
Toxin Production by Bacillus cereus Dairy Isolates in Milk...
6
Vol. 55, No. 10 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, OCt. 1989, p. 2595-2600 0099-2240/89/102595-06$02.00/0 Copyright X 1989, American Society for Microbiology Toxin Production by Bacillus cereus Dairy Isolates in Milk at Low Temperatures ANDERS CHRISTIANSSON,1 A. SATYANARAYAN NAIDU,2 INGRID NILSSON,3 TORKEL WADSTROM,3 AND HANS-ERIK PETTERSSON1* Swedish Dairies' Association, Box 205, S-201 22 Malmo1; Department of Medical Microbiology, Malmo General Hospital, University of Lund, S-214 01 Malmo2; and Department of Medical Microbiology, University of Lund, S-223 62 Lund3; Sweden Received 6 February 1989/Accepted 1 June 1989 A total of 136 strains of Bacillus cereus isolated from milk and cream were evaluated for toxin production based on HeLa S3, Vero, and human embryonic lung (HEL) cell cytotoxicity in vitro. HEL cell monolayers were more susceptible than the other two cell lines. The percentage of isolates exhibiting HEL cytotoxicity was similar (43.0 and 48.4%) when the strains were grown in brain heart infusion broth containing 0.1% glucose (BHIG) at 7 and 24 h, respectively, at 30°C. In milk, only 21.8% of isolates showed HEL cytotoxicity at 7 h, and the number increased significantly to 73.2% at 24 h at 30°C. Further, 102 toxin-positive isolates were acclimatized to grow at 8°C in milk. Ninety-four (92.2%) of the strains produced HEL cytotoxicity of various degrees with no strict correlation to bacterial cell numbers and also elicited vascular permeability reaction in rabbit skin. Under aerated growth conditions (agitation, 200 rpm) B. cereus elicited cytotoxicity in BHIG and in milk at temperatures of 30, 15, and 8°C. However, in nonaerated (stagnant) cultures toxin production was diminished (BHIG) or completely lost (milk) at all temperatures. Toxin production at 8°C was evaluated in two different types of commercial cardboard milk packages by inoculation with a potent toxigenic dairy isolate. No detectable HEL cytotoxicity was observed in milk in any of the packages either at stagnant conditions or during mechanical shaking. However, the same strain produced cytotoxin in whipped cream at 8°C. Bacillus cereus is a widely distributed organism in the environment and undoubtedly the most important of the aerobic sporeforming species found in milk (1, 5). B. cereus is one of the organisms responsible for the spoilage of pasteurized milk and cream referred to as "sweet curdling" (23) and "bitty cream" (3). B. cereus is generally the organism that sets the limits for the keeping quality of Swedish consumer milk, particularly during the summer season. Despite low initial numbers of B. cereus in raw milk, the legal limit of 103/ml on the day labeled "best before" is often easily reached due to the presence of psychrotrophic strains that grow rapidly at 8°C. This temperature is the highest legally permitted for the storage of milk, and 104 B. cereus cells per ml is the ultimate limit of expiry. Thus, from public health and economic points of view, the monitoring and control of B. cereus are of high importance for the dairy industry. B. cereus has been implicated in two distinct forms of food-borne illnesses: a delayed-onset (8 to 16 h) diarrheal syndrome characterized by symptoms of enterotoxicosis, abdominal cramps, and rectal tenesmus, usually resolving within 24 h, and a rapid-onset (1 to 5 h) emetic syndrome with nausea, vomiting, and occasionally diarrhea (8-10, 19, 20). Two different toxins were recognized with these ill- nesses. First is an unstable enterotoxin (30) with a molecular weight of approximately 50,000 and a pl of 4.9 (34). More recent studies have indicated that this toxin consists of several subunits slightly differing in molecular weights and pI (13, 32). Second, a heat-tolerant emetic toxin with a molecular weight of <5,000 has also been identified (18, 34). The pathologic role, if any, of cereolysin, protease(s), and phospholipase C complex in B. cereus is still not known (13). * Corresponding author. Laboratory conditions for optimum production of B. cereus toxins have been defined (6). Toxin production is strongly dependent on culture medium and bacterial growth condi- tions (6, 29, 31). The production of emetic toxin seems to occur during sporulation, but the relation between sporula- tion and enterotoxin production is unclear (33). Sporeforming and psychrotrophic properties enable B. cereus to survive pasteurization as well as to grow in milk at refrigerated storage conditions (5). Despite a high worldwide occurrence of B. cereus in milk, surprisingly few reports on food poisoning have been reported (11, 14, 17, 33, 36). The toxin-producing ability of B. cereus in milk under different dairy processing and storage conditions is not known. There- fore, we have investigated for the production of cytotoxins, hemolysins, and amylases by strains of B. cereus grown in milk under different conditions. Selected psychrotrophic strains were further tested for toxin production by vascular permeability reaction (VPR) tests. MATERIALS AND METHODS Bacterial strains. Samples of milk and cream stored at 8°C for 6 to 7 days were analyzed for the presence of B. cereus. Strains showing typical B. cereus colony morphology and hemolysis on agar plates containing 5% bovine blood and 0.001% polymyxin B sulfate (Sigma Chemical Co., St. Louis, Mo.) were isolated. Species identity was confirmed by mannitol fermentation test and egg yolk reaction on mannitol egg yolk phenol red agar (21) and by microscopic examination. Selected strains were further confirmed by API 50CH and API 20E (15). Finally, a total of 136 isolates were identified as B. cereus. These strains originated from milk and cream produced at 37 different Swedish dairy plants. Laboratory reference strains for enterotoxin production 2595 on June 20, 2018 by guest http://aem.asm.org/ Downloaded from
-
Upload
truongcong -
Category
Documents
-
view
215 -
download
0
Transcript of Toxin Production by Bacillus cereus Dairy Isolates in Milk...

Vol. 55, No. 10APPLIED AND ENVIRONMENTAL MICROBIOLOGY, OCt. 1989, p. 2595-26000099-2240/89/102595-06$02.00/0Copyright X 1989, American Society for Microbiology
Toxin Production by Bacillus cereus Dairy Isolates in Milkat Low Temperatures
ANDERS CHRISTIANSSON,1 A. SATYANARAYAN NAIDU,2 INGRID NILSSON,3TORKEL WADSTROM,3 AND HANS-ERIK PETTERSSON1*
Swedish Dairies' Association, Box 205, S-201 22 Malmo1; Department of Medical Microbiology,Malmo General Hospital, University of Lund, S-214 01 Malmo2; and Department of
Medical Microbiology, University of Lund, S-223 62 Lund3; Sweden
Received 6 February 1989/Accepted 1 June 1989
A total of 136 strains of Bacillus cereus isolated from milk and cream were evaluated for toxin productionbased on HeLa S3, Vero, and human embryonic lung (HEL) cell cytotoxicity in vitro. HEL cell monolayerswere more susceptible than the other two cell lines. The percentage of isolates exhibiting HEL cytotoxicity wassimilar (43.0 and 48.4%) when the strains were grown in brain heart infusion broth containing 0.1% glucose(BHIG) at 7 and 24 h, respectively, at 30°C. In milk, only 21.8% of isolates showed HEL cytotoxicity at 7 h,and the number increased significantly to 73.2% at 24 h at 30°C. Further, 102 toxin-positive isolates wereacclimatized to grow at 8°C in milk. Ninety-four (92.2%) of the strains produced HEL cytotoxicity of variousdegrees with no strict correlation to bacterial cell numbers and also elicited vascular permeability reaction inrabbit skin. Under aerated growth conditions (agitation, 200 rpm) B. cereus elicited cytotoxicity in BHIG andin milk at temperatures of 30, 15, and 8°C. However, in nonaerated (stagnant) cultures toxin production wasdiminished (BHIG) or completely lost (milk) at all temperatures. Toxin production at 8°C was evaluated in twodifferent types of commercial cardboard milk packages by inoculation with a potent toxigenic dairy isolate. Nodetectable HEL cytotoxicity was observed in milk in any of the packages either at stagnant conditions or duringmechanical shaking. However, the same strain produced cytotoxin in whipped cream at 8°C.
Bacillus cereus is a widely distributed organism in theenvironment and undoubtedly the most important of theaerobic sporeforming species found in milk (1, 5). B. cereusis one of the organisms responsible for the spoilage ofpasteurized milk and cream referred to as "sweet curdling"(23) and "bitty cream" (3). B. cereus is generally theorganism that sets the limits for the keeping quality ofSwedish consumer milk, particularly during the summerseason. Despite low initial numbers of B. cereus in raw milk,the legal limit of 103/ml on the day labeled "best before" isoften easily reached due to the presence of psychrotrophicstrains that grow rapidly at 8°C. This temperature is thehighest legally permitted for the storage of milk, and 104 B.cereus cells per ml is the ultimate limit of expiry. Thus, frompublic health and economic points of view, the monitoringand control of B. cereus are of high importance for the dairyindustry.
B. cereus has been implicated in two distinct forms offood-borne illnesses: a delayed-onset (8 to 16 h) diarrhealsyndrome characterized by symptoms of enterotoxicosis,abdominal cramps, and rectal tenesmus, usually resolvingwithin 24 h, and a rapid-onset (1 to 5 h) emetic syndromewith nausea, vomiting, and occasionally diarrhea (8-10, 19,20). Two different toxins were recognized with these ill-nesses. First is an unstable enterotoxin (30) with a molecularweight of approximately 50,000 and a pl of 4.9 (34). Morerecent studies have indicated that this toxin consists ofseveral subunits slightly differing in molecular weights andpI (13, 32). Second, a heat-tolerant emetic toxin with amolecular weight of <5,000 has also been identified (18, 34).The pathologic role, if any, of cereolysin, protease(s), andphospholipase C complex in B. cereus is still not known (13).
* Corresponding author.
Laboratory conditions for optimum production of B. cereustoxins have been defined (6). Toxin production is stronglydependent on culture medium and bacterial growth condi-tions (6, 29, 31). The production of emetic toxin seems tooccur during sporulation, but the relation between sporula-tion and enterotoxin production is unclear (33).
Sporeforming and psychrotrophic properties enable B.cereus to survive pasteurization as well as to grow in milk atrefrigerated storage conditions (5). Despite a high worldwideoccurrence of B. cereus in milk, surprisingly few reports onfood poisoning have been reported (11, 14, 17, 33, 36). Thetoxin-producing ability of B. cereus in milk under differentdairy processing and storage conditions is not known. There-fore, we have investigated for the production of cytotoxins,hemolysins, and amylases by strains of B. cereus grown inmilk under different conditions. Selected psychrotrophicstrains were further tested for toxin production by vascularpermeability reaction (VPR) tests.
MATERIALS AND METHODSBacterial strains. Samples of milk and cream stored at 8°C
for 6 to 7 days were analyzed for the presence of B. cereus.Strains showing typical B. cereus colony morphology andhemolysis on agar plates containing 5% bovine blood and0.001% polymyxin B sulfate (Sigma Chemical Co., St.Louis, Mo.) were isolated. Species identity was confirmedby mannitol fermentation test and egg yolk reaction onmannitol egg yolk phenol red agar (21) and by microscopicexamination. Selected strains were further confirmed by API50CH and API 20E (15). Finally, a total of 136 isolates wereidentified as B. cereus. These strains originated from milkand cream produced at 37 different Swedish dairy plants.Laboratory reference strains for enterotoxin production
2595
on June 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2596 CHRISTIANSSON ET AL.
(4433/73, 4581B/76, 4096/73, ATCC-14579, and NCDO-1771)and a reference strain for emetic toxin production (4810/72)were also included in this study. The origin of the referencestrains is described elsewhere (18, 34, 35).Chemicals and media. Blood agar base no. 2 was from
Oxoid (London, England). Brain heart infusion (BHI) brothand Bacto-Peptone were purchased from Difco. Sodiumcaseinate was purchased from U.S. Biochemical Corpora-tion, Cleveland, Ohio. BHI supplemented with 0.1% glucose(pH 8.0) (BHIG) was prepared according to Glatz andGoepfert (6). Milk (9.4% dry matter) was reconstituted fromskim milk powder obtained from Scandmilk AB, Stockholm,Sweden, and was sterilized at 110°C for 15 min. Tissue cellculture medium was prepared using minimal essential me-dium with 10% fetal calf serum and was supplemented with1% L-glutamine and 0.1% gentamicin, all from Flow Labo-ratories, Herts, England. All chemicals used in the prepara-tion of buffer solutions were of analytical grade.Media and growth conditions. Strains were grown static in
skim milk overnight at 30 and 15°C or for 7 days at 8°C. A 1%inoculum of this culture was added to 10 to 15 ml of sterileskim milk in 150-ml Erlenmeyer flasks and cultivated at 30,15, and 8°C with an agitation of 200 rpm. After centrifugationat 17,000 rpm for 15 min, the cell-free liquid between the fat(top phase) and casein-containing bacterial cell mass (bottomphase) was aspirated aseptically and tested for the presenceof toxin. Toxin production was also tested in BHIG accord-ing to Glatz and Goepfert (6). In both culture conditions,cell-free supernatants were collected at different time inter-vals ranging from 5 to 96 h of growth and tested.B. cereus BC-131 was further tested for toxin production
in pasteurized milk in two different types of commercialcardboard packages (1-liter Purepak and 1-liter Tetra Brik).The strain was grown at 8°C in sterile skim milk as describedabove, and an inoculum giving a final concentration of 4 x104 to 6 x 104 cells per ml was injected into the milkpackages. The packages were stored at 8°C under threedifferent conditions: (i) sealed and stagnant, (ii) sealed withshaking at 300 rpm, and (iii) opened (half emptied) andstagnant. Toxin production by the same strain was alsotested in whipped cream at 8°C and in skim milk withaeration at the same temperature.
Hemolytic activity. The hemolytic activity of 50 B. cereusBHIG culture supernatants were tested by well diffusiontechnique on blood agar plates containing human, sheep, andrabbit washed erythrocytes. Based on the zone size, hemol-ysis results were scored as +++, ++, +, +/-, and nega-tive.
Starch and salicin hydrolysis. The ability of B. cereusstrains to hydrolyze starch was tested on nutrient agar platescontaining 0.95% soluble starch. After incubation at 30°C for24 h, plates were flooded with 3% Lugol's iodine solution.The test was considered positive when a clear zone appearedaround the bacterial growth (24). Limited hydrolysis beneaththe bacterial growth was scored as negative. The salicinfermentation test was performed on a basal ammonium saltmedium containing yeast extract and 0.5% salicin withbromothymol blue as a pH indicator (24). Upon acid fermen-tation, yellow color was developed within 7 days at 30°C.
Cytotoxicity. The cytotoxicity of B. cereus culture super-natants from skim milk and BHIG was examined on HeLaS3, Vero, and human embryonic lung (HEL; also known asMRC-5) cell monolayers. Supernatants were diluted (two-fold from 1:5 to 1:640) in incomplete tissue cell culturemedium, and 100 RId of each dilution in duplicate was addedto microtiter wells. Each well contained 1.4 x 105 HeLa, 3.8
TABLE 1. Hemolysin production and carbohydrate utilizationby strains of B. cereus isolated from milk
No. positive/no. tested (%)
HemolysisaHuman erythrocytes ....................................... 14/50 (28.0)Sheep erythrocytes ........................................ 11/50 (22.0)Rabbit erythrocytes ....................................... 13/50 (26.0)
Carbohydrate utilizationStarch hydrolysisb ........................................ 129/136 (94.9)Salicin fermentation ....................................... 97/136 (71.3)a Cell-free culture supernatants of B. cereus grown in BHIG at 30°C for 5 h
were tested for hemolytic activity on washed erythrocytes obtained fromdifferent sources.
b Ability of the test isolate to hydrolyze soluble starch incorporated in agarmedium at 30'C after 24 h.
Results were scored after 7 days of incubation at 30°C.
x 105 Vero, or 1.5 x 105 HEL cells. Microtiter platescontaining HeLa, Vero, or HEL cells were placed in acarbon dioxide incubator (7% C02) at 37°C for 12 or 48 h.Plates were washed with 0.7 M phosphate-buffered saline(pH 7.2). HeLa cells were fixed with 2% Formalin for 1 min,and Vero and HEL cells were fixed with methanol for 10min. Cells were stained with 0.13% crystal violet (Merck,Darmstadt, Federal Republic of Germany) solution for 20min. After three washings with phosphate-buffered saline,the stained cells retained in the wells were dissolved in 200p.1 of 1% sodium dodecyl sulfate solution. The absorbancewas measured on a Titertek Multiscan (Flow Laboratories,Svenska AB, Solna, Sweden) at 595 nm. Sterile BHIG brothand skim milk were used as negative controls.VPR. The vascular permeability reaction (VPR) test was
performed according to Glatz et al. (7). Skim milk and BHIGcell-free culture supernatants (0.1 ml) were injected intrader-mally into rabbits weighing 2.5 to 3.0 kg. After 3 h, 4 ml of2% Evans blue dye solution was injected intravenously.Measurements of perpendicular diameters of the zones oflight and dark blue, and necrosis when present, were madeafter 1 h. Total reaction was calculated as the sum of themean radii of the blue zones.
RESULTS
Hemolytic pattern and carbohydrate utilization. Fifty B.cereus dairy isolates were grown in BHIG according tostandard procedures, and the supernatants were tested forhemolytic activity on human, sheep, and rabbit erythrocytes(Table 1). Hemolysin activity was found among 22 to 28% ofthe isolates. Human erythrocytes were more susceptible,with stronger lytic reactions than to sheep and rabbit.Hemolysin production showed no strict correlation with cellmonolayer toxicity assays. In carbohydrate utilization tests,B. cereus reference strain F-4810/72, positive for emetictoxin, was unable to hydrolyze starch or ferment salicin,whereas the diarrheagenic reference strains were able toutilize these compounds. Of 136 dairy isolates tested, only 7(5.1%) did not hydrolyze starch, whereas 39 (28.7%) did notferment salicin.
Screening of toxin-producing strains of B. cereus isolatedfrom milk. A total of 136 B. cereus dairy isolates and sixreference strains (positive for toxin production) were ini-tially screened for toxin production under optimal cultureconditions as described in the literature (i.e., growth inBHIG at 30°C with aeration and a harvest time at 5 h). B.cereus NCDO-1771 produced high levels of toxin (HeLa
APPL. ENVIRON. MICROBIOL.
on June 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

B. CEREUS TOXINS IN LOW-TEMPERATURE MILK 2597
TABLE 2. pH-dependent Vero cell cyBHIG culture superm
Strain Harvest% Cy
tie() 6.5 7.0
BC-SO 5 0 07 0 0
NCDO-1771 5 0 07 0 67.7
a Cytotoxicity was determined in microtiter pland Methods. Each well contained 1.4 x 10iPercent cytotoxicity was calculated at 1:20 4
Complete lysis of the cell monolayer (at 1:21cytotoxicity, and the additional percentage at t.1:40) was summed up.
S3/Vero cell cytotoxicity titer, 1:8Cprocedure was suitable for the referethe same parameters were applied toa low proportion (5/136; 3.7%) demoVero cell cytotoxicity. However, onproduced cytotoxin at 7 h (Table 2)time of 5 h did not seem suitable for thupon this observation, further toxicit'dairy isolates were performed withScreening was repeated for toxin pro(37.8%) showed HeLa S3, 50 (37.0%(43.0%) showed HEL cell cytotoxicit)Toxin production in milk and BHIG
further investigated the ability of daitoxin in milk during a time coursestrains positive for toxin production iiproduce detectable amounts of toxinphase of growth (i.e., at 7 h). The inplication rate in BHIG and milk woduring the late stationary growth p1than 50% of the strains demonstratecytotoxicity, while 73.2% showed cyi
100
H-La Vero MRC-5
Cytotoxicity
FIG. 1. Cytotoxin production by B. ce
or BHIG during time course. A total oigrown at 30°C in milk or BHIG, and cecollected at 5, 7, or 24 h and tested for in v
S3, Vero, and HEL (MRC-5) cell moiMaterials and Methods. Higher number oin milk than in BHIG, and the HEL csusceptible to the extracellular cytotoxic
btotoxicity of B. cereus cells when grown in milk. In contrast, B. cereus strainsatants showed no differences in cytotoxicity production when
(totoxicitya at pH: grown in BHIG for 7 or 24 h.When grown in BHIG, enterotoxin-positive laboratory
7.5 8.0 8.5 reference strains showed different cytotoxicity profiles.
0 0 0 Strain 4096/3 demonstrated HEL cytotoxicity only at 7 h,68.5 167.3 163.6 and strain 4433/73 showed a decreased cytotoxicity at a 24-h
harvest time, while strains NCDO-1771 and ATCC-145790 187.9 188.3 showed higher HEL cytotoxicity at 24 h compared to a 7-h
123.3 188.4 187.9 harvest time. However, when grown in milk all the referenceates as described in Materials strains demonstrated a high HEL cytotoxicity at both har-Vero cells in a monolayer. vest times. On the other hand, 4810/72, a reference strain for
dilution of the supematant. emetic toxin, failed to produce any detectable cytotoxicity in'0) was considered as 10OTo BHIG; however, the same strain demonstrated a high HELhe next further dilution (i.e., cytotoxicity in milk at both harvest times.
Toxin production at low temperatures with and withoutaeration. B. cereus reference strain NCDO-1771 and one
)), indicating that this dairy isolate, BC-50, were tested for toxin production atnce strains. But when 15°C in milk and BHIG during a time course with aeration bythe dairy isolates, only mechanical shaking at 200 rpm. Strain NCDO-1771 demon-nstrated HeLa S3 and strated a moderate HeLa S3 and a high Vero cell cytotoxic-e dairy isolate, BC-50, ity during late stationary growth phase (68 to 92 h) in milk, and thus the harvest and BHIG. The cytotoxicity pattern of the dairy isolateie dairy isolates. Based BC-50 was similar to that of NCDO-1771 when grown iny experiments with the milk; however, in BHIG medium the BC-50 strain showed aharvest time at 7 h. low Vero cell cytotoxicity at 92.5 h and no HeLa S3 cell
luction, and 51 isolates cytotoxicity. When grown in milk or BHIG medium without) showed Vero, and 58 mechanical shaking (i.e., aeration), neither of the two strainsy at a 7-h growth phase. produced any detectable levels of cytotoxins.at 7 and 24 h. We have We have further investigated toxin production at lowiry isolates to produce temperatures, i.e., at 8°C. Strains NCDO-1771 and BC-50(Fig. 1). Most of the did not acclimatize to this temperature. One strain, BC-179,n BHIG were unable to was selected and studied in detail. BC-179 was grown within milk during the log and without aeration in BHIG or milk, and the cells reachedioculum and the multi- stationary phase during 72 to 82 h of growth under bothere similar. However, culture conditions. Cytotoxic activity in the aerated BHIGhase (i.e., 24 h), more culture supernatant appeared during this time and reached ad HeLa and Vero cell maximum during 72 to 96 h. In the absence of aeration,totoxicity against HEL cytotoxicity titers were low during the early 72 h, with some
increase at 120 h. When grown in milk, cytotoxicity titerswere low during late logarithmic phase and gradually in-
Milkcreased to a titer of 1:128 during 72 to 96 h. BC-179demonstrated no detectable cytotoxicity in nonaerated milk(Table 3).A total of 102 strains positive for extracellular cytotoxin
production at 30°C in milk and BHIG at 7 and 24 h were
further tested for cytotoxin production at 8°C in milk after 72h (Table 4). Of the three cell lines tested for cytotoxicity,HEL seemed more susceptible (7.8% negativity compared to24.5 and 31.4% negativity of Vero and HeLa S3 cytotoxici-ty). Thus, in the HEL cytotoxicity assay, 75 (73.5%) strainsdemonstrated a strong toxicity while 19 (18.6%) strainsshowed a low toxicity.Toxin production in milk at 8°C (measured as HEL
cytotoxicity) was further investigated in relation to thegrowth of B. cereus strains (Fig. 2). Slow-growing strains,
HILa Vero MRC-5 i.e., <2 x 107 CFU/ml, demonstrated no HEL cytotoxicity.The cluster of toxin-producing strains showed values be-tween 1 x 108 and 3 x 108 CFU/ml; however, the cytotox-
!reus dairy isolates in milk icity titers widely varied between individual strains. Four off 136 dairy isolates were the dairy isolates (HEL cytotoxicity positive at 30°C),ll-free supernatants were though acclimatized to grow in milk at 80C with CFU values'itro cytotoxicity on HeLa similar to those of toxigenic strains, failed to produce HELnolayers as described in cytotoxtysof isolates produced toxins cytotoxicity.Sell line was found more The nonaerated culture supernatants at a 72-h harvest timefactors. at 8°C had a pH of approximately 6.6, whereas the aerated
VOL. 55, 1989
r-e0
on June 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
2598 CHRISTIANSSON ET AL.
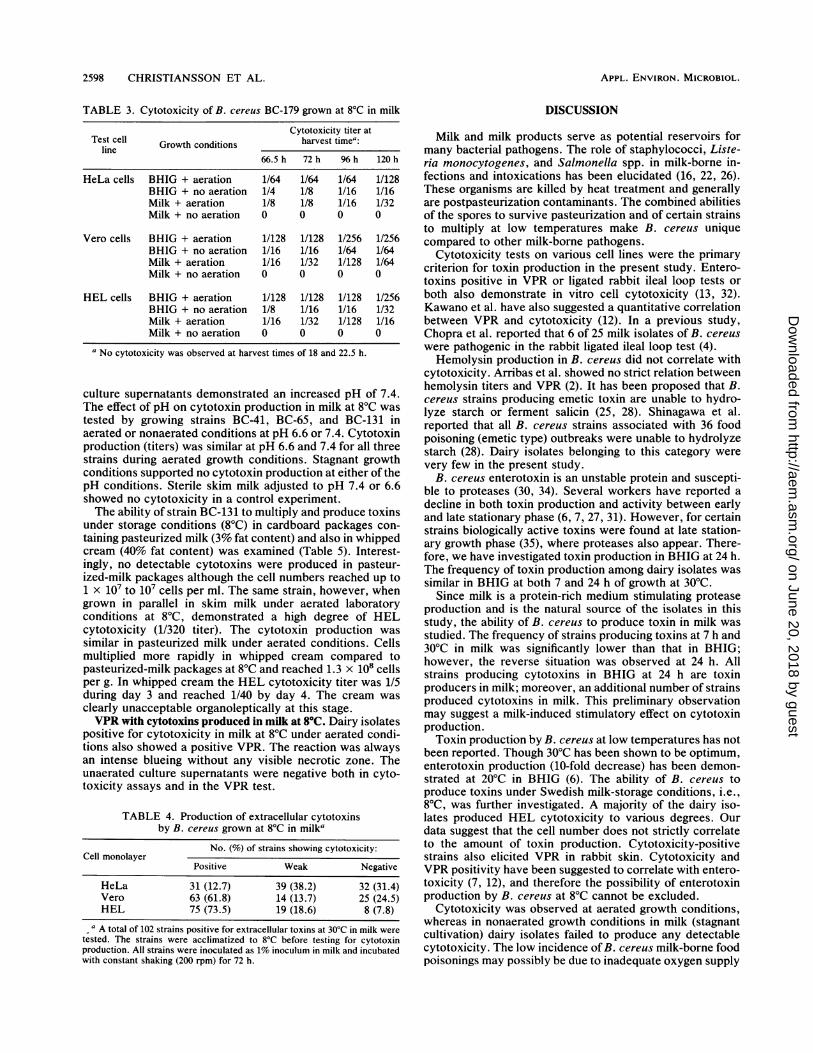
TABLE 3. Cytotoxicity of B. cereus BC-179 grown at 8°C in milk
Cytotoxicity titer atTest cell Growth conditions harvest timea:
line66.5h 72h 96h 120h
HeLa cells BHIG + aeration 1/64 1/64 1/64 1/128BHIG + no aeration 1/4 1/8 1/16 1/16Milk + aeration 1/8 1/8 1/16 1/32Milk + no aeration 0 0 0 0
Vero cells BHIG + aeration 1/128 1/128 1/256 1/256BHIG + no aeration 1/16 1/16 1/64 1/64Milk + aeration 1/16 1/32 1/128 1/64Milk + no aeration 0 0 0 0
HEL cells BHIG + aeration 1/128 1/128 1/128 1/256BHIG + no aeration 1/8 1/16 1/16 1/32Milk + aeration 1/16 1/32 1/128 1/16Milk + no aeration 0 0 0 0
a No cytotoxicity was observed at harvest times of 18 and 22.5 h.
culture supernatants demonstrated an increased pH of 7.4.The effect of pH on cytotoxin production in milk at 8°C wastested by growing strains BC-41, BC-65, and BC-131 inaerated or nonaerated conditions at pH 6.6 or 7.4. Cytotoxinproduction (titers) was similar at pH 6.6 and 7.4 for all threestrains during aerated growth conditions. Stagnant growthconditions supported no cytotoxin production at either of thepH conditions. Sterile skim milk adjusted to pH 7.4 or 6.6showed no cytotoxicity in a control experiment.The ability of strain BC-131 to multiply and produce toxins
under storage conditions (8°C) in cardboard packages con-taining pasteurized milk (3% fat content) and also in whippedcream (40% fat content) was examined (Table 5). Interest-ingly, no detectable cytotoxins were produced in pasteur-ized-milk packages although the cell numbers reached up to1 X 107 to 107 cells per ml. The same strain, however, whengrown in parallel in skim milk under aerated laboratoryconditions at 8°C, demonstrated a high degree of HELcytotoxicity (1/320 titer). The cytotoxin production wassimilar in pasteurized milk under aerated conditions. Cellsmultiplied more rapidly in whipped cream compared topasteurized-milk packages at 8°C and reached 1.3 x 108 cellsper g. In whipped cream the HEL cytotoxicity titer was 1/5during day 3 and reached 1/40 by day 4. The cream wasclearly unacceptable organoleptically at this stage.VPR with cytotoxins produced in milk at 8°C. Dairy isolates
positive for cytotoxicity in milk at 8°C under aerated condi-tions also showed a positive VPR. The reaction was alwaysan intense blueing without any visible necrotic zone. Theunaerated culture supernatants were negative both in cyto-toxicity assays and in the VPR test.
TABLE 4. Production of extracellular cytotoxinsby B. cereus grown at 8°C in milka
No. (%) of strains showing cytotoxicity:Cell monolayer
Positive Weak Negative
HeLa 31 (12.7) 39 (38.2) 32 (31.4)Vero 63 (61.8) 14 (13.7) 25 (24.5)HEL 75 (73.5) 19 (18.6) 8 (7.8)
, a A total of 102 strains positive for extracellular toxins at 30'C in milk weretested. The strains were acclimatized to 8'C before testing for cytotoxinproduction. All strains were inoculated as 1% inoculum in milk and incubatedwith constant shaking (200 rpm) for 72 h.
DISCUSSION
Milk and milk products serve as potential reservoirs formany bacterial pathogens. The role of staphylococci, Liste-ria monocytogenes, and Salmonella spp. in milk-borne in-fections and intoxications has been elucidated (16, 22, 26).These organisms are killed by heat treatment and generallyare postpasteurization contaminants. The combined abilitiesof the spores to survive pasteurization and of certain strainsto multiply at low temperatures make B. cereus uniquecompared to other milk-borne pathogens.
Cytotoxicity tests on various cell lines were the primarycriterion for toxin production in the present study. Entero-toxins positive in VPR or ligated rabbit ileal loop tests orboth also demonstrate in vitro cell cytotoxicity (13, 32).Kawano et al. have also suggested a quantitative correlationbetween VPR and cytotoxicity (12). In a previous study,Chopra et al. reported that 6 of 25 milk isolates of B. cereuswere pathogenic in the rabbit ligated ileal loop test (4).Hemolysin production in B. cereus did not correlate with
cytotoxicity. Arribas et al. showed no strict relation betweenhemolysin titers and VPR (2). It has been proposed that B.cereus strains producing emetic toxin are unable to hydro-lyze starch or ferment salicin (25, 28). Shinagawa et al.reported that all B. cereus strains associated with 36 foodpoisoning (emetic type) outbreaks were unable to hydrolyzestarch (28). Dairy isolates belonging to this category werevery few in the present study.
B. cereus enterotoxin is an unstable protein and suscepti-ble to proteases (30, 34). Several workers have reported adecline in both toxin production and activity between earlyand late stationary phase (6, 7, 27, 31). However, for certainstrains biologically active toxins were found at late station-ary growth phase (35), where proteases also appear. There-fore, we have investigated toxin production in BHIG at 24 h.The frequency of toxin production among dairy isolates wassimilar in BHIG at both 7 and 24 h of growth at 30°C.
Since milk is a protein-rich medium stimulating proteaseproduction and is the natural source of the isolates in thisstudy, the ability of B. cereus to produce toxin in milk wasstudied. The frequency of strains producing toxins at 7 h and30°C in milk was significantly lower than that in BHIG;however, the reverse situation was observed at 24 h. Allstrains producing cytotoxins in BHIG at 24 h are toxinproducers in milk; moreover, an additional number of strainsproduced cytotoxins in milk. This preliminary observationmay suggest a milk-induced stimulatory effect on cytotoxinproduction.Toxin production by B. cereus at low temperatures has not
been reported. Though 30°C has been shown to be optimum,enterotoxin production (10-fold decrease) has been demon-strated at 20°C in BHIG (6). The ability of B. cereus toproduce toxins under Swedish milk-storage conditions, i.e.,8°C, was further investigated. A majority of the dairy iso-lates produced HEL cytotoxicity to various degrees. Ourdata suggest that the cell number does not strictly correlateto the amount of toxin production. Cytotoxicity-positivestrains also elicited VPR in rabbit skin. Cytotoxicity andVPR positivity have been suggested to correlate with entero-toxicity (7, 12), and therefore the possibility of enterotoxinproduction by B. cereus at 8°C cannot be excluded.
Cytotoxicity was observed at aerated growth conditions,whereas in nonaerated growth conditions in milk (stagnantcultivation) dairy isolates failed to produce any detectablecytotoxicity. The low incidence ofB. cereus milk-borne foodpoisonings may possibly be due to inadequate oxygen supply
APPL. ENVIRON. MICROBIOL.
on June 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

B. CEREUS TOXINS IN LOW-TEMPERATURE MILK 2599
320_
C0-=
02
0
U
-I
o
0
160_
80_
40_
20-
10-
5-
0
S
S
*-00* *§-
*~~~
@0
S.0@
0@000 S
0@@0
* See-0 90 000 0 S
_0
* 00@@00000
i B
5 6 7 8 9 10
Growth in milk at 8°C (log 10 CFU / ml)
FIG. 2. Relation between bacterial cell number and toxin titers of B. cereus dairy isolates grown in milk at 8°C. A total of 102 strains weregrown in milk at 80C according to conditions described in Materials and Methods. After 72 h, bacterial cell numbers were determined by platecounts and the cell-free supernatants were tested for HEL cytotoxicity at different dilutions. Strains produced no detectable cytotoxicitywhen growth was below 2 x 107 CFU/ml. Variable degree of cytotoxicity was observed among strains showing cell counts between 1 x 108and 2.5 x 108 CFU/ml.
in milk packages. This hypothesis was evaluated by tests fortoxin production in milk packages with a potent toxigenicdairy isolate, BC-131. Despite a high inoculum, BC-131failed to produce any cytotoxicity in shaken and stagnantmilk packages. On the other hand, BC-131 produced asizeable cytotoxicity in whipped cream. Therefore, aeratedconditions allowing oxygen transfer seem important fortoxin production in a milk environment as for BHIG, assuggested by Glatz et al. (7).
In conclusion, we have shown that the incidence oftoxigenic B. cereus is high in milk and cream. Milk is a
TABLE 5. Growth and toxin production by B. cereus BC-131 at8°C in commercial cardboard packages and in whipped creama
Growth and toxin titerb at time:
Condition Day 0 Day 3 Day 4Toxin Toxin ToxinCFU/ml titer CFU/ml titer CFU/ml titer
Package, sealed/ 4 x 104 0 1 X 107 0 1 X 107 0stagnant
Package, sealed/ 6 x 104 0 3 x 107 0 4 x 107 0shakenc
Package, open/ 6 x 104 0 2 x 107 0 2 x 107 0stagnant
Skim milk, aerated 2 x 106 0 2 x 108 1/320Whipped creamd 5 x 104 0 5 x 107 1/5 1 X 108 1/40
a B. cereus BC-131 was grown in skim milk at 8°C for 7 days. A 0.3%inoculum was injected into the packages with a sterile syringe.bHEL cytotoxicity titer, standard toxin assay conditions.c Mechanical shaking at 300 rpm.d Inoculum was added to the cream before whipping.
suitable medium for B. cereus to produce toxins even at 8°C.However, these toxins were only produced in stronglyaerated conditions. The very low incidence of milk-bornefood poisonings caused by B. cereus is probably due toinsufficient aeration for toxin production under normal stor-age conditions. The toxin production by B. cereus at 8°Creported in this study raises questions on such implicationsin other well-aerated refrigerated foods. However, it remainsto be established whether the toxins produced at 8°C are"true" enterotoxins.
ACKNOWLEDGMENTS
We thank Rangne Fonden for critical reading of the manuscript;Irena Ollermark, Kerstin Ekelund, and Lena Jonsson for excellenttechnical assistance; and Ingvar Frick for photography.
LITERATURE CITED1. Ahmed, A. A., M. K. Moustafa, and E. H. Marth. 1983.
Incidence of Bacillus cereus in milk and some milk products. J.Food. Prot. 46:126-128.
2. Arribas, M. L. G., C. J. Plaza, M. C. de la Rosa, and M. A.Mosso. 1988. Characterization of Bacillus cereus strains isolatedfrom drugs and evaluation of their toxins. J. Appl. Bacteriol.64:257-264.
3. Billing, E., and W. A. Cuthbert. 1958. "Bitty" cream. Theoccurrence and significance of Bacillus cereus spores in rawmilk supplies. J. Appl. Microbiol. 21:65-78.
4. Chopra, P., A. Singh, and M. S. Kalra. 1980. Occurrence ofBacillus cereus in milk and milk products. Indian J. Dairy Sci.33:248-252.
5. Coghill, D., and H. S. Juffs. 1979. Incidence of psychrotrophicbacteria in pasteurized milk and cream products and effect oftemperature on their growth. Aust. J. Dairy Technol. 34:
VOL. 55, 1989
on June 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2600 CHRISTIANSSON ET AL.
150-153.6. Glatz, B. A., and J. M. Goepfert. 1976. Defined conditions for
synthesis of Bacillus cereus enterotoxin by fermentor-growncultures. Appl. Environ. Microbiol. 32:400-404.
7. Glatz, B. A., W. M. Spira, and J. M. Goepfert. 1974. Alterationof vascular permeability in rabbits by culture filtrates of Bacilluscereus and related species. Infect. Immun. 10:299-303.
8. Goepfert, J. W., W. M. Spira, and H. U. Kim. 1972. Bacilluscereus: food poisoning organism. A review. J. Milk FoodTechnol. 35:213-227.
9. Hauge, S. 1950. Matforgiftninger fremkalt av Bacillus cereus.Nord. Hyg. Tidskr. 31:189-206.
10. Hauge, S. 1955. Food poisoning caused by aerobic spore form-ing bacilli. J. Appl. Bacteriol. 18:591-595.
11. Johnsson, K. M. 1984. Bacillus cereus foodborne illness-anupdate. J. Food Prot. 47:145-153.
12. Kawano, J., A. Shimizu, K. Fujii, S. Takeuchi, and S. Kimura.1987. Characterization of Bacillus cereus haemolysis pattern,cytotoxicity and vascular permeability factor activity. Sci. Rep.Fac. Agric. Kobe Univ. 17:273-281.
13. Kramer, J. 1984. Bacillus cereus exotoxins: production, isola-tion, detection and properties, p. 385-386. In J. E. Alouf,J. H. Freer, F. J. Fehrenbach, and J. Jeljaszewicz (ed.), Bac-terial protein toxins. Academic Press, Inc. (London), Ltd.,London.
14. Kristensen, K. K. 1981. Et tilfaelde af Bacillus cereuslevnedsmiddelforgiftning. Dansk Vet. Tidsskr. 64:766-768.
15. Logan, N. A., and R. C. W. Berkeley. 1984. Identification ofBacillus strains using the API system. J. Gen. Microbiol.130:1871-1882.
16. Marth, E. H. 1969. Salmonellae and salmonellosis associatedwith milk and milk products. A review. J. Dairy Sci. 52:283-315.
17. McSwiggan, D. A., R. J. Gilbert, and F. W. T. Fowler. 1975.Food poisoning associated with pasteurized cream. PHLS Com.Dis. Rep. 43.
18. Melling, J., and B. J. Capel. 1978. Characteristics of Bacilluscereus emetic toxin. FEMS Microbiol. Lett. 4:133-135.
19. Melling, J., B. J. Capel, P. C. B. Turnbull, and R. J. Gilbert.1976. Identification of a novel enterotoxigenic activity associ-ated with Bacillus cereus. J. Clin. Pathol. 29:938-940.
20. Mortimer, P. R., and G. McCann. 1974. Food-poisoning epi-sodes associated with Bacillus cereus in fried rice. Lanceti:1043-1045.
21. Mossel, D. A. A., M. J. Koopman, and E. Jongerius. 1967.Enumeration of Bacillus cereus in foods. Appl. Microbiol.15:650-653.
22. Naidu, A. S., and K. Rajyalakshmi. 1985. Enterotoxicological
and epidemiological investigation of staphylococcal food poi-soning outbreaks, p. 194-199. In Proceedings of the 2nd WorldCongress on Foodborne Infections and Intoxications, Berlin,vol. 2.
23. Overcast, W. W., and K. Atmaram. 1974. The role of Bacilluscereus in sweet curdling of fluid milk. J. Milk Food Technol.37:233-236.
24. Parry, J. M., P. C. B. Turnbull, and J. R. Gibson. 1983. Acolour atlas of Bacillus species. Wolfe Medical PublicationsLtd., London.
25. Raevuori, M., T. Kiutamo, and A. Niskanen. 1977. Comparativestudies of Bacillus cereus strains isolated from various foodsand food poisoning outbreaks. Acta Vet. Scan. 18:397-407.
26. Rosenow, E. M., and E. H. Marth. 1987. Listeria, listeriosis anddairy foods: a review. Cult. Dairy Prod. J. 22:13-17.
27. Sharma, P. L., and R. C. Dogra. 1983. Bacillus cereus entero-toxin and its production in different foods. J. Food Sci. Technol.20:223-227.
28. Shinagawa, K., N. Matsusaka, H. Konuma, and H. Kurata. 1985.The relation between the diarrheal and other biological activitiesof Bacillus cereus involved in food poisoning outbreaks. Jpn. J.Vet. Sci. 47:557-565.
29. Spira, W. M., and J. M. Goepfert. 1972. Bacillus cereus-inducedfluid accumulation in rabbit ileal loops. Appl. Microbiol. 24:341-348.
30. Spira, W. M., and J. M. Goepfert. 1975. Biological characteris-tics of an enterotoxin produced by Bacillus cereus. Can. J.Microbiol. 21:1236-1246.
31. Spira, W. M., and G. J. Silverman. 1979. Effects of glucose, pH,and dissolved-oxygen tension on Bacillus cereus growth andpermeability factor production in batch culture. Appl. Environ.Microbiol. 37:109-116.
32. Thompson, N. E., M. J. Ketterhagen, M. S. Bergdoll, and E. J.Schantz. 1984. Isolation and some properties of an enterotoxinproduced by Bacillus cereus. Infect. Immun. 43:887-894.
33. Turnbull, P. C. B. 1986. Bacillus cereus toxins, p. 397-448. In F.Dorner and J. Drews (ed.), Pharmacology of bacterial toxins.Pergamon Press, London.
34. Turnbull, P. C. B., J. M. Kramer, K. Jorgensen, R. J. Gilbert,and J. Melling. 1979. Properties and production characteristicsof vomiting, diarrheal, and necrotizing toxins of Bacillus cereus.Am. J. Clin. Nutrition 32:219-228.
35. Turnbull, P. C. B., J. F. Nottingham, and A. C. Ghosh. 1977. Asevere necrotic enterotoxin produced by certain food, foodpoisoning and other clinical isolates of Bacillus cereus. Br. J.Exp. Pathol. 58:273-280.
36. VIad, A., and A. VIad. 1972. Toxiinfectie alimentara provocatade Bacillus cereus. Microbiol. Parazitol. Epidemiol. 18:531-536.
APPL. ENVIRON. MICROBIOL.
on June 20, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from


















