
Torres 1998
22
American Journal of Primatology 44:255–275 (1998) © 1998 Wiley-Liss, Inc. Chromosome Diversity of the Genus Aotus From Colombia OLGA MARÍA TORRES, 1 * SANDRA ENCISO, 2 FRANCISCO RUIZ, 2 ELIZABETH SILVA, 3 AND IVÁN YUNIS 2 1 Instituto de GenØtica, Universidad Nacional, SantafØ de BogotÆ, Colombia 2 Laboratorio de InmunogenØtica, Instituto Nacional de Salud, SantafØ de BogotÆ, Colombia 3 Laboratorio de GenØtica, Instituto Nacional de Salud, SantafØ de BogotÆ, Colombia Description of six Colombian karyomorphs is completed through an ex- tensive cytogenetic characterization of 35 Aotus (owl monkeys) specimens. The description of a new karyomorph for Colombian Aotus by chromo- some on Q, G, R, and C, sequential banding is included. Pairs of karyomorphs 2 and 3 and 6 and 9 with 2n of 54, and 50, respectively, as well as karyomorphs 7 and 8 with 46 and 58 chromosomes were strongly suspected to represent different species on the grounds of large karyo- typic differences. A proposal for a chromosome nomenclature of Aotus karyomorphs that aims to clarify Aotus taxonomy is presented which achieves a precise correspondence of different banding patterns, based on Q, G, R, and C sequential banding and chromosome measurements. Although our contribution is not a universal nomenclature system, unique criteria for chromosome denomination within Aotus karyomorphs are es- tablished. Previous systems of chromosome nomenclature have not suc- cessfully addressed the nomenclature of chromosomes of the same karyotype. Am. J. Primatol. 44:255–275, 1998. © 1998 Wiley-Liss, Inc. Key words: Cebidae; Aotus; cytotaxonomy; karyomorph; karyotype; Colombian owl monkey INTRODUCTION Taxonomy of neotropical monkeys of the genus Aotus (owl monkeys) is com- plicated due to the difficulty of differentiating species using morphological char- acters, the diversity of chromosome numbers within the genus, and the chromosome number polymorphism of several species. Karyological data concerning the genus Aotus reveal many intra- and inter- specific chromosomal variations, both numerical and structural. Cytogenetic stud- ies carried out by conventional Giemsa staining and banding techniques show chromosome diploid numbers ranging from 46–58 in 18 karyotypes that can be assigned to general karyotypically defined taxa. Table I summarizes the results of cytogenetic studies available to date, including ours. Contract grant sponsor: Instituto Nacional de Salud; Contract grant sponsor: Universidad Nacional de Colombia. *Correspondence to: Olga María Torres, Instituto de Genética, Universidad Nacional de Colombia Santafé de Bogotá, Colombia. Received for publication 3 November 1996; revision accepted 9 November 1997.
Transcript of Torres 1998

American Journal of Primatology 44:255–275 (1998)
© 1998 Wiley-Liss, Inc.
Chromosome Diversity of the Genus AotusFrom Colombia
OLGA MARÍA TORRES,1* SANDRA ENCISO,2 FRANCISCO RUIZ,2 ELIZABETH SILVA,3
AND IVÁN YUNIS21Instituto de Genética, Universidad Nacional, Santafé de Bogotá, Colombia2Laboratorio de Inmunogenética, Instituto Nacional de Salud, Santafé de Bogotá, Colombia3Laboratorio de Genética, Instituto Nacional de Salud, Santafé de Bogotá, Colombia
Description of six Colombian karyomorphs is completed through an ex-tensive cytogenetic characterization of 35 Aotus (owl monkeys) specimens.The description of a new karyomorph for Colombian Aotus by chromo-some on Q, G, R, and C, sequential banding is included. Pairs ofkaryomorphs 2 and 3 and 6 and 9 with 2n of 54, and 50, respectively, aswell as karyomorphs 7 and 8 with 46 and 58 chromosomes were stronglysuspected to represent different species on the grounds of large karyo-typic differences. A proposal for a chromosome nomenclature of Aotuskaryomorphs that aims to clarify Aotus taxonomy is presented whichachieves a precise correspondence of different banding patterns, basedon Q, G, R, and C sequential banding and chromosome measurements.Although our contribution is not a universal nomenclature system, uniquecriteria for chromosome denomination within Aotus karyomorphs are es-tablished. Previous systems of chromosome nomenclature have not suc-cessfully addressed the nomenclature of chromosomes of the samekaryotype. Am. J. Primatol. 44:255–275, 1998. © 1998 Wiley-Liss, Inc.
Key words: Cebidae; Aotus; cytotaxonomy; karyomorph; karyotype;Colombian owl monkey
INTRODUCTIONTaxonomy of neotropical monkeys of the genus Aotus (owl monkeys) is com-
plicated due to the difficulty of differentiating species using morphological char-acters, the diversity of chromosome numbers within the genus, and thechromosome number polymorphism of several species.
Karyological data concerning the genus Aotus reveal many intra- and inter-specific chromosomal variations, both numerical and structural. Cytogenetic stud-ies carried out by conventional Giemsa staining and banding techniques showchromosome diploid numbers ranging from 46–58 in 18 karyotypes that can beassigned to general karyotypically defined taxa. Table I summarizes the resultsof cytogenetic studies available to date, including ours.
Contract grant sponsor: Instituto Nacional de Salud; Contract grant sponsor: Universidad Nacionalde Colombia.
*Correspondence to: Olga María Torres, Instituto de Genética, Universidad Nacional de ColombiaSantafé de Bogotá, Colombia.
Received for publication 3 November 1996; revision accepted 9 November 1997.

256 / Torres et al.
TAB
LE
I. S
umm
ary
of K
aryo
logi
cal D
ata
Rep
orte
d fo
r A
otus
Acc
ordi
ng to
Spec
ies
Her
shko
vitz
[198
3]B
andi
ng T
echn
ique
seC
ount
ryA
otu
sA
otu
sPh
a2n
bK
Mc
Kd
CM
QG
CN
RB
Gof
ori
gin
Ref
eren
cesf
A. l
emu
rin
us
lem
uri
nu
sB
56–5
51
VII
I+
+Pa
nam
a1,
2,3
IXA
. tri
virg
atu
s gr
isei
mem
bra
A. l
emu
rin
us
gris
eim
embr
aB
54–5
22
IIa
++
++
++
Col
ombi
a2,
4,5,
6,7
IVA
. tri
virg
atu
s tr
ivir
gatu
sA
. bru
mba
cki
506
++
Col
ombi
a5
A. t
rivi
rgat
us
voci
fera
ns
A. v
ocif
eran
sB
467
V+
+C
olom
bia
2,4,
9,10
,11
48–4
6X
Perú
XIB
razi
lA
. her
shko
viti
zig
588h
++
Col
ombi
a7,
8A
. sp
509h
Col
ombi
ah
A. t
rivi
rgat
us
triv
irga
tus
A. n
ancy
mai
A54
3I
++
+C
olom
bia
2,4,
10,1
1Pe
rúB
razi
lA
. tri
virg
atu
s n
igri
ceps
A. n
igri
ceps
C52
/51
4V
II+
++
++
Perú
2,4,
9,13
A. t
rivi
rgat
us
azar
aeA
. aza
rae
boli
vien
sis
D50
/49
5V
I+
++
+B
oliv
ia2,
4,9,
14,1
5A
. tri
virg
atu
s A
zara
eA
. aza
rae
azar
aeD
50/4
9+
++
Arg
entin
a16
A. i
nfu
latu
sD
50/4
910
h+
+B
razi
l17
A. (
Ron
don
ia)
D50
/59
Bra
zil
18A
. tri
virg
atu
s50
–54
+19
–28
a Ph, p
heno
type
as
defin
ed b
y M
a et
al [
1976
].b 2n
, dip
loid
num
ber.
c KM
, kar
yom
orph
den
omin
atio
n as
def
ined
by
Reu
mer
and
De
Boe
r [1
980]
.d K
, kar
yoty
pe d
enom
inat
ion
as d
efin
ed b
y M
a et
al [
1976
].e C
M, c
hrom
osom
e m
easu
res.
Q, Q
ban
ds; G
, G b
ands
;R, R
ban
ds; C
, C b
ands
; N, N
ban
ds, n
ucle
olar
org
aniz
er r
egio
ns. R
BG
, R b
ands
, lat
e re
plic
atio
n by
bro
mod
eoxi
urid
ine
inco
rpor
atio
n.f 1,
Ma
et a
l. [1
978]
; 2, M
a [1
981]
; 3, B
ogar
t [1
978]
; cit
ed b
y R
eum
er a
nd D
e B
oer
[198
0]; 4
, Ma
et a
l. [1
976]
; 5, Y
unis
et
al. [
1977
]; 6,
Mill
er e
t al
. [19
77];
7, G
iral
do e
t al
.[1
986]
; 8, G
iral
do e
t al.
[198
3]; 9
, Reu
mer
and
De
Boe
r [1
980]
; 10,
Ma
et a
l. [1
985]
; 11,
Pie
czar
ka e
t al.
[199
2]; 1
2, D
utri
llaux
and
Cou
turi
er [1
981]
; 13,
Ma
et a
l. [1
980]
; 14,
Min
ezaw
a an
d Va
ldiv
ia [
1984
]; 15
, Cam
befo
rt a
nd M
oro
[197
8]; 1
6, M
udry
et
al. [
1984
]; 17
, Pie
czar
ka a
nd N
agam
achi
[19
88];
18, P
iecz
arka
et
al. [
1993
]; 19
, Chu
and
Ben
der
[196
1]; 2
0, C
hiar
elli
[196
1]; 2
1, E
gozc
ue [1
971]
; 22,
Bru
mba
ck e
t al
. [19
71];
23, B
rum
back
[197
5a];
25, B
rum
back
[197
5b];
26, D
e B
oer
[197
4]; 2
7, K
oiffm
ann
and
Sald
anha
[197
4]; 2
8, D
esca
illea
ux e
t al.
[199
0].
g Gro
ves
[199
3].
h This
rep
ort.

Chromosome Diversity of Colombian Aotus / 257
In spite of extensive research of the genus Aotus, systematic and phyloge-netic studies cannot take full advantage of karyotypical information, for cytoge-netic studies have often been rather superficial, incomplete, and in some instancesdisconnected from taxonomic relationships. Furthermore, much confusion ariseson the chromosome nomenclature of those karyological types.
Taxonomy of the genus Aotus has been highly controversial. Aotus, regardedas a single species with allopatric subspecies [Hershkovitz, 1949; Hernandez-Comacho & Cooper, 1975], may actually be comprised of a heterogeneous assem-blage of karyotypically different animals [Ma et al., 1978; Ma, 1981]. It remainsto be explained why phenotypic similarities occur between groups, while cytoge-netically distinct forms are found in the same geographical areas.
A taxonomic review of this genus [Hershkovitz, 1983] proposes that nine sepa-rate species and four subspecies can be defined on the basis of variation of pel-age coloration, karyotype, and geographic origin. These species belong to twomain groups: the gray-neck group north of the Amazon River and the red-neckgroup south of the Amazon River. However, more recent data challenge this view[Pieczarka et al., 1993].
Ford [1994] recognized only two species of the gray-neck group and five spe-cies of the red-neck group, based on multivariate analysis of two data sets: cra-nial and pelage features.
Cytogenetic characterization of Colombian owl monkeys from different loca-tions may help clarify taxonomic relationships. We propose a chromosome no-menclature for each karyomorph of the genus. Using this proposed nomenclature,our results show six karyomorphs north of the Amazon River. The termkaryomorph (KM) is used here according to the definition of Reumer and DeBoer [1980].
METHODSAnimals
This study covers 35 owl monkeys form different geographical areas of Co-lombia shown in Figure 1. Twenty-two specimens were fully characterized,whereas information was supplemented on 13 specimens which had been reportedpreviously [Yunis et al., 1977; Giraldo et al, 1983]. Identification, place of cap-ture, and sex of specimens are presented in Table II as well as the animal fateand current location. Some animals are housed at Bioterio Central InstitutoNacional de Salud (BINS), Centro de Rehabilitación World Society for the Pro-tection of Animals (WSPA), and Centro Experimental de Primates (CEP)(Universidad del Valle/Zoológico de Cali/Instituto Nacional de Salud). The skinand skulls of several specimens were preserved and catalogued at Instituto deCiencias Naturales (ICN), Museo de Historia Natural Universidad Nacional deColombia, and at Instituto Humboldt (IU) (Instituto Nacional de los RecursosNaturales Renovables y del Ambiente [INDERENA]).
CytogeneticsWhole blood samples were cultured according to a modification of the method
of Moorhead et al. [1960] using RPMI 1640 culture medium supplemented with20% fetal bovine serum. Phytohaemagglutinin P (Difco, Detroit, MI), a crudeextract of lectin from Vicia fava seeds (Favin), or pure Favin (Sigma, St. Louis,MO) was used as a mitogen. Favin crude extract gave the best results.

258 / Torres et al.
The Favin crude extract was prepared in our laboratory according to Arangoand Moreno [1977]. Favin was used at 5–20 µg/ml in a constant volume of wholeblood of 0.04 ml/ml. Incubations were carried out at 36.5°C for 63–79 hours.Colcemid (Sigma) (1 µg/ml) was added 40 min before harvesting. Four cultureswere performed for each specimen, two of them for late DNA replication patterns(RBH-FPG) after a 5-bromodeoxyuridine (BrdU) (50 µg/ml) terminal pulse treat-ment during the last 5 and 7 h [Willard & Latt, 1976].
Air-dried slides were prepared by conventional methods. Chromosomes werebanded with quinacrine dichloride for QFQ identification [Caspersson et al., 1970].Giemsa solid staining or banding techniques were carried out on slides that hadbeen Q-banded. Banding techniques included general and regional banding. Thegeneral banding techniques were R banding RHG [Sehested, 1974], G bandingGTG [Seabright, 1971], and R late replication banding (RBH-FPG) [Willard &Latt, 1976; Goto et al., 1975]. Regional banding techniques were C banding CBGmodified [Arrighi & Hsu, 1971; Sumner, 1972] and Ag staining NOR [Goodpasture& Bloom, 1975]. Photographs were taken of each metaphase stained by Q band-ing and of the same metaphase stained by the others (R, G, C, Ag banding) orsolid staining methods. Double karyotypes were prepared by cutting out printsof the same metaphase stained simultaneously by two methods.
Fig. 1. Colombian map showing the regions of karyomorphs location. 2, KM2, Costa Altantica; 3, KM3,Amazonas; 6, KM6, Meta; 7, KM7, Caquetá; 8, KM8, Boyacá; 9, KM9, Quindío.

Chromosome Diversity of Colombian Aotus / 259
Chromosome measurements were made on two to ten Giemsa karyotypes(Table II) from each animal by two independent observers using a graduated magni-fying glass (0.1 mm). In order to balance the known inter- and intraindividual vari-ability, we made chromosome length measurements on at least ten karyotypes (TableII) from each karyomorph. Average chromosome lengths are reported.
TABLE II. Aotus Specimens Studied
Identification Current locationa Cultb Place of capture Sex KMc 2n Numberd Dead Alive
2 67008–72 Bolivar Tiquisio F 53 5 INDERENA060–74 Bolivar Tiquisio M 54 5 INDERENA062–74 Bolivar Tiquisio M 54 5 INDERENA064–74 Bolivar Tiquisio M 54 5 INDERENA065–74 Bolivar Tiquisio F 52 5 INDERENA066–77 N. Santander (unknown) F 54 3 U. SALLE067–77 N. Santander (unknown) M 54 3 U. SALLE
F1.4 056–94 Sucre B8 × A57 F 54 3 INC 14020F1.59 001–94 Sucre A62 × A71 M 54 2 BINSF1.15 086–94 Sucre A86 × A67 M 54 3 BINSF1.74 085–94 Sucre A88 × A21 F 53 2 BINSF1.43 058–94 Sucre D48 × D47 M 53 2 BINS642 021–95 (Unknown) F 52 5 WSPAB.1 002–94 Sucre (San Marcos) M 54 2 BINSB.4 013–94 Sucre (San Marcos) F 52 5 BINSD.36 003–94 Sucre (San Marcos) F 53 4 BINSD.70 004–94 Sucre (San Marcos) F 54 4 ICN14022F1.63 011–95 Sucre (unknown) M 54 2 ICN 14016F1.136 057–94 Sucre B16 × A53 F 54 2 ICN 14017
3 19A1 014–05 Amazonas (Leticia) M 54 3 BINSA2 015–95 Amazonas (Leticia) F 54 5 ICN 14019A3 088–94 Amazonas (Leticia) M 54 3 BINSA4 089–94 Amazonas (Leticia) F 54 3 BINSA5 080–94 Amazonas (Leticia) M 54 5 BINS
6 25039–73 Meta (Villavicencio) F 50 5 IH 0354040–73 Meta (Villavicencio) F 50 5 IH 2806
V66 009–94 (Unknown) F 50 5 CEP728 020–95 (Unknown) M 50 5 WSPA905 022–95 (Unknown) M 50 5 WSPA
7 10V32 010-94 Caquetá (unknown) F 46 10 CEP
8 15L1 080–82 Boyacá (Pajarito) M 58 6 LostL2 084–83 Boyacá (Pajarito) F 58 3 ICN 8880L3 089–83 Boyacá (Pajarito) M 58 4 LostL4 081–82 Boyacá (Pajarito) F 58 2 IH 4140
9 10Q2 079–94 Quindio (unknown) F 50 10 ICN 14023aIdentification number of specimen.bIdentification number of kidney or blood culture-year.cKaryomorph. Located as shown in Figure 1.dNumber of karyotypes scored from each animal and karyomorph.

260 / Torres et al.
Chromosome length is expressed as the percentage of the X-containing hap-loid complement length (% TCL). The arm ratio is the length of the long armdivided by the length of the short arm (q/p).
The karyotypes were ordered on the basis of chromosome measurements (sizeand centromeric position) and the heterochromatic nature of the short arm, assuggested by Miller et al., [1977], with Tjio and Levan’s [1956] ranges. The auto-somes were classified in three groups defined according to the mean values ofarm ratio and arranged in a decreasing size order within each group as follows:metacentric arm ratio, 1–1.9; submetacentric plus subtelocentric arm ratio, 2–4.9;acrocentric plus telocentric arm ratio, >5. All chromosomes with whole heterochro-matic short arms were included in the third group independently of arm ratio.
For the nomenclature proposed herein, each chromosome is defined by itsbanding patterns in addition to its size and morphology. Each karyotype wasarranged with three major morphological groups clearly defined. C banding letsus easily distinguish whole heterochromatic short arm chromosomes which areresponsible for the major interindividual variability in Aotus chromosomes. Eachgroup was ordered by decreasing chromosome size.
RESULTSPhenotypes
The specimens from Amazonas belong to the red-neck group, whereas thosefrom Costa Atlántica, Meta, Caquetá, Boyacá, and Quindío belong to the gray-neck species group. Following the classification proposed by Hershkovitz [1983],specimens from Costa Atlántica, Meta, Caquetá and Amazonas would be namedAotus lemurinus griseimembra, Aotus brumbacki, Aotus vociferans, and Aotusnancymai, respectively.
Following phenotypic descriptions from Ma et al. [1976], specimens from CostaAtlántica would belong to phenotype B and those from Amazonas to phenotypeA; the specimen from Quindío, those from Meta, and the four from Boyacá couldnot be assigned to either group. Salient phenotypic characters of the unassignedspecimens are as follows.
The specimen from Quindío (Fig. 2) shows some peculiar features: the linearmarks on the forehead are wide and well defined, while lateral ones run forwardfrom the top of the head going down like a semicircle at the cheek level, and thecommissure. The neck is dark gray, except for a yellowish tuft present at thethroat level. The abdominal and thoracic ventral regions are covered by longlight yellowish fur, which is more evident at the order line which separates theagouti color from the ventral area. A distinctive characteristic for this specimenis that the pattern reaches the wrist on the forelimbs and the ankle on thehindlimbs, in contrast with phenotypes A and B, in which the pattern reachesonly the elbow and knee or halfway, respectively. In the specimens from Meta,the ventral color also extends as far as the elbows and knees to the inside of thearms and legs, and the hair color is pale orange. The gray-neck specimens fromBoyacá show a rather dull yellow hair color in the limbs’ underside reaching theelbows and knees; the hair coat is long and soft [Espinal et al., 1984].
CytogeneticsEight different karyotypes were identified among the 35 specimens, which
were distributed in six karyomorphs: 2, 3, 6, 7, 8, and 9. Chromosomal struc-tures of those karyomorphs are shown in Table III.

Chromosome Diversity of Colombian Aotus / 261
Chromosome relative length and arm ratio measurements from karyomorphsstudied are shown in Table IV. Chromosome numbers are unique for eachkaryomorph. Numbers were assigned to a chromosome as a result of its length,morphology, and sequencing banding patterns; consequently, a number does notrepresent the same chromosome in different karyomorphs.
Every chromosome in the karyomorphs analyzed showed constitutive hetero-chromatin (C banding) at pericentromeric regions, but only some chromosomeshave a completely heterochromatic short arm, which sometimes is absent fromthe homologous chromosome. Variations in the amount of heterochromatin be-tween homologous chromosomes and among specimens were observed (data notshown). At least three classes of constitutive heterochromatin were present. 1)C-positive–Q-brilliant (Fig. 8, chromosome 5, for example), 2) C-positive–Q-pale(Fig. 7, chromosome 13, for example), and 3) C-negative–Q-brilliant (Fig. 6, chro-mosome 3, for example).
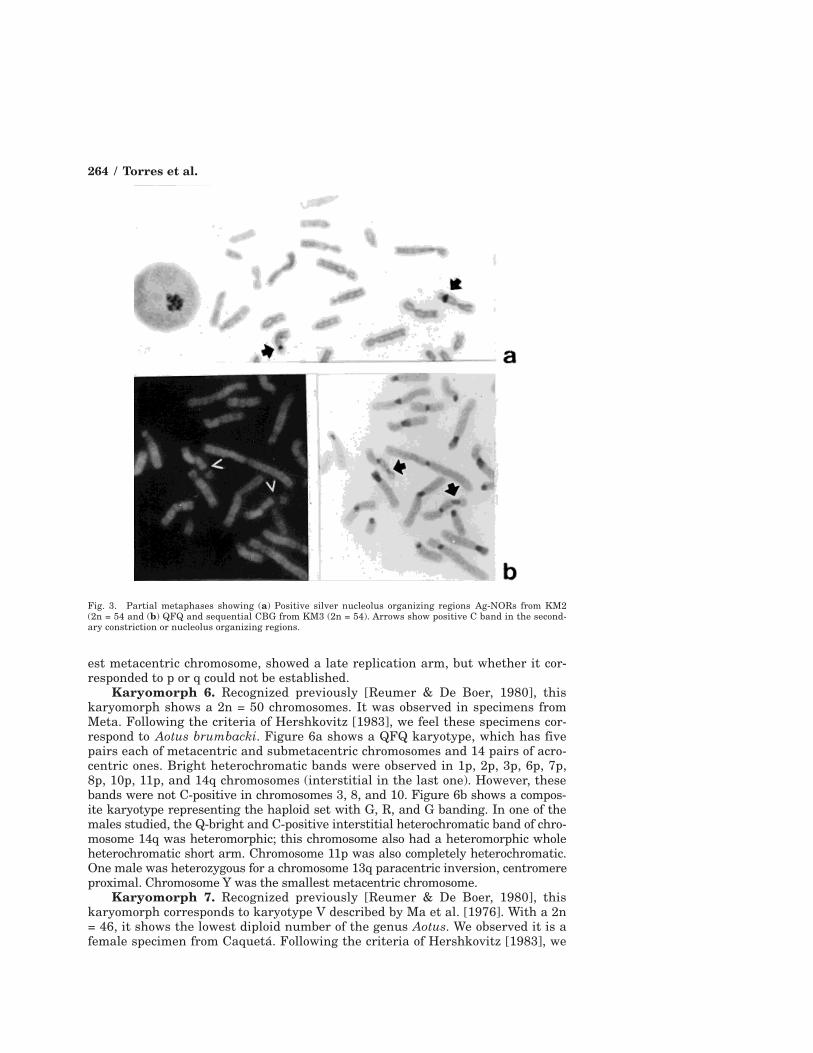
All specimens studied showed a metacentric chromosome with an achromaticregion in q, the marker chromosome characteristic of the Aotus genus [Egozcueet al., 1969]. No specimen showed consistent CBG-positive staining in this re-gion, except occasionally in a few cells, and this positive band, significantly lighter(Fig. 3), could represent NOR activity. This observation occurred in allkaryomorphs studied.
Fig. 2. Aotus phenotype from Quindío (karyomorph 9, KM9, 2n = 50).

262 / Torres et al.
Although the Q- and G-banding patterns were usually similar remarkabledifferences were the variable heterochromatic Q-bright bands at the ends of theshort arms and several interstitial bright bands, usually C-positive but not al-ways G-positive.
The R-banding patterns from bromodeoxiuridine terminal treatment (RBH-FPG) did not represent in all cases the reverse of the G-banding patterns (GTG);several chromosomes showed some similar and corresponding R-positive and G-positive bands. Good quality R bands (RHG) were not usually obtained, and opti-mal conditions could be reproduced only in isolated cases.
Karyomorph 2. This karyomorph was defined from three different diploidnumbers, 54, 53, and 52 [Reumer and De Boer, 1980], corresponding to the karyo-types II, III, and IV of Ma et al. [1976]. We observed it in specimens from theCosta Atlántica. Following the criteria of Hershkowitz [1983], we feel these speci-mens correspond to Aotus lemurinus griseimembra. Figure 4a shows a QFQ karyo-type, which has five pairs each of metacentric and submetacentric chromosomesand 16 pairs of acrocentric ones. The chromosome numbering was determinedaccording to our proposed criteria. Bright heterochromatic terminal bands wereobserved in chromosomes 1p, 2p, 5p, 6p, 7p, 8p, 9p, and 10p; all of them wereheteromorphic. Figure 4b shows a composite karyotype representing the haploidset with G, R, and C banding. The X chromosome was metacentric as in otherkaryomorphs, and the Y chromosome, the smallest metacentric chromosome,showed interindividual morphologic variations due to the resolution level.
Karyomorph 3. This karyomorph exhibits a 2n = 54 [Reumer & De Boer,1980]; it is equivalent to the karyotype I of Ma et al. [1976]. We observed it inthe specimens from Amazonas. Following the criteria of Hershkovitz [1983], wefeel these specimens correspond to Aotus nancymai. Figure 5a shows a QFQ karyo-type. It has nine pairs of metacentric, two pairs of submetacentric, and 15 pairsof acrocentric chromosomes; however, herein the chromosome numbering changesaccording to our proposed criteria. Terminal bright heterochromatic bands wereobserved in chromosomes 12p, 13p, 15p, 17p, and 24p. Figure 5b shows a com-posite karyotype representing the haploid set with G, R, and C banding. Exceptfor whole heterochromatic short arms (chromosomes 12, 13, 14, 15, 17, 18, and24), no terminal or iterstitial constitutive heterochromatin was observed in thespecimens carrying this karyomorph. The centromeric heterochromatin of chro-mosomes 1, 3, 4, and 6 showed heteromorphisms. The Y chromosome, the small-
TABLE III. Chromosomal Structure of Colombian Karyomorphs Studied
Pairs of autosomes Sex chromosomesMb Sb Ab
Karyomorph 2na 1–1.9c 2–4.9c >5c X RLd Y RLd
2 54/52 5/6 5 16/14 Mb 5.5 Mb 0.83 54 9 2 15 M 5.1 M 0.66 50 5 5 14 M 5.1 M 0.97 46 7 5 10 M 5.4 NDe —8 58 4 4 20 M 5.3 M 1.49 50 9 3 12 M 5.0 NDe —a2n, diploid number.bChromosome morphology: M, metacentric; S, submetacentric; A, acrocentric.cRange of arm ratio (q/p).dRL, relative length expressed as % TCL. Total X-containing haploid complement length.eND, not determined (since only females were studied).

Chromosome Diversity of Colombian Aotus / 263
TAB
LE
IV.
Chr
omos
ome
Mea
sure
men
ts F
rom
Col
ombi
an K
aryo
mor
phs
Stud
ied*
Kar
yom
orph
2K
aryo
mor
ph 3
Kar
yom
orph
6K
aryo
mor
ph 7
Kar
yom
orph
8K
aryo
mor
ph 9
Chro
mo-
% T
CLAr
m ra
tioCh
rom
o-%
TCL
Arm
ratio
Chro
mo-
% T
CLAr
m ra
tioCh
rom
o-%
TCL
Arm
ratio
Chro
mo-
% T
CLAr
m ra
tioCh
rom
o-%
TCL
Arm
ratio
som
ex
sx
sso
me
xs
xs
som
ex
sx
sso
me
xs
xs
som
ex
sx
sso
me
xs
xs
15,
50,
311,
40,
141
8,8
0,34
1,0
0,05
16,
40,
301,
50,
131
8,9
0,36
1,2
0,08
13,
80,
101,
20,
081
9,2
0,52
1,2
0,09
24,
20,
181,
90,
292
8,3
0,45
1,1
0,08
25,
70,
221,
20,
082
6,6
0,33
1,7
0,17
23,
60,
181,
20,
122
8,5
0,44
1,2
0,08
33,
80,
291,
30,
233
5,5
0,30
1,3
0,14
34,
30,
341,
90,
233
5,9
0,22
1,1
0,11
33,
20,
221,
10,
153
5,4
0,32
1,3
0,19
43,
30,
171,
10,
144
4,0
0,23
1,9
0,18
44,
00,
141,
40,
154
5,1
0,37
1,0
0,09
42,
60,
131,
10,
044
5,0
0,41
1,2
0,11
52,
40,
171,
00,
075
3,7
0,27
1,7
0,20
53,
70,
221,
40,
145
4,4
0,20
1,6
0,20
56,
10,
222,
50,
235
4,4
0,24
1,6
0,21
65,
90,
403,
40,
586
3,7
0,20
1,4
0,28
65,
90,
393,
50,
796
4,2
0,22
1,6
0,23
65,
00,
182,
70,
286
3,9
0,33
1,6
0,35
75,
30,
203,
50,
357
3,7
0,28
1,2
0,21
75,
00,
302,
80,
347
3,8
0,28
1,3
0,17
74,
80,
192,
30,
227
3,8
0,26
1,9
0,29
85,
20,
193,
30,
438
3,3
0,27
1,1
0,15
84,
50,
183,
20,
368
6,0
0.19
3,9
0,65
84,
20.
082,
80,
268
3,7
0,20
1,2
0,14
94,
60,
243,
20,
279
2,3
0,25
1,9
0,42
94,
20,
232,
30,
609
4,9
0,30
3,1
0,20
94,
90,
1869
,230
,78
93,
30,
411,
10,
1110
4,2
0,31
3,2
0,30
105,
50,
564,
50,
9010
3,9
0,25
3,4
0,53
104,
50,
193,
30,
7010
4,4
0,11
3,0
0,21
105,
70,
283,
40,
4911
4,5
0,21
6,6
1,57
113,
80,
303,
20,
5011
5,7
0,29
8,0
2,20
114,
10,
323,
90,
9511
4,3
0,14
26,3
17,1
411
4,0
0,29
3,3
0,49
124,
50,
2554
,727
,96
124,
80,
164,
70,
6712
4,8
0,28
12,3
8,66
123,
70,
332,
70,
3112
3,9
0,20
14,6
11,6
412
3,4
0,28
3,3
0,31
134,
40,
358,
212
,64
133,
90,
394,
20,
8213
3,9
0,24
12,7
7,06
135,
90,
315,
81,
5413
3,6
0,15
21,1
8,78
134,
50,
466,
11,
6014
3,9
0,22
6,1
1,40
143,
50,
296,
01,
5214
3,9
0,23
4,0
0,71
144,
20,
192,
70,
4814
3,4
0,14
15,4
14,7
314
4,3
0,30
7,6
2,50
153,
90,
196,
53,
4115
3,3
0,17
4,7
1,62
153,
60,
2514
,411
,05
153,
40,
1819
,910
,43
153,
20,
1127
,019
,22
154,
00,
396,
31,
8516
3,4
0,19
14,5
16,7
416
3,2
0,20
20,1
21,2
016
3,5
0,30
10,7
9,59
163,
30,
2029
,514
,59
163,
10,
0931
,815
,42
163,
80,
316,
62,
4717
3,3
0,26
37,3
22,3
317
3,0
0,20
6,0
1,46
173,
40,
2112
,77,
8017
3,2
0,18
17,8
9,83
173,
10,
1025
,320
,73
172,
90,
365,
22,
4218
3,2
0,26
18,5
14,0
818
2,9
0,29
5,1
4,68
182,
90,
2815
,26,
3718
2,6
0,15
19,9
10,5
718
3,1
0,19
2,3
0,28
182,
70,
226,
82,
8019
2,7
0,24
18,8
16,0
219
2,8
0,89
9,5
7,24
192,
90,
2115
,011
,46
192,
50,
2521
,010
,72
193,
00,
2026
,419
,31
192,
60,
2911
,64,
4220
2,6
0,33
24,3
18,0
820
2,7
0,22
6,1
2,77
202,
70,
3014
,310
,84
202,
50,
2217
,98,
8920
2,7
0,14
19,7
11,6
420
2,4
0,27
12,5
5,87
212,
60,
2121
,416
,62
212,
60,
186,
55,
3821
2,6
0,22
15,5
11,7
221
2,5
0,19
12,5
10,0
121
2,7
0,16
24,3
15,4
421
2,3
0,27
15,3
7,32
222,
50,
2426
,416
,90
222,
50,
145,
72,
4822
2,6
0,28
15,2
11,7
522
2,4
0,22
14,9
6,04
222,
70,
2828
,113
,51
222,
10,
2614
,65,
4123
2,4
0,40
22,8
16,8
923
2,1
0,16
7,0
3,89
232,
50,
3021
,713
,39
X5,
40,
451,
30,
0823
2,5
0,12
15,4
9,37
231,
60,
188,
02,
7624
2,2
0,09
28,3
14,4
424
2,1
0,29
3,1
1,01
242,
40,
2513
,28,
1524
2,4
0,17
17,1
10,7
624
1,5
0,15
11,4
3,35
252,
20,
1727
,911
,26
251,
80,
1912
,38,
78X
5,1
0,29
1,2
0,12
252,
30,
1620
,311
,80
X5,
00,
321,
20,
1426
1,8
0,15
25,6
9,19
261,
20,
209,
27,
15Y
0,9
0,12
1,1
0,14
162,
20,
1417
,89,
84X
5,2
0,26
1,3
0.09
X5,
10,
191,
30,
1027
2,1
0,12
16,8
4,91
Y0,
80,
141,
10,
09Y
0,6
0,32
1,1
0,53
281,
80,
157,
87,
49X
5,3
0,10
1,3
0,08
Y1,
40,
161,
20,
14
*%TC
L, r
elat
ive
leng
th (%
tota
l chr
omos
ome
leng
th);
x, m
eans
; s, s
tand
ard
devi
atio
n.
264 / Torres et al.
Fig. 3. Partial metaphases showing (a) Positive silver nucleolus organizing regions Ag-NORs from KM2(2n = 54 and (b) QFQ and sequential CBG from KM3 (2n = 54). Arrows show positive C band in the second-ary constriction or nucleolus organizing regions.
est metacentric chromosome, showed a late replication arm, but whether it cor-responded to p or q could not be established.
Karyomorph 6. Recognized previously [Reumer & De Boer, 1980], thiskaryomorph shows a 2n = 50 chromosomes. It was observed in specimens fromMeta. Following the criteria of Hershkovitz [1983], we feel these specimens cor-respond to Aotus brumbacki. Figure 6a shows a QFQ karyotype, which has fivepairs each of metacentric and submetacentric chromosomes and 14 pairs of acro-centric ones. Bright heterochromatic bands were observed in 1p, 2p, 3p, 6p, 7p,8p, 10p, 11p, and 14q chromosomes (interstitial in the last one). However, thesebands were not C-positive in chromosomes 3, 8, and 10. Figure 6b shows a compos-ite karyotype representing the haploid set with G, R, and G banding. In one of themales studied, the Q-bright and C-positive interstitial heterochromatic band of chro-mosome 14q was heteromorphic; this chromosome also had a heteromorphic wholeheterochromatic short arm. Chromosome 11p was also completely heterochromatic.One male was heterozygous for a chromosome 13q paracentric inversion, centromereproximal. Chromosome Y was the smallest metacentric chromosome.
Karyomorph 7. Recognized previously [Reumer & De Boer, 1980], thiskaryomorph corresponds to karyotype V described by Ma et al. [1976]. With a 2n= 46, it shows the lowest diploid number of the genus Aotus. We observed it is afemale specimen from Caquetá. Following the criteria of Hershkovitz [1983], we

Chromosome Diversity of Colombian Aotus / 265
Fig. 4. Banding patterns from karyomorph KM 2 (2n = 54): (a) Q banding and (b) G, R, and C bandingfrom a haploid set (including sex chromosomes).

266 / Torres et al.
Fig. 5. Banding patterns from karyomorph KM 3 (2n = 54): (a) Q banding and (b) G, R, and C bandingfrom a haploid set (including sex chromosomes).

Chromosome Diversity of Colombian Aotus / 267
Fig. 6. Banding patterns from karyomorph KM 6 (2n = 50): (a) Q banding and (b) G, R, and C bandingfrom a haploid set (including sex chromosomes).
feel this specimen corresponds to Aotus vociferans. Figure 7a shows a QFQ karyo-type, which has seven pairs of metacentric, five pais of submetacentric, and tenpairs of acrocentric chromosomes. Bright heterochromatic bands were observedin chromosomes 2p, 5p, 8p, and 9p. These bands were also C-positive and hetero-morphic. Figure 7b shows a composite karyotype representing the haploid setwith G, R, and C banding. The X chromosome was identified by its late replica-

268 / Torres et al.
Fig. 7. Banding patterns from karyomorph KM 7 (2n = 46): (a) Q banding and (b) G, R, and C bandingfrom a haploid set (including sex chromosomes).
tion. The whole heterochromatic short arms of chromosomes 13 and 14 were het-eromorphic. Chromosomes 2, 3, 5 and 6 show prominent centromeric bands.
Karyomorph 8. This karyomorph exhibits the highest diploid number ofthe genus Aotus, 2n = 58 chromosomes. It was observed in four specimens fromBoyacá [Giraldo et al., 1983]. These specimens were considered Aotus hershkovitziby Groves [1993]. Figure 8a shows a QFQ karyotype, which presents four pairs

Chromosome Diversity of Colombian Aotus / 269
Fig. 8. Banding patterns from karyomorph KM 8 (2n = 58): (a) Q banding and (b) G, R, and C bandingfrom a haploid set (including sex chromosomes).

270 / Torres et al.
each of metacentric and submetacentric chromosomes and 20 pairs of acrocentricones. Bright heterochromatic bands were observed, interstitial bands in the 20qchromosome and terminal bands in chromosomes 2p, 5p, 6p, 7p, 8p, and 25q.Two of four animals, a male and a female, presented a heteromorphic terminalband in the Xp chromosome. Figure 8b shows a composite karyotype represent-ing the haploid set with G, R, and C banding. Constitutive heterochromatin waslocalized pericentromerically (in all chromosomes), terminally 10p, 4q, 5p, 6p,7p, 8p, 25q, and Yq), and interstitially (20q, 27q). CBG heteromorphic bands wereobserved (chromosomes 2, 3, 4, 5, 6, 7, 10, 18, 20, and 27), showing size varia-tions, or sometimes were absent. The whole heterochromatic short arms of chro-mosomes 10 and 18 do not have very late replication. Of the karyomorphs studied,this karyomorph presented the longest Y chromosome, which was also the small-est metacentric chromosome.
Karyomorph 9. This previously undescribed karyomorph exhibits a diploidnumber of 50 chromosomes, with a chromosome constitution different from allpublished Aotus karyotypes. It was observed in a female specimen from Quindío,(Aotus sp.). Figure 9a shows a QFQ karyotype, which has nine pairs of metacen-tric chromosomes, as well as three pairs of submetacentric and 12 pairs of acro-centric ones. Intense fluorescent terminal bands were present in the 8p, 10p,11p, 12p, 13p, 14p, and 15p chromosomes. For chromosomes 8, 13, 14, and 15,these bands correspond to C-positive constitutive heterochromatin. Figure 9bshows a composite karyotype representing the haploid set with G, R, and C band-ing. As in other karyomorphs, the X chromosome was identified by its late repli-cation and band pattern. Chromosomes 13q and 22q show interstitial C bands,proximal to the centromere, while chromosomes 13–18 show heteromorphic wholeheterochromatic short arms. The centromeric heterochromatic band of chromo-some 5p is also heteromorphic.
A comparison of chromosome nomenclature equivalence with previous reportsis presented in Table V.
DISCUSSIONThe existence of six species of Aotus in Colombia is clearly possible from the
results presented here as well as from the comparisons of these karyomorphswith those reported in the literature. We have identified eight karyotypes thatfit six different karyomorphs in Colombian Aotus based on chromosomal diver-sity. Metacentric chromosomes ranged from four to nine pairs, submetacentricsfrom two to five, and acrocentrics from 10–20.
To date, there is not a uniform system of cytogenetic nomenclature for thegenus. We intended to establish a uniform nomenclature system that followsunique criteria for chromosome denomination in the karyomorphs studied. Thisdoes not imply, however, that one particular chromosome maintains the samedenomination in different karyomorphs. This was not possible because of chro-mosome diversity and the possibility of representing different species.
The major stumbling block for developing a single general standard number-ing system for all karyomorphs observed was the great variability in chromo-somal structure. For example, karyomorphs 2 and 3 have equal diploid numbersof chromosomes and yet show differences that were impossible to consolidateunder the same numbering system. The same situation occurs for karyomorphs6 and 9. Identical homologous chromosomes cannot conserve the same denomi-nation in different karyomorphs without causing confusion regarding the rear-ranged chromosomes. The equivalences for identical homologous chromosomes of

Chromosome Diversity of Colombian Aotus / 271
Fig. 9. Banding patterns from karyomorph KM 9 (2n = 50): (a) Q banding and (b) G, R, and C bandingfrom a haploid set (including sex chromosomes).
the different karyomorphs of the genus found in Colombia are reported by Torreset al. [in press].
We did, however, develop a numbering system for each karyomorph or chromo-some taxon in which each chromosome has a unique designation based on sequen-tial banding patterns and chromosome measurements (size and morphology).

272 / Torres et al.
Previous nomenclature systems of Ma et al. [1976] and Reumer and De Boer[1980] do not meet our analytical needs.
The nomenclature system of Ma et al. [1976] poses a few problems. First thecriterion for establishing the A and B chromosome groups does not define thelimits in arm ratio, and consequently there are mixtures of chromosomes witheuchromatic and heterochromatic short arms in the B group. Second, some de-nominations are not represented in certain karyotypes, and thus the chromo-some denomination does not keep a relationship with the diploid number. Finally,it is difficult to distinguish a numeric polymorphism within one karyomorph fromthe chromosome variation found in different karyomorphs.
Reumer and De Boer [1980] did not succeed in unifying the nomenclatureaccording to a standard karyotype for two reasons. First, their approach did nottake into account differences in karyotype structure (i.e., karyomorphs with similardiploid numbers but qualitatively different chromosomes or karyotypes having a
TABLE V. Aotus Karyomorph Found in Colombia: Chromosome NomenclatureEquivalences With Previous Reports
Karyomorph
KM2 KM3 KM7 KM64b 9b 9b 4b 9b 4b 9b
TRa II 6b K2 (standard) TRa I 11b 12b (standard) TRa V 11b (standard) TRa 5b
25p1 2 1 2 18q 1 2 2 1i 6/7 1 1 1 8/11 1 112 3 2 3 1 2 1 1 2i 8/11 2 2 2 9/25 2 53 4 3 4 2 3 3 3 4 4/15 3 3 3 4/15 3 124 5 4 5 3 4 4 4 6 4 4 4 21/22 4 135 6 5 6 4 5 5 5 5 5 5 5 1 5 16 7 6 7 5 6 6 6 8 3 6 7 7 20/27 6 27 8 7 9 6 7 7 7 7 2 7 8 8 2 7 38 9 8 8 7 8 8 8 9 8 10 10 10 8 49 12 9 12 10 9 10 10 23 9 12 12 6 9 1010 11 10 13 12 10 11 11 3 5 10 11 11 12 10 611 10 12 10 9 11 14 14 11 12 11 13 13 3 11 1412 13 11 1q 8 12 12 12 10 9 12 6 6 7 12 1513 14 13 1p 11 13 13 13 13 13 9 9 13 1614 15 14 11 16 14 16 18 15 14 17 17 14 1915 16 15 14 13 15 15 17 16 18 15 15 15 15 1816 17 16 15 17 16 19 19 12 16 16 16 16 1717 18 17 16 14 17 18 15 14 17 14 14 14 17 2018 19 19 17 15 18 17 9 18 18 21 20 18 2219 21 20 19 19 19 23 21 19 19 18 19 19 2120 23 23 20 22 22 17 20 22 21 20 2321 24 21 21 20 21 20 23 21 21 19 18 21 822 22 22 20 21 22 21 16 20 22 20 22 22 923 20 18 23 23 23 9 20 22 23 724 26 24 22 22 24 24 24 24 25 2425 25 25 24 26 25 25 25 2526 27 26 25 27 26 26 26 26X X X X X X X X X X X X XY Y Y Y Y Y YaTR, this report.bReferences as cited in Table I: 4, Ma et al. [1976]; 5, Yunis et al. [1977]; 6, Miller et al. [1977]; 9, Reumerand De Boer [1980]; 11, Pieczarka et al. [1992]; 12, Dutrillaux and Couturier [1981].

Chromosome Diversity of Colombian Aotus / 273
higher number than the standard karyotype), making it difficult to define somekaryomorphs. Second, the difficulty of identifying several chromosomes withintrachromosomal rearrangements is not addressed. In this system, chromosomeswhich have suffered inversion or other intrachromosome changes keep the samedenomination corresponding to the standard element; thus, fundamentally differentchromosomes have the same denomination. Although those chromosomes were de-nominated as ‘‘variant chromosomes’’ ( a term that is theoretically incorrect, as theauthors themselves pointed out), this feature makes the nomenclature ambiguous.Moreover, the authors recognize a weakness in their nomenclature when they findone or two putative chromosome pairs in karyomorph 3 which are not present inany other karyomorph.
It is well known that there are not direct relationships among banding pat-terns. In independent studies with different chromosome identification methods,one particular chromosome of the same established karyotype could have differ-ent identity. For example, chromosome 2 (GTG) karyotype I of Ma et al., [1976]does not correspond to chromosome 2 (RTBA) karyotype I of Dutrillaux and Cou-turier [1981]. Such ambiguity can be solved by establishing a chromosome no-menclature system based on sequential banding. In this proposal, chromosomedenomination is based on G, R, Q, and C sequential banding patterns and chro-mosome measurements. Through this methodology, a precise correspondence ofdifferent banding patterns is achieved.
CONCLUSIONS1. Six different Aotus karyomorphs were found in Colombia. In order to elu-
cidate whether they represent different species, it is necessary to observe a largersample, both in the number of specimens and the number of regions.
2. It was not possible to generate a standard, single, cytogenetic nomencla-ture for the genus Aotus because of the chromosomal differences of the observedkaryomorphs, but a nomenclature for each one of the recognized karyomorphswould be effective.
3. The nomenclature system based on the sequential banding pattern size,and morphology of the chromosomes proposed for each one of the six karyomorphsaccommodates all existing information to date.
ACKNOWLEDGMENTSWe are very grateful to the genetics group of the Instituto Nacional de Salud
for collaboration, to Camenza Murillo for the Favin extract preparation to LuisFernando Cadavid and Mario Contreras, who participated in the earlier part ofthe work, to Gloria Pérez for her collaboration in the preparation and measure-ment of karyotypes, to Cecilia Ramirez from the Instituto de Ciencias Naturalesfor her collaboration in the preservation of the specimens, to Jorge Gardeazábalfrom Zoológico de Cali, to Fernando Nassar and Juanita Roda from Centro deRehabilitación WSPA (World Society for the Protection of Animals) and to TobiasMojica, Ph.D., for his critical reading of the manuscript.
The work was supported by the Instituto Nacional de Salud and by theUniversidad Nacional de Colombia (OMT).
REFERENCESArango, M.: Moreno, M.C. Propiedades
mitogénicas y leucoaglutinantes en lin-focitos humanos de la lectina del haba
(Vicia fava). Tesis de grado, Depto deQuímica Facultad de Ciencias, Univer-sidad Nacional de Colombia, 1977.

274 / Torres et al.Arrighi, F.E.; Hsu, T.C. Localization of het-
erochromatin in human chromosomes. CY-TOGENETICS 10:81–86, 1971.
Bogart, M.H. Chromosomes of an owl mon-keys. ZOONOOZ 51:15, 1978.
Brumback, R.A. A third species of the owlmonkey (Aotus). JOURNAL OF HEREDI-TAS 65:321–323, 1974.
Brumback, R.A. Fluorescent banding of thekaryotype of Aotus trivirgatus (Humboldt1812): A preliminary study. JOURNAL OFHEREDITAS 66:247–248, 1975a.
Brumback, R.A. Giemsa banding pattern ofthe karyotype of Aotus griseimembra(Eliot, 1912): A preliminary study. JOUR-NAL OF HUMAN EVOLUTION 4:385–385, 1975b.
Brumback, R.A.; Staton, R.D.; Benjamin,S.A.; Lang, C.M. The chromosomes ofAotus trivirgatus Humboldt 1812. FOLIAPRIMATOLOGICA 15:264–273, 1971.
Cambefort, Y.; Moro, F. Cytogenetics andtaxonomy of some south Bolivian monkeys.FOLIA PRIMATOLOGICA 29:307–314,1978.
Caspersson, T.; Zech, L.; Johansson, C.Diffeential binding of alkylating fluoro-chromes in human chromosomes. EXPERI-MENTAL CELL RESEARCH 60:315–319,1970.
Chiarelli, B. Primi risultati di ricerche digenetica e cariologia comparata in primatie loro interesse evolutivo. RIVISTA DIANTROPOLOGIA 50:87–124, 1963.
Chu, E.H.; Bender, M.A. Chromosome cy-tology and evolution in primates. SCI-ENCE 133:1399–1405, 1961.
De Boer, L.E. Cytotaxonomy of the Platyr-rhini (Primates). GENEN EN PHAENEN17:1–115, 1974.
Descailleaux, J.; Fujita, R.; Rodriguez, L.A.;Aquino, R.; Encarnación, F. Reearregloscromosómicos y variabilidad cariotípica delgénero Aotus (Cebidae: Platyrrhini). Pp.572–577 in LA PRIMATOLOGIA EN ELPERÚ: TRABAJOS DEL CENTRO DEREPRODUCCIÓN Y CONSERVACIÓNDE PRIMATES TEMA, Vol. 4., 1990.
Dutrillaux, B.; Couturier, J. The ancestralkaryotype of platyrrhini monkeys. CYTO-GENETIC AND CELL GENETICS 30:232–242, 1981.
Egozcue, J. A note on the chromosomes ofAotus trivirgatus (Humboldt 1812). FOLIAPRIMATOLOGICA 15:274–276, 1971.
Egozcue, J.; Perkins, E.; Hagemenas, F. Thechromosomes of Saguinus fuscicollis illig-rei (Pucheran, 1845) and Aotus trivirgatus(Humboldt, 1811). FOLIA PRIMATOLO-GICA 11:154–159, 1969.
Espinal, C.; Moreno, E.; Umaña, J.; Ramirez,J.; Montilla, M. Susceptibility of differentpopulations of Colombian owl monkey to theFCB-1 strain of Plasmodium falciparum.
THE AMERICAN JOURNAL OF TROPI-CAL MEDICINE AND HYGIENE 33:777–782, 1984.
Ford, S.M. Taxonomy and distribution of theowl monkey. Pp. 1–57 in THE OWL MON-KEY. J. Baer; R. Weller; I. Kakoma, eds.London, Academic Press Inc, 1994.
Goodpasture, C.; Bloom S.E. Visualizationof nucleolar organizer regions in mamma-lian chromosomes using silver staining.CHROMOSOMA 53:37–50, 1975.
Giraldo, A.; Torres, O.M.; Bueno, M.L.;Silva, E.; Fraser, C. de C., Ramirez, J.;Espinal, C.A. Patrones de bandas Q y Cdel cariotipo de una nueva especie deAotus (2n = 58). VI Congreso Latino-americano de Genética. Maracaibo, Ven-ezuela. 1983.
Giraldo, A.; Bueno, M.; Silva, E.; Ramírez,J.; Umaña, J.; Espinal, C. Estudio citoge-nético de 288 Aotus colombianos, BIO-MEDICA 6:5–13, 1986.
Goto, K.; Akematsu, T.; Shimazu, H.; Sugi-yama, T. Simple differential Giemsa stain-ing of sister chromatids after treatmentwith photosensitive dyes and exposure tolight and the mechanism of staining.CHROMOSOMA 53:223–230, 1975.
Groves, C.P. Order Primates. Pp. 243–277in MAMMAL SPECIES OF THE WORLD:a TAXONOMIC AND GEOGRAPHICREFERENCE. D.E. Wilson; D.M. Reeder,eds. Washington, London, Smithsonian In-stitution Press, 1993.
Hernandez-Camacho, J.; Cooper, R. Thenon-human primates of Colombia. Pp 35–69 in FIELD STUDIES AND CONSERVA-TION. R. Thoringthon; P. Heltne, eds.Washington, DC, Academy of Sciences,1975.
Hershkovitz, P. Mammals of northern Co-lombia. Preliminary report #4. Monkeys(Primate) with taxonomic revision of someforms. PROCEEDINGS OF THE UNITEDSTATES NATIONAL MUSEUM 98:323–427, 1949.
Hershkovitz, P. Two new species of night mon-keys, genus Aotus (Cebidae Platyrryni): Apreliminary report of Aotus taxonomy.AMERICAN JOURNAL OF PRIMATOL-OGY 4:209–243, 1983.
Koiffmann, C.; Saldanha, P.H. Cytogeneticsof Brazilian monkeys. JOURNAL OF HU-MAN EVOLUTION 3:275–282, 1974.
Ma, N.S.F. Chromosome evolution in the owlmonkey, Aotus. AMERICAN JOURNAL OFPHYSICAL ANTHROPOLOGY 54:293–303,1981.
Ma, N.S.F.; Jones T.C.; Miller, A.; Morgan,L.; Adams, E. Chromosome polymorphismand banding patterns in the owl monkey(Aotus). LABORATORY ANIMAL SCI-ENCE 26:1022–1036, 1976.
Ma, N.S.F.; Rossan, R.N.; Kelley, S.T.;

Chromosome Diversity of Colombian Aotus / 275Harper, J.S.; Bedard, M.T.; Jones, T.C.Banding patterns of the chromosomes oftwo new karyotypes of the owl monkey,Aotus, captured in Panamá. JOURNAL OFMEDICAL PRIMATOLOGY 7:146–155,1978.
Ma, N.S.F.; Renquist, D.M.; Hall, R.; Sebgal,P.K.; Simone, T.; Jones, T.C. XX/XO sexdetermination system in a population ofPeruvian owl monkey, Aotus. JOURNALOF HEREDITY 71:336–342, 1980.
Ma, N.S.F.; Aquino, R.; Collins, W.E. Two newkaryotypes in the Peruvian owl monkey(Aotus trivirgatus). AMERICAN JOURNALOF PRIMATOLOGY 9:333–341, 1985.
Miller, C.K.; Miller, D.A.; Miller, O.J.; Tan-travahi, R.; Reese, R.T. Banded chromo-somes of the owl monkey, Aotus trivirgatus.CYTOGENETICS AND CELL GENETICS19:215–226, 1977.
Minezawa, M.; Valdivai, J. Cytogeneticstudy of the Bolivian monkeys: I. Prelimi-nary report on karyotypes of Cebus apella,Saimiri sciureus, Aotus azarae and Saguin-us labiatus. Kyoto University OverseasReseach. REPORTS OF NEW WORLDMONKEYS 4:53–67, 1984.
Moorhead P.S.; Nowell, P.C.; Mellman, W.J.;Battips, D.M.; Hungerford, D.A. Chromo-some preparations of leukocytes culturesfrom human peripheral blood. EXPERI-MENTAL CELL RESEARCH 20:613–616,1960.
Mudry, M.P.; Colillas, O.J.; Brieux, S.S. TheAotus of northern Argentina. PRIMATES25:530–537, 1984.
Pieczarka J.C.; Nagamachi C.Y. Cytogeneticstudies of Aotus from eastern Amazonia:Y/autosome rearrangement. AMERICANJOURNAL OF PRIMATOLOGY 14:255–263, 1988.
Pieczarka, J.C.; Barros, R.M.; Nagamachi
C.Y.; Rodriguez, R.; Espinel, A. Aotusvociferans × Aotus nancymai: Sympatrywithout chromsomal hybridation. PRI-MATE 33:239–245, 1992.
Pieczarka, J.C.; Barros, R.M.; Faria J.R.;Nagamachi, C.Y. Aotus from the south-western Amazon region is geographicallyand chromosomally intermediate betweenA. azarae boliviensis and A. infulatus. PRI-MATES 34:197–204, 1993.
Reumer, J.W.; De Boer, L.E. Standardizationof Aotus chromosome nomenclature with de-scriptions of the 2n = 49–50 karyotype andthat of a new hybrid. JOURNAL OF HU-MAN EVOLUTION 9:461–482, 1980.
Seabright, M. A rapid banding technique forhuman chromosomes. LANCET 2:971,1971.
Sehested, J. A simple method for R-band-ing of human chromosomes showing a pH-dependent connection between R and Gbands. HUMAN GENETICS 21:55–58,1974.
Sumner, A.T. A simple technique for dem-onstrating centromeric heterochromatin.EXPERIMENTAL CELL RESEARCH75:304–306, 1972.
Tjio, J.H.; Levan, A. The chromosome num-ber of man. HEREDITAS 42:1–6, 1956.
Torres, O.M.; Mojica, T.; Ruiz F.; Enciso, S.;Yunia, I. Chromosomal intrepretation ofAotus phylogeny. AMERICAN JOURNALOF PRIMATOLOGY (submitted).
Willard, H.; Latt, S. Analyses of deoxyri-bonucleic acid replication in human ×chromosomes by microscopy fluorescence.AMERICAN JOURNAL OF HUMAN GE-NETICS 28:213–227, 1976.
Yunis, E.; Torres, O.M.deC.; Ramirez, C.Genus Aotus: Q and G-band karyotypesand natural hybrids. FOLIA PRIMATOL-OGICA 27:165–177, 1977.



















