Translocation of constitutively expressed heat shock protein Hsc70 to synapse-enriched areas of the...
8
Translocation of Constitutively Expressed Heat Shock Protein Hsc70 to Synapse-Enriched Areas of the Cerebral Cortex After Hyperthermic Stress Sheng Chen and Ian R. Brown * Centre for the Neurobiology of Stress, University of Toronto at Scarborough, Ontario, Canada Hsp70 is a multi-gene family composed of stress-induci- ble members (Hsp70) and other members that are consti- tutively expressed (Hsc70). The heat shock proteins Hsp70 and Hsc70 exhibit similar molecular structure and biochemical functions. Constitutively expressed Hsc70 is enriched in the mammalian nervous system compared with non-neural tissues and present at high levels in neu- ronal cell bodies. After thermal stress, Hsc70 is translo- cated to synapse-enriched areas of the cerebral cortex where it associates with Hsp40 to form a complex that can refold denaturated proteins. These results suggest that the heat shock response in the nervous system in- volves not only the synthesis of stress-inducible Hsps but also the translocation of constitutively expressed Hsc70 to synapse-enriched areas where it could participate in neuroprotective mechanisms that preserved synaptic func- tion during times of stress. V V C 2007 Wiley-Liss, Inc. Key words: heat shock proteins; constitutively expres- sed Hsc70; Hsp40; protein refolding Heat shock ‘preconditioning,’ that is exposure to sub- lethal temperatures, induces the expression of heat shock proteins (Hsps) and protects tissues and organisms from cell death that would normally result from exposure to lethal temperatures and other forms of stress (Morimoto et al., 1997). Prior heat shock has been reported to protect neural cells against subsequent stressful stimuli in both tissue cul- ture systems and in the intact nervous system (Walsh et al., 1987; Barbe et al., 1988; Walsh et al., 1989; Rordorf et al., 1991; Mayer and Brown, 1994; Tytell et al., 1994; Yenari, 2002). An interesting question that has received compara- tively little attention is whether heat shock preconditioning protects physiological processes in the nervous system such as synaptic function. Synapses are critical points of information transfer and their functionality must be preserved during stressful conditions to prevent communication breakdown. It is increasingly accepted that synapses are vulnerable regions of neurons (Mattson and Magnus, 2006). Using a macropatch electrode to record synaptic activity at individ- ual synaptic boutons in the Drosophila larval neuromuscular junction, our collaborative studies have shown that prior heat shock protects synaptic transmission from subsequent high test temperatures through both presynaptic and postsy- naptic modifications (Karunanithi et al., 1999). An enhance- ment of the level of synaptic protection is observed in trans- genic Drosophila that have been engineered to overexpress Hsp70 (Karunanithi et al., 2002). Thermal precondition- ing has also been reported to protect synaptic transmission events recorded in mammalian brain slices (Kelty et al., 2002). Hence the protective scenario in the nervous system conferred by heat shock preconditioning has been extended to synaptic function. Hsp70 is a multi-gene family composed of some members that are stress-inducible (Hsp70) and other mem- bers that are constitutively expressed (Hsc70). It is notewor- thy that Hsp70 and Hsc70 proteins exhibit almost identical molecular structure and indistinguishable biochemical func- tions (Hightower et al., 1994). Heat shock pre-conditioning paradigms trigger the synthesis of stress-inducible Hsp70, hence protective mechanisms have tended to focus on this stress-inducible Hsp. However, Hsc70 has also been hypo- thesized to play a role in thermotolerance and stress resist- ance (diIorio et al., 1996). Increased levels of Hsc70 corre- late with increased thermotolerance in species of lizards (Ulmasov et al., 1992) and desert-adapted humans (Lyashko et al., 1994). In vitro studies on purified mammalian Hsc70 have reported that the protein undergoes a conformational change that activates its peptide/unfolded-protein binding activity as temperature is increased (Leung et al., 1996). This heat-induced Hsc70 conformational change may be associated with acquired thermotolerance (Leung et al., 1996). We have noted that constitutive Hsc70 protein is enriched in the mammalian nervous system compared with Contract grant sponsor: NSERC Canada. *Correspondence to: Dr. Ian R. Brown, Centre for the Neurobiology of Stress, University of Toronto at Scarborough, 1265 Military Trail, Toronto, Ontario, Canada, M1C 1A4. E-mail: [email protected] Received 1 August 2006; Revised 6 September 2006; Accepted 20 September 2006 Published online 3 January 2007 in Wiley InterScience (www. interscience.wiley.com). DOI: 10.1002/jnr.21124 Journal of Neuroscience Research 85:402–409 (2007) ' 2007 Wiley-Liss, Inc.
-
Upload
sheng-chen -
Category
Documents
-
view
212 -
download
0
Transcript of Translocation of constitutively expressed heat shock protein Hsc70 to synapse-enriched areas of the...

Translocation of ConstitutivelyExpressed Heat Shock Protein Hsc70 toSynapse-Enriched Areas of the CerebralCortex After Hyperthermic Stress
Sheng Chen and Ian R. Brown*
Centre for the Neurobiology of Stress, University of Toronto at Scarborough, Ontario, Canada
Hsp70 is a multi-gene family composed of stress-induci-ble members (Hsp70) and other members that are consti-tutively expressed (Hsc70). The heat shock proteinsHsp70 and Hsc70 exhibit similar molecular structure andbiochemical functions. Constitutively expressed Hsc70 isenriched in the mammalian nervous system comparedwith non-neural tissues and present at high levels in neu-ronal cell bodies. After thermal stress, Hsc70 is translo-cated to synapse-enriched areas of the cerebral cortexwhere it associates with Hsp40 to form a complex thatcan refold denaturated proteins. These results suggestthat the heat shock response in the nervous system in-volves not only the synthesis of stress-inducible Hsps butalso the translocation of constitutively expressed Hsc70to synapse-enriched areas where it could participate inneuroprotective mechanisms that preserved synaptic func-tion during times of stress. VVC 2007 Wiley-Liss, Inc.
Key words: heat shock proteins; constitutively expres-sed Hsc70; Hsp40; protein refolding
Heat shock ‘preconditioning,’ that is exposure to sub-lethal temperatures, induces the expression of heat shockproteins (Hsps) and protects tissues and organisms from celldeath that would normally result from exposure to lethaltemperatures and other forms of stress (Morimoto et al.,1997). Prior heat shock has been reported to protect neuralcells against subsequent stressful stimuli in both tissue cul-ture systems and in the intact nervous system (Walsh et al.,1987; Barbe et al., 1988; Walsh et al., 1989; Rordorf et al.,1991; Mayer and Brown, 1994; Tytell et al., 1994; Yenari,2002).
An interesting question that has received compara-tively little attention is whether heat shock preconditioningprotects physiological processes in the nervous system such assynaptic function. Synapses are critical points of informationtransfer and their functionality must be preserved duringstressful conditions to prevent communication breakdown.It is increasingly accepted that synapses are vulnerableregions of neurons (Mattson and Magnus, 2006). Using amacropatch electrode to record synaptic activity at individ-ual synaptic boutons in the Drosophila larval neuromuscularjunction, our collaborative studies have shown that prior
heat shock protects synaptic transmission from subsequenthigh test temperatures through both presynaptic and postsy-naptic modifications (Karunanithi et al., 1999). An enhance-ment of the level of synaptic protection is observed in trans-genic Drosophila that have been engineered to overexpressHsp70 (Karunanithi et al., 2002). Thermal precondition-ing has also been reported to protect synaptic transmissionevents recorded in mammalian brain slices (Kelty et al.,2002). Hence the protective scenario in the nervous systemconferred by heat shock preconditioning has been extendedto synaptic function.
Hsp70 is a multi-gene family composed of somemembers that are stress-inducible (Hsp70) and other mem-bers that are constitutively expressed (Hsc70). It is notewor-thy that Hsp70 and Hsc70 proteins exhibit almost identicalmolecular structure and indistinguishable biochemical func-tions (Hightower et al., 1994). Heat shock pre-conditioningparadigms trigger the synthesis of stress-inducible Hsp70,hence protective mechanisms have tended to focus on thisstress-inducible Hsp. However, Hsc70 has also been hypo-thesized to play a role in thermotolerance and stress resist-ance (diIorio et al., 1996). Increased levels of Hsc70 corre-late with increased thermotolerance in species of lizards(Ulmasov et al., 1992) and desert-adapted humans (Lyashkoet al., 1994). In vitro studies on purified mammalian Hsc70have reported that the protein undergoes a conformationalchange that activates its peptide/unfolded-protein bindingactivity as temperature is increased (Leung et al., 1996).This heat-induced Hsc70 conformational change may beassociated with acquired thermotolerance (Leung et al.,1996).
We have noted that constitutive Hsc70 protein isenriched in the mammalian nervous system compared with
Contract grant sponsor: NSERC Canada.
*Correspondence to: Dr. Ian R. Brown, Centre for the Neurobiology of
Stress, University of Toronto at Scarborough, 1265 Military Trail,
Toronto, Ontario, Canada, M1C 1A4. E-mail: [email protected]
Received 1 August 2006; Revised 6 September 2006; Accepted 20
September 2006
Published online 3 January 2007 in Wiley InterScience (www.
interscience.wiley.com). DOI: 10.1002/jnr.21124
Journal of Neuroscience Research 85:402–409 (2007)
' 2007 Wiley-Liss, Inc.

non-neural tissue and is present at high levels in neuronalcell bodies (Manzerra et al., 1993, 1997; Manzerra andBrown, 1996). In the current study, we report that consti-tutively expressed Hsc70 is translocated to synapse-enrichedareas of the cerebral cortex after thermal stress. This suggeststhat the heat shock response in the nervous system involvesnot only the synthesis of stress-inducible Hsps but also trans-location of constitutively expressed Hsc70 to synaptic ter-minals where it could participate in neuroprotective mecha-nisms that preserve synaptic function during times of stress.
MATERIALS AND METHODS
Treatment of Animals
All procedures using animals were approved by the Ani-mal Care Committee of the University of Toronto and werein accordance with the guidelines established by the CanadianCouncil on Animal Care. The body temperature of adultmale Wistar rats (35 days old) was raised 3.58C above normal(388C) by using a dry incubator set at 458C. The body tem-perature of the animals was monitored with a rectal thermalprobe (Physitemp Instruments, Clifton, NJ) and maintained atthe elevated temperature for 1 hr. After incubation, the rats wereplaced at room temperature for recovery until being sacrificed.
Immunohistochemistry
Rats were anesthetized with sodium pentobarbital andperfused intracardially with cold 0.1 M phosphate-buffered sa-line (PBS, pH 7.4), followed by 4% paraformaldehyde in PBS.Brain tissue was removed and fixed overnight in 4% parafor-maldehyde in PBS at 48C. Tissue was then equilibrated with20% sucrose in PBS, mounted in OCT embedding compound(Miles, Elkhart, IN), and frozen at�708C until use. Cryostat sec-tions (10 lm) were collected on gelatin-coated microscopeslides and air-dried for 3 hr. After rehydration in PBS-Tbuffer (0.1 M PBS, pH 7.4, 0.2% Triton X-100, 0.1% BSA),tissue sections were blocked in 10% goat serum in PBS-T for1 hr. Sections were incubated overnight with primary anti-bodies diluted in PBS-T consisting of 1:500 mouse anti-NeuN (MAB377; Chemicon, Temecula, CA) in combinationwith either 1:1,000 rabbit anti-Hsp40 (SPA-400; Stressgen),or 1:1,000 rat-anti-Hsc70 (SPA-815, Stressgen Biotechnolo-gies, Victoria, Canada), or 1:1,000 rabbit-anti-Hsp70 (SPA-812, Stressgen) antibodies, or 1:500 rabbit anti-synaptophysin(RB-1461, Lab Vision, Freemont, CA). After washing in PBS-T,sections were incubated for 2 hr with secondary antibodies(Jackson ImmunoResearch Laboratories, West Grove, PA) di-luted in PBS-T consisting of 1:100 Cy5 labeled anti-mouse IgGfor NeuN in combination with 1) 1:200 Cy3 labeled anti-rabbitIgG for Hsp40, Hsp70, or synaptophysin; or 2) Rhodamine-red-X labeled anti-rat IgG for Hsc70. For nucleic acid staining, slideswere then incubated with 2 lM Yo-pro-1 (Invitrogen, Carlsbad,CA) in PBS-T for 30 min. After washes in PBS-T, sections weremounted with anti-fading fluorescent mounting media (Dako,Mississauga, Canada).
For the colocalization studies, rat brain sections wereincubated overnight with a combination of the following pri-mary antibodies: 1:1,000 rat-anti-Hsc70 and 1:300 mouse-anti-synaptophysin (S5768, Sigma, St. Louis, MO), or 1:1,000
rat-anti-Hsc70 and 1:1,000 rabbit-anti-Hsp40, or 1:100mouse-anti-Hsp70 (SPA-810, Stressgen) and 1:1,000 rabbit-anti-Hsp40, or 1:1,000 rabbit-anti-Hsp70 and 1:300 mouse-anti-synaptophysin. Sections were then incubated with sec-ondary antibodies (Jackson ImmunoResearch Laboratories) for2 hr: 1:200 Cy5-anti-rat IgG and 1:100 Cy2-anti-mouse IgG(for Hsc70 and synaptophysin), or 1:200 Cy5-anti-rat IgG and1:200 Cy2-anti-rabbit IgG (for Hsc70 and Hsp40), or Cy5-anti-mouse IgG and Cy2-anti-rabbit IgG (for Hsp70 andHsp40), or Cy5-anti-rabbit IgG and Cy2-anti-mouse IgG (forHsp70 and synaptophysin). After washes in PBS-T, sectionswere mounted with anti-fading fluorescent mounting media(Dako). Data representative of six experimental repeats are shown.
Image Analysis
Quantitative immunohistochemistry with confocal mi-croscopy and image analysis software has been used widely inbiomedical research to compare fluorescent intensities repre-senting proteins levels in selected image areas (Kretz et al.,2004; Buttini et al., 2005). Slides were analyzed using a ZeissLSM510 confocal microscope. Detection configurations wereadjusted to obtain images with pixel intensity within a linearrange and were kept constant for different slides. Backgroundfluorescent intensities were determined by scanning the nega-tive control slides (omission of primary antibodies) with thesame detection configurations.
Quantitative colocalization analysis is a powerful tool forthe reliable estimation of the colocalization of proteins (Zin-chuk et al., 2005). Fifty regions of interest (ROI) with an areaof approximately 20 lm2 from each time point were ran-domly selected and used for quantitative colocalization analysiswith the Zeiss LSM510 software (version 3.2 with servicepack 2). The colocalization coefficient, which indicates therelative number of colocalizing pixels as compared with thetotal number of pixels above the threshold (or background),was used as a quantitative parameter. For instance, for a coloc-alization analysis of synaptophysin and Hsc70, pixels in eachROI were classified into four categories: a) for pixels that areonly positive for synaptophysin; b) for pixels that are onlypositive for Hsc70; c) for pixels that are positive for both syn-aptophysin and Hsc70, representing colocalization; and d) forpixels that are neither positive for synaptophysin nor Hsc70.The colocalization coefficient for synaptophysin in this ROI isthen expressed as c/(a + c).
Western Blotting
Cerebral cortex tissue was harvested and kept at �708Cuntil use. Tissue was homogenized in ice-cold 0.32 M sucrosewith a protease inhibitor cocktail (5 lg/ml of antipain, aproti-nin, and leupeptin). Protein concentration of the homogenateswas determined with the Bio-Rad protein assay kit (Hercules,CA). Equal amounts of protein samples were solubilized byboiling in 23 SDS loading buffer (100 mM Tris, pH 7.4, 4%SDS, 200 mM DTT, 20% glycerol, 0.2% bromophenol blue),separated by SDS-PAGE with 5% stacking gel and 10% sepa-rating gel, and transferred to a nitrocellulose membrane. Afterblocking for 1 hr with 5% fat-free milk powder in TBST(10 mM Tris, 250 mM NaCl, 0.5% Tween-20, pH 7.4), blotswere incubated overnight at room temperature with primary
Translocation of Hsc70 to Synaptic Areas 403
Journal of Neuroscience Research DOI 10.1002/jnr
antibodies in TBST diluted by 1:30,000 for Hsc70 (SPA-815,Stressgen), 1:5,000 for Hsp70 (SPA-810, Stressgen), 1:5,000 forHsp40 (SPA-400, Stressgen) and 1:10,000 for synaptophysin(S5768, Sigma). After washes, blots were incubated with HRP-conjugated secondary antibodies (Sigma) for 2 hr in TBST atroom temperature. Immunoactivity was visualized with ECLWestern blot detection reagents (Amersham, Piscataway, NJ)and quantified with Fluorchem (Alpha Innotech, San Leandro,CA). Data representative of six experimental repeats are shown.
Coimmunoprecipitation
Cerebral cortex tissue was homogenized in ice-cold ho-mogenization buffer (150 mM NaCl, 20 mM Tris, pH 7.4,10 mM EDTA, 0.2% Triton X-100, 5 lg/ml of antipain,aprotinin and leupeptin). Samples were centrifuged at 10,0003 g for 20 min at 48C. The protein concentration of the su-pernatant was determined using the Bio-Rad protein assay kit.Protein samples (1 mg) were incubated with 10 lg of non-specific rat or mouse IgG (Sigma) and 40 ll of protein A/Gplus agarose slurry (Santa Cruz Biotechnology, Santa Cruz, CA)on a rotating wheel at 48C for 2 hr. After centrifugation (2,0003 g for 5 min), 10 lg of Hsc70 antibody (SPA-815, Stressgen)or Hsp70 antibody (SPA-810, Stressgen) and 40 ll protein A/Gplus agarose slurry were added to the supernatants. The reactionmixture was placed on a rotating wheel at 48C overnight. Theprecipitates were collected after centrifugation at 2,000 3 g for5 min, and washed 3 times with cold homogenization buffer.Protein in the precipitate was eluted by boiling the precipitatein 40 ll of 13 SDS loading buffer (50 mM Tris, pH 7.4, 2%
SDS, 100 mM DTT, 10% glycerol, 0.1% bromophenol blue),and Western blot analysis was carried out. Data representative ofthree experimental repeats are shown.
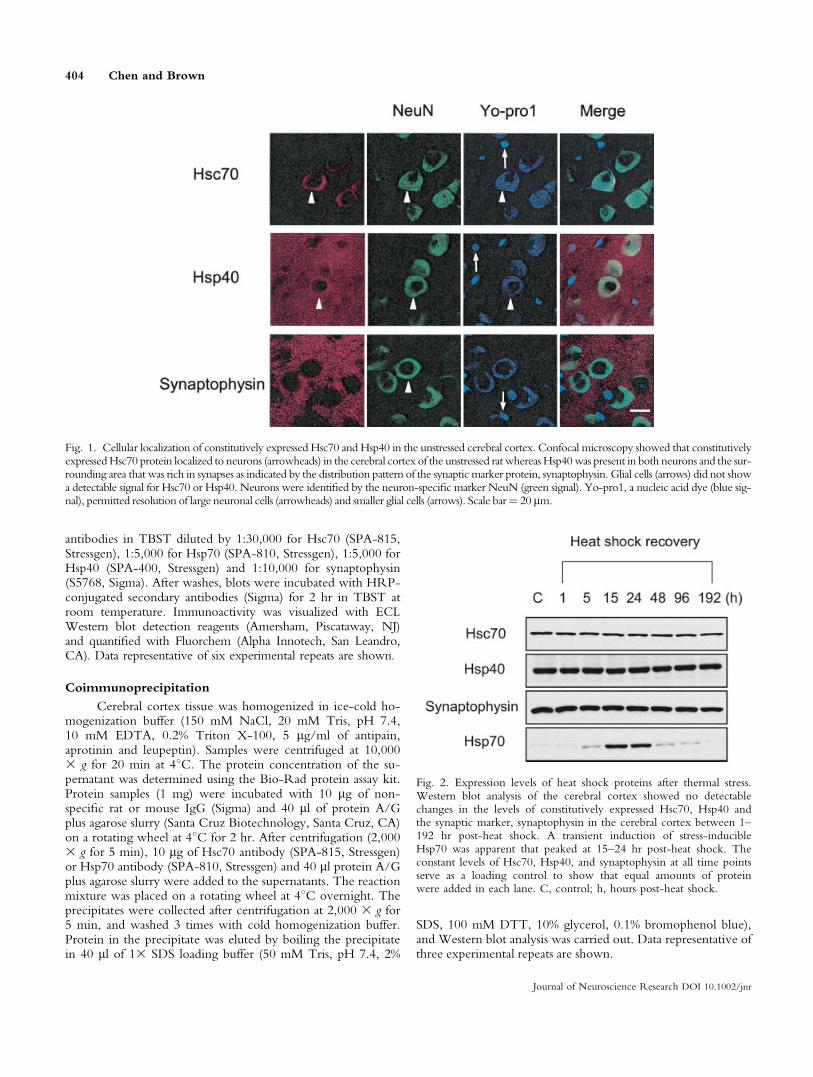
Fig. 1. Cellular localization of constitutively expressed Hsc70 and Hsp40 in the unstressed cerebral cortex. Confocal microscopy showed that constitutivelyexpressedHsc70 protein localized to neurons (arrowheads) in the cerebral cortex of the unstressed rat whereas Hsp40was present in both neurons and the sur-rounding area that was rich in synapses as indicated by the distribution pattern of the synaptic marker protein, synaptophysin. Glial cells (arrows) did not showa detectable signal for Hsc70 or Hsp40. Neurons were identified by the neuron-specific marker NeuN (green signal). Yo-pro1, a nucleic acid dye (blue sig-nal), permitted resolution of large neuronal cells (arrowheads) and smaller glial cells (arrows). Scale bar¼ 20 lm.
Fig. 2. Expression levels of heat shock proteins after thermal stress.Western blot analysis of the cerebral cortex showed no detectablechanges in the levels of constitutively expressed Hsc70, Hsp40 andthe synaptic marker, synaptophysin in the cerebral cortex between 1–192 hr post-heat shock. A transient induction of stress-inducibleHsp70 was apparent that peaked at 15–24 hr post-heat shock. Theconstant levels of Hsc70, Hsp40, and synaptophysin at all time pointsserve as a loading control to show that equal amounts of proteinwere added in each lane. C, control; h, hours post-heat shock.
404 Chen and Brown
Journal of Neuroscience Research DOI 10.1002/jnr

Statistic Analysis
The results are expressed as the mean 6 SD and the sta-tistical significance was assessed by one-way ANOVA.
RESULTS
Distribution Pattern of Constitutively ExpressedHsc70 and Hsp40 in the Unstressed Rat CerebralCortex
As shown in Figure 1, constitutively expressed Hsc70protein localized to neurons (red signal indicated by arrow-heads) in the cerebral cortex of the unstressed rat that wereidentified by the neuron-specific marker protein NeuN(green signal) (Mullen et al., 1992). Hsp40 localized to neu-rons and was also present in a punctuate pattern in the sur-rounding area that is rich in synapses as indicated by distri-bution pattern of synaptophysin, a widely used synapticmarker protein (McMahon et al., 1996; Kretz et al., 2004;Buttini et al., 2005). Smaller glial cells (indicated by arrows)do not show a detectable signal for Hsc70 or Hsp40. Yo-pro1, a marker for nucleic acids stained both the large neu-rons (arrowheads) and the smaller glial cells (arrows).
Effect of Thermal Stress on Expression Levelsof Heat Shock Proteins
A time-course Western blot analysis showed that heatshock did not change the overall levels of constitutivelyexpressed Hsc70 or Hsp40 protein in the rat cerebral cortexnor did it alter levels of the synaptic marker protein synap-tophysin from 1–192 hr post-stress (Fig. 2). However, thewhole body hyperthermic treatment triggered the transientinduction of stress-inducible Hsp70 that appeared at 5 hrpost-stress and peaked at 15–24 hr.
Heat Shock Induces a Translocationof Constitutively Expressed Hsc70 toSynapse-Enriched Areas
As shown in the upper panel of Figure 3A for theunstressed rat, constitutively expressed Hsc70 localized toneuronal cell bodies (red signal) in the cerebral cortex andwas not detectable in the surrounding field that was rich insynaptophysin, a synaptic marker protein (green signal).The merged image of these two signals showed no overlapin the unstressed cerebral cortex. After heat shock, no over-
Fig. 3. Translocation of constitu-tively expressed Hsc70 to synapseenriched areas after thermal stress.A: Immunocytochemistry showedthe translocation of constitutivelyexpressed Hsc70 (red signal) tosynapse enriched areas at 24 hrpost-heat shock. Localization ofsynaptophysin, a synaptic markerprotein, is indicated by the greensignal. The merged image (yellowsignal) shows the pattern of coloc-alization of Hsc70 and synapto-physin in the synaptic enrichedarea surrounding the neurons at24 hr post-heat shock. Scale bar ¼20 lm. B: Quantitative analysisindicating that the transient coloc-alization of Hsc70 and synapto-physin in synapse-enriched areaspeaked at 15–48 hr after thermalstress.
Translocation of Hsc70 to Synaptic Areas 405
Journal of Neuroscience Research DOI 10.1002/jnr

all change in levels of constitutively expressed Hsc70 pro-tein were detected (Fig. 2). However, at 24 hr post-heatshock (lower panel in Fig. 3A), Hsc70 protein translocatedto the synapse-enriched field as indicated by the appearanceof a punctuate Hsc70 signal in the area surrounding theneuronal cell bodies that colocalized with the synapticmaker synaptophysin (as indicated by the yellow signal inthe merged image). A quantitative time course analysisshowed that the colocalization of constitutively expressedHsc70 protein and the synaptic marker protein was transientand peaked at 15–48 hr after heat shock in the synapse-enriched field (Fig. 3B).
Increased Interaction of Hsc70 and Hsp40After Heat Shock
The functioning of Hsc70 protein requires the assis-tance of Hsp40 that delivers protein candidates to Hsc70and stimulates the ATPase activity of Hsc70 to mediate cor-rect protein folding (Fan et al., 2003). The association ofHsc70 with Hsp40 was investigated by coimmunoprecipita-tion analysis. This showed that Hsc70 and Hsp40 are associ-ated in the unstressed cerebral cortex, as expected, inHsc70/Hsp40 chaperoning machinery engaged in the fold-ing process of newly synthesized proteins (control lane C in
Fig. 4A). Heat shock induces protein unfolding (Lepock,2005). The association of Hsc70 and Hsp40 in the cerebralcortex showed a transient increase after heat shock (Fig. 4A)that peaked at 15–48 hr post-stress (Fig. 4B). This likelyreflected increased cooperation of Hsc70 and Hsp40 to han-dle the appearance of an increased amount of unfolded pro-teins induced by the thermal stress.
Increased Colocalization of Hsc70 and Hsp40in Synapse-Enriched Areas After Heat Shock
As shown in the top panel of Figure 5A, in theunstressed cerebral cortex, Hsc70 was localized to the neu-ronal cell body (red signal) whereas Hsp40 was present inboth the neuronal cell body and the surrounding synapse-enriched field (green signal). The merged signal (yellow)showed the colocalization of the Hsc70 and Hsp40 in thecytoplasm of the neuronal cell bodies in the unstressed cere-bral cortex. After heat shock (bottom panel, Fig. 5A), apunctuate Hsc70 signal appeared in the synapse-enrichedfield and colocalized with Hsp40 as indicated by the yellowsignal in the merged image. Quantitative analysis of thecolocalization of Hsc70 and Hsp40 in the synaptic fieldshowed that the association was transient and peaked at15–48 hr post-stress (Fig. 5B). This could reflect the tran-sient association of Hsc70 and Hsp40 in chaperone machin-ery located at the synapse that refolds synaptic proteinsthat have been susceptible to stress-induced unfolding.In addition, the thermal stress induced an increased asso-ciation of Hsc70 and Hsp40 in the cytoplasm of the neuro-nal cell body as reflected by the increased colocalizationsignal (yellow) in the merged image in the lower panel ofFigure 5A.
Induction Pattern of Stress-Inducible Hsp70in the Cerebral Cortex
As shown by Western blotting in Figure 2, heat stresstriggered a transient induction of stress-inducible Hsp70protein in the rat cerebral cortex. Immunocytochemistry(shown in Fig. 6A) showed that this Hsp70 induction (redsignal) occurred in glial cells (indicated by arrows) and notin neurons (indicated by arrowheads). Hsp70 was notdetected in the unstressed cerebral cortex by either Westernblot analysis (Fig. 2) or at the cellular level by immunocyto-chemistry (Fig. 6A). Stress-inducible Hsp70 that wasinduced in glial cells was found to colocalize with Hsp40 at24 hr post-heat shock (Fig. 6B).
DISCUSSION
In response to stressful stimuli, cells induce the classic‘heat shock response’ in which heat shock proteins areinduced that play important roles in cellular repair and pro-tective mechanisms (Morimoto et al., 1997). Our laboratoryand others have reported that glial cells and neuronal cells inboth tissue culture systems and in the intact nervous systemrespond differentially to thermal stress. For example, glialcells exhibit a strong induction of stress-inducible Hsp70whereas most neuronal cell types, particularly differentiated
Fig. 4. Coimmunoprecipitation analysis of the association of Hsc70and Hsp40 after heat shock. A: Using an Hsc70 specific antibody,coimmunoprecipitation analysis showed that the association of Hsc70with Hsp40 in the cerebral cortex increased transiently after heatshock. Western blots of coimmunoprecipitation products for Hsc70and Hsp40 are shown. The constant level of Hsc70 at all time pointsserves as a loading control to show that equal amounts of proteinwere added in each lane. h, hours after heat shock; negative control(coimmunoprecipitation with non-specific rat IgG) is labeled as (�).B: Quantitative analysis indicating that the transient association ofHsc70 with Hsp40 in the cerebral cortex peaked at 15–48 hr afterheat shock.
406 Chen and Brown
Journal of Neuroscience Research DOI 10.1002/jnr

Fig. 5. Colocalization analysis ofHsc70 and Hsp40 after heat shock.A: Immunocytochemistry showed thetranslocation of Hsc70 (red signal) tosynapse-enriched areas at 24 hr post-heat shock where it colocalized withHsp40 (green signal). The mergedimage of the two signals is indicatedin yellow. Scale bar ¼ 20 lm. B:Quantitation of the time course ofthe transient colocalization of Hsc70and Hsp40 in synapse-enriched areasshowed that the association peaked at15–48 hr post-heat shock.
Fig. 6. Analysis of cell types ex-pressing stress-inducible Hsp70in the cerebral cortex. A:Induction of Hsp70 in glialcells. Immunocytochemistry sh-owed that heat shock inducedHsp70 in glia cells (red signal)and not in neurons (indicatedby the green NeuN signal). B:The stress-inducible Hsp70(red signal) that was expressedin glial cells colocalized withHsp40 (green signal) as indi-cated by the yellow mergedimage. Arrowheads indicateneurons. Arrows indicate glialcells. Scale bars ¼ 20 lm.
Translocation of Hsc70 to Synaptic Areas 407
Journal of Neuroscience Research DOI 10.1002/jnr

neurons, do not (Manzerra and Brown, 1992; Manzerraet al., 1993; Marcuccilli et al., 1996; Foster and Brown,1997; Batulan et al., 2003).
Hsp70 is a multi-gene family composed of somemembers that are stress-inducible, (Hsp70) and other mem-bers that are constitutively expressed in unstressed cells(Hsc70). Interestingly Hsp70 and Hsc70 proteins exhibitalmost identical molecular structure and indistinguishablebiochemical functions (Hightower et al., 1994). We havenoted that constitutively expressed Hsc70 is enriched in themammalian nervous system compared with non-neural tis-sues and is present at high levels in the cytoplasm of neuro-nal cell bodies (Manzerra et al., 1993, 1997; Manzerra andBrown, 1996).
The present study indicates that levels of constitutiveHsc70 do not change in the cerebral cortex of the rat brainafter whole body hyperthermia. However a pronouncedtranslocation of Hsc70 to synapse-enriched areas occurs af-ter thermal stress. This translocation is transient and colocal-ization of Hsc70 with the synaptic marker synaptophysinpeaks at 15–48 hr after the thermal stress.
At the functional level Hsc70 is known to specificallyinteract with Hsp40 to form a chaperone machine that mayalso include proteins such as Hip (Hohfeld et al., 1995).Hsp40 delivers protein candidates to Hsc70 and stimulatesthe ATPase activity of Hsc70 to mediate correct proteinfolding (Fan et al., 2003). Coimmunoprecipitation analysisshowed that the association of Hsc70 with Hsp40 transi-ently increased after heat shock, likely reflecting increasedcooperation of Hsc70 and Hsp40 to deal with the appear-ance of the increased amount of unfolded proteins that areinduced by thermal stress (Lepock, 2005). Confocal immu-nocytochemistry showed a transient increase in the colo-calization of Hsc70 and Hsp40 in synapse-enriched areasafter heat shock, suggesting the formation of a chaperonemachine located at the synapse that could refold synapticproteins that are susceptible to stress-induced unfolding.
These results suggest that the heat shock response inthe nervous system involves not only the synthesis of stress-inducible Hsps but also the translocation of constitutivelyexpressed Hsc70 to synapse-enriched areas where it parti-cipates with Hsp40 in neuroprotective mechanisms thatpreserve synaptic function during times of stress. Hence thedefinition of the heat shock response requires extension toinclude stress-induced redeployment of constitutively ex-pressed Hsc70 involving its translocation to intracellular sitesthat are particularly vulnerable to stress. This protective sce-nario may be particularly relevant to differentiated neuronsthat characteristically exhibit high levels of Hsc70 and donot synthesize stress-inducible Hsp70 after thermal stress(Manzerra and Brown, 1992; Manzerra et al., 1993; Mar-cuccilli et al., 1996; Foster and Brown, 1997; Batulan et al.,2003).
Prior heat shock involving exposure to sub-lethaltemperatures induces Hsps and protects neural cells fromcell death that would normally result from exposure to le-thal temperatures and other forms of stress (Walsh et al.,1987, 1989; Barbe et al., 1988; Rordorf et al., 1991; Mayerand Brown, 1994; Tytell et al., 1994; Yenari, 2002). Heat
shock preconditioning also confers functional protection tosynapses as shown in both Drosophila and mammalian sys-tems (Karunanithi et al., 1999, 2002; Kelty et al., 2002).Neuroprotective mechanisms have tended to focus onstress-inducible Hsp70 that is induced by the precondition-ing paradigms. The present results suggest that the transloca-tion of constitutively expressed Hsc70 to synapses and itscolocalization at these sites with Hsp40 could play a role insynaptic protective mechanisms that preserve neurotrans-mission processes during times of stress. Interestingly thetranslocation of Hsc70 to synapses and its colocalizationwith Hsp40 at these sites is transient and peaks at 15–48 hrafter thermal stress suggesting that the window of synapticprotection conferred by prior heat shock could be substan-tial in duration.
ACKNOWLEDGMENTS
This study was supported by grants to I.B. fromNSERC Canada who also holds a Canada Research Chair.We thank R. Or for technical advice on confocal micros-copy and S. Lively, R. Cheung, and A. Chow for helpfulsuggestions.
REFERENCES
Barbe MF, Tytell M, Gower DJ, Welch WJ. 1988. Hyperthermia pro-
tects against light damage in the rat retina. Science 241:1817–1820.
Batulan Z, Shinder GA, Minotti S, He BP, Doroudchi MM, Nalbanto-
glu J, Strong MJ, Durham HD. 2003. High threshold for induction of
the stress response in motor neurons is associated with failure to activate
HSF1. J Neurosci 23:5789–5798.
Buttini M, Masliah E, Barbour R, Grajeda H, Motter R, Johnson-Wood
K, Khan K, Seubert P, Freedman S, Schenk D, Games D. 2005. Beta-
amyloid immunotherapy prevents synaptic degeneration in a mouse
model of Alzheimer’s disease. J Neurosci 25:9096–9101.
diIorio PJ, Holsinger K, Schultz RJ, Hightower LE. 1996. Quantitative
evidence that both Hsc70 and Hsp70 contribute to thermal adaptation
in hybrids of the livebearing fishes Poeciliopsis. Cell Stress Chaperones
1:139–147.
Fan CY, Lee S, Cyr DM. 2003. Mechanisms for regulation of Hsp70
function by Hsp40. Cell Stress Chaperones 8:309–316.
Foster JA, Brown IR. 1997. Differential induction of heat shock mRNA
in oligodendrocytes, microglia, and astrocytes following hyperthermia.
Brain Res Mol Brain Res 45:207–218.
Hightower L, Sadis S, Takenaka I. 1994. Interaction of vertebrate Hsc70
and Hsp70 with unfolded proteins and peptides. In: Morimoto RI,
Tissieres A, Georgopoulos C. editors. The biology of heat shock
proteins and molecular chaperones. New York: Cold Spring Harbor
Laboratory Press. p 179–208.
Hohfeld J, Minami Y, Hartl FU. 1995. Hip, a novel cochaperone
involved in the eukaryotic Hsc70/Hsp40 reaction cycle. Cell 83:589–
598
Karunanithi S, Barclay JW, Robertson RM, Brown IR, Atwood HL.
1999. Neuroprotection at Drosophila synapses conferred by prior heat
shock. J Neurosci 19:4360–4369.
Karunanithi S, Barclay JW, Brown IR, Robertson RM, Atwood HL.
2002. Enhancement of presynaptic performance in transgenic Drosoph-
ila overexpressing heat shock protein Hsp70. Synapse 44:8–14.
Kelty JD, Noseworthy PA, Feder ME, Robertson RM, Ramirez JM.
2002. Thermal preconditioning and heat-shock protein 72 preserve
synaptic transmission during thermal stress. J Neurosci 22:RC193.
Kretz O, Fester L, Wehrenberg U, Zhou L, Brauckmann S, Zhao S,
Prange-Kiel J, Naumann T, Jarry H, Frotscher M, Rune GM. 2004.
408 Chen and Brown
Journal of Neuroscience Research DOI 10.1002/jnr

Hippocampal synapses depend on hippocampal estrogen synthesis.
J Neurosci 24:5913–5921.
Lepock JR. 2005. How do cells respond to their thermal environment?
Int J Hyperthermia 21:681–687.
Leung SM, Senisterra G, Ritchie KP, Sadis SE, Lepock JR, Hightower
LE. 1996. Thermal activation of the bovine Hsc70 molecular chaperone
at physiological temperatures: physical evidence of a molecular ther-
mometer. Cell Stress Chaperones 1:78–89.
Lyashko VN, Vikulova VK, Chernicov VG, Ivanov VI, Ulmasov KA,
Zatsepina OG, Evgen’ev MB. 1994. Comparison of the heat shock
response in ethnically and ecologically different human populations.
Proc Natl Acad Sci USA 91:12492–12495.
Manzerra P, Brown IR. 1992. Distribution of constitutive- and hyper-
thermia-inducible heat shock mRNA species (hsp70) in the Purkinje
layer of the rabbit cerebellum. Neurochem Res 17:559–564.
Manzerra P, Brown IR. 1996. The neuronal stress response: nuclear
translocation of heat shock proteins as an indicator of hyperthermic
stress. Exp Cell Res 229:35–47.
Manzerra P, Rush SJ, Brown IR. 1993. Temporal and spatial distribution
of heat shock mRNA and protein (hsp70) in the rabbit cerebellum in
response to hyperthermia. J Neurosci Res 36:480–490.
Manzerra P, Rush SJ, Brown IR. 1997. Tissue-specific differences in
heat shock protein hsc70 and hsp70 in the control and hyperthermic
rabbit. J Cell Physiol 170:130–137.
Marcuccilli CJ, Mathur SK, Morimoto RI, Miller RJ. 1996. Regulatory
differences in the stress response of hippocampal neurons and glial cells
after heat shock. J Neurosci 16:478–485.
Mattson MP, Magnus T. 2006. Ageing and neuronal vulnerability. Nat
Rev Neurosci 7:278–294.
Mayer J, Brown IR. 1994. Heat shock proteins in the nervous system.
London: Academic. 297 p.
McMahon HT, Bolshakov VY, Janz R, Hammer RE, Siegelbaum SA, Sud-
hof TC. 1996. Synaptophysin, a major synaptic vesicle protein, is not essen-
tial for neurotransmitter release. Proc Natl Acad Sci USA 93:4760–4764.
Morimoto RI, Kline MP, Bimston DN, Cotto JJ. 1997. The heat-shock
response: regulation and function of heat-shock proteins and molecular
chaperones. Essays Biochem 32:17–29.
Mullen RJ, Buck CR, Smith AM. 1992. NeuN, a neuronal specific nu-
clear protein in vertebrates. Development 116:201–211.
Rordorf G, Koroshetz WJ, Bonventre JV. 1991. Heat shock protects cul-
tured neurons from glutamate toxicity. Neuron 7:1043–1051.
Tytell M, Barbe MF, Brown IR. 1994. Induction of heat shock (stress)
protein 70 and its mRNA in the normal and light-damaged rat retina
after whole body hyperthermia. J Neurosci Res 38:19–31.
Ulmasov KA, Shammakov S, Karaev K, Evgen’ev MB. 1992. Heat shock
proteins and thermoresistance in lizards. Proc Natl Acad Sci USA
89:1666–1670.
Walsh DA, Klein NW, Hightower LE, Edwards MJ. 1987. Heat shock
and thermotolerance during early rat embryo development. Teratology
36:181–191.
Walsh DA, Li K, Speirs J, Crowther CE, Edwards MJ. 1989. Regulation
of the inducible heat shock 71 genes in early neural development of
cultured rat embryos. Teratology 40:321–334.
Yenari MA. 2002. Heat shock proteins and neuroprotection. Adv Exp
Med Biol 513:281–299.
Zinchuk V, Zinchuk O, Okada T. 2005. Experimental LPS-induced cholesta-
sis alters subcellular distribution and affects colocalization of Mrp2 and Bsep
proteins: a quantitative colocalization study. Microsc Res Tech 67:65–70.
Translocation of Hsc70 to Synaptic Areas 409
Journal of Neuroscience Research DOI 10.1002/jnr