
Trophic importance of subtidal metazoan meiofauna ... · Trophic importance of subtidal metazoan...
12
Mar Biol (2007) 152:339–350 DOI 10.1007/s00227-007-0696-y 123 RESEARCH ARTICLE Trophic importance of subtidal metazoan meiofauna: evidence from in situ exclusion experiments on soft and rocky substrates Roberto Danovaro · Mariaspina Scopa · Cristina Gambi · Simonetta Fraschetti Received: 29 January 2007 / Accepted: 28 March 2007 / Published online: 24 April 2007 © Springer-Verlag 2007 Abstract In coastal marine ecosystems, predation might aVect spatial distribution and population dynamics of ben- thic assemblages. Here, by means of experimental exclu- sion of potential predators, we compared the eVects of epibenthic predation on metazoan meiofaunal assemblages on soft and rocky substrates. DiVerent patterns of abun- dance were observed in uncaged versus caged plots, across habitats. In caged soft substrates, the abundance of Nema- todes, Copepods and Polychaetes increased by 56, 45, 57%, respectively, in the Wrst 3 months. An increase in the num- ber of meiofaunal taxa was also observed. The exclusion of predators from rocky substrates showed less clear patterns. It did not aVect the number of taxa while a decrease in mei- ofaunal abundance was observed. Our results suggest that the exclusion of epibenthic predators had clear eVect on total metazoan meiofaunal abundance and on the number of taxa, only in soft bottoms. The diVerent impact of predation across habitats can be potentially explained by diVerences in terms of spatial variability and substrate complexity. We estimated that, coarsely, more than 75% of total metazoan meiofaunal production can be channeled to higher trophic levels through predation on soft-bottoms. Among meiofa- unal taxa, Polychaetes and Nematodes provided the major contribution to benthic energy transfers. These results sug- gest the trophic relevance of metazoan meiofauna in coastal food webs and claim for the reWnement of further experi- ments for the quantiWcation of its role in diVerent ecologi- cal systems. Introduction Predation is considered a key factor aVecting patterns of distribution of populations and assemblages, at local scale (Constable 1999; Menge 2000; Underwood 2000; Lenihan and Micheli 2001). Predators inXuence on coastal systems has been quantiWed through a vast array of manipulative experiments, in a wide range of habitats, including salt marshes (e.g. Sarda et al. 1998), intertidal Xats (e.g. Reise 1979; Fernandes et al. 1999; Richards et al. 1999; Thrush 1999), coastal lagoons (e.g. Irlandi and Mehlich 1996; McArthur 1998), subtidal mud-substrates (e.g. Hines et al. 1990), eelgrass beds (Reise 1978), seagrass beds (e.g. Macia 2000; Spitzer et al. 2000; Hindell et al. 2002), kelp forests (e.g. Kennelly 1991), mangroves (e.g. Schrijvers and Vincx 1997; Schrijvers et al. 1998), coral reefs (e.g. Connell 1997), rocky substrates (e.g. Choat 1982; Stocker 1986) and artiWcial structures (e.g. Connell 2001; Einbinder et al. 2006). The exclusion of epibenthic predators (such as Wshes, shrimps and crabs) determines, in some cases, the increase of macrofaunal abundance by a factor of two or more (Posey et al. 2006). However, results are highly vari- able in space and time and might depend on spatial and temporal scales of investigation, and on the size of preda- tors (Quammen 1984; Micheli 1997; Felsinga et al. 2005; Como et al. 2004; Langlois et al. 2006). Metazoan meiofauna are potentially a relevant food source for higher trophic levels, particularly for macrofa- una, small Wshes, juveniles of larger Wshes and other Communicated by R. Cattaneo-Vietti. R. Danovaro (&) · M. Scopa · C. Gambi Department of Marine Science, Polytechnic University of Marche, Via Brecce Bianche, 60131 Ancona, Italy e-mail: [email protected] S. Fraschetti Department of Biological and Environmental Science and Technology, University of Lecce, Prov.le Lecce-Monteroni, 73100 Lecce, Italy
Transcript of Trophic importance of subtidal metazoan meiofauna ... · Trophic importance of subtidal metazoan...

Mar Biol (2007) 152:339–350
DOI 10.1007/s00227-007-0696-yRESEARCH ARTICLE
Trophic importance of subtidal metazoan meiofauna: evidence from in situ exclusion experiments on soft and rocky substrates
Roberto Danovaro · Mariaspina Scopa · Cristina Gambi · Simonetta Fraschetti
Received: 29 January 2007 / Accepted: 28 March 2007 / Published online: 24 April 2007© Springer-Verlag 2007
Abstract In coastal marine ecosystems, predation mightaVect spatial distribution and population dynamics of ben-thic assemblages. Here, by means of experimental exclu-sion of potential predators, we compared the eVects ofepibenthic predation on metazoan meiofaunal assemblageson soft and rocky substrates. DiVerent patterns of abun-dance were observed in uncaged versus caged plots, acrosshabitats. In caged soft substrates, the abundance of Nema-todes, Copepods and Polychaetes increased by 56, 45, 57%,respectively, in the Wrst 3 months. An increase in the num-ber of meiofaunal taxa was also observed. The exclusion ofpredators from rocky substrates showed less clear patterns.It did not aVect the number of taxa while a decrease in mei-ofaunal abundance was observed. Our results suggest thatthe exclusion of epibenthic predators had clear eVect ontotal metazoan meiofaunal abundance and on the number oftaxa, only in soft bottoms. The diVerent impact of predationacross habitats can be potentially explained by diVerencesin terms of spatial variability and substrate complexity. Weestimated that, coarsely, more than 75% of total metazoanmeiofaunal production can be channeled to higher trophiclevels through predation on soft-bottoms. Among meiofa-unal taxa, Polychaetes and Nematodes provided the majorcontribution to benthic energy transfers. These results sug-
gest the trophic relevance of metazoan meiofauna in coastalfood webs and claim for the reWnement of further experi-ments for the quantiWcation of its role in diVerent ecologi-cal systems.
Introduction
Predation is considered a key factor aVecting patterns ofdistribution of populations and assemblages, at local scale(Constable 1999; Menge 2000; Underwood 2000; Lenihanand Micheli 2001). Predators inXuence on coastal systemshas been quantiWed through a vast array of manipulativeexperiments, in a wide range of habitats, including saltmarshes (e.g. Sarda et al. 1998), intertidal Xats (e.g. Reise1979; Fernandes et al. 1999; Richards et al. 1999; Thrush1999), coastal lagoons (e.g. Irlandi and Mehlich 1996;McArthur 1998), subtidal mud-substrates (e.g. Hines et al.1990), eelgrass beds (Reise 1978), seagrass beds (e.g.Macia 2000; Spitzer et al. 2000; Hindell et al. 2002), kelpforests (e.g. Kennelly 1991), mangroves (e.g. Schrijversand Vincx 1997; Schrijvers et al. 1998), coral reefs (e.g.Connell 1997), rocky substrates (e.g. Choat 1982; Stocker1986) and artiWcial structures (e.g. Connell 2001; Einbinderet al. 2006). The exclusion of epibenthic predators (such asWshes, shrimps and crabs) determines, in some cases, theincrease of macrofaunal abundance by a factor of two ormore (Posey et al. 2006). However, results are highly vari-able in space and time and might depend on spatial andtemporal scales of investigation, and on the size of preda-tors (Quammen 1984; Micheli 1997; Felsinga et al. 2005;Como et al. 2004; Langlois et al. 2006).
Metazoan meiofauna are potentially a relevant foodsource for higher trophic levels, particularly for macrofa-una, small Wshes, juveniles of larger Wshes and other
Communicated by R. Cattaneo-Vietti.
R. Danovaro (&) · M. Scopa · C. GambiDepartment of Marine Science, Polytechnic University of Marche, Via Brecce Bianche, 60131 Ancona, Italye-mail: [email protected]
S. FraschettiDepartment of Biological and Environmental Science and Technology, University of Lecce, Prov.le Lecce-Monteroni, 73100 Lecce, Italy
123

340 Mar Biol (2007) 152:339–350
epibenthic predators (Chardy and Dauvin 1992) as alsosuggested by the gut content analyses of XatWshes, crabsand shrimps (de Morais and Bodiou 1984; Hoyt et al. 2000;Feller 2006). On soft substrates, the eVects of predation onsubtidal meiofauna have received scarce attention (maybebecause this ecological compartment is less tractable froman experimental point of view), but there are evidences ofboth direct and indirect eVects of predators on meiofaunalassemblages (Henry and Jenkins 1995; Gregg and Fleeger1997). Direct eVects of predation result in lowering meiofa-unal abundance and changing community structure, whilstindirect eVects are related with the capability of changingthe physical–chemical characteristics of the sediments(Warwick et al. 1990; OlaVson and Elmgren 1991; Titaet al. 2000). Other studies suggest that Wsh predation onmetazoan meiofauna can be almost completely negligiblebecause prey populations are large and the number of pred-atory Wshes comparatively small (Gibbons 1988; Coull1990; Shaw and Jenkins 1992). Furthermore, the continu-ous reproduction and recruitment and fast growth rates ofseveral metazoan meiofaunal taxa might represent an obsta-cle in quantifying eVects of predation (Hicks 1984). Onrocky substrates, Wshes are considered the major predatorson epibiota (reviews: Choat 1982; Choat and Kingett 1982;Hixon 1997), but information on the potential impact onmeiofauna is extremely scarce also due to the few dataavailable on meiofauna living in this habitat. It has beenhypothesized that habitat complexity of rocky substratescan alter predation eYciency and inXuence faunal distribu-tion and diversity (Coull and Wells 1982; Gibbons 1988).
The scarce available information on the fraction of meio-faunal production removed through predation makes inves-tigations of the potential transfer of energy associated withmeiofauna a priority to better understand their role in thecomplexity of the benthic marine food webs, in both softand rocky habitats. Previous investigations demonstratedthat soft and rocky substrates are inhabited by the samemetazoan meiofaunal taxa, but with substantial diVerencesin terms of overall abundance (typically lower on rockysubstrates) and relative importance of the diVerent taxo-nomic groups (typically rocky substrates are generallydominated by Harpacticoid Copepods, Isopods and Amphi-pods, whereas soft bottom are dominated by Nematodes;Danovaro and Fraschetti 2002). Such diVerences mighthave important implications on benthic food web function-ing, but a formal comparison of predatory eVects acrosshabitats has never been carried out.
In this study, we compared the potential eVects ofepibenthic predation on metazoan meiofaunal abundance,number of taxa, assemblage structure and production in softand rocky substrates, through experimental caging manipu-lations. The hypothesis of the present work is that the exclu-sion of epibenthic predators aVects metazoan meiofauna both
in terms of abundance, assemblage structure and produc-tion, determining signiWcant diVerences between caged anduncaged plots. We also hypothesize that the exclusionexperiment could have a diVerent impact according to thetype of substrate investigated, with soft substrates revealingmore evident changes than rocky substrates.
Materials and methods
Study area
The investigation was carried out in a coastal area of theNorthern Adriatic Sea (Conero Promontory; 43.57 Lat N,13.62 Long W; Fig. 1). The Northern Adriatic Sea is fea-tured by shallow depth (on average ca. 35 m in the northernsector), large nutrient inputs coming from the Po River and,in the study area, almost negligible tidal excursion (< 30 cm).This is a coastal area easy to be manipulated by SCUBAdivers (9 m depth), characterized by the presence of bothsoft bottoms and rocky substrates in close proximity, allow-ing the test of the eVect of predation across the two habitats.During the study period, current speeds were typically <10 cm s¡1 and surface water temperatures ranged from 24to 27°C and transparency from 2 to 4 m (Secchi Disk).
Fig. 1 Study area (upper half) and location of the sites utilized forin situ experiments on rocky and soft substrates (lower half)
Adriatic Sea
13.513.4 13.4 13.5 13.6 13.6 13.7 13.7 13.843.4
43.6
43.5
43.7
Longitude
Lat
itu
de
Trave Cliff0 m
2.5 m
8.0 m Hard Substrates
Soft Substrates9.0 m
13.5
123

Mar Biol (2007) 152:339–350 341
Sediments are composed by Wne sands (Danovaro and Fras-chetti 2002). Visual census on Wsh abundances in the studyarea provided evidence of the presence of predators of mei-ofauna, such as small Wshes (Blennidae, Mullidae and Soleaspp.; typically <5 cm in length) and crabs (i.e. Eriphiaverrucosa and Pilumnus hirtellus). Previous studies carriedout in this area reported meiofaunal abundance rangingfrom 130 to 974 and from 820 to 6,298 ind. 10 cm¡2 onrocky and soft substrates, respectively (Danovaro and Fras-chetti 2002).
Field experiments
The experiment was carried out in two habitat types: softand rocky substrates. In each habitat, the eVects of preda-tion on metazoan meiofaunal abundance, number of taxa,assemblage structure and production were quantiWedcomparing one caged with one uncaged (control) plot. Inboth habitats, the uncaged plot was located at the samedepth of the cage, about 15 m far apart. The cages for theexclusion of predators were made of zinc-iron (length70 cm £ width 70 cm £ height 12 cm on soft substrates,and length 150 cm £ width 150 cm £ height 12 cm onrocky substrates). Since we did not have any speciWchypothesis on which predator species among those visuallyencountered in situ could be responsible for possible diVer-ences of metazoan meiofaunal assemblages between cagedand uncaged plots, a typical cage (with top and lateral sidescovered by a plastic net of 1-mm mesh to allow the immi-gration and emigration of meiofauna) was utilized toexclude larger epibenthic predators. Before cage deploy-ment, the substrates were accurately checked to make surethat epibenthic predators were not included. The lateralpanels of the cages were inserted into soft sediments pene-trating by ca. 20 cm and Wxed using four metal nails driveninto the substratum down to a depth of ca. 50 cm. Consider-ing that cages can alter the hydrodynamic regime at the sed-iment–water interface, modifying sediment transport withpossible consequences on meiofauna (Commito and Tita2002), four vertical metal panels were inserted into the sed-iment (down to ca. 20 cm depth) at each side of the cage.These panels, as observed during our diving activities,resulted successful in avoiding sediment alteration (e.g., byerosion) around and within the cage. On rocky substrates,only metal nails were utilized, but the basis of the cage wasequipped with a thick rubber ring allowing a perfect adher-ence to the substrate.
We are aware that the present experimental design (onecage vs. one uncaged plot) limits data interpretation anddiVerences among treatments are diYcult to discriminate inthe absence of a rigorous quantiWcation of spatial variabil-ity of prey and predators. A second aspect is the lack ofprocedural controls, with the possibility that the eVects of
Wsh exclusion could be confounded with caging eVects (e.g.shading, reduced water movement, presence of the struc-ture). As a consequence, results are discussed with caution,in the light of a suite of factors which could have inXuencedthe outcome of the experiment.
A pre-survey in both habitats showed that there were noinitial signiWcant diVerences among structure and abun-dance of benthic assemblages between the location selectedfor the deployment of the cages and control areas. Duringthe experiment, no particular concentration of Wsh wasobserved around the experimental plots.
Sampling of metazoan meiofauna was carried out at sixdates from June to September by SCUBA divers. The WrstWve samplings were performed every 15 days and the lastsamples were collected at 90 days from the beginning ofthe experiment. At each sampling date, meiofaunal sampleswere collected in three replicates in caged and uncagedplots in the both soft and rocky substrates. Soft-bottommeiofauna were sampled penetrating plexiglas corers (i.d.,3.7 cm) into the sediment down to a depth of 14 cm. Forrocky-bottom meiofauna, sampling was carried out bymeans of a modiWed manual corer (internal diameter, i.d.8.5 cm, 14 cm length, see Danovaro and Fraschetti 2002 fordetails). In caged plots, samples were collected opening thenet on the top of the cage without disturbing the surface ofthe sampling area. Sampling was never carried out in prox-imity of the four sides to avoid potential edge eVects (Belland Woodin 1984). At each sampling date, samples weretaken in points selected with random numbers and nevercoincident with sampling points of previous sampling datesto guarantee data independency.
Each sample was Wxed with buVered formaldehyde (4%Wnal volume), stained with Bengal Rose and samples weresonicated from 3 min (Branson Sonifer, 60 W, three timesfor 1 min with 30 s intervals). Meiofauna, sieved through a1-mm mesh and retained by 30 �m mesh net, was added toLudox HD 40 (density arranged to 1.15) for density centri-fugation extraction (10 min, 800£ g, three times). Thesame extraction and preservation procedures were used forsoft- and rocky-bottoms samples. All meiobenthic animalswere counted and classiWed per taxon under a stereo micro-scope.
Meiofaunal biomass and production
Meiofaunal biomass was estimated from individual bio-mass of Nematodes, Copepods, Polychaetes, Turbellariansand others taxa reported according with the proceduredescribed by Danovaro et al. (2002, for details). The frac-tion of biomass removed by predation was calculated as thediVerences between the average biomass (estimated on thewhole period of the experiment) of each selected taxonfrom caged and uncaged sediments.
123

342 Mar Biol (2007) 152:339–350
Estimates of metazoan meiofaunal production on annualbasis were obtained using diVerent values of the Productionto Biomass (P:B) ratio (Heip et al. 1982). In order to mini-mize the associated error and to make our results compara-ble with those available in the literature, we utilized fourdiVerent approaches: (1) a P:B ratio = 9, as average valuefor all taxa (Gerlach 1971); (2) a P:B ratio = 10 as averagevalue for all taxa (McIntyre 1964); (3) a P:B ratio speciWcfor each taxon (i. e. P:B = 8.4 for Nematodes, 18 for Cope-pods, 5 for Turbellarians and Annelids and 9.12 for othertaxa; Warwick et al. 1979); (4) and another fractionatedapproach (P:B = 9.75 for Nematodes, 21 for Copepods,11.3 or Turbellarians, 9.7 for Annelids and 9.12 for othertaxa; Lasserre et al. 1976). Methods based on annual P:Bratios are subjected to problems of accuracy (Warwick1980). Although P:B ratios are variable within each taxon,the methods used here remain the only way to provide com-parable values of total meiofaunal secondary production(Chardy and Dauvin 1992; Chardy et al. 1993) and havebeen previously applied to the evaluation of metazoan mei-ofaunal contribution to energy transfer in other systems(Danovaro et al. 2002).
Statistical analyses
Analyses were carried out separately for each habitat. Theexperimental design includes two factors: time (six levels,Wxed) and treatment (two levels, Wxed and orthogonal totime) with three replicates per combination of factors.
To investigate the eVect of the treatment on the meio-faunal assemblages as a whole (36 observations for 14variables for soft substrate and 17 variables for rockysubstrate), and on the meiofaunal biomass inhabiting softsediments (36 observations for 8 variables), the experimen-tal design was analyzed using permutational multivariateanalysis of variance (PERMANOVA; Anderson 2001a;McArdle and Anderson 2001). The analyses were basedon Bray–Curtis dissimilarities. Data were all square-roottransformed (Bray and Curtis 1957). All tests used 4999permutations of the appropriate units (see Anderson 2001b;Anderson and ter Braak 2003). The analyses were doneusing the computer program PERMANOVA.exe (Ander-son 2005). The interaction term time £ treatment wasfound signiWcant in the analyses relative to the experimenton soft substrates. Pairwise comparisons were used to testfor diVerences of the factor treatment within each level ofthe factor time.
Graphical representations of multivariate patterns wereobtained by non-metric multi-dimensional scaling (nMDS)ordinations using the PRIMER 5 program (PlymouthMarine Laboratory, UK) (Clarke and Warwick 1994). Forrocky substrates, a single ordination was produced, plottingcentroids of the n = 3 replicates for each sampling time.
Centroids were calculated using principal coordinates(Anderson 2001). For soft substrates, since the interactionterm time £ treatment was found signiWcant, separatenMDS plots were obtained, plotting all the replicate obser-vations (Anderson 2003).
The similarity percentages procedure (PRIMER, SIM-PER routine, Clarke 1993) was used to identify the percent-age contribution that each taxon made to the observed valueof the Bray–Curtis dissimilarity between caged and uncagedplots, when pairwise multivariate comparison showed evi-dence of signiWcant diVerences. A cut-oV criterion was usedfor the identiWcation of a subset of taxa whose cumulativepercentage contribution reached 90% of the dissimilarityvalue.
Analysis of variance (ANOVA) was used to test fordiVerences in total meiofaunal abundance, total numberof taxa and abundance of dominant taxa between cagedand uncaged plots. Analyses of variance were followedby a posteriori Student–Newman–Keuls tests (SNK tests)when time £ treatment was found to be signiWcant withP < 0.05. Homogeneity of variances was tested using Coch-ran’s C test (Winer et al. 1991).
Results
Meiofaunal assemblages
A total of 14 taxa were collected in soft sediments. Nema-todes were the dominant taxon, on average 80–84% of totalmetazoan meiofaunal abundance, in uncaged and cagedareas respectively. On rocky substrates, a total of 17 taxawere collected and Harparticoid Copepods dominated, onaverage 33–34% of meiofaunal abundance, in uncaged andcaged areas, respectively (Fig. 2).
Caging experiments
PERMANOVA analyses showed diVerent results for thetwo habitats. On soft bottoms, a signiWcant multivariateinteraction between the factors time £ treatment wasobserved. Assemblages under the cage were signiWcantlydiVerent from those in the uncaged plot even thoughthose diVerences varied with time (see Table 1). Pairwisemultivariate comparisons showed that there were no initialsigniWcant diVerences between caged and uncaged assem-blages (as observed in the pre-survey), and these diVerencesbecame apparent after 45 days from the beginning of theexperiment. The nMDS analysis conWrmed the resultsobtained from the above analyses (Fig. 3 a, b, c, d, e and f).SIMPER analyses (Table 2) revealed that four groups(Nematodes, Copepods, Nauplii and Polychaetes) wereimportant in diVerentiating the assemblages inside caged
123

Mar Biol (2007) 152:339–350 343
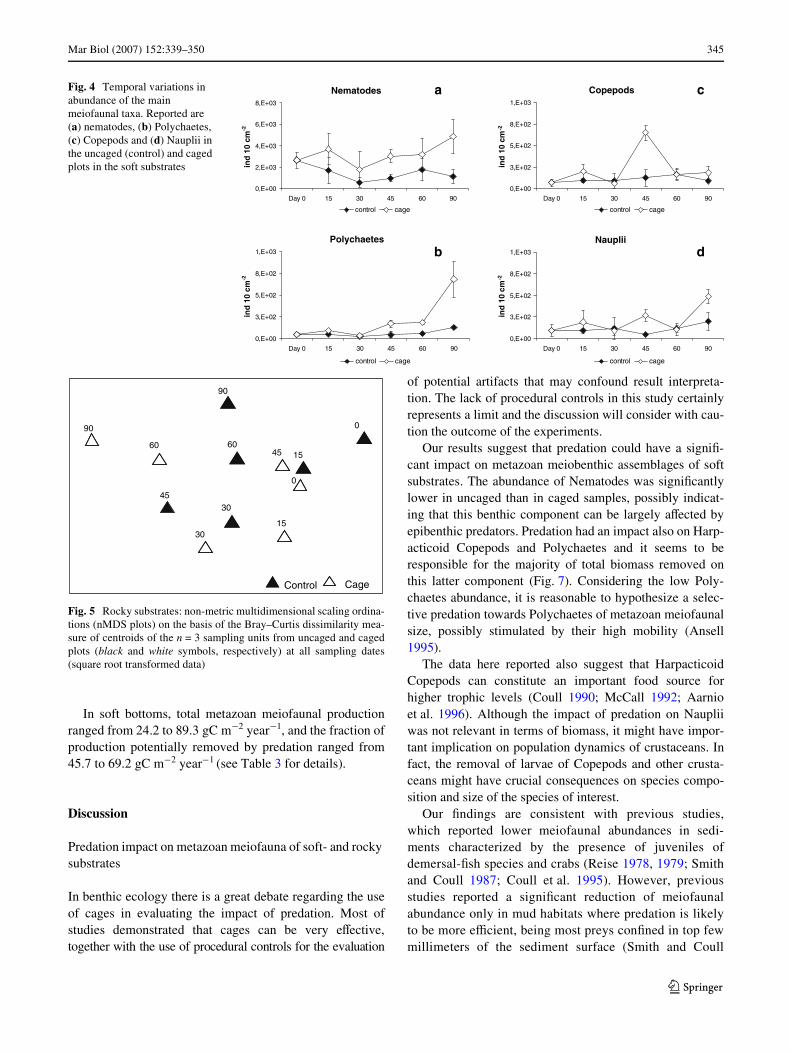
versus uncaged plots. These four groups were presentin both plots, but with lower mean abundance in theuncaged areas. ANOVA carried out on single variablesrevealed that Nematode abundance showed signiWcantdiVerences between caged and uncaged plots consistentlyin time (time £ treatment, F5,24, P = 0.39). Copepods, theirNauplii and Polychaetes were characterized by signiWcantdiVerences in the interaction term time £ treatment andSNK tests showed that diVerences between cage anduncaged plots were apparent after 45 days from the begin-ning of the experiment. The abundance of Nematodes,Copepods and Polychaetes decreased by 56, 45, 57%,respectively at 90 days (Fig. 4). Therefore, at the end of theexperiment, the overall abundance of metazoan meiofaunain uncaged plot was approximately 50% lower than thatin caged sediments. ANOVA on the number taxa alsorevealed signiWcant diVerences in the interaction termtime £ treatment (F5,24, P = 0.013). Also in this case, SNKtest revealed that there were no diVerences in the number oftaxa until 30 days from the beginning of the experiment
when some minor taxa (such as Kinorhynchs, Oligochaetes,Amphipods and Termosbanaceans) disappeared from theuncaged plot.
On rocky substrates, PERMANOVA analysis showed noclear eVect of the caging experiment: signiWcant diVerencesbetween caged assemblages and uncaged plots wereobserved consistently in time (Table 1), with lower meanabundance in the caged plot. This possibly means that justafter 15 days after the beginning of the experiment, theassemblages under the cage already diVered from assem-blages in the uncaged plot. The nMDS analysis conWrmedthe diVerences already evidenced by PERMANOVA(Fig. 5). SIMPER analysis revealed that the group mainlyresponsible for the observed diVerences was represented byCopepods, more abundant in uncaged plot than under thecage (i.e., on average a contribution of ca. 13% to the aver-age dissimilarity between caged and uncaged plot). TheANOVA did not show any diVerence in the number of taxa,revealing only signiWcant diVerences in time (F5,24 = 12.12,P < 0.0002).
Fig. 2 Meiofaunal community structure in uncaged (control a, b) and caged plots (c, d) at soft and rocky bottoms, respectively
Day 0 15 30 45 60 90 Day 0 15 30 45 60 90
Rocky-bottom meiofauna
0%
20%
40%
60%
80%
100%
50%
60%
70%
80%
90%
100%
Nematodes Copepods NaupliiPolychaetes Bivalves Others
Soft-bottom meiofauna
50%
60%
70%
80%
90%
100%
Day 0 15 30 45 60 90 Day 0 15 30 45 60 90
0%
20%
40%
60%
80%
100%
Nematodes Copepods NaupliiPolychaetes Bivalves Others
a b
c d
Table 1 Results of the PERMANOVA test on meiofaunal abundance of soft and rocky substrates and on meiofaunal biomass of soft substrates(data were all square root transformed)
P values were obtained using 4999 permutations of a given permutable unit. In the analyses relative to soft substrates, the results of the interactionterm are the only reported in the table, since time £ treatment was found signiWcant in the analysis
Source df Meiofaunal abundance Meiofaunal biomass
Soft substrates Rocky substrates Soft substrates
MS F P MS F P MS F P
Time = T 5 1243.4 1205.6 3.9 0.0002 1380.5
Treatment (Tr) = Tr 1 2931.0 1597.9 5.1 0.0006 2934.1
T £ Tr 5 452.8 1.86 0.04 355.2 1.1 0.28 497.3 2.7 0.003
Residual 24 242.4 309.1 184.6
Total 35
123

344 Mar Biol (2007) 152:339–350
Since controversial interpretations about estimates ofbiomass transfer to the higher trophic levels could resultfrom the experiment on rocky substrates, we estimated thetransfer of metazoan meiofaunal biomass and productiononly in soft sediments. In soft sediments, we selected Nem-atodes, Copepods, Polychaetes, Bivalves, Cladocerans andGastrotrichs as these groups had shown an increase underthe cage (related to total metazoan meiofaunal abundance)higher than 0.1%.
Data on total metazoan meiofaunal biomass and on eachspeciWc taxon biomass in caged and uncaged soft sedi-ments, and the fraction potentially removed by predationwere shown in Fig. 6.
PERMANOVA analyses showed diVerences betweencaged and uncaged plots in the value of biomass on softsediments, varying with time (see Table 1). As observedin the analysis of total abundance, pairwise multivariatecomparisons showed that there were no initial signiWcantdiVerences between caged and uncaged assemblages forbiomass. Also in this case, these diVerences became appar-ent after 45 days from the beginning of the experiment.Considering the contribution of each taxon to the total bio-mass potentially removed through predation, we identiWedNematodes and Polychaetes as the major contributors(Fig. 7).
Fig. 3 Soft substrates: non-metric multidimensional scaling ordina-tions (nMDS plots) on the basis of the Bray–Curtis dissimilaritymeasure of square root transformed data from uncaged and caged plots(black and white symbols, respectively) separately for each samplingdate. Reported are (a) beginning of the experiment, (b) 15 days, (c)30 days, (d) 45 days, (e) 60 days and (f) 90 days
Control CageControl Cage
a
c
e
b
d
f
Tab
le2
Ave
rage
abu
ndan
ce o
f the
mai
n ta
xa c
ontr
ibut
ing
to th
e B
ray–
Cur
tis d
issi
mila
rity
val
ue b
etw
een
cont
rol (
unca
ged)
and
cag
ed p
lots
at 4
5, 6
0 an
d 90
days
resp
ecti
vely
fro
m th
e be
ginn
ing
of th
e ex
peri
men
t in
soft
sed
imen
ts
Soft
bot
tom
45da
ys60
days
90da
ys
Ave
rage
abu
ndan
ceC
ontr
ibut
ion
%C
umul
ativ
e %
Ave
rage
abu
ndan
ceC
ontr
ibut
ion
%C
umul
ativ
e %
Ave
rage
abu
ndan
ceC
ontr
ibut
ion
%C
umul
ativ
e %
Con
trol
Cag
eC
ontr
olC
age
Con
trol
Cag
e
Nem
atod
es30
.854
.834
.334
.340
.755
.636
.736
.732
.369
.043
.943
.9
Cop
epod
s10
.525
.421
.555
.712
.612
.55.
771
.39.
213
.14.
589
.1
Nau
plii
6.9
16.3
13.3
69.0
10.4
9.7
5.8
65.5
13.7
21.9
9.8
71.1
Poly
chae
tes
6.1
12.9
9.7
78.7
7.6
13.5
12.6
49.3
11.3
25.9
17.4
61.3
Oth
ers
2.9
6.3
4.9
89.6
1.1
5.9
10.4
59.7
0.0
2.5
3.1
92.2
123
Mar Biol (2007) 152:339–350 345
In soft bottoms, total metazoan meiofaunal productionranged from 24.2 to 89.3 gC m¡2 year¡1, and the fraction ofproduction potentially removed by predation ranged from45.7 to 69.2 gC m¡2 year¡1 (see Table 3 for details).
Discussion
Predation impact on metazoan meiofauna of soft- and rocky substrates
In benthic ecology there is a great debate regarding the useof cages in evaluating the impact of predation. Most ofstudies demonstrated that cages can be very eVective,together with the use of procedural controls for the evaluation
of potential artifacts that may confound result interpreta-tion. The lack of procedural controls in this study certainlyrepresents a limit and the discussion will consider with cau-tion the outcome of the experiments.
Our results suggest that predation could have a signiW-cant impact on metazoan meiobenthic assemblages of softsubstrates. The abundance of Nematodes was signiWcantlylower in uncaged than in caged samples, possibly indicat-ing that this benthic component can be largely aVected byepibenthic predators. Predation had an impact also on Harp-acticoid Copepods and Polychaetes and it seems to beresponsible for the majority of total biomass removed onthis latter component (Fig. 7). Considering the low Poly-chaetes abundance, it is reasonable to hypothesize a selec-tive predation towards Polychaetes of metazoan meiofaunalsize, possibly stimulated by their high mobility (Ansell1995).
The data here reported also suggest that HarpacticoidCopepods can constitute an important food source forhigher trophic levels (Coull 1990; McCall 1992; Aarnioet al. 1996). Although the impact of predation on Naupliiwas not relevant in terms of biomass, it might have impor-tant implication on population dynamics of crustaceans. Infact, the removal of larvae of Copepods and other crusta-ceans might have crucial consequences on species compo-sition and size of the species of interest.
Our Wndings are consistent with previous studies,which reported lower meiofaunal abundances in sedi-ments characterized by the presence of juveniles ofdemersal-Wsh species and crabs (Reise 1978, 1979; Smithand Coull 1987; Coull et al. 1995). However, previousstudies reported a signiWcant reduction of meiofaunalabundance only in mud habitats where predation is likelyto be more eYcient, being most preys conWned in top fewmillimeters of the sediment surface (Smith and Coull
Fig. 4 Temporal variations in abundance of the main meiofaunal taxa. Reported are (a) nematodes, (b) Polychaetes, (c) Copepods and (d) Nauplii in the uncaged (control) and caged plots in the soft substrates
Nematodes
0,E+00
2,E+03
4,E+03
6,E+03
8,E+03
Day 0 15 30 45 60 90
mc 01
dni
2-
control cage
Copepods
0,E+00
3,E+02
5,E+02
8,E+02
1,E+03
Day 0 15 30 45 60 90
mc 01
dni
2-
control cage
Polychaetes
0,E+00
3,E+02
5,E+02
8,E+02
1,E+03
Day 0 15 30 45 60 90
mc 01
dni
2 -
control cage
Nauplii
0,E+00
3,E+02
5,E+02
8,E+02
1,E+03
Day 0 15 30 45 60 90
mc 0 1
dni
2 -
control cage
a c
b d
Fig. 5 Rocky substrates: non-metric multidimensional scaling ordina-tions (nMDS plots) on the basis of the Bray–Curtis dissimilarity mea-sure of centroids of the n = 3 sampling units from uncaged and cagedplots (black and white symbols, respectively) at all sampling dates(square root transformed data)
0
0
15
15
30
30
45
45
6060
90
90
Control Cage
123

346 Mar Biol (2007) 152:339–350
1987; Gregg and Fleeger 1997). Our experimental resultssuggest that these Wndings could be also extended tosandy habitats.
The presence of multiple-trophic levels can complicatethe interpretation of results. For instance, the lack of epi-benthic predators can also increase the abundance infaunal(endobenthic) predators, which can overexploit other infau-nal taxa (e.g. Ambrose 1984a, b; Commito and Shrader1985). Several studies reported also that the exclusion ofpredators can determine a concomitant reduction in theabundance of preys’ conspeciWc juveniles (e.g. RaVaelliand Milne 1987; Thrush et al. 1994). Kent and Day (1983)demonstrated that increased abundances of the adults ofnereid polychaetes in cages can reduce the abundance ofjuvenile conspeciWcs, and this eVect counter-balance theeVects of predation exclusion (i.e. birds and Wshes). How-ever, in our study, we observed that the abundance of theNauplii of Copepods in caged areas initially increased withrespect to uncaged (control) sediments, as a result of thepotentially lower predatory pressure on Copepod femalesbearing eggs.
Our data indicate that predation did modify both meta-zoan meiofaunal community structure and the number oftaxa (here utilized as an estimate of meiofaunal diversity),which was signiWcantly higher in caged than in uncagedsoft substrates. A moderate epibenthic predation (i.e., in ourcase in the uncaged plot) is expected to increase benthicdiversity as a result of the reduced competition (Peterson1979; BonsdorV et al. 1986) and such an eVect has beenobserved on diVerent trophic levels (Mattila 1992; Mattilaand BonsdorV 1998). However, the opposite pattern wasobserved in this study. A strong and/or selective predationmay lead to a reduced species diversity (Peterson 1979).Here we report that epibenthic predation on sandy sub-strates could result in a reduced metazoan meiofaunal bio-diversity due to the disappearance of minor taxa such asKinorhynchs, Termosbanaceans, Oligochaetes and Amphi-pods, and we cannot exclude that increasing the taxonomicdetail of this analysis could lead to partially diVerentresults.
The exclusion of predators from rocky substrates dis-played controversial patterns as the predation had no eVects
Fig. 6 Biomass of the main meiofaunal taxa in uncaged (control), caged plots and the fraction of the biomass of each taxon potentially removed by predation in soft sediments
0
4
8
12
16
eNm
sedota
sdopepoC loP
hcya
tese
Blavi
sev
C
snarecodal Gtsar
tori
shcO
hte
sr atoTl
m Cg
2-
control cagecage-control
Fig. 7 Contribution of meiofaunal taxa to biomass transfer in soft substrates. Reported are nematodes, copepods, polychaetes, bivalves, cladocerans and gastrotrichs
40,9%
6,4%
47,6%
0,7%
0,3%
1,3%
2,8%
Nematodes Copepods Polychaetes Bivalves
Cladocerans Gastrotrichs Others
123

Mar Biol (2007) 152:339–350 347
on the number of metazoan meiofaunal taxa and we alsoobserved the decrease of meiofaunal abundance in cagedplots. The Wrst result might be partly due to the characteris-tics of the substrate. Comparisons of predation eVectsbetween unvegetated soft-sediment communities and adja-cent vegetated habitats generally indicate that predatorshave a greater impact in the absence of biogenic structure(Lenihan and Micheli, 2001). The eVects of predation couldbe altered by spatial heterogeneity and by the presence ofspatial refuges (Reise 1978; Gilinsky 1984; Skilleter 1994).For example, macroalgal cover increases the spatial com-plexity of the habitat, oVering refuges from predation andcontributing to reduce the impact of epibenthic predationon rocky bottom metazoan meiofauna. In a previous studycarried out at the same site, Danovaro and Fraschetti (2002)demonstrated that the presence of macroalgae on theserocky substrates (dominated by Rhodophyceae, with analgal biomass ranged up to 25.1 g 10 cm¡2) increased the
fractal complexity of the system, which could act as refugefor metazoan meiofaunal preys (Coull and Wells 1982).The interpretation of the decrease of meiofaunal abundancein caged plot is controversial and an implementation of theexperimental design is strongly needed to quantify theeVects of predation in this habitat.
Meiofaunal production and potential energy transfer to the benthic food webs
Our estimates of metazoan meiofaunal production in theuncaged plot in soft sediments were comparable to valuesreported (using the same procedure) for estuarine environ-ments (Escaravage et al. 1989) and two to four times higherthan those reported in the North Sea and the Ligurian Sea(Heip et al. 1983; Danovaro et al. 2002). Since our studyindicated that soft bottom meiofauna were subjected to apotentially signiWcant predatory pressure, we estimated the
Table 3 Meiofaunal biomass and production (estimated using diVerent P:B ratios) in uncaged (a) and caged (b) soft sediments and calculation ofthe biomass potentially removed by predation (c)
Biomass gC m¡2
Gerlach (1971) P:B = 9 gC m¡2 year¡1
McIntyre (1964) P:B = 10 gC m¡2 year¡1
Warwick et al. (1979) P:B fractionated gC m¡2 year¡1
Lasserre et al. (1976) P:B fractionated gC m¡2 year¡1
(a) Uncaged sediments
Nematodes 1.18 10.63 11.81 9.92 11.51
Copepods 0.28 2.55 2.84 5.11 5.96
Polychaetes 1.43 12.90 14.33 7.17 13.90
Bivalves 0.04 0.39 0.44 0.40 0.40
Gastrotriches 0.03 0.25 0.28 0.26 0.26
Cladocerans 0.00 0.02 0.02 0.02 0.02
Others 0.14 1.28 1.42 1.28 1.30
Total 3.11 28.02 31.14 24.15 33.35
(b) Caged sediments
Nematodes 2.60 23.37 25.97 21.82 25.32
Copepods 0.56 5.03 5.59 10.05 11.73
Polychaetes 4.98 44.79 49.77 24.88 48.28
Bivalves 0.10 0.94 1.04 0.95 0.95
Gastrotriches 0.07 0.62 0.69 0.63 0.63
Cladocerans 0.04 0.37 0.41 0.37 0.37
Others 0.22 2.02 2.24 2.03 2.05
Total 8.57 77.14 85.71 60.74 89.33
(c) Biomass removed
Nematodes 1.75 15.71 17.46 14.66 17.02
Copepods 0.38 3.45 3.83 6.89 8.04
Polychaetes 4.26 38.34 42.60 21.30 41.33
Bivalves 0.08 0.76 0.85 0.77 0.77
Gastrotriches 0.05 0.46 0.51 0.46 0.46
Cladocerans 0.05 0.42 0.46 0.42 0.42
Others 0.13 1.16 1.29 1.17 1.18
Total 6.57 59.13 65.70 45.69 69.23
123

348 Mar Biol (2007) 152:339–350
eYciency in the energy transfer, as the fraction of totalmetazoan meiofaunal production potentially transferred tohigher trophic levels (epibenthic predators).
Our estimates indicate that Polychaetes and Nematodeswere the major contributors to such energy transfer(accounting for 59 and 28% respectively of productionremoved), likely as the result of their large individual bio-mass (for Polychaetes) and high abundance (for Nema-todes). Overall the secondary production removed throughpredation ranged from 45.7 to 69.2 gC m¡2 year¡1 (accord-ing to the approach utilized). This means that, during theexperimental period, most of metazoan meiofaunal produc-tion (75–77%) was channeled to higher trophic levelsthrough predation. These results suggest that metazoanmeiofauna are a key component of the coastal benthos, con-tributing signiWcantly to energy transfer to higher trophiclevels, especially in soft substrates. The present study pro-vides new quantitative information for incorporating themetazoan meiofauna on ecological models of benthic foodwebs. The reWnement of the experimental design, with anadequate replication in space and time, will further clarifythe functional role of metazoan meiofauna and their impor-tance in benthic food webs.
Acknowledgments This work has been carried out within the frameof the Network of Excellence MARBEF (Marine Biodiversity andEcosystem Functioning) and has been Wnancially supported by theMIUR, project FIRB 2001 (n. RBAU 012 KXA_009).
References
Aarnio K, BonsdorV E, Rosenback N (1996) Food and feeding habitsof juvenile Xounder, Platichthys Xesus (L.), and turbot, Scoph-thalmus maximus L., in the Aland archipelago, northern BalticSea. J Sea Res 36:311–320
Ambrose WG Jr (1984a) InXuence of predatory Polychaetes on thestructure of a soft-bottom community in a Maine estuary. J ExpMar Biol Ecol 81:115–145
Ambrose WG Jr (1984b) Role of predatory infauna in structuring ma-rine soft bottom communities. Mar Ecol Prog Ser 17:109–115
Anderson MJ (2001a) A new method for non-parametric multivariateanalysis of variance. Aust Ecol 26:32–46
Anderson MJ (2001b) Permutation tests for univariate or multivariate anal-ysis of variance and regression. Can J Fish Aquat Sci 58:626–639
Anderson MJ (2003) PCO: a FORTRAN computer program for prin-cipal coordinate analysis. Department of Statistics, University ofAuckland, New Zealand
Anderson MJ (2005) PERMANOVA: a FORTRAN computer pro-gram for permutational multivariate analysis of variance. Depart-ment of Statistics, University of Auckland, New Zealand
Anderson MJ, ter Braak CJF (2003) Permutation tests for multi-facto-rial analysis of variance. J St Cmp Sm 73:85–113
Ansell AD (1995) Surface activity of some benthic invertebrate prey inrelation to the foraging activity of juvenile XatWshes. In: Elefthe-riou A, Ansell AD, Smith CJ (eds) Biology and ecology of shal-low coastal waters, Proceedings of the 28th European MarineBiology Symposium, Institute of Marine Biology, Crete. Fredens-borg: Olsen and Olsen, pp 245–252
Bell SS, Woodin SA (1984) Community unity: experimental evidencefrom meiofauna and macrofauna. J Mar Res 42:605–632
BonsdorV E, Mattila J, Ronn C, Osterman CS (1986) Multidimensionalinteractions in shallow soft-bottom ecosystems: testing the com-petitive exclusion principle. Ophelia Suppl 4:37–44
Bray JR, Curtis JT (1957) An ordination of the upland forest commu-nities of southern Wisconsin. Ecol Monogr 27:325–349
Chardy P, Dauvin JC (1992) Carbon Xows in a subtidal Wne sand com-munity from the western English Channel: a simulation analysis.Mar Ecol Prog Ser 81:147–161
Chardy P, Gros P, Mercier H, Monbet Y (1993) Benthic carbon bud-gets for the Bay of Sant Brieuc (western channel): application ofinverse method. Oceanol Acta 16:687–694
Choat JH (1982) Fish feeding and the structure of benthic communitiesin temperate waters. Ann Rev Ecol Syst 13:31–42
Choat JH, Kingett PD (1982) The inXuence of Wsh predation on theabundance cycles of an algal turf invertebrate fauna. Oecologia54:88–95
Clarke KR (1993) Non-parametric multivariate analyses of changes incommunity structure. Aust J Ecol 181:103–122
Clarke KR, Warwick RM (1994) Change in marine communities: anapproach to statistical analysis and interpretation. Natural Envi-ronment Research Council, Plymouth
Commito JA, Shrader PB (1985) Benthic community response toexperimental additions of the polychaete Nereis virens. Mar Biol86:101–107
Commito JA, Tita G (2002) DiVerential dispersal rates in an intertidalmeiofauna assemblage. J Exp Mar Biol Ecol 268:237–256
Como S, Rossi F, Lardicci C (2004) Response of deposit-feeders toexclusion of epibenthic predators in a Mediterranean intertidalXat. J Exp Mar Biol Ecol 303:157–171
Connell SD (1997) Exclusion of predatory Wsh on a coral reef: theanticipation, pre-emption and evaluation of some caging artifacts.J Exp Mar Biol Ecol 213:181–198
Connell SD (2001) Predatory Wsh do not always aVect the earlydevelopment of epibiotic assemblages. J Exp Mar Biol Ecol260:1–12
Constable AJ (1999) Ecology of benthic macro-invertebrates in soft-sediment environments: a review of progress towards quantitativemodels and predictions. Aust J Ecol 24:452–476
Coull BC (1990) Are members of the meiofauna food for higher tro-phic levels? Trans Am Microsc Soc 109:233–246
Coull BC, Wells JBJ (1982) Refuges from Wsh predation: experimentswith phytal meiofauna from the New Zealand rocky intertidal.Ecology 64:1599–1609
Coull BC, Greenwood JG, Fielder DR, Coull BA (1995) SubtropicalAustralian juvenile Wsh eat meiofauna: experiments with winterwhiting Sillago maculata and observations on other species. MarEcol Prog Ser 125:13–19
Danovaro R, Fraschetti S (2002) Meiofaunal vertical zonation onrocky-bottoms: comparison with soft-bottom meiofauna. MarEcol Prog Ser 230:159–169
Danovaro R, Gambi C, Mirto S (2002) Meiofaunal production and en-ergy transfer eYciency in a seagrass Posidonia oceanica bed inthe western Mediterranean. Mar Ecol Prog Ser 234:95–104
Einbinder S, Perelberg A, Ben-Shaprut O, Foucart MH, Shashar N(2006) EVects of artiWcial reefs on Wsh grazing in their vicinity:Evidence from algae presentation experiments. Mar Environ Res61:110–119
Escaravage V, Garcia ME, Castel J (1989) The distribution ofmeiofauna and its contribution to detritic pathway inn tidal Xats(Arcachon Bay, France). In: Ros JD (ed) Topics in marine Biol-ogy. Sci Mar 53:551–559
Felsinga M, Glencrossa B, Telferb T (2005) Preliminary study on theeVects of exclusion of wild fauna from aquaculture cages in ashallow marine environment. Aquaculture 243:159–174
123

Mar Biol (2007) 152:339–350 349
Fernandes TF, Huxham M, Piper SR (1999) Predator caging experi-ments: a test of the importance of scale. J Exp Mar Biol Ecol241:137–154
Feller RJ (2006) Weak meiofaunal trophic linkages in Crangon cran-gon and Carcinus maenas. J Exp Mar Biol Ecol 330:274–283
Gerlach SA (1971) On the importance of marine meiofauna for ben-thos communities. Oecologia 6:176–190
Gibbons MJ (1988) Impact of predation by juvenile Clinus supercilio-sus on phytal meiofauna: are Wsh important as predators? MarEcol Prog Ser 45:13–22
Gilinsky E (1984) The role of Wsh predation and spatial heterogeneityin determining benthic community structure. Ecology 65:455–468
Gregg JC, Fleeger JW (1997) Importance of emerged and suspendedmeiofauna tot the diet of the darter goby (Gobionellus bolesomaJordan and Gilbert). J Exp Mar Biol Ecol 209:123–142
Heip C, Herman PM, Coomans A (1982) The productivity of the ma-rine meiobenthos. Academiae Analecta (Klasse der Wetenschap-pen) 44:1–20
Heip C, Herman PM, Vincx M (1983) Variability and production ofmeiobenthos in the Southern Bight of the North Sea. Proc VerbReun Int Explor Mer 183:51–56
Henry BA, Jenkins GP (1995) The impact of predation by the gridledgoby, Nesogobius sp1 on abundances of meiofauna and smallmacrofauna. J Exp Mar Biol Ecol 191:223–238
Hicks GFR (1984) Spatio-temporal dynamics of a meiobenthic cope-pod and the impact of predation-disturbance. J Exp Mar Biol Ecol8:47–72
Hindell JJ, Jenkins GJ, Keough MJ (2002) Variability in the numbersof post-settlement King George whiting (Sillaginidae: Sillagin-odes punctata, Cuvier) in relation to predation, habitat complexityand artiWcial cage structure. J Exp Mar Biol Ecol 268:13–31
Hines AH, Haddon AM, Wiechert LA (1990) Guild structure and for-aging impact of blue crab and epibenthic Wsh in a sub-estuary ofChesapeake Bay. Mar Ecol Prog Ser 67:105–126
Hixon MA (1997) EVects of reef Wshes on corals and algae. In: Birke-land C (ed) Life and death of coral reefs. Chapman and Hall, NewYork, pp 230–248
Hoyt M, Fleeger JW, Seibeling R, Feller RJ (2000) Serological estima-tion of prey-protein gut-residence time and quantiWcation of mealsize for grass shrimp consuming meiofaunal copepods. J Exp MarBiol Ecol 248:105–119
Irlandi EA, Mehlich ME (1996) The eVect of tissue cropping and dis-turbance by browsing Wshes on growth of two species of suspen-sion feeding bivalves. J Exp Mar Biol Ecol 197:279–293
Lasserre P, Renaud-Mornand J, Castel J (1976) Metabolic activities ofmeiofaunal communities in a semi-enclosed lagoon: possibilitiesof trophic competition between meiofauna and mugil Wsh. In:Persoone G, Jasper E (eds)Proceedings of the 10th EuropeanMarine Biology Symposium. Universa Press, Wetteren 2, pp393–414
Langlois TJ, Anderson MJ, Babcock RC, Kato S (2006) Marine re-serves demonstrate trophic interactions across habitats. Oecologia147:134–140
Kennelly SJ (1991) Caging experiments to examine the eVects of Wsheson understorey species in a sublittoral kelp community. J Exp MarBiol Ecol 147:207–230
Kent AC, Day RW (1983) Population dynamics of an infaunal poly-chaete: The eVects of predators on an adult-recruit interaction.J Exp Mar Biol Ecol 73:185–203
Lenihan HS, Micheli F (2001) Soft-sediment communities. In: Bert-ness MD, Gaines SM, Hixon ME (eds) Marine community ecol-ogy. Sinauer Associates, Sunderland, pp 253–287
Macia S (2000) The eVects of sea urchin grazing and drift algal bloomson a subtropical seagrass bed community. J Exp Mar Biol Ecol246:53–67
Mattila J (1992) Can Wsh predators regulate benthic communities onshallow soft bottoms in the Baltic Sea? The role of perch, ruVeand roach. PhD thesis, Abo Akademi University
Mattila J, BonsdorV E (1998) Predation by juvenile Xounder (Platich-thys Xesus L.): a test of prey vulnerability, predator preference,switching behavior and functional response. J Exp Mar Biol Ecol227:221–236
McArdle BH, Anderson MJ (2001) Fitting multivariate models tosemi-metric distances: a comment on distance-based redundancyanalysis. Ecology 82:290–297
McArthur VE (1998) Predation and the survival of juvenile Cerasto-derma glaucum Bruguiere (Mollusca: Bivalvia) in a coastallagoon. J Exp Mar Biol Ecol 225:79–97
McCall JN (1992) Source of harpacticoid copepods in the diet of juve-nile starry Xounder. Mar Ecol Prog Ser 86:41–50
McIntyre AD (1964) Meiobenthos of sublitoral muds. J Mar BiolAssoc UK 44:665–674
Menge BA (2000) Top-down and bottom-up community regulation inmarine rocky intertidal habitats. J Exp Mar Biol Ecol 250:257–289
Micheli F (1997) EVects of predator foraging behavior on patterns ofprey mortality in marine soft bottoms. Ecol Monogr 67:203–224
de Morais T, Bodiou JY (1984) Predation on meiofauna by juvenileWsh in a Western Mediterranean XatWsh nursery ground. Mar Biol82:209–215
OlaVson E, Elmgren R (1991) EVects of biological disturbance by ben-thic amphipods Monoporeira aVinis on meiobenthic communitystructure: a laboratory approach. Mar Ecol Prog Ser 74:99–107
Peterson CH (1979) Predation, competitive exclusion and diversity inthe soft-sediment benthic communities of estuaries and lagoons.In: Livingston RJ (ed) Ecological processes in coastal and marinesystems. Plenum, London
Posey MH, Alphin TD, Cahoon L (2006) Benthic community respons-es to nutrient enrichment and predator exclusion: inXuence ofbackground nutrient concentrations and interactive eVects. J ExpMar Biol Ecol 330:105–118
Quammen ML (1984) Predation by shorebirds, Wsh and crabs on inver-tebrates in intertidal mudXats: an experimental test. Ecology65:529–537
RaVaelli D, Milne H (1987) An experimental investigation of theeVects of shorebird and XatWsh predation on estuarine food web.Estuar Coast Shelf Sci 28:149–160
Reise K (1978) Experiments on epibenthic predation in the WaddenSea. Helgoland Mar Res 31:55–101
Reise K (1979) Moderate predation on meiofauna by the macrobenthosof the Wadden Sea. Helgoland Mar Res 32:453–465
Richards MG, Huxham M, Bryant A (1999) Predation: a causal mech-anism for variability in intertidal bivalve populations. J Exp MarBiol Ecol 241:159–177
Sarda R, Foreman K, Werme CE, Valiela I (1998) The impact of epi-faunal predation on the structure of macroinfaunal invertebratescommunities of tidal saltmarsh creeks. Estuar Coast Shelf Sci46:657–669
Schrijvers J, Vincx M (1997) Cage experiments in an East Africanmangrove forest: a synthesis. J Sea Res 38:123–133
Schrijvers J, Camargo MG, Pratiwi R, Vincx M (1998) The infaunalmacrobenthos under East African Cerips tagal mangroves impact-ed by epibenthos. J Exp Mar Biol Ecol 222:175–193
Shaw M, Jenkins GP (1992) Spatial variation in feeding, prey distribu-tion and food limitation of juvenile Xounder Rhombosolea tapirinaGünther. J Exp Mar Biol Ecol 165:1–21
Skilleter GA (1994) Refuges from predation and the persistence ofestuarine clam populations. Mar Ecol Prog Ser 109:29–42
Smith LD, Coull BC (1987) Juvenile spot (Pisces) and grass shrimppredation on meiobenthos in muddy and sandy substrata. J ExpMar Biol Ecol 105:123–136
123

350 Mar Biol (2007) 152:339–350
Spitzer PM, Mattila J, Heck KL Jr (2000) The eVects of vegetationdensity on the relative growth rates of juvenile pinWsh, Lagodonrhomboids (Linneaus), in Big Lagoon, Florida. J Exp Mar BiolEcol 244:67–86
Stocker LJ (1986) Artefactual eVects of caging on the recruitment andsurvivorship of a subtidal colonial invertebrate. Mar Ecol ProgSer 34:305–307
Thrush SF (1999) Complex role of predators in structuring soft-sedi-ment macrobenthic communities: implications of changes in spa-tial scale for experimental studies. Aust J Ecol 24:344–354
Thrush SF, Pridmore RD, Hewitt JE, Cummings VJ (1994) The impor-tance of predators on a sandXat: interplay between seasonalchanges in prey density and predators eVects. Mar Ecol Prog Ser107:211–222
Tita G, Desrosiers G, Vincx M, Nozais C (2000) Predation and sedi-ment disturbance eVects of the intertidal polychaetes Nereisvirens (Sars) on associated meiofaunal assemblages. J Exp MarBiol Ecol 243:261–282
Underwood AJ (2000) Experimental ecology of rocky intertidal habi-tats: what are we learning? J Exp Mar Biol Ecol 250:51–76
Warwick RM (1980) Population dynamics and secondary productionof the benthos in an estuarine environment. In: Tenore KR, CoullBC (eds) Marine benthic dynamics. University of South CarolinaPress, Columbia, pp 1–24
Warwick RM, Joint IR, Radford PJ (1979) Secondary production ofthe benthos in an estuarine environment. In: JeVeries RL, DaveyAJ (eds) Ecological processes in coastal environments. BlackwellScientiWc, Oxford, pp 429–450
Warwick RM, Clarke KR, Gee JM (1990) The eVect of disturbance bysoldier crabs Mictyris platycheles H. Milne Edwards on meioben-thic community structure. J Exp Mar Biol Ecol 135:19–33
Winer BJ, Brown DR, Michels KM (1991) Statistical principles inexperimental brown alga Pseudolithoderma sp. Journal design.Experimental marine biology and ecology, 3rd edn. McGraw-Hill, USA, 1057 p
123


















