
TRANSLOCATIONS IN DICTYOSTELIUM DISCOIDEUMTRANSLOCATIONS IN DICTYOSTELIUM DISCOIDEUM D. L. WELKER'...
24
Copyright 0 1985 by the Genetics Society of America TRANSLOCATIONS IN DICTYOSTELIUM DISCOIDEUM D. L. WELKER' AND K. L. WILLIAMS' Max Planck Institut fur Biochemie, 8033 Martinsried bei Munchen, Federal Republic of Germany Manuscript received March 1, 1984 Revised copy accepted October 5, 1984 ABSTRACT Fourteen translocations of independent origin were identified in Dictyoste- lium discoideum on the basis of segregation anomalies of, diploids heterozygous for these chromosome rearrangements, all of which led to the cosegregation of unlinked markers. Many of these translocations were discovered in strains mutagenized with MNNG or in strains carrying mutations affecting DNA re- pair; however, spontaneous translocations were also obtained. Haploid mitotic recombinants of the rearranged linkage groups were produced from diploids heterozygous for the translocations at frequencies of up to 5% of viable haploid segregants; this is at least a ten-fold higher frequency than that seen with diploids not heterozygous for translocations (-0.1 %). These haploid recombi- nants included both translocated and nontranslocated strains. The T354(II,VIZ) translocation and possibly the T357(IV,VII) translocation reduce the chromo- some number to n = 6; haploids carrying 11 other translocations all have karyotypes with n = 7. Genetic characterization of the T357(IV,VII) transloca- tion showed that the bwnA and whiC loci normally found on linkage group IV were physically linked to the linkage group VI1 loci couA, PhgA, bsgB and cobA. OLITARY amoebas of the cellular slime mould Dictyostelium discoideum feed S by phagocytosis of bacteria but on starvation aggregate to form a multi- cellular fruiting body consisting of spores, stalk and basal disc cells. This tran- sition between unicellular and multicellular stages has been the subject of numerous developmental and biochemical studies (LOOMIS 1982). In the last decade, parasexual genetic analysis of this eukaryotic microorganism has ad- vanced to the stage where more than 100 genetic markers have been assigned to six well-defined linkage groups (NEWELL 1982; GODFREY and SUSSMAN 1982). Two recent reports describe mutations that may map to a seventh group (WALLACE and NEWELL 1982a; KASBEKAR, MADIGAN and KATZ 1983). More than 40 genetic markers have been ordered on five linkage groups (WELKER and WILLIAMS 1982a; WALLACE and NEWELL 1982a). The availability of gene order information plus the capacity for cytological examination (ROBSON and WILLIAMS 1977; WELKER and WILLIAMS 1980a) aids the characterization of chromosome abnormalities. Aneuploidy (WILLIAMS, ROBSON and WELKER 1980; D. L. WELKER, unpublished results), fusion chro- mosomes (WELKER and WILLIAMS 1981a) and a tandem duplication (WELKER, I Present address: Departnient of Biology, Utah State University, UMC 55, Logan, Utah. Present address: School of Biological Sciences, Macquarie University, North Ryde, 21 13 New South Wales, Australia. Genetics 109 341-364 February, 1985.
Transcript of TRANSLOCATIONS IN DICTYOSTELIUM DISCOIDEUMTRANSLOCATIONS IN DICTYOSTELIUM DISCOIDEUM D. L. WELKER'...

Copyright 0 1985 by the Genetics Society of America
TRANSLOCATIONS IN DICTYOSTELIUM DISCOIDEUM
D. L. WELKER' AND K. L. WILLIAMS'
Max Planck Institut f u r Biochemie, 8033 Martinsried bei Munchen, Federal Republic of Germany
Manuscript received March 1, 1984 Revised copy accepted October 5, 1984
ABSTRACT
Fourteen translocations of independent origin were identified in Dictyoste- lium discoideum on the basis of segregation anomalies of, diploids heterozygous for these chromosome rearrangements, all of which led to the cosegregation of unlinked markers. Many of these translocations were discovered in strains mutagenized with MNNG or in strains carrying mutations affecting DNA re- pair; however, spontaneous translocations were also obtained. Haploid mitotic recombinants of the rearranged linkage groups were produced from diploids heterozygous for the translocations at frequencies of up to 5% of viable haploid segregants; this is at least a ten-fold higher frequency than that seen with diploids not heterozygous for translocations (-0.1 %). These haploid recombi- nants included both translocated and nontranslocated strains. The T354(II,VIZ) translocation and possibly the T357(IV,VII) translocation reduce the chromo- some number to n = 6; haploids carrying 11 other translocations all have karyotypes with n = 7. Genetic characterization of the T357(IV,VII) transloca- tion showed that the bwnA and whiC loci normally found on linkage group IV were physically linked to the linkage group VI1 loci couA, PhgA, bsgB and cobA.
OLITARY amoebas of the cellular slime mould Dictyostelium discoideum feed S by phagocytosis of bacteria but on starvation aggregate to form a multi- cellular fruiting body consisting of spores, stalk and basal disc cells. This tran- sition between unicellular and multicellular stages has been the subject of numerous developmental and biochemical studies (LOOMIS 1982). In the last decade, parasexual genetic analysis of this eukaryotic microorganism has ad- vanced to the stage where more than 100 genetic markers have been assigned to six well-defined linkage groups (NEWELL 1982; GODFREY and SUSSMAN 1982). Two recent reports describe mutations that may map to a seventh group (WALLACE and NEWELL 1982a; KASBEKAR, MADIGAN and KATZ 1983). More than 40 genetic markers have been ordered on five linkage groups (WELKER and WILLIAMS 1982a; WALLACE and NEWELL 1982a).
The availability of gene order information plus the capacity for cytological examination (ROBSON and WILLIAMS 1977; WELKER and WILLIAMS 1980a) aids the characterization of chromosome abnormalities. Aneuploidy (WILLIAMS, ROBSON and WELKER 1980; D. L. WELKER, unpublished results), fusion chro- mosomes (WELKER and WILLIAMS 1981a) and a tandem duplication (WELKER,
I Present address: Departnient of Biology, Utah State University, UMC 55, Logan, Utah. Present address: School of Biological Sciences, Macquarie University, North Ryde, 21 13 New South Wales,
Australia.
Genetics 109 341-364 February, 1985.

342 D. L. WELKER AND K. L. WILLIAMS
METZ and WILLIAMS 1982) have already been identified. In addition, a number of translocations have been proposed on the basis of cosegregation of normally unlinked markers (WELKER, METZ and WILLIAMS 1982; NEWELL and ROSS 1982). These cosegregation patterns are thought to arise due to the production of inviable duplication-deficiency progeny, which carry a duplication for part of one affected linkage group but only an incomplete copy of the second. Such inviable progeny are formed by the segregation into the same haploid nucleus of one normal chromosome and one rearranged chromosome. In this report we examine the origin of these previously reported translocations and several new ones. This leads to some generalizations about how translocations arise in D. discoideum. A detailed examination of a translocation of spontaneous origin [T357(IV,VZI)] is also presented.
MATERIALS AND METHODS
Culture of D. discoideum: All strains were maintained in association with Klebsiella aerogenes on SM agar at 21 & l o as described previously (WELKER and WILLIAMS 1982a). Genotypes of haploid strains are provided in Table 1; all were derived solely from the NC4 wild-type strain.
Parasexual genetic analyses: Established parasexual genetic techniques were used for diploid construction, mitotic recombination, segregation of haploids and determination of genotypes (NEW- ELL 1982; WELKER and WILLIAMS 1982a). In this work either benlate (20-35 pglml) or thiaben- dazole (2pg/mI) was used to haploidize diploids (WILLIAMS and BARRAND 1978; WELKER and WILLIAMS 1980a). Some diploids were selected using dominant cobalt resistance (cob-354) associated with the endogenous Ddpl plasmid (WILLIAMS 1978; METZ et al. 1983). For these crosses the well-marked tester strain HU1628 was used as a haploid parent and the diploids were selected using growth in association with a cobalt-resistant Bacillus subtilis strain, BSU2 (isolated by S. KAYMAN in this laboratory), at 21 & 1" on SM agar plates containing 250 pg/ml of cobaltous chloride. The markers used in this work have been described previously except agg-361 and tsgV1826; their established linkage relationships and phenotypes are summarized in Table 2. The tsgV1826 and agg-361 mutations were isolated in strains H u l l 1 6 and HU1098, respectively, after mutagenesis of strain AX3 with N-methyl-N'-nitro-N-nitrosoguanidine (MNNG) to 30% survival (B. A. METZ and K. L. WILLIAMS, unpublished results). Nomenclature for translocations follows that proposed by WELKER, METZ and WILLIAMS (1982) and takes the form Tn(x,y), where n is a number designation unique to each translocation and x and y denote the linkage groups involved.
Chromosome staining: Amoebas were removed from colonies on growth plates, dispersed in 1 ml of salt solution and added to 4 ml of pregrown E. coli B/r (-109/ml) in phosphate-buffered saline (DEERING et al. 1970) in a 50-ml Erlenmeyer flask. Thiabendazole (final concentration 10 rg/ml) was added and the flask shaken at 180 rpm on an orbital shaker at 21 f 1". After 2.5 hr the amoebas were fixed and the chromosomes stained as described previously (WELKER and WILLIAMS 1980a).
RESULTS
Origins of translocations in D. discoideum: In D. discoideum as in other eu- karyotes there is an association between the occurrence of translocations and prior treatment with mutagenic agents (KAFER 1977). In particular, many strains in which translocations have been found were previously mutagenized with MNNG. Six of the 15 translocations listed in Table 3 [T35Z(III,VI), T352(I,III), T353(III,VI), T359(I,IV), T36O(II,VZI) and T367(I,W)] were found in strains that were isolated after MNNG treatment; in addition, two putative nonreciprocal translocations that were recovered after MNNG mutagenesis are

TRANSLOCATIONS IN D. DISCOZDECJM 343
being characterized (D. L. WELKER, unpublished results). These results strongly indicate that these translocations were induced by MNNG and are consistent with results obtained with MNNG-treated mammalian cells (DIPAOLO, AMS- BAUGH and POPESCU 1980; KINSELLA and RADMAN 1980; DIPAOLO and Po-
Several translocations have been discovered in strains with defective DNA repair. Three translocations [T351(ZZZ,VZ), T354(ZZ,VZZ), and T359(Z,ZV)] and also the D35O(ZZZ,ZZZ) tandem duplication (WELKER, METZ and WILLIAMS 1982) were found in a radBl3 genetic background, whereas two translocations [T362(ZZZ,VZZ) and T366(ZV,VZ)] were discovered in a radG5 genetic background. Note that the T351(ZZZ,VZ) and T359(Z,ZV) translocations may have been MNNG induced as they have been traced to the original radBl3 strain, yS13, which was isolated after MNNG mutagenesis.
However, five of the translocations that we have studied cannot be attributed to any known mutagenic treatment or genetic background. Two of these [ T357(W,VZZ) and T365(ZZZ,VZZ)] clearly arose in haploid stocks being main- tained by clonal passaging. Three others, first discovered in diploids [T361(Z,VZ), T363(ZZZ,ZV)] or in a segregant haploid [X23, T364(Z,ZZZ,VZ)], may also have arisen in haploid stocks maintained by clonal passaging.
The origins of the translocations that we have studied will be given. All were discovered by us except the T353(ZZZ,VZ) translocation (NEWELL and Ross 1982; Ross and NEWELL 1979) and are of certain independent origin except the T354(ZZ,VZZ) and T355(ZZ,VZZ) translocations which are probably multiple isolates of a single event.
1. The T351(ZZZ,VZ) translocation was traced to the MNNG-induced, radBl3 DNA repair mutant, $313; it is not present in other repair mutants isolated at the same time from the same parent (NC4). This was established by crossing $31 3, $35 and several derivatives of other repair mutants with well-marked untranslocated tester strains such as HU413 or XM1. Only diploids derived from $313 cosegregated linkage groups I11 and VI (DU1244, Table 4a).
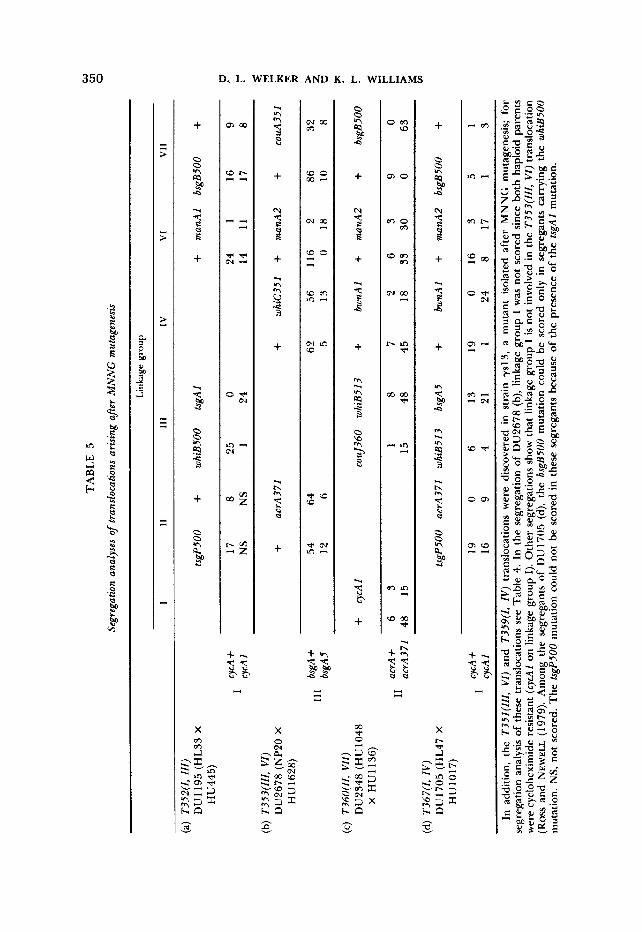
2. The T352(Z,ZZZ) translocation was discovered in strain HL33, a white- spored (whiB5OO) mutant isolated by MORRISSEY, WHEELER and LOOMIS (1980) after MNNG mutagenesis; it is not present in other white mutants isolated from the same parent (HL21) such as HL47 (but note that HL47 carries a different translocation, Table 5d). A typical segregation pattern for the T352(Z,ZZZ) translocation is presented in Table 5a.
3. The T353(ZZZ,VZ) translocation, which is of certain independent origin to the T351(ZZZ,VZ) translocation, has now been traced by us as far back as strain NP20, a cycloheximide-resistant mutant derived by MNNG mutagenesis (P. C. NEWELL, personal communication). This was established by crossing NP20 with strain HU 1628, a well-marked, nontranslocated, tester strain carrying the dom- inant cobalt resistance selector cob-354 (Table 5b). This translocation is present in many strains, including NP187, XP55, XP222 and their derivatives which all show similar segregation patterns (NEWELL and ROSS 1982; ROSS and NEW- ELL 1979; WALLACE and NEWELL 1982a; WELKER and WILLIAMS 1983).
4. The T354(ZZ,VZZ) and T355(ZZ,VZI) fusion chromosomes were previously
PESCU 1977).
344 D. L. WELKER AND K. L. WILLIAMS
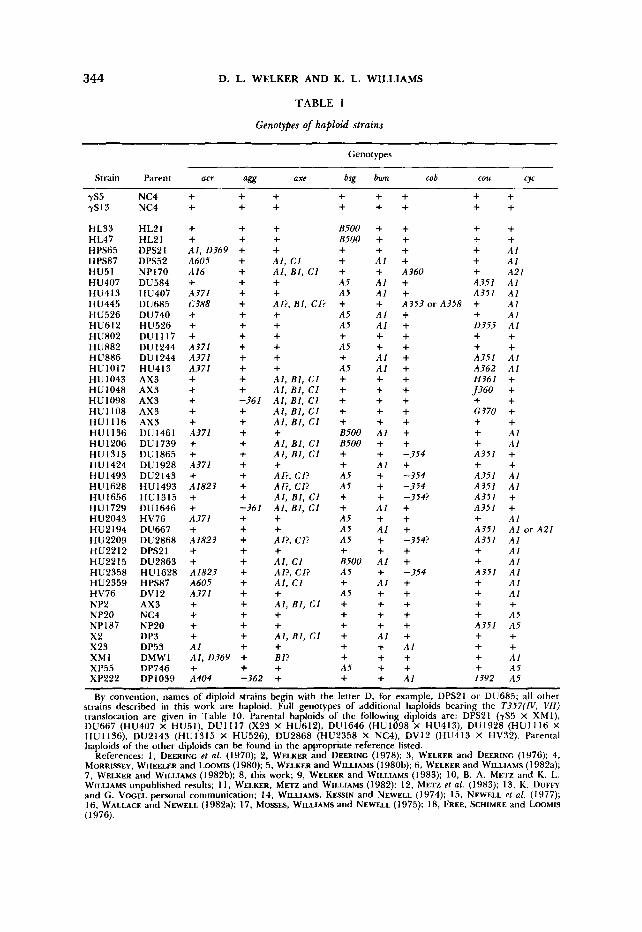
TABLE 1
Genotypes of haploid strains
Genotypes
Strain Parent acr agg axe bsg bwn cob cou C Y
YS5 7.513
HL33 HL47 HPS65 HPS87 HU51 HU407 HU413 HU445 HU526 HU612 HU802 HU882 HU886 HU1017 HU1043 HU1048 HU1098 HUI 108 HU1116 HU1136 HU1206 HU1315 HU 1424 HU1493 HU1628 HU1656 HU1729 HU2043 HU2194 HU2209 HU2212 HU2215 HU2358 HU2359 HV76 N P2 NP20 NP187 x 2 X23
NC4 NC4
HL21 HL21 DPS2 1 DPS52 NP170 DU584 HU407 DU685 DU740 HU526 DUI 117 DU 1244 DU 1244 HU413 AX3 AX3 AX3 AX3 AX3 DUI461 DUI739 DU 1865 DU 1928 DU2143 HU1493 HU1315 DU 1646 HV76 DU667 DU2868 DPS2 1 DU2863 HU1628 HPS87 DV12 AX3 NC4 NP20 DP3 DP53
+ + + + A I , 0 3 6 9 A605 A16 + A371 C388 + + + A371 A371 A371 + + + + + A371 + + A371 + A1823 + + A371 + A I 8 2 3 + + A1823 A605 A371 + + + + AI
+ + + + + + + + + + + A I , CI + A l . BI , C1 + + + + + AI?, B l , C l ? + + + + + + + + + + + + + A I , B I , C l + A I , BI , C l -361 A I , B l , C l + A I , B I , CI + A I , 8 1 , CI + + + A l . BI. C l + A I , 8 1 , CI + + + AI?, CI? + AI?, Cl? + A l , B I , CI -361 A I , B I , CI + + + + + AI?, C l ? + + + A I , CI + AI?, Cl? + A I , C1 + + + A I , B l , C l + + + + + A I , B I , CI + +
XMI DMWl A I , 0 3 6 9 + BI? XP55 DP746 + + + XP222 DP1039 A404 -362 +
+ + 8500 8500 + + + A5 A5 + A5 A5 + A5 + A5 + + + + + 8500 8500 + + A5 A5 + + A5 A5 A5 + 8500 A5 + A5 + + + + + + A5 +
+ + + + + AI + A1 AI + A 1 A I + + AI A1 + + + + + AI + + AI + + + AI + AI + + AI + A1 + + + + A1 + + + +
+ + + + + + + + + + + + A360 + + A351 + A351 A353 or A358 + + + + 0 3 5 5 + + + + + A35 I + A362 + H361 + J360 + + + G370 + + + + + + -354 A351 + + -354 A35 I -354 A351 -354? A351 + A351 + + + A351 -354? A35 I + + + + -354 A35 1 + + + + + + + + + A351 + + A1 + + + + + A1 1392
+ + + + A1 A1 A21 A1 A1 A1 A1 A1 + + A1 AI + + + + + AI A1 + + AI A1 + + AI A1 or A21 A1 A 1 A I A1 A1 AI + A5 A5 + + A1 A5 A5
By convention, names of diploid strains begin with the letter D, for example, DPS21 o r DU685; all other strains described in this work are haploid. Full genotypes of additional haploids bearing the T357(IV, V l l ) translocation are given in Table 10. Parental haploids of the following diploids are: DPS2l (7S5 x XMI) , DU667 (HU407 X HU51), DUI117 (X23 X HU612). DUI646 (HU1098 X HU413), DUI928 (HU1116 X HU1136), DU2143 (HU1315 X HU526). DU2868 (HU2358 X NC4), DV12 (HU413 X HV32). Parental haploids of the other diploids can be found in the appropriate reference listed.
References: 1, DEERINC et al. (1970); 2, WELKER and DEERINC (1978); 3, WELKER and DEERINC (1976); 4, MORRISSEY, WHEELER and LOOMIS (1980); 5. WELKER and WILLIAMS (1980b); 6, WELKER and WILLIAMS (1982a); 7, WELKER and WILLIAMS (1982b); 8, this work; 9, WELKER and WILLIAMS (1983); 10, B. A. METZ and K. L. WILLIAMS unpublished results; 11, WELKER, M m and WILLIAMS (1982); 12, METZ et al. (1983); 13, K. DUFFY and G. VWEL personal communication; 14, WILLIAMS, KESSIN and NEWELL (1974); 15, NEWELL et al. (1977); 16, WALLACE and NEWELL (1982a); 17, MOSSES, WILLIAMS and NEWELL (1 975); 18, FREE, SCHIMKE and LOOMIS (1976).
TRANSLOCATIONS IN D. DISCOIDEUM 345
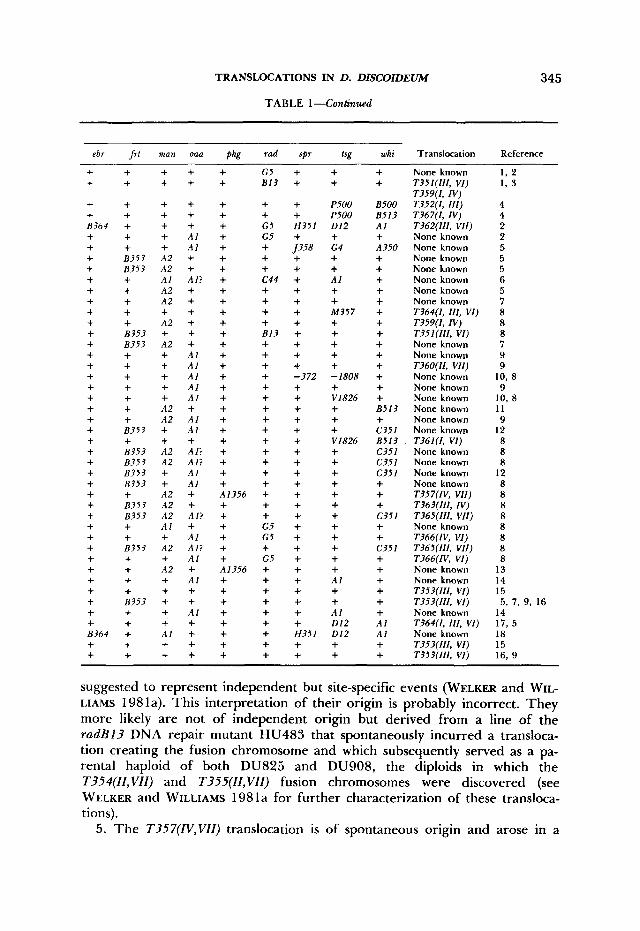
TABLE 1-Continued
ebr frf man oaa phg rad spr tsg whi Translocation Reference
i i i-i i i i i i i
i i i f i i i i + i 8364 i i i i i i i A1 i i i i A1 i i 8353 A2 i i i 8353 A.? i i i i AI Al? i i i A 2 i i i i A . ? + i i i i f i i i A 2 i i i 8353 i i i i 8353 A2 i i i i i A1 i i i i A i i i i i A i i i i i A1 i i i i A1 i i i A 2 i i i i A2 A1 i i E353 i A1 i i + + i i i 8353 A2 Al? i i 8353 A2 A / ? + i 8353 i A1 i i 8353 i AI i i i A2 i A1356 i 8353 A2 i i i 8353 A2 AI? i i i A I + i i i + A1 i i 8353 A2 A17 i i i i A1 i i i A2 i A1356 i i i A1 i i + i i i i 8353 i i i i i i A1 i i i i i i 8364 i A1 i i i i i f i i i i i i
G5 813
i i G5 G5 i i i c44 i i i i E13 i i i + i i i i i i + + + i + i i G5 G5 i G5 + + i i i i + i i
i i
i i H35 1 i J358 i i i i i i + + i i i -372 + i i i + i i i i i i i i i i i i i i i i i i H351 i i
i + P500 P500 Dl2 + G4 i + A1 + i M357 i i i + i -1808 i VI826 i + i V1826 i i i + i i i i i i i i A1 i i A1 0 1 2 0 1 2 i i
i i
E500 E513 A1 i A350 + i i + + i i i i i i i i i E513 + C35l E513 C351 C351 C351 i i i C351 i i C35 1 i i i i i i A1 AI i i
None known T351(III, VI) T359(1, IV) T352(I, 111) T367(1, IV) T362(111, VII) None known None known None known None known None known None known None known T364(1, 111, VI) T359(1, IV) T35 l(II1, VI) None known None known T360(II, VII) None known None known None known None known None known None known T36 1 (I , VI) None known None known None known None known T357(IV, VII) T363(111, N) T365(111, VII) None known T366(IV, VI) T365(111, VII) T366(IV, VI) None known None known T353(111, VI) T353(111, VI) None known T364(1, I l l , VI) None known T353(III, VI) T353(III, VI)
1, 2 1, 3
4 4 2 2 5 5 5 6 5 7 8 8 8 7 9 9
10, 8 9
10, 8 11 9
12 8 8 8
12 8 8 8 8 8 8 8 8
13 14 15 5, 7 , 9, 16
14 17, 5 18 15 16, 9
suggested to represent independent but site-specific events (WELKER and WIL- LIAMS 1981a). This interpretation of their origin is probably incorrect. They more likely are not of independent origin but derived from a line of the radBI3 DNA repair mutant HU483 that spontaneously incurred a transloca- tion creating the fusion chromosome and which subsequently served as a pa- rental haploid of both DU825 and DU908, the diploids in which the T354(II, VIZ) and T355(II, VIZ) fusion chromosomes were discovered (see WELKER and WILLIAMS 198 l a for further characterization of these transloca- tions).
5. The T357(IV,VII) translocation is of spontaneous origin and arose in a

346 D. L. WELKER AND K. L. WILLIAMS
TABLE 2
Linkage relationships and phenotypes of the marker loci
Linkage ! P u p Marker loci in strains used
Total loci
I coul cycA 15 I 1 acrA axeA axeC oaaA tsgD tsgP whiA 26
I11 acrC axeB bsgA COUD COUG COUJ radB radC 20
1V acrD bwnA couH ebrB fr tA sprH sprJ whiC 20 radC tsgA whiB
VI manA tsgV 3 VI1 bsgB cobA couA frtB phgA tsgG tsgM 9
Phenotypes of niutations at these loci are: acrA, altered resist- ance to acriflavin (100 rg/nil), methanol (2-3%) and/or benzim- idazole derivatives; acrC and acrD, altered resistance to acriflavin (100 pg/ml) and/or benzimidazole derivatives; agg, aggregation defective; axe, ability to grow in axenic media in the absence of bacteria; bsg, inability to use B. subfilis as a food source; bwnA, production of brown pigment during development; cob, resistance to cobaltous chloride (300 pglnil); COUA, COUG, couH and COUJ, sensitivity to coumarin (1.3 1 1 1 ~ ) with pleiotropic temperature sen- sitivity; couD and c o d , sensitivity to coumarin (1.3 mM); cycA, resistance to cycloheximide (500 pglml); ebrB, resistance to ethid- ium bromide (25 pg/nil); frtB, distribution of fruiting bodies in concentric rings; manA, a-mannosjdase-1 deficient; oaaA, devel- opment in the presence of w-aminocarboxylic acids; phgA, defec- tive phagocytosis (K. DUFFY and G. VOGEL, personal communi- cation; V ~ C E L et al. (1980); rad, sensitivity to irradiation with UV light or 7-rays; sprH, round spore shape; sprJ, failure of spores to mature; tsg, temperature sensitive for growth; whi, absence of the normal yellow spore pigment.
The number of loci per linkage group is from NEWELL (1982). The assignment of the phgA locus to linkage group VI1 will be described elsewhere (K. DUFFY and G. VOGEL, unpublished re- sults). The agg-361 mutation maps to linkage group I and the agg- 3 6 2 mutation maps to either linkage group 111 or VI (WELKER and WILLIAMS 1983), but their relationships to previously mapped agg loci were not established. In the interests of simplicity, we have used couA?51 to designate the coumarin sensitivity (couA35I) and temperature sensitivity (previously tsgK2I) which we proposed to be pleiotropic manifestations of a single mutation (WELKER and WILLIAMS 1982b; WELKER and WILLIAMS 1983). In the data pre- sented here, the temperature sensivity was always scored and in most cases the coumarin sensitivity as well. We have never ob- served a coumarin-sensitive (couA351) strain that was not also temperature sensitive; although WALLACE and NEWELL (1 982a) have claimed separation of the two phenotypes by mitotic recom- bination (in 10% of recombination events proximal to the cobA cobalt resistance locus of linkage group VII). We have not been able to duplicate their work; in more than 100 independent co- balt-resistant, mitotic recombinants of linkage group VII, no s e p aration was observed (D. L. WELKER unpublished results).
line of strain HV76 that was clonally passaged on SM plates once per week. This stock has been named HU2043. A typical segregation pattern for a dip- loid heterozygous for this translocation is presented in Table 6a (DU258 1). Strain HV76 (bwnA+, phgA1356) clearly does not carry this translocation, and diploids constructed with HV76 segregate linkage groups IV (bwnA+) and VI1

TRANSLOCATIONS IN D. DISCOIDEUM 347
TABLE 3
Translocations in D. discoideum
Frequency of re- Strain of combinant segre- Markers scored in
Translocation origin gants recombinants Probable origin Reference
T35 I ( I I I , VI ) yS13 1% (5/823) radB, manA
T352(1, I I I ) T353(1II, V I ) T354(II, VU)
T357(IV, VU) T359(1, IV)
T360(II, V U ) T361(1, VI ) T362(III, VII)
T363(III, IV)
HL33 4% (18/406) NP2O 2% (2/136) DU825 5% (5/98)
DU908 2% (1/39)
HU2043 4% (6/148) $313 3% (3/95)
HU1048 Cl%? (0/72) DU1928 5% (6/121) HPS65 <1%? (0/51)
DU667 2% (2/99) T364(1, III , V I ) X23 3% (3/95) T365(III, VII) HU2358 1% (1/111) T366(IV, V I ) HU2359 1% (1/105)
T367(1, IV) HL47 3% (3/123)
cycA, whiB bsgA, manA whiA, cobA
whiA, cobA
bwnA, phgA cycA, bwnA
acrA, bsgB cycA, manA radG, bsgB
bsgA, bwnA cycA, bsgA, manA bsgA, couA bwnA, manA
cycA, bwnA
MNNG-induced or defective DNA re- pair
MNNG-induced M N NG-induced Spontaneous or de-
fective DNA re- pair
Spontaneous or de- fective DNA re- pair
Spontaneous MNNG-induced or
defective DNA re- pair
MNNG-induced Spontaneous Spontaneous or de-
fective DNA re- pair
Spontaneous Spontaneous Spontaneous Spontaneous or de-
fective DNA re- pair
MNNG-induced
1, 2, 3, 4
3, 4 3, 4, 5 3, 6
3, 6
4 3, 4
7 4, 8 4
3, 4, 9 4, 10 4 4
4
These translocations lead to the apparent cosegregation of markers known to map on different linkage groups, which are indicated in the name of the translocation. Diploids heterozygous for - - these translocations produce only two major classes of segregants, having parental combinations of markers on the affected linkage groups, rather than the expected four classes of segregants. The missing nonparental segregants would normally account for 50% of total segregants. Hence, these translocations are most likely reciprocal rather than nonreciprocal or insertional, except the T354(II, VU) and T355(II, VU) translocations, which were shown to be fusion chromosomes (WELKER and WILLIAMS 1981a) and the T357(IV, VU) translocation which may also be a fusion chromosome. Recombinant segregants with nonparental combinations of markers on the affected linkage groups are produced at up to 5% of total segregants. Such recombinants arise by mitotic recombination and include both translocated and nontranslocated strains.
All strains listed in this table are haploid except DU825, DU908, DU1928 and DU667, which are diploids. Translocation-bearing haploid segregants derived from these diploids are, respectively: HU538 (WELKER and WILLIAMS 1981a), HU628 (WELKER and WILLIAMS 1981a), HU1424 and HU2194. Strain yS13 carries two translocations which have been separated in segregants of DU1244 to produce HU886 and HU882 which carry the T351(III, VI) and T359(1, IV) translo- cations, respectively.
References: 1, WELKER and DEERINC (1976); 2, D. L. WELKER, unpublished data; 3, WELKER, METZ and WILLIAMS (1982); 4, this work; 5 , NEWELL and Ross (1982); 6, WELKER and WILLIAMS (1981a); 7, WELKER and WILLIAMS (1983); 8, S. KAYMAN, personal communication; 9, WELKER and WILLIAMS (1980b); 10, WELKER and WILLIAMS (198213).

348 D. L. WELKER AND K. L. WILLIAMS
h e
c 2 - 0
C U
c U

TRANSLOCATIONS IN D. DISCOIDEUM 349
8 v, cq - $ 9 9
+ * z cv
O n 3 In E
+ r o o
0 w + v
Q Z 2 -U 2
+ --$
v, 0 \o
T m m
2
- + 2 2 22 5
0 -
g z
3 z g z a , E = ' a Z . . Z n c
g s 2 z s
X 3 Z s n 3 - 3 2 p m X
cc > E
h n - v
W C L C "e,
s g . 2 z 5 p 9 g q 5 3 4 . g g $ ? 2 4 4 - arq r e i
" % g y : ! r u c 8,; a.2 b m ' 3 e, gy
'Z 3 g 2;:o U
8 - 0 u g D z . c 4 p.3: m u p @ % z 3 4 3 U & , x L E 2 g % L E -%am 23 wy &93 & * 0.5 2 e, e,439 E 5 g ESS 0.c & c e,*&- U .u;S8 .- c c- e, 0 o s ,=: 8 e , ' , ? 4 g 2 e ..S c r u . ' l m % E ? M S " Q C : 5 2 2 $ U c k g U Q P C .B.s SD&C E 8. f 52 5.22 $ 2 E g g . j 9 s s a 0
g2 ?j & a 2 2 2 U E S y z m o 3 B E . 5 +' $ E 82 g T 5 . c g: 8 ;&-".8-1
CI-., .i 0 4 gg p m s b u T 2 c o
g ? g scg $ 4
z??sfm;z8 8 a23 ,9 8 ; g --xu 82 2 g > 0 g % L 3 . 2 c s = - = e s 2 g q p&&& U z-- U " ? $4 y ~ . c % 0 1 0 ~ c e,, E & L L &.a E * * % a G & e , a% 72 g2 g 5 g-&:b e, := 2 g 3 ;"e;s;a,+ L 42 .2 e ; i t; E - m s z ; = c * 0
' Z 2 &as,- e, 5 0 a' UT. c
z $ . e , L $ L 2 :& ESeTG 3 3 & 2 5,;: m - c ? 2 02* - P - m 2 5 e,$:
L L @-cJ-UC_cI
c 2 3 E &.?3 4
a as .Y s c Ea y) 3 3 ' 2 $ 2
P r m o ,
0 aa, q1;;
k.2 ES 2 M 8 5
&,$% $ & E g
$ u u E ' E 8."
% . c - M X U U
.- Ea 3 g E Z % %
- C T S 2 % b
350 D. L. WELKER AND K. L. WILLIAMS
In w el m 2
+ v \ T T 9 9 9 9
I - I
e
0 O 2 9 9
+
ry
-2 9 + 1
-2 3 4
+
2 U\ 2
s 3
0 \o
s
1
'c, 2.
+
h
v
X P-
h
9

TRANSLOCATIONS IN D. DZSCOZDEUM 351
1
2 s 8
v, v\
2 2 + ry
T E
+ v\ 7 9 4
+ 1 t\ T 7 $ 1
% i? -. s 6
h m m r.
3 z X
h
v m h e

352 D. L. WELKER AND K. L. WILLIAMS
(phgA1356) normally (K. DUFFY and G. VOGEL, personal communication; D. L. WELKER, unpublished results). For example, with diploid DV43 (HV76 X HUll36) of 34 segregants: four were bwnA+, phgA1356; six were bwnA+, phgA+; 11 were bwnA1, phgA1356 and 13 were bwnA1, phgA+.
6. The T359(Z,ZV) translocation has been traced to the MNNG-induced, radBl3 DNA repair mutant $31 3 which also carries the T351(ZZZ,VZ) translo- cation (Table 4a and b). The T359(Z,ZV) translocation is not present in other DNA repair mutants isolated from the same parent at the same time. This was demonstrated in the same experiments used to establish the origin of the T351 (ZZZ, VZ) translocation.
7. The T36O(ZZ,VZZ) translocation was discovered in a MNNG-induced cou- marin-sensitive mutant of AX3, HU1048 (DU2348, Table 5c; see also WELKER and WILLIAMS 1983). It is not present in AX3 or in other coumarin-sensitive or temperature-sensitive mutants (HU 1043, HU 1 108 and HU 1 1 16) derived from AX3 at the same time as HU1048 (WELKER and WILLIAMS 1983; Table 7b).
8. The T361(Z,VZ) translocation was found in DUI928 (Table 7a) and is due to rearrangement of linkage groups I and VI derived from HUl116, a parental haploid of DU1928. This was confirmed by crossing a haploid segregant of DU 1928, HU 1424, which carries the T361(Z,VZ) translocation to the non- translocated tester strain HU526 (Table 7a). However, HU 1 1 16 (Table 7b), its parent AX3 and other mutants derived from AX3 at the same time as H u l l 16 do not contain this translocation (WELKER and WILLIAMS 1983) so it must be of spontaneous origin either in DU1928 or a line of H u l l 1 6 that was maintained by clonal passaging on SM plates and from which DU1928 was derived.
9. The T362(ZZZ,VZZ) translocation was discovered in the radG5 DNA repair mutant HPS65 (Table 4c). It is not present in yS5, the original radG5 mutant or in DPS21, the diploid from which HPS65 was derived, since a diploid constructed with another segregant of DPS2l (HU2212) did not cosegregate linkage groups I11 and VI1 (DU2935, Table 4d).
10. The T363(ZZZ,ZV) translocation was discovered in diploid DU667 (Table 7c) and shown to affect linkage groups 111 and IV derived from strain HU407. This translocation is of spontaneous origin and may have arisen in DU667 itself or in the line of HU407 used to construct DU667. It is not present in strain HU407 (see segregation of DU788 (HU407 X HU510), WELKER and WILLIAMS 1980b).
11. The T364(Z,ZZZ,VZ) translocation was discovered in strain X23 and its derivatives, for example HU802 (Table 7d). This translocation is of sponta- neous origin since a diploid constructed with another segregant derived from the same diploid as X23 and which carried the same linkage groups I, 111 and possibly VI, as X23 did not cosegregate linkage groups I, I11 and VI when crossed to an appropriate nontranslocated tester strain (D. L. WELKER, unpub- lished results). This complex translocation appears to break frequently, and some diploids, such as DU1088, which should be heterozygous for this rear- rangement do not cosegregate all three linkage groups. In particular with

TRANSLOCATIONS IN D. DISCOZDEUM
B e M
0 Y
3
0 % rq c .a
+
cy T I
+
I
I +
2 9 v\
-c 3
+
I h n\ f t
+
* 7 B +
X
2 -
OIn m
o w
5 - >
0 s c .a
+
cy 3 E
+
I
3 +
2 "\ 2 3
+
-.I h n\ f
5
+
I
f B +
e.
+I f f P P
+
w
2 B
cy T E
+
L-.
3 +
U\ f 1
+
+
s s -c 3
z X m m x 0 a
or) m
a

354 D. L. WELKER AND K. L. WILLIAMS
DU1088 only linkage groups I and VI cosegregated (Table 7d; see also WELKER and WILLIAMS 1982b).
12. The T365(ZZZ,VZZ) translocation is of spontaneous origin and was discov- ered in a line of HU1628 that was passaged weekly on SM plates containing 300 pg/ml of cobaltous chloride. The translocated line of HU1628 has been named HU2358 (Table Sa). This translocation is clearly not carried by HU1628 itself (Table 8b).
13. The T366(ZV,VZ) translocation arose in a line of the radG5 DNA repair mutant HPS87. The translocated version of HPS87 has been named HU2359. In the present work, diploids heterozygous for this translocation had very skewed segregation patterns with only a few translocation-bearing segregants being produced (Table 4e); the cause of this phenomenon is not clear since the translocation-bearing segregants that were obtained did not have abnor- mally slow growth rates. This translocation was not present in other radG5 mutants such as HPS65 (DU2861, Table 4c) or in other strains derived from HPS87 (WELKER and WILLIAMS 1980~).
14. The T367(Z,ZV) translocation was discovered in strain HL47 (Table 5d), a whiB513 mutant derived from HL21 by MNNG mutagenesis (MORRISSEY, WHEELER and LOOMIS 1980). This translocation is not in other white mutants derived from HL21 such as HL33 [see segregation data presented by MOR- RISSEY, WHEELER and LOOMIS (1980) and by WELKER and WILLIAMS (1980c)l.
Characterization of the T357(IV,VII) translocation: To substantiate that spon- taneously arising cosegregation patterns involve chromosome rearrangements, we characterized in detail the T357(ZV,VZZ) translocation which involves two well- marked chromosomes. Diploids heterozygous for this translocation coseg- regated markers on linkage groups IV and VI1 and, like diploids heterozygous for other translocations (Table 3), in turn produced segregant haploid mitotic recombinants at higher than normal frequency (-3%). Among 147 independ- ently derived, thiabendazole-induced segregants of two diploids heterozygous for the T357(ZV,VZZ) translocation (DU2499 and DU258 1), four haploid mitotic recombinants were found (Table 6a). The frequency at which mitotic recombi- nants were obtained was increased about two-fold by irradiation with ultraviolet light (UV) (1 30 J/m2, 254 nm). Of 197 thiabendazole-induced haploid segregants obtained from irradiated DU2581 populations, 13 were recombinant (Table 6b). This increase was consistent with earlier reports of UV-induced mitotic recom- bination in D. discoideum (WALLACE and NEWELL 1982b; WELKER, METZ and WILLIAMS 1982).
The origin and expected phenotypes for haploid mitotic recombinants derived from DU2581 are illustrated in Table 9. We recovered four of the six possible classes. These four arise from crossovers between the translocation breakpoint and the genetic markers on linkage groups IV and VII. The two classes of recombinants that were not recovered involve crossovers between the COUA and phgA loci on linkage group VII. From backcrosses of eight recombinants to nontranslocated tester strains (NP2, X2, HU526, HU 1656) we found, on the basis of cosegregation of linkage groups IV and VII, that two were translocated and six carried a normal pair of linkage groups IV and VI1 (Table 9). Of four

TRANSLOCATIONS IN D. DISCOZDEUM
+ 1
2 s s
+ cy
T E
+ 1
2 2 a
+ .-r h n, 1 4 + 1
s 2.
h
v
v \ + 11 22
I H I
+ 1 v\ n, s s
+ cy
-2 2
+ 1
2 r; -e a
+ 1 p\ n, 3 s
+ -4
5 P
355

356 D. L. WELKER AND K. L. WILLIAMS
TABLE 9
Recombination in a diploid, DU2581, heterozygous for the T357(IV, VII) translocation
Normal linkage Group IV
Rearranged linkage Group IV-VI1
Normal linkage Group VI1
Rearranged linkage Group VII-IV
Recombi. nation interval
Phenotypes of haploid mitotic recombinants
Centromeres No. Genotype in segregants Translocated analysed
1 bwnA+ phgA+ couA35l B, C Nontranslocated 2 1 bwnAl phgA1356 couA+ A, D Translocated 1 2 bwnA+ phgA+ couA351 B, D Translocated 1 2 bwnAl phgAl356 couA+ C, A Nontranslocated 4 3 bwnA+ phgA1356 couA351 B, D Translocated 0 3 bwnAl phgA+ couA+ C, A Nontranslocated 0
The translocation is shown here as reciprocal, although no known markers have been assigned to a rearranged linkage group VII-IV; preliminary cytological data indicate that this translocation may in fact be a fusion chromosome, possibly even a Robertsonian translocation. The order of the phgA and couA loci on linkage group VI1 are hypothetical and for illustrative purposes only; however, if the translocation is reciprocal, then both loci are distal to the bwnA locus on the rearranged linkage group IV-VI1 (Table 11). The translocation breakpoints are indicated by the * symbols. T o clarify the diagram, linkage group IV is represented by dashed lines and linkage group VI1 by solid lines. The origin of the centromeres in the haploid mitotic recombinants is indicated by the capital letters. The chromosome whose centromere is listed on the left has undergone a mitotic exchange, and the centromere of the unexchanged chromosome is shown on the right.
haploid mitotic recombinants obtained from other diploids heterozygous for the 7'357(ZV, VU) translocation, two were normal and two were translocated.
Determination of the gene order on the rearranged T357(IV,VII) linkage groups: T o determine the position of the translocation breakpoint(s), we first constructed a series of translocation-bearing haploids that contained appropriate genetic markers for linkage groups IV and VII. These, which were obtained as haploid mitotic recombinants in the manner described before, carry either the couA35I or bsgB5OO marker for linkage group VI1 in place of the phgAZ356 mutation found on the original translocated chromosome, whereas linkage group IV was

TRANSLOCATIONS IN D. DZSCOIDEUM 357
marked by insertion of either the bwnA2 or whiC352 mutations (Table 10). In addition, spontaneous cobalt resistance mutations at the cobA locus of linkage group VI1 were selected in some well-marked strains. Crosses between these haploids and nontranslocated tester strains established that they retained the T357(ZV,VZZ) translocation on the basis of cosegregation of linkage groups IV and VII. When diploids were constructed between pairs of the translocation- bearing haploids, the linkage group IV markers (6wnAI and whiC35Z) and the linkage group VI1 markers (phgA2356, couA352 and cobA39I) again segregated as a unit. This is the expected result for diploids homozygous for the T357(ZV,VZZ) translocation if the bwnAl and whiC352 markers are physically linked to the linkage group VI1 markers. Thus, the bwnA, whiC, phgA, couA, 6sgB and c06A loci must be on the same rearranged chromosome. Of 64 haploid segregants of these diploids, no haploid mitotic recombinants were obtained. If the transloca- tion is reciprocal, then, since D. discoideum chromosomes are acrocentric, either the bwnA and whiC loci are proximal to the linkage group VI1 markers or the linkage group VI1 markers are proximal to bwnA and whiC.
To determine which of these two gene orders was correct, diploid mitotic recombinants were selected from DU2498 using the recessive cobA392 cobalt resistance mutation. Diploid DU2498, which was homozygous for the T357(ZV,VZZ) translocation, was heterozygous at the bwnA, phgA, couA and cobA loci on the rearranged chromosome. If these markers are linked to the linkage group VI1 centromere with 6wnA distal to the linkage group VI1 markers, then diploid recombinants of DU2498 should be homozygous for the bwnA2 mutation (as in the absence of multiple recombination events all markers distal to the selected crossover become homozygous). If they are linked to the linkage group IV centromere with the linkage group VI1 markers distal to bwnA, then recom- binant diploids which remain heterozygous at bwnA are expected, as in this configuration 6wnA is proximal to the selector c06A. The second possibility was observed. Of 1 1 independent, cobalt-resistant, diploid mitotic recombinants, selected on the basis of homozygosity for the co6A392 cobalt resistance mutation (Table 1 I), one was homozygous at both the 6wnA and phgA loci (and presumed homozygous for the wild-type allele at the couA locus), eight remained hetero- zygous at the 6wnA locus but were homozygous at the couA and phgA loci and two remained heterozygous at the bwnA, COUA and PhgA loci. The first diploid could have arisen by nondisjunction, whereas the last two could have arisen without mitotic recombination by new mutation at the c06A locus. However, the remaining eight diploids, together with known gene orders on linkage groups IV and VI1 (WELKER and WILLIAMS 1982a; WALLACE and NEWELL 1982a), prove that, if the translocation is reciprocal, then the gene order on the rearranged chromosome must be: centromere IV, bwnA, whiC, breakpoint IV/VII, couA, 6sgB cobA, telomere VI1 with the phgA locus of linkage group VI1 proximal to cobA but not ordered. The second rearranged chromosome produced by a reciprocal translocation would thus consist of centromere VII, breakpoint VII/ IV, telomere IV. However, we have not established this or that any known locus was transferred from linkage group IV to the linkage group VI1 centromere. In fact preliminary cytological evidence suggests that the T357(ZV,VZZ) translocation

358 D. L. WELKER AND K. L. WILLIAMS
1 v\ 9 U
+ + + $
+ + +
1
4 + + 2
d 9 i 9-c 9 9 3 s

TRANSLOCATIONS IN D. DISCOIDEUM
TABLE 11
Gene orders on the T357(IV, VII) rearranged linkage group IV-VII determined from diploid mitotic recombinants selected from DU2498, a diploid homozygous for the
translocation
359
~~
DU2498 (HU2175 X HU2129) parental genotype on the rearranged chromosome:
bwnAl phgAl356 couA+ cobA391
-o-----+------ * I I I I I
(+l+) (-2 -)(- 3 -)(- 4 -) bwnA+ P W + cow4351 cobA +
-o------+------ * I I I I I I
Diploid mitotic recombinants obtained by selection for cobalt resistance
Interval 1 (or nondisjunction):
bwnA 1 phgA1356 couA+ cobA391 No. observed
-o------+------- * 1 I I I I
-o----+------- * I I I I bwnAl PhgA 1356 couA + cobA391 1
Interval 2:
bwnAl phgA1356 couA+ cobA391
bwnA+ phgA 1356 couA+ cobA39l
-o------+------- * I I I I I
Interval 3:
bwnA 1 phgA1356 couA + cobA391
a
bwnA+ &A+ couA + cobA39 1 0 -o------+------- * I I I
I
Interval 4 (or new mutation at the cobA locus):
bwnA 1 phgA1356 cowl+ cobA391
-o------+------- * I I I I I I
bwnA + P k A + couA351 cobA391 2
Total 11
For this diagram the T357(IV, VII) translocation is assumed to be reciprocal; however, prelim- inary cytological evidence is also consistent with this rearrangement being a fusion chromosome, possibly even a Robertsonian translocation. The available recombination data are not inconsistent with such an event. The genotypes of the diploid recombinants were determined by examination

3 60 D. L. WELKER AND K. L. WILLIAMS
TABLE 1 1 -Continued of the phenotypes of the diploid recombinants and of haploid segregants derived from them. Except for segregants of the interval 1 recombinant, which were all bwnA1, phgA1356, couA+, cobA391, the segregants carrying the nonrecombined version of the IV-VI1 rearranged linkage group were distinguishable from those carrying the recombined linkage group on the basis of the presence or absence of the bwnAl marker. The gene order of the phgA and couA loci has not been established and is assumed here for illustrative purposes only. N o recombination events (in either haploids or diploids) between the phgA and couA loci were observed in this work (total of 22 haploid and 11 diploid recombinants), suggesting that these loci are relatively close together on linkage group VII. The translocation breakpoints are indicated by the * symbols. T o clarify the diagram, linkage group 1V DNA is indicated by the dashed lines and linkage group VI1 DNA by the solid lines.
may be an example of a fusion chromosome, possibly even a Robertsonian fusion. This recombination data, considering the possibility of nondisjunction, does not exclude such an event.
Cytological examination of translocation-bearing haploids: Cytological characteri- zation has already been useful in the analysis of strains bearing the D35O(ZZZ,ZZZ) tandem duplication (WELKER, METZ and WILLIAMS 1982) and the fusion chro- mosome T354(ZZ,VZZ) (WELKER and WILLIAMS 1981a); therefore, we made a preliminary cytological examination of haploids bearing 13 of the transloca- tions listed in Table 3. The following haploids had karyotypes containing seven chromosomes: $313 [T35Z(ZZZ,VZ), T359(Z,ZV)]; HL33 [T352(Z,ZZZ)]; NP20 [T353(ZZZ,VZ)]; HU 1048 [T36O(ZZ,VZZ)]; HU 1424 [T36Z(Z,VZ)]; HPS65 [ T362(ZZZ, VU)]; H U 2 1 94 [ T363(ZZZ,ZV)]; H U 8 02 [ T364(Z,ZZZ, VZ)] ; H U 2 209 [T365(ZZl,VZZ)]; and HU22 15 [T366(ZV,VZ)]. Hence, these translocations do not reduce the chromosome number as does the T354(ZZ,VZZ) [and T355(ZZ,VZZ)] translocation and are likely to be reciprocal translocations (or possibly, but less likely, nonreciprocal or insertional translocations).
Cytological examination of HU2043 [T357(ZV,VZZ)] was less conclusive. These preparations were unusual in that most mitotic figures were scored as having six chromosomes, but a minority appeared to have seven chromosomes. However, no unusually large fusion chromosome was observed.
DISCUSSION
Translocations can be identified in D. discoideum on the basis of altered segregation patterns in parasexual genetic analyses of diploids heterozygous for these chromosome rearrangements. In particular, the apparent cosegregation of markers known to map to different linkage groups was used to identify 14 independently derived translocations (Table 3). We propose that, in D. discoi- deum, as in other eukaryotes such as A. nidulans (KAFER 1962, 1965, 1977; MA and KAFER 1974), these cosegregation patterns arise due to the production of nonviable duplication/deficiency progeny. Although more complex explanations for this phenomenon, for example, incompatibility mutations, can be and have been proposed (NEWELL 1978; ROSS and NEWELL 1979), there are now two unambiguous cases in which such cosegregation patterns have been shown to reflect the presence of translocations. The T354(ZZ,VZZ) translocation was estab- lished by cytological and genetic tests to be a translocation of most or all of

TRANSLOCATIONS IN D. DISCOIDEUM 361
linkage group VI1 to the noncentromeric telomere of linkage group I1 (WELKER and WILLIAMS 198 la); whereas the T357(ZV,VZZ) translocation, which may be another example of a fusion chromosome, was established in this work by studies involving mitotic recombination. In addition, the demonstration of transloca- tions, including T357(ZV,VZZ), that arose spontaneously is inconsistent with an alternative hypothesis involving incompatibility mutations (NEWELL 1978; ROSS and NEWELL 1979), since such explanations for the cosegregation phenomenon require multiple mutations.
In D. discoideum, as in A. nidulans (KAFER 1977), the frequency of mitotic recombination, detected by the production of haploid mitotic recombinants, was increased in diploids heterozygous for translocations. Frequencies of haploid mitotic recombinants as high as 5% of all haploid segregants were observed (Table 3). This phenomenon, although not understood, could involve chromo- some breakage of the rearranged chromosomes in the heterozygous diploids via chromosome bridges during mitosis, followed by recombination of the chromo- some fragments with their untranslocated homologs.
If the T357(ZV,VZZ) translocation is another example of a fusion chromosome, then, in order to explain the cytological data, it may break at a high frequency. However, the data obtained from genetic analyses of haploid strains carrying this translocation do not indicate frequent curing of the translocation, which might result from such breakage. No example is known of a strain that should have carried this translocation but that no longer cosegregated linkage group IV and VI1 markers. If the nuclei with apparently seven chromosomes do arise by breakage of the rearranged chromosome, one explanation may be that these products are inviable. A precedent for this is the work of BRODY and WILLIAMS (1 974) in which numerous aneuploid nuclei were seen in cytological preparations of diploids but in which no viable aneuploids were recovered. Unlike the earlier identified fusion chromosome (WELKER and WILLIAMS 1981a), there was no unusually large chromosome in preparations of strains carrying the T357(ZV,VZZ) translocation. However, this can be explained if the chromosome corresponding to linkage group IV is small. One possibility, not inconsistent with the genetic and cytological data, is that this chromosome rearrangement is a Robertsonian translocation. Mitotic figures might be misinterpreted as having seven rather than six chromosomes if both chromosome arms of a metacentric fusion chro- mosome were scored as separate acrocentric chromosomes.
The work reported here establishes that new translocations arise in D. discoi- deum at easily detectable frequencies. An approximate estimate is that 1 % or more of all strains will carry a new spontaneous translocation. Once identified, the translocations can be maintained and carried through genetic analyses, including those involving mitotic recombination of the affected linkage groups (Tables 10 and 11). Mutagenesis with MNNG and defective DNA repair are both implicated in the origin of translocations. In particular, defects at the radB and radG loci may be important. I t is interesting to note that both the radB and radC gene products are required for normal cell growth, since mutations in either locus lead to pleiotropic DNA repair defects and temperature sensitivities (WELKER and WILLIAMS 198 1 b).

362 D. L. WELKER AND K. L. WILLIAMS
Of the 14 translocations of certain independent origin, five affect linkage group I, two affect linkage group 11, seven affect linkage group 111, five affect linkage group IV, five affect linkage group VI and five affect linkage group VII. If one assumes that the number of known markers on each linkage group (Table 2) reflects its size, then linkage group I1 is underrepresented and linkage groups VI and VI1 are overrepresented. This suggests that translocations in D. discoideum do not occur randomly. Moreover, particular pairs of linkage groups tend to rearrange (e.g., linkage group I with linkage group IV; linkage group I1 with linkage group VII ; linkage group 111 with linkage group VI). This suggests that DNA sequences near the translocation breakpoints were important in the for- mation of the rearrangements, possibly due to the presence of transposable elements (GOLDBERG et al. 1983).
We have shown that there were no detectable chromosome rearrangements among pairs of the six established linkage groups between a nonmutated stock of NC4 close to the original wild isolate and a well-marked tester strain derived from NC4, HU1628 (Table 8b). Tester strain HU1628 carries genetic markers on each of the six established linkage groups plus the dominant cobalt resistance mutation cob-354, which is associated with the endogenous D. discoideum plasmid Ddpl (METZ et al. 1983). With HU1628 and similar strains it is now possible to study the extent of chromosome rearrangements between NC4 and other wild isolates of D. discoideum. Not only are gross changes in chromosome structure detectable but also rearrangements involving single genes using DNA restriction fragment length polymorphisms (D. L. WELKER, unpublished data). Such studies are expected to greatly expand our knowledge of genome organization and of gene regulation in D. discoideum.
We wish to t h a n k KAREN DUFFY ant1 G. VOCEI. (both of the Gesamtliochschule Wuppertal, Federal Republic of Gerniany), P. C. NEWELL (University of Oxford, United Kingdom) and S. KAYMAN (Albert Einstein College of Medicine, New York) for personal communications of unpublished results. We thank G. RUGER and H. DAHNE for technical assistance, S. DVI'HORN for typing the manuscript and PETER HUGHES for help with word processing. This investiption was made possible by support from the Max Planck Gesellschaft.
LITERATURE CITED
BRODY, T. and K. I. . WII.I.IAMS, 1974 Cytological analysis of the parasexual cycle in Dictyostelium discoideum. J. Gen. Microbiol. 82: 37 1-383.
DEERING, U. A., M. S. SMITH, B. K. THOMPSON arid A. C. ADOLF, 1970 Ganirna-ray-resistant and - sensitive strains of slime niold (Dictyostelium discoideum). Radiat. Res. 43: 7 1 1-728.
DIPAOLO, J. A., S. C . AMSRAUGH and N. C . POPESCU, 1980 Antipain inhibits N-methyl-N'-nitro-N- iiitrosoguanidine-induced transformation and increases rhroniosonial aberrations. Proc. Natl. Acad. Sri. USA 77: 6649-6653.
DIPAOLO, J. A. and N. C. POPESCU, 1977 Banding pattern analysis of initial structural chromosome alterations induced by N-niethyl-N'-nitro-N-nitrosoguanidine in Syrian hamster cells. Mutat. Res. 44: 3.59-368.
FREE, S. 1 . . U. '1'. SCHIMKE and W. F. LOOMIS, 1976 Dictyostelium discoideum. Genetics 84: 159-1 74.
Annu. Rev. Genet. 16: 385-404.
The structural gene for a-niannosidase i n
GODFREY, S . S . and M. SUSSMAN, 1982 The generics o f development in Dictyostelium discoideum.

TRANSLOCATIONS IN D. DISCOIDEUM 363
GOLDBERG, M. L., J-Y. SHEEN, W. J. GEHRING and M. M. GREEN, 1983 Unequal crossing-over associated with asymmetrical synapsis between nomadic elements in the Drosophila melanogaster genome. Proc. Natl. Acad. Sci. USA 8 0 5017-5021.
KAFER, E., 1962
KAFER, E., 1965
KAFER, E., 1977
Translocations in stock strains of Aspergillus nidulans. Genetics 33: 59-68.
Origins of translocations in Aspergillis nidulans. Genetics 52: 21 7-232.
Meiotic and mitotic recombination in Aspergillus and its chromosomal aberrations.
Use of nystatin-resistant mutations in
Adv. Genet. 19: 33-1 3 1.
KASREKAR, D. P., S. MADICAN and E. R. KATL, 1983 parasexual genetic analysis in Dictyostelium discoideum. Genetics 104: 27 1-277.
KINSELLA, A. K. and M. RADMAN, 1980 Inhibition of carcinogen-induced chromosomal aberrations by an anticarcinogenic protease inhibitor. Proc. Natl. Acad. Sci. USA 77: 3544-3547.
i,oOMIS, W. F. (editor), 1982 The Development of Dictyostelium discoideum. Academic Press, New York.
M A , G. L. C. and E. KAFER, 1974 Genetic analysis of the reciprocal translocation T2(1,Vll) of Aspergillus using the technique of mitotic mapping i n homoiygous translocation diploids. Genetics 77: 11-23,
Identification of an endogenous METL, B. A., T. E. WARD, D. L. WELKER and K. L. WILLIAMS, 1983 pl;rsniid i n Dictyostelium discoideum. EMBO J. 2: 5 15-5 19.
MORRISSEY, J. H., S. WHEELER and W. F. LOOMIS, 1980 New loci in Dictyostelium discoideum determining pignient formation and growth on Bacillus subtilis. Genetics 9 6 1 15-1 23.
MOSSES, D . , K. L. WILLIAMS and P. C. NEWELL, 1975 l h e use of mitotic crossing-over for genetic analysis in Dictyostelium discoideum: mapping of linkage group 11. J. Gen. Microbiol. 90: 247- 2
NEWEI.]., P. C., 1978
NEWEI.~., P. C., 1982
Genetics of the cellular slime molds. Annu. Rev. Genet. 12: 69-93.
Genetics. pp. 35-70. In: The Development of Dictyostelium discoideum, Edited by W. F. LOOMIS. Academic Press, New York.
NEWEL]., P. C., R. F. HENDERSON, D. MossEsand D. I . RATNER, 1977 Sensitivity toBacillus subtilis: a novel system for selection of hetero7ygous diploids of Dictyostelium discoideum. J. Gen. Microbiol. 1 0 0 207-2 1 1.
NEWELI., P. C. and F. M. ROSS, 1982 Genetic analysis of the slug stage of Dictyostelium discoideum. J . Gen. Microbiol. 128: 1639-1652.
KORSON, G. E. and K. L. WII.I.IAMS, 1977 The mitotic chromosomes of the cellular slime mould Dzctyostelium discoideum: a karyotype based on giemsa banding. J. Gen. Microbiol. 99: 19 1-200.
ROSS, F. M. and P. C. NEWELL, 1979 Genetics of aggregation pattern mutations in the cellular slinie mould Dictyostelium discoideum. J. Gen. Microbiol. 115: 289-300.
VOCEI., G., L. THILO, H. SCHWARTZ and R. STEINHART, 1980 Mechanism of phagocytosis i n Dictyostelium discoideum: phagocytosis is mediated by different recognition sites as disclosed by niutants with altered phagocytotic properties. J. Cell Biol. 8 6 456-465.
Genetic analysis by mitotic recombination in Dictyostelium discoideum of growth and developmental loci on linkage group VI]. J. Gen. Microbiol. 128: 953- 964.
Stimulation of mitotic recombination i n Dictyostelium
Genetic analysis of radiation-sensitive mutations in the
WALLACE, J . S. and P. C . NEWELL, 1982a
U'AI.I.ACE, J. S. and P. C. NEWELL, 1 982b discoidrum by ultraviolet irradiation. FEMS Microbiol. Lett. 14: 37-4 1.
slinic mould Dictyostelium discoideum. J. Gen. Microbiol. 97: 1-1 0. U'EI.KF.R, D. 1.. and R. A. DEERING, 1976
WELKER, I). 1.. and K. A. DEERIN(;, 1978 Genetics of radiation sensitivity in the slime mould Dictyostplium discoideum. J. (;en, Mi(-1-obiol. 109: 1 1-23.

3 64 D. L. WELKER AND K. L. WILLIAMS
WELKER, D. L., B. A. METZ and K. L. WILLIAMS, 1982 Chromosome rearrangements in Dictyostelium
Mitotic arrest and chromosome doubling using thiabendazole, cambendazole, nocodazole and benlate in the slime mould Dictyostelium discoideum. J. Gen. Microbiol. 116: 397-407.
The assignment of four new loci, including the coumarin sensitivity locus couA, to linkage group VI1 of Dictyostelium discoideum. J. Gen. Microbiol. 120: 149-159.
WELKER, D. L. and K. I-. WILLIAMS, 1980c Bacillus subtilis sensitivity loci in Dictyostelium discoideum.
Genetic and cytological characterisation of fusion
WELKER, D. L. and K . L. WILLIAMS, 1981b Temperature-sensitive D N A repair mutations in the
A genetic map of Dictyostelium discoideum based on
WELKER, D. L. and K. L. WILLIAMS, 1982b Genetic analysis and phenotypic characterization of effects on the cytoskeleton of coumarin-sensitivity mutations in Dictyostelium discoideum. J. Gen. Microbiol. 128: 1329-1343.
discoideum. Genetics 102: 71 1-723.
WELKER, D. L. and K. L. WILLIAMS, 1980a
WELKER, D. L. and K. L. WILLIAMS, 1980b
FEMS Microbiol. Lett. 9 179-183.
WELKER, D. I.. and K. L. WILLIAMS, 1981a chroniosonies of Dictyostelium discoideum. Chromosoma (Berl.) 82: 32 1-332.
cellular slime mould Dictyostelium discoideum. Curr. Genet. 3: 167-1 7 1.
WELKER, D. L. and K . 1.. WILLIAMS, 1982a mitotic recombination. Genetics 102: 691-710.
WELKER, D. L. and K. L. WILLIAMS, 1983 Genetic loci associated with altered resistance to microtubule inhibitors and with spore shape in Dictyostelium discoideum. J. Gen. Microbiol. 1 2 9 2207-22 16.
Characterization of dominant resistance to cobalt chloride in Dictyostelium
Parasexual genetics in the cellular slime mould Dictyostelium
WILIJAMS, K . I>. , 1978 discoideum and its use in parasexual genetic analysis. Genetics 90: 37-47.
WILLIAMS, K . L. and P. BARRAND, 1978 discoideum: haploidisation of diploid strains using benlate. FEMS Microbiol. Lett. 4: 155-1 59.
WILLIAMS, K . L., R. H. KESSIN and P. C. NEWELL, 1974 Parasexual genetics in Dictyostelium discoideum: mitotic analysis of acriflavin resistance and growth in axenic medium. J. Gen. Microbiol. 84: 59-69.
WILLIAMS, K . Is . , G. E. ROBSON and D. L. WELKER, 1980 Chromosome fragments in Dictyostelium discoideum obtained from parasexual crosses between strains of different genetic background. Genetics 95: 289-304.
Communicating editor: S. L. ALLEN


















