
Uccelli Antonio - Clinical Applications of Cellular ... · Antonio Uccelli Neuroimmunology Unit...
43
Antonio Uccelli Neuroimmunology Unit Dept. Of Neurosciences University of Genoa, Italy Workshop on Transplant and Cellular Therapy for Autoimmune Diseases CIBMTR, April 19-20 2013, Milwaukee, WI
-
Upload
vuongthuan -
Category
Documents
-
view
215 -
download
0
Transcript of Uccelli Antonio - Clinical Applications of Cellular ... · Antonio Uccelli Neuroimmunology Unit...

Antonio UccelliNeuroimmunology UnitDept. Of NeurosciencesUniversity of Genoa, Italy
Workshop on Transplant and Cellular Therapy for Autoimmune
DiseasesCIBMTR, April 19-20 2013,
Milwaukee, WI

HSC InfusionChemotherapy
MSC
Induction of immune tolerance
Neuroprotection
Induction of oligodendrogenesis
Induction of neurogenesis
Transdifferentiation and neuroprotection
Immunomodulation
NPC
HSC
HSC Mobilization
Eradication of autoimmunity
Reset of the immune system
Immune reconstitution
Uccelli and Mancardi, Curr Opin Neurol 2010
Stem Cells Portfolio for the treatment of MS

The hematopoietic stem cell niche is the site where HSCs home in the BM in close contact with osteoblasts, endothelial and stromal cells, a rare cell population of non-heamatopoietic origin(representing < 0.01% of total BM cells) named mesenchymal stem cells (MSC)
Uccelli A, et al, Nat Rev Immunol 2008

Why should we use MSC for the treatment of brain repair?

Neural differentiation: prolonged (30 days) and specific
nestin
MAP2 GABA-A
GFAP
In vitro neuronal (trans)differentiation of MSC:is this biologically relevant in vivo?
Anghileri et al, 2008

Mesenchymal stem cells protect CNS neurons against glutamate excitotoxicity
Voulgari-Kokota et al Exp Neurol 2012

Mesenchymal stem cells instruct oligodendrogenic fate decision on adult neural stem cells.
Rivera et al., Stem Cells, 2006
Control MSC-CM
GalC
GFAP MBP

MSC promote in vivo neurogenesis
• MSCs promote proliferation, migration, and survival of mouse NSCs• Implantation of human MSCs increased synthesis of trophic factors
Munoz et al, PNAS 2006

MSC inhibit T cell proliferation arresting cell division
Benvenuto et al, Stem Cells 2007

MSC inhibit B cell functions
Corcione et al, Blood 2005
B cells B cells +MSC 1:1
B cells B cells +MSC 1:1
B cells B cells +MSC 1:1
***
cpm
0
20000
40000
60000
80000 B cells
B cells + MSC 1:1
B cells + MSC 1:2
MSC
IgM IgG IgA
ng/m
l
0
50
100
150
200
250
300
350
IgM IgG IgA
B cells
B cells + MSC 1:1 **
***
MSC inhibit B cell proliferation
MSC inhibit immunoglobulins production
MSC inhibit B cells differentiation to antibody secreting cells
In collaboration with V. Pistoia

CD11c CD86
A
100 101 102 103 104FL1-H
100 101 102 103 104FL2-H
100 101 102 103 104FL1-H
100 101 102 103 104FL2-H
100 101 102 103 104FL3-H
0
50
0
50
0
50
0
50
CD80 CD40
MHC cl.I MHC cl.II
CCR7 CD49d 1
78% vs 51% 72% vs 59 %
41% vs 20%
68% vs 32% 89% vs 75%
45% vs 16%
55% vs 22% 48% vs 19%
Control DC
MSC treated DC
MSC impair LPS-induced maturation of DC
Chiesa et al, PNAS 2011

A DC + CD4+ TMSC exposed DC +
CD4+ T
Cel
l num
ber
500
0
500
0
50
0
CFSE
CD
69
CFSE
DC + CD8+ TMSC exposed DC
+ CD8+ T
48h
B
CFSE
69885 16035
67755 57210
4500 2610C
ell n
umbe
r
48h
72h
DC + CD8+ TMSC exposed DC
+ CD8+ T400
400
0
0
MSC impair DC antigen presentation to DO11.10 CD4+ T cells and cross-presentation to OT.1 CD8+ T cells
20800 2226
78110 2428
2MOVA
MOVA
0.2MOVA
Chiesa et al, PNAS 2011

The complex interplay between MSCs and the immune system
Uccelli et al, Nat Rev Immunol 2008

Do all these results translate into relevant in vivo biological effects
in experimental models of neurological diseases?

0
1000
2000
3000
4000
5000
6000
7000
8000
T ce
lls p
rolif
erat
ion
(CPM
)
Mog 1 M Mog 3 M Mog 10 M
Inhibition of Ag-specific T and B cell response
MSCs ameliorate EAE inducing in vivo tolerance to myelin antigens inside secondary lymphoid organs (Zappia et al 2005; Gerdoni et al 2007)
Zappia et al, Blood 2005
50 m
T cell
MSC
A B
Demyelination
T cells
Macrophages
B cells
Axonal loss
EO4 A
DC
B
GFAP NeuN
GFP
Neuroprotection without transdifferentiation
Inhibition of T cell encephalitogenic potential
Amelioration of EAE
Lymph nodes engraftment

Is MSCs therapeutic effect based also on long term engraftment
and cell replacement?

MSC are effective across MHC constraints“are they immunoprivileged?”
• MSC T cell inhibition overcome inter and intraspecies barriers
• Syngeneic, allogeneic and xenogeneic MSC can inhibit EAE
Zappia and Casazza, unpublished resultsRafei et al, Mol Ther 2010

Allogeneic marrow stromal cells are immune rejected by MHC class I– and class II–mismatched recipient mice
Eliopoulos et al, Blood 2005

MSCs are killed by cells of the innate immunity
Spaggiari et al, Blood 2006
Prigione et al, Stem Cells 2009
NK cells
γδ cells

Bone Marrow-Derived Mesenchymal Stem Cells after Infusion principally distribute to the lungs
Gao et al, 2001 Barbash et al, 2003 Lee et al, 2009

0
0.5
1
1.5
2
2.5
3
1° 3° 5° 7° 9° 11° 13° 15° 17° 19° 21° 23° 25° 27° 29° 31° 33° 35° 37° 39° 41° 43° 45°
ctrl iv
ctrl ic
msc iv
msc ic
*P < 0.05: **P < 0.01
Identification of the optimal route of administration
ctrl iv msc iv ctrl ic msc ic0
10
20
30
40
50
60
Dem
yelin
atio
n ar
eas
(%)
* *
10X
MSC I.C.MSC I.V.
10X10X
10X
CTRL I:V: CTRL I.C.
Morando et al., Stem Cell Res & Ther 2012

MAP2 MSC 40x
MAP2 CTRL 40x
Demyelination
T cells
Macrophages
B cells
Axonal loss
MSCs do not transdifferentiate but
protect neurons in vivo
EO4 A
DC
B
GFAP
NeuN
GFP
Gerdoni et al, Ann Neurol 2007

Conditioned growth medium from human
MSCs biases the development of
neurosphere derived cells toward
oligodendrocytes and neurons, and
promotes functional recovery in EAE
Bai et al Nat Neurosci 2012
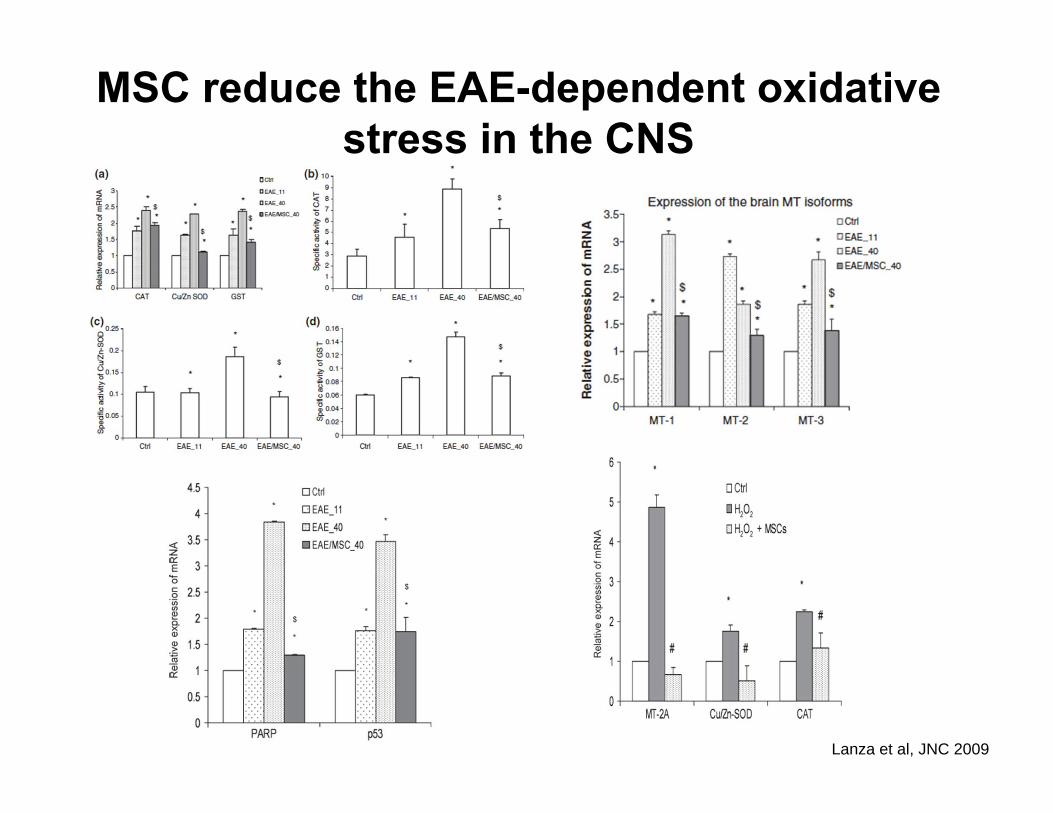
MSC reduce the EAE-dependent oxidative stress in the CNS
Lanza et al, JNC 2009

GFAP
IB4
MSCs reduces astrogliosis and microglia activation in EAE
SPINAL CORD CTR 40X SPINAL CORD MSC 40X
Casazza and Principato unpublished results

Perry, V. H. et al. (2010) Nat. Rev. Neurol.
Microglia dynamically changes phenotype and functions depending on environmental cues

Microglial plays a key role in CNS homeostasis
Neumann H et al. Brain 2009;132:288-295
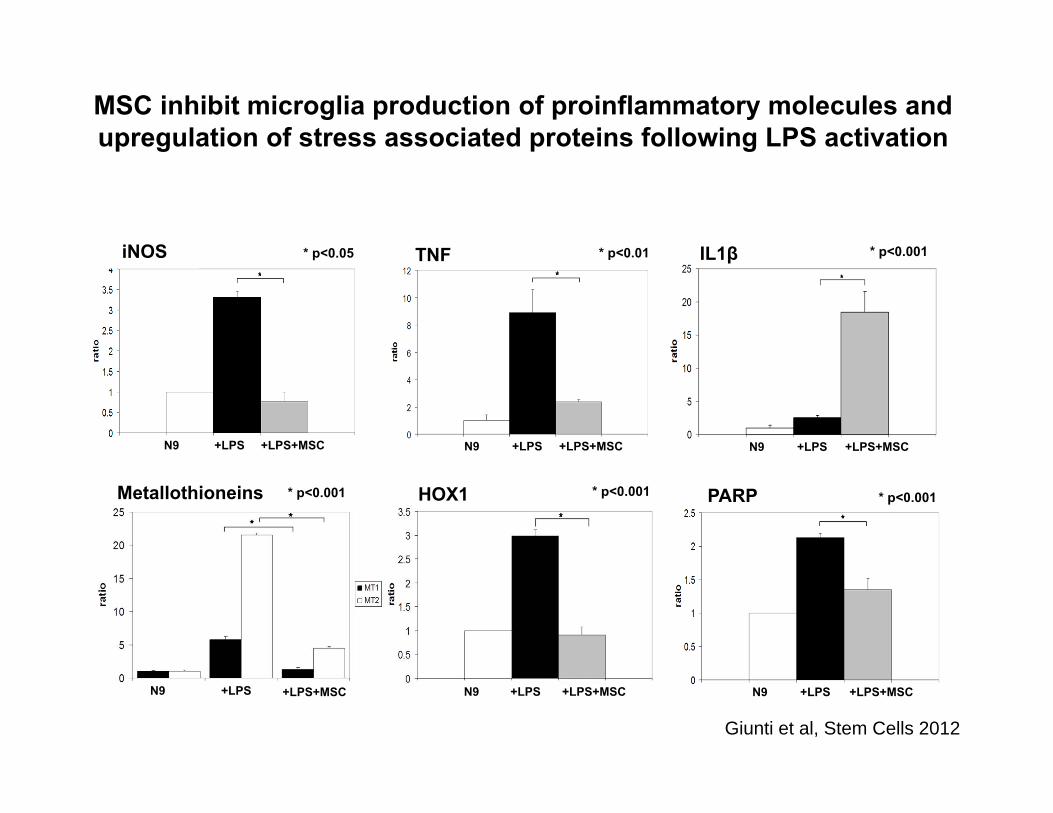
iNOS
HOX1
TNF
PARP
N9 +LPS +LPS+MSCN9 +LPS +LPS+MSC
N9 +LPS +LPS+MSC
IL1β
N9 +LPS +LPS+MSC
Metallothioneins * p<0.001
N9 +LPS +LPS+MSC
N9 +LPS +LPS+MSC
* p<0.05 * p<0.01 * p<0.001
* p<0.001* p<0.001
MSC inhibit microglia production of proinflammatory molecules and upregulation of stress associated proteins following LPS activation
Giunti et al, Stem Cells 2012

MSC reverse and enhance the LPS-mediated decrease of molecules associated with neuroprotection
IGF1
CX3CR1 CD200RNURR1
N9 +LPS +LPS+MSC
* p<0.05
N9 +LPS +LPS+MSC
* p<0.05
N9 +LPS +LPS+MSCN9 +LPS +LPS+MSC
* p<0.05
EP2 * p<0.05
N9 +LPS +LPS+MSC
* p<0.05
Giunti et al, Stem Cells 2012

MSC induce functional changes on microglia
N9 +LPS +LPS+MSC
Intracellular Ca2+ * p<0.001
N9 +LPS +LPS+MSC
Phagocytosis * p<0.001 TREM2 * p<0.001
N9 +LPS +LPS+MSC
N9 +LPS +LPS+MSC
Giunti et al, Stem Cells 2012

The disruption of CX3CL1-CX3CR1 signaling induces neurotoxicity in mouse models of neurodegeneration (Cardona et al, Nat Neurosci, 2006)
CX3CL1 expressed by neurons modulates the activity of CX3CR1-positive microglia

MSC express and produce fractalkine under inflammatory conditions
MSC +IFN +IFN+LPS
+LPS MSC +IFN +IFN+LPS
+LPS
CX3CL1 production * p<0,05 * p<0,01CX3CL1 expression
Giunti et al, Stem Cells 2012

CX3CL1 silencing reverts MSC effect on the microglia expression of inflammatory and neuroprotective genes
IL1β
* p<0.01** p<0.01
* p<0.05** p<0.05NURR1CX3CR1
* p<0.01** p<0.01
* p<0.001** p<0.05TNF
N9 +LPS +LPS+MSC
+LPS+MSCCX3CL1si
N9 +LPS +LPS+MSC
+LPS+MSCCX3CL1si
N9 +LPS +LPS+MSC
+LPS+MSCCX3CL1si
N9 +LPS +LPS+MSC
+LPS+MSCCX3CL1si
N9 +LPS +LPS+MSC
+LPS+MSCCX3CL1si
EP2 * p<0.01** p<0.01
Giunti et al, Stem Cells 2012

Silencing of fractalkine in reverts functional changes exerted by MSCs on microglia
N9 +LPS +LPS+MSC +LPS+MSC+ aCX3CL1
Phagocytosis * p<0.001** p<0.001
N9 +LPS +LPS+MSC
+LPS+MSC+aCX3CL1
Intracellular Ca2+ * p<0.001** p<0.001
N9 +LPS +LPS+MSC
+LPS+MSC+aCX3CL1
TREM2 * p<0.001** p<0.001
N9 +LPS +LPS+MSC
+LPS+MSCsiCX3CL1
Giunti et al, Stem Cells 2012

CONCLUSIONS
• MSC – microglia interactions lead to the inhibition of the production of proinflammatory molecules and enhances molecules associated to neuroprotection.
• MSC promote microglia’s functions involved in CNS homeostasis• CX3CL1 produced by MSC plays a major role in the induction of a
functional switch of microglia from a detrimental one to a beneficial one further pointing out a major role for molecules released by MSC upon interaction with the inflammatory environment.

Uccelli et al, Mol Med 2012
MSCs improve survival and motor functions in SOD1/G93A(+) mice through inhibition of glutamate driven excitotoxicity

Mesenchymal stem cells improves experimental epilepsy rescuing hippocampal neurons from apoptosis
Voulgari-Kokota et al, Exp Neurol 2012

Systemic administration of MSC increases neuronal survival in global cerebral ischemia
Perasso et al 2010

T
B
+
+
-
-
-
dem
yelin
ated
axo
n
TNF-
IFN-
NO
MSC inhibit the autoimmune attack to the CNS and promote neural cell survival
Adapted from Uccelli et al Trends Immunol 2007
NeuronsNeural precursor cells
Oligodendrocytes
Astrocytes
MicrogliaROI
++
Neuron

• Transdifferentiation potential ✗• Engraftment/cell contact mechanisms ✗• Direct inhibition of the effector functions of cells of the
immune system ✔• By-stander effect through the release of:
– Anti-apoptotic and anti-oxidant molecules ✔– Trophic factors ✔– Factors inducing the mobilization and induction of
proliferation of local precursors (committed) cells ✔• Non-cell autonomous mechanisms mediated by:
– Dendritic cells – Monocytes/macrophages– microglia
MoA of MSC for brain repair: take home messeage

"Stemness" Does Not Explain the Repair of Many Tissues by
Mesenchymal Stem/Multipotent Stromal Cells (MSCs)
There has recently been an explosion of interest in adult stem/progenitor cells that have the potential to repair
tissues, with over 3,000 citations to publications (PubMed) and numerous announcements of clinical trials in which the cells are used to treat individuals
with a broad range of diseases. At the same time, the data present “a paradox” as the cells originally
attracting attention because of their stem-cell like properties, frequently repair injured tissues without
much evidence of either engraftment or differentiation.Darwin Prockop 2007

The studies on MSC have been supported by grants of the Italian Foundation for Multiple Sclerosis (FISM), Istituto Superiore di Sanità (ISS) “National Program on Stem Cells”, by the Ministero della Salute, Ricerca Finalizzata, by the Ministero dell’Università e della Ricerca, Progetti PRIN, by the
Fondazione CARIGE, by the Fondazione CARIPLO and the Regione LIGURIA
• Neuroimmunology Unit – University of Genoa– F. Benvenuto – S. Casazza– D. Giunti– L. Lovato– F. Ivaldi– S. Morando– B. Parodi– L. Serpero– T. Vigo– A. Uccelli
• Dept. Of Neurosciences,- University of Genoa– GL. Mancardi
• ISPE San Raffaele Hospital• G. Martino
• Department of Biology University of Genoa• L. Vergani
• Dept. Of Nuclear Medicine University of Genoa– G. Sambuceti
Institute of Anatomy University of BariD. Virgintino
Dept of biomedical sciences - University of TeramoM. Maccarrone
• Hematology – S. Martino Hospital, Genoa– F. Gualandi
• University of California San Francisco– J. Oksenberg– S. Baranzini
• Gaslini Insitute, Genoa– F. Frassoni– S. Chiesa– G. Candiano – E. Traggiai– L. Moretta– V. Pistoia
• Dept. of Biochemistry – University of Genoa– A. De Flora
• Dept. Neuropharmacology, University Genoa– GB. Bonanno
– CNR Genoa– Cesare Usai
– Tor Vergata University – Rome– D. Centonze– A. Urbani
– University of Verona – Italy– B. Bonetti
– Ospedali Riuniti Bergamo – Italy– M. Introna

www.ici2013.org


















