
Translational control of the synthesis of the Euglena light harvesting chlorophyll a/b binding...
11
Plant Science, 85 (1992) 79-89 79 Elsevier Scientific Publishers Ireland Ltd. Translational control of the synthesis of the Euglena light harvesting chlorophyll a/b binding protein of photosystem II Ram Kishore and Steven D. Schwartzbach School of Biological Sciences, University of Nebraska, Lincoln, NE 68588 (USA) (Received March 23rd, 1992; revision received May 15th, 1992; accepted May 19th, 1992) Synthesis of the precursor to the light harvesting chlorophyll a/b binding protein of photosystem II (LHCPII) increases 50-100-fold upon exposure of dark grown resting Euglena to light. LHCPII mRNA levels increase approximately two-fold upon light exposure indicating that LHCPII synthesis is controlled at the translational level. Approximately 68% of cellular LHCPII mRNA was associated with polysomes in dark grown resting cells. Light exposure had little effect on the relative amount of LHCPII polysomes indicating that light regulates pLHCPII synthesis at the level of polypeptide chain elongation. LHCPII synthesis is undetectable in the bleached Euglena mutants W3BUL and W10BSmL which lack detectable protochlorophyll(ide), the chloroplast photoreceptor and most if not all of the chloroplast genome. Over 60% of LHCPII mRNA was associated with polysomes in bleached mutants. LHCPII polysomes were membrane bound in wild type Euglena and bleached mutants maintained in the dark or the light. Photocon- trol of Euglena LHCPII synthesis is probably mediated through the synthesis of a chloroplast generated signal which is required for release of signal recognition particle dependent arrest of LHCPII mRNA translation by membrane-bound polysomes. Key words: elongation; Euglena; LHCPII; photoinduction; polysome; translation Introduction Exposure of dark grown resting Euglena to light induces the development of the proplastid into a fully functional chloroplast (reviewed in Ref. 1). The accumulation of proteins within the develop- ing chloroplast is the result of an increase in their rate of synthesis rather than a decrease in their rate of degradation [2]. A comparison by two-dimen- sional gel electrophoresis of the most abundant in Correspondence to: Dr. Steven D. Schwartzbach, School of Biological Sciences, 303 Lyman Hall, University of Nebraska, Lincoln, NE 68588-0343, USA. Abbreviations: DEPC, diethylpyrocarbonate; EDTA, ethylenediaminetetraacetic acid; Kb, kilobase; LHCPII, the light harvesting chlorophyll a/b-binding protein of photosystem II; pLHCPII, the precursor to the light harvesting chlorophyll a/b-binding protein of photosystem II; lYrE, polyoxyethylene 10-tridecyl ether; SDS~ sodium dodecyl sulfate; SSC, contains i 50 mM NaCI, 15 mM sodium citrate (pH 7.0); SSPE, contains 150 mM NaCI, 1 mM EDTA, 10 mM Na2HPO4 (pH 7.4). vivo and in vitro Euglena translation products indicates that translation rather than transcription is the major site of photoregulation [2,3]. LHCPII is an abundant cytoplasmically syn- thesized thylakoid localized protein [4]. Exposure of dark-grown resting Euglena to light increases the rate of pLHCPII synthesis 50-100-fold [5] while steady state LHCPII mRNA levels only in- crease twofold [6,7]. Bleached Euglena mutants lacking most if not all of the chloroplast genome contain normal levels of LHCPII mRNA but LHCPII synthesis is undetectable [7,8]. Euglena pLHCPII is a polyprotein which is post- translationally processed into 8-10 copies of LHCPII [9,10]. The half-life of pLHCPII is ap- prox. 20 min in the light or the dark [5,10]. The long half-life of pLHCPII allows direct measurements of the rate of pLHCPII synthesis while avoiding complications arising from rapid degradation of LHCPII in the absence of chlorophyll synthesis and insertion into the 0168-9452/92/$05.00 © 1992 Elsevier Scientific Publishers Ireland Ltd. Printed and Published in Ireland
-
Upload
ram-kishore -
Category
Documents
-
view
212 -
download
0
Transcript of Translational control of the synthesis of the Euglena light harvesting chlorophyll a/b binding...

Plant Science, 85 (1992) 79-89 79 Elsevier Scientific Publishers Ireland Ltd.
Translational control of the synthesis of the Euglena light harvesting chlorophyll a/b binding protein of photosystem II
Ram Kishore and Steven D. Schwartzbach
School of Biological Sciences, University of Nebraska, Lincoln, NE 68588 (USA)
(Received March 23rd, 1992; revision received May 15th, 1992; accepted May 19th, 1992)
Synthesis of the precursor to the light harvesting chlorophyll a/b binding protein of photosystem II (LHCPII) increases 50-100-fold upon exposure of dark grown resting Euglena to light. LHCPII mRNA levels increase approximately two-fold upon light exposure indicating that LHCPII synthesis is controlled at the translational level. Approximately 68% of cellular LHCPII mRNA was associated with polysomes in dark grown resting cells. Light exposure had little effect on the relative amount of LHCPII polysomes indicating that light regulates pLHCPII synthesis at the level of polypeptide chain elongation. LHCPII synthesis is undetectable in the bleached Euglena mutants W3BUL and W10BSmL which lack detectable protochlorophyll(ide), the chloroplast photoreceptor and most if not all of the chloroplast genome. Over 60% of LHCPII mRNA was associated with polysomes in bleached mutants. LHCPII polysomes were membrane bound in wild type Euglena and bleached mutants maintained in the dark or the light. Photocon- trol of Euglena LHCPII synthesis is probably mediated through the synthesis of a chloroplast generated signal which is required for release of signal recognition particle dependent arrest of LHCPII mRNA translation by membrane-bound polysomes.
Key words: elongation; Euglena; LHCPII; photoinduction; polysome; translation
Introduction
Exposure of dark grown resting Euglena to light induces the development of the proplastid into a fully functional chloroplast (reviewed in Ref. 1). The accumulation of proteins within the develop- ing chloroplast is the result of an increase in their rate of synthesis rather than a decrease in their rate of degradation [2]. A comparison by two-dimen- sional gel electrophoresis of the most abundant in
Correspondence to: Dr. Steven D. Schwartzbach, School of Biological Sciences, 303 Lyman Hall, University of Nebraska, Lincoln, NE 68588-0343, USA. Abbreviations: DEPC, diethylpyrocarbonate; EDTA, ethylenediaminetetraacetic acid; Kb, kilobase; LHCPII, the light harvesting chlorophyll a/b-binding protein of photosystem II; pLHCPII, the precursor to the light harvesting chlorophyll a/b-binding protein of photosystem II; lYrE, polyoxyethylene 10-tridecyl ether; SDS~ sodium dodecyl sulfate; SSC, contains i 50 mM NaCI, 15 mM sodium citrate (pH 7.0); SSPE, contains 150 mM NaCI, 1 mM EDTA, 10 mM Na2HPO 4 (pH 7.4).
vivo and in vitro Euglena translation products indicates that translation rather than transcription is the major site of photoregulation [2,3].
LHCPII is an abundant cytoplasmically syn- thesized thylakoid localized protein [4]. Exposure of dark-grown resting Euglena to light increases the rate of pLHCPII synthesis 50-100-fold [5] while steady state LHCPII mRNA levels only in- crease twofold [6,7]. Bleached Euglena mutants lacking most if not all of the chloroplast genome contain normal levels of LHCPII mRNA but LHCPII synthesis is undetectable [7,8]. Euglena pLHCPII is a polyprotein which is post- translationally processed into 8-10 copies of LHCPII [9,10]. The half-life of pLHCPII is ap- prox. 20 min in the light or the dark [5,10]. The long half-life of pLHCPII allows direct measurements of the rate of pLHCPII synthesis while avoiding complications arising from rapid degradation of LHCPII in the absence of chlorophyll synthesis and insertion into the
0168-9452/92/$05.00 © 1992 Elsevier Scientific Publishers Ireland Ltd. Printed and Published in Ireland

80
thylakoid membrane [11,12]. The lack of correla- tion between LHCPII mRNA levels and pLHCPII synthesis rate in wild type and bleached Euglena indicates that LHCPII is one of the light-induced Euglena proteins whose synthesis is controlled at the translational rather than the transcriptional level.
Changes in the amount of LHCPII mRNA associated with ribosomes and the size of LHCPII polysomes can allow one to distinguish photocon- trol of polypeptide chain initiation from photo- control of polypeptide chain elongation [13]. If pLHCPII synthesis is controlled at the level of polypeptide chain initiation, the amount of ribosome-associated LHCPII mRNA and size of LHCPII polysomes would increase upon light exposure [13]. If on the other hand pLHCPII syn- thesis is controlled at the level of polypeptide chain elongation, the amount of ribosome- associated LHCPII mRNA and size of LHCPII polysomes would be unaltered by light exposure [13]. This paper shows that LHCPII mRNA is translated on membrane-bound polysomes. Ex- posure of dark grown resting Euglena to light had little effect on the amount and size of LHCPII polysomes. Over 60% of the LHCPII mRNA was associated with polysomes in bleached mutants even though pLHCPII synthesis is undetectable in these mutants [8]. The increased rates of pLHCPII synthesis in the absence of an increase in the amount and size of LHCPII polysomes indicates that light regulates pLHCPII synthesis at the level of polypeptide chain elongation. A chloroplast generated signal appears to be required for the release of signal recognition particle dependent ar- rest of LHCPII mRNA translation on membrane- bound polysomes.
Materials and Methods
Euglena gracilis Klebs var. bacillaris Cori main- tained in the dark in our laboratory for many years was used throughout this work. The bleached mutants W3BUL and WI0BSmL derived from this strain lack detectable protochlorophyll(ide) and most if not all of their chloroplast genome [1]. Conditions for cell growth, preparation of resting cells and light-induced chloroplast development have been described [5,141.
Polysome isolation and mRNA extraction Polysomes were prepared by a modification of
the procedure of Freyssinet and Schiff [15]. Cyclo- heximide (100 tzg/ml) was added to polysome isola- tion buffers to prevent polysome runoff during isolation and sucrose gradient centrifugation. Preliminary experiments indicated that cyclo- heximide addition did not artifactually increase polysome levels. All buffers and glassware were treated with DEPC and/or autoclaved to eliminate RNase activity. RNase free chemicals were used when available. All steps for polysome isolation were performed at 0-4°C
Resting cells (150 ml, 2 × 106 cells/ml) were harvested by centrifugation for 4 min at 1000 x g. Cell pellets were washed by resuspension in 30 ml of 27% (w/v) sucrose prepared in polysome isola- tion buffer (containing 10 mM Tris-HC1 pH 7.5, 30 mM KCI, 2 mM magnesium acetate and I00 ~g/ml cycloheximde). The washed cells were resus- pended in 4.5 ml of 27% (w/v) sucrose prepared in polysome isolation buffer. Cells were broken by passage through a French pressure cell at 2000 lb/in. 2, the pressate was collected into 6.0 ml polysome isolation buffer and unbroken cells were removed by centrifugation for 5 min at 1300 x g yielding the cell-free extract. To release membrane-bound polysomes [16], PTE (final con- centration 2%) and proteinase K (final concentra- tion 5 mg/ml) were added to the cell-free extract, the extract was incubated for 10 min and centrifug- ed for 10 min at 15 000 x g. Aliquots (1 ml) of the 15 000 x g supernatant fraction were layered onto 10 ml 15-50% (w/v) linear sucrose gradients prepared in polysome isolation buffer. Polysomes were separated by centrifugation for 2 h at 37 000 rev./min in a Beckman SW41 rotor. Eleven 1-ml fractions were collected using an Isco 740 gradient fractionator connected to an Isco UA-5 absor- bance monitor. Room temperature extraction buffer (1 ml) (containing 50 mM Tris-HCl pH 8.0, 50 mM EDTA and 1% SDS) was immediately added to each fraction [17] and RNA was depro- teinized by extraction with water-saturated phenol/chloroform (1:1, v/v). RNA from each sucrose gradient fraction was recovered by ethanol precipitation, the RNA pellets were dissolved in 2 /~1 of DEPC-treated water and stored at -70°C. For EDTA dissociation of ribosomes, EDTA (50

mM final concentration) was added at the time of PTE and proteinase K addition to the 1300 x g cell free extract.
Measurement of RNA levels by Northern and dot blotting
Northern blots were prepared by denaturing ali- quots of RNA from each fraction with formalde- hyde, subjecting the RNA to electrophoresis on 1.2% agarose/2.2 M formaldehyde gels and blot- ting the gels to supported nitrocellulose mem- branes (BA-5; Schleicher and Schull) as described previously [7,9]. Dot blots were prepared by denaturing aliquots of RNA from each fraction with formaldehyde and applying the RNA to sup- ported nitrocellulose using a dot blot apparatus (Bio-Rad) as described [18]. LHCPII mRNA levels were determined by hybridization to a 0.7- Kb exon from a Euglena LHCPII genomic clone [9]. Small subunit rRNA levels were determined by hybridization to a 1.6-Kb BamHI-SalI frag- ment from a Euglena 20S rRNA genomic clone [19]. Probes were labeled with [32p]CTP using a random primed DNA labeling kit (Boehringer, Mannheim).
Blots were prehybridized for 2-4 h at 42°C in hybridization solution (containing 50% (v/v) for- mamide, 0.1% (w/v) polyvinyl pyrrolidone, 0.1% (w/v) ficoll, 0.1% (w/v) bovine serum albumin, 5 x SSPE, 0.2% SDS, 200 #g/ml denatured salmon sperm DNA) and hybridized for 18 h at 42°C in fresh hybridization solution containing 32p-labeled probe. Blots were washed once for 15 min at 45°C with 4 x SSC, 0.2% (w/v) SDS, for 30 min at 50°C with 2 x SSC, 0.1% (w/v) SDS, once for 30 min at 50°C with 1 × SSC, 0.1% (w/v) SDS and once for 40 min at 63°C with 0.1 x SSC and 0.1% (w/v) SDS. Autoradiography was per- formed at -70°C using Kodak X-Omat AR film and two intensifying screens. Quantitative analysis of dot blots was performed using an Ambis radioanalytical imaging system using software ver- sion 2.0,
Results
The distribution of LHCPII mRNA and rRNA on polysome gradients loaded with extracts from cells exposed to light is shown in Fig. 1. Fraction
81
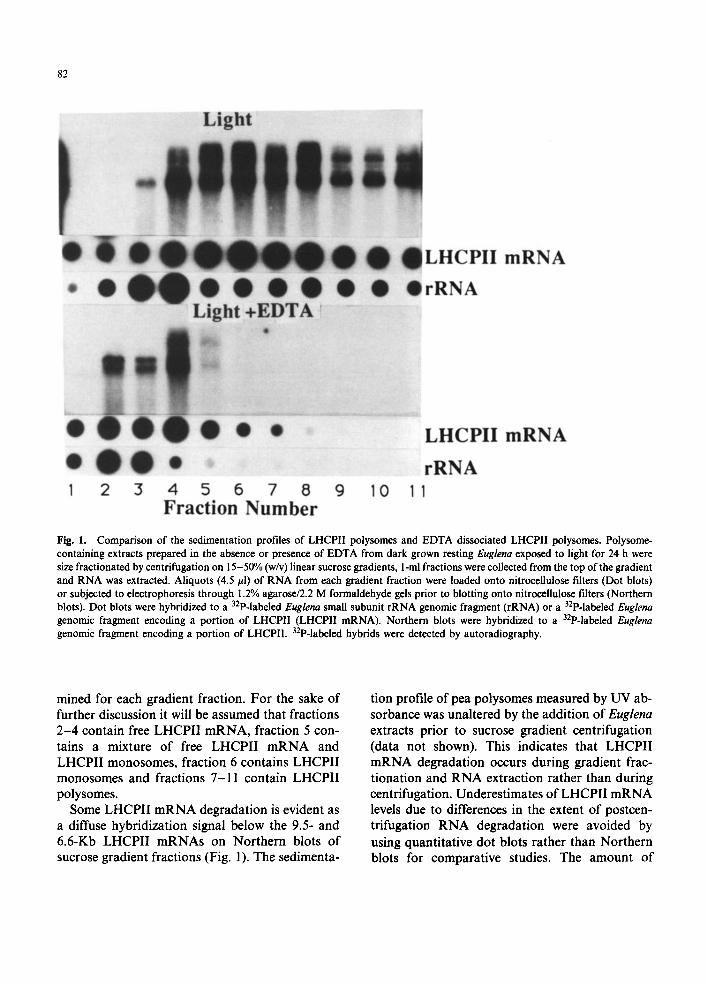
4 contains the bulk of UV absorbing material (data not shown) and the largest amount of small subunit rRNA (Fig. 1). The Mg 2+ chelator EDTA dissociates ribosomes into subunits releasing polysome-bound mRNAs while having no effect on the sedimentation of free LHCPII mRNA. EDTA dissociation of ribosomes decreases the amount of small subunit rRNA associated with material sedimenting to sucrose gradient fraction 4 and increases the amount associated with material sedimenting to fractions 2-3 which are closer to the top of the gradient (Fig. 1). The decrease in the amount of rRNA in fraction 4 after EDTA treat- ment identifies fraction 4 as the 89S monosomal region, fractions 2-3 as the ribosomal subunit region and fractions 5-11 as the polysomal region of the sucrose gradient.
The distribution of LHCPII mRNA on polysome gradients differs significantly from the small subunit rRNA distribution. The greatest amount of 6.6- and 9.5-Kb LHCPII mRNA is recovered in fraction 6 with additional material present in faster sedimenting regions (Fig. 1). Lit- tle LHCPII mRNA is recovered in the ribosomal subunit or 89S monosome region of the sucrose gradient (Fig. 1). Due to the large size of the LHCPII mRNAs, 6.6 and 9.5 Kb [10], LHCPII polysomes could contain over 40 ribosomes and pellet to the bottom of the sucrose gradient. An LHCPII mRNA-containing pellet was not found (data not shown). An analysis of LHCPII mRNA distribution in the sucrose gradient fractions thus provides an accurate representation of the size distribution of LHCPII polysomes.
Dissociation of ribosomes with EDTA decreases the amount of LHCPII mRNA in fractions 6-11 and increases the amount of 6.6- and 9.5-Kb LHCPII mRNA in fraction 4. Intact 6.6- and 9.5- Kb LHCPII mRNA are recovered in gradient frac- tion 4 indicating that the decreased sedimentation of LHCPII mRNA results from EDTA dependent release of LHCPII mRNA from polysomes rather than from activation of a latent RNase activity. Based on the EDTA dependent shift in LHCPII mRNA distribution, it appears that free LHCPII mRNA cosediments with 89S monosomes and ribosomal subunits. LHCPII mRNA bound to ribosomes is recovered in fractions 6-11. The ac- tual number of bound ribosomes cannot be deter-
82
Light
O
O
O i 2 :3
LHCPII mRNA
rRNA
LHCPII m R N A
• rRNA 4 5 6 7 8 9 I0 II Fraction Number
Fig. 1. Comparison of the sedimentation profiles of LHCPII polysomes and EDTA dissociated LHCPII polysomes. Polysome- containing extracts prepared in the absence or presence of EDTA from dark grown resting Euglena exposed to light for 24 h were size fractionated by centrifugation on 15-50% (w/v) linear sucrose gradients, l-ml fractions were collected from the top of the gradient and RNA was extracted. Aliquots (4.5/zl) of RNA from each gradient fraction were loaded onto nitrocellulose filters (Dot blots) or subjected to electrophoresis through 1.2% agarose/2.2 M formaldehyde gels prior to blotting onto nitrocellulose filters (Northern blots). Dot blots were hybridized to a 32p-labeled Euglena small subunit rRNA genomic fragment (rRNA) or a 32p-labeled Euglena genomic fragment encoding a portion of LHCPII (LHCPII mRNA). Northern blots were hybridized to a 32p-labeled Euglena genomic fragment encoding a portion of LHCPII. 32p-labeled hybrids were detected by autoradiography.
mined for each gradient fraction. For the sake o f further discussion it will be assumed that fractions 2 - 4 contain free L H C P I I m R N A , fraction 5 con- tains a mixture o f free L H C P I I m R N A and L H C P I I monosomes, fraction 6 contains L H C P I I monosomes and fractions 7-11 contain L H C P I I polysomes.
Some L H C P I I m R N A degradat ion is evident as a diffuse hybridizat ion signal below the 9.5- and 6.6-Kb L H C P I I m R N A s on Nor thern blots o f sucrose gradient fractions (Fig. 1). The sedimenta-
tion profile o f pea polysomes measured by UV ab- sorbance was unaltered by the addit ion o f Euglena extracts prior to sucrose gradient centrifugation (data not shown). This indicates that L H C P I I m R N A degradat ion occurs during gradient frac- t ionation and R N A extraction rather than during centrifugation. Underestimates o f L H C P I I m R N A levels due to differences in the extent o f postcen- trifugation R N A degradat ion were avoided by using quantitative dot blots rather than Nor thern blots for comparat ive studies. The amount o f

83
0
n"
0 I-- b
0
(U ._o "O
0 I--
0
40
20
40
20
rRNA Dark
o ~ o • • • ,
Light
• O 0 • • • • • . •
0 ¸
.2
0 I--
0
20
10
• •
LHCPII mRNA Dark
• • • • • • • • •
0 ~ L i g h t
• • o O O O O 0 • .g
o ~ 10
0 1 2 3 4 5 6 7 8 9 10 11 1 2 3 4 5 6 7 8 9 10 11
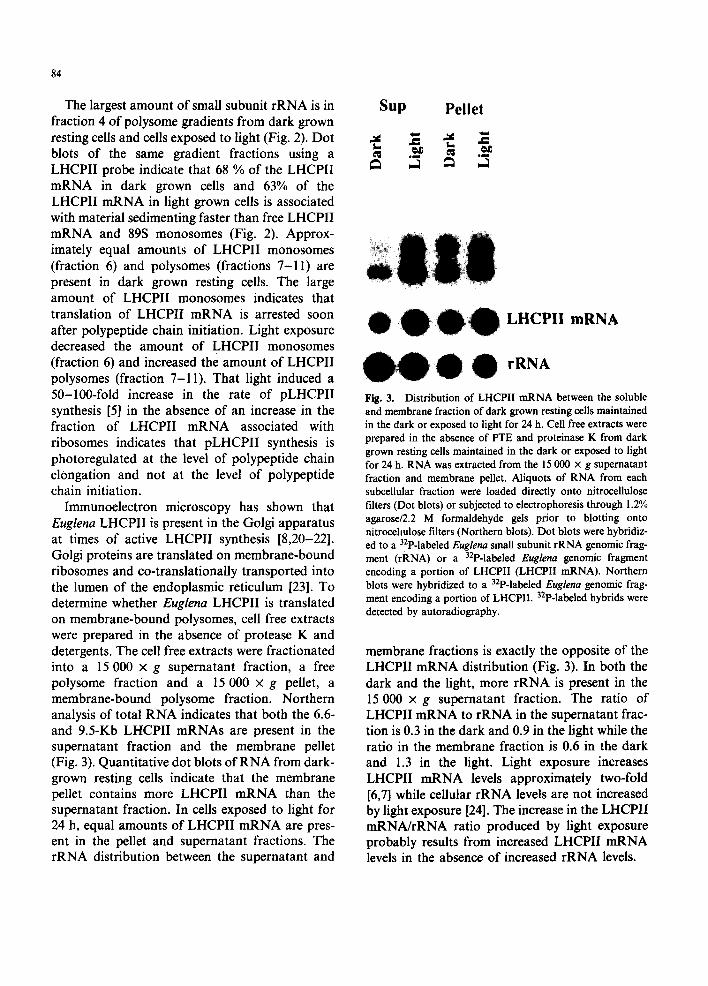
Fract ion N u m b e r Fract ion Number Fig. 2. Sedimentation analysis of LHCPII polysomes in dark grown resting Euglena and in cells exposed to light for 24 h. Polysome containing extracts prepared from dark grown resting Euglena maintained in the dark or exposed to light for 24 h were size frac- donated by ccntrifugation on 15-50% (w/v) linear sucrose gradients, l-ml fractions were collected from the top of the gradient and RNA was extracted. Dot blots were prepared by loading 4.5-/~1 aliquots of RNA from each gradient fraction onto nitrocellulose filters. Dot blots were hybridized to a 32P-labeled Euglena small subunit rRNA genomic fragment (rRNA) or a 32p-labeled Euglena genomic fragment encoding a portion of LHCPII (LHCPII mRNA) and hybrids were detected by autoradiography. Quantitative analysis of the dot blots was performed using an Ambis radioanalytical imaging system. The amount of 32p-labeled rRNA probe or 32p-labeled LHCPII probe hybridized to each gradient fraction is plotted as a percentage of the total 32p-labeled rRNA probe or 32p-labeled LHCPII probe hybridized across the gradient.
LHCPII mRNA in each gradient fraction was nor- malized to the total amount of LHCPII mRNA recovered from the gradient in order to correct for variations in the amount of LHCPII mRNA actu-
ally loaded. Dot blots of rRNA provided an inter- nal sedimentation standard for localization of free LHCPII mRNA, LHCPII monosomes and LHCPII polysomes.
84
The largest amount of small subunit rRNA is in fraction 4 of polysome gradients from dark grown resting cells and cells exposed to light (Fig. 2). Dot blots of the same gradient fractions using a LHCPII probe indicate that 68 % of the LHCPII mRNA in dark grown cells and 63% of the LHCPII mRNA in light grown cells is associated with material sedimenting faster than free LHCPII mRNA and 89S monosomes (Fig. 2). Approx- imately equal amounts of LHCPII monosomes (fraction 6) and polysomes (fractions 7-11) are present in dark grown resting cells. The large amount of LHCPII monosomes indicates that translation of LHCPII mRNA is arrested soon after polypeptide chain initiation. Light exposure decreased the amount of LHCPII monosomes (fraction 6) and increased the amount of LHCPII polysomes (fraction 7-11). That light induced a 50-100-fold increase in the rate of pLHCPII synthesis [5] in the absence of an increase in the fraction of LHCPII mRNA associated with ribosomes indicates that pLHCPII synthesis is photoregulated at the level of polypeptide chain elongation and not at the level of polypeptide chain initiation.
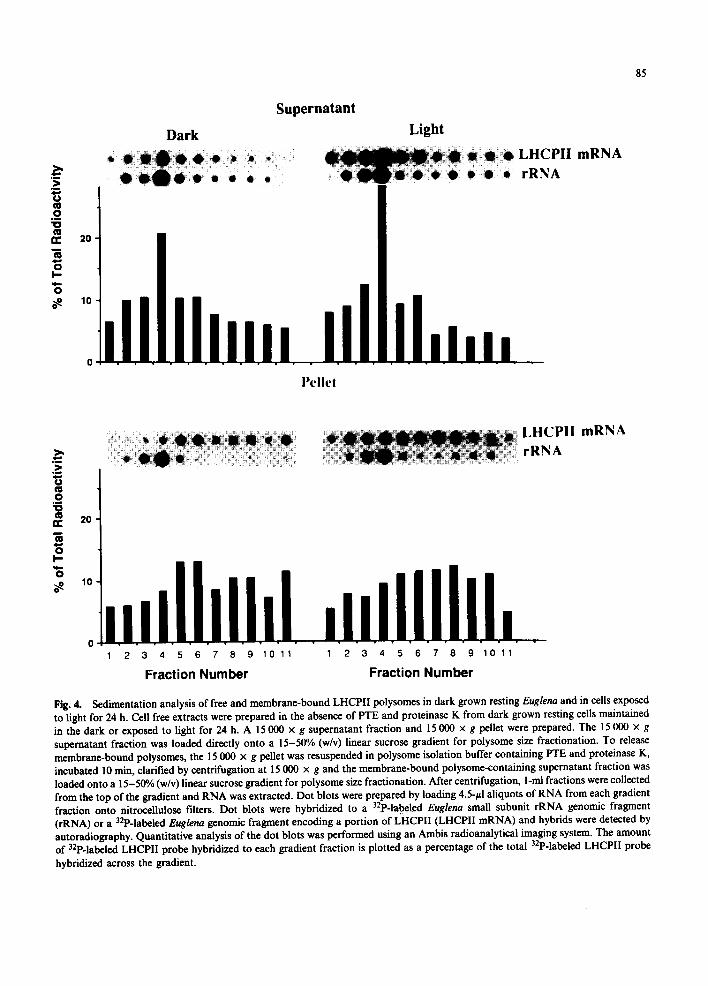
Immunoelectron microscopy has shown that Euglena LHCPII is present in the Golgi apparatus at times of active LHCPII synthesis [8,20-22]. Golgi proteins are translated on membrane-bound ribosomes and co-translationally transported into the lumen of the endoplasmic reticulum [23]. To determine whether Euglena LHCPII is translated on membrane-bound polysomes, cell free extracts were prepared in the absence of protease K and detergents. The cell free extracts were fractionated into a 15 000 x g supernatant fraction, a free polysome fraction and a 15 000 × g pellet, a membrane-bound polysome fraction. Northern analysis of total RNA indicates that both the 6.6- and 9.5-Kb LHCPII mRNAs are present in the supernatant fraction and the membrane pellet (Fig. 3). Quantitative dot blots of RNA from dark- grown resting cells indicate that the membrane pellet contains more LHCPII mRNA than the supernatant fraction. In cells exposed to light for 24 h, equal amounts of LHCPII mRNA are pres- ent in the pellet and supernatant fractions. The rRNA distribution between the supernatant and
Sup Pellet
e l m l o m
LHCPII m R N A
rRNA
Fig. 3. Distribution of LHCPII mRNA between the soluble and membrane fraction of dark grown resting cells maintained in the dark or exposed to light for 24 h. Cell free extracts were prepared in the absence of PTE and proteinase K from dark grown resting cells maintained in the dark or exposed to light for 24 h. RNA was extracted from the 15 000 x g supernatant fraction and membrane pellet. Aliquots of RNA from each subceUular fraction were loaded directly onto nitrocellulose filters (Dot blots) or subjected to electrophoresis through 1.2% agarose/2.2 M formaldehyde gels prior to blotting onto nitrocellulose filters (Northern blots). Dot blots were hybridiz- ed to a 32p-labeled Euglena small subunit rRNA genomic frag- ment (rRNA) or a 32p-labeled Euglena genomic fragment encoding a portion of LHCPII (LHCPII mRNA). Northern blots were hybridized to a 32p-labeled Euglena genomic frag- ment encoding a portion of LHCPII. 32P-labded hybrids were detected by autoradiography.
membrane fractions is exactly the opposite of the LHCPII mRNA distribution (Fig. 3). In both the dark and the light, more rRNA is present in the 15 000 x g supernatant fraction. The ratio of LHCPII mRNA to rRNA in the supernatant frac- tion is 0.3 in the dark and 0.9 in the light while the ratio in the membrane fraction is 0.6 in the dark and 1.3 in the light. Light exposure increases LHCPII mRNA levels approximately two-fold [6,7] while cellular rRNA levels are not increased by light exposure [24]. The increase in the LHCPII mRNA/rRNA ratio produced by light exposure probably results from increased LHCPII mRNA levels in the absence of increased rRNA levels.
85
.>_
m .o_ "o re
o I.-
O
Q
20
10
Supernatant
Dark Light
LHCPII mRNA rRNA
l )cl lct
LHCPll mRNA £ r R N A
¢~ 20
0 1 2 3 4 5 6 7 8 9 10 11 1 2 3 4 5 6 7 8 9 10 11
Fraction Number Fraction Number
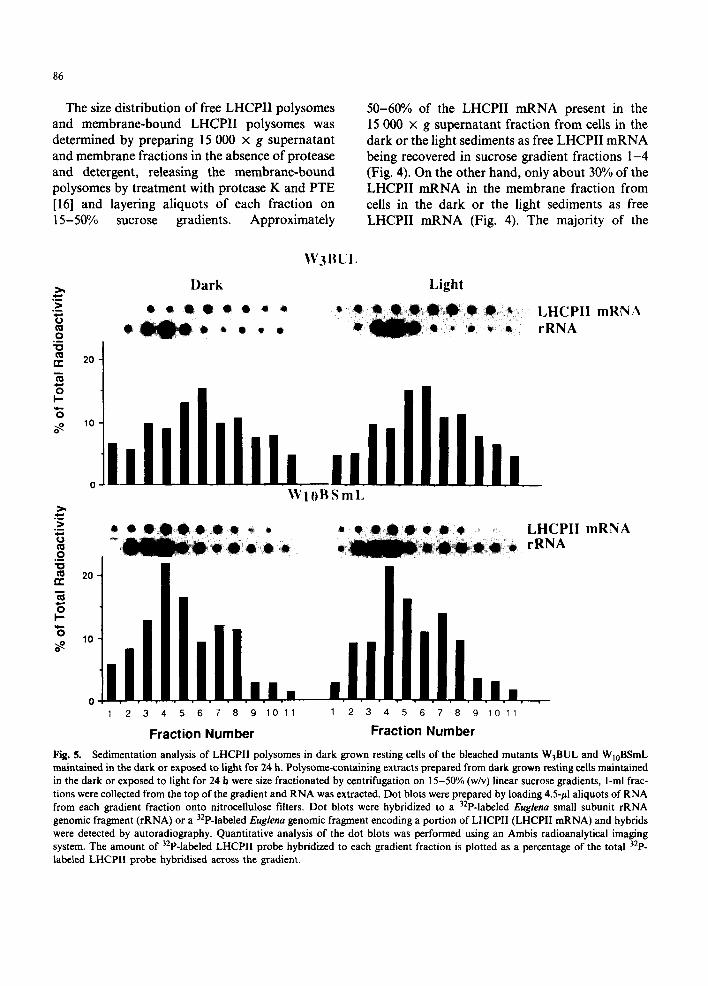
Fig. 4. Sedimentation analysis of free and membrane-bound LHCPII polysomes in dark grown resting Euglena and in cells exposed to light for 24 h. Cell free extracts were prepared in the absence of PTE and proteinase K from dark grown resting cells maintained in the dark or exposed to light for 24 h. A 15 000 x g snpernatant fraction and 15 000 × g pellet were prepared. The 15 000 x g supernatant fraction was loaded directly onto a 15-50% (w/v) linear sucrose gradient for polysome size fractionation. To release membrane-bound polysomes, the 15 000 x g pellet was resuspended in polysome isolation buffer containing PTE and proteinase K, incubated 10 rain, clarified by centrifugation at 15 000 x g and the membrane-bound polysome-containing supernatant fraction was loaded onto a 15-50% (w/v) linear sucrose gradient for polysome size fractionation. After centrifugation, l-m1 fractions were collected from the top of the gradient and RNA was extracted. Dot blots were prepared by loading 4.5-pl aliquots of RNA from each gradient fraction onto nitrocellulose filters. Dot blots were hybridized to a 32p-labeled Euglena small subunit rRNA genomic fragment (rRNA) or a 32p-labeled Euglena gcnomic fragment encoding a portion of LHCPII (LHCPII mRNA) and hybrids were detected by autoradiography. Quantitative analysis of the dot blots was performed using an Ambis radioanalyticai imaging system. The amount of 32p-labeled LHCPII probe hybridized to each gradient fraction is plotted as a percentage of the total 32p-labeled LHCPII probe
hybridized across the gradient.
86
The size distribution of free LHCPII polysomes and membrane-bound LHCPII polysomes was determined by preparing 15 000 x g supernatant and membrane fractions in the absence of protease and detergent, releasing the membrane-bound polysomes by treatment with protease K and PTE [16] and layering aliquots of each fraction on 15-50% sucrose gradients. Approximately
50-60% of the LHCPII mRNA present in the 15 000 x g supernatant fraction from cells in the dark or the light sediments as free LHCPII mRNA being recovered in sucrose gradient fractions 1-4 (Fig. 4). On the other hand, only about 30% of the LHCPII mRNA in the membrane fraction from cells in the dark or the light sediments as free LHCPII mRNA (Fig. 4). The majority of the
.>__
0
t~ rr
I.-
"6
20
10
0
W3 B UI,
D a r k
• • • • 4J • • •
h • • • • •
L i g h t
LHCPII mRNA rRNA
>,
._o "o ¢0 ¢w
t~ o I--
O
20
10
W I l) B S ill 1,
LHCPI I mRNA rRNA
0 ~ 1 2 3 4 5 6 7 8 9 10 11 1 2 3 4 5 6 7 8 9 10 11
Fraction Number Fraction Number
Fig. 5. Sedimentation analysis of LHCPII polysomes in dark grown resting cells of the bleached mutants W3BUL and WIoBSmL maintained in the dark or exposed to light for 24 h. Polysome-containing extracts prepared from dark grown resting cells maintained in the dark or exposed to light for 24 h were size fractionated by centrifugation on 15-50% (w/v) linear sucrose gradients, l-mi frac- tions were collected from the top of the gradient and RNA was extracted. Dot blots were prepared by loading 4.5-/zl aliquots of RNA from each gradient fraction onto nitrocellulose filters. Dot blots were hybridized to a 32p-labeled Euglena small subunit rRNA genomic fragment (rRNA) or a 32p-labeled Euglena genomic fragment encoding a portion of LHCPII (LHCPII mRNA) and hybrids were detected by autoradiography. Quantitative analysis of the dot blots was performed using an Ambis radioanalytical imaging system. The amount of 32p-labeled LHCPII probe hybridized to each gradient fraction is plotted as a percentage of the total 32p. labeled LHCPII probe hybridised across the gradient.

LHCPII mRNA released from the membrane pellet is present as LHCPII monosomes (fraction 6) and polysomes (fractions 7-11) in both the dark and light (Fig. 4). The enrichment of LHCPII mRNA (Fig 3) and LHCPII polysomes (Fig. 4) in the membrane fraction indicates that pLHCPII is synthesized on membrane-bound polysomes in Euglena rather than free polysomes as found in higher plants.
The plastidless mutants W3BUL and WIoBSmL have the same amount of LHCPII mRNA as dark grown resting Euglena [7]. pLHCPII synthesis is however undetectable in these mutants [8]. Quan- titative dot blot analysis of sucrose gradient frac- tions shows that in the dark or the light, approximately 60% of the LHCPII mRNA in W3BUL and approx. 40% of the LHCPII mRNA in W10BSmL is associated with LHCPII monosomes and LHCPII polysomes (Fig. 5). The 15 000 x g membrane fraction of W3BUL and W10BSmL was enriched in LHCPII mRNA (data not shown). The association of LHCPII mRNA with monosomes and polysomes in the absence of detectable pLHCPII synthesis provides further evidence for translational control of pLHCPII synthesis at the level of polypeptide chain elonga- tion on membrane-bound polysomes.
Discussion
Light exposure increases the rate of Euglena pLHCPII synthesis 50-100-fold [5] while having little effect on the amount of LHCPII polysomes. pLHCPII synthesis is undetectable in the bleached mutants W3BUL and W10BSmL [8]. The amount of LHCPII polysomes and LHCPII mRNA [7] in these bleached mutants are, however, similar to the levels in wild type cells. Chlorophyll-deficient plants also have reduced or undetectable rates of LHCPII synthesis even though LHCPII mRNA is associated with polysomes I11]. Measurements of LHCPII turnover indicate that LHCPII is rapidly degraded in the absence of pigment binding and membrane insertion [12]. Synthesis of barley chloroplast-encoded, chlorophyll-binding apopro- teins is undetectable in the dark even though nor- mal levels of apoprotein polysomes are present on the thylakoid membrane [25]. Light induces
87
apoprotein synthesis in the absence of increased polysome levels. It appears that apoprotein syn- thesis is undetectable in dark grown plants because the apoproteins are degraded prior to completion of the nascent chain [26]. Light induces chloro- phyll synthesis, the nascent apoprotein is stabil- ized through chlorophyll binding and an increased rate of apoprotein synthesis is seen [26]. Thus for higher plant LHCPII and chloroplast-encoded chlorophyll-binding apoproteins, the lack of cor- relation between polysome levels and apoprotein synthesis rates reflects post-translational control of apoprotein stability rather than translational control of apoprotein synthesis [l 1,12,25,26].
In contrast to pLHCPII of higher plants and other algae, Euglena pLHCPII is a polyprotein which is post-translationally processed into 8-10 LHCPIIs [9,10]. The in vivo half-life of Euglena pLHCPII, 20 rain in the light or the dark [5,10], permits direct measurements of pLHCPII syn- thesis rates avoiding complications caused by rapid degradation in the absence of pigment bind- ing or thylakoid assembly Ill,12]. The light- induced 50-100-fold increase in pLHCPII syn- thesis rate does not result from a decreased rate of pLHCPII degradation [5]. The absence of detec- table pLHCPII synthesis in the bleached mutants WaBUL and WIoBSmL [8] is not the result of rapid degradation in the absence of chloroplasts and chlorophyll synthesis. The absence of a cor- relation between the rate of pLHCPII synthesis and the fraction of LHCPII mRNA associated with polysomes indicates that light regulates pLHCPII synthesis at the level of polypeptide chain elongation. Since a large amount of LHCPII mRNA is associated with monosomes, transla- tional arrest appears to occur soon after polypep- tide chain initiation.
Membrane-bound LHCPII polysomes were found in wild type Euglena and in the bleached mutants. Imuno-electron microscopy has localized Euglena LHCPII in both the Golgi apparatus and chloroplast at times of active pLHCPI! synthesis [8,20-22]. An LHCPII immunoreaction is seen only over the chloroplast in cells not actively syn- thesizing pLHCPII [8,20-22] indicating that pLHCPII is transported through the ER prior to localization within the chloroplast. The 135 amino

88
acid N-terminal extension of the pLHCPII polyprotein contains a domain having the characteristics of a signal sequence (Muchhal and Schwartzbach, unpublished). Similar N-terminal extensions with signal sequence domains are found on the Euglena precursors to the small subunit of ribulose bisphosphate carboxylase [27] and hydroxymethylbilane synthase [28] suggesting that all cytoplasmically synthesized chloroplast localiz- ed proteins are co-translationally transported into the ER prior to chloroplast localization.
Photoinduction of most Euglena chloroplast proteins [2,3] including pLHCPII [5-7], the small subunit of ribulose bisphosphate carboxylase [29] and hydroxymethylbilane synthase [30] is transla- tionally regulated. Perhaps the best characterized example of translational control is the arrest of polypeptide chain elongation resulting from the in- teraction of signal recognition particle and the N- terminal signal sequence of ER targeted proteins [13,31]. Light-mediated release of signal recogni- tion particle dependent translational arrest pro- vides a likely mechanism for translational control of the synthesis of pLHCPII, the small subunit of ribulose bisphosphate carboxylase, hydroxy- methylbilane synthase and other Euglena chloroplast proteins. The binding of signal recognition particle to the N-terminal signal pep- tide of nascent cytoplasmically synthesized chloroplast localized Euglena proteins would arrest polypeptide chain elongation soon after polypeptide chain initiation [31]. Since pLHCPII synthesis is arrested in dark grown cells and bleached mutants lacking chloroplasts even though LHCPII mRNA is associated predomin- antly with membrane-bound polysomes, polypep- tide chain elongation is not resumed upon the binding of the signal recognition factor ribosome complex to the ER membrane. Resumption of polypeptide chain elongation in Euglena appears to require a light-induced effector molecule syn- thesized within the chloroplast. The proposed linkage between co-translational transport of cytoplasmically synthesized chloroplast proteins through the ER membrane aiad the synthesis of a light-induced effector molecule within the chloroplast provides a general mechanism for translational control of cytoplasmically synthesiz- ed chloroplast localized proteins in Euglena.
Acknowledgments
This work was supported by National Science Foundation grant DCB-8817250, National Insti- tute of Health biomedical research support grant RRo07055 and funds from the University of Nebraska Lincoln Research Council. We thank Dr. G. Freyssinet for the gift of a Euglena small subunit rRNA genomic clone, Umesh Muchhal for the Euglena LHCPII genomic clone, Dr. E. Davies for samples of pea polysomes and Tom Babbit for assistance in developing polysome isolation procedures.
References
1 S.D. Schwartzbach, Photocontrol of organdie biogenesis in Euglena. Photochem. Photobiol., 51 (1990) 231-254.
2 A.F. Monroy, S.A. McCarthy and S.D. Schwartzbach, Evidence for translational regulation of chloroplast and mitochondrial biogenesis in Euglena. Plant Sci., 51 (1987) 61-76.
3 S.A. McCarthy and S.D. Schwartzbach, Absence of photoregulation of abundant mRNA levels in Euglena. Plant. Sci. Lett., 35 (1984) 61-66.
4 F.X. Cunningham Jr. and J.A. Schiff, Chlorophyll- protein complexes from Euglena gracilis and mutants de- ficient in chlorophyll b. II. Polypeptide composition. Plant Physiol., 80 (1986) 231-238.
5 A. Rikin and S.D. Schwartzbach, Regulation by light and ethanol of the synthesis of the light harvesting chlorophyll a/b-binding protein of photosystem II in Euglena. Planta, 178 (1989) 76-83.
6 C. Weiss, G. Houln~, M-L. Schantz and R. Schantz, Pho- toinduction of the synthesis of chloroplast membrane pro- teins in Euglena gracilis. J. Plant Physiol., 133 (1988) 521-528.
7 R. Kishore and S.D. Schwartzbach, Photo and nutritional regulation of the light-harvesting chlorophyll a/b-binding protein of photosystem II mRNA levels in Euglena. Plant Physiol., 98 (1992) 808-812.
8 J.A. Schiff, S.D. Schwartzbach, T. Osafune and E. Hase, Photocontrol and processing of LHCPII apoprotein in Euglena: possible role of Golgi and other cytoplasmic sites. J. Photochem. Photobiol. B. Biol., 11 (1991) 219-236.
9 U.K. Muchhal and S.D. Schwartzbach, Characterization of the Euglena gene encoding a polyprotein precusor to the light harvesting chlorophyll a/b-binding protein of photosystem II. Plant Mol. Biol., 18 (1992) 287-299.
l0 A. Rikin and S.D. Schwartzbach, Extremely large and slowly processed precursors to the Euglena light- harvesting chlorophyll a/b binding protein of photo- system II. Proc. Natl. Acad. Sci. USA, 85 (1988) 5117-5121.

11 K. Apel and K. Kloppstech, The effect of light on the biosynthesis of the light-harvesting chlorophyll a/b pro- tein. Evidence for the requirement of chlorophyll a for the stabilization of the apoprotein. Planta, 150 (1980) 426-430.
12 T. Terao and S. Katoh, Synthesis and breakdown of the apoprotein of light-harvesting chlorophyll a/b protein in chlorophyll b-deficient mutants of rice. Plant Cell Physiol., 30 (1989) 571-580.
13 J.W.B. Hershey, Translational control in mammalian cells. Annu. Rev. Biochem., 60 (1991) 717-755.
14 M.A. Horrum and S.D. Schwartzbach, Nutritional regulation of organelle biogenesis in Euglena: Photo and metabolite induction of mitochondria. Planta, 149 (1980) 376-383.
15 G. Freyssinet and J.A. Schiff, The chloroplast and cytoplasmic ribosomes of Euglena. II. Characterization of ribosomal proteins. Plant Physiol., 53 (1974) 543-554.
16 S. Abe and E. Davies, Quantitative isolation ofundegrad- ed polysomes from aged pea tissue in the absence of con- taminants and artifacts. Plant Cell Physiol., 26 0985) 1499-1509.
17 J.O. Berry, D.E. Breiding and D.F. Klessig, Light mediated control of translational initiation of ribulose- 1,5-bisphosphate carboxylase in Amaranth colyledons. Plant Ceil, 2 (1990) 795-803.
18 J. Sambrook, E.F. Fritsch and T. Maniatis, Molecular Cloning: A Laboratory Manual, 2nd edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1989, pp. 7.53-7.55
19 Neyret-Djossou, G. Freyssinet, P. Raval-Chapius and P. Heizmann, Comparison between the organization of nuclear ribosomal DNA unit of Euglena gracilis Z and var. Bacillaris. Plant Mol. Biol., 6 (1986) l l l - l l 7 .
20 T. Osafune, J.A. Schiff and E. Hase, Immunogold localization of LHCPII apoprotein in Golgi of Euglena. Cell Structure and Function, 15 (1990) 99-105.
21 T. Osafune, S. Sumida, J.A. Schiff and E. Hase, Im- munolocalization of LHCP II apoprotein in the Goigi during light induced chloroplast development in non- dividing Euglena cells. J. Electron Microsc., 40 (1991) 41-47.
22 T. Osafune, J.A. Schiff and E. Hase, Stage dependent localization of LHCPII apoprotein in Golgi of syn- chronized cells of Euglena gracilis by immunogold elec- tron microscopy. Exp. Cell Res., 193 (1991) 320-330.
89
23 P. Walter and V.R. Lingappa, Mechanism of protein translocation across the endoplasmic reticulum. Annu. Rev. Cell Biol., 2 (1986) 499-516.
24 D. Cohen and J.A. Schiff, Events surrounding the early development of Euglena chloroplasts. Photoregulation of the transcription of chloroplastic and cytoplasmic ribosomal RNAs. Arch. Biocbem. Biophys. 177 (1976) 201-216.
25 R.R. Klein, H.S. Mason and J.E. Mullet, Light-regulated translation of chloroPlast proteins. I. Transcripts of PsaA-PsaB, PsbA and PbcL are associated with polysomes in dark grown and illuminated barley seed- lings. J. Biol. Chem., 262 (1988) 286-301.
26 J.E. Mullet, P.G. Klein and R.R. Klein, Chlorophyll regulates accumulation of the plastid-encoded chlorophyll apoprotein CP43 and D1 by increasing apoprotein stabili- ty. Proc. Natl. Sci. Acad. USA, 87 (1990) 4038-4042.
27 R.L. Chart, M. Keller, J. Canady, J.H. Weil and P. Im- bault, Eight small subunits of Euglena ribulose-l,5- bisphosphate carboxylase/oxygenase are translated from a large mRNA as a polyprotein. EMBO J. 3 (1990) 333-338.
28 A.L. Sharif, A.G. Smith and G. Abell, Isolation and characterization of a cDNA clone for a chlorophyll syn- thesis enzyme from Euglena gracilis. The chloroplast enzyme hydroxymethylbilane synthase (porphobilinogen deaminase) is synthesized with a very long transit peptide in Euglena. Eur. J. Biol. Chem., 184 (1989) 353-359.
29 M. Keller, R.L. Chan, L.H. Tessier, J.H. Weil and P. Im- bault, Post-transcriptional regulation by light of the biosynthesis of Euglena ribulose-l,5-bisphosphate car- boxylase/oxygenase small subunit. Plant Mol. Biol., 17 (1991) 73-82.
30 L.S. Schashidhara and A.G. Smith, Expression and subcellular location of the tetrapyrrole synthesis enzyme porphobilinogen deaminase in light-grown Euglena gracilis and three nonchlorophyllous cell lines. Proc. Natl. Acad. Sci. USA, 88 (1991) 63-67.
31 P. Walter and G. Blobel, Translocation of proteins across the endoplasmic reticulum. IlL Signal recognition protein (SRP) causes signal sequence-dependent and site-specific arrest of chain elongation that is released by microsomal membranes. J. Cell Biol., 91 (1981) 557-561.





![Developmental Changes of the Photochemical …Chlorophyll fluorescence emanates primarily from chlorophyll a in photosystem II [21] and leaf photosynthetic status, as indicated by](https://static.fdocuments.in/doc/165x107/5f0c18fe7e708231d433baa2/developmental-changes-of-the-photochemical-chlorophyll-fluorescence-emanates-primarily.jpg)












