
Ttez.sdu.edu.tr/Tezler/TF02403.pdf · In Denizli roosters collagen fibers are under the lateral...
55
T.C SÜLEYMAN DEMĠREL ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ DENĠZLĠ HOROZU VE TAVUĞU (Gallus gallus domesticus) ĠLE SÜLÜN (Phasianus colchicus) SĠRĠNKSLERĠNDE BAĞ DOKUSU FĠBRĠLLERĠNĠN DAĞILIMI ÖZNUR ÖNAL DanıĢman: Yrd. Doç. Dr. Kenan ÇINAR YÜKSEK LĠSANS TEZĠ BĠYOLOJĠ ANABĠLĠM DALI ISPARTA - 2013
Transcript of Ttez.sdu.edu.tr/Tezler/TF02403.pdf · In Denizli roosters collagen fibers are under the lateral...

T.C
SÜLEYMAN DEMĠREL ÜNĠVERSĠTESĠ
FEN BĠLĠMLERĠ ENSTĠTÜSÜ
DENĠZLĠ HOROZU VE TAVUĞU (Gallus gallus domesticus) ĠLE
SÜLÜN (Phasianus colchicus) SĠRĠNKSLERĠNDE BAĞ DOKUSU
FĠBRĠLLERĠNĠN DAĞILIMI
ÖZNUR ÖNAL
DanıĢman: Yrd. Doç. Dr. Kenan ÇINAR
YÜKSEK LĠSANS TEZĠ
BĠYOLOJĠ ANABĠLĠM DALI
ISPARTA - 2013

© 2013 [Öznur ÖNAL]



i
ĠÇĠNDEKĠLER
Sayfa
İÇİNDEKİLER .......................................................................................................... i
ÖZET ....................................................................................................................... ii ABSTRACT ............................................................................................................ iii
TEŞEKKÜR .............................................................................................................iv ŞEKİLLER DİZİNİ ................................................................................................... v
1. GİRİŞ .................................................................................................................... 1 2. KAYNAK ÖZETLERİ .......................................................................................... 7
3. MATERYAL ve YÖNTEM ................................................................................. 13 4.ARAŞTIRMA BULGULARI................................................................................ 14
4.1. Histolojik Bulgular ........................................................................................ 14 4.1.1. Denizli Horozu ....................................................................................... 14
4.1. 2. Denizli tavuğu ....................................................................................... 15 4.1.3. Erkek sülün ............................................................................................ 16
4.1.4. Dişi sülün ............................................................................................... 16 4.2. Histokimyasal Bulgular ................................................................................. 16
4.2.1. Denizli Horozu ....................................................................................... 18 4.2.2. Denizli tavuğu ........................................................................................ 23
4.2.3. Erkek sülün ............................................................................................ 27 4.2.4. Dişi sülün ............................................................................................... 31
5. TARTIŞMA VE SONUÇ ..................................................................................... 36 6. KAYNAKLAR .................................................................................................... 41
ÖZGEÇMİŞ............................................................................................................. 45

ii
ÖZET
Yüksek Lisans Tezi
DENĠZLĠ HOROZU VE TAVUĞU (Gallus gallus domesticus) ĠLE SÜLÜN
(Phasianus colchicus) SĠRĠNKSLERĠNDE BAĞ DOKUSU FĠBRĠLLERĠNĠN
DAĞILIMI
Öznur ÖNAL
Süleyman Demirel Üniversitesi
Fen Bilimleri Enstitüsü
Biyoloji Anabilim Dalı
DanıĢman: Yrd. Doç. Dr. Kenan ÇINAR
Bu tez çalımasında Denizli horozu ve tavuğu (Gallus gallus domesticus) ile sülün
(Phasianus colchicus) sirinkslerinde bağ dokusu fibrillerinin dağılımı ve
düzenlenmelerinin belirlenmesi amaçlandı. Bu amaçla 4’er adet erişkin Denizli
horozu ve tavuğu ile dişi ve erkek sülünlerden alınan sirinksler materyal olarak
kullanıldı. Denizli horozu ve tavuğu ile dişi sülünlerde sirinksin yapısına son 1
trakeyal siringeyal halka ile ilk 5 bronşiyal siringeyal kıkırdak halka katılırken; erkek
sülünlerde son 1 trakeyal ve ilk 3 bronşiyal siringeyal kıkırdağın katıldığı dolayısıyla
çalışılan hayvan türlerinde sirinkslerin trakeyobronşiyal tipte olduğu tespit edildi.
LTM’nin son trakeyal siringeyal halka ile birinci bronşiyal siringeyal kıkırdak
halkalar arasında, mediyal timpanik membranın pessulus ile ligamentum
interbronchiale arasında yer aldığı tespit edildi.
Denizli horozunda kollagen fibrillerin lateral timpanik membran epiteli altından bağ
dokusu derinlerine doğru geniş bir alanda kalın demetler halinde olduğı; özellikle
bağ dokusunun derin kısımlarında çok daha sıkı biçimde yerleşim gösterdikleri
saptandı.
MTM’a ait epitel altında kollagen fibrillere nazaran daha geniş bir alanda kalın
fibriller şeklinde yerleşim gösteren elastik fibrillerin Denizli tavuğunda bağ doku
derinlerine doğru daha yoğun biçimde düzenlendikleri gözlendi.
Erkek sülünlerde LTM epitelinin altında, bronşiyal kıkırdak halkalar arasında ve
pessulus lateralindeki epitel altında kollagen fibrillerin sıkı biçimde bir araya gelerek
ince demetler oluşturduğu gözlendi.
Anahtar Kelimeler: Bağ dokusu, Fibril, Denizli horozu, Sülün, Sirinks.
2013, 45 sayfa

iii
ABSTRACT
M.Sc. Thesis
DISTRIBUTION OF CONNECTIVE TISSUE FIBERS IN SYRINX DENĠZLĠ
COCK AND CHICKEN (Gallus gallus domesticus) OF PHEASANT (Phasianus
colchicus)
Öznur ÖNAL
Süleyman Demirel University
Graduate School of Applied and Natural Sciences
Department of Biology
Supervisor: Asst. Prof. Dr. Kenan ÇINAR
The objective of this thesis is to determine the arrangement and distribution of
connective tissue fibers in the Denizli rooster and chicken (Gallus gallus domesticus)
and pheasant (Phasianus colchicus). For this purpose, 4 adult male and 4 female
Denizli rooster and chicken, plus 4 male and 4 female pheasants were used as
material for the syrinx. The structure of the syrinx of the Denizli rooster / chicken
and the female pheasants consists of one tracheal ring and the first 5 bronchial rings
of cartilage. The male pheasants syrinx has one trachea ring and 3 bronchial rings of
cartilage. All the studied animal species where found to have tracheobronchial
syrinx’s.
LTM is located between the last siringeyal ring and the first tracheal siringeyal
bronchial cartilage. Medial tympanic membran is located between pessulus and
ligamentum interbronchiale.
In Denizli roosters collagen fibers are under the lateral tympanic membran
epithelium bunched deeper into a broad range of connective tissue, especilay the
deeper parts where the fibers are broader and tighter.
MTM epithelial collagen fibers are spread over a wider area than the settlement of
the elastic fibers in the form of thick fibers of connective tissue deeper into Denizli
chicken of tissue was more intensive.
Male pheasants LTM under the epithelium of the bronchial epithelium in the lateral
cartilage rings and pessulus thin collagen fibrils formed bundles was met firmly.
Keywords: Connective tissue, Fiber, Denizli rooster, pheasant, syrinx.
2013, 45 pages

iv
TEġEKKÜR
Bu tez çalışmasında konunun bulunmasında, yürütülmesinde büyük emeği geçen,
karşılaştığım zorlukları bilgi ve tecrübesi ile aşmamda yardımcı olan değerli
danışman hocam Yrd. Doç. Dr. Kenan ÇINAR’a teşekkürlerimi sunarım.
Ayrıca 3311- YL1-12 No’lu proje ile tezime maddi desteğinden dolayı Süleyman
Demirel Üniversitesi Bilimsel Araştırma Projeleri Yönetim Birimi Başkanlığı’na
teşekkürler ederim.
Öznur ÖNAL
ISPARTA, 2013

v
ġEKĠLLER DĠZĠNĠ
Sayfa
Şekil 4. 1. Trakeyal sirinks. Submukoza (ince ok) ve kıkırdak halka arası (Kalın
ok) kollagen fibril. Van Gieson. Bar: 80 µm. ........................................... 19 Şekil 4. 2. LTM. Epitel altı (ince ok) ve submukozada (Kalın ok) kollagen fibril.
Van Gieson. Bar: 80 µm.......................................................................... 19 Şekil 4. 3. Bronşiyal sirinks. Epitel altı (ince ok) ve kıkıdak halka arası (kalın ok)
elastik fibril. Weigert. Bar: 80 µm. .......................................................... 20 Şekil 4. 4. MTM. Epitel altı (oklar) elastik fibril. Weigert. Bar: 80 µm. ................... 21
Şekil 4. 5. LTM. Epitel altı (ince ok) ve sub mukozada (kalın ok) retikulum
fibrilleri. Gordon&Sweet. 60 µm............................................................. 22
Şekil 4. 6. Bronşiyal sirinks. Epitel altı (kalın ok) ve kıkırdak halka arası (ince ok)
retiku lum fibril. Gordon&Sweet. 60 µm. ................................................ 22
Şekil 4. 7. Bronşiyal sirinks. Kıkırdak halka arası (oklar) kollagen fibril.Van
Gieson. Bar: 80 µm. ................................................................................ 23
Şekil 4. 8. MTM. Bağ dokusu içeri sinde kollagen fibriller (oklar). Van Gieson.
Bar:80 µm ............................................................................................... 24
Şekil 4. 9. LTM. Epitel altı (oklar) elastik fibriller. Weigert. Bar: 60 µm. ................ 25 Şekil 4. 10. MTM. Epitel altı (*) ve submu kozada (oklar) elastik fibriller.
Weigert. Bar:80 µm. ............................................................................. 25 Şekil 4. 11. Bronşiyal sirinks. Kıkırdak halka arası (oklar) retikulum fibrilleri.
Gordon & Sweet. 80 µm. ...................................................................... 26 Şekil 4. 12. MTM. Epitel altı (*) ve submukoza (oklar) elastik fibriller.
Gordon&Sweet. Bar:60 µm. ................................................................. 27 Şekil 4. 13. Bronşiyal sirinks. Kıkırdak halka arası (oklar) kollagen fibril. Van
Gieson. Bar:80 µm. ............................................................................... 28 Şekil 4. 14. MTM. Submukozada (oklar) kollagen fibril. Van Gieson. Bar: 60 µm. . 28
Şekil 4. 15. Trakeyal sirinks. Epitel altı (*) ve kıkırdak halka arası (ok) elastik
fibril. Weigert. Bar:60 µm..................................................................... 29
Şekil 4. 16. MTM. Epitel altı (oklar) elastik fibril. Weigert. Bar:60 µm. .................. 29 Şekil 4. 17. Bronşiyal sirinks. Kıkırdak halka halka arası(oklar) retikulum
fibrilleri. Gordon & Sweetµm. Bar: 60 µm. ........................................... 30 Şekil 4. 18. MTM. Epitel altı (oklar) retikulum fibrilleri. Gordon&Sweet. Bar: 80
µm. ....................................................................................................... 31 Şekil 4. 19. LTM. Epitel altı (oklar) kollagen fibrilleri. Van Gieson. Bar: 60 µm. .... 32
Şekil 4. 20. MTM. Epitel altı (oklar) kollagen fibriller. Van Gieson. Bar: 60 µm. .... 32 Şekil 4. 21. Bronşiyal sirinks. Epitel altı (*) ve kıkırdak halka arası (ok) elastik
fibriller. Weigert. Bar: 80 µm................................................................ 33 Şekil 4. 22. MTM. Submukozada elastik fibriller (oklar). Weigert. Bar:60 µm. ....... 34
Şekil 4. 23. LTM. Submukoza (oklar) retikulum fibrilleri. Gordon&Sweet. Bar:
60 µm. .................................................................................................. 35
Şekil 4. 24. Bronşiyal sirinks. Epittel altı (*) ve kıkırdak halka arası (ok)
retikulum fibrilleri. Gordon & Sweet. Bar:80 µm. ................................. 35

vi
ÇĠZELGELER DĠZĠNĠ
Sayfa
Çizelge 4. 1. Denizli horozu ve Denizli tavuğu ile dişi ve erkek sülün
sirinkslerinde bağ dokusu fibrilleri ..................................................... 17

1
1. GĠRĠġ
Kuşlarda solunum sisteminin başlıca fonksiyonları, vücutta oksijen ve karbondioksit
değişimini sağlamak, vücut ısısını dengelemek, solunum yüzeyi dehidrasyonundan korumak
ve sesin oluşumuna katkıda bulunmaktadır (Bell ve Freeman, 1971; Fedde, 1998; Demirsoy,
1995).
Solunum sistemi, organizma ile dış ortam arasında gaz alışverişini sağlayan bir ucu açık diğer
ucu kapalı bir sistemdir (Öber ve İzzetoğlu, 2006; Junqueira ve Carneiro, 2009). Bu sistem
fonksiyonel olarak respiratorik ve iletici bölüm olmak üzere iki kısımdan oluşur (Junqueira
vd., 1998; Tanyolaç, 1999; Yaman, 1999). İletici bölüm burun boşlukları, sinüsler,
nasofarinks, orofarinks, larinks, trake, bronkus ve bronkuluslardan; respiratorik bölüm ise
respiratuvar bronşiyaller, alveolar kanalları, alveolar keseler ve alveollerden oluşur (Junqueira
ve Carneiro, 2009). Kuşlarda üst solunum yolları burun delikleri, burun boşluğu, orofarinks
ve trakeden; alt solunum yolları ise sirinks, bronşlar, akciğerler ve hava keselerinden oluşur
(Taşbaş vd., 1994).
İletici bölümün iki önemli görevi vardır. Bunlar; havanın akciğerlere gidip gelebileceği bir
kanal sağlamak ve solunan havayı uygun hale getirmektir. Kıkırdak, elastik ve kollagen lifler
ile düz kas sayesinde iletici bölüme sert bir yapısal destek sağlanırken, bükülebilme ve
uzayabilme yeteneği ile hava iletiminin kesintisiz olması güvence altına alınır (Junqueira vd.,
1998; Junqueira ve Carneiro, 2009). Hem iletici bölüm hem de solunum bölümü, yapısına
esneklik kazandıran ve gerildikten sonra tekrar eski durumuna dönmesini sağlayan bol
miktarda elastik lifler ile donatılmıştır. İletici bölümde elastik lifler lamina propriada bulunur;
yerleşimleri genelde longitudinaldir. Elastik lif yoğunluğu iletici tüpün çapı ile ters orantılıdır
(yani en küçük bronşiyoller en yüksek oranda elastik life sahiptir. Trakeadan alveol
kanallarına kadar olan tüpleri düz kas demetleri kuşatır (Junqueira vd., 1998).
Solunum sisteminin başlangıcını oluşturan burun boşluğu (cavum nasi), önde nares denilen
burun delikleri ile dış ortama, arkada koana (choana) denilen iki açıklıkla nasofarinkse
bağlanır (Öber ve İzzetoğlu, 2006). Burun boşluğu daha sonra genişleyerek septum nasi
denilen (kıkırdak ve kemik septumla) bir bölme ile uzunlamasına ortadan iki yarıma
ayrılmıştır (Demirsoy, 1995; Özcan, 2011).

2
Burun boşluğu epitelin karakterine göre; ön (vestibuler bölge= regio vestibularis), arka
(respiratuvar bölge= regio respiratoria) ve üst (olfaktif bölge= regio olfaktoria) olmak üzere
üç bölgeye ayrılmıştır (Tanyolaç, 1999; Öber ve İzzetoğlu, 2006; Özcan, 2011).
Burun boşluğu girişini oluşturan regio vestibularis kutan mukoza ile kaplı olup, çok katlı yassı
epitele sahiptir. Lamina propriyasında yağ ve ter bezleri içerir. Ayrıca bu bölüm kan
damarları ve sinirler bakımından da zengindir (King ve Mclelland, 1984; Özcan, 2011). Regio
respiratoria, burun boşluğunun en geniş bölümüdür. Glandular mukozaya sahiptir. Yalancı
çok katlı prizmatik epitelle örtülüdür. Epitel hücreleri arasında goblet hücreleri boldur.
Kinosilyumların hareketi larinkse doğrudur (Tanyolaç, 1999). Lamina propriyasında çok
sayıda kollagen ve elastik lifler ile kan damarları ve sinir telleri bulunur (Junqueira vd., 1998).
Regio olfaktoria, koku duyusu ile ilgili bölümdür. Bu mukoza glandular özelliktedir ve regio
respiratoria’dakinden daha kalındır (Tanyolaç, 1999).
Burun boşluğunun görevi, havadaki yabancı cisimleri vestibulde bulunan uzun kıllarla tutmak
ve nazal mukozaya geçişlerine engel olmak ve nem-ısıyı ayarlamaktır (Erkoçak, 1982).
Kuşlarda burun boşluğunun memelilerden farklı olarak, su ve enerji tasarrufunda önemli rolü
bulunur. Burun boşluğunun ön kısmının arka kısmına oranla daha soğuk olması nedeniyle,
soluk verme sırasında vücudu terk edecek olan havadaki nemin suya dönüşerek dışarı atılması
engellenmiş olur. Bu su tasarrufu özellikle göçmen kuşlar ile çölde yaşayan kuşlar için büyük
önem taşır (Getty, 1975).
Burun boşluğundan sonra gelen farinks, sindirim sisteminin ağız boşluğu ile solunum
sisteminin burun boşluğunun kesiştiği bir bölgedir (Zık, 2011). İç burun delikleri ile larinks ve
özafagusun giriş kısımları arasında yer alır (Martini vd., 2000). Mukozasının çeşitli hücresel
kompozisyonları nedeniyle üç bölgeye ayrılır. Bunlardan nazofarinks respiratorik mukoza ile
örtülüdür ve epitel hücreleri arasında goblet hücreleri bulunur. Nazofarinks yutkunma
sırasında yumuşak damak ile küçük dil nazofarinksinin kapanmasını sağlayarak, besinlerin
burun boşluğuna geçişini önler (Tanyolaç, 1999; Zık, 2011). Orofarinks ağız boşluğunun
posteriorundadır ve onun devamı halindedir. Kutan mukoza özelliğine sahiptir. Lamina
epitelyalisi çok katlı yassı epitelden oluşmuştur ve lamina propriyasında ise bezler yoktur
(Tanyolaç, 1999; Özcan, 2011). Orofarinks yutkunma sırasında, hem besinlerin hem de
solunan havanın geçişini sağlar. Farinksin son bölümünü oluşturan laringofarinks çok katlı

3
yassı epitel ile kaplıdır ve orofarinks gibi besinler ve hava için bir geçiş yeri oluşturur
(Marieb, 2001).
Larinks (gırtlak) farinks’i trakeye bağlayan düzensiz bir boru şeklindedir (Tanyolaç, 1999;
Junqueria ve Carnerio, 2009). Bu borunun duvarı lümenden itibaren mukoza, kıkırdak ve
kaslardan oluşur. Larinks aynı zamanda ses çıkarma ile ilgili yapısal özelliklere de sahiptir.
Larinksin mukozası gırtlağın başlangıç bölümünde (vestibulum laringis) ve sesin meydana
gelmesinde şiddetli zorlamalara uğrayan pilika vokalis’lerde (mukoza dürümleri) kutan, geri
kalan bölümde (kavum laringis propriyum) respiratorik karakterdedir. Plika içinde epitelin
altında ses telleri demetleri (ligamentum vokale) ve onun da altında iskelet kasları (muskulus
vokalis) bulunur (Tanyolaç, 1999; Zık, 2011). Larinks kıkırdaklarından geniş ve tek olan üç
tanesi (tiroid, krikoid, epiglottis) larinksin gövdesini şekillendirir (Marieb, 2001). Tiroid ve
krikoid kıkırdakların tamamı hiyalin kıkırdaktan meydana gelir (Tanyolaç, 1999; Junqueria ve
Carnerio, 2009; Özcan, 2011). Kıkırdaklar, destek sağlamanın yanı sıra, katı ve sıvı besinlerin
trakeaya kaçmasını önleyici bir kapak görevi üstlenirler. Ayrıca ses oluşumuna da katkıda
bulunurlar (Junqueira vd., 1998; Junqueria ve Carnerio, 2009, Özcan, 2011). Epiglottis,
larinksin üst bölümündedir ve bulunduğu yer ve görev bakımından büyük önem taşır.
Yutkunma esnasında epiglottis, glottisin üstünü örtecek şekilde geriye katlanarak besin
maddelerinin hava yoluna geçmesini önler (Tanyolaç, 1999). Epiglottisin altında, mukoza
larinksin lümenine uzanan iki çift katlantı yapar. Üstteki çift yalancı ses tellerini (vestibüler
katlanmalar) oluşturur. Bunlar solunum epiteli ile döşelidir. Alttaki çift katlanma ise gerçek
ses telleridir. Vokal bağı oluşturan iri demetler halinde birbirine paralel elastik lifler, çok katlı
yassı epitelle döşeli vokal katlanmaların içinde yer alır. Bağlara paralel olarak, katlanma ve
bağlardaki gerilimi düzenleyen ve iskelet kası demetlerinden oluşan ses kasları yer alır. Hava
katlanmalar arasında sıkıştırıldığı zaman bu kaslar değişik frekanslarda seslerin oluşmasını
sağlar (Junqueira vd, 1998; Junqueria ve Carnerio, 2009). Yalancı ses telleri kas
içermediğinden dolayı ses oluşumunda görev almaz, ancak ses rezonansını (tınısını) sağlar
(Eşrefoğlu, 2009). Sesin oluşumu glottisten geçen havanın ses tellerini titretmesi ile
gerçekleşir. Sesin özelliği bu vokal tellerin çapına, büyüklüğüne ve gerginliğine bağlıdır
(Ayvaz, 2010).
Kuşlarda memelilerde bulunan tiroid kıkırdakları ile epiglottis bulunmaz (King ve Mclelland,
1984; Taşbaş vd., 1986).

4
Larinksin hareketleri (daralıp genişlemesi) kıkırdakların çevresindeki çizgili kaslarla sağlanır
ve bunun sonucunda da larinks boşluğunun şekli değişir. Ayrıca ses çıkarmada gerekecek
çabuk reaksiyon da bu kaslarla sağlanmaktadır (Öber ve İzzetoğlu, 2006). Plika vokalislerin
az ya da çok sıkışıp gevşemesi ve akciğerlerden gelen havanın etkisiyle de çeşitli tonlarda
sesler çıkarılır. Kasların kontraksiyonuyla akciğerlere yabancı maddelerin geçişi önlenmeye
çalışılır. Yabancı maddeleri uzaklaştırmaya yarayan şiddetli öksürük refleksi yine bu kasların
kontraksiyonuyla gerçekleştirilir (Tanyolaç, 1999).
Kuşlarda larinks, larinks kraniyalis ve larinks kaudalis (sirinks) olmak üzere iki kısımda
incelenir. Larinks kraniyalis trakenin başlangıcında bulunan yalnızca solunumla ilgili
bölümdür (Tanyolaç, 1999, Zık, 2011). Duvar yapısında içten dışa doğru mukoza, kıkırdak
halkalar ve kaslar yer alır. Mukoza epiteli tek katlı prizmatik ve yalancı çok katlı prizmatik
epitel karışımıdır. Kuşlarda larinks kıkırdakları, cartilago cricoidea, cartilago procricoidea ve
iki adet cartilago arytenoidea’dan oluşur. Kısmen kemikleşmiş olan bu dört adet kıkırdak bağ
doku ile birbirine bağlanır. Kuşlarda ses telleri bulunmaz (Özcan, 2011).
Larinks ile bronkuslar arasında uzanan trake tüp şeklinde bir organdır. Dorsalinde yemek
borusu bulunur (Tanyolaç, 1999; Eşrefoğlu, 2009). Solunum sisteminin iletici bölümünün
büyük bir kısmını oluşturur. Lümenden itibaren trakenin duvar yapısına katılan oluşumlar
mukoza, kıkırdak halkalar ile ligamentler, muskularis ve adventisyadır (Tanyolaç, 1999).
Lamina epitelyalis yalancı çok katlı prizmatik epitel ile örtülüdür. Goblet hücreleri sayıca
fazladır. Lamina propria gevşek bağ dokusu karakterindedir (Tanyolaç, 1999; Junqueria ve
Carnerio, 2009; Özcan, 2011). Submukoza tabakasının altında yer alan 16-20 adet C
şeklindeki kıkırdak halkaları lümenin açık kalmasını sağlar. C şekilli trakeal kıkırdakların
açık kısımları posterior olarak özofagusa bakar (Tanyolaç, 1999; Eşrefoğlu, 2009).
Kuşlarda memelilerden farklı olarak trake halkaları tam bir halka şeklindedir (Taşbaş vd.,
1994; Erdost, 2011) ve türlerin boyun uzunluklarına bağlı olarak kıkırdak halkaların sayısı
değişmektedir. Genellikle trake, 108-126 adet kıkırdak halkadan oluşur (King ve Mclelland,
1984; Taşbaş vd., 1986). Trakede kıkırdak halkaların sağ ve sol yarımları geniş olup,
medianlarında birer çentik bulunur. Kıkırdak halkaların geniş kısımları da mediandaki çentik
aracılığıyla birbiri üzerine geçer. Bu nedenle ligamentum anulare de bulunmaz (Piperno ve
Peirone, 1975).

5
Kuşların trakesinde, m. tracheolateralis, m. sternotrachealis, m. cleidotrachealis, m.
sternohyoidideus olmak üzere toplam 4 adet kas bulunur (Dursun, 2002).
Kuşlarda akciğerin yapısı memeli akciğerinden tamamıyla farklıdır. Memeli akciğerinde, hava
kanallarının alveollerde sonlanan ağaç benzeri dallanmaları kanatlı akciğerinde yoktur.
Kuşlarda alveol yerine hava kapillarları bulunur (Hodges, 1974). Ayrıca kuşlarda akciğerler
lopsuz olarak şekillenir ve diyafram da bulunmaz. Vücut boşluğunu dolduran akciğerler,
kuşlarda pleura’ya karşılık bir zarla örtülmez, thorax duvarına fibröz bir bağ doku aracılığıyla
sıkıca bağlanır. Kuşların solunum sisteminde memelilerden farklı olarak sirinks ve hava
keseleri bulunur (Demirsoy, 1995).
Kuşların akciğerleri, bir primer bronş (bronchus, mesobronchus), dört adet sekonder bronş
grubu, çok sayıda tersiyer bronş (parabronchus) ile bunlarla ilişkili atria ve hava kapillarından
oluşur (Bell ve Freeman, 1971; King ve Mclelland, 1984; Taşbaş vd., 1994).
Akciğerlere giren primer bronşlar akciğerleri boydan boya geçtikten sonra abdominal hava
kesesine açılır. Primer bronşlar seyri boyunca akciğer içerisine kollar verir. Bunlar sekonder
bronşlardır. Sekonder bronşlar dorsal ve ventral olmak üzere iki gruptur. Bu iki grup sekonder
bronşları birbirine bağlayan tersiyer bronşlardır (parabronşlar). Atriyaların dip kısımlarında
hava kapilları ve bu kapillarların etrafında da kan papillaları vardır. Kuşlarda gaz alışverişi de
bu kısımda gerçekleşir.
Primer bronşların duvar yapısı mukoza, kıkırdak, kas ve adventisyadan oluşur. Epitel kat
yalancı çok katlı prizmatik yapıdadır. Yine mukoza katmanında basit alveolar müköz bezler
yer alır. Primer bronşların ilerleyen bölümlerinde epitel kalınlığında azalma ve bezlerin
ortadan kalkması söz konusudur. Kıkırdakların dış tarafında elastik ipliklerden zengin bağ
dokusu bulunur. Sekonder bronşlarda epitel katman basık prizmatik kinosilyumlu epitel
hücrelerine dönüşür. Epiteli alt taraftan az miktarda elastik ve kollagen iplik içeren bağ
dokusu sarar. Tersiyer bronşlarda (parabronşlar) iyice yassılaşmış epitel katman, etrafından
yoğun elastik iplik ağıyla desteklenmiştir. Tersiyer bronşlara açılan atriyum epiteli, iri
çekirdekli kübik hücrelerden oluşur. Epitel etrafında elastik ve kollagen iplik yoğunluklu bağ
doku yer alır. Kuşların solunum sistemine ait ince duvarlı, kese görünümündeki yapılar hava
keseleridir. Bronkusların ekstra pulmonal genişlemeleri ile şekillenmişlerdir. Fonksiyonel
olarak 9 adet hava keseciği, anterior keseciklere (interklavikular) ve posterior (abdominal)

6
keseciklere ayrılabilir. Hava keseleri tek katlı yassı örtü epiteline sahiptir. Ayrıca bu keselerin
gaz değişimi ile doğrudan bir rolü yoktur. Vücut ağırlığının azaltılmasında ve iç buharlaşma
ile vücut sıcaklığının sabit tutulmasında ve sesin oluşumuna yardımcı olmada etkindir (Özcan,
2011). Ayrıca testislerle de bağlantılı olduklarından, memelilerin scrotomuna benzer bir
görevle bu kısımların sıcaklığının azaltılmasını ve spermaların oluşmasını da sağlarlar. Sucul
kuşların su üstünde kalabilmeleri ve bazı kuşların kur yapma sırasında vücut tüylerinin
kabartılması da hava keselerinin sayesinde olur ( Demirsoy, 1995; Kuru, 2009). Kuşlarda
havanın solunum sistemindeki sirkülasyonu iki inspirasyon ve iki ekspirasyon olmak üzere
dört aşamada gerçekleşir. İlk inspirasyon ile alınan hava solunum yolunu izleyerek kaudal
hava keselerine gelir ve ilk ekspirasyon sırasında sıkışan hava kaudal hava kesesi aracılığıyla
akciğerlere geçer. İkinci inspirasyonla akciğerlerde bulunan hava cranial hava keselerine girer
ve hava ikinci ekspirasyon ile vücuttan dışarı atılır. Kuşların akciğerlerinde hava ve dolaşım
zıt yönde olur ve buna ‘ters akım alış-veriş sistemi’ adı verilir. Bu sistem sayesinde kan
akciğeri terk etmek üzere iken, oksijen yönünden zengi olan havadan da oksijen alır (Noyan,
1993).
Kuşlarda ses çıkarma (ötme), toplu yaşayan ve sürü oluşturan kuşlarda bir araya toplanmayı
sağlamak, kuluçkaya yatma alanlarını başka kuşlara tanıtmak, üreme mevsiminde karşı cinsin
ilgisini çekmek, anne, baba ve yavrular arasındaki haberleşmeyi sağlamak ve bir tehlikenin
varlığını diğer kuşlara bildirmek için kullanılır (Demirsoy, 1995; Kuru, 2009).
Kuşlarda çağrı ve şarkı sesi olmak üzere iki tür ses vardır. Bu canlılar çıkarttıkları son derece
güçlü seslerle birbirlerine çağrıda bulunurlar. Çağrı seslerinin çok özel işlevleri vardır ve şarkı
seslerinden farklı olarak daha çok içgüdüseldir. Bu sesler daha çok cinsiyet belirlenmesinde
kullanılır. Kuşlar hayatlarının çok erken dönemlerinde, hatta yumurtadan çıkmadan önce
seslerini kullanmaya başlarlar. Aynı zamanda yavrularda yumurta içerisindeyken
ebeveynlerinin seslerini dinlerler. Daha yumurtadan çıkmadan ebeveyniyle bu yolla iletişim
kurarak onları tanıyabilirler.
Ayrıca kuşlarda ses, bir kuşun türünü tanımlamak, kuşun yerini belirlemek, kuluçkaya yatma
alanlarını korumak, besin kaynağı olduğunu bildirmek, sürüyü bir arada tutmak, düşmanın
varlığı hakkında uyarmak, düşmanları korkutmak gibi durumları belirtmek için de kullanılır
(Kuru, 2009).

7
2. KAYNAK ÖZETLERĠ
Kuşlarda ses çıkarma organı olan sirinks (larynx caudalis) göğüs boşluğunda, trakenin
bifurkasyon yerinde primer bronşlar üzerinde bulunur (King, 1989; Taşbaş vd., 1994; Yıldız
vd., 2003). Sirinks trakenin devamı niteliğinde olup, trakeyal ve bronşiyal kıkırdak halkaların
birleşmesiyle şekillenir. Sirinks iskeleti timpanum ile trakeyal siringeyal ve bronşiyal
siringeyal kıkırdaklardan oluşur. Genellikle trakeyal siringeyal kıkırdakların uç kısmındaki
kıkırdaklar timpanum şeklini almıştır. Timpanum, son iki veya daha fazla trakeyal siringeyal
kıkırdağın kaynaşması ile şekillenir.
Trakenin son halkaları kümes hayvanlarında yanlardan yassılaşmış ve daralmış bir şekilde
görünür (Doğuer ve Erençin, 1964). Kazlarda ise son trake halkaları hemen hemen tam olarak
kaynaşmış bir durumdadır. Timpanumu oluşturan kıkırdak halkalarının çapı trakeyal kıkırdak
halkalardan biraz daha büyüktür (Getty, 1975). Bronşiyal siringeyal kıkırdaklar genellikle çift
halde bulunur ve ‘C’ biçimindeki kıkırdak halkalardan oluşmuştur (Baumel vd., 1993).
Sirinksin büyük bir bölümünü oluşturan ve timpanumun kaudomedialinde yer alan pessulus,
trakenin bifurkasyon yeri ile bronkusların başlangıcı arasında dorsa-ventral pozisyonda
yerleşmiş üçgen biçimindeki yapıdır (Taşbaş vd., 1994; Rodentio, 1998; Kuru, 2009).
Pessulus’un güvercinde (Yıldız vd., 2005) bağ dokusundan, Denizli horozu (Taşbaş vd.,
1994), şahin (Kabak vd., 2007), bıldırcın (Çevik-Demirkan vd., 2007) ve martıda (Gezer İnce
vd., 2012) da kemik dokudan oluştuğu belirtilmiştir. Ancak Bayram ve Liman (2000)
bıldırcında başlangıçta hiyalin kıkırdak karakterinde olan pessulusta yumurtadan çıkışın 42.
gününden itibaren kemik doku odakları oluştuğunu bildirmişlerdir. Ayrıca yeşilbaş ördek
(Frank vd., 2006; 2007) sirinksinde pessulusun sadece uç kısmının hiyalin kıkırdaktan, diğer
kısımlarının kemik dokusundan oluştuğu belirtilirken; devekuşu (Yıldız vd., 2003) sirinksinde
pessulusun kıkırdak veya kemik doku içermediği bildirilmiştir. Pessulus şekil itibariyle evcil
kanatlılarda (Getty, 1975; Taşbaş vd., 1994) kama, Japon bıldırcınlarında (Çevik-Demirkan
vd., 2007) yarı prizma, erkek ördekte (Frank vd., 2006) uzunlamasına oval görünümdedir.
Sirinksin oluşumuna trakeyal veya bronşiyal kıkırdakların katılmasına göre sirinks;
trakeyobronşiyal, trakeyal ve bronşiyal tiplere ayrılır (Demirsoy, 1995; Frank vd., 2007;
Kabak vd., 2007). Ötücü kuşlar genelde trakeyobronşiyal tip sirinkse sahiptirler (Larsen ve
Goller, 2002).

8
Sirinkste ses oluşumu ile ilgili lateral (laterik timpanik membran = LTM) ve mediyal
(mediyal timpanik membran = MTM) konumlu iki çift membran bulunmaktadır. Membranlar
birbirinden bağımsız çalışır ve sirinksi çevreleyen ekstrinsik ve intrinsik kasların gergin halde
olmasını sağlar (Taşbaş vd., 1994; Yıldız vd., 2005). Bu kaslar kıkırdak halkaları farklı
derecelerde hareket ettirerek, ses oluşumunda etkin olan membranların gerginliğini değiştirir;
böylece sesin tonu ve ritmi de ayarlanmış olur (Taşbaş vd., 1994; Yıldız vd., 2003; Kabak vd.,
2007). Lateral (Membrana timpaniformis lateralis) ve mediyal (Membrana timpaniformis
medialis) membranların sirinks iskeletinde yerleşim yerleri genellikle kuş türleri arasında
farklılık göstermektedir. Denizli horozu (Taşbaş vd., 1994), bıldırcın (Bayram ve Liman,
2000; Çevik-Demirkan vd., 2007), güvercin (Yıldız vd, 2005) ve baykuşta (Çevik-Demirkan
ve Özdemir, 2011) LTM’ın son trakeyal kıkırdak halka ile ilk bronşiyal kıkırdak halka
arasında yer aldığı bildirilmiştir.
MTM’ın şahinde (Kabak vd, 2007) pessulusun kaudali ile bronşiyal kıkırdaklar arasında,
baykuşta (Çevik-Demirkan ve Özdemir, 2011) ilk trakeyal siringeyal kıkırdak halka ile son
bronşiyal siringeyal kıkırdak halka arasında, kazda (Onuk vd., 2010) pessulus ile 2. bronşiyal
kıkırdak arasında ve martıda (Gezer İnce vd., 2012) bronşiyal kıkırdakların mediyalinde
bulunduğu bildirilmiştir.
En gelişmiş sirinksi içeren ötücü kuşlar ile diğer bazı kuşların (Goller ve Suthers, 1996)
sirinkslerinde sirinks membranlarının titreşimini sağlayan özel sirinks kasları bulunur.
Bunların yanında solunum organı kasları da ses çıkarma olayına yardımcı olurlar. Bu
kapsamda Kahverengi Batöz ve Kanarya gibi ötücü kuşlarda (Larsen ve Goller, 2002) 5 çift,
orman kargasında ise 7 çift özel siringeyal kas vardır. Diğer kuşların sirinksinde ise özel
kaslar yoktur, bu organın çalışması trake ve bronkus kasları ile sağlanır (Rodentio, 1988).
Tavuk (Gallus gallus) ötücü olmayan bir kuştur ve ses ile ilgili sadece 3 çift trakeyal kasa
sahiptir (Gaunt ve Gaunt, 1977). Ayrıca güvercin (Yıldız vd., 2005) ve malard ördekte de
(Lockner ve Youngren, 1976) siringeyal kas yoktur. Sadece 2 veya 3 çift trakeyal kas
bulunur. Bu bakımdan da bu kuşlarda çok zayıf bir ses meydana getirilir. İntrinsik kaslar;
musculus tracheobronchialis dorsalis, musculus tracheobroncialis brevis, musculus
tracheobroncialis ventralis, musculus syringealis dorsalis, musculus syringealis ventralis,
musculus vocalis dorsalis, musculus vocalis ventralis, musculus obliquus ventralis, musculus
obliquus lateralis, musculus syringealis superficialis, musculus syringealis profundus,

9
musculus syringealis caudalis ve musculus sternotrachealis’dir. Ekstrinsik syringeal kaslar ise
musculus tracheolateralis ve musculus sternotrachealis’tir (Baumel vd., 1993).
Erkek kuşların sirinksleri dişi kuşlarınkine kıyasla çok fazla gelişmiştir. Ötücü kuşlarda cinse l
farklılıklar özellikle siringeyal kasların boyutundan kaynaklanır. Dişilerde sternotrakeyal
kaslar erkeklerdekine nazaran oldukça küçüktür (Myers, 2010). Erkek kuşların ses
repertuvarları genellikle dişilerden daha fazladır. Bazı türlerde örneğin; zebra ispinozlarında
sadece erkekler şarkı söyler. Ayrıca dişi kuşlarda şarkı repertuvarları ve şarkı süreleri
erkeklerden daha kısadır.
Kuşun ötüşü sırasında meydana gelen sesler, kasların yardımıyla sirinks halkalarının ve
dolayısıyla membranların birbirlerine yaklaşıp uzaklaşması, pessulusun titreştirilmesi ile
meydana gelen hava titreşimleri ile oluşur (Rodentio, 1988).
Bağ dokusu vücudun en önemli dokularından birisidir. Vücut şeklinin sağlanması ve
devamlılığından sorumludur. Bağ dokuları mekanik bir rol üstlenerek hücre ve organları
birbirine bağlamaya yarayan bir matriks oluştururlar ve sonuçta vücuda destek sağlarlar.
Bağ dokusu çok değişik işlevlere sahiptir, bunların en dikkat çekici olanı ise yapısal işlevidir.
Vücudun organlarını ve onların hücrelerini destekleyen iç kurguyu çevreleyen kapsüller, bağ
dokusundan oluşurlar. Bu doku aynı zamanda tendonler, ligamentler ve organlaın arasındaki
boşlukları dolduran areolar dokuları da oluşturur. Kemik, yağ dokusu ve kıkırdak dokusu
yumuşak dokuları destekleme görevlerini yerine getirmek ve yağları depolamak için özel
şekilde değişmiş bağ dokusu tipleridir (Junqueria vd., 1998). Ayrıca bağ dokusu organizma
için savunucu, koruyucu ve tamir edici bir sistemdir (kan ve lenf doku). Bağ dokusu diğer
dokuları beslemede de rol oynar. Bağ dokusunun bu rolü kan damarları ile yakın ilişkisinden
kaynaklanır. Bağ dokusu matriksi, hücrelerle onların kanlanma yolları arasında besinlerin ve
metabolik atıkların değiş-tokuş edildiği yerdir, değiştirilen besin ve metabolik atıklar için bir
ortam oluşturma işlevi görür (Junqueria vd., 1998; Öber ve İzzetoğlu, 2006).
Yapısal bileşimi bakımından bağ dokusu üç alt bileşenden oluşur. Bunlar; hücreler, lifler ve
ara maddedir. Esas olarak hücrelerin oluşturduğu diğer doku tiplerinden farklı olarak bağ
dokusunun ana bileşeni hücre dışı maddedir. Hücre dışı madde de protein lifleri (kollagen,
elastik, retiküler) ve ara maddenin farklı bileşimlerinden oluşur. Vücut içinde oldukça çok
çeşitli şekillerde bulunan bağ dokusunun tipleri, bu üç bileşenin kompozisyonu ve

10
miktarındaki farklılıklarla belirlenir. Bağ dokusunda bulunan lifler kollagen, elastik ve
retiküler liflerdir (Eşrefoğlu, 2009).
Ara madde, hücrelerin yüzeyindeki reseptör proteinlerine (integrinler) ve diğer matriks
bileşenlerine bağlanarak matriksin dayanıklılığına ve sertliğine katkıda bulunan anyonik
makromoleküller (glikozaminoglikanlar ve proteoglikanlar) ve çoklu yapışkan proteinlerden
(laminin, fibronektin) oluşan yüksek düzeyde su sever (hidrofilik) kıvamlı bir bileşiktir
(Junqueria ve Carnerio, 2009). Bu madde bağ dokusunun hücre ve lifleri arasındaki boşlukları
doldurur, visközdür ve kayganlaştırıcı işlevinin yanı sıra dokulara yabancı partiküllerin
girmesine karşı engel teşkil eder (Junqueria vd., 1998; Akay, 2006).
Embriyonik kök hücrelerden yani mezenşim hücrelerinden gelişen ve bulundukları dokuya
özel olan bağ dokusu hücreleri sabit hücreler (mezenkim hücreleri, fibroblast, retiküler
hücreler, yağ hücreleri ve plazma hücreleri) ve canlı yapının gereksinimlerine göre dolaşan
hücreler (mast hücreleri, makrofajlar, lenfosit, lökositler) olmak üzere iki gruba ayrılırlar
(Cireli, 1999; Akay, 2006; Öber ve İzzetoğlu, 2006).
Bağ doku fibrillerinden kollagen fibriller; bütün bağ doku türleri için en karakteristik
fibrilleridir. Gevşek bağ dokularında uzun, düz ve dalgalı seyirleri ile kendilerini belli ederler.
Kollagen fibriller gevşek bağ dokuda yan yana gelerek 1-12 mikron kalınlığında demetler
oluştururlar. Bu fibriller çekilmelere karşı çok dayanıklıdırlar, uzayamaz ve genişleyemezler,
elastiki özellikleri yoktur. Kollagen fibriller hücrede ergasitoplazmada salgılanan
proteinlerden önce tropokollagen olarak oluşur. Hücre yüzeyinde mikrofibriller halinde
kondanse olurlar. Fibril oluşumu sitoplazma içinde başlar. Mikrofibrillerin kristalizasyonu ile
de fibriller oluşur. Kollageni oluşturan temel aminoasitler glisin, prolin, hidroksiprolin ve
hidroksilizindir. Zincirde genellikle glisinin önünde hidroksiprolin ve hidroksilizin, arkasında
prolin yer alır. Zincirdeki şeker gruplarının varlığı dolayısı ile kollagen, glikoprotein olarak
tanımlanır. 27 farklı kollagen tipi tanımlanmıştır ve bu kollagen tiplerinin yerleşim yerleri de
birbirinden farklı olabilir. Tip I, II, III, V, XI kollagenler bantlaşma gösteren kollagen
tipleridir. Daha esnek olan Tip IX, XII, XIV, XIX, XX ve XXI diğer liflerin yüzeyinde
bulunur. Bu lifler bir ölçüde bağlayıcı özellik gösterirler. Tip XIII, XVII, XXIII ve XXV
transmembran kollagenlerdir. Tip XV ve XVIII bazal membran zonunda bulunurlar. Tip IV,
VI ve VII kollagenler ise bazal membran kollagenlerdir. Tip I, II ve III kollagen molekülleri
sıkıca bir araya gelerek kollagen fibrilleri oluştururlar. Sıkı ve gevşek bağ dokularında en sık

11
rastlanan kollagen, Tip I kollagendir. Toplam vücut kollageninin %90’ nı oluşturan Tip I
kollagen, fibröz bağ dokularında, deri (dermis tabakası), diş (dentin tabakası), kemik, tendon,
ligament ve organ kapsülünde bulunur. Tip II kollagen, hiyalin ve elastik kıkırdakta, Tip III
kollagen gevşek bağ dokularında, uterus, karaciğer, dalak, böbrek gibi organlarda, deride, kan
damarlarında, kas ve retiküler bağ dokusunda bulunur. Tip VI kollagen bazal membran
kollagenidir. Tip V kollagen retiküler ağla bağlantılı olabilir. Tip VI kollagen kondrositlerin
hemen altındaki kıkırdak matriksini oluşturur. Tip VII kollagen deri, göz, uterus ve
özofagustaki bağlayıcı liflerde bulunur. Tip VIII kollagen endotel hücrelerinin ürünüdür. Tip
IX kollagen Tip II kollagenle beraber kıkırdakta bulunur. Tip XI kollagen Tip II kollagenle
birlikte kondrositler tarafından üretilir. Tip XII kollagen mekanik basınca yüksek oranda
maruz kalan dokulardan, deri ve plasentadan izole edilmiştir. Tip XIII kollagen kemik,
kıkırdak, bağırsak, deri, plasenta ve çizgili kasta bulunan nadir bir transmembran proteinidir.
Tip XV kollagen mezenkim kaynaklı dokularda, kalp ve iskelet kasında yer alır. Tip XVI
kollagenin geniş bir dağılımı vardır. Fibroblast ve arteriyal düz kasların bulunduğu yerlerde,
ancak Tip I kollagen liflerden uzak bulunur. Tip XVII epitelyal hücre membranlarında
bulunan nadir bir transmembran proteinidir. Tip XVIII ise epitelyal ve vasküler bazal
membranlarda yer alır. Tip XIX karaciğerde, Tip XX kornea epitelinde, sternum kıkırdağında
ve tendonlarda, Tip XXI ise kalp ve iskelet kasında bulunur (Cireli, 1999; Öber ve İzzetoğlu,
2006, Eşrefoğlu, 2009).
Elastik fibriller; gerilme ve çekmeye dayanıklılıkları nedeniyle normal boylarının bir veya bir
buçuk katına kadar kopmadan uzayabilen bu lifler doğal hallerinde kıvrımlıdır. Bağ dokunun
elastikiyetini veren de büyük oranda bu liflerdir. Gevşek lifli bağ dokuda ince uzun ve yer yer
dallanmış olarak bulunurlar. Omurgada bulunan bazı ligamentlerde ve büyük kan
damarlarının duvarlarında ise kalın demetler oluşturdukları görülmektedir. Kısmen kollagen
ve retiküler fibril oluşturan hücrelerce (tendon, ligament ve dermiste yer alan fibroblastlarca)
üretilmeleri yanında kan damarlarının düz kas hücrelerince ve farklılaşmamış mezenşimal
hücrelerce de sentezlenmektedir (Öber ve İzzetoğlu, 2006). Elastik fibrillerin esas maddesi
elastin adı verilen bir tür proteindir. Elastik liflerin oluşumunda önce mikrofibriller oluşur,
elastin daha sonra mikrofibril yüzeyinde birikir. Elastinin lif oluşturmasında fibrillin
mikrofibrilleri çok önemli rol oynarlar (Eşrefoğlu, 2009) Bu madde vücut proteinleri arasında
kimyasal etkilere ve ısıya karşı en dayanıklıdır. Elastik iplikler gevşek bağ dokusunda
kollajen ipliklere nazaran daha az bulunurlar ve oldukça uzundurlar (Cireli, 1999). Elastik

12
fibriller akciğer, mide, damar etrafı, ligament, ses telleri, mezenter, mesane gibi gerilmelerle
karşı karşıya kalan vücut kısımlarında bulunur (Akay, 2006).
Retiküler lifler; organizmanın hücresel düzenlenişinde farklı organ ve dokuların yapısına içte
bölmeler ya da dışta örtü olarak katılırlar. Gevşek paketlenmiş proteoglikan ile
glikoproteinlerden yapılmışlardır. Tipik olarak ince fibriler demetler halindedir. Bu
makromoleküller ince köprülerle birbirlerine bağlanarak ağsı yapılanmaya neden olurlar. Ağsı
yapıları nedeniyle hareketli hücrelerin geçişine olanak tanırlar (Öber ve İzzetoğlu, 2006).
Retiküler lifleri oluşturacak olan reticulin, özellikle retiküler hücre ve fibroblastlar tarafından
üretilmekte olup, ayrıca schwann hücreleri de gerektiğinde senteze katılmaktadır (Junqueria
vd., 1998; Öber ve İzzetoğlu, 2006).
Retiküler lifler; embriyonik bağ doku ile gevşek lifli bağ dokuda, özellikle epitel dokuda, yağ
doku hücreleri, düz kas hücreleri, kan damarları ve sinirleri çevresinde yoğun olarak
bulunmaktadır. Ayrıca hemopoitik organlardan lenf düğümü, dalak, kırmızı kemik iliği gibi
yapıların etrafında ağ oluşturacak şekilde bol miktarda bulunurlar (Junqueria vd., 1998; Cireli,
1999; Akay, 2006; Öber ve İzzetoğlu, 2006; Junqueria ve Carnerio, 2009).
Yapılan literatür taramasında farklı kuş türlerinde sirinksin histolojik yapısını belirlemeye
yönelik çalışmaların bulunmasına karşılık bu çalışmada Denizli horozu ve tavuğu ile dişi ve
erkek sülün sirinkslerinde bağ dokusu fibrillerinin belirlenmesine yönelik çalışmanın
bulunmaması konunun önemini artırmaktadır. Bu çalışmanın Kuş Fizyoloji ve Histolojisi
alanında yapılacak çalışmalara katkı sağlayacağı düşünülmektedir.

13
3. MATERYAL ve YÖNTEM
Çalışmada araştırma materyali olarak Süleyman Demirel Üniversitesi Ziraat Fakültesi Çiftçi
Eğitim ve Tarımsal Uygulama kümesinden temin edilen 4 adet 2500-3000 g ağırlığında canlı
erişkin Denizli horozu ve 4 adet 2000-2500 g ağırlığında canlı Denizli tavuğu (Gallus gallus
domesticus) ile 4’er adet erişkin dişi ve erkek sülün (Phasianus colchisus) kullanıldı. Ketamin
HCL (ketalar-50 mg/kg)’in intramusküler yolla verilmesiyle horoz, tavuk ve sülünler derin
anestezi altına alındı. Anestezi altında dekapitasyon yöntemi uygulanarak hayvanların
boyunlarından itibaren anüslerine doğru vücut boşlukları açıldı. Trake ile trakenin
bifurkasyon bölgesi de dahil olmak üzere bronşlarla birlikte örnekler çıkarıldı. Alınan
örnekler Bouin solüsyonunda 18 saat süreyle tespit edildikten sonra rutin histolojik doku
takibi aşamalarından geçirilerek parafinde bloklandı. Parafin bloklardan 5-6 mikrometre
kalınlığında alınan kesitlere aşağıdaki boyama yöntemleri uygulandı.
1. Genel histolojik yapının belirlenmesi için Hematoksilen-Eozin boyama yöntemi
(Culling vd., 1976),
2. Kollagen fibrillerin belirlenmesi için Van Gieson’un bağ dokusu boyama yöntemi
(Van Gieson, 1899),
3. Elastik fibrillerin belirlenmesi için Wiegert’in rezorsin-fuksin boyama yöntemi
(Weigert, 1898)
4. Retikulum fibrillerinin belirlenmesi için de Gordon&Sweet’in retikulum fibril boyama
yöntemi (Gordon&Sweet,1936)
Hazırlanan preperatlar Olympus CX 41 tipi ışık mikroskobunda incelenerek ilgili kısımlardan
fotoğraf çekimi yapıldı.

14
4.ARAġTIRMA BULGULARI
4.1. Histolojik Bulgular
Denizli horozu ve Denizli tavuğu ile dişi ve erkek sülünlere ait sirinkslerin yapısına trakeyal
ve bronşiyal kıkırdak halkalar ile lateral timpanik membran (LTM), mediyal timpanik
membran (MTM) ve pessulusun katıldığı belirlendi.
Ayrıca Denizli horozu ve Denizli tavuğu ile erkek sülün sirinksinde sağ ve sol bronşlar
arasında ligamentum interbronchiale’nin varlığı gözlendi.
Denizli horozu ve tavuğu ile dişi sülünlerde sirinksin yapısına son 1 trakeyal siringeyal halka
ile ilk 5 bronşiyal siringeyal kıkırdak halka katılırken; erkek sülünlerde son 1 trakeyal ve ilk 3
bronşiyal siringeyal kıkırdağın katıldığı dolayısıyla çalışılan hayvan türlerinde sirinkslerin
trakeyobronşiyal tipte olduğu tespit edildi. Ayrıca Denizli horozu ve Denizli tavuğunda
timpanumun son dört trakeya halkasından oluştuğu belirlenirken; dişi ve erkek sülünlerde son
3-4 trakeyal halkadan oluştuğu tespit edildi.
LTM’nin son trakeyal siringeyal halka ile birinci bronşiyal siringeyal kıkırdak halkalar
arasında, MTM’nin pessulus ile ligamentum interbronchiale arasında yer aldığı tespit edildi.
Çalışılan her iki türe ait hayvanlarda sirinksin trakeye ait kısmının yalancı çok katlı prizmatik
örtü epiteline sahip olduğu ve epitel hücreleri arasında goblet hücrelerinin bulunduğu tespit
edildi.
4.1.1. Denizli Horozu
Denizli horozu sirinksinin trakeyal kısmının lamina propriası gevşek bağ doku, trakeyal
kıkırdak halka arasının ise sıkı bağ doku yapısında olduğu tespit edildi.
LTM’nin trake tarafındaki epitelin yalancı çok katlı prizmatik örtü epiteli ile örtülü olduğu,
son 2 trakeyal kıkırdak halka hizasından itibaren çok katlı yassı karakter kazanan epitelin,
birinci bronşiyal kıkırdak halka hizasında tek katlı yassı örtü epiteline dönüştüğü saptandı.
Ayrıca LTM’nın sıkı bağ doku yapısında olduğu ve bu bölgede kan damarlarının trakeya

15
nazaran sayıca az olduğu gözlendi. Pessulusun kranialine ait epitelin altında sıkı bağ dokusu,
bu bölge ile kemik doku arasında gevşek bağ dokusu, pessulus lateralinde ise sıkı bağ
dokusunun bulunduğu gözlendi.
MTM epitelinin pessulus bitiminden birinci bronşiyal kıkırdak halka hizasına kadar yalancı
çok katlı prizmatik, birinci ile ikinci bronşiyal kıkırdak halka arasında çok katlı yassı
karakterde olduğu; ikinci bronşiyal siringeyal kıkırdak halkadan itibaren tekrar yalancı çok
katlı prizmatik epitele dönüştüğü belirlendi.
Pessulusu kuşatan epitelin yalancı çok katlı prizmatik örtü epiteli yapısında olduğu ve epitel
hücreleri arasında az sayıda goblet hücrelerinin bulunduğu saptandı. Ayrıca pessulusun üçgen
şekilde ve iç bölgenin kompakt kemik yapısında olduğu saptandı.
4.1. 2. Denizli tavuğu
Denizli tavuğunda LTM’nin trake tarafındaki epitelin yalancı çok katlı prizmatik epitelle
örtülü olduğu, sondan 7. trakeyal halka hizasında çok katlı yassı karakter kazanan epitelin
birinci bronşiyal kıkırdak halka hizasından itibaren tekrar yalancı çok katlı prizmatik epitele
dönüştüğü belirlendi.
Ayrıca Denizli tavuğu sirinksinin trakeyal kısmındaki kıkırdak halkaların trake bifurkasyon
noktasında yassılaşmış şekilde bulunduğu; trakeyal siringeyal kıkırdak halkalar arasında sıkı
bağ dokusunun bulunduğu ve bu dokunun geniş bir alan kapladığı saptandı.
Denizli tavuğunda MTM epiteli yapısının Denizli horozu ile benzer karakterde olduğu
gözlendi.
Pessulusun yalancı çok katlı prizmatik örtü epiteli yapısında olduğu ve epitel hücreleri
arasındaki goblet hücrelerinin sayıca Denizli horozuna nazaran daha fazla olduğu belirlendi.
Ayrıca pessulusun üçgen şekilli ve iç bölgenin süngerimsi kemik yapısında olduğu gözlendi.

16
4.1.3. Erkek sülün
Erkek sülünlerde trakeyal siringeyal kıkırdak halkaların yerleşim olarak birbirine çok yakın
yerleşim gösterdikleri gözlendi.
LTM epiteli karakterinin Denizli horozunun bulguların benzer olduğu saptandı.
MTM epitelinin pessulus bitiminden birinci bronşiyal kıkırdak halka hizasına kadar çok katlı
yassı karakterde olduğu; birinci bronşiyal kıkırdak halka hizasından itibaren yalancı çok katlı
prizmatik epitele dönüştüğü belirlendi. Ayrıca MTM epitelinde çok sayıda tubuler biçimli bez
hücresinin bulunduğu saptandı.
Pessulus epitelinin yalancı çok katlı prizmatik karakterde olduğu ve goblet hücrelerinin çok az
sayıda olduğu belirlendi.
Ayrıca pessulusun oval şekilli ve iç bölgenin hiyalin kıkırdak yapısında olduğu görüldü.
4.1.4. DiĢi sülün
Dişi sülünlerde LTM’nin trake tarafında bulunan epitelinin yalancı çok katlı prizmatik epitelle
örtülü olduğu, epitelin birinci ve ikinci bronşiyal kıkırdak halkalar arasında tek katlı yassı,
ikinci bronşiyal halka hizasında çok katlı yassı karakterde olduğu; üçüncü bronşiyal kıkırdak
halkadan itibaren ise tekrar yalancı çok katlı epitele dönüştüğü belirlendi.
MTM epitelinin yapı olarak erkek sülünlere benzer şekilde olduğu saptandı. Ayrıca MTM’nin
devam eden derin kısımlarında epitel hücreleri arasında çok sayıda bez hücresi görüldü.
Pessulusu kuşatan epitel hücreleri arasında bulunan goblet hücrelerin sayıca az olduğu ve iç
bölgenin hiyalin kıkırdak yapısında olduğu gözlendi.
4.2. Histokimyasal Bulgular
Uygulanan bağ dokusu fibril boyama yöntemlerini bir tablo halinde özetleyecek olursak:

17
Çizelge 4. 1. Denizli horozu ve Denizli tavuğu ile dişi ve erkek sülün sirinkslerinde bağ
dokusu fibrilleri
D, Dar alan; G, Geniş alan; K, Kalın fibril; İ, İnce fibril; s, Sıkı biçimli; g, Gevşek biçimde
düzenlenmiş; * , çok sıkı biçimde düzenlenmiş.
DENĠZLĠ
HOROZU
DENĠZLĠ
TAVUĞU
ERKEK SÜLÜN
DĠġĠ SÜLÜN
Koll. Els. Ret.
Koll. Els. Ret.
Koll. Els. Ret.
Koll. Els. Ret
Trakeyal örtü
epiteli altı
D İ
Gg
Gg
D İ
Gg
Gg
D İ
sK
Dg
Dİ
Gg
Dg
Trakeyal
kıkırdaklar arası
sK
sK
Dg
Gg
sK*
Dg
gK
sİ
Dg
sK
sİ
Gs
LTM örtü epiteli
altı
gK
Gg
Gg
sK
Gg
Gs*
sİ
Gs*
Gs*
sK
Gg
Gs*
BronĢiyal örtü
epiteli altı
DK
Ds
Ds*
DK
Ds
Gg
DK*
Ds
Ds*
gK*
Ds
Ds*
BronĢiyal
kıkırdak arası
sK
Ds
Ds
sK
Ds
Gg
sİ
Ds
Ds*
sK
Ds
Ds*
Pessulus epitel
altı
Gg
Dİ
Gs
gK
sİ
Dİ
sİ
sİ
Dİ
Ds
Dİ
Gs
MTM örtü epiteli
altı
GK
Gs
Dİ
GK
Gs
Ds
GK
Gs
Ds
sK
Gs
Ds

18
4.2.1. Denizli Horozu
Kollagen Fibriller
Sirinksin trakeya ait bölümünde lamina epitelyalisin hemen altında dar bir alanda sıkı biçimde
yerleşim gösteren kollagen fibrillerin, bağ dokusu derinlerine doğru geniş bir alanda seyrek
biçimde düzenlendikleri; trakeyal kıkırdak halkalar arasında ise kalın demetler halinde sıkı
biçimde bulundukları belirlendi (Şekil 1).
Kollagen fibrillerin LTM epiteli altından bağ dokusu derinlerine doğru geniş bir alanda kalın
demetler halinde olduğu; özellikle bağ dokusunun derin kısımlarında çok daha sıkı biçimde
yerleşim gösterdikleri saptandı (Şekil 2).
Sirinksin bronşiyal kısmına ait epitel altında yerleşim gösteren kollagen fibrillerin trakeyal
kısımdaki fibrillere nazaran daha dar bir alanda daha sıkı biçimde düzenlendikleri, bronşiyal
kıkırdak halkalar arasında ise kalın demetler halinde bulundukları görüldü.
Pessulusun uç ve lateral kısmına ait epitel altında geniş bir alanda gevşek tarzda yerleşim
gösteren kollagen fibrillerin kemik dokuya doğru gidildikçe dar bir alanda sıkı demetler
halinde bulundukları tespit edildi.
MTM’ın bağ dokusunda bulunan kollagen fibrillerin dağılım ve düzenlemesinin LTM bağ
dokusunda bulunan fibrillerle dağılım ve yoğunluk olarak benzerlik gösterdiği saptandı.

19
Şekil 4. 1. Trakeyal sirinks. Submukoza (ince ok) ve kıkırdak halka arası (Kalın ok) kollagen
fibril. Van Gieson. Bar: 80 µm.
Şekil 4. 2. LTM. Epitel altı (ince ok) ve submukozada (Kalın ok) kollagen fibril. Van Gieson.
Bar: 80 µm.
Elastik fibriller
Sirinksin trakeyal kısmında epitel altından bağ dokusunun derinlerine doğru dağınık şekilde
yerleşim gösteren elastik fibrillerin geniş bir alanda gevşek tarzda düzenlendikleri, trakeyal
kıkırdak halkalar arasında ise daha sıkı ve kalın demetler oluşturdukları gözlendi.
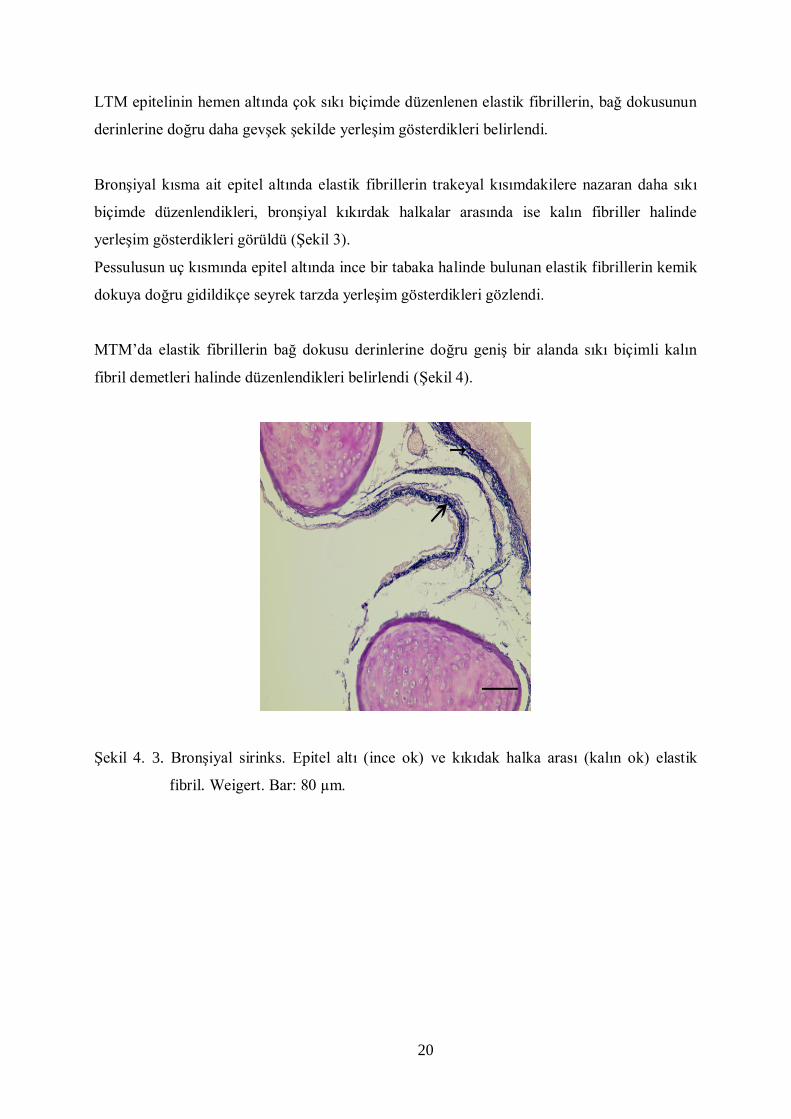
20
LTM epitelinin hemen altında çok sıkı biçimde düzenlenen elastik fibrillerin, bağ dokusunun
derinlerine doğru daha gevşek şekilde yerleşim gösterdikleri belirlendi.
Bronşiyal kısma ait epitel altında elastik fibrillerin trakeyal kısımdakilere nazaran daha sıkı
biçimde düzenlendikleri, bronşiyal kıkırdak halkalar arasında ise kalın fibriller halinde
yerleşim gösterdikleri görüldü (Şekil 3).
Pessulusun uç kısmında epitel altında ince bir tabaka halinde bulunan elastik fibrillerin kemik
dokuya doğru gidildikçe seyrek tarzda yerleşim gösterdikleri gözlendi.
MTM’da elastik fibrillerin bağ dokusu derinlerine doğru geniş bir alanda sıkı biçimli kalın
fibril demetleri halinde düzenlendikleri belirlendi (Şekil 4).
Şekil 4. 3. Bronşiyal sirinks. Epitel altı (ince ok) ve kıkıdak halka arası (kalın ok) elastik
fibril. Weigert. Bar: 80 µm.

21
Şekil 4. 4. MTM. Epitel altı (oklar) elastik fibril. Weigert. Bar: 80 µm.
Retikulum fibrilleri
Retikulum fibrillerinin sirinksin trakeyal kısmına ait epitel altında sıkı demetler halinde
yerleşim gösterdiği; bağ dokusunun daha derin kısımlarında gevşek şekilde düzenlendikleri
belirlendi.
Trakeyal kıkırdak halkalar arasında retikulum fibrillerinin ince demetler halinde seyrek
biçimde yerleşim gösterdikleri tespit edildi. Son üç trakeyal kıkırdak halka arasında ise
fibrillerin kalın demetler halinde sıkı biçimde düzenlendikleri gözlendi.
Retikulum fibrillerinin LTM’a ait epitelinin altında trakeye nazaran daha geniş bir alanda
gevşek tarzda yerleşim gösterdiği; bağ dokusu derinlerine doğru ise giderek sıkılaştığı ve
kalın demetler halinde düzenlendikleri saptandı (Şekil 5).
Sirinksin bronşiyal kısmına ait epitel altında dar bir alanda sıkı demetler halinde yerleşim
gösteren retikulum fibrillerinin kıkırdak halkalara doğru giderek seyrekleştiği belirlendi
(Şekil 6).

22
Pessulusun uç kısmına ait epitel altında dar bir alanda ince demetler halinde yerleşim gösteren
retikulum fibrillerinin pessulus lateralinde geniş bir alanda sıkı biçimde düzenlendikleri;
kemik dokuya doğru gidildikçe geniş bir alanda gevşek tarzda bulundukları tespit edildi.
Retikulum fibrillerinin MTM’a ait epitel altında geniş bir alanda ince demetler halinde
bulundukları; bağ doku derinlerine doğru giderek sıkılaşıp kalınlaştığı ve kalın demetler
halinde yerleşim gösterdikleri belirlendi.
Şekil 4. 5. LTM. Epitel altı (ince ok) ve sub mukozada (kalın ok) retikulum fibrilleri.
Gordon&Sweet. 60 µm.
Şekil 4. 6. Bronşiyal sirinks. Epitel altı (kalın ok) ve kıkırdak halka arası (ince ok) retiku lum
fibril. Gordon&Sweet. 60 µm.

23
4.2.2. Denizli tavuğu
Kollagen fibril
Sirinksin trakeyal kısmına ait epitel altında dar bir alanda demetler halinde yerleşim gösteren
kollagen fibrillerin trakeyal kıkırdak halkalara doğru gidildikçe gevşek tarzda düzenlendikleri;
trakeyal kıkırdak halkalar arasında ise geniş bir alanda gevşek şekilde bulundukları belirlendi.
LTM’a ait epitel altında ve bronşiyal kıkırdak halkalar arasında (Şekil 7) kollagen fibrillerin
trakeyal kıkırdak halkalara nazaran kalın demetler halinde sıkı biçimde düzenlendikleri
saptandı.
Pessulusun uç kısmına ait epitel altında düzgün bir yapılanma ve kalın demetler halinde
yerleşim gösteren fibrillerin pessulus laterali epitelinden kemik dokuya doğru gidildikçe
gevşek tarzda düzenlendikleri görüldü.
Pessulusun MTM’a doğru devam eden kısmında ise fibrillerin dar bir alanda çok sıkı biçimde
düzenlenmiş oldukları saptandı (Şekil 8).
Bağ doku derinlerinde geniş bir alanda kalın demetler halinde yerleşim gösteren fibrillerin
MTM’nin devam eden derin kısımlarında yoğunluk olarak arttığı belirlendi.
Şekil 4. 7. Bronşiyal sirinks. Kıkırdak halka arası (oklar) kollagen fibril.Van Gieson. Bar: 80
µm.

24
Şekil 4. 8. MTM. Bağ dokusu içeri sinde kollagen fibriller (oklar). Van Gieson. Bar:80 µm
Elastik fibril
Sirinksin trakeyal kısmına ait epitelinin altında geniş bir alanda fibriler tarzda yerleşim
gösteren elastik fibrillerin bağ doku derinlerine doğru ve trakeyal kıkırdak halkalar arasında
gevşek şekilde düzenlendikleri gözlendi. Son üç trakeyal kıkırdak halkalar arasında ise geniş
bir alanda çok sıkı biçimde yerleşim gösterdikleri belirlendi.
LTM’ a ait epitel altında kollagen fibrillere nazaran daha geniş bir alanda gevşek tarzda
yerleşim gösteren elastik fibrillerin bağ dokusu derinlerine doğru sıkı biçimde düzenlenip
helezonlaştığı gözlendi (Şekil 9).
Sirinksin bronşiyal kısmına ait epitel altında LTM’na nazaran daha dar bir alanda çok sıkı
biçimde yerleşim gösteren elastik fibrillerin bronşiyal kıkırdak halkalar arasında sıkı biçimli
fibril demetleri halinde bulundukları tespit edildi.
Pessulusun uç ve lateral kısmında sıkı biçimde yerleşim gösteren fibrillerin kemik dokuya
doğru gidildikçe yoğunlaştığı ve gevşek tarzda düzenlendikleri belirlendi.

25
MTM’a ait epitel altında kollagen fibrillere nazaran daha geniş bir alanda kalın fibriller
şeklinde yerleşim gösteren elastik fibrillerin kas tabakasına doğru daha yoğun biçimde
düzenlendikleri gözlendi (Şekil 10).
Şekil 4. 9. LTM. Epitel altı (oklar) elastik fibriller. Weigert. Bar: 60 µm.
Şekil 4. 10. MTM. Epitel altı (*) ve submu kozada (oklar) elastik fibriller. Weigert. Bar:80
µm.

26
Retikulum fibrilleri
Trakeyal kısma ait epitel altında ve trakeyal kıkırdak halkalar arasında retikulum fibrillerin
geniş bir alanda gevşek şekilde düzenlendikleri; son üç trakeyal kıkırdak halkalar arasında ise
kalın demetler halinde sıkı biçimde yerleşim gösterdikleri gözlendi.
LTM’a ait epitel altından bağ doku derinlerine doğru retikulum fibrillerinin geniş bir alanda
sıkı biçimde düzenlendikleri görüldü.
Sirinksin bronşiyal bölümüne ait epitelin altında ve bronşiyal kıkırdak halkalar arasında
fibrillerin dar bir alanda sıkı biçimli fibril demetleri halinde yerleşim gösterdikleri belirlendi
(Şekil 11).
Pessulusun uç ve lateral kısmına ait epitel altında dar bir alanda ince demetler halinde bulunan
retikulum fibrillerin kemik dokuya doğru gidildikçe yoğunluklarının arttığı görüldü.
MTM’a ait epitel altında retikulum fibrillerinin dar bir alanda ince demetler halinde yerleşim
gösterdikleri; membranın derin kısımlarına doğru ise yoğunluklarının azaldığı gözlendi
(Şekil 12).
Şekil 4. 11. Bronşiyal sirinks. Kıkırdak halka arası (oklar) retikulum fibrilleri. Gordon &
Sweet. 80 µm.

27
Şekil 4. 12. MTM. Epitel altı (*) ve submukoza (oklar) elastik fibriller. Gordon&Sweet.
Bar:60 µm.
4.2.3. Erkek sülün
Kollagen fibril
Trake epitelin altında dar bir alanda ince demetler halinde yerleşim gösteren kollagen
fibrillerin trakeyal kıkırdak halkalar arasında kalın demetler halinde sıkılaşıp helezonlaştığı
gözlendi.
LTM epitelinin altında, bronşiyal kıkırdak halkalar arasında (Şekil 13) ve pessulus
lateralindeki epitel altında kollagen fibrillerin sıkı biçimde bir araya gelerek ince demetler
oluşturduğu gözlendi.
MTM’a ait epitel altında ise fibrillerin geniş bir alanda kalın demetler halinde bulundukları ve
oldukça sıkı biçimde düzenlendikleri gözlendi (Şekil 14).

28
Şekil 4. 13. Bronşiyal sirinks. Kıkırdak halka arası (oklar) kollagen fibril. Van Gieson. Bar:80
µm.
Şekil 4. 14. MTM. Submukozada (oklar) kollagen fibril. Van Gieson. Bar: 60 µm.
Elastik fibril
Trakeyal kısma ait epitelin hemen altında ve trakeyal kıkırdak halkalar arasında elastik
fibrillerin sıkı biçimde yerleşim gösterdiği belirlendi (Şekil 15).
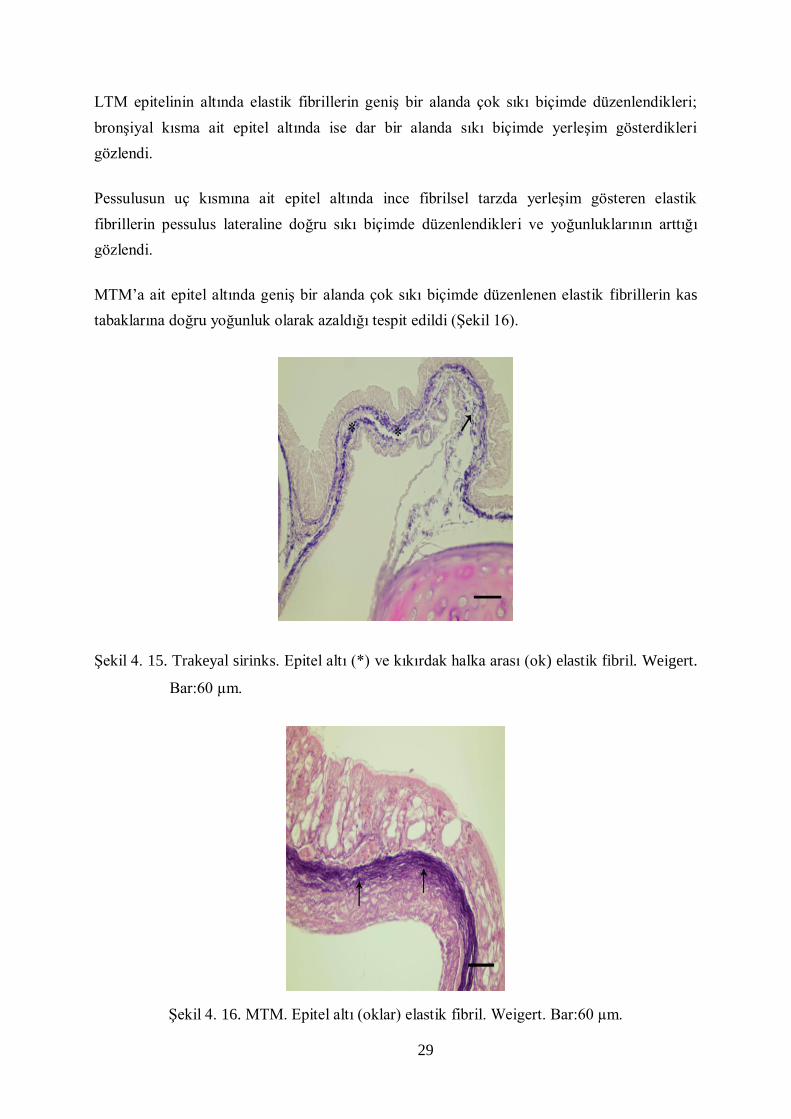
29
LTM epitelinin altında elastik fibrillerin geniş bir alanda çok sıkı biçimde düzenlendikleri;
bronşiyal kısma ait epitel altında ise dar bir alanda sıkı biçimde yerleşim gösterdikleri
gözlendi.
Pessulusun uç kısmına ait epitel altında ince fibrilsel tarzda yerleşim gösteren elastik
fibrillerin pessulus lateraline doğru sıkı biçimde düzenlendikleri ve yoğunluklarının arttığı
gözlendi.
MTM’a ait epitel altında geniş bir alanda çok sıkı biçimde düzenlenen elastik fibrillerin kas
tabaklarına doğru yoğunluk olarak azaldığı tespit edildi (Şekil 16).
Şekil 4. 15. Trakeyal sirinks. Epitel altı (*) ve kıkırdak halka arası (ok) elastik fibril. Weigert.
Bar:60 µm.
Şekil 4. 16. MTM. Epitel altı (oklar) elastik fibril. Weigert. Bar:60 µm.

30
Retikulum fibrilleri
Retikulum fibrillerinin sirinksin trakeyal kısmına ait epitel altında ve trakeyal kıkırdak
halkalar arasında dar bir alanda gevşek tarzda düzenlendikleri gözlendi.
LTM’a ait epitel altında fibrillerin çok sıkı biçimde düzenlendikleri; bronşiyal kıkırdak
halkalara doğru ise geniş bir alanda gevşek şekilde yerleşim gösterdikleri saptandı.
Sirinksin bronşiyal kısmına ait epitel altında geniş bir alanda çok sıkı biçimde yerleşim
gösteren retikulum fibrillerinin; pessulusun lateral kısımlarında dar bir alanda sıkı biçimde
düzenlendikleri belirlendi (Şekil 17).
MTM’nin başlangıcına ait epitel altında dar bir alanda sıkı biçimde yerleşim gösteren
fibrillerin (Şekil 18) membranın derinlerine doğru devam eden kas tabakasına doğru geniş bir
alanda oldukça sıkı biçimde uzun fibril demetleri halinde bulundukları görüldü.
Şekil 4. 17. Bronşiyal sirinks. Kıkırdak halka halka arası(oklar) retikulum fibrilleri. Gordon &
Sweetµm. Bar: 60 µm.

31
Şekil 4. 18. MTM. Epitel altı (oklar) retikulum fibrilleri. Gordon&Sweet. Bar: 80 µm.
4.2.4. DiĢi sülün
Kollagen fibril
Trakeyal epitelin hemen altında dar bir alanda ince demetler halinde yerleşim gösteren
kollagen fibrillerin, trakeyal kıkırdak halkalar arasında kalın demetler halinde sıkı biçimde
düzenlendikleri belirlendi.
LTM’a ait epitel altında oldukça sıkı ve kalın demetler halinde yerleşim gösteren fibrillerin
(Şekil 19) bronşiyal kıkırdak halkalara doğru demetler halinde gevşek tarzda yerleşim
gösterdiği gözlendi.
Bronşiyal kıkırdak halkalar arasında yer alan kollagen fibrillerin trakeyal kıkırdak halkalar
arasındakilere nazaran kalın demetler halinde sıkı biçimde düzenlendikleri görüldü.
Pessulusun uç ve lateral bölgelerine ait epiteller altında dar bir alanda sıkı biçimde
düzenlenmiş olan kollagen fibrillerin submukoza doğru ve MTM epiteli altında (Şekil 20)
kalın demetler halinde daha sıkı biçimde düzenlendikleri tespit edildi.

32
Şekil 4. 19. LTM. Epitel altı (oklar) kollagen fibrilleri. Van Gieson. Bar: 60 µm.
Şekil 4. 20. MTM. Epitel altı (oklar) kollagen fibriller. Van Gieson. Bar: 60 µm.
Elastik fibril
Sirinksin trakeyal kısmına ait epitel altında ince fibriler tarzda yerleşim gösteren elastik
fibrillerin bağ doku derinlerine doğru geniş bir alanda seyrek şekilde yerleşim gösterdiği
gözlendi.

33
Trakeyal kıkırdak halkalar arasında ise elastik fibrillerin daha sıkı biçimde düzenlendikleri
belirlendi.
LTM epitelinin hemen altında yerleşim gösteren elastik fibrillerin gevşek şekilde
düzenlendikleri ve helezonlaşma gösterdikleri tespit edildi.
Bronşiyal kıkırdak halkalar arasında ise elastik fibrillerin trakeyal kıkırdak halkalara nazaran
dar bir alanda sıkı biçimde düzenlendikleri görüldü (Şekil 21).
Pessulusun uç kısmına ait epitel altında sıkı biçimde yerleşim gösteren elastik fibrilerin
pessulus lateraline doğru seyrekleştiği ve MTM’na doğru ise sıkılaşıp kalınlaştığı belirlendi.
MTM’a ait epitel altından kas tabakalarına doğru elastik fibrillerin yoğunlaştığı ve kollagen
fibrillere nazaran daha seyrek şekilde yerleşim gösterdiği saptandı (Şekil 22).
Şekil 4. 21. Bronşiyal sirinks. Epitel altı (*) ve kıkırdak halka arası (ok) elastik fibriller.
Weigert. Bar: 80 µm.

34
Şekil 4. 22. MTM. Submukozada elastik fibriller (oklar). Weigert. Bar:60 µm.
Retikulum fibriller
Retikulum fibrillerinin trakeyal kısma ait epitel altında dar bir alanda ince fibril demetleri
halinde bulundukları görüldü.
Trakeyal kıkırdak halkalar arası alanda ve LTM’a ait epitel altında ise retikulum fibrillerinin
geniş bir alanda sıkı biçimde düzenlenmiş fibril demetleri halinde bulundukları gözlendi
(Şekil 23)
Sirinksin bronşiyal kısmına ait retikulum fibril dağılım ve düzenlenmesinin erkek sülündeki
bulgularla benzer olduğu tespit edildi (Şekil 24).
Pessulusun uç ve lateral kısmına ait epitel altında belirgin bir şekilde gözlenemeyen retikulum
fibrillerinin kemik dokunun bitiminden MTM’a doğru devam eden bölgede sıkı biçimde
düzenlendikleri belirlendi.
MTM epitelinin altında dar bir alanda sıkı biçimde yerleşim gösteren retikulum fibrillerinin
membranın derinlerine doğru geniş bir alanda yerleşim gösterdikleri tespit edildi.

35
Şekil 4. 23. LTM. Submukoza (oklar) retikulum fibrilleri. Gordon&Sweet. Bar: 60 µm.
Şekil 4. 24. Bronşiyal sirinks. Epittel altı (*) ve kıkırdak halka arası (ok) retikulum fibrilleri.
Gordon & Sweet. Bar:80 µm.

36
5. TARTIġMA VE SONUÇ
Kanatlılarda ses çıkarmayı sağlayan sirinksin morfolojik yapısı bıldırcın (Bayram ve Liman,
2000), devekuşu (Yıldız vd., 2003), güvercin (Yıldız vd., 2005), şahin (Kabak vd., 2007), kaz
(Onuk vd., 2010), baykuş (Çevik-Demirkan ve Özdemir, 2011) ve martı (Gezer İnce vd.,
2012) türlerinde araştırılmıştır.
Bu çalışmadan elde edilen bulgularla benzer şekilde penguen (Taşbaş vd., 1986), Denizli
horozu (Taşbaş vd., 1994), bıldırcın (Bayram ve Liman, 2000; Çevik-Demirkan vd., 2007),
devekuşu (Yıldız vd., 2003), güvercin (Yıldız vd., 2005), yeşilbaş ördek (Frank et al., 2006;
2007), şahin (Kabak vd., 2007), kaz (Onuk vd., 2010), baykuş (Çevik-Demirkan ve Özdemir,
2011) ve martılarda (Gezer İnce vd., 2012), trakeyal ve bronşiyal kıkırdakların her ikisi de
sirinks oluşumuna katıldığı için, sirinksin trakeyobronşiyal tipte olduğu bildirilmiştir.
Sirinksin penguende (Taşbaş vd., 1986) son 3 trakeyal, ilk 2 bronşiyal; bıldırcında son 2
trakeyal, ilk 2 bronşiyal (Bayram ve Liman, 2000) ve son 2-3 trakeyal, ilk 2-3 bronşiyal
(Çevik-Demirkan vd., 2007); devekuşunda (Yıldız vd., 2003) son 4 trakeyal, ilk 3 bronşiyal;
güvercinde (Yıldız vd., 2005) son 4 trakeyal, ilk 5 bronşiyal; şahinde (Kabak vd., 2007) son
5-6 trakeyal, ilk 4 bronşiyal; kazda (Onuk vd., 2010) son 4 trakeyal, ilk 6 bronşiyal; baykuşta
(Çevik-Demirkan ve Özdemir, 2011) son 4-5 trakeyal, ilk 2 bronşiyal ve martıda (Gezer İnce
vd., 2012) ise son 5 trakeyal ile ilk 7 bronşiyal kıkırdaktan oluştuğu bildirilmiştir. Bu
çalışmada Denizli horozu, Denizli tavuğu ve dişi sülünlerde sirinksin yapısına son 1 trakeyal
halka ile ilk 5 bronşiyal siringeyal kıkırdak halka katılırken; erkek sülünlerde son 1 trakeyal
ve ilk 3 bronşiyal kıkırdak halkanın katıldığı belirlendi.
Timpanumu şekillendiren trakeyal halka sayısının tavukgillerde (Getty, 1975) 3-4, kızıl
şahinde (Kabak vd., 2006) ve devekuşunda (Yıldız vd., 2003) ise son 3 adet kıkırdaktan
oluştuğu bildirilmiştir. Bu çalışmada ise Denizli horozu ve Denizli tavuğunda timpanumun
son dört trakeyal halkadan oluştuğu belirlenirken; dişi ve erkek sülünlerde son 3-4 trakeyal
halkadan oluştuğu tespit edildi.
Sirinksin bir bölümünü oluşturan pessulusun evcil kanatlılarda kama şeklinde bir kıkırdak
(Taşbaş vd., 1994), serçegillerde üçgen şeklinde olduğu ve tarla kuşunda (King ve Mclelland,
1984) bulunmadığı bildirilmesine karşın; bu çalışmada Denizli horozu ve tavuğunda

37
pessulusun üçgen şekilde kemik yapısında olduğu, sülünlerde ise yapının oval şekilli hiyalin
kıkırdak yapısında olduğu bildirilmiştir.
Ancak Bayram ve Liman (2000) bıldırcında başlangıçta hiyalin kıkırdak karakterinde olan
pessulusta yumurtadan çıkışın 42. gününden itibaren kemik doku odakları oluştuğunu
bildirmişlerdir. Ayrıca yeşilbaş ördek (Frank vd., 2006; 2007) sirinksinde pessulusun sadece
uç kısmının hiyalin kıkırdaktan, diğer kısımlarının kemik dokusundan oluştuğu belirtilirken;
devekuşu (Yıldız vd., 2003) sirinksinde pessulusun kıkırdak veya kemik doku içermediği
bildirilmiştir.
Şahin (Kabak vd., 2007), kaz (Onuk vd., 2010) ve martı (Gezer İnce vd., 2012) sirinkslerinde
yapılan çalışmalarda pessulus ile ligamentum interbronchiale arasında foramen
interbronchiale bulunduğu belirlendi. Bu çalışmada Denizli horozu, Denizli tavuğu ve erkek
sülünlerin sirinkslerinde sağ ve sol bronşlar arasında ligamentum interbronchiale’nin
bulunduğu gözlenirken, dişi sülün sirinksinde bu yapının bulunmadığı saptandı.
Şahin (Kabak vd., 2007) ve martıda (Gezer İnce vd., 2012) LTM’ın birinci ve ikinci bronşiyal
siringeyal kıkırdaklar arasında yer aldığı bildirilmiştir. Bıldırcın (Bayram ve Liman, 2000;
Çevik-Demirkan vd., 2007), güvercin (Yıldız vd, 2005) ve baykuşta (Çevik-Demirkan ve
Özdemir, 2011) ise LTM’nin son trakeyal kıkırdak halka ile ilk bronşiyal kıkırdak halka
arasında yer aldığı belirtilmiştir. Bu çalışmada da benzer bulgular elde edildi.
MTM’ın şahinde (Kabak vd, 2007) pessulusun kaudali ile bronşiyal kıkırdaklar arasında,
baykuşta (Çevik-Demirkan ve Özdemir, 2011) ilk trakeyal siringeyal kıkırdak halka ile son
bronşiyal siringeyal kıkırdak halka arasında, kazda (Onuk vd., 2010) pessulus ile 2. bronşiyal
kıkırdak arasında ve martıda (Gezer İnce vd., 2012) bronşiyal kıkırdakların mediyalinde
bulunduğu bildirilmiştir. Bununla birlikte Bayram ve Liman (2000) bıldırcın sirinksinde
MTM’ın pessulusun yan taraflarından başlayıp 1. bronşiyal kıkırdak halkanın serbest ucuna
kadar uzandığını bildirmelerine karşın; Çevik-Demirkan vd. (2007) pessulusun kaudalinde
başlayıp, primer bronkusları çevrelediğini belirtmişlerdir. Bu çalışmada ise MTM’nin
pessulusun kaudali ile ligamentum interbronchiale arasında yer aldığı tespit edildi.
Bu çalışmada elde edilen bulgulara benzer olarak trakeyal sirinksin devekuşu (Yıldız vd.,
2003) ve bıldırcında (Bayram ve Liman, 2000) yalancı çok katlı prizmatik epitel ile örtülü

38
olduğunun bildirilmesine karşın; güvercinde (Yıldız vd., 2005) bu epitelin çok katlı yassı non-
keratinize karaktere sahip olduğu belirtilmiştir.
Bayram ve Liman (2000) LTM epitelinin yalancı çok katlı prizmatik örtü epiteli ile başlayıp,
tek katlı kübik örtü epiteline dönüştüğünü bildirmişlerdir. Bu çalışmada Denizli horozu,
Denizli tavuğu ve erkek sülünlerde LTM epitelinin yalancı çok katlı prizmatik epitelle örtülü
olduğu, son 2 trakeyal kıkırdak halka hizasından itibaren çok katlı yassı karakter kazanan
epitelin, birinci bronşiyal kıkırdak halka hizasında tek katlı yassı örtü epiteline dönüştüğü
belirlenirken; dişi sülünde LTM’nin trake tarafında bulunan epitelin yalancı çok katlı
prizmatik epitelle örtülü olduğu, birinci ve ikinci bronşiyal kıkırdak halkalar arasında tek
katlı yassı, ikinci bronşiyal halka hizasında çok katlı yassı karakterde bulunan epitelin üçüncü
bronşiyal kıkırdak halkadan itibaren tekrar yalancı çok katlı epitele dönüştüğü belirlendi.
Aynı araştırıcılar (Bayram ve Liman, 2000) MTM’ın çok katlı kübik epitele sahip olduğunu
bildirirlerken; pekin ördeklerinde (Scala vd., 1990) bu epitelin yalancı çok katlı prizmatik
epitelden oluştuğunu belirtmişlerdir. Bu çalışmada da MTM epitelinin Denizli horozu ve
Denizli tavuğunda pessulus bitiminden birinci bronşiyal kıkırdak halka hizasına kadar yalancı
çok katlı prizmatik, birinci bronşiyal kıkırdak halka ile ikinci kıkırdak halka arasında çok katlı
yassı karakterde olduğu; ikinci bronşiyal siringeyal kıkırdak halkadan itibaren tekrar yalancı
çok katlı prizmatik epitele dönüştüğü belirlenirken; dişi ve erkek sülünlerde epitelin pessulus
bitiminden birinci bronşiyal kıkırdak halka hizasına kadar çok katlı yassı karakterde olduğu;
birinci bronşiyal kıkırdak halka hizasından itibaren tekrar yalancı çok katlı prizmatik epitele
dönüştüğü belirlendi.
Devekuşu (Yıldız vd., 2003), güvercin (Yıldız vd., 2005) ve bıldırcın (Çevik-Demirkan vd.,
2007) trakeyal sirinksinde lamina propria ve submukozanın gevşek bağ dokusu karakterinde
olduğu bildirilmiştir. Bu çalışmada Denizli horozu sirinksinin trakeyal kısmının gevşek bağ
dokusu, trakeyal kıkırdak halkalar arasının ise sıkı bağ dokusu yapısında olduğu tespit edildi.
Bu çalışmada da elde edilen bulgulara benzer şekilde devekuşu (Yıldız vd., 2003) ve
güvercinde (Yıldız vd., 2005) elastik fibrillerin submukozada, bağ dokunun derinlerinde ve
trakeyal kıkırdak halkalar arasında bulunduğu belirtilmiştir.

39
Ayrıca Denizli tavuğunda sirinksin trakeyal kısmına ait epitel altında geniş bir alanda fibrilsel
tarzda yerleşim gösteren elastik fibrillerin bağ doku derinlerine doğru ve trakeyal kıkırdak
halkalar arasında gevşek şekilde düzenlendikleri gözlendi. Son üç trakeyal kıkırdak halkalar
arasında ise fibrillerin geniş bir alanda çok sıkı biçimde yerleşim gösterdikleri belirlendi.
Frank vd (2006) elastik fibrillerin yeşilbaş ördekte LTM örtü epiteli altında gevşek dağılım
gösterdiklerini, MTM epiteli altında ise sıkı ağlar oluşturduklarını bildirmişlerdir. Bu
çalışmada Denizli horozunda LTM epitelinin hemen altında çok sıkı biçimde düzenlenen
elastik fibrillerin, submukozaya doğru daha gevşek yerleşim gösterdikleri belirlendi.
MTM’da elastik fibrillerin submukozaya doğru geniş bir alanda sıkı biçimli kalın fibril
demetleri halinde düzenlendikleri belirlendi.
Denizli tavuğunda LTM ve MTM’na ait epitel altında daha geniş bir alanda gevşek tarzda
yerleşim gösteren elastik fibrillerin bağ dokusu derinlerine doğru dalgalı bir görünüm
oluşturdukları, erkek sülünde ise elastik fibrillerin geniş bir alanda çok sıkı biçimde
düzenlendikleri; bronşiyal kısma ait epitel altında ise dar bir alanda sıkı biçimde yerleşim
gösterdikleri gözlendi. Dişi sülünlerde ise LTM epitelinin hemen altında yerleşim gösteren
elastik fibrillerin erkek sülündeki fibrillere nazaran daha gevşek düzenlendikleri ve
helezonlaşma gösterdikleri tespit edildi.
Bayram ve Liman (2000) bıldırcında yumurtadan çıkışın 21. gününden itibaren LTM örtü
epiteli altında kollagen fibrillerin demetler halinde düzenlendiğini belirtmişlerdir. Bu
çalışmada Denizli horozu, Denizli tavuğu ve dişi sülünlerde kollagen fibrillerin LTM epiteli
altından bağ dokusu derinlerine doğru geniş bir alanda kalın demetler halinde olduğu;
özellikle submukozada sıkı ve kalın demetler halinde yerleşim gösterdikleri saptanırken;
erkek sülünlerde LTM’a ait örtü epiteli altında kollagen fibrillerin sıkı biçimde düzenlenmiş
ince demetler oluşturduğu gözlendi.
Serçe sirinksine ait LTM ve MTM epiteli altında kollagen fibrillerin gevşek şekilde yerleşim
gösterdikleri bildirilmiştir (Riede ve Goller, 2009). Bu çalışmada da erkek ve dişi sülünde
MTM’a ait epitel altında kollagen fibrillerin geniş bir alanda kalın demetler halinde
bulundukları ve oldukça sıkı biçimde düzenlendikleri gözlendi.

40
Onuk vd. (2010) kaz sirinksinde bronşiyal kıkırdak halkalar arasında ve kıkırdaklara komşu
bağ dokusunda kollagen fibrillerin bulunduğunu bildirmişlerdir. Bu çalışmada da benzer
bulgular belirlendi.
Denizli horozu ve Denizli tavuğu türlerinin sirinksleri karşılaştırıldığında kollagen, elastik ve
retikulum fibrillerinin dağılım ve yoğunluğunun benzer şekilde olduğu görüldü. Ancak erkek
sülünlere ait sirinks bölümlerinde fibril dağılım ve yoğunluklarının dişi sülünlerdekine
nazaran daha geniş bir alanda daha sıkı biçimde düzenlendikleri tespit edildi.
Sirinksin histolojik yapısı az sayıda kuş türünde incelenmiş olup, bağ dokusunun tüm
fibrillerinin bir arada araştırıldığı çalışmaya rastlanmamıştır. Denizli horozu, Denizli tavuğu,
dişi ve erkek sülün sirinkslerinin bazı özelliklerinin ortaya konmasına yönelik yapılan bu
çalışmadan elde edilen bulgulara göre; sirinksin histolojik yapısının, özellikle trakeyal sirinks
mukozasının bıldırcın (Bayram ve Liman, 2000; Çevik-Demirkan vd., 2007) ve devekuşu
(Yıldız vd., 2003) ile benzerlik gösterdiği belirlenirken, diğer kuş türlerinden farklılıkların
olduğu saptandı. Bununla birlikte sirinkste kollagen ve elastik fibrillerin dağılımının devekuşu
(Yıldız vd., 2003), güvercin ( Yıldız vd., 2005) ve kaz (Onuk vd., 2010) türlerinde benzerlik
gösterdiği sonucuna varıldı.

41
6. KAYNAKLAR
Akay, T., 2006. Genel Histoloji. 6. Baskı, Palme Yayıncılık, 47-75, Ankara.
Ayvaz, A., 2010. İnsan Fizyolojisi. (Aktümsek, A., ed.) Nobel Yayın Dağıtım, Bölüm E.
115-148, Ankara.
Baumel, J. J., King, A. S., Breazile, J. E., Evans, H. E., Vanden Berge, J. C., 1993. Nomina
Anatomica Avium. Publications of The Nuttall Ornithological, 23, Cambridge.
Bayram, G., Liman, N., 2000. Bıldırcınlarda sirinksin postnatal gelişimi üzerine morfolojik
araştırmalar. Turkısh Journal of Veterinary and Animal Sciences, 24, 381-392.
Bell, D.J., Freeman, B.M., 1971. Physiology and Biochemistry of the Domestic Fowl.
Academic Pres, 101-121, London, New York.
Cireli, E., 1999. Genel Histoloji. 6. Baskı, Palme Yayıncılık, 183-199, İzmir.
Culling, C.F.A., Reid, and Dunn, W.L., 1976. A new histochemical method for the
identification and visualization of both side chain acylated and non-acylatedsialic
acids. Journal of Histochemistry and Cytochemistry, 24, 1225-1230.
Çevik-Demirkan, A., Hazıroğlu, RM., Kürtül, I., 2007. Gross morphological and histological
features of larynx, trachea and syrinx in japanese quail. Anatomia Histologia
Embryologia, 36, 215-219.
Çevik-Demirkan, A., Özdemir, V., 2011. A study on anatomical structures of the larynx,
trachea and syrinx in Eurasion eagle owl (Bubo bubo). Journal of Animal and
Veterinary Advances 10 (24), 3218-3224.
Demirsoy, A., 1995. Yaşamın Temel Kuralları- Omurgalılar/Amniyota, Meteksan A.Ş., Cilt
III/Kısım II, 199- 228, Ankara.
Doğuer, S., Erençin, Z., 1964. Evcil Kuşların Komparativ Anatomisi. Ankara Üniversitesi
Yayınları, 45-60, Ankara.
Dursun, N., 2002. Evcil Kuşların Anatomisi, Medisan Yayınevi, 1. baskı, 93-95, Ankara.
Erdost H., 2011. Veteriner Özel Histoloji. (Özer, A., ed) Nobel Yayıncılık, 3. Baskı, 145-147,
Ankara.
Erkoçak, A. 1982. Özel Histoloji. Ankara Üniversitesi Tıp Fakültesi Yayınları, 4. Baskı, 280
s. Ankara.
Eşrefoğlu, M., 2009. Genel Histoloji, Medipres Matbaacılık Yayıncılık, 131-135, Malatya.
Fedde, M.R., 1998. Relationship of structure and function of the avian respiratory system to
disease susceptibility. Poultry Science, 77, 1130-1138.

42
Frank, T., Probst, A., König, H.E., Walter, I., 2007. The syrinx of the male mallard (Anas
platyrhynchos): Speecial anatomical features. Anatomia Histologia Embryologia, 36,
121-126.
Frank, T., Walter, I., Probst, A., König, H.E., 2006. Histological aspect of the syrinx of the
male mallard (Anas platyrhynchos). Anatomia Histologia Embryologia, 35, 396-401.
Gaunt, A.S., Gaunt, L.L., 1977. Mechanics of the syrinx in Gallus gallus. II.
Electromyographic studies of ad libitum vocalizations. Journal of Morphology, 152
(1), 1-19.
Getty, R., 1975. Sison and Grossman’s. The Anatomy of the Domestic Animals. W.B
Saunders Company, Philadelphia.
Gezer İnce, N., Pazvant, G., Alpak, H., 2012. Anatomical features of the syrinx in sea gulls.
Ankara Üniversitesi Veteriner Fakültesi Dergisi, 59, 1-3.
Goller, F., Suthers, R.A., 1996. Role of syringeal muscles in gating airflow and sound
production in singing brown thrashers. Journal of Neurophysiol 75, 867–76.
Gordon, H.& Sweets, H.H., 1936. A simple method for the silver impregnation of reticulum.
American Journal of Pathology, 12, 545.
Hodges, R. D., 1974. The Histology of the Fowl. Academic Press, 113-149, London.
Junqueria, L.C., Carnerio, J., 2009. Temel Histoloji (Solakoğlu, S., Aytekin, Y., ed) 1. Baskı,
Nobel Tıp Kitabevleri. 91- 12, İstanbul.
Junqueria, L.C., Carnerio, J., Kelley, R.O., 1998. Temel Histoloji. (Aytekin, Y., Solakoğlu,
S., Ahıshalı, B., ed). Beta A.Ş. Yayıncılık. 88-102, İstanbul.
Kabak, M., Orhan, I.O., Hazıroğlu, R.M., 2007. The gross anatomy of larynx, trachea and
syrinx in the long-legged buzzard (Buteo rufinus). Anatomia Histologia Embryologia,
36, 27-32.
King, A.S., 1989. Functional anatomy of the syrinx. In: Form and Function in Birds. London:
Academic Press, pp. 105-182.
King, A.S., McLelland, J., 1984. Birds their Structure and Function. Second Edition, Bailliere
Tindall, London, Philadelphia, Toronto, Mexico City, Rio De Janeiro, 121- 130,
Sydney, Tokyo, Hong Kong.
Kuru, M., 2009. Omurgalı Hayvanlar. Palme Yayıncılık, 9. Baskı, 425-454, Ankara.
Larsen, O.N., Goller, F., 2002. Direct observation of syringeal muscle function in songbirds
and a parrot. The Journal of Experimental Biology, 205, 25-35.
Lockner, F. R., Youngren, O. M., 1976. Functional syringeal anatomy of the mallard. In situ
electromyograms during ESB elicitated calling. American University in Kosova, 93,
324–342.

43
Marieb, E.N., 2001. Human Anatomy & Physiology. Fifth edition, Addison Wesley,
Longman, pp. 1249. USA.
Martini, F.H., Timmans, M.J., McKinley, M.P., 2000. Human Anatomy. Third edition,
prentice Hall, pp. 866, New Jersey.
Myers, A. J., 1917. Studies of the syrinx of Gallus domesticus. Journal of Morphology. 29,
165–215.
Noyan, A., 1993. Kuşlarda Solunum. Yaşam ve Hekimlikte Fizyoloji, Meteksan A.Ş, 10.
baskı, 581-588, Ankara.
Onuk, B., Hazıroğlu, R.M., Kabak, M., 2010. The gross anatomy of larynx, trachea and syrinx
in goose (Anser anser domesticus). Kafkas Üniversitesi Veteriner Fakültesi Dergisi 16,
443-450.
Öber, A., İzzetoğlu G.T., 2006. Histoloji. Nobel Yayın Dağıtım. 55- 74, Ankara
Özcan, A., 2011. Veteriner Özel Histoloji. (Özer, A., ed) Nobel Yayıncılık, 3. Baskı, 135-
136, Ankara.
Piperno, E., Peirone, S. 1975. Morpholojical characteristics and mutual relatinships of the
tracheal cartilaginous rings in Gallus Gallus. Anatomia Histologia Embryologia, 4,
172-178.
Riede, T., Goller, F., 2010. Functional morphology of the sound-generating labia in the syrinx
of the two songbird species. Journal of Anatomy, 216, 23-36.
Rodentio, Ö.M., 1988. Omurgalı Hayvanların Karşılaştırmalı Anatomisi. İstanbul Üniversitesi
Basımevi, 215-217.
Scala, G., Corono, M., Pelagalli, G.V., 1990. The structure of the syrinx in the duck (Anas
platyrhinchos). Anatomia Histologia Embryologia, 19 (2), 135-42.
Tanyolaç, A., 1999. Özel Histoloji, Yorum Basın Yayın Sanayi Ltd. Şti., 213, Ankara.
Taşbaş, M., Hazıroğlu, R.M, Çakır, A., Özer, M., 1994. Denizli horozunun solunum
sisteminin morfolojisi. II. Larynx, trachea, syrinx. Ankara Üniversitesi Veteriner
Fakültesi Dergisi, 41, 135-153.
Taşbaş, M., Özcan, Z., Hazıroğlu, M., 1986. Penguenin dili ve ön solunum yollarının (larynx
cranialis, trachea, syrinx) anatomik ve histolojik yapısı üzerinde bir çalışma. Ankara
Üniversitesi Veteriner Fakültesi Dergisi, 33, 240-261.
Van Gieson, I., 1899. Laboratory notes of technical methods for the nervous system. New
York Medical Journal. 50,57.
Weigert, C., 1898. Ueber enine methode zur Farbung elastischer Fasern. Zentrablatt für
Allagemeine Pathologie und Pathologische Anatomie, 9-289.

44
Yaman, K., 1999. Fizyoloji. VİPAŞ A.Ş., 3. Baskı, 564, Bursa.
Yıldız, H., Bahadır, A., Akkoç, A., 2003. A study on the morphological structure of syrinx in
ostriches (Struthio camelus). Anatomia Histologia Embryologia, 32, 187-191.
Yıldız, H., Yılmaz, B., Arıcan, I., 2005. Morhological structure of the syrinx in the Bursa
roller pigeon (Columba lıvıa). Bulletin of the Veterinary Instıtute Pulawy, 49, 323-
327.
Zık, B., 2011. Veteriner Özel Histoloji. (Özer, A., ed) Nobel Yayıncılık, 3. Baskı, 137-139,
Ankara.

45
ÖZGEÇMĠġ
Adı Soyadı : Öznur ÖNAL
Doğum Yeri ve Yılı: Antalya, 1989
Medeni Hali : Bekar
Yabancı Dili : İngilizce
E-posta : [email protected]
Eğitim Durumu
Lise : 2003-2006 Antalya Kepez Lisesi
Lisans : 2007-2011 SDÜ, Fen Edebiyat Fakültesi, Biyoloji Bölümü
Yüksek Lisans : 2011-2013 SDÜ, Fen Bilimleri Enstitüsü, Biyoloji Anabilim Dalı
Yayınlar
1. Önal, Ö., Türk, S., Çınar, K. 2013. Dişi ve Erkek Keklik (Alectoris chukar) Üropigial
Bezinin Histolojik ve Histokimyasal Özellikleri. F.Ü.Sağ. Bil. Vet. Derg. 2013; 27 (1):
7-12.
2. Önal, Ö., Çınar, K. 2013. Keklik (Alectoris Chukar) Harder Bezi Üzerine Histolojik
Ve Histokimyasal Çalışmalar. F.Ü.Sağ. Bil. Vet. Derg. 2013; 27 (1): 13-17
Taranmış
Fotoğraf
(3.5cm x 3cm)















![Retroflex versus bunched [r] in compensation for ...linguistics.berkeley.edu/phonlab/documents/2011/retrobunch_paper.pdf · Retroflex versus bunched [r] in compensation for coarticulation](https://static.fdocuments.in/doc/165x107/5b09bbee7f8b9a992a8e2cb3/retroflex-versus-bunched-r-in-compensation-for-versus-bunched-r-in-compensation.jpg)


