
tRNA methyltransferases during embryogenesis in Musca domestica
4
DEVELOPMENTAL BIOLOGY 54, 304-307 (1976) tRNA Methyltransferases during Embryogenesis in Musca domestica RONALD E. LAW, ADOLPH J. FERRO, MICHAEL R. CUMMINGS, AND STANLEY K. SHAPIRO Department of Biological Sciences, University of Illinois at Chicago, Chicago, Illinois 60680 Accepted July 12,1976 Activity and diversity of the tRNA methyltransferases were examined during embryogene- sis of the housefly, Musca domestica. A rapid rise in the activity of the tRNA methyltransfer- ases was observed during the first 3 hr of embryogenesis. Activity increased slowly until the tenth hour of embryogenesis and then declined until hatching at 12 hr. The greatest diversity of tRNA methyltransferases, as indicated by extent of methylation, existed at 6 hr of embryo- genesis; the least diversity was observed in 1-hr embryos, while 12-hr embryos showed intermediate levels. Inhibition of embryonic tRNA methyltransferases at high concentrations of enzyme was observed in all extracts examined. INTRODUCTION Translational-level control of protein synthesis plays an important role in regu- lating early stages of embryonic develop- ment (5). In the sea urchin, when RNA synthesis is prevented by actinomycin D treatment, both protein synthesis and morphological development proceed nor- mally from fertilization to the swimming blastula stage (6). Cytoplasmic mRNA synthesized during oogenesis, therefore, contains sufficient information to direct early events in embryogenesis. Borek (2) has suggested that the methyl- ation of tRNA may participate in the regu- lation of development at the translational level. He observed that tRNA methyl- transferases are invariably constant in stable biological systems, while a variety of systems undergoing rapid cell prolifera- tion and/or differentiation exhibit marked quantitative and qualitative alterations in the tRNA methyltransferases (3). Regula- tory functions attributed to the methyla- tion of tRNA include the interaction of tRNA’s with aminoacyl tRNA synthetases (14) and tRNA coding response (4). Limited information exists concerning tRNA methyltransferases during embryo- genesis. In this communication we extend such studies through an investigation of tRNA methyltransferases during embryo- genesis of the housefly, Musca domestica. MATERIALS AND METHODS S-Adenosyl-L[14CH,lmethionine (sp act, 50 mCi/mmole) was purchased from Amer- sham-Searle. Escherichia coli B tRNA was the product of General Biochemicals. Collection of Embryos M. domestica of the Orlando wild-type strain were maintained in cages at 29°C as previously described (7). For the collection of embryos, an oviposition substrate con- sisting of a black cloth soaked in saturated ammonium carbonate placed over a finger bowl containing CSMA (Ralston Purina Co.) was introduced into the colony cage. Embryos collected over a 2-hr period were removed from the black cloth, weighed, and homogenized. Since the average age of embryos collected over a 2-hr period is 1 hr, these embryos have been designated as 1-hr embryos. Embryos were maintained at 29°C and embryogenesis was complete at 12 hr. Preparation of Enzyme Extracts A cell-free extract, from embryos of M. domestica, was prepared by homogenizing with 4 vol of 0.02 M potassium phosphate 304 Copyright 0 1976 by Academic Press, Inc. All rights of reproduction in any form reserved.
Transcript of tRNA methyltransferases during embryogenesis in Musca domestica

DEVELOPMENTAL BIOLOGY 54, 304-307 (1976)
tRNA Methyltransferases during Embryogenesis in Musca domestica
RONALD E. LAW, ADOLPH J. FERRO, MICHAEL R. CUMMINGS, AND STANLEY K. SHAPIRO
Department of Biological Sciences, University of Illinois at Chicago, Chicago, Illinois 60680
Accepted July 12,1976
Activity and diversity of the tRNA methyltransferases were examined during embryogene- sis of the housefly, Musca domestica. A rapid rise in the activity of the tRNA methyltransfer- ases was observed during the first 3 hr of embryogenesis. Activity increased slowly until the tenth hour of embryogenesis and then declined until hatching at 12 hr. The greatest diversity of tRNA methyltransferases, as indicated by extent of methylation, existed at 6 hr of embryo- genesis; the least diversity was observed in 1-hr embryos, while 12-hr embryos showed intermediate levels. Inhibition of embryonic tRNA methyltransferases at high concentrations of enzyme was observed in all extracts examined.
INTRODUCTION
Translational-level control of protein synthesis plays an important role in regu- lating early stages of embryonic develop- ment (5). In the sea urchin, when RNA synthesis is prevented by actinomycin D treatment, both protein synthesis and morphological development proceed nor- mally from fertilization to the swimming blastula stage (6). Cytoplasmic mRNA synthesized during oogenesis, therefore, contains sufficient information to direct early events in embryogenesis.
Borek (2) has suggested that the methyl- ation of tRNA may participate in the regu- lation of development at the translational level. He observed that tRNA methyl- transferases are invariably constant in stable biological systems, while a variety of systems undergoing rapid cell prolifera- tion and/or differentiation exhibit marked quantitative and qualitative alterations in the tRNA methyltransferases (3). Regula- tory functions attributed to the methyla- tion of tRNA include the interaction of tRNA’s with aminoacyl tRNA synthetases (14) and tRNA coding response (4).
Limited information exists concerning tRNA methyltransferases during embryo- genesis. In this communication we extend such studies through an investigation of
tRNA methyltransferases during embryo- genesis of the housefly, Musca domestica.
MATERIALS AND METHODS
S-Adenosyl-L[14CH,lmethionine (sp act, 50 mCi/mmole) was purchased from Amer- sham-Searle. Escherichia coli B tRNA was the product of General Biochemicals.
Collection of Embryos
M. domestica of the Orlando wild-type strain were maintained in cages at 29°C as previously described (7). For the collection of embryos, an oviposition substrate con- sisting of a black cloth soaked in saturated ammonium carbonate placed over a finger bowl containing CSMA (Ralston Purina Co.) was introduced into the colony cage. Embryos collected over a 2-hr period were removed from the black cloth, weighed, and homogenized. Since the average age of embryos collected over a 2-hr period is 1 hr, these embryos have been designated as 1-hr embryos. Embryos were maintained at 29°C and embryogenesis was complete at 12 hr.
Preparation of Enzyme Extracts
A cell-free extract, from embryos of M. domestica, was prepared by homogenizing with 4 vol of 0.02 M potassium phosphate
304 Copyright 0 1976 by Academic Press, Inc. All rights of reproduction in any form reserved.
BRIEF NOTES 305
(pH 6.8), 1 M dextrose, 0.004 M P-mercap- of 14CH, groups of S-adenosylmethionine toethanol, and 0.0004 M EDTA in a Ten- into E. coli B tRNA in the presence of broeck homogenizer. The 30,OOOg superna- saturating levels of enzyme (1). While tant was used as the source of tRNA meth- tRNA methyltransferase activity is a yltransferases. quantitative measurement, the extent of
Assay of tRNA Methyltransferases methylation reflects qualitative differ- ences among the individual base-, se- quence-, or tRNA-specific enzymes present in the heterologous population of tRNA methyltransferases.
The tRNA methyltransferases were as- sayed according to the method of Sharma et al. (13). The standard reaction mixture contained 5 pmoles of Tris-HCl, pH 8.2, 5 pmoles of MgCl*, 5 pmoles of P-mercapto- ethanol, 100 pg of E. coli B tRNA, 2 nmoles of (0.1 &i) S-adenosyl-L [‘4CH,]methionine, and 0.5 mg of protein in a total volume of 0.5 ml. Control tubes contained no tRNA. After incubation for 30 min at 37”C, the reaction was termi- nated by adding 2 ml of 10% TCA. Precipi- tates were collected on Whatman GF/C filters and washed 4x with 5 ml of cold 10% TCA and 5 ml of cold 95% ethanol. Filters were dried and counted in a Beck- man LS 250 liquid scintillation counter. Protein was determined by the method of Lowry et al. (9) using bovine serum albu- min as a standard.
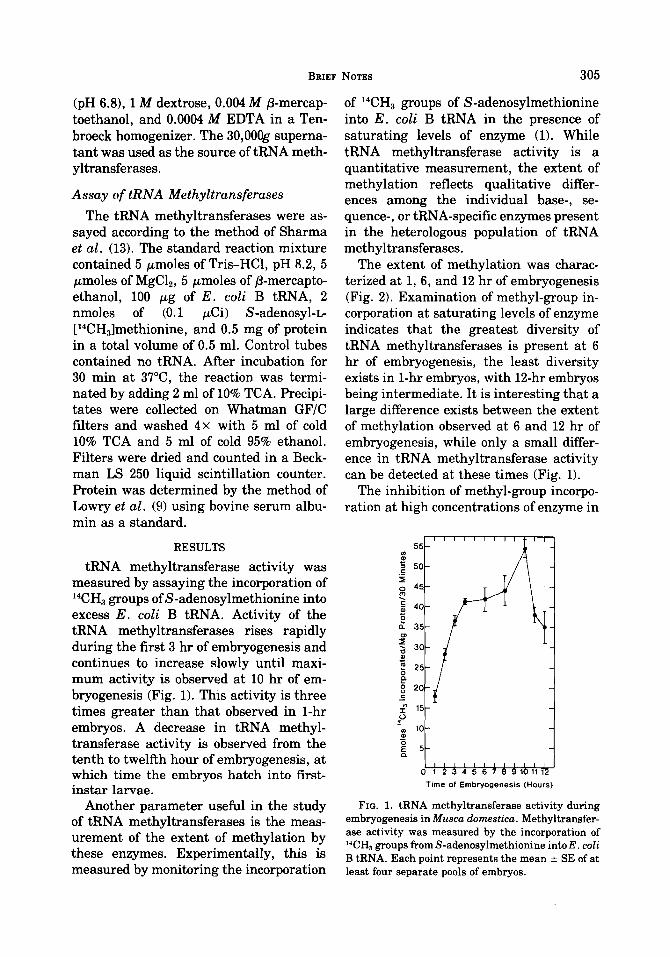
The extent of methylation was charac- terized at 1, 6, and 12 hr of embryogenesis (Fig. 2). Examination of methyl-group in- corporation at saturating levels of enzyme indicates that the greatest diversity of tRNA methyltransferases is present at 6 hr of embryogenesis, the least diversity exists in 1-hr embryos, with 12-hr embryos being intermediate. It is interesting that a large difference exists between the extent of methylation observed at 6 and 12 hr of embryogenesis, while only a small differ- ence in tRNA methyltransferase activity can be detected at these times (Fig. 1).
The inhibition of methyl-group incorpo- ration at high concentrations of enzyme in
RESULTS
tRNA methyltransferase activity was measured by assaying the incorporation of 14CH, groups ofS-adenosylmethionine into excess E. coli B tRNA. Activity of the tRNA methyltransferases rises rapidly during the first 3 hr of embryogenesis and continues to increase slowly until maxi- mum activity is observed at 10 hr of em- bryogenesis (Fig. 1). This activity is three times greater than that observed in 1-hr embryos. A decrease in tRNA methyl- transferase activity is observed from the tenth to twelfth hour of embryogenesis, at which time the embryos hatch into first- instar larvae.
Another parameter useful in the study of tRNA methyltransferases is the meas- urement of the extent of methylation by these enzymes. Experimentally, this is measured by monitoring the incorporation
Time of Embryogenesis (Hours)
FIG. 1. tRNA methyltransferase activity during embryogenesis in Musca domestica. Methyltransfer- ase activity was measured by the incorporation of WH, groups from S-adenosylmethionine into E. coli B tRNA. Each point represents the mean ? SE of at least four separate pools of embryos.

306 DEVELOPMENTAL BIOLOGY VOLUME 54, 1976
I I I I I
0.5 1.0 1.5 2.0 2.5 3.0
Protein Concentration (mg)
FIG. 2. Extent of tRNA methylation. The incor- poration of WH, groups from S-adenosylmethioninc into E. coli B tRNA was measured at 1 (O-O), 6 (O-O), and 12 (0-O) hr of embryogenesis at various protein concentrations. Each point repre- sents the average of two separate pools of embryos.
all embryonic extracts examined is also noteworthy (Fig. 2). This suggests the presence of an inhibitor whose activity de- pends on its absolute concentration. Simi- lar inhibition of tRNA methyltransferases at high concentrations of enzyme has been observed in the slime mold Dictyostelium discoideum (111, the sea urchin Strongylo- centrotus purpuratus (131, and adult rat liver (8). Sharma et al. (12) have suggested that the tRNA methyltransferases in de- velopment may be regulated through the presence or absence of such inhibitors.
DISCUSSION
Although elevated tRNA methyltrans- ferase activities have been reported in a wide variety of systems undergoing devel- opment and/or differentiation (3), this is the first observation of increased activity during embryogenesis. Our experiments on the extent of methylation suggest that elevated embryonic tRNA methyltransfer- ase activity is due, at least in part, to the appearance of novel-base and/or site-spe- cific enzymes during embryogenesis. The tRNA methyltransferases have previously been studied during sea urchin embryo- genesis by Sharma et al. (13), who mea- sured the extent of methylation in extracts from 15 min, and 4,24, and 48 hr postfertil-
ization. Their observation of a decrease in the extent of methylation during the course of embryogenesis differs from our observations in Musca. It appears, there- fore, that tRNA methyltransferase diver- sity decreases during sea urchin embryo- genesis. In Musca, however, a more com- plex pattern of tRNA methyltransferase diversity is observed. Six-hour embryos have a much greater tRNA methyltrans- ferase diversity than 1-hr embryos, while 12-hr embryos exhibit an intermediate di- versity.
In the sea urchin, inhibition of the tRNA methyltransferases at a high con- centration of enzyme was observed for all examined stages of embryogenesis (13). We report a similar observation in em- bryos of M. domestica. No information ex- ists concerning the nature of the inhibitors of embryonic tRNA methyltransferases.
Autoradiographic studies by Pietrushka and Bier (10) demonstrated that protein synthesis increases during the first 3 hr of embryogenesis in M. domestica. Addition- ally, the percentage of active ribosomes more than doubles during this period (Kinniburgh and Cummings, unpub- lished). Coincident increases in the activ- ity and diversity of tRNA methyltransfer- ases suggest that these enzymes partici- pate in the turn-on of embryonic protein synthesis in M. domestica.
This research was supported by Public Health Service Grant AM 14133 from the National Institute of Arthritis and Metabolic Diseases and by National Science Foundation Grant GB 36861.
REFERENCES
1. BALIGA, B. S., and BOREH, E. (1974). Endocri- nology 94, 815-821.
2. BOREK, E. (1963). Cold Spring Harbor Symp. Qua&. Biol. 28, 139-148.
3. BOREK, E., and KERR, S. J. (1972). Adv. Cancer Res. 15, 163-190.
4. CAPRA, J. D., and PETERKOFSKY, A. (1968). J. Mol. Biol. 33, 591-607.
5. GROSS, P. R. (1967). Curr. Top. Develop. Biol. 2, l-43.
6. GROSS, P. R. (1968). Ann. Rev. Biochem. 37,631- 660.
7. HALL, T. J., SANDERS, S. M., and CUMMINGS, M.

BRIEF NOTES 307
R. (1976). Insect Biochem. 6, 13-18. 8. KERR, S. J. (1970). B~OC~CWZ~S~F~ 9, 690-695. 9. LOWRY, 0. H., ROSENBROUGH, M., FARR, A., and
RANDALL, R. J. (1951). J. Biol. Chem. 193, 265-278.
10. PIETRUSCHKA, F., and BIER, K. (1972). Wilhelm Roux Archiu. 169, 56-69.
11. PILLENGER, D., and BOREK, E. (1969). PFOC. Nut.
Acao!. Sci. USA 62, 1145-1150. 12. SHARMA, 0. K., KERR, S. J., LIPSHITZ-WIESNER,
R., and BOREK, E. (1971). Fed. PFOC. 30, 167- 176.
13. SHARMA, 0. K., LOEB, L. A., and BOREK, E. (1971). Biochim. Biophys. A& 240,558-563.
14. SHUGART, L., NOVELLI, G. D., and STULBERG, M. P. (1968). Biochim. Biophys. Ada 157, 83-90.


















