
Transient motion of visual texture delays saccadic eye movements
12
acta psychologica ELSEVIER Acta Psychologica 92 (1996) 251-262 Transient motion of visual texture delays saccadic eye movements T.J. Crawford * Department of Psychology, Lancaster Unit, ersi~', Lancaster LA I 4YF, UK Received 19 September 1994; revised 21 April 1995; accepted 27 April 1995 Abstract Neurophysiological research has established that a transient stimulus, spatially located outside the receptive field of visual cells, can activate some cells and inhibit firing in others. However, the significance of this phenomenon for behavioural responses in man is unclear. This study investigated the effect of a transient peripheral event on the initiation of saccadic eye movements to a luminance increment. In Experiment 1 human saccadic eye movements to targets that varied in luminance were compared on 'shift' trials, in which the saccadic target was timed to coincide with a step displacement of a vertical grating in the background, and on 'no-shift' trials, when the background remained stationary. The results showed a significant delay in mean saccadic latencies on 'shift trials' compared to 'no-shift' trials. Saccadic latencies were reduced in both conditions with increasing target intensity. Measurement of visual sensitivity showed a small non-significant increase in thresholds in the background 'shift' condition. A second experiment manipulated visuo-temporal information by varying both target duration and intensity. This experiment revealed significant effects of target duration and signal intensity on saccadic latency; and a 3-way interaction showing that saccades suffered the greatest delay in the background 'shift' condition at the lowest duration and intensity targets. These results show that the peripheral motion of visual texture delays the programming of saccadic eye movements and there is a particularly marked effect for targets of low signal strength. PsyclNFO classification: 2300; 2330; 2320; 2346 Keywords: Saccadic latency; Target brightness; Shift-effect; Sustained; Transient; Magnocellular; Parvocellu- lar visual channels; Visual thresholds * Tel.: +44 1524 593761, Fax: +44 1524 593744. 0001-6918/96/$15.00 Copyright © 1996 Elsevier Science B.V. All rights reserved. SSD10001-6918(95)00015-1
-
Upload
tj-crawford -
Category
Documents
-
view
213 -
download
0
Transcript of Transient motion of visual texture delays saccadic eye movements

acta psychologica
ELSEVIER Acta Psychologica 92 (1996) 251-262
Transient motion of visual texture delays saccadic eye movements
T.J. C r a w f o r d *
Department of Psychology, Lancaster Unit, ersi~', Lancaster LA I 4YF, UK
Received 19 September 1994; revised 21 April 1995; accepted 27 April 1995
Abstract
Neurophysiological research has established that a transient stimulus, spatially located outside the receptive field of visual cells, can activate some cells and inhibit firing in others. However, the significance of this phenomenon for behavioural responses in man is unclear. This study investigated the effect of a transient peripheral event on the initiation of saccadic eye movements to a luminance increment. In Experiment 1 human saccadic eye movements to targets that varied in luminance were compared on 'shift' trials, in which the saccadic target was timed to coincide with a step displacement of a vertical grating in the background, and on 'no-shift' trials, when the background remained stationary. The results showed a significant delay in mean saccadic latencies on 'shift trials' compared to 'no-shift' trials. Saccadic latencies were reduced in both conditions with increasing target intensity. Measurement of visual sensitivity showed a small non-significant increase in thresholds in the background 'shift' condition. A second experiment manipulated visuo-temporal information by varying both target duration and intensity. This experiment revealed significant effects of target duration and signal intensity on saccadic latency; and a 3-way interaction showing that saccades suffered the greatest delay in the background 'shift' condition at the lowest duration and intensity targets. These results show that the peripheral motion of visual texture delays the programming of saccadic eye movements and there is a particularly marked effect for targets of low signal strength.
PsyclNFO classification: 2300; 2330; 2320; 2346
Keywords: Saccadic latency; Target brightness; Shift-effect; Sustained; Transient; Magnocellular; Parvocellu- lar visual channels; Visual thresholds
* Tel.: +44 1524 593761, Fax: +44 1524 593744.
0001-6918/96/$15.00 Copyright © 1996 Elsevier Science B.V. All rights reserved. SSD10001-6918(95)00015-1

252 T.J. Cra~ford / Acta Psychologica 92 (1996) 251-262
1. Introduction
Under photopic conditions saccadic eye movements are normally generated within a visual context of pattern and motion signals. The control of saccadic eye movements involves a complex interaction of passive spatial and temporal filters at the early stages of visual processing, and neurocognitive mechanisms at later stages in the selection of an appropriate saccadic programme. Several studies, in recent years, have attempted to explore the neurological mechanisms involved in the filtering of distracting signals from those which require selective attention in the programming of saccadic eye movements (see Guitton et al., 1985; Crawford et al., 1989a,b; Pierrot-Deseilligny et al., 1991; Crawford and MiJller, 1992). However, much of this research has focused on the final motor programming stages, confining investigations to the primary saccades aimed towards a secluded target in the dark. Much of the brain's selective visual processing is therefore, artificially sacrificed on the grounds of experimental rigour (but see e.g. Findlay, 1987; Lawden et al., 1995). Further more ecologically valid studies are required in which saccadic eye movements are examined in a rich visual environment that includes systematic control of visual texture and motion signals.
Mcllwain (1964, 1966) showed that retinal ganglion cells are sensitive to high contrast stimuli occurring well outside classically defined receptive fields, an area in the cat that was typically less than 8 ° in diameter. This sensitivity to remote visual perturbations, termed the periphery or shift-effect, has been observed in retinal ganglion (e.g. Fischer et al., 1975) and lateral geniculate (LGN) (Fischer and Kriiger, 1974) neurones in both cat (Kriiger and Fischer, 1973; Fischer et al., 1975) and monkey (Kriiger et al., 1975). In man, under similar conditions, a form of visual masking is observed: visual sensitivity to a foveal target is reduced in the presence of simultaneous movement of a peripheral pattern surround (Brooks and Impelman, 1981; Derrington, 1984). The primary aim of this study was to investigate the possible significance of these observations for behavioural responses in man.
Two experiments are described that examine the effect of a background displacement on the initiation of visually-guided saccadic eye movements. Saccade latencies to the sudden onset of a visual target were compared in two conditions: in one condition a target was presented within a stationary, i.e. no-shift (NS), but patterned background; in the second condition a step displacement of the background supplied a transient visual input, i.e. shift (S) of the visual surround, that coincided with the onset of the target.
2. Experiment 1
2.1. Methods
Eleven subjects, who were all naive to the purposes of the experiment, served as subjects for the study. All subjects had emmetropic vision.
Fig. l shows the target display used in this study. A high contrast (0.8) rectangular wave grating of spatial frequency 0.2 c / ° was projected on a screen I. 1 metres from the observer. The central region (16°X 6 °) was occluded and illuminated by a second

T.J. Crawford / Acta Psychologica 92 (1996) 251-262 253
11111 • • <~ • •
lllJJ. I I I I I I I I I I 8degs I I I ~ I
I 40.6 deos I
Fig. 1. Stimulus display showing the central LEDs targets and surrounding background display. The LEDs targets were only visible when illuminated.
projector system in order avoid complex visual interactions in the central and back- ground visual field (see e.g. Davidson et al., 1992). The mean luminance of the central field was equal to the space average luminance of the grating (10.8 c d / m 2). The targets for the saccades consisted of LEDs spaced at +_ 2.5 ° and + 5 ° to the right and left of the central fixation point. The four targets were used to reduce the likelihood of anticipatory saccades. The LEDs were embedded in a translucent screen such that they were only visible when illuminated. However, as there was no a priori hypotheses regarding target laterality, the number of replications would be increased by collapsing the data across this factor.
A trial commenced with the onset of a central fixation LED target for a randomised foreperiod (900, 1050 or 1200 msecs). Subjects were instructed to maintain fixation at the centre until the illumination of a peripheral target, and then respond with a saccadic eye movement to the target, which was illuminated for 1000 msecs. The direction and amplitude of the target movement were randomized to avoid predictive saccades. On 50% of the trials ( 'S trials') a General Scanning mirror galvanometer (G300PO), in the projection path of the surrounding grating, caused it to be rapidly displaced by 1/2 a cycle. This contrast reversal occurred simultaneously with the onset of the target LED. S

254 T.J. Crawford / Acta Psychologica 92 (1996) 251-262
trials were randomly interleaved with NS trials. On each trial the intensity of the target was randomly assigned to one of the values 0.7, 0.8, 1.4 and 4.4 c d / m 2. An experimental session for a single subject consisted of 288 trials.
Eye movements were recorded binocularly using an infra-red scleral reflection device (EMS 130) with a linearity range of + 10 ° and a bandwidth of 100 Hz. Eye movements were sampled at 200 Hz which, experience in our laboratory has shown, is sufficient for the reliable measurement of saccadic latencies. The target paradigm and eye movement sampling were controlled by an LSI 2 / 2 0 minicomputer. Eye movement analysis was conducted off-line with an interactive software program. The latency of the primary saccade was measured from the onset of the peripheral LED based on a saccade detection algorithm with a velocity criterion of 30°/sec in addition to a minimum acceleration over three consecutive samples. The interactive procedure ensured that artifacts due to blinks were rejected from the analyses. Head movements were restrained by the use of an adjustable chin rest. The experiments were conducted in the dark.
It was necessary to determine whether any effects on saccades were attributable to reduced visual sensitivity induced by the display, therefore visual detection thresholds were obtained for three subjects prior to the eye movement task. Subjects were required to maintain fixation at the centre of the screen and respond with a manual key-press on detection of a target. The format for target presentation was identical to the eye movement condition. Visual thresholds were determined using a 4-alternative forced- choice procedure according to a modified PEST adaptive staircase (Findlay, 1978) which aimed at a threshold level of 75%. S trials were randomly interleaved with NS trials.
2.1.1. Statistical analyses The study used a 2-way repeated measures design consisting of 'shift' condition (2
levels) and target intensity (4 levels). Univariate analyses of repeated measures requires the assumption of sphericity (see O'Brien and Kaiser, 1985); this was conducted using Mauchly's sphericity test. As a conservative procedure, to protect against false positives, univariate analyses were then conducted using corrected degrees of freedom using both Huynh-Feldt and the Greenhouse-Geisser calculations. The effects were also subjected to multivariate analysis (using Pillais, Hotelling-T and Wilks lamda) as a final analysis of the robustness of the effect since this requires the assumption of homogeneity of variance but not that of sphericity. All statistical analyses were conducted using the SPSS software statistics package.
2.2. Results
Mean latencies, shown in Table 1, of saccades on S and NS trials tabulated against target intensity satisfied the assumption of sphericity according to the Mauchly test (W = 0.7278, p = 0.735, ns). Saccade latencies were significantly longer on S compared to NS trials (F(I ,10) = 7.96, p = 0.018). There was a tendency for this effect to decline with target intensity which just failed to be significant on the Huynh-Feldt (F(3,30) = 2.58, p = 0.072). There was a marked main effect of target intensity with saccadic latency decreasing as a function of increasing target intensity (Huynh-Feldt, F(3,30) =
T.J. Crawford / Acta Psychologica 92 (1996) 251-262
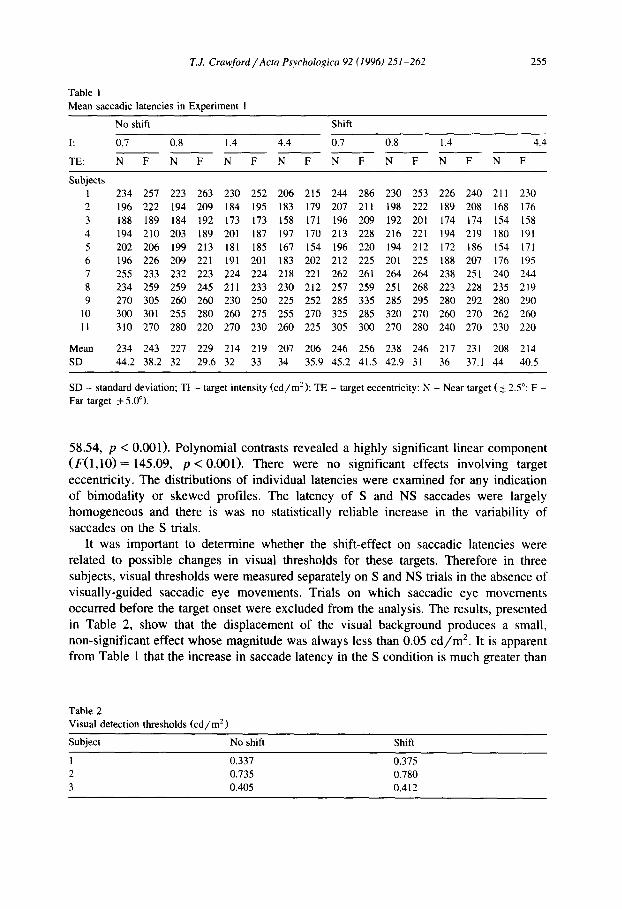
Table 1 Mean saccadic latencies in Experiment 1
255
No shift Shift
I: 0.7 0.8 1.4 4.4 0.7 0.8 1.4 4.4
TE: N F N F N F N F N F N F N F N F
Subjects 1 234 257 223 263 230 252 206 215 244 286 230 253 226 240 211 230 2 196 222 194 209 184 195 183 179 207 211 198 222 189 208 168 176 3 188 189 184 192 173 173 158 171 196 209 192 201 174 174 154 158 4 194 210 203 189 201 187 197 170 213 228 216 221 194 219 180 191 5 202 206 199 213 181 185 167 154 196 220 194 212 172 186 154 171 6 196 226 209 221 191 201 183 202 212 225 201 225 188 207 176 195 7 255 233 232 223 224 224 218 221 262 261 264 264 238 251 240 244 8 234 259 259 245 211 233 230 212 257 259 251 268 223 228 235 219 9 270 305 260 260 230 250 225 252 285 335 285 295 280 292 280 290
10 300 301 255 280 260 275 255 270 325 285 320 270 260 270 262 260 11 310 270 280 220 270 230 260 225 305 300 270 280 240 270 230 220
Mean 234 243 227 229 214 219 207 206 246 256 238 246 217 231 208 214 SD 44.2 38.2 32 29.6 32 33 34 35.9 45.2 41.5 42.9 31 36 37.1 44 40.5
SD - standard deviation; TI - target intensity (cd/m 2 ); TE - target eccentricity: N - Near target (_+ 2.5°: F - Far target 5: 5.0°).
58 .54, p < 0.001) . P o l y n o m i a l con t ras t s r evea l ed a h igh ly s ign i f i can t l inea r c o m p o n e n t
( F ( 1 , 1 0 ) = 145.09, p < 0 . 0 0 1 ) . The re were no s ign i f i can t e f fec ts i n v o l v i n g target
eccent r ic i ty . The d i s t r ibu t ions o f ind iv idua l l a t enc ies were e x a m i n e d for any ind ica t ion
o f b i m o d a l i t y or s k e w e d profi les . T he la tency o f S and N S saccades were la rgely
h o m o g e n e o u s and there is was no s ta t is t ical ly re l iab le inc rease in the var iab i l i ty of
saccades on the S trials.
I t was i m p o r t a n t to d e t e r m i n e w h e t h e r the sh i f t -e f fec t on saccadic la tenc ies were
re la ted to poss ib l e c h a n g e s in v isua l t h re sho lds for these targets . The re fo re in three
subjec ts , v i sua l t h re sho lds were m e a s u r e d separa te ly on S and NS trials in the absence o f
v i sua l l y -gu ided saccadic eye m o v e m e n t s . Tr ia ls on w h i c h saccadic eye m o v e m e n t s
o c c u r r e d be fo re the ta rge t onse t were exc luded f rom the analysis . The results , p r e sen t ed
in T a b l e 2, show that the d i s p l a c e m e n t o f the v isual b a c k g r o u n d p roduces a smal l ,
n o n - s i g n i f i c a n t e f fec t w h o s e m a g n i t u d e was a lways less than 0.05 c d / m 2. It is appa ren t
f rom T a b l e 1 tha t the inc rease in saccade la tency in the S cond i t i on is m u c h grea te r than
Table 2 Visual detection thresholds (cd/m 2)
Subject No shift Shift
I 0.337 0.375 2 0.735 0.780 3 0.405 0.412

256 T.J. Crawford/Acta Psychologica 92 (1996) 251-262
might be attributed to the effect on visual detection thresholds in the absence of saccadic eye movements.
3. Discussion
The current findings can be briefly summarised: 1. Saccadic eye movements were delayed to visual targets presented with a simultane-
ous movement of a textured background in the peripheral field compared to visually- guided saccades with a stationary background.
2. The mean latencies of saccades, with or without a background movement, showed an inverse linear trend against target intensity.
3. There was a small, non-significant increase in visual threshold for detection of the targets with the background movement, compared to the no background movement trials, showing that the origin of the saccadic effect was unlikely to lie exclusively within the visual pathways.
However, it remained possible that the latency effect might be sensitive to stimulus factors. Two aspects of the present findings indicated that visuo-temporal summation might be a critical factor. First, there was a slight tendency for the effect to decline with increasing target intensity. Second, although reaction times were not measured in the threshold detection condition, it is well known that reaction times for manual are typically longer than for saccadic eye movements (by about 100-300 msecs, e.g. see Crawford and Miiller, 1992). Thus, the longer time available for the detection of the stimuli remains as a possible significant factor in the visual detection results. Experiment 2 was designed to address this issue.
4. Experiment 2
In this experiment the saccadic targets were varied systematically by manipulating both the intensity and temporal persistence.
4.1. Methods
Five subjects, four of whom (Subjects 1-4) had participated in Experiment 1, were tested in both the S and NS conditions. LED targets could vary between 3 possible duration's (10, 100 and 1000 msecs) and two intensity levels (low - 0.7 c d / m 2 and high - 4.4 c d / m 2) and were presented in a fully randomized and counterbalanced order. Subjects were required to fixate the target LED as quickly and as accurately as possible. In this and all other respects the experimental conditions were identical to Experiment 1.
4.2. Results
Mean latency data for individual subjects are shown in Table 3. Univariate tests with corrected degrees of freedom (Huynh-Feldt epsilons) for repeated measures revealed a
T.J. Crawford / Acta Psychologica 92 (1996) 251-262
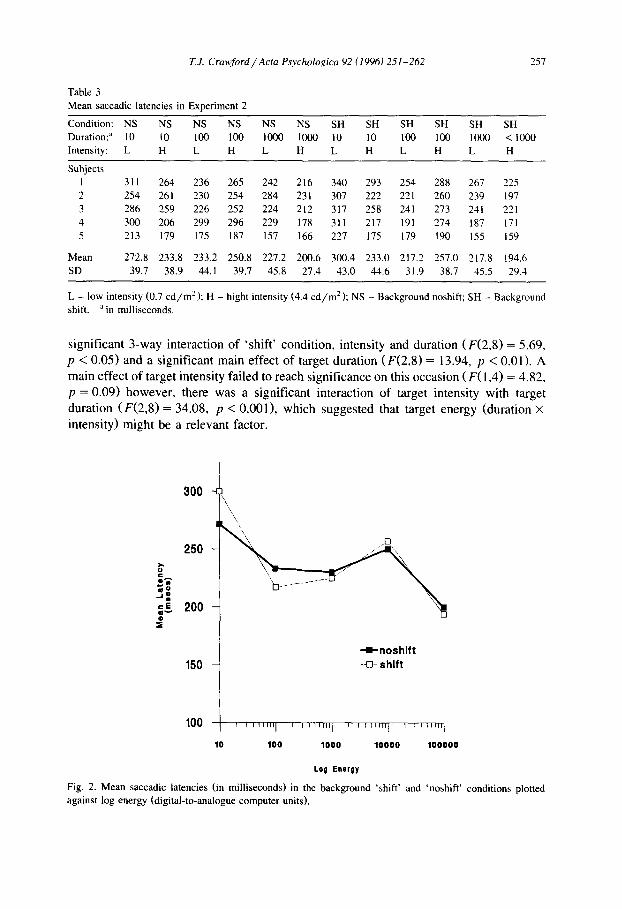
Table 3 Mean saccadic latencies in Experiment 2
257
Condition: NS NS NS NS NS NS SH SH SH SH SH SH Duration: a 10 10 100 100 1000 1000 10 10 100 100 1000 < 1000 Intensity: L H L H L H L H L H L H
Subjects 1 311 264 236 265 242 216 340 293 254 2 254 261 230 254 284 231 307 222 221 3 286 259 226 252 224 212 317 258 241 4 300 206 299 296 229 178 311 217 191 5 213 179 175 187 157 166 227 175 179
Mean 272.8 233.8 233.2 250.8 227.2 200.6 300.4 233.0 217.2 SD 39.7 38.9 44.1 39.7 45.8 27.4 43.0 44.6 31.9
288 267 225 260 239 197 273 241 221 274 187 171 190 155 159
257.0 217.8 194.6 38.7 45.5 29.4
L - low intensity (0.7 cd/m2); H - hight intensity (4.4 cd/m2); NS - Background noshift; SH - Background shift, ain milliseconds.
significant 3-way interaction of 'shift' condition, intensity and duration ( F ( 2 , 8 ) = 5.69, p < 0.05) and a significant main effect of target duration (F(2 ,8 ) = 13.94, p < 0.01). A main effect o f target intensity failed to reach significance on this occasion (F(1 ,4 ) = 4.82, p = 0.09) however, there was a significant interaction of target intensity with target duration ( F ( 2 , 8 ) = 34.08, p < 0.001), which suggested that target energy (duration x intensity) might be a relevant factor.
c
= E Ii V o
300
250
200
150
100
.,\ I "\\
\
\
.D
-m-noshift -~- shift
' '"'"1 ''"'"1 ' ' '""q ' ''"'"1 10 100 1 0 0 0 1 0 0 0 0 1 0 0 0 0 0
Log Energy
Fig. 2. Mean saccadic latencies (in milliseconds) in the background 'shift' and 'noshift' conditions plotted against log energy (digital-to-analogue computer units).
258 T.J. Crawford / Acta Psychologica 92 (1996) 251-262
Fig. 2 shows saccadic latencies plotted against target energy and illustrates a small shift-effect that was evident only at the weakest target signal. Saccadic latencies are maximal at the weakest signal level but reached an asymptote level thereafter, with little further decline at the high energy targets.
The 3-way interaction of 'shift ' condition, duration and intensity was apparently due to a shift-effect that was limited to targets at the lowest signal level (recall that the brightness × intensity interaction just failed significance in Experiment 1). An initially puzzling finding, however, was the absence of main effect of background 'shift ' at other target levels, in contrast to the previous experiment. Post-hoc analyses indicated that this might be partially attributed to a decline in the effect in the latter half of the session in some subjects and an absence of the effect in one highly experienced subject. Note that the subjects, who were naive to the stimulus conditions of Experiment 1, were experienced when tested in Experiment 2. In an attempt to analyse in more detail the possibility that extensive exposure to these background movement might be a critical short-term variable in determining the size of the effect, the experiment was subdivided into two halves; a first and second phase. Saccadic latencies in the first phase of the
30
20
H
o I I
-lO
-20
-30
-40
-5o - ~ l - - l - - l - - l - - l - - l - - l ~ ~
1 2 3 4 S ° 1 2 3 4 5"
Subjects
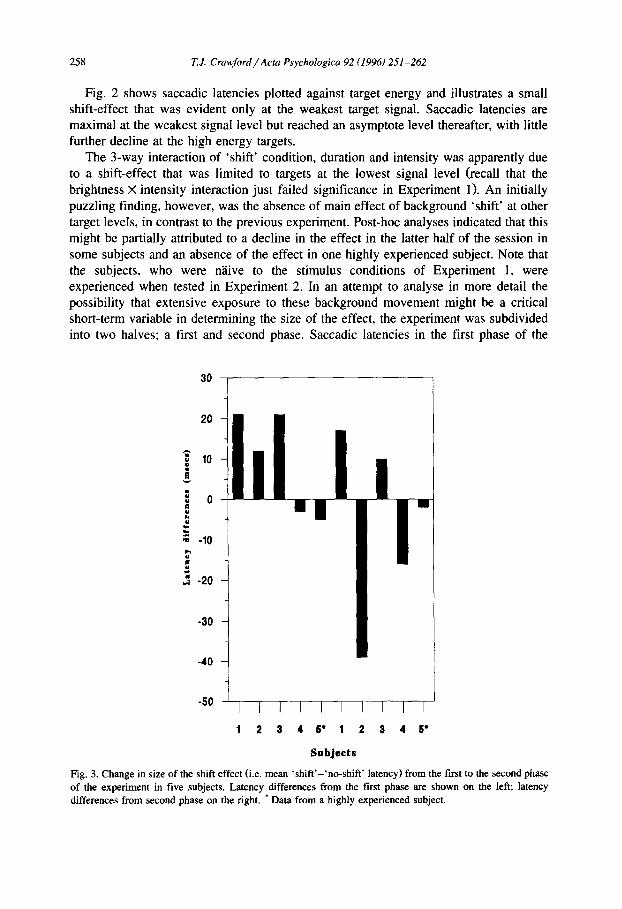
Fig. 3. Change in size of the shift effect (i.e. mean 'shift'-'no-shift' latency) from the first to the second phase of the experiment in five subjects. Latency differences from the first phase are shown on the left; latency differences from second phase on the right. * Data from a highly experienced subject.

T.J. Cra~ford / Acta Psychologica 92 (1996) 251-262 259
experiment were compared separately for each subject with latencies in the second phase. If stimulus exposure were a critical factor the shift-effect would be expected to decline from the first to the second phase of the experiment. The individual data for the mean latency differences (i.e. ' sh i f t ' - ' no-sh i f t ' ) are shown in Fig. 3. Subjects 1-3 showed a greater effect of the background in the first compared to the second phase. A fourth subject showed a slight negative effect in the first phase which, became even more negative in the second phase. Interestingly, Subject 5 who showed no sign of the effect was the author, who had spent many hours generating numerous saccades under these conditions and had apparently developed an erstwhile immunity to the shifting background!
5. Discussion
The primary finding of this study can be simply stated: saccadic eye movements in the context of the peripheral movement of a visual background are delayed compared to a stationary background. However, this finding is subject to at least two qualifications. (1) The effect is more robust with 'weak ' visual targets, and (2) the effect is likely to be influenced by adaptive mechanisms such that the size of the effect may decline with repeated exposure. Although somewhat speculative at present, the findings would seem to support processing by an adaptive spatial filter, resulting in reduced distraction on visual attention and the programming of saccadic eye movements. Rapid saccadic adaptation to the effects of peripheral non-target stimuli has previously been shown by Findlay and Crawford (1983). A subject presented with double-targets within the same hemifleld will frequently generate an initial saccade that lands at an intermediate position between the two targets, the so-called 'centre of gravity' (Findlay, 1982). Findlay and Crawford (1983) showed that extended exposure to these targets resulted in a marked reduction in the 'centre-of-gravity' effect. Such an adaptive mechanism is of obvious biological advantage, particularly in conditions where central visual attention might be distracted by the sudden movement of an object in the periphery. However, the finding that saccades were slightly delayed to targets of low signal energy suggests that the processing of weak visual stimuli may be especially resistant to such adaptive
1 processes. The studies on simultaneous double-target presentation have provided important
insights with regard to the effect of distracting target on saccadic latencies. In the Findlay (1982) study, saccadic latency was only increased in the condition when the target and non-target were simultaneously presented in different hemifields. A similar finding was reported by Zeevi et al. (1988). Lueck et al. (1990) showed that, compared
t On a recent train journey I noticed a scenario that seemed to parallel the laboratory experiments: during an attempt to read a book while the express train was in motion there was a distinct impression that my reading speed was slowed compared to when the train was stationary; the effect did not seem to be entirely explained by induced body, head and compensatory eye movements since when background motion from the environment was occluded the effect on reading speed was reduced. However, after some time the distracting effect of the background was less noticeable, presumably as adaptive processes emerged.

260 T.J. Crawford / Acta Psychologica 92 (1996) 251-262
to a saccade to an auditory target alone, a saccade towards an auditory target when simultaneously paired with a visual non-target, was similarly delayed, providing the target was displayed in the opposite hemifield. These findings provide support for the separation of the 'where' (i.e. spatial) and 'when' (i.e. temporal) signals in the programming of saccades (Becker and Jiirgens, 1979), and suggest that the distracting effect of non-target information is likely to disturb the control of the ' when' mechanism if the saccadic system is presented with ambiguous directional cues, while the 'where' system is sensitive to ambiguous amplitude signals within the same hemi-field. The findings of the present study adds to the known forms of 'distractors' that can delay the initiation of saccades and together with previous studies leads to the hypothesis that bi-visual field interactions may be a critical factor in this effect. However, further studies are required to determine whether the distraction effects are more apparent when saccades are generated in a context of bilateral compared to unilateral background movement.
5.1. Implications for uisual pathways
There is increasingly convincing evidence for two functionally distinct visual streams (magnocellular (M) and parvocellular (P)) projecting from the retina to lateral geniculate nucleus and through association cortex (DeYoe and Van Essen, 1988). Despite a continuing debate on the separation of the two pathways it is widely accepted that colour information is conveyed predominantly by the P-stream and that the M-stream processes transient and high velocity stimuli of low spatial frequency (see e.g. Schiller et al., 1990; Merigan et al., 1991). Burr et al. (1994) showed that saccades selectively suppress the M-pathway while sparing or even enhancing the P-pathway. As the M-pathway projects to visual attention areas in parietal cortex, the suppression of magnocellular channels might therefore inhibit the disturbing attentional effect of visual motion in the periphery.
Derrington (1984) showed that high spatial frequencies are relatively unimpaired by the peripheral motion of a visual pattern in comparison to the detection of low spatial frequencies implying that the shift-effect may primarily affect M-pathways. Saccades can be generated towards targets of high spatial frequency information, although they are delayed (relative to low spatial frequency targets) suggesting that visually-guided saccades are preferentially tuned to low spatial frequency channels (Deubel et al., 1985). Together these studies suggest that the shift-effect may constrain saccades to use the available intact, but slower, P-pathway.
5.2. Cortico-tectal mechanisms
Can the saccadic shift-effect can be explained in terms of neural activation within purely sensory or motor pathways or a combination of both? Visual thresholds, measured in the absence of saccades, showed little change in visual sensitivity demon- strating that the effect did not lie exclusively within sensory channels. The absence of any effect on the variance of saccade latencies implies that it's effect on saccades was unlikely to be due to temporal uncertainty caused by increased neural noise on detection channels. Further evidence of increased signal noise such as multi-stepping saccades

T.J. Cra~ford / Acta Psychologica 92 (1996) 251-262 261
(Crawford, 1991), were also markedly absent. Increased signal noise may, however, be filtered at a later stage in the processing of the visuo-motor information data stream. This could, for example, occur as a result of signal averaging within a distributed neural network (see Findlay, 1987).
One important site of sensory-motor interaction, with particular relevance to sac- cades, lies in the superior colliculus. The superficial layers are organised in a topograph- ical retinotopic map and with many cells that have large receptive fields. An interesting finding is that the cells with large receptive fields discharge before more superficial cells with smaller receptive fields (Mohler and Wurtz, 1976). This identifies a putative mechanism whereby stimulation over a large field could interact with detection in more focal channels.
The experimental manipulations in this study would have generated a perception of relative movement, it is therefore important to consider whether this factor may have had some influence on the saccadic eye movements. Recently (Davidson and Bender, 1991) it has been found that many cells in the superior colliculus are selective for relative motion between a focal target moving within the classic receptive field and a textured background moving outside the central receptive field. Moreover, it has been shown that this effect has a cortical origin, since a lesion restricted to the cortico-tectal pathway severely diminished this sensitivity (Davidson et al., 1992). Further evidence supporting the important contribution of cortical mechanisms to the processing of relative motion between a central target and a structured background comes from the finding that impaired smooth pursuit in this context is associated with lesions of inferior temporal cortex (Lawden et al., 1995). However the question of whether the saccadic shift-effect is dependent on cortico-tectal neural pathways or on more peripheral neural activation at the retino-geniculate level (Krtiger and Fischer, 1973; Fischer and Kriiger, 1974; Fischer et al., 1975; Kriiger et al., 1975) is at present unresolved.
Acknowledgements
The author would like to thank Drs. John Findlay, Andrew Derrington and two anonymous referees for their helpful comments on an earlier draft. This work was partly funded by the Medical Research Council and the Wellcome Trust. The experiments were conducted in the Department of Psychology, at the University of Durham, UK.
References
Becker, W. and R. Jtirgens, 1979. An analysis of the saccadic system by means of double step stimuli. Vision Research 19, 967-983.
Brooks, B.A. and D.M. Impelman, 1981. Suppressive effects of a peripheral grating displacement during saccadic eye movements and during fixation. Experimental Brain Research 42, 289-492.
Burr, D.C., M.C. Morrone and J. Ross, 1994. Selective suppression of the magnocellular visual pathway during saccadic eye movements. Nature 376, 511-513.
Crawlbrd, T.J., L. Henderson and C. Kennard, 1989a. Abnormalities of nonvisually-guided eye movements in Parkinson's disease. Brain 112, 1573-1586.
Crawford, T.J., S. Goodrich, L. Henderson and C. Kennard, 1989b. Anticipatory responses to predictable

262 T.J. Crawford / Acta Psychologica 92 (1996) 251-262
visual signals in Parkinson's disease: Manual keypresses and saccadic eye-movements. Journal Neurology, Neurosurgery and Psychiatry 52, 1033-1042.
Crawford, T.J., 1991. Multi-stepping saccadic sequences in humans. Acta Psychologica 76, 11-29. Crawford, T.J. and H.J. Miiller, 1992. Spatial and temporal effects of spatial attention on human saccadic eye
movements. Vision Research 32(2), 293-304. Davidson, R.A. and D.B. Bender, 1991. Selectivity for relative motion in the monkey superior colliculus.
Journal of Neurophysiology 65, 1115-1133. Davidson, R.A., T.J. Joly and D.B. Bender, 1992. Effect of corticotectal tract lesions on relative motion
selectivity in the monkey superior colliculus. Experimental Brain Research 92, 246-258. Derrington, A.M., 1984. Spatial frequency selectivity of remote pattern masking. Vision Research 24,
1965-1968. Deubel, H., J.M. Findlay, A.M. Jacobs and D. Brogan, 1985. 'Saccadic eye movements to targets defined by
structure differences'. In: G. Luer, U. Lass and J. Shallo-Hoffman (Eds.), Eye movement research: Physiological and psychological aspects. G~ttingen: C.J. Hogrefe.
DeYoe, E.A. and D.C. van Essen, 1988. Concurrent processing streams in monkey visual cortex. Trends in Neurosciences 11,219-226.
Findlay, J.M., 1978. Estimates on probability functions: A more virulent PEST. Perception and Psychophysics 23, 181-185.
Findlay, J.M., 1982. Global visual processing for saccadic eye movement. Vision Research 22, 1033-1045. Findlay, J.M., 1987. Visual computation and saccadic eye movements. Spatial Vision 2, 175-189. Findlay, J.M. and T.J. Crawford, 1983. 'The visual control of saccadic eye movements: Evidence for limited
plasticity'. In: R. Groner, C. Menz, D.F. Fisher and R.A. Monty (Eds.), Eye movements and psychological functions: International views. Hillsdale, NJ: Erlbaum.
Fischer, B. and J. Kriiger, 1974. The shift-effect in the cat's lateral geniculate neurones. Experimental Brain Research 21,225-227.
Fischer, B., J. Kriiger and W. Droll, 1975. Quantitative aspects of the shift-effect in cat retinal ganglion cells. Brain Research 83, 391-403.
Guitton D., H.A. Buchtal and R.M. Douglas, 1985. Frontal lobe lesions in man cause difficulties in suppressing reflexive glances and in generating goal directed saccades. Experimental Brain Research 58, 455-474.
KrUger, J., B. Fischer and R. Barth, 1975. The shift-effect in retinal ganglion cells of the rhesus monkey. Experimental Brain Research 23, 443-446.
Kriiger, J. and B. Fischer, 1973. Strong periphery effect in cat retinal ganglion cells. Excitatory responses in ON- and OFF-center neurons to single grid displacements. Experimental Brain Research 18 316-318.
Lawden, M.C., H. Bagelmann, T.J. Crawford, T.D. Matthews and C. Kennard, 1995. An effect of structured backgrounds on smooth pursuit eye movements in patients with cerebral lesions. Brain 118, 37-48.
Lueck, C.J., T.J. Crawford, C.J. Savage and C. Kennard, 1990. Auditory-visual interaction in the generation of saccades in man. Experimental Brain Research 82, 149-157.
Mcllwain, J.T., 1964. Receptive fields of optic tract axons and lateral geniculate cells: Peripheral extent and barbiturate sensitivity. Journal of Neurophysiology 27, 1154-1173.
Mcllwain, J.T., 1966. Some evidence concerning the physiological basis of the periphery effect in cat's retina. Experimental Brain Research 1,265-271.
Merigan, W.H., C.E. Byrne and J.H.R. Maunsell, 1991. Does primate motion perception depend on the magnocellular pathway? Joumal of Neuroscience 11, 3422-3429.
Mohler, C.W. and R.H. Wurtz, 1976. Organization of monkey superior colliculus: intermediate layers cells discharging before eye movements. Journal of Neurophysiology 39, 722-744.
O'Brien, R.G. and M.K. Kaiser, 1985. MANOVA method for analyzing repeated measures designs: An extensive primer. Psychological Bulletin 97(2), 316-333.
Schiller, P.H., N.K. Logothetis and E.R. Charles, 1990. Functions of the colour-opponent and broad-band channels of the visual system. Nature 343, 68-70.
Pierrot-Deseilligny, C.H., S. Rivaud, B. Gaymaud and Y. Agid, 1991. Cortical control of reflexive visually- guided saccades. Brain 114, 1473-1485.
Zeevi, Y., P.A. Wetzel and G.A. Geri, 1988. Preferences and asymmetries in saccadic responses to delayed bi-hemifield stimuli. Vision Research 28(10), 1145-1155.


















