
Treating seeds in menadione sodium bisulphite primes salt tolerance in Arabidopsis by inducing an...
8
Environmental and Experimental Botany 109 (2015) 23–30 Contents lists available at ScienceDirect Environmental and Experimental Botany jo ur nal homep ag e: www.elsevier.com/locate/envexpbot Treating seeds in menadione sodium bisulphite primes salt tolerance in Arabidopsis by inducing an earlier plant adaptation David Jiménez-Arias a , José A. Pérez b , Juan C. Luis c , Vanesa Martín-Rodríguez a , Francisco Valdés-González c , Andrés A. Borges a,∗ a Instituto de Productos Naturales y Agrobiología – CSIC, Avda. Astrofísico Francisco Sánchez 3, P.O. Box 195, 38206 La Laguna, Tenerife, Canary Islands, Spain b Instituto Universitario de Enfermedades Tropicales y Salud Pública, Universidad de La Laguna, Área de Genética. Avda. Astrofísico Francisco Sánchez s/n, 38271 La Laguna, Tenerife, Canary Islands, Spain c Grupo de Biología Vegetal Aplicada (GBVA), Departamento de Biología Vegetal – Facultad de Farmacia, Universidad de La Laguna, Avda. Astrofísico Francisco Sánchez s/n, 38071, La Laguna, Tenerife, Canary Islands, Spain a r t i c l e i n f o Article history: Received 25 March 2014 Received in revised form 29 May 2014 Accepted 28 July 2014 Available online 12 August 2014 Keywords: Abiotic stress Priming Proline accumulation Salt stress Soaking seeds a b s t r a c t For the majority of crops, salinity is one of the most important abiotic stresses, since about 20% of irrigated agricultural land is adversely affected by it. Menadione sodium bisulphite (MSB), a water-soluble vitamin K 3 o menadione derivative, has been previously reported as a plant defence activator against several pathogens in a number of species. We have further explored the MSB effects on salt tolerance. In this study, Arabidopsis thaliana wild ecotype Col-0 plants were exposed to prolonged salt (50 mM) stress. Salt treatment resulted in severe growth inhibition. This detrimental effect was lower in terms of relative growth rate (RGR) in plants from seeds soaked in 20 mM of MSB. In these plants, the drop in RGR was nearly 30% lower than untreated plants after 7 days in salt. Furthermore, we found that the salt stress imposed was not enough to disturb photosystem II or induce the expression of several detoxification genes. These functional impairments are characteristic of ionic injuries due to high levels of reactive oxygen species (ROS). At the end of the second week of the experiment, salt-treated plants recover RGR levels close to those of the control. Under our experimental conditions plants seem to be challenged by an osmotic stress with a minimum ionic imbalance. Those from MSB-treated seeds were primed to induce an earlier proline accumulation. Although no significant expression of ROS detoxification genes was found, several transcription factors involved in ROS signalling were detected after salt addition. In this context, MSB treatment was able to prime these transcription factors, resulting in an early adaptation of plants in response to salt stress. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Throughout evolution, plants have developed numerous defence mechanisms to endure environmental abiotic stresses such as drought, water excess, salinity, heat, cold and wounding, and to combat challenges arising from biotic stress (Bohnert et al., 2006). Abiotic stresses are considered to be the main cause of decreased agricultural yield (Ashraf et al., 2008). Drought and soil salinity are two major environmental stresses that limit crop pro- duction, affecting nearly 37% of the total agricultural land in the world (Munns and Tester, 2008; Reynolds and Tuberosa, 2008). Plants exposed to a mild chronic stress are able to induce specific ∗ Corresponding author. Tel.: +34 922 256 847; fax: +34 922 260 135. E-mail addresses: [email protected], [email protected] (A.A. Borges). morphogenic stress responses. These responses are characterized by a blockage of cell division in the main meristem tissues, an inhi- bition of cell elongation, and a redirected development of lateral organs (Potters et al., 2007). These stress induced morphogenic responses are part of a general adaptation strategy, whereby plants redirect their growth when exposed to stress. These stress responses are also characterized by the presence of antioxidants that prevent damage caused by reactive oxygen species (ROS). For instance, the accumulation of foliar anthocyanin acts as a mod- ulator of stress signals. Another strategy is the accumulation of compatible low molecular weight osmolytes, such as the amino acid L-proline (Steyn et al., 2002; Szabados and Savouré, 2009). Progress in plant genetic transformation and the availability of potentially useful genes from different sources has encouraged the use of transgenic approaches to minimize yield loss in many crops (Hirayama and Shinozaki, 2010). It is well known that salt http://dx.doi.org/10.1016/j.envexpbot.2014.07.017 0098-8472/© 2014 Elsevier B.V. All rights reserved.
Transcript of Treating seeds in menadione sodium bisulphite primes salt tolerance in Arabidopsis by inducing an...

Ti
DFa
Sb
3c
F
a
ARRAA
KAPPSS
1
dat2dsdwP
h0
Environmental and Experimental Botany 109 (2015) 23–30
Contents lists available at ScienceDirect
Environmental and Experimental Botany
jo ur nal homep ag e: www.elsev ier .com/ locate /envexpbot
reating seeds in menadione sodium bisulphite primes salt tolerancen Arabidopsis by inducing an earlier plant adaptation
avid Jiménez-Ariasa, José A. Pérezb, Juan C. Luisc, Vanesa Martín-Rodrígueza,rancisco Valdés-Gonzálezc, Andrés A. Borgesa,∗
Instituto de Productos Naturales y Agrobiología – CSIC, Avda. Astrofísico Francisco Sánchez 3, P.O. Box 195, 38206 La Laguna, Tenerife, Canary Islands,painInstituto Universitario de Enfermedades Tropicales y Salud Pública, Universidad de La Laguna, Área de Genética. Avda. Astrofísico Francisco Sánchez s/n,8271 La Laguna, Tenerife, Canary Islands, SpainGrupo de Biología Vegetal Aplicada (GBVA), Departamento de Biología Vegetal – Facultad de Farmacia, Universidad de La Laguna, Avda. Astrofísicorancisco Sánchez s/n, 38071, La Laguna, Tenerife, Canary Islands, Spain
r t i c l e i n f o
rticle history:eceived 25 March 2014eceived in revised form 29 May 2014ccepted 28 July 2014vailable online 12 August 2014
eywords:biotic stressrimingroline accumulationalt stressoaking seeds
a b s t r a c t
For the majority of crops, salinity is one of the most important abiotic stresses, since about 20% of irrigatedagricultural land is adversely affected by it. Menadione sodium bisulphite (MSB), a water-soluble vitaminK3 o menadione derivative, has been previously reported as a plant defence activator against severalpathogens in a number of species. We have further explored the MSB effects on salt tolerance. In thisstudy, Arabidopsis thaliana wild ecotype Col-0 plants were exposed to prolonged salt (50 mM) stress. Salttreatment resulted in severe growth inhibition. This detrimental effect was lower in terms of relativegrowth rate (RGR) in plants from seeds soaked in 20 mM of MSB. In these plants, the drop in RGR wasnearly 30% lower than untreated plants after 7 days in salt. Furthermore, we found that the salt stressimposed was not enough to disturb photosystem II or induce the expression of several detoxificationgenes. These functional impairments are characteristic of ionic injuries due to high levels of reactiveoxygen species (ROS). At the end of the second week of the experiment, salt-treated plants recover RGRlevels close to those of the control. Under our experimental conditions plants seem to be challenged by an
osmotic stress with a minimum ionic imbalance. Those from MSB-treated seeds were primed to induce anearlier proline accumulation. Although no significant expression of ROS detoxification genes was found,several transcription factors involved in ROS signalling were detected after salt addition. In this context,MSB treatment was able to prime these transcription factors, resulting in an early adaptation of plantsin response to salt stress.© 2014 Elsevier B.V. All rights reserved.
. Introduction
Throughout evolution, plants have developed numerousefence mechanisms to endure environmental abiotic stresses suchs drought, water excess, salinity, heat, cold and wounding, ando combat challenges arising from biotic stress (Bohnert et al.,006). Abiotic stresses are considered to be the main cause ofecreased agricultural yield (Ashraf et al., 2008). Drought and soilalinity are two major environmental stresses that limit crop pro-
uction, affecting nearly 37% of the total agricultural land in theorld (Munns and Tester, 2008; Reynolds and Tuberosa, 2008).lants exposed to a mild chronic stress are able to induce specific
∗ Corresponding author. Tel.: +34 922 256 847; fax: +34 922 260 135.E-mail addresses: [email protected], [email protected] (A.A. Borges).
ttp://dx.doi.org/10.1016/j.envexpbot.2014.07.017098-8472/© 2014 Elsevier B.V. All rights reserved.
morphogenic stress responses. These responses are characterizedby a blockage of cell division in the main meristem tissues, an inhi-bition of cell elongation, and a redirected development of lateralorgans (Potters et al., 2007). These stress induced morphogenicresponses are part of a general adaptation strategy, wherebyplants redirect their growth when exposed to stress. These stressresponses are also characterized by the presence of antioxidantsthat prevent damage caused by reactive oxygen species (ROS). Forinstance, the accumulation of foliar anthocyanin acts as a mod-ulator of stress signals. Another strategy is the accumulation ofcompatible low molecular weight osmolytes, such as the aminoacid L-proline (Steyn et al., 2002; Szabados and Savouré, 2009).
Progress in plant genetic transformation and the availabilityof potentially useful genes from different sources has encouragedthe use of transgenic approaches to minimize yield loss in manycrops (Hirayama and Shinozaki, 2010). It is well known that salt

2 and E
t(staarIgtdd
esPblepPps2sTm2vaa2r2
c(sdsdiepatbdtoiowt
2
2
iiast
4 D. Jiménez-Arias et al. / Environmental
olerance is a complex trait involving the function of many genesMunns and Tester, 2008), however the transgenic approaches pur-ue the development of salinity tolerant cultivars based on specificraits controlled by only one gene, e.g. a transcription factor orn important ion channel (Munns, 2011). Most of these strategiesre highly criticized due to poor evaluation methodology on car-ying out such studies under field conditions (Ashraf et al., 2008).n addition, although constitutive expression of a specific trans-ene can increase tolerance to stress conditions, this usually leadso a decrease in yield (Heil and Baldwin, 2002). It would be highlyesirable that defence genes were expressed only under stress con-itions.
Plants are able to develop numerous defence strategies tondure stress. Amongst them higher plants are capable of demon-trating some stress ‘memory’, or stress imprinting (Conrath, 2011;astor et al., 2013). Stress imprinting is usually defined as genetic oriochemical modifications induced by a first stress exposure that
eads to enhanced resistance to a later stress. Preliminary stressxposure is indeed known to enhance the stress tolerance of thelant through induction of adaptation responses (Conrath, 2011;astor et al., 2013), while tolerance can be linked to an array of mor-hological, physiological and biochemical responses that decreasetress damage or facilitate repair in damaged systems (Potters et al.,007). By using this “memory” or imprinting mechanism it is pos-ible to induce responses to a range of biotic and abiotic stresses.his is known as priming and is observed in plants and also in ani-als (Pham et al., 2007; Beckers and Conrath, 2007; Jung et al.,
009). Typically, primed plants display faster and/or stronger acti-ation of defence mechanism, which are induced in response tobiotic stresses or following attack by microbial pathogens (Beckersnd Conrath, 2007; Pham et al., 2007; Conrath, 2011 Pastor et al.,013). Consequently, priming could provide low-cost protection inelatively high-stress environmental conditions (Van-Hulten et al.,006).
Several molecules have been tested as priming agents whichan increase plant resistance to different and unrelated stressesMaffei et al., 2012; Filippou et al., 2013). Among them, menadioneodium bisulphite (MSB), a water-soluble vitamin K3 o menadioneerivative, is capable of inducing priming in Arabidopsis by foliarpray 24 h prior to inoculation with the virulent strain of Pseu-omonas syringae pv. tomato DC3000 or against downy mildewn pre-treated pearl millet seeds (Borges et al., 2009; Pushpalathat al., 2007), respectively. Additionally, our research group hasatented practical applications of MSB in agriculture (Borges-Péreznd Fernández-Falcón, 1995, 1996), including one for inducingolerance to salt stress (Borges et al., 2010) and several other MSB-ased formulations have been commercially developed as plantefence elicitors. Here we present MSB as a molecule which pro-ects plants against detrimental salt stress from the seed stagenward, through priming of existing defence mechanisms, avoid-ng undesirable resource allocation, energy costs and manipulationf the genome (Heil, 2002; Van-Hulten et al., 2006). The aim of thisork was to understand how the priming agent MSB potentiates
he Arabidopsis response to salinity stress (50 mM NaCl).
. Materials and methods
.1. In vitro conditions and MSB concentration
Arabidopsis thaliana ecotype Columbia-0 was used in the exper-ments. The MSB treatments were performed on seeds by soaking
n the MSB solution during seed stratification for 2 days at 4 ◦Ct the desired concentration (2, 20 and 40 mM of MSB). Controleeds were stratified with distilled water under identical condi-ions and both experimental treatments were sterilized accordingxperimental Botany 109 (2015) 23–30
to Martinez-Zapater and Salinas (1998). MSB effects on germina-tion ratio were studied in vitro on MS medium supplemented with0.7% agar in Petri dishes. Plates were placed in a growth chamberat 22 ◦C, 16 h light (100–110 �mol m−2 s−1) and 60–70% relativehumidity. Germination rates were assessed after 1 week of growth,while fresh weights of the above-ground plants were measuredafter 2 weeks. Each MSB treatment was applied to 50 seeds per plateand repeated in four independent experiments. Statistical analyseswere performed by one-way ANOVA and the significance of dif-ferences between experimental treatments was calculated using aTamhane post hoc test (IBM SPSS 20).
2.1.1. Hydroponic conditionsSalinity studies were performed under hydroponic conditions.
The containers (Araponics®, Liège, Belgium) were 1.9 L black plas-tic boxes accommodating 18 seed-holders with one plants each(Supplementary Fig. 1). A mixture of two granulometry sizes ofriver sand was used as a physical substrate. Seeds, each sown inseed-holders, were germinated in nursery trays filled with moist-ened river sand for 1 week in a growth chamber at 22 ◦C, 16 hlight (100–110 �mol m−2 s−1) and 100% relative humidity. Seed-holders with seedlings were transferred to the hydroponic systemin the same growth conditions but 60–70% relative humidity. Fol-lowing the previously optimized conditions, seedlings remainedwithout aeration during the first week, and then the nutri-ent solution was renewed every 7 days and gentle aerationapplied. The solution used was: KNO3 1.25 mM, KH2PO4 0.5 mM,MgSO4 × 7H2O 0.75 mM, Ca (NO3)2 × 4H2O 0.75 mM, H3BO350 �M, MnSO4 × H2O 10 �M, ZnSO4 × 7H2O 2 �M, CuSO4 × 5H2O1.5 �M, (NH4)6Mo7O24 × 4H2O 0.075 �M, Sequestrene® 44.8 �M(Syngenta, USA). Salinity treatments were applied 2 weeks aftertransferring seed-holders, by adding 50 mM NaCl to the nutrientsolution.
Supplementary Fig. 1 related to this article can befound, in the online version, at http://dx.doi.org/10.1016/j.envexpbot.2014.07.017.
2.2. Growth, chlorophylls, anthocyanin and prolinedeterminations
All these quantifications were restricted to the aerial part ofthe plants. After fresh weight (FW) measurement, harvested plantswere dried in a hot air oven at 70 ◦C for 72 h and dry weight (DW)determined. The relative growth rate (RGR) was calculated by usingthe following formula: RGR = [Ln (DWD2) − Ln (DWD1)]/D2 − D1,where D1 and D2 are the initial and the final time-points of theanalysis expressed in days.
Chlorophylls were analyzed in fully expanded leaves (50 mgFW), which were immediately grounded with liquid nitrogen andextracted with ice-cold acetone/water 85% (v/v) and 100% ace-tone. The extract was then centrifuged at 15,000 × g for 5 min andkept at −20 ◦C until analysis. Chlorophyll pigments were quanti-fied according to Porra (2002) and anthocyanin following Mita et al.(1997). Approximately 100 mg fresh tissue was collected in Eppen-dorf tubes, flash-frozen in liquid N2 and ground into powder. Oneml of 1% (v/v) hydrochloric acid in methanol was added to eachsample and the tubes were vigorously vortexed. After 1 day of incu-bation at 4 ◦C, the mixture was centrifuged at 15,000 × g for 15 minand absorbance of the supernatant was measured at 530 nm and657 nm. Proline content was determined as Bates et al. (1973) withminor modifications as follows. Briefly, proline was extracted fromapproximately 100 mg fresh tissue, which was immediately ground
in liquid nitrogen and extracted with 4 ml of 3% sulphosalicylic acid.Two �l of extract was reacted with 2 ml of acidic ninhydrin for60 min at 100 ◦C, stopping the reaction in an ice-bath. The reac-tion mixture was extracted with 4 ml of toluene, and vortexed.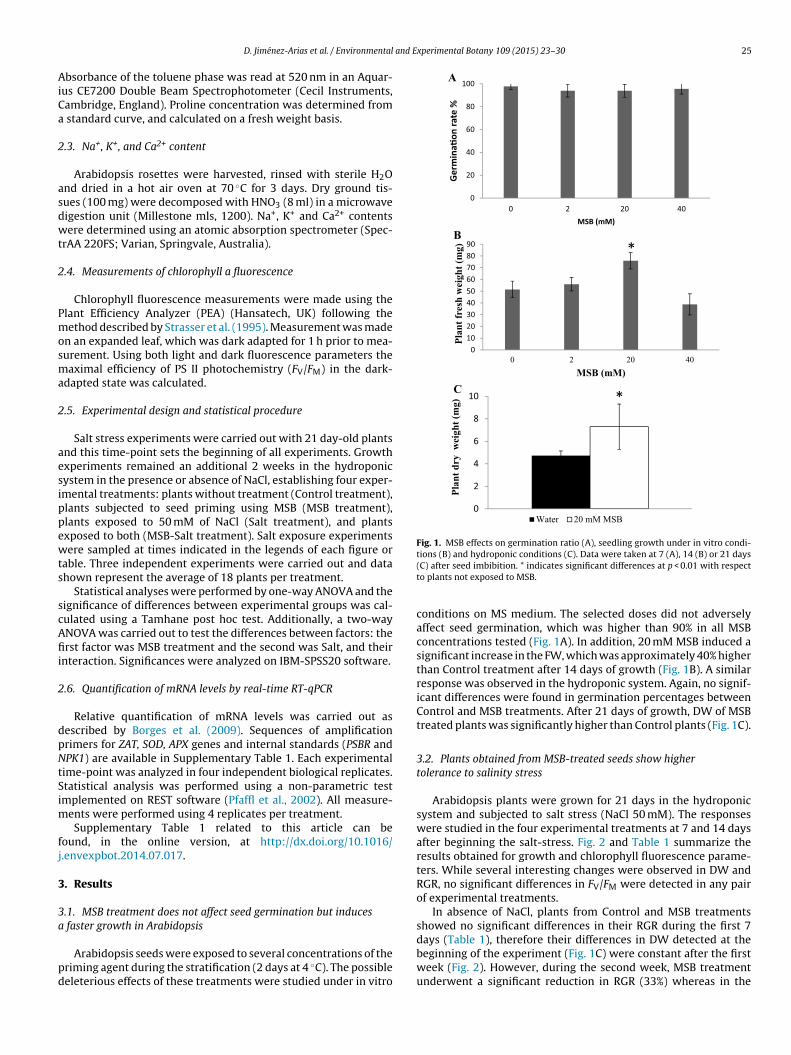
and Experimental Botany 109 (2015) 23–30 25
AiCa
2
asdwt
2
Pmosma
2
aesippewts
scAfii
2
dpNtSim
fj
3
3a
pd
0
20
40
60
80
100
0 2 20 40
Ger
min
a�on
rate
%
MSB (mM)
0102030405060708090
0 2 20 40
Plan
t fre
sh w
eigh
t (m
g)
MSB (mM)
*
0
2
4
6
8
10
Plan
t dry
wei
ght (
mg)
Water 20 mM MS B
*
A
B
C
Fig. 1. MSB effects on germination ratio (A), seedling growth under in vitro condi-tions (B) and hydroponic conditions (C). Data were taken at 7 (A), 14 (B) or 21 days
D. Jiménez-Arias et al. / Environmental
bsorbance of the toluene phase was read at 520 nm in an Aquar-us CE7200 Double Beam Spectrophotometer (Cecil Instruments,ambridge, England). Proline concentration was determined from
standard curve, and calculated on a fresh weight basis.
.3. Na+, K+, and Ca2+ content
Arabidopsis rosettes were harvested, rinsed with sterile H2Ond dried in a hot air oven at 70 ◦C for 3 days. Dry ground tis-ues (100 mg) were decomposed with HNO3 (8 ml) in a microwaveigestion unit (Millestone mls, 1200). Na+, K+ and Ca2+ contentsere determined using an atomic absorption spectrometer (Spec-
rAA 220FS; Varian, Springvale, Australia).
.4. Measurements of chlorophyll a fluorescence
Chlorophyll fluorescence measurements were made using thelant Efficiency Analyzer (PEA) (Hansatech, UK) following theethod described by Strasser et al. (1995). Measurement was made
n an expanded leaf, which was dark adapted for 1 h prior to mea-urement. Using both light and dark fluorescence parameters theaximal efficiency of PS II photochemistry (FV/FM) in the dark-
dapted state was calculated.
.5. Experimental design and statistical procedure
Salt stress experiments were carried out with 21 day-old plantsnd this time-point sets the beginning of all experiments. Growthxperiments remained an additional 2 weeks in the hydroponicystem in the presence or absence of NaCl, establishing four exper-mental treatments: plants without treatment (Control treatment),lants subjected to seed priming using MSB (MSB treatment),lants exposed to 50 mM of NaCl (Salt treatment), and plantsxposed to both (MSB-Salt treatment). Salt exposure experimentsere sampled at times indicated in the legends of each figure or
able. Three independent experiments were carried out and datahown represent the average of 18 plants per treatment.
Statistical analyses were performed by one-way ANOVA and theignificance of differences between experimental groups was cal-ulated using a Tamhane post hoc test. Additionally, a two-wayNOVA was carried out to test the differences between factors: therst factor was MSB treatment and the second was Salt, and their
nteraction. Significances were analyzed on IBM-SPSS20 software.
.6. Quantification of mRNA levels by real-time RT-qPCR
Relative quantification of mRNA levels was carried out asescribed by Borges et al. (2009). Sequences of amplificationrimers for ZAT, SOD, APX genes and internal standards (PSBR andPK1) are available in Supplementary Table 1. Each experimental
ime-point was analyzed in four independent biological replicates.tatistical analysis was performed using a non-parametric testmplemented on REST software (Pfaffl et al., 2002). All measure-
ents were performed using 4 replicates per treatment.Supplementary Table 1 related to this article can be
ound, in the online version, at http://dx.doi.org/10.1016/.envexpbot.2014.07.017.
. Results
.1. MSB treatment does not affect seed germination but induces faster growth in Arabidopsis
Arabidopsis seeds were exposed to several concentrations of theriming agent during the stratification (2 days at 4 ◦C). The possibleeleterious effects of these treatments were studied under in vitro
(C) after seed imbibition. * indicates significant differences at p < 0.01 with respectto plants not exposed to MSB.
conditions on MS medium. The selected doses did not adverselyaffect seed germination, which was higher than 90% in all MSBconcentrations tested (Fig. 1A). In addition, 20 mM MSB induced asignificant increase in the FW, which was approximately 40% higherthan Control treatment after 14 days of growth (Fig. 1B). A similarresponse was observed in the hydroponic system. Again, no signif-icant differences were found in germination percentages betweenControl and MSB treatments. After 21 days of growth, DW of MSBtreated plants was significantly higher than Control plants (Fig. 1C).
3.2. Plants obtained from MSB-treated seeds show highertolerance to salinity stress
Arabidopsis plants were grown for 21 days in the hydroponicsystem and subjected to salt stress (NaCl 50 mM). The responseswere studied in the four experimental treatments at 7 and 14 daysafter beginning the salt-stress. Fig. 2 and Table 1 summarize theresults obtained for growth and chlorophyll fluorescence parame-ters. While several interesting changes were observed in DW andRGR, no significant differences in FV/FM were detected in any pairof experimental treatments.
In absence of NaCl, plants from Control and MSB treatmentsshowed no significant differences in their RGR during the first 7
days (Table 1), therefore their differences in DW detected at thebeginning of the experiment (Fig. 1C) were constant after the firstweek (Fig. 2). However, during the second week, MSB treatmentunderwent a significant reduction in RGR (33%) whereas in the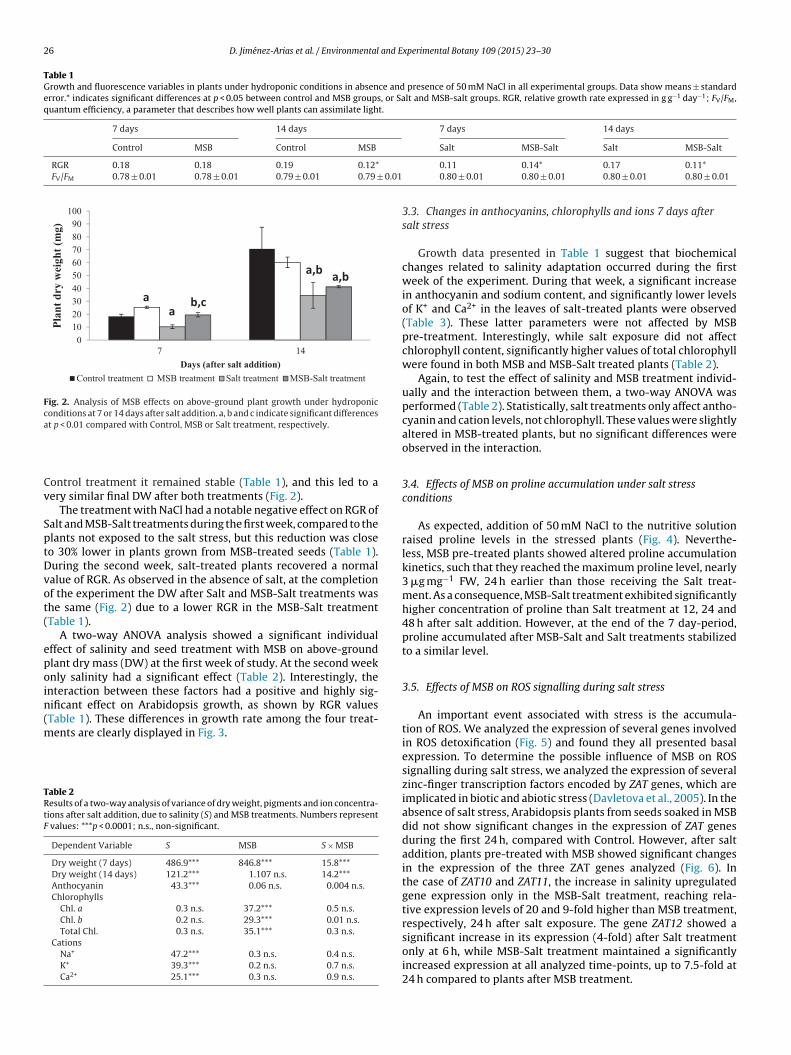
26 D. Jiménez-Arias et al. / Environmental and Experimental Botany 109 (2015) 23–30
Table 1Growth and fluorescence variables in plants under hydroponic conditions in absence and presence of 50 mM NaCl in all experimental groups. Data show means ± standarderror.* indicates significant differences at p < 0.05 between control and MSB groups, or Salt and MSB-salt groups. RGR, relative growth rate expressed in g g−1 day−1; FV/FM,quantum efficiency, a parameter that describes how well plants can assimilate light.
7 days 14 days 7 days 14 days
Control MSB Control MSB Salt MSB-Salt Salt MSB-Salt
RGR 0.18 0.18 0.19 0.12*
FV/FM 0.78 ± 0.01 0.78 ± 0.01 0.79 ± 0.01 0.79 ± 0.01
0102030405060708090
100
7 14
Plan
t dry
wei
ght (
mg)
Days (after salt addition)Control treatment MSB treatment Salt treatment MSB-Salt treatment
aa
a,b
b,c
a,b
Fig. 2. Analysis of MSB effects on above-ground plant growth under hydroponicconditions at 7 or 14 days after salt addition. a, b and c indicate significant differencesa
Cv
SptDvot(
epoin(m
TRtF
t p < 0.01 compared with Control, MSB or Salt treatment, respectively.
ontrol treatment it remained stable (Table 1), and this led to aery similar final DW after both treatments (Fig. 2).
The treatment with NaCl had a notable negative effect on RGR ofalt and MSB-Salt treatments during the first week, compared to thelants not exposed to the salt stress, but this reduction was closeo 30% lower in plants grown from MSB-treated seeds (Table 1).uring the second week, salt-treated plants recovered a normalalue of RGR. As observed in the absence of salt, at the completionf the experiment the DW after Salt and MSB-Salt treatments washe same (Fig. 2) due to a lower RGR in the MSB-Salt treatmentTable 1).
A two-way ANOVA analysis showed a significant individualffect of salinity and seed treatment with MSB on above-groundlant dry mass (DW) at the first week of study. At the second weeknly salinity had a significant effect (Table 2). Interestingly, thenteraction between these factors had a positive and highly sig-ificant effect on Arabidopsis growth, as shown by RGR valuesTable 1). These differences in growth rate among the four treat-
ents are clearly displayed in Fig. 3.
able 2esults of a two-way analysis of variance of dry weight, pigments and ion concentra-ions after salt addition, due to salinity (S) and MSB treatments. Numbers represent
values: ***p < 0.0001; n.s., non-significant.
Dependent Variable S MSB S × MSB
Dry weight (7 days) 486.9*** 846.8*** 15.8***Dry weight (14 days) 121.2*** 1.107 n.s. 14.2***Anthocyanin 43.3*** 0.06 n.s. 0.004 n.s.Chlorophylls
Chl. a 0.3 n.s. 37.2*** 0.5 n.s.Chl. b 0.2 n.s. 29.3*** 0.01 n.s.Total Chl. 0.3 n.s. 35.1*** 0.3 n.s.
CationsNa+ 47.2*** 0.3 n.s. 0.4 n.s.K+ 39.3*** 0.2 n.s. 0.7 n.s.Ca2+ 25.1*** 0.3 n.s. 0.9 n.s.
0.11 0.14* 0.17 0.11* 0.80 ± 0.01 0.80 ± 0.01 0.80 ± 0.01 0.80 ± 0.01
3.3. Changes in anthocyanins, chlorophylls and ions 7 days aftersalt stress
Growth data presented in Table 1 suggest that biochemicalchanges related to salinity adaptation occurred during the firstweek of the experiment. During that week, a significant increasein anthocyanin and sodium content, and significantly lower levelsof K+ and Ca2+ in the leaves of salt-treated plants were observed(Table 3). These latter parameters were not affected by MSBpre-treatment. Interestingly, while salt exposure did not affectchlorophyll content, significantly higher values of total chlorophyllwere found in both MSB and MSB-Salt treated plants (Table 2).
Again, to test the effect of salinity and MSB treatment individ-ually and the interaction between them, a two-way ANOVA wasperformed (Table 2). Statistically, salt treatments only affect antho-cyanin and cation levels, not chlorophyll. These values were slightlyaltered in MSB-treated plants, but no significant differences wereobserved in the interaction.
3.4. Effects of MSB on proline accumulation under salt stressconditions
As expected, addition of 50 mM NaCl to the nutritive solutionraised proline levels in the stressed plants (Fig. 4). Neverthe-less, MSB pre-treated plants showed altered proline accumulationkinetics, such that they reached the maximum proline level, nearly3 �g mg−1 FW, 24 h earlier than those receiving the Salt treat-ment. As a consequence, MSB-Salt treatment exhibited significantlyhigher concentration of proline than Salt treatment at 12, 24 and48 h after salt addition. However, at the end of the 7 day-period,proline accumulated after MSB-Salt and Salt treatments stabilizedto a similar level.
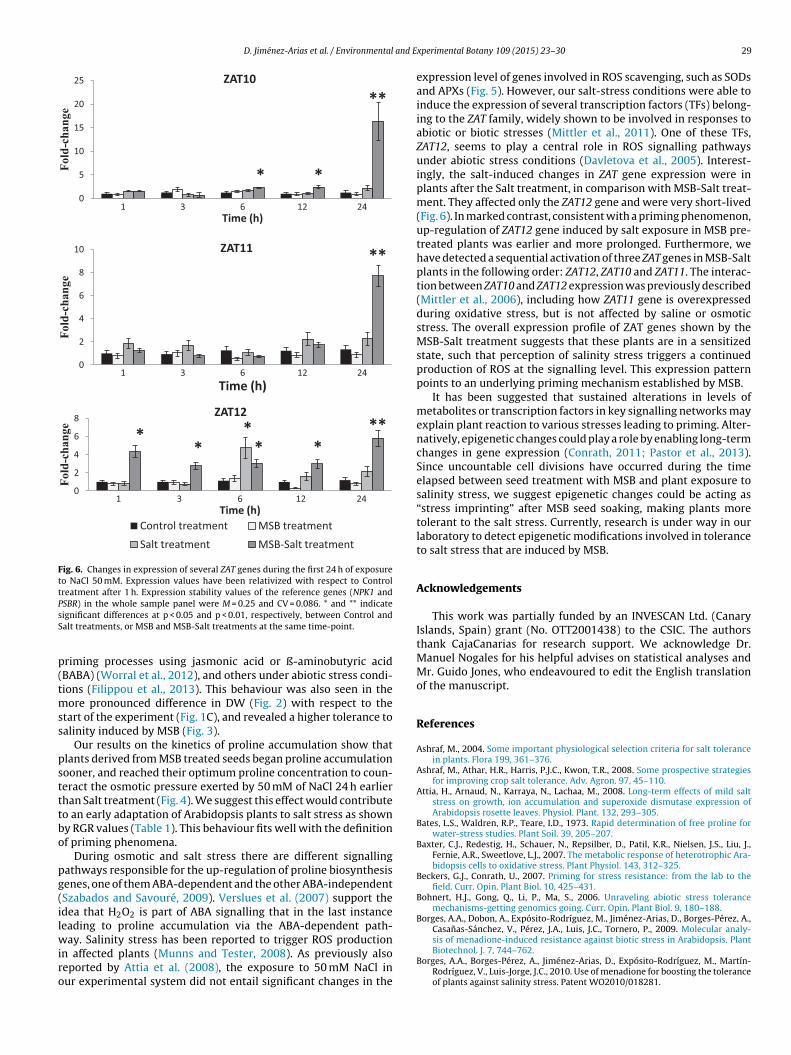
3.5. Effects of MSB on ROS signalling during salt stress
An important event associated with stress is the accumula-tion of ROS. We analyzed the expression of several genes involvedin ROS detoxification (Fig. 5) and found they all presented basalexpression. To determine the possible influence of MSB on ROSsignalling during salt stress, we analyzed the expression of severalzinc-finger transcription factors encoded by ZAT genes, which areimplicated in biotic and abiotic stress (Davletova et al., 2005). In theabsence of salt stress, Arabidopsis plants from seeds soaked in MSBdid not show significant changes in the expression of ZAT genesduring the first 24 h, compared with Control. However, after saltaddition, plants pre-treated with MSB showed significant changesin the expression of the three ZAT genes analyzed (Fig. 6). Inthe case of ZAT10 and ZAT11, the increase in salinity upregulatedgene expression only in the MSB-Salt treatment, reaching rela-tive expression levels of 20 and 9-fold higher than MSB treatment,respectively, 24 h after salt exposure. The gene ZAT12 showed a
significant increase in its expression (4-fold) after Salt treatmentonly at 6 h, while MSB-Salt treatment maintained a significantlyincreased expression at all analyzed time-points, up to 7.5-fold at24 h compared to plants after MSB treatment.
D. Jiménez-Arias et al. / Environmental and Experimental Botany 109 (2015) 23–30 27
F salt at
4
saegodmsT
TCnst
ig. 3. Appearance of plants from different experimental treatments after 7 days ofreatment (D).
. Discussion
Despite being considered a salinity-sensitive species, Arabidop-is thaliana can endure long periods of exposure to 50 mM NaClnd still accomplish its life cycle (Attia et al., 2008). After sev-ral days of salinity stress, a partial recovery of hydration androwth rate is generally observed, depending on the efficiencyf the adaptive mechanisms that protect the plant against the
etrimental effects of salt (Munns, 2011). In order to study theseechanisms we designed our experiments for a long-term expo-ure to a non-lethal salt concentration, to facilitate their expression.he effect of salt on plant growth in our experimental system was
able 3hanges in anthocyanin (units ml−1), chlorophyll (mg ml−1) and cation (mmol mg−1 dry wormal and salt stress conditions. Data show the mean values plus their standard deviatioalt treatments or MSB and MSB-salt treatments are indicated by * and **, and significanreatments are shown by † .
Control Salt
Anthocyanin 1.05 ± 0.46 3.91 ± 1.40Chlorophylls
Chl. a 0.44 ± 0.14 0.38 ± 0.10Chl. b 0.14 ± 0.05 0.13 ± 0.05Total Chl. 0.59 ± 0.19 0.52 ± 0.15
CationsNa+ 0.014 ± 0.001 2.07 ± 0.57K+ 12.46 ± 1.04 1.58 ± 0.08Ca2+ 17.8 ± 0.90 9.73 ± 1.12
ddition. Control treatment (A), MSB treatment (B), Salt treatment (C) and MSB-Salt
not long-lasting, as indicated by the recovery of RGR during thesecond week of NaCl exposure in plants grown after the Salt treat-ment, reaching a value close to Control plants (Table 1). This is theusual behaviour in the osmotic phase of salt stress (Munns andTester, 2008). Several of the monitored parameters indicate thatan ionic imbalance occurred due to the increased Na+ level, seenin decreased K+ and Ca2+ concentrations and anthocyanin accu-mulation (Munns, 2011). However, these stress indicators did not
lead to a significant reduction in chlorophyll levels or FV/FM val-ues (Tables 1 and 3, respectively), as would be expected under asevere ionic stress (Ashraf, 2004). Moreover, the basal expressionlevel of several genes involved in ROS detoxification in all analyzedeight) content of MSB treated and untreated Arabidopsis plants after 7 days undern. Significant differences at p < 0.05 and p < 0.01, respectively, between control and
t differences at p < 0.05 between MSB and control treatments, or MSB-salt and salt
MSB MSB-Salt
** 1.13 ± 0.52 4.04 ± 1.02**
0.72 ± 0.11† 0.73 ± 0.12†
0.25 ± 0.05† 0.25 ± 0.02†
0.98 ± 0.16† 0.98 ± 0.16†
** 0.012 ± 0.001 2.42 ± 0.37**
** 8.36 ± 0.99 2.01 ± 0.05**
* 20.7 ± 1.11 12.1 ± 1.01*

28 D. Jiménez-Arias et al. / Environmental and E
0
1
2
3
4
5
0 12 24 48 72 16 8
Prol
ine
(µM
)/FW
(mg)
Time (h)
Control treatment
MSB treatme nt
Salt tr eatment
MSB-Sal t treatme nt*
*
**
Fig. 4. Kinetics of proline accumulation in Salt and MSB-Salt treatments. * and **ip
thdes
eb1Mtwb
ponic system induced a decrease in the RGR in stressed plants
Ft
ndicate significant differences between Salt and MSB-Salt treatments at p < 0.05 and < 0.01, respectively.
reatments indicated that the level of toxic oxygen radicals was notigh after 50 mM salt addition (Fig. 5). These results are in accor-ance with Attia et al. (2008). Overall, the data suggest that in ourxperimental conditions plants are basically subjected to salinitytress with minimal oxidative injury.
In the present work we study the capacity of MSB to prime tol-rance to salt stress in Arabidopsis from the seed stage. MSB haseen described as an inducer of ROS stress in plants (Sun et al.,999; Baxter et al., 2007; Lehmann et al., 2008), but none of theSB treatments used in our optimization experiment had a nega-
ive effect on germination rate. Thus, there seems to be a relativelyide range of MSB concentrations with no toxic effects on Ara-
idopsis seeds. Moreover, plants derived from seeds exposed to
0
1
2
3
12 24
Fold
Cha
nge
Hours
APX1
0123
12 24
Fold
Cha
nge
Hour s
CSD1
0
1
2
12
Fold
Cha
nge
FSD
Control treatmentSalt tr eatment
ig. 5. Changes in expression of several APX and SOD genes during the first 24 h of exposureatment after 1 h. Expression stability values of the reference genes (NPK1 and PSBR) in
xperimental Botany 109 (2015) 23–30
20 mM MSB showed increased growth at least during the first weekof the experiment, both in fresh and dry weight (Figs. 1B and 2).This behaviour recalls a report of accelerated growth induced byMSB treatment in several agronomic species (Rama-Rao et al.,1985). Interestingly, we found that in plants from seeds exposedto MSB, total chlorophyll was significantly higher than in plantsfrom untreated seeds (Table 3). Leaf area and cell number increasedexponentially during most of the leaf development (Granier andTardieu, 1998). Moreover, vegetative growth parameters in Ara-bidopsis, such as leaf area, leaf number (Granier et al., 2002) andchlorophylls (Johansson and Nyström, 2004) increase continuallyuntil they reach a maximum value. These values remain constantuntil the plants enter inflorescence, the final stage of development.Chlorophyll levels and the significantly lower RGR observed in theMSB treatment during the last week of the experiment (Table 1)could be related to an earlier completion of vegetative growth inthese plants. Indeed, the rate of flowering in Arabidopsis plantsfrom MSB-treated seed is higher than untreated controls at the endof the experiment (approximately 70% vs. 20%). This is consistent itha previous study demonstrating that foliar-spray MSB treatmentsaccelerate flowering in banana crops (Borges-Pérez and Fernández-Falcón, 1996). Completing the life cycle in a shorter time is highlyadvantageous under stress conditions. This is because it reducesthe developmental period and may help to ensure the survival ofthe species, constituting a valuable ecological tolerance mechanism(Yaish et al., 2011).
In accordance with data published by other researchers (Attiaet al., 2008), we found that addition of NaCl 50 mM to the hydro-
during the first week of exposure to salt (Table 1). Neverthe-less, this reduction was significantly lower in MSB pre-treatedplants (approximately 30%). This increased salt tolerance resembles
0123
12 24Fold
Cha
nge
Hours
APX2
0123
12 24Fold
Cha
nge
Hours
CSD2
24Hour s
1
MSB treatmentMSB-Salt tr eatment
re to NaCl 50 mM. Expression values have been relativized with respect to Control the whole sample panel were M = 0.25 and CV = 0.086.
D. Jiménez-Arias et al. / Environmental and E
0
5
10
15
20
25
1 3 6 12 24
Fold
-cha
n ge
Time (h)
ZAT10
0
2
4
6
8
10
1 3 6 12 24
Fold
-cha
n ge
Time (h)
ZAT11
0
2
4
6
8
1 3 6 12 24
Fold
-cha
nge
Time (h)
ZAT12
Control treatme nt MSB treatme ntSalt treatment MSB -Sal t treatment
*
*
*
*
**
**
* ***
*
Fig. 6. Changes in expression of several ZAT genes during the first 24 h of exposureto NaCl 50 mM. Expression values have been relativized with respect to Controltreatment after 1 h. Expression stability values of the reference genes (NPK1 andPsS
p(tmss
pstttbo
pg(ilwiro
SBR) in the whole sample panel were M = 0.25 and CV = 0.086. * and ** indicateignificant differences at p < 0.05 and p < 0.01, respectively, between Control andalt treatments, or MSB and MSB-Salt treatments at the same time-point.
riming processes using jasmonic acid or ß-aminobutyric acidBABA) (Worral et al., 2012), and others under abiotic stress condi-ions (Filippou et al., 2013). This behaviour was also seen in the
ore pronounced difference in DW (Fig. 2) with respect to thetart of the experiment (Fig. 1C), and revealed a higher tolerance toalinity induced by MSB (Fig. 3).
Our results on the kinetics of proline accumulation show thatlants derived from MSB treated seeds began proline accumulationooner, and reached their optimum proline concentration to coun-eract the osmotic pressure exerted by 50 mM of NaCl 24 h earlierhan Salt treatment (Fig. 4). We suggest this effect would contributeo an early adaptation of Arabidopsis plants to salt stress as showny RGR values (Table 1). This behaviour fits well with the definitionf priming phenomena.
During osmotic and salt stress there are different signallingathways responsible for the up-regulation of proline biosynthesisenes, one of them ABA-dependent and the other ABA-independentSzabados and Savouré, 2009). Verslues et al. (2007) support thedea that H2O2 is part of ABA signalling that in the last instanceeading to proline accumulation via the ABA-dependent path-
ay. Salinity stress has been reported to trigger ROS productionn affected plants (Munns and Tester, 2008). As previously alsoeported by Attia et al. (2008), the exposure to 50 mM NaCl inur experimental system did not entail significant changes in the
xperimental Botany 109 (2015) 23–30 29
expression level of genes involved in ROS scavenging, such as SODsand APXs (Fig. 5). However, our salt-stress conditions were able toinduce the expression of several transcription factors (TFs) belong-ing to the ZAT family, widely shown to be involved in responses toabiotic or biotic stresses (Mittler et al., 2011). One of these TFs,ZAT12, seems to play a central role in ROS signalling pathwaysunder abiotic stress conditions (Davletova et al., 2005). Interest-ingly, the salt-induced changes in ZAT gene expression were inplants after the Salt treatment, in comparison with MSB-Salt treat-ment. They affected only the ZAT12 gene and were very short-lived(Fig. 6). In marked contrast, consistent with a priming phenomenon,up-regulation of ZAT12 gene induced by salt exposure in MSB pre-treated plants was earlier and more prolonged. Furthermore, wehave detected a sequential activation of three ZAT genes in MSB-Saltplants in the following order: ZAT12, ZAT10 and ZAT11. The interac-tion between ZAT10 and ZAT12 expression was previously described(Mittler et al., 2006), including how ZAT11 gene is overexpressedduring oxidative stress, but is not affected by saline or osmoticstress. The overall expression profile of ZAT genes shown by theMSB-Salt treatment suggests that these plants are in a sensitizedstate, such that perception of salinity stress triggers a continuedproduction of ROS at the signalling level. This expression patternpoints to an underlying priming mechanism established by MSB.
It has been suggested that sustained alterations in levels ofmetabolites or transcription factors in key signalling networks mayexplain plant reaction to various stresses leading to priming. Alter-natively, epigenetic changes could play a role by enabling long-termchanges in gene expression (Conrath, 2011; Pastor et al., 2013).Since uncountable cell divisions have occurred during the timeelapsed between seed treatment with MSB and plant exposure tosalinity stress, we suggest epigenetic changes could be acting as“stress imprinting” after MSB seed soaking, making plants moretolerant to the salt stress. Currently, research is under way in ourlaboratory to detect epigenetic modifications involved in toleranceto salt stress that are induced by MSB.
Acknowledgements
This work was partially funded by an INVESCAN Ltd. (CanaryIslands, Spain) grant (No. OTT2001438) to the CSIC. The authorsthank CajaCanarias for research support. We acknowledge Dr.Manuel Nogales for his helpful advises on statistical analyses andMr. Guido Jones, who endeavoured to edit the English translationof the manuscript.
References
Ashraf, M., 2004. Some important physiological selection criteria for salt tolerancein plants. Flora 199, 361–376.
Ashraf, M., Athar, H.R., Harris, P.J.C., Kwon, T.R., 2008. Some prospective strategiesfor improving crop salt tolerance. Adv. Agron. 97, 45–110.
Attia, H., Arnaud, N., Karraya, N., Lachaa, M., 2008. Long-term effects of mild saltstress on growth, ion accumulation and superoxide dismutase expression ofArabidopsis rosette leaves. Physiol. Plant. 132, 293–305.
Bates, L.S., Waldren, R.P., Teare, I.D., 1973. Rapid determination of free proline forwater-stress studies. Plant Soil. 39, 205–207.
Baxter, C.J., Redestig, H., Schauer, N., Repsilber, D., Patil, K.R., Nielsen, J.S., Liu, J.,Fernie, A.R., Sweetlove, L.J., 2007. The metabolic response of heterotrophic Ara-bidopsis cells to oxidative stress. Plant Physiol. 143, 312–325.
Beckers, G.J., Conrath, U., 2007. Priming for stress resistance: from the lab to thefield. Curr. Opin. Plant Biol. 10, 425–431.
Bohnert, H.J., Gong, Q., Li, P., Ma, S., 2006. Unraveling abiotic stress tolerancemechanisms-getting genomics going. Curr. Opin. Plant Biol. 9, 180–188.
Borges, A.A., Dobon, A., Expósito-Rodríguez, M., Jiménez-Arias, D., Borges-Pérez, A.,Casanas-Sánchez, V., Pérez, J.A., Luis, J.C., Tornero, P., 2009. Molecular analy-
sis of menadione-induced resistance against biotic stress in Arabidopsis. PlantBiotechnol. J. 7, 744–762.Borges, A.A., Borges-Pérez, A., Jiménez-Arias, D., Expósito-Rodríguez, M., Martín-Rodríguez, V., Luis-Jorge, J.C., 2010. Use of menadione for boosting the toleranceof plants against salinity stress. Patent WO2010/018281.

3 and E
B
B
C
D
F
G
G
H
H
H
J
J
L
M
M
M
M
M
0 D. Jiménez-Arias et al. / Environmental
orges-Pérez, A., Fernández-Falcón, M.J., 1995. Compositions for inducing resistanceto tracheomycosis in plants. Patent WO95/0372.
orges-Pérez, A., Fernández-Falcón, M.J., 1996. Utilization of compositions whichcontain menadione for the stimulation of plant metabolism in order to inducetheir resistance to pathogen and pest and/or accelerate their blooming. PatentWO96/28026.
onrath, U., 2011. Molecular aspects of defence priming. Trends Plant Sci. 16,524–531.
avletova, S., Schlauch, K., Coutu, J., Mittler, R., 2005. The zinc-finger protein Zat12plays a central role in reactive oxygen and abiotic stress signaling in Arabidopsis.Plant Physiol. 139, 847–856.
ilippou, P., Tanou, G., Molassiotis, A., Fotopoulos, V., 2013. Plant acclimation to envi-ronmental stress using priming agents. In: Narendra, T., Sarvajeet, S.G. (Eds.),Plant Acclimation to Environmental Stress. Springer, New York, pp. 1–28.
ranier, C., Tardieu, F., 1998. Is thermal time adequate for expressing the effect oftemperature on sunflower leaf development? Plant Cell Environ. 21, 695–703.
ranier, C., Massonet, C., Turc, O., Muller, B., Chenu, K., Tardie, F., 2002. Individual leafdevelopment in Arabidopsis thaliana: a stable thermal-time based programme.Ann. Bot. (Lond.) 89, 595–604.
eil, M., 2002. Ecological cost of induced resistance. Curr. Opin. Plant Biol. 5,5345–5350.
eil, M., Baldwin, T., 2002. Fitness cost of induced resistance: emerging experimentalsupport for a slippery concept. Trends Plant Sci. 7, 61–67.
irayama, T., Shinozaki, K., 2010. Research on plant abiotic stress responses in thepost-genome era: past, present and future. Plant J. 61, 1041–1052.
ohansson, E., Nyström, T., 2004. Progression and specificity of protein oxidation inthe life cycle of Arabidopsis thaliana. J. Biol. Chem. 279, 22204–22208.
ung, H.W., Tschaplinski, T.J., Wang, L., Glazebrook, J., Greenberg, J.T., 2009. Primingin systemic plant immunity. Science 324, 89–91.
ehmann, M., Schwarzlander, M., Obata, T., Sirikantaramas, S., Burow, M., Olsen,C.E., Tohge, T., Fricker, M.D., Moller, B.L., Fernie, A.R., Sweetlove, L.J., Laxa, M.,2008. The metabolic response of Arabidopsis roots to oxidative stress is distinctfrom that of heterotrophic cells in culture and highlights a complex relationshipbetween the levels of transcripts, metabolites, and flux. Mol. Plant 3, 390–406.
affei, M.E., Gen-Ichiro, A., Mithöfer, A., 2012. Natural elicitors, effectors and mod-ulators of plant responses. Nat. Prod. Rep. 29, 1288–1303.
artinez-Zapater, J.M., Salinas, J., 1998. Arabidopsis Protocols. Humana Press; NewJersey.
ita, S., Murano, N., Akaike, M., Nakamura, K., 1997. Mutants of Arabidopsis thalianawith pleiotropic effects on the expression of the gene for beta-amylase andon the accumulation of anthocyanin that are inducible by sugars. Plant J. 11,841–851.
ittler, R., Kim, Y.S., Song, L., Coutu, J., Coutu, A., Ciftci-Yilmaz, S., Lee, H., Stevenson,
B., 2006. Gain and loss-of-function mutations in Zat10 enhance the tolerance ofplants to abiotic stress. FEBS Lett. 580, 6537–6542.ittler, R., Vanderauwera, S., Suzuki, N., Miller, G., Tognetti, V.B., Vandepoele, K.,Gollery, M., Shulaev, V., Van-Breusegem, F., 2011. ROS signalling: the new wave?Trends Plant Sci. 16, 300–309.
xperimental Botany 109 (2015) 23–30
Munns, R., 2011. Plant adaptations to salt and water stress: differences and com-monalities. Adv. Bot. Res. 57, 2–26.
Munns, R., Tester, M., 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol.59, 651–681.
Pastor, V., Luna, E., Mauch-Mani, B., Ton, J., Flors, V., 2013. Primed plants do notforget. Environ. Exp. Bot. 94, 46–56.
Pfaffl, M.W., Horgan, W., Dempfle, L., 2002. Relative expression software tool (REST)for group-wise comparison and statistical analysis of relative expression resultsin real-time PCR. Nucleic Acids Res. 30, e36.
Pham, L.N., Dionne, M.S., Shirasu-Hiza, M., Schneider, D.S., 2007. A specific primedimmune response in Drosophila is dependent on phagocytes. PLoS Pathog. 3,e26.
Porra, R.J., 2002. The chequered history of the development and use of simultaneousequations for the accurate determination of chlorophylls a and b. Photosynth.Res. 73, 149–156.
Potters, G., Pasternak, T.P., Guisez, Y., Palme, K.J., Jansen, A.K., 2007. Stress-inducedmorphogenic responses: growing out of trouble? Trends Plant Sci. 12, 98–105.
Pushpalatha, H., Mythrashree, S., Shetty, R., Geetha, N., Sharathchandra, R.,Amruthesh, K., Shetty, H., 2007. Ability of vitamins to induce downy mildewdisease resistance and growth promotion in pearl millet. Crop Prot. 26,1674–1681.
Rama-Rao, A.V., Ravichandra, K., David, S.B., Ranade, S., 1985. Menadione sodiumbisulphite: a promising plant growth regulator. Plant Growth Regul. 3,111–118.
Reynolds, M., Tuberosa, R., 2008. Translational research impacting on crop produc-tivity in drought-prone environments. Curr. Opin. Plant Biol. 11, 171–179.
Steyn, W.J., Wand, S.J.E., Holcroft, D.M., Jacobs, G., 2002. Anthocyanins in vegeta-tive tissues: a proposed unified function in photoprotection. New Phytol. 155,349–361.
Strasser, R.J., Srivastava, A., Govindjee, G., 1995. Polyphasic chlorophyll a fluores-cence transient in plants and cyanobacteria. Photochem. Photobiol. 61, 32–42.
Sun, Y.L., Zhao, Y., Hong, X., Zhai, Z.H., 1999. Cytochrome c release and cas-pase activation during menadione-induced apoptosis in plants. FEBS Lett. 462,317–321.
Szabados, L., Savouré, A., 2009. Proline: a multifunctional amino acid. Trends PlantSci. 15, 89–97.
Van-Hulten, M., Pelser, M., Van-Loon, L.C., Pieterse, C.M., Ton, J., 2006. Costs andbenefits of priming for defence in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 103,5602–5607.
Verslues, P.E., Kim, Y.S., Zhu, J.K., 2007. Altered ABA, proline and hydrogen peroxidein an Arabidopsis glutamate: glyoxylate aminotransferase mutant. Plant Mol.Biol. 64, 205–217.
Worral, D., Holroyd, G.H., Moore, J.P., Glowacz, M., Croft, P., Taylor, J.E., Paul, N.D.,
Roberts, M.R., 2012. Treating seeds with activators of plant defence generateslong lasting priming of resistance to pests and pathogens. New Phytol. 193,770–778.Yaish, M.W., Colosanti, J., Rothstein, S.J., 2011. The role of epigenetic processes incontrolling flowering time in plants exposed to stress. J. Exp. Bot. 62, 3727–3735.


















