of Camponotus vagus
(1...16, 17...34...37...49...51 – numbers of baits).
Figure 2. Study area seen along the transect from the Formicasanguinea territory (photo W. Czechowski).
Figure 3. Small stump-side Formica polyctena nest, typical of the studied area (photo W. Czechowski).
for clarifying the position of each ant species in thecommunity and to study the foraging pattern of antsaround their nests as well (e.g. Reznikova 1975, 1980,Vepsäläinen and Savolainen 1990, Czechowski andVepsäläinen 2001, Markó and Czechowski 2004, Cze-chowski and Markó 2005).
On the pattern of the previous study (Czechowskiand Vepsäläinen 2001) a transect of 51 fruit syrup baits
10 m

was established within the studied area, i.e., along the margin of the vast part of the forest plantation and within the narrow strip between the forest and the young coppice. Baits were put on the ground, every 2 m, in a line (bifurcated at the end) that was constructed according to the topography of the area.The transect crossed both the territories of F. poly-ctena and F. sanguinea, which intermingled each other on this area; the F. polyctena territory crossedthe F. sanguinea territory on a small section of thetransect (baits 35 and 36) (Fig. 4). The territory bor-ders were determined by (1) observing the distributionof foragers of each of the two territorial species beforeand during the baiting experiment, and (2) their pres-ence/absence on particular baits.
Twelve series of readings were carried out at differ-ent times of the day, from late morning (ca. 1030 a.m.)till evening (ca. 700 p.m.), within four days (23–26 July),2–4 series per day at at least three-hour intervals.Baits were replenished when necessary. The presenceand numbers of workers of the ant species wererecorded for each bait. As F. fusca is so called oppor-tunistic species, submissive to foragers of dominantspecies, which are present at a given bait, also those F. fusca foragers were recorded, which were observedaround a bait (thus presumably interested in it) withina 10 cm radius.
Data analysis
The abundance of F. fusca (and other subordinateants) recorded on baits can be used to estimate the sizeof the population in a given area, i.e. within the F. po-lyctena’s or F. sanguinea’s territory. It is not a fault-less estimate, however, it appears to be fairly suitablein the case of relatively big areas (see Czechowski andVepsäläinen 2001). The smaller the area, the smallerthe accuracy of this measure is because of the increasein the probability of recording foragers coming fromneighbouring areas as residents by mistake, especiallyin the case of such mobile ants as F. fusca. That iswhy, the data from baits 35 and 36 (the narrow strip of
F. polyctena territory close to colony FP-XI; Fig. 4)was omitted from the analysis. The data from baits1–16 were used for an appraisal of the situation withinthe territory of F. polyctena, and from baits 17–34 and37–51 for that within the territories of F. sanguinea(see Fig. 4).
The number of individuals was pooled and aver-aged by the number of replicates (n = 12) separa-tely for baits on the territories of F. polyctena and F. sanguinea. The mean number of individuals perreplicate per bait was used further on, with the ex-ception of the analyses of occurrences and co-occur-rences of different species with territorial species onbaits. In these cases every observation was handledseparately.
The datasets were not normally distributed, thusnon-parametric tests (Mann-Whitney U-test, Spearmanrank-correlation) were used for statistical analyses.The evenness of the presence of specific ant species onbaits was estimated on the basis of Shannon-Wienerentropy (log2) calculated over the 12 replicates for eachbait, then the average evenness was taken into consid-eration (max. possible HSh = 3.58).
RESULTS
Species and their occurrences
Altogether eight species were observed on baits(Table 1). Most frequent were: F. polyctena, F. sangu-inea and F. fusca. Besides these three other speciesexploited baits quite frequently: Camponotus vagus(Scop.) (from the only nest present within the studyarea; Fig. 4), Myrmica ruginodis Nyl. and Tetramor-ium caespitum (L.). Two additional species occurredon baits sporadically: five Leptothorax acervorum(F.) foragers on F. polyctena’s territory, and one forager of L. acervorum and Lasius (Lasius s. str.)sp. (most probably L. platythorax Seifert) on the territory of F. sanguinea. Lasius fuliginosus whichnested in the forest interior did not reach the line ofbaits.
542 W. CZECHOWSKI and B. MARKÓ
Table 1. Presence of the most frequent ant species on baits and their mean rate of occurrence separately on the territories of Formica polyctenaand F. sanguinea (Fp – F. polyctena, Fs – F. sanguinea; for data on Leptothorax acervorum and Lasius sp. see the text). * – no. ofobservations = 192 and 396 for the territories of F. polyctena and F. sanguinea respectively; ** – max. no. of occurrences in each replicate = 16
and 33 baits for the territories of F. polyctena and F. sanguinea respectively.
Species F. polyctena F. sanguinea F. fusca C. vagus M. ruginodis T. caespitum
Territory Fp Fs Fp Fs Fp Fs Fp Fs Fp Fs Fp FsTotal number of foragers 1941 6 – 1101 654 649 160 – 82 632 2491 435
Mean number of foragers 10.63 0.02 – 2.78 3.58 1.64 0.85 – 0.42 1.6 12.91 1.1per bait and SD* 12.28 0.26 4.23 4.89 2.12 1.99 2.58 4.56 66.98 4.2
Mean occurrence rate** 0.7 0.01 – 0.72 0.68 0.61 0.2 – 0.09 0.31 0.14 0.19
The species’ mean rate of occurrence on baits gen-erally correlated with the above pattern (Table 1): ter-ritorial Formica species occurred mostly or exclusi-vely (e.g. F. sanguinea) on their own territory, where-as F. fusca exploited sources almost equally fre-quent on territories of both species. Interestingly, inspite of its sporadically high number observed on baits,Tetramorium caespitum occurred with low frequen-cy on baits (Table 1). This may be due to competition orto the fact that the distances of its nests to the baitswere generally too great.
The Shannon-Wiener evenness of the presence ofdifferent ant species offered basically the same picturewith slight adjustments (Table 2). The evenness of F. polyctena’s and F. sanguinea’s presence was thehighest on their territories among all species, followedby F. fusca, whereas, though it was observed in lowernumbers, a high evenness was shown by Camponotusvagus on the territory of F. polyctena, as well. Conse-quently, C. vagus appeared in low numbers on a fewbaits, but its presence on those sources was even overthe observation period. The low evenness of Tetramo-rium caespitum’s presence emphasized its sporadicpresence on sources, along the low rate of occurrence.
Between territory differences
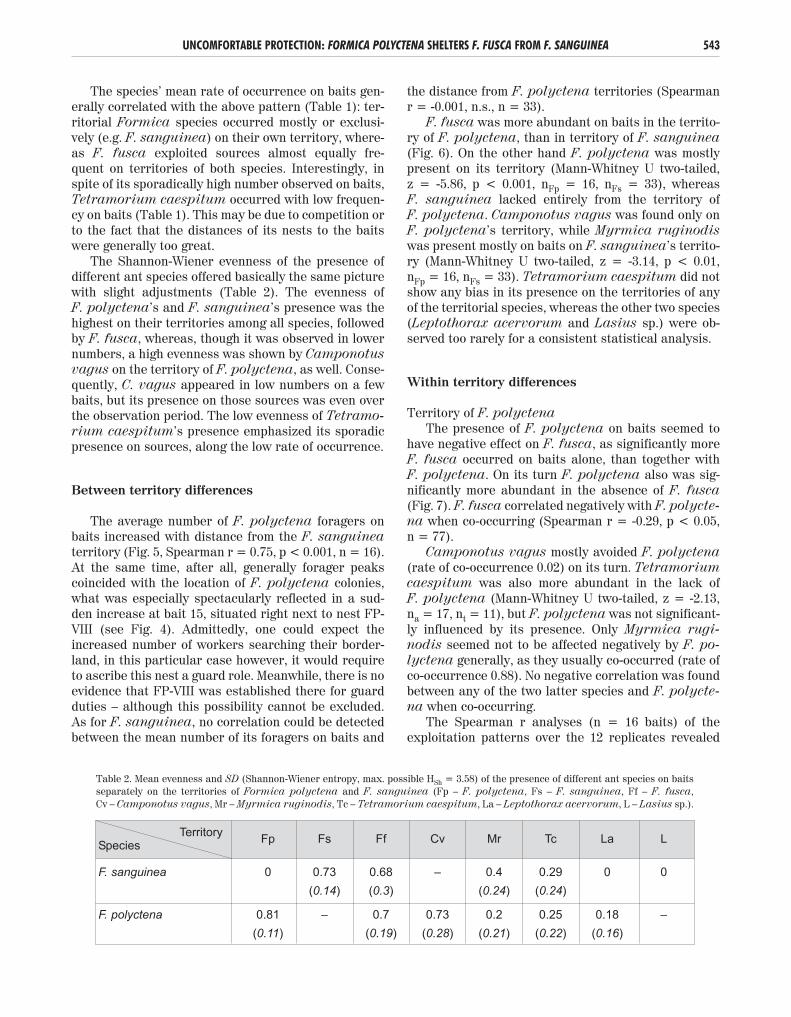
The average number of F. polyctena foragers onbaits increased with distance from the F. sanguineaterritory (Fig. 5, Spearman r = 0.75, p < 0.001, n = 16).At the same time, after all, generally forager peakscoincided with the location of F. polyctena colonies,what was especially spectacularly reflected in a sud-den increase at bait 15, situated right next to nest FP-VIII (see Fig. 4). Admittedly, one could expect theincreased number of workers searching their border-land, in this particular case however, it would requireto ascribe this nest a guard role. Meanwhile, there is noevidence that FP-VIII was established there for guardduties – although this possibility cannot be excluded.As for F. sanguinea, no correlation could be detectedbetween the mean number of its foragers on baits and
the distance from F. polyctena territories (Spearmanr = -0.001, n.s., n = 33).
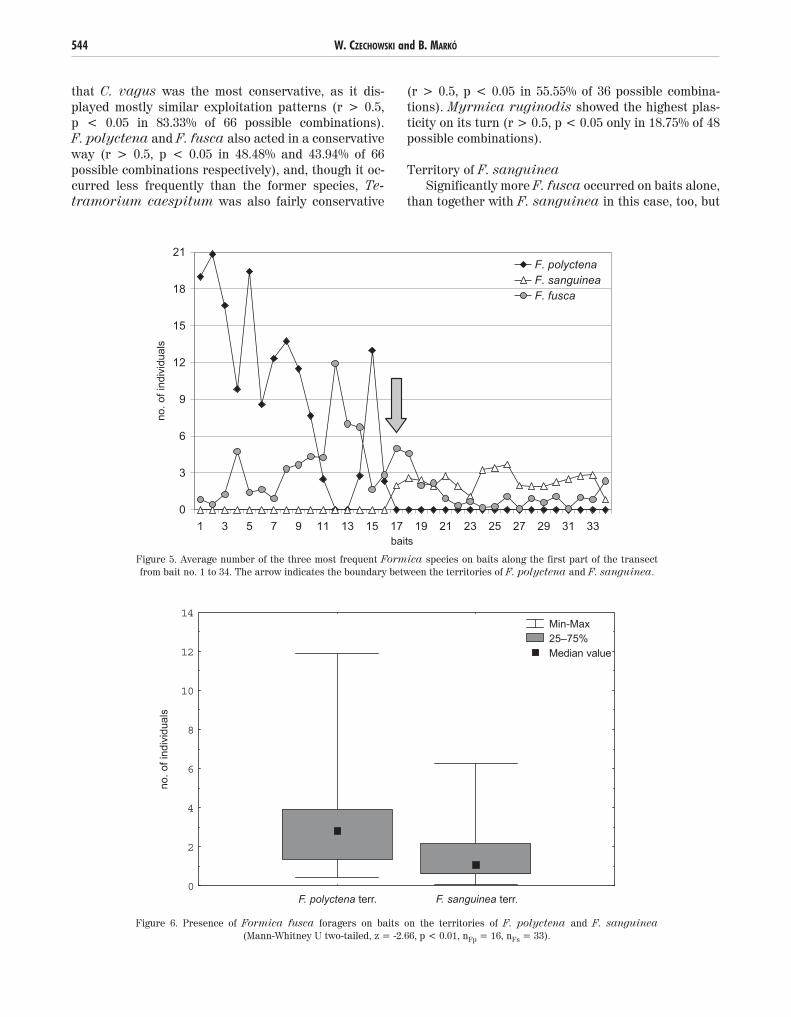
F. fusca was more abundant on baits in the territo-ry of F. polyctena, than in territory of F. sanguinea(Fig. 6). On the other hand F. polyctena was mostlypresent on its territory (Mann-Whitney U two-tailed, z = -5.86, p < 0.001, nFp = 16, nFs = 33), whereas F. sanguinea lacked entirely from the territory of F. polyctena. Camponotus vagus was found only onF. polyctena’s territory, while Myrmica ruginodiswas present mostly on baits on F. sanguinea’s territo-ry (Mann-Whitney U two-tailed, z = -3.14, p < 0.01, nFp = 16, nFs = 33). Tetramorium caespitum did notshow any bias in its presence on the territories of anyof the territorial species, whereas the other two species(Leptothorax acervorum and Lasius sp.) were ob-served too rarely for a consistent statistical analysis.
Within territory differences
Territory of F. polyctenaThe presence of F. polyctena on baits seemed to
have negative effect on F. fusca, as significantly moreF. fusca occurred on baits alone, than together with F. polyctena. On its turn F. polyctena also was sig-nificantly more abundant in the absence of F. fusca(Fig. 7). F. fusca correlated negatively with F. polycte-na when co-occurring (Spearman r = -0.29, p < 0.05, n = 77).
Camponotus vagus mostly avoided F. polyctena(rate of co-occurrence 0.02) on its turn. Tetramoriumcaespitum was also more abundant in the lack of F. polyctena (Mann-Whitney U two-tailed, z = -2.13,na = 17, nt = 11), but F. polyctena was not significant-ly influenced by its presence. Only Myrmica rugi-nodis seemed not to be affected negatively by F. po-lyctena generally, as they usually co-occurred (rate ofco-occurrence 0.88). No negative correlation was foundbetween any of the two latter species and F. polycte-na when co-occurring.
The Spearman r analyses (n = 16 baits) of theexploitation patterns over the 12 replicates revealed
UNCOMFORTABLE PROTECTION: FORMICA POLYCTENA SHELTERS F. FUSCA FROM F. SANGUINEA 543
Table 2. Mean evenness and SD (Shannon-Wiener entropy, max. possible HSh = 3.58) of the presence of different ant species on baitsseparately on the territories of Formica polyctena and F. sanguinea (Fp – F. polyctena, Fs – F. sanguinea, Ff – F. fusca, Cv – Camponotus vagus, Mr – Myrmica ruginodis, Tc – Tetramorium caespitum, La – Leptothorax acervorum, L – Lasius sp.).
Territory Fp Fs Ff Cv Mr Tc La LSpecies
F. sanguinea 0 0.73 0.68 – 0.4 0.29 0 0(0.14) (0.3) (0.24) (0.24)
F. polyctena 0.81 – 0.7 0.73 0.2 0.25 0.18 –(0.11) (0.19) (0.28) (0.21) (0.22) (0.16)
that C. vagus was the most conservative, as it dis-played mostly similar exploitation patterns (r > 0.5, p < 0.05 in 83.33% of 66 possible combinations). F. polyctena and F. fusca also acted in a conservativeway (r > 0.5, p < 0.05 in 48.48% and 43.94% of 66 possible combinations respectively), and, though it oc-curred less frequently than the former species, Te-tramorium caespitum was also fairly conservative
(r > 0.5, p < 0.05 in 55.55% of 36 possible combina-tions). Myrmica ruginodis showed the highest plas-ticity on its turn (r > 0.5, p < 0.05 only in 18.75% of 48possible combinations).
Territory of F. sanguineaSignificantly more F. fusca occurred on baits alone,
than together with F. sanguinea in this case, too, but
544 W. CZECHOWSKI and B. MARKÓ
Figure 6. Presence of Formica fusca foragers on baits on the territories of F. polyctena and F. sanguinea(Mann-Whitney U two-tailed, z = -2.66, p < 0.01, nFp = 16, nFs = 33).
0
2
4
6
8
10
12
14M in-M ax
25% -75%
M edian value
Figure 5. Average number of the three most frequent Formica species on baits along the first part of the transectfrom bait no. 1 to 34. The arrow indicates the boundary between the territories of F. polyctena and F. sanguinea.
baits
F. polyctena terr. F. sanguinea terr.
no. o
f ind
ivid
uals
no. o
f ind
ivid
uals
Min-Max25–75%Median value

no differences were found in the case of F. sanguinea(Fig. 8) with regard to the absence or presence of F. fusca on baits. No negative correlation was foundbetween F. fusca and F. sanguinea when co-occurring.
On the other hand Myrmica ruginodis was notaffected by the presence of F. sanguinea (Mann-Whitney U two-tailed, z = -1.37, n.s., na = 41, nt = 80),however, less F. sanguinea individuals occurred onbaits exploited by M. ruginodis, than in the lack of
this species (Mann-Whitney U two-tailed, z = -2.79, p <0.01, na = 205, nt = 80). There were not found any sig-nificant differences in the amount of F. sanguinea andTetramorium caespitum foragers in each othersabsence or presence (Mann-Whitney U two-tailed, z = -0.45, n.s., na = 246, nt = 39 for F. sanguinea, na = 36, nt = 39 for T. caespitum). No negative corre-lations were found between any of these species and F. sanguinea when co-occurring.
UNCOMFORTABLE PROTECTION: FORMICA POLYCTENA SHELTERS F. FUSCA FROM F. SANGUINEA 545
Figure 7. Presence of Formica fusca and F. polyctena foragers on baits in the absence of each other and together on the territory of F. polyctena (Mann-Whitney U, two-tailed): F. polyctena z = -3.46, p < 0.01
(na = 57, ncooc = 78); F. fusca z = -6.34, p < 0.001 (na = 54, ncooc = 78).
Figure 8. Presence of Formica sanguinea and F. fusca foragers on baits in the absence of each other andtogether on the territory of F. sanguinea (Mann-Whitney U, two-tailed): F. sanguinea z = -1.89, n.s. (na = 107,
ncooc = 178); F. fusca z = -2.72, p < 0.05 (na = 64, ncooc = 178).
0
10
20
30
40
50
60
70
80
90
100
110F. polyctena
F. fusca
0
5
10
15
20
25
30
35
40
45F. sanguinea
F. fusca
alone co-occurrence
alone co-occurrence
no. o
f ind
ivid
uals
no. o
f ind
ivid
uals
F. polyctenaF. fusca
F. sanguineaF. fusca

F. fusca showed an extremely high stability (Spear-man r > 0.34, p < 0.05 in 92.42% of the 66 possible combinations) of the exploitation pattern over the 12replicates. Myrmica ruginodis and Tetramoriumcaespitum were less conservative (r > 0.34, p < 0.05in 36.36% and 28.79% of 66 possible combinationsrespectively). Interestingly the exploitation of F. san-guinea changed to a great extent (r > 0.34, p < 0.05 in21.21% of 66 possible combinations).
DISCUSSION
The success of ant species depends mostly on theirplasticity, which helps them persist under differentant-community circumstances (Wilson and Hölldobler1990, Detrain et al. 1999). This is even more valid fornon-territorial species, which must bear different typesof competitive pressure from dominant species. Ste-baev and Reznikova (1972) and Reznikova (1975, 1982)were the first to study the role of behavioural plasticityin relations between dominating Formica s. str. andsubordinate Serviformica species. Though direct for-aging success (e.g. amount and quality of retrievedfood) could be considered to be the key to survival of a social insect colony, direct success is in fact stronglyinfluenced by other conditions, e.g., predatory and/orcompetitive pressure (Savolainen 1990, 1991, Detrainet al. 1999, Herbers and Banschbach 1999, Korb andLinsenmair 2002, Jarau et al. 2003, Markó and Cze-chowski 2004). In such trade-off situations – biggerfood quantity and better quality alongside considerableexposure to competitive/predatory damage – the‘choice’ offering longer colony life and safer develop-ment can be considered to be the successful strategy.Thus long-term advantages should be ‘preferred’(Creswell et al. 2000). For intraspecific competition inwood ants which leads through mutual predation to a long-term advantage see also de Bruyn and Mabelis(1972, 1979).
What are the survival chances of a non-territorialand potential slave ant species living on neighbouringterritories of F. polyctena and F. sanguinea? Are thedifferences between the competitive and life-historystrategies of both territorial species reflected in thecolonial success of F. fusca? The territory of F. poly-ctena would certainly offer safety from the periodicalrobbing activity of F. sanguinea. Consequently, F. fusca colonies could persist there and grow big-ger. On the other hand F. polyctena severely decreas-es the foraging success and alters the foraging strate-gy of F. fusca (Czechowski 1985, Savolainen and Vepsäläinen 1989, Savolainen 1990, 1991, Czechowskiand Markó 2005), and occasionally even destroyswrong-placed colonies. In addition, its dependent colo-ny-foundation causes F. fusca colonies to die out,
though at a slower rate than F. sanguinea. All thesefactors hinder the development of big F. fusca coloniesas proved by Savolainen (1990, 1991) and noticed alsoby Mabelis (1984). F. sanguinea, in turn, is a lousiercompetitor than many territorial species (Czechowski1999, 2000), which means that it exerts less pressureon the foraging success of F. fusca. Nevertheless itsraids cause severe damage to slave ant colonies. Thusthe question remains whether relatively successful for-aging activity (F. sanguinea version) or relativelypeaceful colony development (F. polyctena version) re-sults in greater survival chances (greater forager num-bers) for F. fusca.
On the basis of the results of the present study, itseems that F. polyctena territories offer more oppor-tunity for F. fusca to develop as the relative abundanceof the latter turned out to be more than twice higherwithin the territory of F. polyctena than within the ter-ritories of F. sanguinea, which could be a fair esti-mate of F. fusca colony size and number. Of course, itshould be remembered that the abundance of ants atbaits represents only an approximation of actual popu-lation size (see Data Analysis and also Czechowski andVepsäläinen 2001). The factors determining such spec-tacular quantitative differentiation within the localpopulation of F. fusca, i.e. the slavery pressure of F. sanguinea on the one hand and the indirect protec-tion by F. polyctena against this pressure on the oth-er, were probably very powerful as at the same timethere were two side factors operating with a potentialto reduce the differences. They were (1) the presenceof a colony of the obligatory slave-making ant Polyer-gus rufescens in the neighbourhood of the F. polycte-na territory, and (2) the possible presence of F. fuscaworkers enslaved by F. sanguinea. P. rufescens veryeffectively ravages colonies of F. fusca (see e.g. Cze-chowski 2005a, 2006). A marginal part of the F. po-lyctena territory under study encompassing the first4–5 baits lay within the range of P. rufescens raids(see Fig. 4). One successful raid during the study sea-son and a few raids in the following year were observedin that area, with no counter-action of F. polyctena(W. Czechowski, unpubl. data; see also Czechowski2002). It means that the F. fusca population within theterritory of F. polyctena was at least occasionally sub-jected to slavery pressure from P. rufescens. On theother hand, the estimated size of the F. fusca popula-tion on the F. sanguinea side could be an overesti-mate due to the presence of F. fusca slaves from F.sanguinea colonies. Ecologically, such enslaved indi-viduals, being F. sanguinea nestmates, representcolonies of their social parasite rather than those oftheir own species. What is more, because of the protec-tive role of F. polyctena, one can expect that a consid-erable part of the slaves originated from outside thestudy area. Thus presumably the difference between
546 W. CZECHOWSKI and B. MARKÓ

the number of non-slave F. fusca foragers on the terri-tory of F. polyctena and F. sanguinea was evengreater in reality than that suggested by these results.
Savolainen (1990, 1991) presents clear evidencethat F. polyctena negatively influences F. fusca’s for-aging success by reducing the size, quality andretrieval rate of F. fusca’s prey items. Czechowski andMarkó (2005) also present indirect evidence for theinhibition of foraging activity of F. fusca by F. rufa.The negative effect of wood ant species exerted on theforaging success and strategy of Myrmica Latr. spe-cies also emphasizes their strong competitive pressure(Vepsäläinen and Savolainen 1990). The results of bait-ing experiments support the occurrence of a significantreduction of F. fusca’s foraging activity in the presenceof F. polyctena. F. sanguinea, in turn, seemed not toinfluence the foraging activity of F. fusca so dramati-cally, though in this case the problem of a lack of differ-ences between slave and non-slave individuals still hin-dered more reliable conclusions regarding F. san-guinea’s policy towards non-slave F. fusca. The lackof differences in the number of other ant species pres-ent in baits on the territory of F. sanguinea in its pres-ence or absence, and the respective significant differ-ences found on the territory of F. polyctena (Cam-ponotus vagus, Tetramorium caespitum) alsoemphasize that F. sanguinea could indeed be a lesseffective competitor than its territorial rival, being thusa more preferable ruler for non-territorial ants. Conse-quently, M. ruginodis appeared to prefer F. san-guinea’s territory. In fact Myrmica ruginodis, whenpresent in baits in large numbers, may have had a neg-ative effect on F. sanguinea, which is consistent withthe plasticity of M. ruginodis’ competitive behaviour(Markó and Kiss, in prep.), a phenomenon also knownin the related M. rubra L. (Czechowski 1985), and inother species such as Lasius psammophilus Seifert(Markó and Czechowski 2004).
In response to competitive pressure, F. fusca andCamponotus vagus exhibited relatively high spatialfidelity for already successfully exploited baits. F. po-lyctena showed also a certain degree of predictabi-lity in this respect, whereas F. sanguinea’s foragingstrategy was characterized by low spatial fidelity, orrather randomness. As C. vagus is known to protect itsfood sources, and to be an aggressive species (Cze-chowski 2005b), it is not surprising that almost no co-occurrence with F. polyctena was observed within theterritory of the latter. Presumably C. vagus can existand forage successfully only in gaps (spots with lowcompetitive pressure) on the territory of wood ants.
Although the data indicate that location within anarea dominated by the lousier competitor, F. sangu-inea, would be more favourable for the foraging successof a non-territorial ant than living on F. polyctenaterritory, the slave-making pressure of F. sanguinea
drives F. fusca to succeed in developing bigger (andconsequently more stable) colonies on F. polyctena’sterritory despite the higher local and momentary pressure. The situation is clearly ‘uncomfortable’ for F. fusca but it still offers long-term advantages, whichmake its survival possible. In this respect it would beinteresting to compare colony persistence and movingrates in F. fusca on both territories.
ACKNOWLEDGEMENTS
We are grateful to the referees, Joanna Pętal forgeneral comments, Bram Mabelis for his precise cor-rections and supplements of the manuscript, and thethird referee for supplement of the references, as wellas methodological advices bossible to be taken in thenear future. The field study was supported by theMuseum and Institute of Zoology PAS, Warsaw.
REFERENCES
Bruyn, G. J. de and A. A. Mabelis. 1972. Predation and aggres-sion as possible regulatory mechanisms in Formica.Ekologia Polska, 20: 93–101.
Buschinger, A. 1986. Evolution of social parasitism in ants.Trends in Ecology and Evolution, 1: 155–160.
Buschinger, A. 1990. Sympatric speciation and radiative evo-lution of socially parasitic ants – heretic hypotheses andtheir factual background. Zeitschrift für Zoologische Sys-tematik und Evolutionsforschung, 28: 241–260.
Buschinger, A., Ehrhardt, W. and U. Winter. 1980. The organ-ization of slave raids in dulotic ants. A comparative study(Hymenoptera; Formicidae). Zeitschrift für Tierpsycholo-gie, 53: 245–264.
Cresswell, J. E., Osborne, J. L. and D. Goulson. 2000. An eco-nomic model of the limits to foraging range in centralplace foragers with numerical solutions for bumblebees.Ecological Entomology, 25: 249–255.
Czechowski, W. 1985. Competition between Myrmica laevi-nodis Nyl. and Lasius niger (L.) (Hymenoptera, Formi-coidea). Annales Zoologici, 39: 153–173.
Czechowski, W. 1999. Lasius fuliginosus (Latr.) on a sandydune – its living conditions and interference during raidsof Formica sanguinea Latr. (Hymenoptera, Formicidae).Annales Zoologici, 49: 117–123.
Czechowski, W. 2000. Interference of territorial ant species in the course of raids of Formica sanguinea Latr. (Hy-menoptera, Formicidae). Annales Zoologici, 50: 35–38.
Czechowski, W. 2002. Polyergus rufescens (Latr.), an oblig-atory slave-keeper, in the Białowieża Forest. Parki Naro-dowe i Rezerwaty Przyrody, 21: 29–32. (In Polish).
Czechowski, W. 2005a. Integration raids in the Amazon antPolyergus rufescens (Hymenoptera, Formicidae). In-sectes Sociaux, 52: 103–104.
Czechowski, W. 2005b. Nest competition between Campo-notus vagus (Scopoli, 1763) and Camponotus hercu-leanus (Linnaeus, 1758) (Hymenoptera: Formicidae) in
UNCOMFORTABLE PROTECTION: FORMICA POLYCTENA SHELTERS F. FUSCA FROM F. SANGUINEA 547

the Białowieża Forest (Poland). Myrmecologische Nach-richten, 7: 43–45.
Czechowski, W. 2006. The route of Formica polyctena Först.as a factor promoting emancipation of Formica fusca L.slaves from colonies of Polyergus rufescens (Latr.) (Hy-menoptera: Formicidae). Polish Journal of Ecology, 54:159–162.
Czechowski, W. In press. Do Polyergus rufescens (Latr.),and Formica sanguinea (Latr.) (Hymenoptera: Formici-dae) share resources of slave species with each other?Polish Journal of Ecology, 55.
Czechowski, W. and B. Markó. 2005. Competition betweenFormica cinerea Mayr (Hymenoptera: Formicidae) andco-occurring ant species, with special reference to Formi-ca rufa L.: direct and indirect interferences. Polish Jour-nal of Ecology, 53: 467–487.
Czechowski, W. and B. Pisarski. 1994. How do wood antqueens (Hymenoptera, Formicidae) score their reproduc-tive success? Przegląd Zoologiczny, 38: 79–85. (In Polish).
Czechowski, W. and K. Vepsäläinen. 2001. Formica rufa pro-tects indirectly F. fusca against raids of F. sanguinea.Annales Zoologici, 51: 267–273.
Detrain, C., Deneubourg, J. L. and J. M. Pasteels. 1999. Deci-sion-making in foraging by social insects. In: C. Detrain(ed.). Information Processing in Social Insects. BirkhäuserVerlag, Basel, p. 331–354.
Gallé, L. J. 1975. Factors stabilizing the ant populations (Hy-menoptera, Formicidae) in the grass associations of theTisza Basin. Tisza, 10: 61–66.
Gallé, L. J. 1978. Dispersion of the nests of ant species (Hyme-noptera, Formicidae). Acta Biologica, Szeged, 24: 105–109.
Herbers, J. M. and V. S. Banschbach. 1999. Plasticity of socialorganization in a forest ant species. Behavioral Ecologyand Sociobiology, 45: 451–465.
Hölldobler, B. and E. O. Wilson. 1990. The Ants. Harvard Uni-versity Press, Cambridge, MA, 732 pp.
Jarau, S., Hrncir, M., Schmidt, V. M., Zucchi, R. and F. G.Barth. 2003. Effectiveness of recruitment behavior in stin-gless bees (Apidae, Meliponini). Insectes Sociaux, 50:365–374.
Korb, J. and K. E. Linsenmair. 2002. Evaluation of predationrisk in the collectively foraging termite Macrotermes bel-licosus. Insectes Sociaux, 49: 264–269.
Kutter, H. 1968. Liste sozialparasitischer Ameisen. Archives.Institut Grand-Ducal de Luxembourg, 33: 201–210.
Mabelis, A. A. 1984. Interference between wood ants and oth-er ant species (Hymenoptera, Formicidae). NetherlandsJournal of Zoology, 34: 1–20.
Mabelis, A. A. 1979. Wood ant wars. The relationship betweenaggression and predation in the red wood ant (Formicapolyctena Först.). Netherlands Journal of Zoology, 29:451–620.
Markó, B. and W. Czechowski. 2004. Lasius psammophilusSeifert and Formica cinerea Mayr (Hymenoptera: Formi-cidae) on sand dunes: conflict and coexistence. AnnalesZoologici, 54: 365–378.
Pisarski, B. 1973. Social structure of Formica (Coptofor-
mica) exsecta Nyl. (Hymenoptera: Formicidae) and itsinfluence on morphology, ecology and ethology of the species. Zoological Institute PAS, Warsaw, 134 pp. (In Pol-ish with French summary).
Pisarski, B. 1982 Territoires et territorialisme de Formica(Coptoformica) exsecta Nyl. Memorabilia Zoologica, 38:163–203.
Pisarski, B. and W. Czechowski. 1994. Ways to reproductivesuccess of wood ant queens. Memorabilia Zoologica, 48:181–186.
Pisarski, B. and K. Vepsäläinen. 1989. Competitive hierarchyin ant communities (Hymenoptera, Formicidae). AnnalesZoologici, 42: 321–329.
Punttila, P., Haila, Y., Pajunen, T., and H. Tukia 1991. Coloni-sation of clearcut forests by ants in the southern Finnishtaiga: a quantitative survey. Oikos, 61: 250–262.
Punttila, P., Haila, Y., and H. Tukia. 1996. Ant communities intaiga clearcuts: habitat effects and species interactions.Ecography, 19: 16–28.
Reznikova, J. [Zh.] 1982. Interspecific communication amongants. Behaviour, 80: 84–95.
Reznikova, Zh. 1999. Ethological mechanisms of populationdensity control in coadaptive complexes of ants. RussianJournal of Ecology, 30: 187–192.
Reznikova, Zh. I. 1975. Non-antagonistic relations betweenant species occupying similar ecological niches. Zoo-logicheskij Zhurnal, 54: 1020–1031. (In Russian with Eng-lish summary).
Reznikova, Zh. I. 1980. Interspecific hierarchy in steppe ants.Zoologicheskij Zhurnal, 59: 1168–1176. (In Russian withEnglish summary).
Reznikova, Zh. I. 1983. Interspecific relations in ants. Nauka,Novosibirsk, 206 pp. (In Russian).
Savolainen, R. 1990. Colony success of the submissive antFormica fusca within territories of the dominant Formi-ca polyctena. Ecological Entomology, 15: 79–85.
Savolainen, R. 1991. Interference by wood ant influences sizeselection and retrieval rate of prey by Formica fusca.Behavioral Ecology and Sociobiology, 28: 1–7.
Savolainen, R., and K. Vepsäläinen. 1988. A competition hier-archy among boreal ants: impact on resource partitioningand community structure. Oikos, 51: 135–155.
Savolainen, R. and K. Vepsäläinen. 1989. Niche differentiationof ant species within territories of the wood ant Formicapolyctena. Oikos, 56: 3–16.
Savolainen, R., Vepsäläinen, K. and H. Wuorenrinne. 1989.Ant assemblages in the taiga biome: testing the role of ter-ritorial wood ants. Oekologia, 81: 481–486.
Stebaev, I. V. and J. [Zh.] Reznikova. 1972. Two interactiontypes of ants living in steppe ecosystems in South Siberia.Ekologia Polska, 20: 103–109.
Vepsäläinen, K. and B. Pisarski. 1982. Assembly of island ant communities. Annales Zoologici Fennici, 19: 327–335.
Vepsäläinen, K. and R. Savolainen. 1990. The effect of inter-ference by Formicine ants on the foraging of Myrmica.Journal of Animal Ecology, 59: 643–654.
548 W. CZECHOWSKI and B. MARKÓ
Received: May 5, 2006Accepted: July 15, 2006


















