
Topoisomerase Type II Structures Comparison & Implications of the ’07 Berger & Dong Model.
31
Topoisomerase Type II Structures Comparison & Implications of the ’07 Berger & Dong Model
-
Upload
regan-wilmer -
Category
Documents
-
view
217 -
download
1
Transcript of Topoisomerase Type II Structures Comparison & Implications of the ’07 Berger & Dong Model.

Topoisomerase Type II Structures
Comparison & Implications of the ’07 Berger & Dong Model

Topoisomerase: what is it?
• Enzymes within cells that remove excessive supercoiling—and thereby torsional stress—while maintaining sufficient supercoiling to ensure a stable compact genome.– These enzymes are apparent in every cellular function involved with
DNA—transcription, replication, recombination, and chromosome separation.
– The enzymes complete their job in any of the above function by cleavage; that is, cutting.
– Type I topoisomerase cut only 1 strand of DNA, Type II topoisomerase cut both (2) strands of DNA.

What’s Our Type?
• Type I Isomerase – lA: links to 5’-phosphate
terminus, relaxes negatively supercoiled DNA, introduces transient breaks in ss region of DNA
– IB: links 3’ terminus covalently to tyrosol group, relaxes both negatively and positively supercoiled DNA, segment can rotate with respect to protein to relieve torsional stress
– Neither require ATP energy– Both cut only 1 strand of DNA
• Type II Isomerase– IIA: found in eukaryotes,
highly expressed in cancerous cells
– IIB: structurally and biochemically distinct from Type IIA, found in archaea and some higher plants
– Both require ATP energy to activate
– Both perform a double-stranded cut of DNA

What’s Our Type?
• Type I Isomerase – lA: links to 5’-phosphate
terminus, relaxes negatively supercoiled DNA, introduces transient breaks in ss region of DNA
– IB: links 3’ terminus covalently to tyrosol group, relaxes both negatively and positively supercoiled DNA, segment can rotate with respect to protein to relieve torsional stress
– Neither require ATP energy– Both cut only 1 strand of DNA
• Type II Isomerase– IIA: found in eukaryotes,
highly expressed in cancerous cells
– IIB: structurally and biochemically distinct from Type IIA, found in archaea and some higher plants
– Both require ATP energy to activate
– Both perform a double-stranded cut of DNA

Topoisomerase Type IIA
• Mechanism: A two-gate structure, Type II topoisomerases work by cleaving and opening one DNA duplex, passing a second duplex through the opening, and then resealing the break.

Topoisomerase Type IIA
• Function: Type II topoisomerase performs a cycle of DNA cleavage, transport, and religation (coupled with ATP binding and hydrolysis) to preserve the topological invariant of the DNA– That is, it relaxes positively and negatively charged supercoiled
DNA and performs cantenations/decantenations to prevent torsional stress and keep the cell’s compact genetic structure
• This is an essential function for cell functionality and survival, and so all organisms have at least one type of Type II topoisomerase!

Berger & Dong 2007 Model
• Summary• Conclusions• Impact• Limitations• Comparison to Previous Models
– Berger et al Model 1996– Rybenkov et al Model 1997– Vologodskii et al Model 2001

Berger and Dong 2007 Model
Summary• The molecular understanding of DNA binding
to Topo II was examined• Crystallization of the Topo II-DNA complex
was performed• Crystallography determines the arrangement of
atoms in solids

Berger & Dong 2007 Model
• Summary• Conclusions• Impact• Limitations• Comparison to Previous Models
– Berger et al Model 1996– Rybenkov et al Model 1997– Vologodskii et al Model 2001

Berger and Dong 2007 Model
Conclusions• Upon binding, Topo II
distorts the DNA at ~150 degree global angle
• The mechanism of this bend very similar to remodeling proteins
• Along with the change in DNA’s conformation, Topo II undergoes a conformational change
• This change forms a catalytic site for the ‘cutting’ reaction
Bending of DNAChange in Top2’s conformation (shape)

Berger & Dong 2007 Model
• Summary• Conclusions• Impact• Limitations• Comparison to Previous Models
– Berger et al Model 1996– Rybenkov et al Model 1997– Vologodskii et al Model 2001
Berger and Dong 2007 Model
• The researchers concluded that their findings:– Corroborated previous biochemical findings about
Topo II– Established the structure as mechanism intermediate– Discovery of DNA cleavage controls– ***Control of DNA topology
• Is Topo II attracted to curvature OR does it cause the curvature? (Which came first, the chicken or the egg?)
• Question is resolved….Topo II CAUSES curvature! (via bending)
• This bending provides a means to decatenation and relaxation
• Preferred binding sites @ highly curved DNA crossovers and loops in vivo probably arise from intrinsic deforms DNA

Berger & Dong 2007 Model
• Summary• Conclusions• Impact• Limitations• Comparison to Previous Models
– Berger et al Model 1996– Rybenkov et al Model 1997– Vologodskii et al Model 2001

Berger and Dong 2007 Model
Limitations• Only a portion of Topo II was examined
– Fragment used was ~720 aa long (residues 408-1177)
• Experiment performed in vitro• Model organism used was S. cerevisiae, or
yeast (is structure/binding/bending conserved with other organisms?

Berger & Dong 2007 Model
• Summary• Conclusions• Impact• Limitations• Comparison to Previous Models
– Berger et al Model 1996– Rybenkov et al Model 1997– Vologodskii et al Model 2001

Berger Model 1996
• One of the researchers of Dong/Berger model published a report on the Structure and Mechanism of Topo II 12 years ago
• The Topo II examined was also S. cerevisiae of similar size to 2007 model
• In 1996, researchers thought conformation changes occurred after passage of T-segment, not upon binding of G-segment
• Bending of DNA upon binding with Topo II was not detected
1996 cartoon
2007 cartoon

Rybenkov Model 1997
• This study didn’t specifically look at the structure of Topo II, but it examined implications of its mechanism
• Rybenkov et al concluded that Topo II doesn’t simply catalyze the passing of one DNA strand through another, changing its local conformation and topology, but it controls the global property of DNA as well
• Suggested Topo II catalyzed the passage of one strand through another if resulting crossing angles were favorable
• Suggested Topo II shortened DNA • Topo II binds three (3) DNA
segments (crossover complex)
Top2 bound to crossover

Vologodskii Model 2001
• This study attempted to use its model to explain previous experimental data
• Computer Analysis of enzyme performed– Discrete worm-like chain– Monte Carlo procedure
simulated possible DNA conformations
– Found that simplification of DNA topology by Topo II not possible without distortion of DNA
– Suggested that Topo II create asymmetry in DNA upon binding
Simulation of DNA
DNA distortion by Topo II

Applications of the Berger & Dong 2007 Model
• Summary of Topoisomerase Type II pharmaceuticals
• Existing limitations with current Topoisomerase Type II-focused medications• Increasing drug efficiency with Berger &
Dong’s current Topoisomerase Type II model

Pharmaceuticals Summary
• Mammalian type II topoisomerases are targets of anti-tumor drug classes
- aminoacridines, anthracenediones, ellipticines, and epipodophyllotoxins.
- many of them interact with DNA by an intercalative mode
• Bacterial type II toposiomerases are targets of antibacterial gents
- quinolones and flouroquinolones.

• DNA intercalators are used in chemotherapeutic treatment to inhibit DNA replication in rapidly growing cancer cells.
Ethidium intercalated between two adenine-uracil base pairs.

How does drug work on DNA?
• The drug ,localized precisely at the site of DNA cleavage, prevents topoisomerase from rejoining DNA breaks.
• Cleavage complexes themselves are not sufficient to cause cell death.
• Active cellular processes (transcription/replication) are necessary to transform cleavage complexes into cytotoxic lesions
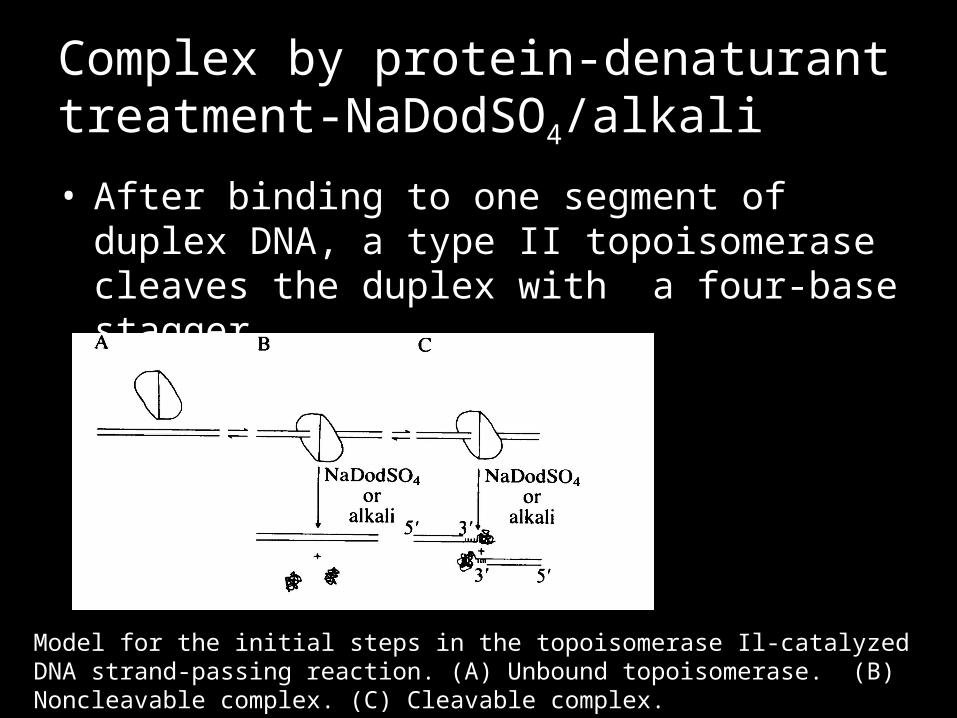
Complex by protein-denaturant treatment-NaDodSO4/alkali
• After binding to one segment of duplex DNA, a type II topoisomerase cleaves the duplex with a four-base stagger.
Model for the initial steps in the topoisomerase Il-catalyzed DNA strand-passing reaction. (A) Unbound topoisomerase. (B) Noncleavable complex. (C) Cleavable complex.

Current Limitations
• Multi-drug resistance
- Altered topoisomerase II complex activities
- results in decreased ability to form a stable ternary complex
- current measurements of topoisomerase II not reveal the relevant qualitative change

Current Limitations
• Unrecognized cofactors may influence topoisomerase II activity and confound analyses.
• Drug-topoisomerase II interactions and the precise defects in cells( altered activity) are not understood completely.

Pharmaceutical Impact
• With the current Berger and Dong model for Topoisomerase Type II, we can:– Examine complex DNA and see if its
geometry is changed by experimental drugs– Test inhibitors for Topoisomerase Type II to
prevent DNA bending and kill the cell– Prevent rigidity in Topoisomerase Type II
treatments due to built-up resistance

Further Research

Future Research
• Multicellular Confirmation– Is Berger and Dong’s
model conserved when the experiment is repeated on multicellular organisms
• Enzyme Shift– Following Berger and
Dong’s model bending DNA, does the enzyme undergo another conformational change prior to cutting the DNA

Future Research
• Chemical basis of DNA cleavage
• Benefits of unidirectional transport – not understood
• Visualization of DNA cleavage

References- “Structure and mechanism of DNA topoisomerase II”
+ James Berger & Ken Dong; Nature, Vol. 379, Jan. 1996, pp. 225 – 232- “Simplification of DNA Topology Below Equilibrium Values by Type II Topoisomerases”
+ Valentin Rybenkov et al; Science; Vol. 277; Aug. 1997; pp. 690 – 693- “Mechanism of topology simplification by type II DNA topoisomers”
+ Alexander Vologodskii et al; PNAS; Vol. 98 No. 6; Mar. 2001; pp. 3045 – 3049
- “An Antitumor Drug-Induced Topoisomerase Cleavage Complex Blocks a Baccteriophage T4 Replication Fork in Vivo”
+ George Hong & Kenneth Kreuzer; Molecular and Cellular Biology; Vol 20 No. 2; Jan. 2000; pp.594-603
- “Involvement of DNA topoisomerase II in the selective resistance of a mammalian cell mutant to DNA minor groove ligands: lingand-induced DNA—protein crosslinking and responses to topoisomerase poisons
+ Paul J. Smith et al; Carcinogenesis; Vol 11 No. 4; pp.659-665- “Multidrug Resistance Associated With Altered Topoisomerase II Activity—Topoisomerases II
as Targets for Rational Drug Design”+ Kenneth Cowan & Charles Marrow; National Cancer Institute; February 28, 1990.
- “Mechanism of antitumor drug action: Poisoning of mammalian DNA topoisomerase II on DNA by 4’-(9-acridinylamino)-methanesulfon-m-anisidide”
+ Eric M. Nelson et al; Proc. Natl. Acd. Sci. USA; Vol 81; Mar 1984; pp.1361-1365- “Design of DNA Intercalators To Overcome Topoisomerase II-Mediated Multidrug Resistance”
+ Bruce C. Baguley et al; Journal of the National Cancer Institute; Vol 82; Mar. 1990; pp.398-402













![Arrest of Replication Forks by Drug-stabilized Topoisomerase ......(CANCER RESEARCH 49, 5077-5082. September 15. 1989] Arrest of Replication Forks by Drug-stabilized Topoisomerase](https://static.fdocuments.in/doc/165x107/602f037beadb407535717aaf/arrest-of-replication-forks-by-drug-stabilized-topoisomerase-cancer-research.jpg)





