TP receptor-mediated release of eosinophil chemotactic activity from human bronchial smooth muscle...
7
Immunopharmacology and Inflammation TP receptor-mediated release of eosinophil chemotactic activity from human bronchial smooth muscle cells ☆ Yusuke Suzuki, Koichiro Asano ⁎, Kyoko Niimi, Jun Miyata, Yoshiki Shiraishi, Koichi Fukunaga, Tetsuya Shiomi, Takeshi Nakajima, Tsuyoshi Oguma, Koichi Sayama, Akitoshi Ishizaka Division of Pulmonary Medicine, Department of Medicine, Keio University School of Medicine, 35 Shinanomachi, Shinjuku-ku, Tokyo 160-8582, Japan abstract article info Article history: Received 8 July 2008 Received in revised form 22 September 2008 Accepted 29 September 2008 Available online 8 October 2008 Keywords: Asthma Interleukin-4 Chemotaxis CCL11 CCR3 There are reports indicating that thromboxane A 2 receptors (TP receptors) may stimulate the eosinophil accumulation in the lower airways of asthmatics, however, the mechanisms behind such an effect remain unknown. We quantified the synthesis of eosinophil chemotactic activity and eosinophilic CC chemokines, including CCL5, CCL7, CCL8, CCL11, CCL13, CCL24, and CCL26 in primary cultures of human bronchial smooth muscle cells (BSMC) stimulated with a prostanoid TP receptor agonist, IBOP (10 - 9 –10 - 7 M). The activation of prostanoid TP receptors in BSMC induced the release of potent eosinophil chemoattractant(s) in the presence of interleukin (IL)-4. CCL11/eotaxin-1 was the only synthesis significantly increased by IBOP co-stimulated with IL-4, and pretreatment with an anti-CCL11 antibody abrogated the eosinophil chemotactic activity released from IBOP/IL-4-stimulated BSMC. The effect of IBOP was also completely blocked by pretreatment with a prostanoid TP receptor-specific antagonist, AA-2414. IBOP had no effect on the expression of IL-4 receptor-α, or on the IL-4-induced phosphorylation of STAT6 in BSMC. In conclusion, activation of prostanoid TP receptors in a Th2-dominant microenvironment might exacerbate the eosinophilic inflammation of the airways by synthesis and release of CCL11 from BSMC. © 2008 Elsevier B.V. All rights reserved. 1. Introduction The airways of allergic patients synthesize excessive amounts of thromboxane (Tx)A 2 , as manifested by an increased concentrations of TxB 2 and 11-dehydro-TxB 2 in urine, bronchoalveolar lavage fluid, or exhaled air condensate recovered from patients with asthma and other allergic diseases (Huszar et al., 2005; Kumlin et al., 1992; Montuschi and Barnes, 2002; Narita et al., 1995). Furthermore, other ligands of TxA 2 receptor (TP) such as prostaglandin D 2 and 8- isoprostane F 2 α, are abundantly present in asthmatic airways (Cole- man and Sheldrick, 1989; Janssen et al., 2000). The activation of prostanoid TP receptors accentuates the airflow limitation in the lower airways by mucosal edema, smooth muscle contraction, and disturbance of mucociliary clearance (Allen et al., 2006; Martin et al., 2000; Richter et al., 2007; Xiang et al., 2002), while pharmacological blockade of prostanoid TP receptors attenuates bronchial hyper- responsiveness and alleviates the respiratory symptoms of asthmatic patients (Nagai et al., 2006; Rolin et al., 2006). We and others have also recently reported that prostanoid TP receptor agonists accelerate the proliferation of bronchial smooth muscle cells (BSMC) (Capra et al., 2003; Suzuki et al., 2004), suggesting a link between prostanoid TP receptor activation and airway remodeling. In contrast, the role of prostanoid TP receptor-mediated signals on the pathogenesis of eosinophilic inflammation in the airways, a cardinal characteristic of asthma, is less well known. Previous studies have shown that prostanoid TP receptor antagonists can efficiently downregulate eosinophilic airway inflammation in animals and in asthmatics (Fukuoka et al., 2003; Hoshino et al., 1999; Itoh et al., 1993, 1996; Matsumoto et al.,1994; Shi et al., 1998). The anti-inflammatory effect of prostanoid TP receptor antagonists in patients with asthma was first observed in a study by Hoshino et al., who examined the pathology of bronchial mucosa before and after 4 months of treatment with a prostanoid TP receptor antagonist, AA-2414 [7-(3,5,6- trimethyl-1,4-benzoquinon-2-yl)-7-phenylheptanoic acid], or placebo in 31 asthmatics (Hoshino et al., 1999). They observed a significant decrease in the number of EG2-positive eosinophils in the airways treated with AA-2414. Fukuoka et al. also reported that treatment with AA-2414 for 12 weeks significantly decreased the concentration of eosinophil cationic protein in sputum, which returned to baseline after 12 weeks of discontinuation of AA-2414 (Fukuoka et al., 2003). Similar effects were observed with other prostanoid TP receptor antagonists in experimental models of allergic airway inflammation (Itoh et al., 1996; Shi et al., 1998). However, precisely how prostanoid European Journal of Pharmacology 600 (2008) 133–139 ☆ This work was supported by Grants-in-Aid from the Japanese Ministries of Education, Culture, Sports, Science and Technology and of Health, Labor and Welfare, and by the Keio Gijuku Academic Development Fund. ⁎ Corresponding author. Tel.: +813 3353 1211; fax: +81 3 3353 2502. E-mail address: [email protected] (K. Asano). 0014-2999/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2008.09.044 Contents lists available at ScienceDirect European Journal of Pharmacology journal homepage: www.elsevier.com/locate/ejphar
-
Upload
yusuke-suzuki -
Category
Documents
-
view
216 -
download
3
Transcript of TP receptor-mediated release of eosinophil chemotactic activity from human bronchial smooth muscle...

European Journal of Pharmacology 600 (2008) 133–139
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar
Immunopharmacology and Inflammation
TP receptor-mediated release of eosinophil chemotactic activity from humanbronchial smooth muscle cells☆
Yusuke Suzuki, Koichiro Asano ⁎, Kyoko Niimi, Jun Miyata, Yoshiki Shiraishi, Koichi Fukunaga,Tetsuya Shiomi, Takeshi Nakajima, Tsuyoshi Oguma, Koichi Sayama, Akitoshi IshizakaDivision of Pulmonary Medicine, Department of Medicine, Keio University School of Medicine, 35 Shinanomachi, Shinjuku-ku, Tokyo 160-8582, Japan
☆ This work was supported by Grants-in-Aid fromEducation, Culture, Sports, Science and Technology andand by the Keio Gijuku Academic Development Fund.⁎ Corresponding author. Tel.: +81 3 3353 1211; fax: +8
E-mail address: [email protected] (K. Asano
0014-2999/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.ejphar.2008.09.044
a b s t r a c t
a r t i c l e i n f oArticle history:
There are reports indicatin Received 8 July 2008Received in revised form 22 September 2008Accepted 29 September 2008Available online 8 October 2008Keywords:AsthmaInterleukin-4ChemotaxisCCL11CCR3
g that thromboxane A2 receptors (TP receptors) may stimulate the eosinophilaccumulation in the lower airways of asthmatics, however, the mechanisms behind such an effect remainunknown. We quantified the synthesis of eosinophil chemotactic activity and eosinophilic CC chemokines,including CCL5, CCL7, CCL8, CCL11, CCL13, CCL24, and CCL26 in primary cultures of human bronchial smoothmuscle cells (BSMC) stimulated with a prostanoid TP receptor agonist, IBOP (10−9–10−7 M). The activation ofprostanoid TP receptors in BSMC induced the release of potent eosinophil chemoattractant(s) in the presenceof interleukin (IL)-4. CCL11/eotaxin-1 was the only synthesis significantly increased by IBOP co-stimulatedwith IL-4, and pretreatment with an anti-CCL11 antibody abrogated the eosinophil chemotactic activityreleased from IBOP/IL-4-stimulated BSMC. The effect of IBOP was also completely blocked by pretreatmentwith a prostanoid TP receptor-specific antagonist, AA-2414. IBOP had no effect on the expression of IL-4receptor-α, or on the IL-4-induced phosphorylation of STAT6 in BSMC. In conclusion, activation of prostanoidTP receptors in a Th2-dominant microenvironment might exacerbate the eosinophilic inflammation of theairways by synthesis and release of CCL11 from BSMC.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
The airways of allergic patients synthesize excessive amounts ofthromboxane (Tx)A2, as manifested by an increased concentrations ofTxB2 and 11-dehydro-TxB2 in urine, bronchoalveolar lavage fluid, orexhaled air condensate recovered from patients with asthma andother allergic diseases (Huszar et al., 2005; Kumlin et al., 1992;Montuschi and Barnes, 2002; Narita et al., 1995). Furthermore, otherligands of TxA2 receptor (TP) such as prostaglandin D2 and 8-isoprostane F2α, are abundantly present in asthmatic airways (Cole-man and Sheldrick, 1989; Janssen et al., 2000). The activation ofprostanoid TP receptors accentuates the airflow limitation in thelower airways by mucosal edema, smooth muscle contraction, anddisturbance of mucociliary clearance (Allen et al., 2006; Martin et al.,2000; Richter et al., 2007; Xiang et al., 2002), while pharmacologicalblockade of prostanoid TP receptors attenuates bronchial hyper-responsiveness and alleviates the respiratory symptoms of asthmaticpatients (Nagai et al., 2006; Rolin et al., 2006).We and others have also
the Japanese Ministries ofof Health, Labor and Welfare,
1 3 3353 2502.).
l rights reserved.
recently reported that prostanoid TP receptor agonists accelerate theproliferation of bronchial smooth muscle cells (BSMC) (Capra et al.,2003; Suzuki et al., 2004), suggesting a link between prostanoid TPreceptor activation and airway remodeling.
In contrast, the role of prostanoid TP receptor-mediated signals onthe pathogenesis of eosinophilic inflammation in the airways, acardinal characteristic of asthma, is less well known. Previous studieshave shown that prostanoid TP receptor antagonists can efficientlydownregulate eosinophilic airway inflammation in animals and inasthmatics (Fukuoka et al., 2003; Hoshino et al., 1999; Itoh et al., 1993,1996; Matsumoto et al., 1994; Shi et al., 1998). The anti-inflammatoryeffect of prostanoid TP receptor antagonists in patients with asthmawas first observed in a study by Hoshino et al., who examined thepathology of bronchial mucosa before and after 4months of treatmentwith a prostanoid TP receptor antagonist, AA-2414 [7-(3,5,6-trimethyl-1,4-benzoquinon-2-yl)-7-phenylheptanoic acid], or placeboin 31 asthmatics (Hoshino et al., 1999). They observed a significantdecrease in the number of EG2-positive eosinophils in the airwaystreatedwith AA-2414. Fukuoka et al. also reported that treatment withAA-2414 for 12 weeks significantly decreased the concentration ofeosinophil cationic protein in sputum, which returned to baselineafter 12 weeks of discontinuation of AA-2414 (Fukuoka et al., 2003).Similar effects were observed with other prostanoid TP receptorantagonists in experimental models of allergic airway inflammation(Itoh et al., 1996; Shi et al., 1998). However, precisely how prostanoid

134 Y. Suzuki et al. / European Journal of Pharmacology 600 (2008) 133–139
TP receptor antagonists inhibit the recruitment of eosinophils intoallergic airways remains unknown.
We hypothesized that airway resident cells that express prostanoidTP receptors, such as BSMC (Capra et al., 2003; Suzuki et al., 2004),produce eosinophil chemotactic factors such as chemokines inresponse to the activation of prostanoid TP receptors. The chemokinereceptor almost exclusively expressed on eosinophils is CCR3, whichresponds to several of its ligands, including 1) CCL5/regulated onactivation, normal T-cells, expressed and secreted (RANTES), 2) CCL7/monocyte chemoattractant factor-3 (MCP-3), 3) CCL8/MCP-2, 4)CCL11/eotaxin-1, 5) CCL13/MCP-4, 6) CCL24/eotaxin-2, and 7) CCL26/eotaxin-3 (Kaplan, 2001). The present study was designed to test ourhypothesis in primary cultures of human BSMC. We confirmed thatprostanoid TP receptor-mediated signals, in association with Th2cytokines, induce the release of chemotactic activity for eosinophilsvia the synthesis of CCL11.
2. Materials and methods
2.1. Cell cultures
Primary cultures of normal human BSMC were obtained fromCambrex (East Rutherford, NJ), and maintained in culture mediumcontaining 5% fetal bovine serum (FBS), 1 ng/ml of human recombi-nant epidermal growth factor, 2 ng/ml of fibroblast growth factor,10 μg/ml of insulin, and gentamicin and amphotericin B. BSMC at their5th–6th passages, from 5–6 different donors, were used for theexperiments, after a 24 h-depletion of FBS and growth factors.
2.2. Assay of eosinophil chemotactic activity in the supernatant of BSMCcultures
Confluent BSMC were stimulated with a TxA2 mimetic [1S-1α,2β(5Z),3α(1E,3R⁎),4α]]-7-[3-(3-hydroxy-4-(4′-iodophenoxy)-1-butenyl)-7-oxabicyclo-[2.2.1]heptan-2-yl]-5-heptenoic acid (IBOP,10−8Min 0.001% ethanol, Cayman Chemical Co., Ann Arbor, MI) in thepresence or absence of 1–10 ng/ml of interleukin (IL)-4 (PeproTech ECLtd., London, UK). The culture supernatants were collected 24 h later,centrifuged for 15 min at 1710 g at 4 °C, and stored at −80 °C forsubsequent experiments.
Peripheral blood eosinophils were isolated from non-atopic,healthy volunteers using the CD16-negative selection method (Niimiet al., 2007). Eosinophils (99±1%, mean±SEM) were resuspended inRPMI medium supplemented with 10% FBS. The eosinophil chemo-tactic activity was examined, using a slightly modified methoddescribed previously (Niimi et al., 2007). In brief, 300 μl of the culturesupernatants of BSMC stimulated with or without IL-4/IBOP, recom-binant human CCL11 (100 ng/ml, R&D systems, Minneapolis, MN), orplatelet activating factor (10 nM, Avanti Polar-Lipids, Alabaster, AL)was diluted in RPMI medium supplemented with 10% FBS, and wasadded to the lower compartment of a 96-well chemotaxis plate,separated by a 3-µm pore size filter (Neuroprobe Inc., Gaithersburg,MD). In some experiments, the culture supernatants, CCL11, or plateletactivating factor was incubated with 10 μg/ml of anti-CCL11neutralizing antibody (R&D systems) or control antibody (mouseIgG1) at 37 °C, for 1 h before the assay. Eosinophils (5×104 cells in50 µl) were then placed in the upper compartment. As standards foreosinophil peroxidase activity assay, designated number of eosino-phils (5×102–3×104 cells/well) were placed only in the lowercompartment of separated wells. After 1 h of incubation at 37 °C,the medium on the upper surface of the filter was wiped to removethe non-migrated eosinophils, and replaced by 0.5 mM EDTA in PBS.Following another hour of incubation, the plate was centrifuged at430 g at 4 °C for 20 min, the filter was gently removed, and thesupernatant aspirated. The eosinophil peroxidase activity in the lowerchamber was determined by incubating the plate for 10 min with
substrate solution (0.5 mM o-phenylenediamine, 10 mM H2O2, and0.1% Triton X-100 in Tris buffer, pH 8.0), followed by the addition of4 M H2SO4 to stop the reaction, and the absorbance was measured at490 nm.
2.3. Expression of CCR3 ligand mRNA in BSMC
After stimulation for 4–24 h with IBOP (10−8 M), IL-4 (10 ng/ml), orboth, total RNA in BSMC was extracted using a RNeasy Mini Kit(QIAGEN, Hilden, Germany). CCR3 ligand (CCL5, CCL7, CCL8, CCL11,CCL13, CCL24, CCL26) and glycerol-aldehyde dehydrogenase (GAPDH)mRNA concentrations were measured by reverse transcription, usingSuperScript III (Invitrogen Corp., Carlsbad, CA) followed by quantita-tive PCR amplification, using the TaqMan method (ABI PRISM 7000,Applied Biosystems Inc., Foster City, CA). Premixed PCR primers andTaqMan probes for human CCR3 ligands and GAPDH were obtainedfrom Applied Biosystems Inc. (Assay-on-Demand). The conditions forPCR were as follows: 1 cycle at 95 °C for 9 min, 50 cycles at 95 °C for0.5 min and at 60 °C for 1 min, and 1 cycle at 72 °C for 5 min. Thedifference of the threshold cycle (CT) for CCR3 ligands and for GAPDH(ΔCT) was determined for each samples, and then ΔΔCT wasdetermined to calculate the fold increase in the amount of chemokinetranscripts by comparing ΔCT in medium-treated cells (medium) andthat in IBOP/IL-4-stimulated cells. The mRNA levels in the “medium”
groups were defined as 1.0.
2.4. Measurements of CCR3 ligand concentrations in the supernatant ofBSMC cultures
BSMC were stimulated with IBOP, 10−9–10−7 M, or IL-1, 1 ng/ml(R&D systems), in the presence or absence of IL-4, 0.1–10 ng/ml. Insome experiments, the cells were pretreated for 30 min and co-incubated with a prostanoid TP receptor antagonist, AA-2414 (0.1–3 µM in 0.001–0.03% dimethyl sulfoxide) during the stimulationperiod. The cell culture supernatant was then retrieved, and theconcentrations of CCL7, CCL8, CCL11, CCL13, and CCL26weremeasuredby enzyme-linked immunosorbent assay (ELISA), according to themanufacturer's instructions (R&D systems).
2.5. Flow cytometric analysis of IL-4 receptor expression on BSMC
BSMC stimulated for 24 h with IBOP (10−8 M) were harvested,using 0.05% trypsin-EDTA, and resuspended in PBS containing 0.5%BSA. The cells, in a concentration of 107cells/ml, were incubated with2.5 µg/ml of anti-IL-4 receptor antibody (R&D systems) or control IgG(mouse IgG2a) at 4 °C for 30min, washed twice, then incubated at 4 °Cfor 30minwith a FITC-labeled anti-mouse IgG F(ab)2 (DAKO, Glostrup,Denmark). The labeled cells were analyzed by flow cytometry(FACScan, Becton Dickinson, Franklin Lakes, NJ).
2.6. Western blot and in-cell enzyme-linked immunosorbent analysis ofsignal transducer and activator of transcription 6
The activation of signal transducer and activator of transcription(STAT) 6 was determined by Western blot analysis of the phosphory-lated proteins. BSMC were cultured in the presence or absence of10−8 M of IBOP for 24 h, then stimulated with 10 ng/ml of IL-4 for15 min. The reaction was terminated by adding ice-cold PBS, and thecellular proteins were extracted with Cell Lysis Buffer (Cell SignalingTechnology, Beverly, MA) supplemented with 0.1% SDS and 1 mMPMSF. Extracts were mixed with sample buffer (0.05 M Tris–HCl, 2%SDS, 6% β-mercaptoethanol, and 10% glycerol), boiled at 95 °Cfor 5 min, and electrophoresed on 10% polyacrylamide gel. Afterelectrotransfer to a polyvinylidine difluoride membrane, Westernblot analysis was carried out with either an anti-phospho STAT-6(tyrosine 641) monoclonal antibody (Cell Signaling Technology), or
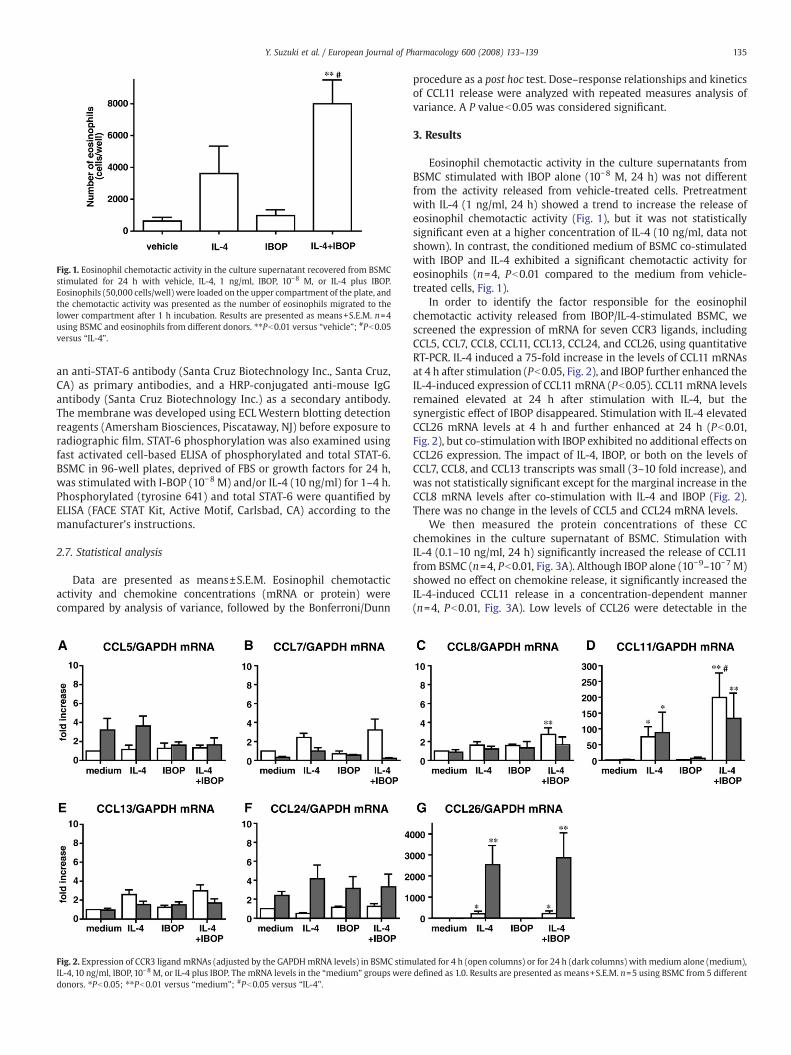
Fig. 1. Eosinophil chemotactic activity in the culture supernatant recovered from BSMCstimulated for 24 h with vehicle, IL-4, 1 ng/ml, IBOP, 10−8 M, or IL-4 plus IBOP.Eosinophils (50,000 cells/well) were loaded on the upper compartment of the plate, andthe chemotactic activity was presented as the number of eosinophils migrated to thelower compartment after 1 h incubation. Results are presented as means+S.E.M. n=4using BSMC and eosinophils from different donors. ⁎⁎Pb0.01 versus “vehicle”; #Pb0.05versus “IL-4”.
135Y. Suzuki et al. / European Journal of Pharmacology 600 (2008) 133–139
an anti-STAT-6 antibody (Santa Cruz Biotechnology Inc., Santa Cruz,CA) as primary antibodies, and a HRP-conjugated anti-mouse IgGantibody (Santa Cruz Biotechnology Inc.) as a secondary antibody.The membrane was developed using ECLWestern blotting detectionreagents (Amersham Biosciences, Piscataway, NJ) before exposure toradiographic film. STAT-6 phosphorylation was also examined usingfast activated cell-based ELISA of phosphorylated and total STAT-6.BSMC in 96-well plates, deprived of FBS or growth factors for 24 h,was stimulated with I-BOP (10−8 M) and/or IL-4 (10 ng/ml) for 1–4 h.Phosphorylated (tyrosine 641) and total STAT-6 were quantified byELISA (FACE STAT Kit, Active Motif, Carlsbad, CA) according to themanufacturer's instructions.
2.7. Statistical analysis
Data are presented as means±S.E.M. Eosinophil chemotacticactivity and chemokine concentrations (mRNA or protein) werecompared by analysis of variance, followed by the Bonferroni/Dunn
Fig. 2. Expression of CCR3 ligand mRNAs (adjusted by the GAPDHmRNA levels) in BSMC stimIL-4, 10 ng/ml, IBOP, 10−8 M, or IL-4 plus IBOP. The mRNA levels in the “medium” groups weredonors. ⁎Pb0.05; ⁎⁎Pb0.01 versus “medium”; #Pb0.05 versus “IL-4”.
procedure as a post hoc test. Dose–response relationships and kineticsof CCL11 release were analyzed with repeated measures analysis ofvariance. A P valueb0.05 was considered significant.
3. Results
Eosinophil chemotactic activity in the culture supernatants fromBSMC stimulated with IBOP alone (10−8 M, 24 h) was not differentfrom the activity released from vehicle-treated cells. Pretreatmentwith IL-4 (1 ng/ml, 24 h) showed a trend to increase the release ofeosinophil chemotactic activity (Fig. 1), but it was not statisticallysignificant even at a higher concentration of IL-4 (10 ng/ml, data notshown). In contrast, the conditioned medium of BSMC co-stimulatedwith IBOP and IL-4 exhibited a significant chemotactic activity foreosinophils (n=4, Pb0.01 compared to the medium from vehicle-treated cells, Fig. 1).
In order to identify the factor responsible for the eosinophilchemotactic activity released from IBOP/IL-4-stimulated BSMC, wescreened the expression of mRNA for seven CCR3 ligands, includingCCL5, CCL7, CCL8, CCL11, CCL13, CCL24, and CCL26, using quantitativeRT-PCR. IL-4 induced a 75-fold increase in the levels of CCL11 mRNAsat 4 h after stimulation (Pb0.05, Fig. 2), and IBOP further enhanced theIL-4-induced expression of CCL11 mRNA (Pb0.05). CCL11 mRNA levelsremained elevated at 24 h after stimulation with IL-4, but thesynergistic effect of IBOP disappeared. Stimulation with IL-4 elevatedCCL26 mRNA levels at 4 h and further enhanced at 24 h (Pb0.01,Fig. 2), but co-stimulationwith IBOP exhibited no additional effects onCCL26 expression. The impact of IL-4, IBOP, or both on the levels ofCCL7, CCL8, and CCL13 transcripts was small (3–10 fold increase), andwas not statistically significant except for the marginal increase in theCCL8 mRNA levels after co-stimulation with IL-4 and IBOP (Fig. 2).There was no change in the levels of CCL5 and CCL24 mRNA levels.
We then measured the protein concentrations of these CCchemokines in the culture supernatant of BSMC. Stimulation withIL-4 (0.1–10 ng/ml, 24 h) significantly increased the release of CCL11from BSMC (n=4, Pb0.01, Fig. 3A). Although IBOP alone (10−9–10−7 M)showed no effect on chemokine release, it significantly increased theIL-4-induced CCL11 release in a concentration-dependent manner(n=4, Pb0.01, Fig. 3A). Low levels of CCL26 were detectable in the
ulated for 4 h (open columns) or for 24 h (dark columns) with medium alone (medium),defined as 1.0. Results are presented as means+S.E.M. n=5 using BSMC from 5 different

Fig. 3. Concentrations of CCL11 (A) and CCL26 (B) proteins in the culture supernatant ofBSMC stimulated for 24 hwith IBOP,10−9–10−7M, in the presence versus absence of IL-4,0.1–10 ng/ml. Results are means+S.E.M. n=4 (CCL11) or n=5 (CCL26) using BSMC from4–5 different donors. (C) Time course of CCL11 synthesis from unstimulated BSMC(unfilled circles) versus BSMC stimulated with IBOP, 10−8 M and IL-4, 10 ng/ml (filledcircles). Results are means+S.E.M. n=5 using BSMC from 5 different donors. ⁎⁎Pb0.01versus time “0”.
Fig. 4. CCL11 release from BSMC stimulated for 24 h with (A) IL-4, 10 ng/ml, (B) IBOP,10−8M+IL-4, or (C) IL-1, 1 ng/ml+IL-4, in the presence or absence of 0.1 to 3 µM of aprostanoid TP receptor antagonist, AA-2414. Results are means+S.E.M. n=3 using BSMCfrom3different donors. ⁎Pb0.05, ⁎⁎Pb0.01 versusmeasurementswith 0 µMof AA-2414.
136 Y. Suzuki et al. / European Journal of Pharmacology 600 (2008) 133–139
supernatant of BSMC stimulated with IL-4, 10 ng/ml but IBOPexhibited no effects on CCL26 release (Fig. 3B). Concentration ofCCL11 in the supernatant of BSMCwas elevated at 4 h after stimulationwith IBOP/IL-4, and further increased over the 24-h period (Fig. 3C).IBOP (10−9–10−7 M) also increased the synthesis of CCL11 induced by10 ng/ml of IL-13 in a concentration-dependent manner (Pb0.01, datanot shown). There was no release of CCL7, CCL8, or CCL13 from eitherunstimulated or IBOP/IL-4-stimulated BSMC.
AA-2414, a prostanoid TP receptor antagonist, in a concentration of0.3–3 µM, inhibited the IBOP-induced enhancement of CCL11 releasefrom IL-4-stimulated BSMC (n=3, Pb0.05–0.01, Fig. 4B). AA-2414,however, did not suppress the effects of IL-4 (10 ng/ml) alone, or IL-4(10 ng/ml) in combination with IL-1 (1 ng/ml) (Fig. 4A and C),confirming that the IBOP-induced release of CCL11 was dependent onthe activation of prostanoid TP receptors.
Based on the expression pattern of the CCR3 ligands, the mainfactor with eosinophil chemotactic activity released from IBOP/IL-4-stimulated BSMC appeared to be CCL11. We further confirmed the roleof CCL11 using a CCL11-neutralizing antibody. The specificity of thisantibody was confirmed by the fact that it suppressed eosinophilmobilization induced by recombinant human CCL11, but not thatinduced by platelet activating factor (Fig. 5). Pretreatment of theconditioned medium from IBOP/IL-4-stimulated BSMC with this anti-CCL11 antibody decreased the eosinophil chemotactic activity by

Fig. 5. Effect of an anti-CCL11 neutralizing antibody on eosinophil chemotactic activityreleased from BSMC stimulated with IL-4, 10 ng/ml, and IBOP, 10−8 M. The conditionedmedium, recombinant human CCL11 (100 ng/ml), or platelet activating factor (10 nM)was pretreated with control IgG1 (control IgG, 10 μg/ml) or an anti-CCL11 neutralizingantibody, 10 μg/ml, at 37 °C, for 1 h. Results are means+S.E.M. n=6 using differentcombination with BSMC from 3 donors and eosinophils from 2 donors. ⁎Pb0.05 versuscontrol IgG1.
Fig. 7. STAT-6 phosphorylation in BSMC. A) Cell-based ELISA of phosphorylated STAT-6 (p-STAT6) and total STAT-6 in BSMC stimulatedwith I-BOP (10−8M) and/or IL-4 (10 ng/ml) for1–4 h. The mean value of the p-STAT6/total STAT6 ratio in the “unstimulated” BSMC wasdefined as 1.0. Results aremeans+S.E.M.n=3 using BSMC from3different donors. ⁎Pb0.05versus “unstimulated” BSMC. B)Western blot analysis of phosphorylated and total STAT-6.BSMC pretreated with IBOP, 10−8 M for 24 h, were stimulated with medium or IL-4, 10 ng/ml for 15 min. Representative data from 3 separate experiments.
137Y. Suzuki et al. / European Journal of Pharmacology 600 (2008) 133–139
N85% (n=6, Pb0.05, Fig. 5). This indicated that the functionaleosinophil chemotactic activity derived from BSMC co-stimulatedwith prostanoid TP receptor agonists and Th2 cytokines were mostlymediated by CCL11.
In order to clarify the mechanism(s) how IBOP had enhanced IL-4-induced CCL11 expression in BSMC, we pretreated the cells with IBOP,10−8 M, for 24 h, and then examined the expression of IL-4 receptor-αon cellular surface, or the activation of its downstream signalingmolecule, STAT-6. A flow cytometric analysis confirmed that the IL-4receptor-α was expressed on the surface of BSMC constitutively,though was not affected by 24-h incubation with IBOP (Fig. 6A). IL-4treatment (10 ng/ml, 1–4 h) significantly induced the phosphorylationof STAT-6 (Pb0.05), but there was no additional or synergistic effect bythe co-stimulationwith IBOP (10−8 M) (Fig. 7A). Longer pre-treatmentwith IBOP (24 h) also failed to modulate the IL-4-induced phosphor-ylation of STAT-6 (Fig. 7B). These observations indicate that prostanoidTP receptor agonism enhanced the CCL11 gene expression induced byIL-4, without modifying the IL-4 receptor/STAT6 pathway.
4. Discussion
This study discovered a mechanism by which activated prostanoidTP receptors in the airways of asthmatics might induce the recruit-ment of eosinophils. The stimulation of prostanoid TP receptors
Fig. 6. Flow cytometric analysis of IL-4 receptor-α expression on the surface of BSMCunstimulated or stimulated for 24 h with IBOP, 10−8 M. Representative data from 3separate experiments.
caused the de novo synthesis and release of CCL11, a potenteosinophilic chemoattractant, in bronchial smooth muscles in a Th2-dominant microenvironment.
PGD2, one of themajormediators released from activatedmast cells,has beendemonstrated to act as a direct chemoattractant for eosinophilsthrough G-protein-coupled receptors, chemoattractant receptor homo-logous molecule expressed on Th2 (CRTH2) (Hirai et al., 2001; Nagataet al., 1999; Shiraishi et al., 2005). Our findings indicate anothermechanism by which PGD2 exacerbates the eosinophilic airwayinflammation, activation of prostanoid TP receptors on bronchialsmooth muscle cells. This alternative pathway could be especiallyimportant in the asthmatic airways, where mast cells are abundantlypresent in the bronchial smooth muscle layers (Brightling et al., 2002).
In the study by Hoshino et al. (1999), treatment with AA-2414suppressed the expression of CCL5 and CCL11 in the airways,suggesting the existence of an association between prostanoid TPreceptors and chemokines. However, whether the activation of thesereceptors induces directly the expression of chemokines was mostlyunknown, except for a report showing the prostanoid TP receptor-mediated induction of CCL2/MCP-1 in vascular endothelial cells(Ishizuka et al., 2000). This study showed that prostanoid TPreceptor-mediated signals do not directly induce the expression ofCC chemokines, but enhance the IL-4/IL-13-induced expression ofCCL11 in BSMC. Interestingly, the prostanoid TP receptor-mediatedinduction is strictly limited to CCL11, and the expression of other CCR3ligands including CCL5 and CCL26 was unchanged.
Like IBOP, cytokines and other molecules, including oncostatin M,transforming growth factor (TGF)-β, IL-1, and double-stranded RNA,induce the synthesis of CCL11 in BSMC in synergy with Th2 cytokines,IL-4 and IL-13 (Faffe et al., 2005; Hirst et al., 2002; Niimi et al., 2007;Zuyderduyn et al., 2004). Oncostatin M can, by itself, induce the

138 Y. Suzuki et al. / European Journal of Pharmacology 600 (2008) 133–139
expression of CCL11 in BSMC via the activation of STAT3, and furtherenhance the IL-4/IL-13-induced synthesis of CCL11 by increasing thenumber of IL-4 receptor-α (Faffe et al., 2005). Double-stranded RNAalso increases the expression of IL-4 receptor in bronchial epithelialcells (Tsuji et al., 2005). In the present study, however, IBOPmodulated neither the expression of the IL-4 receptor-α on thesurface of BSMC nor the IL-4 receptor-mediated phosphorylation ofSTAT-6. Furthermore, the levels of CCL26, which was also inducible inan IL-4-dependent manner in BSMC, were unchanged by stimulationwith IBOP. Thus, it is unlikely that prostanoid TP receptor signalsincrease the expression of CCL11 in BSMC by modulating theexpression of IL-4 receptor.
TGF-β alone did not induce the expression of CCL11 or CCL26 inBSMC, however, in combination with IL-4 or IL-13, it increased theproduction of CCL11, and inhibited the production of Th2 cytokine-induced CCL26 (Zuyderduyn et al., 2004). TGF-β also increased theexpression of IL-13-induced CCL11 in fibroblasts, without changingthe expression of IL-4 receptor-α or phosphorylation of STAT-6(Wenzel et al., 2002), as in IBOP-stimulated BSMC. The differentialmodulation of CCL11 and CCL26, and the lack of direct STAT6activation by TGF-β are close to that observed in the present studywith IBOP. Thus, a common signaling pathway might exist down-stream of the TGF-β and prostanoid TP receptors, which modulatesthe expression of CCL11 in BSMC.
One of the possible signaling molecules shared by the TGF-β andprostanoid TP receptors are mitogen-activated protein (MAP) kinases.TGF-β induces the proliferation of BSMC via MAP kinase-dependent,Smad-independent pathways (Xie et al., 2007). Prostanoid TPreceptor, predominantly coupled to G(i/o) proteins, also activates thep42/p44 kinase pathway in BSMC (Citro et al., 2005) and in vascularsmooth muscle cells (Gallet et al., 2003). However, it is difficult todemonstrate that MAP kinases play a role in the effect of IBOPamplifying the IL-4-induced CCL11 synthesis, because IL-4 itselfinduces CCL11 synthesis in a MAP kinase-dependent manner;blockade of MAP kinases either with U0126 or SB202190 suppressedthe release of CCL11 from IL-4-stimulated BSMC (our unpublishedobservations), as other researchers previously reported (Hirst et al.,2002; Moore et al., 2002).
In conclusion,we have shown that a prostanoid TP receptor agonist,IBOP, in synergy with IL-4, induces the release of CCL11, a functionaleosinophil chemoattractant, from human BSMC, an effect that wasprevented by AA2414, a prostanoid TP receptor antagonist. Thissuggests that prostanoid TP receptor antagonism might mitigate theeosinophilic inflammation in Th2-cytokine dominant asthmatic air-ways, by inhibiting the synthesis and release of CCL11 from BSMC.
Acknowledgments
AA-2414 was a generous gift from Takeda Pharmaceutical Co.(Osaka, Japan). The authors thank Miyuki Yamamoto for her excellenttechnical assistance.
References
Allen, I.C., Hartney, J.M., Coffman, T.M., Penn, R.B., Wess, J., Koller, B.H., 2006.Thromboxane A2 induces airway constriction through an M3muscarinic acetylcho-line receptor-dependent mechanism. Am. J. Physiol., Lung Cell. Mol. Physiol. 290,L526–533.
Brightling, C.E., Bradding, P., Symon, F.A., Holgate, S.T., Wardlaw, A.J., Pavord, I.D., 2002.Mast-cell infiltration of airway smooth muscle in asthma. N. Engl. J. Med. 346,1699–1705.
Capra, V., Habib, A., Accomazzo, M.R., Ravasi, S., Citro, S., Levy-Toledano, S., Nicosia, S.,Rovati, G.E., 2003. Thromboxane prostanoid receptor in human airway smoothmuscle cells: a relevant role in proliferation. Eur. J. Pharmacol. 474, 149–159.
Citro, S., Ravasi, S., Rovati, G.E., Capra, V., 2005. Thromboxane prostanoid receptorsignals through Gi protein to rapidly activate extracellular signal-regulated kinasein human airways. Am. J. Respir. Cell Mol. Biol. 32, 326–333.
Coleman, R.A., Sheldrick, R.L., 1989. Prostanoid-induced contraction of human bronchialsmooth muscle is mediated by TP-receptors. Br. J. Pharmacol. 96, 688–692.
Faffe, D.S., Flynt, L., Mellema, M., Moore, P.E., Silverman, E.S., Subramaniam, V., Jones, M.R.,Mizgerd, J.P., Whitehead, T., Imrich, A., Panettieri Jr., R.A., Shore, S.A., 2005. OncostatinM causes eotaxin-1 release from airway smooth muscle: synergy with IL-4 and IL-13.J. Allergy Clin. Immunol. 115, 514–520.
Fukuoka, T., Miyake, S., Umino, T., Inase, N., Tojo, N., Yoshizawa, Y., 2003. The effect ofseratrodast on eosinophil cationic protein and symptoms in asthmatics. J. Asthma40, 257–264.
Gallet, C., Blaie, S., Levy-Toledano, S., Habib, A., 2003. Epidermal-growth-factor receptorand metalloproteinases mediate thromboxane A2-dependent extracellular-signal-regulated kinase activation. Biochem. J. 371, 733–742.
Hirai, H., Tanaka, K., Yoshie, O., Ogawa, K., Kenmotsu, K., Takamori, Y., Ichimasa, M.,Sugamura, K., Nakamura, M., Takano, S., Nagata, K., 2001. Prostaglandin D2
selectively induces chemotaxis in T helper type 2 cells, eosinophils, and basophilsvia seven-transmembrane receptor CRTH2. J. Exp. Med. 193, 255–261.
Hirst, S.J., Hallsworth, M.P., Peng, Q., Lee, T.H., 2002. Selective induction of eotaxin releaseby interleukin-13 or interleukin-4 in human airway smoothmuscle cells is synergisticwith interleukin-1β and is mediated by the interleukin-4 receptor α-chain. Am.J. Respir. Crit. Care Med. 165, 1161–1171.
Hoshino, M., Sim, J., Shimizu, K., Nakayama, H., Koya, A., 1999. Effect of AA-2414, athromboxane A2 receptor antagonist, on airway inflammation in subjects withasthma. J. Allergy Clin. Immunol. 103, 1054–1061.
Huszar, E., Szabo, Z., Jakab, A., Barta, I., Herjavecz, I., Horvath, I., 2005. Comparativemeasurement of thromboxane A2 metabolites in exhaled breath condensate bydifferent immunoassays. Inflamm. Res. 54, 350–355.
Ishizuka, T., Sawada, S., Sugama, K., Kurita, A., 2000. Thromboxane A2 (TXA2 receptorblockade suppresses monocyte chemoattractant protein-1 (MCP-1) expression bystimulated vascular endothelial cells. Clin. Exp. Immunol. 120, 71–78.
Itoh, K., Mukaiyama, O., Satoh, Y., Yamaguchi, T., Iizuka, Y., 1993. Effects of CS-518,a thromboxane synthase inhibitor, on eosinophil function. Eur. J. Pharmacol. 239,159–169.
Itoh, K., Takahashi, E., Mukaiyama, O., Satoh, Y., Yamaguchi, T., 1996. Effects of athromboxane synthase inhibitor (CS-518) on the eosinophil-dependent lateasthmatic response and airway hyperresponsiveness in guinea pigs. Int. Arch.Allergy Immunol. 109, 79–85.
Janssen, L.J., Premji, M., Netherton, S., Catalli, A., Cox, G., Keshavjee, S., Crankshaw, D.J.,2000. Excitatory and inhibitory actions of isoprostanes in human and canine airwaysmooth muscle. J. Pharmacol. Exp. Ther. 295, 506–511.
Kaplan, A.P., 2001. Chemokines, chemokine receptors and allergy. Int. Arch. AllergyImmunol. 124, 423–431.
Kumlin, M., Dahlen, B., Bjorck, T., Zetterstrom, O., Granstrom, E., Dahlen, S.E., 1992.Urinary excretion of leukotriene E4 and 11-dehydro-thromboxane B2 in response tobronchial provocations with allergen, aspirin, leukotriene D4, and histamine inasthmatics. Am. Rev. Respir. Dis. 146, 96–103.
Martin, C., Ullrich, V., Uhlig, S., 2000. Effects of the thromboxane receptor agonistU46619 and endothelin-1 on large and small airways. Eur. Respir. J. 16, 316–323.
Matsumoto, T., Ashida, Y., Tsukuda, R., 1994. Pharmacological modulation of immediateand late airway response and leukocyte infiltration in the guinea pig. J. Pharmacol.Exp. Ther. 269, 1236–1244.
Montuschi, P., Barnes, P.J., 2002. Exhaled leukotrienes and prostaglandins in asthma.J. Allergy Clin. Immunol. 109, 615–620.
Moore, P.E., Church, T.L., Chism, D.D., Panettieri Jr., R.A., Shore, S.A., 2002. IL-13 and IL-4cause eotaxin release in human airway smooth muscle cells: a role for ERK. Am.J. Physiol., Lung Cell. Mol. Physiol. 282, L847–853.
Nagai, H., Teramachi, H., Tuchiya, T., 2006. Recent advances in the development of anti-allergic drugs. Allergol. Intern. 55, 35–42.
Nagata, K., Hirai, H., Tanaka, K., Ogawa, K., Aso, T., Sugamura, K., Nakamura, M., Takano, S.,1999. CRTH2, an orphan receptor of T-helper-2-cells, is expressed on basophils andeosinophils and responds to mast cell-derived factor(s). FEBS Lett. 459, 195–199.
Narita, S., Asakura, K., Shirasaki, H., Kojima, T., Kataura, A., 1995. Studies onthromboxane B2 level in nasal lavage fluid after antigen challenge in patientswith allergic rhinitis and guinea pig models. Arerugi 44, 1159–1166.
Niimi, K., Asano, K., Shiraishi, Y., Nakajima, T., Wakaki, M., Kagyo, J., Takihara, T., Suzuki, Y.,Fukunaga, K., Shiomi, T., Oguma, T., Sayama, K., Yamaguchi, K., Natori, Y., Matsumoto,M., Seya, T., Yamaya, M., Ishizaka, A., 2007. TLR3-mediated synthesis and release ofeotaxin-1/CCL11 from human bronchial smoothmuscle cells stimulatedwith double-stranded RNA. J. Immunol. 178, 489–495.
Richter, M., Cloutier, S., Sirois, P., 2007. Endothelin, PAF and thromboxane A2 in allergicpulmonary hyperreactivity in mice. Prostaglandins, Leukot. Essent. Fat. Acids 76,299–308.
Rolin, S., Masereel, B., Dogne, J.M., 2006. Prostanoids as pharmacological targets inCOPD and asthma. Eur. J. Pharmacol. 533, 89–100.
Shi, H., Yokoyama, A., Kohno, N., Hirasawa, Y., Kondo, K., Sakai, K., Hiwada, K., 1998.Effect of thromboxane A2 inhibitors on allergic pulmonary inflammation in mice.Eur. Respir. J. 11, 624–629.
Shiraishi, Y., Asano, K., Nakajima, T., Oguma, T., Suzuki, Y., Shiomi, T., Sayama, K., Niimi,K., Wakaki, M., Kagyo, J., Ikeda, E., Hirai, H., Yamaguchi, K., Ishizaka, A., 2005.Prostaglandin D2-induced eosinophilic airway inflammation is mediated by CRTH2receptor. J. Pharmacol. Exp. Ther. 312, 954–960.
Suzuki, Y., Asano, K., Shiraishi, Y., Oguma, T., Shiomi, T., Fukunaga, K., Nakajima, T., Niimi,K., Yamaguchi, K., Ishizaka, A., 2004. Human bronchial smooth muscle cellproliferation via thromboxane A2 receptor. Prostaglandins, Leukot. Essent. Fat.Acids 71, 375–382.
Tsuji, K., Yamamoto, S., Ou, W., Nishi, N., Kobayashi, I., Zaitsu, M., Muro, E., Sadakane, Y.,Ichimaru, T., Hamasaki, Y., 2005. dsRNA enhances eotaxin-3 production throughinterleukin-4 receptor upregulation in airway epithelial cells. Eur. Respir. J. 26,795–803.

139Y. Suzuki et al. / European Journal of Pharmacology 600 (2008) 133–139
Wenzel, S.E., Trudeau, J.B., Barnes, S., Zhou, X., Cundall, M., Westcott, J.Y., McCord, K.,Chu, H.W., 2002. TGF-β and IL-13 synergistically increase eotaxin-1 production inhuman airway fibroblasts. J. Immunol. 169, 4613–4619.
Xiang, A., Uchida, Y., Nomura, A., Iijima, H., Sakamoto, T., Ishii, Y., Morishima, Y.,Masuyama, K., Zhang, M., Hirano, K., Sekizawa, K., 2002. Involvement ofthromboxane A2 in airway mucous cells in asthma-related cough. J. Appl. Physiol.92, 763–770.
Xie, S., Sukkar, M.B., Issa, R., Khorasani, N.M., Chung, K.F., 2007. Mechanisms of inductionof airway smooth muscle hyperplasia by transforming growth factor-β. Am.J. Physiol., Lung Cell. Mol. Physiol. 293, L245–253.
Zuyderduyn, S., Hiemstra, P.S., Rabe, K.F., 2004. TGF-beta differentially regulates TH2cytokine-induced eotaxin and eotaxin-3 release by human airway smooth musclecells. J. Allergy Clin. Immunol. 114, 791–798.