
Transformation of microorganisms for the production of bioethanol from second and third generation...
45
“Second-generation bioethanol” Transformation of promissory microorganisms for the production of bioethanol on an industrial scale Godfrey Idrobo Libreros BSc, PhD. Sci. Biology Stu.
description
The production of fuels of the second generation and third generation needs of new alternative substrate that they do not compete with the food. The xilosa is the most abundant second sugar on the Earth, to be able to take advantage of it there are needed robust microorganisms that could assimilate and produce metabolitos of interest as the ethanol.
Transcript of Transformation of microorganisms for the production of bioethanol from second and third generation...

“Second-generation bioethanol” Transformation of promissory microorganisms for the
production of bioethanol on an industrial scale
Godfrey Idrobo LibrerosBSc, PhD. Sci. Biology Stu.

Utilización de microorganismos para la producción de etanol a partir de azúcares
obtenidos de residuos de caña
MADR
Godfrey Idrobo Libreros, BSc. Biol. Genética, Ph.D


Programa: Producción de etanol a partir de residuos de caña de azúcar_(MADR2008D3210)
Proyecto 1: Hidrólisis de residuos de la caña de azúcar para la obtención de azúcares simples como materia prima para la obtención de etanol (2007D3718-346-07)
Proyecto 2: Utilización de microorganismos para la producción de etanol a partir de azúcares obtenidos de residuos de caña(MADR 2008D3210-3466)
MADR

ProyectoMetodología - Marco lógico - Resultados
Toma de muestras biológicasFormulación de medios de cultivo-revisión bibliografía
Macerado o triturado de muestras molino
Diluciones 1:10 en H2O pep, estéril, y Exudados directos
Siembra en medio de cultivo idóneo Caracterización:
morfológica Gram. y lactofenol.
Purificación (re-asilamiento)
Incubación a (26-27ºC) 28-37ºC
Preparación inóculos frescos (Xilosa, ara, glucosa)
Biomasa (peso seco, absorbancia)Consumo de azúcar (HPLC)Producción de alcohol (HPLC)Cambio de pH
Asimilen, Crezcan y Co-fermenten Xilosa-Ara-Glucosa
Selección de microorganismos
Pruebas de tolerancia a Et-OH
Pruebas de fermentación (10-15 L)Alcohol producido será:
Evaluado (Alcotest), cuantificado (densímetro)
Prueba de Oxido-Fermentación
Fermentación en lote 30ºCCinética de crecimiento C5 y
C61

MetodologíaAislamiento
YEAmod2: Composición por litroP: 0.5 gramosEL: 0.5 gramosAA: 15 gramosXil y/o Ara: 5.0 gramosAgua dd: 1 Litro

Frecuencia de bacterias, levaduras y hongos aislados del muestreo en Bahía Málaga.
8.8 %
35.3 %
55.9 %

Cepas aisladas de Bahía Málaga con capacidad de crecimiento sobre azúcares C5 y C6.
3/34 aislamientos probados presentaron crecimiento a partir de xilosa y/o arabinosa o glucosa.
+: crecimiento

Frecuencia de crecimiento en glucosa (C6), xilosa y arabinosa (C5) de las cepas
aisladas de muestras colectadas en Bahía Málaga.
8.8 %

Aislamientos de sustratos vegetales

Crecimiento arabinosa Crecimiento xilosa
La utilización directa del medio selectivo permitió aislar un total de 6 bacterias, 1 levadura y 2 hongos de los residuos vegetales analizados.
De las cepas Amb 1 a 5 solamente presentó crecimiento en xilosa y arabinosa fue la Amb1.

Resultados preeliminares de prueba OF (Hugh & Leifson)

Contenido• Introducción• Antecedentes• Metodología de clonación (Termófilos
Gram+)• Producción de etanol
(Thermoanaerobacterium saccharolyticum )• Perspectivas

Ford Modelo A 1896

Godfrey
Vehículo Ford adaptado por el INT en el año 1925 para demostraciones del uso de bioetanol puro como combustible (BNDES y CGEE. 2008. Bioetanol de caña de azúcar: energía para el desarrollo Sostenible)

fatt
y ac
id p
athw
ays
Trends in Biotechnology. 2008. 26 (7)
isop
reno
id p
athw
ays
Central metabolic pathways and the potential fuel molecules derived from them
Godfrey
Central metabolic pathways and the potential fuel molecules derived from them. The blue box indicates isoprenoid pathways and isoprenoid-derived molecules.The yellow box indicates fatty acid pathways and fatty acid-derived molecules. Short-chain alcohols are shown in green text. Biomass-derived sugars are indicated in blue text.Abbreviations: 6PG: 6-phosphogluconate; CIT: citrateDHAP: dihydroxyacetone phosphateDMAPP: dimethylallyl pyrophosphateE4P: erythrose-4-phosphateF6P: fructose-6-phosphateFBP: fructose-1,6-bisphosphateFPP: farnesyl pyrophosphateFUM: fumarate; G´1P: galactose-1-phosphate; G1P: glucose-1-phosphate; G3P: glyceraldehyde-3-phosphate; G6P: glucose-6-phosphate;GGPP: geranylgeranyl pyrophosphate; GPP: geranyl pyrophosphate; ICT: isocitrate; IPP: isopentenyl pyrophosphate; M6P: mannose-6-phosphate; MAL: malate; OAA: oxaloacetate; OGA: 2-oxoglutarate; PEP: phosphoenolpyruvate; R5P: ribose-5-phosphate; S7P: sedoheptulose-7-phosphate; SUC: succinate; SUC-CoA: succinyl coenzyme A; X5P: xylulose-5-phosphate.

Nature reviews Microbiology 2009 (7) 715-23

Nature 2008 454: 841-45

Science 2007 315, 804-07
Godfrey
(A)A simplified model showing the interaction of the major polysaccharides in the cell wall. (Lignin is not shown here because its interactions are not well established.) In this system, hemicelluloses are closely associated to the surface of the rigid cellulose crystallite forming the microfibril network. Pectins are cross-linked polysaccharides forming a hydrated gel that “glues” the cell-wall components together. (B) The 36-chain model of the cellulose elementary fibril. Here, the depiction of the glucan chains is based generally on an x-ray structure of cellulose Ib (39). It has been proposed (16) that the cellulose elementary fibril may contain three groups of glucan chains: in group C1 (red) are six true crystalline chains; in group C2 (green) are 12 subcrystalline chains with a small degree of disorder; and in group C3 (blue) are 18 surface chains that are subcrystalline with a large degree of disorder.(C) The intra- and interchain hydrogen-bond network in cellulose I-Beta

Waites et. al 2005
Approximately 6–8 tons of dried leaves called trash is produced from one hectare of sugarcane crop (Singh et al. 2008)40%- celulosa (40-60% BNDES y CGEE 2008)25% hemicelulosa (20-40% BNDES y CGEE 2008)18-20% lignina (10-25% BNDES y CGEE 2008)
25%-85% w/w hemicellulose: xylose is the second much abundant sugar in nature after D-glucose
Godfrey
BNDESBanco Nacional de Desenvolvimento Econômico e Social.CGEECentro de Gestão e Estudos Estratégicos.

Saccharomyces cerevisiae más común Zymomonas mobilis homo fermentador Escherichia coli silvestre etanol + ácidos orgánicos
A well-known problem in S. cerevisiae is that xylose transport into the cell is inhibited by glucose, since xylose and glucose compete for the same transport systems (Kilian et. al 1988, Meinander et al. 1997)
etanol

Godfrey
Two routes to xylose assimilation:When xylose enters Sacchromyces cerevisiae it can be incorporated into the pentose phosphate pathway through either the three-enzyme pathway containing a xylitol intermediate or a two-step process that uses a fungal or bacterial xylose isomerase gene. The two-step process bypasses the need for the reducing power that is incorporated in NAD- and NADP-reducing partners and has been shown to improve ethanol production. Xylulose-5-phosphate is formed by both pathways and can enter into central carbon metabolism through the transketolase and transaldolase reactions.

Nature reviews Microbiology 2007 5 248-50
1. Producción de enzimas sacarolíticas (celulasas y hemicelulasas)
2. Hidrólisis de los polisacáridos presentes en la biomasa pretratada, es decir las rutas metabólicas para poder realizar dicha hidrólisis
3. Fermentación alcohólica de hexosas (glucosa)
4. Fermentación de pentosas Xilosa y Arabinosa



Metodology
Gene Cloning and Plasmid Construction (Desai et al. 2004) 18
Transformation of T. saccharolyticum (Mai et al. 1997, Klapath et al. 1996 ,Tyurin et. al 2005)14-16
Media Composition and Strain Storage (Baskaran et al. 1995) 25
Fermentation Conditions
Analytical Methods (Lu et al. 2006) 28
Enzymatic Assays (Shaw et al. 2008b) 12
Thermoanaerobacterium saccharolyticum L-ldh- (Desai et al. 2004) 18
Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
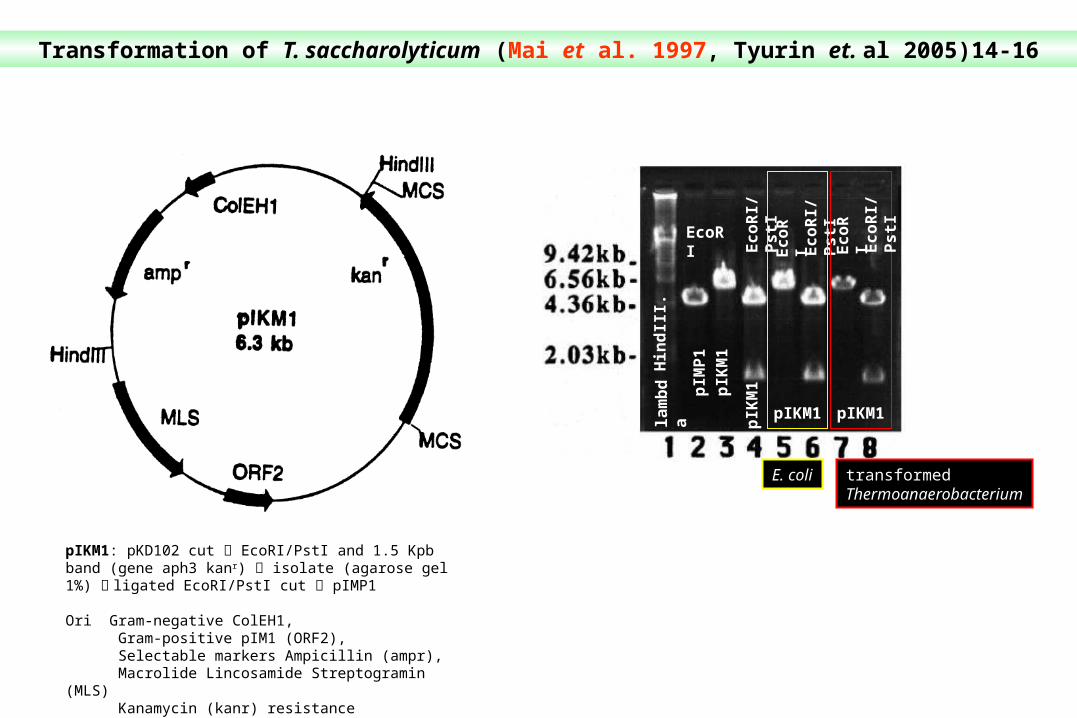
Transformation of T. saccharolyticum (Mai et al. 1997, Tyurin et. al 2005)14-16
pIKM1: pKD102 cut EcoRI/PstI and 1.5 Kpb band (gene aph3 kanr) isolate (agarose gel 1%) ligated EcoRI/PstI cut pIMP1
Ori Gram-negative ColEH1, Gram-positive pIM1 (ORF2), Selectable markers Ampicillin (ampr), Macrolide Lincosamide Streptogramin (MLS) Kanamycin (kanr) resistance
pIM
P1
lam
bda
pIK
M1
EcoRI
pIK
M1
EcoR
I/Pst
I
pIKM1
EcoR
I/Pst
I
EcoR
I
pIKM1
EcoR
I/Pst
I
EcoR
I
E. coli transformed Thermoanaerobacterium
Hin
dIII.
Godfrey
El problema: ausencia de un Ori termoestable en termofilos gran-positivos.Solución:

Immediately before electroporation.
Transformation of T. saccharolyticum (Mai et al. 1997, Tyurin et. al 2005)14-16
Godfrey
Future modi¢cation of pIKM1 including the incorporation of a strong promoter, ribosomebinding region and a multicloning site should enhance its utility for the expression of cloned genes in Thermoanaerobacterium or closely related bacteria.

Transformation of T. saccharolyticum (Mai et al. 1997, Klapath et al. 1996, Tyurin et. al 2005)14-16

Transformation of T. saccharolyticum (Mai et al. 1997, Klapath et al. 1996, Tyurin et. al 2005)14-16
Electropermeabilization• 24Mhz• 10kV• Oscillations are correlated with
high-frequency transformation

Thermoanaerobacterium saccharolyticum L-ldh- and Gene Cloning and Plasmid Construction (Desai et al. 2004) 18
Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
Thermoanaerobacterium saccharolyticum JW/SL-YS485 (Provided by Wiegel).25% glicerol + 75% medio N2
Culture recoveredDMS61 broth, 0.5 g cysteina/l, TransformatiónMYE mineral, YEA 2g/l, Xyl or Glu 5 g/l caracterization
Selectión 50-400µg kanamycin/ ml
Cloning of L-lactate dehydrogenase
Conserved regions from: C acetobutylicum, KPGETR, NPVDIL,Bacillus megaterius GEHGD, KGATYYBacillus sterothermophilus
Degenerated primers were designed CODEHOP program
Genomic DNA Thermoanaerobacterium saccharolyticum
Ldh fragment (Sau3AI) cloned DNA library secuenced GenBank AN: JW/SL-Y485-AY278026


Thermoanaerobacterium saccharolyticum L-ldh- and Gene Cloning and Plasmid Construction (Desai et al. 2004) 18
Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
Knockout vector construction
pSGD8: pUC18Amplification: primers degenerate Ldh (up and down): 1.0 kb and 0.43 KbA-tailed TOPO pCR2.1 pSGD8
KmR: kanamycin R, ApR: bla penicillin R, ldh fragments.
pIKM1

Thermoanaerobacterium saccharolyticum L-ldh- and Gene Cloning and Plasmid Construction (Desai et al. 2004) 18
Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
W TD1pIMM1Primers

Thermoanaerobacterium saccharolyticum L-ldh- and Gene Cloning and Plasmid Construction (Desai et al. 2004) 18
Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
Glu
cose
Xylo
se
Wild TD1 (ldh-)
X : OD□ : Ethanol : Substrate• : Lactate∆ : Acetate

Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
L-ldh- L-lactate dehydrogenase
ack- acetate kinase
pta- phosphateacetyltransferase
X
X
X

Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
L-ldh-
ack- pta-
ack- pta- L-ldh- (Lk1)

Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
1.2- and 0.6-kb regions 5´ and 3´ ofthe pta and ack genes were amplified
K1, K2 Kanamycin promoter region E1, E2 adenine methylase gene conferring erythromycin
resistance

Knockout vector construction
pSGD9: pUC18Amplification: primers degenerate pta and ack (up and down): 1.2 kb and 0.6 KbA-tailed TOPO pCR2.1 pSGD8
KmR: kanamycin R, ApR: bla penicillin R, ldh fragments.
thermostable S. faecalis kanamycin resistance cassette
Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
pta and ack homology regions

Fermentaciones en Batch cepa silvestre, L-ldh, pta/ack, y ALK1
En tubos bajo anaerobiosis (Ngas 5% v/v)4 g/l xilosa, 2.5 g/l extracto de levadura, 10 g/l buffer MES (pH 6.2). Temperatura: 55°C (Sin agitación)
Fermentaciones en continuo ALK1 y ALK2
Bioreactor de 2 L, Vt:0.5L, Applikon Instruments20–70 g/l xilosa10 g/l extracto de levadura 5 g/l triptona. Temperatura: 55°CpH: buffer del medio MTCAnaerobiosis: 30-60 min. Ngas, ResazurinRecuperación de CO2 : contenedor con agua
Fermentaciones en Batch ALK2
Bioreactor de 3 L, Vt:1L, Sartorius12 g/l xilosa, 5 g/l extracto de levadura, pH 5.5, 200 rpmTemperatura: 55°C
Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)

*Peso seco, etanol, ● xilosa, □ ácido acético, ◊ ácido láctico
Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
Godfrey
Fermentación LoteXilose, 55°C, bioreactors controlled at pH 6.0. (A) Wild-type strain(B) ALK2 strain. Xilosa: circulos verdes llenos, Etanol: circulos cafe, Ácido lactico: diamantes rojosÁcido Acético cuadrados azulesPeso seco: *Carbon recovery for the wild-type fermentation is 107% and the ALK2 fermentation is 102%

Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)
Xylose, glucose Mannosegalactose EthanolDry weight (*).

Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)

Thermoanaerobacterium saccharolyticum L-ldh-, ack- pta-, ack- pta- L-ldh-(Shaw et al. 2008)

Perspective• Transition first second generation biofuel: wild-type
genetically engineered variants with superior catabolic properties and homoethanologenic production phenotypes.
• Research strategy in thermophiles by a lack of the same degree of knowledge (Colombia)
• Construction of nonnatural metabolism that allows the biosynthesis, for the first time, of an array of alcohols not readily produced by microorganisms (Higher-chain alcohol)


















