
Tolerance to Lime - Induced Chlorosis and Drought in ...
30
Chapter 9 Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstocks Pavel Pavloušek Additional information is available at the end of the chapter http://dx.doi.org/10.5772/54793 1. Introduction Many woody crop plants such as grapevine are traditionally grown with scion varieties grafted onto rootstocks. The selection of an appropriate rootstock provides a powerful tool to manage the growth and fruiting of the scion (Jones, 2012). In the grapevine propagation, the use of rootstocks is not a new matter. The evidence of the use of rootstocks can be found out even in works written by the Roman author Columella who occupied himself with agriculture and viticulture. However, the use of rootstocks obtained a new dimension after the phylloxera calamity, which destroyed European vineyards in the second half of the 19 th century. Rootstocks were introduced to Europe after the phylloxera invasion, a pest which rapidly spread through vineyards and destroyed large areas of sensitive cultivars. At present, grafting European varieties on pathogen-resistant rootstock is a normal procedure and many rootstock varieties have been developed by plant breeders (Arrigo & Arnold, 2007). When choosing a suitable rootstock it is important to select one with a good tolerance to phylloxera and well as being to the specific climatic conditions and soil characteristics of individual vineyard sites. The rootstock connects the grafted plants with soil and influences mutual relationships. The root system of the rootstock enables the uptake of water and nutrients from soil. The rootstock also shows a marked effect on the growth intensity of grafted plants. When selecting a suitable rootstock, it is important to consider characteristics and parameters of the site. The most important of them are the following: depth of the soil horizon, water-holding capacity of soil, slope and exposure of the site, and climatic conditions. The architecture of the root system of the plant is also very important for its resistance/tolerance to drought. In the case of grapevine, selection and use of a suitable rootstock may help to solve problems of plant protection and of overcoming extreme soil conditions. Adaptability of plants © 2013 Pavloušek; licensee InTech. This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Transcript of Tolerance to Lime - Induced Chlorosis and Drought in ...

Chapter 9
Tolerance to Lime - Induced Chlorosis and Drought inGrapevine Rootstocks
Pavel Pavloušek
Additional information is available at the end of the chapter
http://dx.doi.org/10.5772/54793
1. Introduction
Many woody crop plants such as grapevine are traditionally grown with scion varieties graftedonto rootstocks. The selection of an appropriate rootstock provides a powerful tool to managethe growth and fruiting of the scion (Jones, 2012). In the grapevine propagation, the use ofrootstocks is not a new matter. The evidence of the use of rootstocks can be found out even inworks written by the Roman author Columella who occupied himself with agriculture andviticulture. However, the use of rootstocks obtained a new dimension after the phylloxeracalamity, which destroyed European vineyards in the second half of the 19th century.
Rootstocks were introduced to Europe after the phylloxera invasion, a pest which rapidlyspread through vineyards and destroyed large areas of sensitive cultivars. At present, graftingEuropean varieties on pathogen-resistant rootstock is a normal procedure and many rootstockvarieties have been developed by plant breeders (Arrigo & Arnold, 2007).
When choosing a suitable rootstock it is important to select one with a good tolerance tophylloxera and well as being to the specific climatic conditions and soil characteristics ofindividual vineyard sites. The rootstock connects the grafted plants with soil and influencesmutual relationships. The root system of the rootstock enables the uptake of water andnutrients from soil. The rootstock also shows a marked effect on the growth intensity of graftedplants. When selecting a suitable rootstock, it is important to consider characteristics andparameters of the site. The most important of them are the following: depth of the soil horizon,water-holding capacity of soil, slope and exposure of the site, and climatic conditions. Thearchitecture of the root system of the plant is also very important for its resistance/toleranceto drought. In the case of grapevine, selection and use of a suitable rootstock may help to solveproblems of plant protection and of overcoming extreme soil conditions. Adaptability of plants
© 2013 Pavloušek; licensee InTech. This is an open access article distributed under the terms of the CreativeCommons Attribution License (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use,distribution, and reproduction in any medium, provided the original work is properly cited.

to environmental conditions, e.g. their tolerance to lime, low soil pH, soil humidity, salts etc.,is very important.
Nowadays, grapevine plants are more and more influenced by various kinds of environmentalstress. The most important kinds of abiotic stress are the following: extreme temperatures ortoo high (or too low) irradiation, water logging, drought, lack of minerals in soil (theirdeficiency) and too high salinity of soil (Koyro et al., 2012). Stress can be defined as anenvironmental factor that shows a negative effect on the living organism (Levitt, 1980).
This review tries to summarise data about the adaptation of rootstocks to soil (pedological)conditions, viz. their resistance to lime-induced chlorosis and drought. Regarding the globalwarming, these properties of rootstocks are very important also under conditions of the CentralEuropean viticulture.
2. Root system of grapevine plants
The most important functions of the grapevine root system involve anchoring of plants in soil,storage of reserve substances, uptake and conduction of water and dissolved nutrients withinthe plant and synthesis of growth hormones. The root system consisted not only of older,lignified roots but also of a great number of new ones that are used above all for the uptake ofnutrients. Regarding their diameter, roots can be divided into two groups. Thick roots (withthe diameter above 2 mm) represent a great proportion of root biomass. They create the“architecture“ of the root system, enable the transport of water and nutrients and fulfil thefunction of a reserve organ. Fine roots (with the diameter below 2 mm) enable above all theuptake of water and nutrients. These are above all root hairs that develop on thicker roots.Root tips consist of root caps and apical meristem. This apical meristem assures growth anddevelopment of roots. Root growth takes place in the elongation zone which is approximatelytwo milimeter long. Root tips are also the place of synthesis of plant hormones (gibberelinsand cytokinines). These growth hormones are transported via conductive tissues into theaboveground parts of the plant and participate in processes controlling the balance betweenroots and tops, initiation of flowering, and growth and development of berries. Cell division(and, thus, the growth of roots) is controlled by auxins that are transported from tops of annualshoots into roots via phloem. The elongation growth is influenced by gibberellins that aresynthesised in roots. Root hairs are localised behind the elongation zone of roots and assurethe uptake of water and nutrients. It is capable to release organic compounds into the soil andto participate in propagation and development of microflora existing in the root zone(Pavloušek, 2011a).
The distribution of roots in soil is influenced by soil and environmental factors, e.g. tempera‐ture, degree of aeration, texture, availability of water and nutrients, pH value and frequencyand depth of tillage operations (Richards, 1983, Morlat & Jaquet, 1993).
The root system of grapevine plants is mostly created by the root system of rootstocks. Theroot system of grapevine rootstocks enables the uptake of water and nutrients from greater
Abiotic Stress - Plant Responses and Applications in Agriculture278

soil depths. Distribution of root systems of individual rootstocks enables to identify theiruptake capacity for nutrients. This means that different rootstocks have a different capacity touptake individual nutrients from soil (Somkuwar et al., 2012).
Within the framework of their response to edaphoclimatic conditions individual rootstocksalso show differences in growth capacity of their roots, water uptake, transport of water intoannual shoots, metabolic activity and storage of carbon.
Water uptake and its transport represent one of the most important functions of roots. Thedistribution of roots in soil and the root turnover are the key parameters of water uptake; theyare also important for the hydraulic redistribution (Bauerle et al., 2008a).
Root architecture refers to the spatial configuration of the root system, specifically focusing onthe geometric properties of root axes and laterals, mostly concerned with the entire root systemcharacteristics (Lynch, 1995). Typically, root distribution studies include root biomass or rootlength as a function of soil depth, distance from the plant stem, and position between neigh‐bouring plants (Bassoi et al., 2003).
The available soil volume is probably the most important factor dictating the size and thedistribution of root system (Saayman, 1982). The spatial root distribution is predominantly afunction of the soil environment, while root density is a function of rootstock (Southey &Archer, 1988).
From the viewpoint of tolerance to abiotic factors associated with climatic and soil conditionsit is important to study and understand both vertical and horizontal distribution of roots insoil. For example the Dog Ridge and Salt Creek rootstocks, which belong to Vitis champinii, putforth prolific root systems of thickness of < 2 mm and 2-5 mm in the top depths of 0-30 cm upto 60 cm away from the trunk, and later thicker roots of > 5 mm beyond 60 cm from the trunk.However, St. George, which belongs to Vitis rupestris, has less root length in all categories atall blocks horizontally up to a 150 cm distance from the trunk. In the vertical direction, SaltCreek showed the greatest root length in the category of < 2 mm at a depth of 0-30 cm, whileDog Ridge and St. George were at par for root length. However, at depths of 31-60 cm and61-90 cm, Dog Ridge produced the greatest root length, followed by Salt Creek and St. George(Simkuwar et al., 2012).
The rootstock shows a significant effect not only on the distribution of roots in soil but also onthe architecture of the root system. In Vitis rupestris, major roots create vertically a narrow angleand can penetrate deep into the bottom soil layers. On the other hand, roots of Vitis riparia aredistributed in a wide angle and most of them are situated in a shallow top layer of soil (Perold,1927). Swanepoel & Southey (1989) analysed the distribution of roots within a wider spectrumof rootstocks growing under conditions of South Africa (Table 1).
Also abiotic factors may influence growth and development of the root system. In this case,above all the vertical distribution of roots within the soil horizon is important.
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
279

RootstockNumber of
roots per m2Rooting index
Number of roots per diameter (mm) class
<0.5 0.5-2 2-5 5-10 >10
Berlandieri 13/5 2069 39.6 1792 226 38 10 3
101-14 Mgt 1604 27.1 1210 337 43 11 3
775 P 1006 44.7 839 145 16 4 2
1103 P 2660 41.9 2199 399 53 6 3
99 R 1138 28.9 833 267 30 8 0
110 R 1468 29.9 1103 319 37 9 0
140 Ru 635 20.2 483 122 21 6 3
Table 1. Distribution of roots of different rootstocks cultivated under conditions of South Africa (Swanepoel &Southey, 1989). (Rooting index = number of roots <2 mm/ number of roots ≥2 mm).
To assure a good tolerance of plants to drought and lime is necessary assure a proper devel‐opment of the root system (Figure 1). Deep loosening of soil is also very important (Figure 2).
Figure 1. Root system of a five-year-old vine in clay loam soil.
Abiotic Stress - Plant Responses and Applications in Agriculture280

Figure 2. Deep lossening of soil in clay loam soil.
An adequate care about the root system and its proper development is very important andshould be started immediately after the establishment of a new vineyard. This means that it isnecessary to create and maintain a required ratio between thick and thin roots. The root systemcan be modelled by means of a cut off of roots so that they produce more branches, the totalvolume of root mass increases and the sorption area for the uptake of nutrients grows up.Usually and most frequently, this cut off takes place during the process of deep soil looseningand/or deep supplementary fertilisation. After these operations, the total volume of roots ispartly reduced but the recovery is very quick. If, however, this intervention is too drastic, itmay show a negative effect on the overall growth process of grapevine plants. The cutoff andshortening of roots should be performed at best at the beginning of the growing season, i.e. inthe period of exogene dormancy, and should not be done every year because the optimumdevelopment of roots would be disturbed and plants could be under a permanent stress. It isrecommended to use a deep additional fertilisation in three-year cycles.
A good understanding of effects of a limited availability of water on the growth of the rootsystem and its functions has a principal influence on the selection of a suitable method ofvineyard management (Schultz, 2010) and also on the selection of a suitable rootstock. In bothtemperate and Mediterranean regions, the growth of grapevine root system takes place aboveall within the period starting on flowering and finishing at the beginning of softening of berries(Comas et al., 2010). Drought-resistant rootstocks (e.g. 1103P) show a capability to create new
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
281

roots also during the periods of summer drought (Alsina et al., 2010). In these periods, themajority of roots is formed in depths below 60 cm, where the groundwater is available (Bauerleet al., 2008b).
To understand well to principles and processes of growth and development of the root systemis therefore very important for understanding to stress situations caused by abiotic factors,especially by drought. The architecture of the root system is a genetic characteristic of root‐stocks and soil, climatic and growing conditions modify it only very slightly. In the European„cool climate viticulture“ rootstocks with Vitis rupestris in their pedigree are relatively rare inspite of the fact that their roots penetrate deep into the soil horizon. Practically, the onlyrepresentative of these rootstocks is the Moravian rootstock Schwarzmann that was selectedin Bzenec (Czech Republic) to the end of the 19th century and was very popular in formerCzechoslovakia. Unfortunately, the results of introduction of other rootstocks with Vitisrupestris in their pedigree into the European „cool climate viticulture“ have not been verysuccessful yet.
3. Tolerance of grape rootstocks to lime-induced chlorosis
Lime-induced iron chlorosis, i.e. the condition of a reduced availability of soluble iron to thegrapevine plants due high concentrations of bicarbonate ions in calcareous soils, can seriouslyimpair the health condition of vines. The lime-induced chlorosis affects yield and quality ofgrapevines growing in lots of calcareous areas world-wide (Bavaresco et al., 1994).
A high content of active and total lime in soil can induce symptoms of chlorosis ingrapevine plants and, thus, negatively influence the growth and yielding capacity ofgrapevine plants as well as the quality of harvested grapes. On the other side, however,lime present in soil participates very significantly to the manifestation of sensory proper‐ties of wine. From the geological point of view, it is a very important component of „ter‐roir“, which directly influences the character of produced wine. Wine-growing regionssituated on calcareous subsoils can be considered for localities that are very suitable forproduction of quality wine. As typical examples it is possible to mention French Cham‐pagne wine region or Czech limestone Pálava Hills.
Selection of suitable, lime-tolerant rootstocks represents one of possible ways how to react toan increased content of calcium in soil and, at the same time, to preserve quality of plants andharvested grapes. Although the genetic improvement of tolerance of rootstocks to chlorosisrepresents a very long process, it still represents the best method of fight with iron chlorosis(Nikolic et al., 2000, Pestana et al., 2003). Differences in the uptake and distribution of nutrientswithin the plant may be influenced by the variety of rootstock because it may show a ratherdifferent absorption capacity as far as individual minerals are concerned (Rizk-Alla, et al.,2001). And just these differences in the absorption capacity may be associated also with theoccurrence of the lime-induced chlorosis.
Rootstocks 333 EM and 41B (originating from crossing Vitis berlandieri x Vitis vinifera) belongedto the first rootstocks selected for a high degree of tolerance to calcium so that they could be
Abiotic Stress - Plant Responses and Applications in Agriculture282

used in vineyards established on calcareous soils. A. Ruggeri, an Italian breeder and selec‐tionist, was the author of the rootstock 140 Ru (Vitis berlandieri x Vitis rupestris), which alsoshowed tolerance to lime chlorosis (Fregoni & Bavaresco, 1986). The rootstock Fercal, selectedin France by Pouget and Ottenwaeter (1978) is one of the most modern rootstocks showing avery high degree of tolerance to calcium.
3.1. Deficiency symptoms of lime-induced chlorosis
A correct identification of symptoms of lime-induced chlorosis is one of the first presumptionsof a successful fight against this deficiency. Iron deficiency chlorosis is one of the majorproblems affecting a variety of crop species grown in calcareous soils (Gruben & Kosegarten,2002). Iron deficiency causes various morphological and physiological changes in plants(Bertamini & Nedunchezhian, 2005).
Although the symptoms of lime-induced chlorosis are visible on the whole grapevine plant,the most important ones can be probably observed on leaves.
Iron-deficient plants are characterized by the development of a pronounced intervenialchlorosis similar to that caused by magnesium (Mg) deficiency but occurring first on theyoungest leaves. Intervenial chlorosis is sometimes followed by chlorosis of the veins, causingthe whole leaf to become yellow. In severe cases, the leaves become white with necrotic lesions(Abadia, 1992).
Typical symptoms of lime-induced chlorosis are the inter-vein yellowing of leaves and adecrease in plant biomass because, under conditions of iron (Fe) deficiency, a decreasedphotosynthetic performance of plants is induced by a lower content of chlorophyll in leaves(Bavaresco & Poni, 2003). Chlorosis is a disease manifesting itself by yellowing of young leaves,whereas more mature leaves are frequently green. Plant growth is often considerably de‐pressed, independent of whether young leaves are chlorotic or green (Mengel, Bübl, Scherer,1984). The impaired formation of new leaves and restricted leaf growth is a typical and moresensitive symptom of Fe-deficiency than is leaf chlorosis (Kosegarten et al.,1998)
Chlorotic symptoms also vary from year to year as a result of environmental variables, likeyields, temperature, rains. In soils where shallow layers are less rich in CaCO3 than deeperlayers, it is likely that vines develops chlorosis only when the age and roots explore layers withpoor conditions for Fe uptake (Tagliavini & Rombola, 2001).
As the lime-induced chlorosis is the result of the relationship existing between soil conditionsand grapevine root system, it is manifested also in growth characteristics of grapevines. Thelime-induced chlorosis of grapevine was characterized by a dramatic reduction of shootgrowth, grape production and leaf Fe content, and a distribution of dry matter towards rootsmore than to the clusters (Bavaresco, Giachino, Pezzutto, 2003).
Vines growing on high-bicarbonate soil significantly reduced the dry matter production ofindividual organs and the total plant weight. Lime stress conditions increased the percentdistribution of dry matter in the stem and roots and decreased that one in the fruit (berries andcluster stems) (Bavaresco & Poni, 2003).
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
283

A high content of lime mostly causes a low availability of iron, which is a result of its non-solubility occurring in soils showing higher values of pH. Under such conditions, iron cannotbe uptaken by roots of plants (Hell & Stephan, 2003).
This lime-induced iron deficiency shows a strong effect not only on grapevine plants but alsoon some other economically important fruit species cultivated on calcareous soils. It is quitecommon also in peach, pear, quince-tree, kiwi, and citrus fruit plantations (Tagliavini &Rombola, 2001).
Lime-induced stress conditions show a strong effect on production of grapes and reduce theyield of grapes per vine. When growing grapevine on calcareous soils, a lower number ofgrapes per annual shoot depends on stress conditions existing in the preceding growing season(when the flower buds were differentiated) while a small size of grapes and berries is aconsequence of iron deficiency in the current year (Bavaresco, Presutto, Civardi, 2005).
Because the lime-induced chlorosis affects above all the growth of grapevine plants, influencesthe total leaf area capable of photosynthetic activities and thus also yield and quality of fruitand for that reason it is possible to say that iron deficiency is caused mainly by higher levelsof calcium carbonate and the resulting high contents of bicarbonates in soil. These high levelsof bicarbonate ion are typical just for these calcareous soils (Pestana, Faria, De Varennes,2004; Mengel, Breininget, Bübl, 1984). Under such conditions, the occurrence of chlorosissymptoms is quite common and for that reason this type of chlorosis may be defined as a lime-induced iron chlorosis or, abbreviated, lime-induced chlorosis (Pestana et al., 2004).
The identification of chlorosis symptoms in vineyards is very important because it enables toperform protection of plants against mechanisms that induce this plant disease. The identifi‐cation can be performed by means of leaf analysis that enables to estimate contents of indi‐vidual macro and microelements in leaf blades and/or leaf petioles.
The leaf analysis enables to identify all factors that can influence the availability of nutrientsin soil and their uptake by plants; it also can provide information about the nutrient balanceof plants in the moment of sampling (Pestana et al., 2003).
3.2. Causes of the occurrence of lime-induced chlorosis
Although the reasons of the occurrence of this type of chlorosis seem to be relatively definite,the mechanism of its occurrence is still not explicitly defined. It seems that different forms ofiron present in soil and their availability for plants contribute a lot to the occurrence of thistype of chlorosis. Even in very small amounts, iron represents one of those minerals, whichare utilised by plants to assure their sound growth. Iron is used by plants in two forms, viz. asFe2+ and Fe3+.
Iron chlorosis affects susceptible plants growing on calcareous soils. Different kinds ofcarbonates induce different degree of chlorosis. Chlorosis is high for magnesite, hydromag‐nesite and calcite and low for aragonite and nil for dolomite (Fregoni, 1980). There are severaldifferent views concerning iron concentration in soil and its relationship to the occurrence ofchlorosis. In some cases the lime-induced chlorosis can occur under conditions of a low content
Abiotic Stress - Plant Responses and Applications in Agriculture284

of iron in soil and leaves (Bavaresco et al., 1992) while in other its symptoms may be observedat very high levels of iron in leaves (Mengel et al., 1984b). Iron also plays an important role inactivities of the enzymatic system of plants: it actively participates in photosynthetic reduction-oxidation reactions, respiration, biosynthesis of proteins and chlorophyll, biological bindingof atmospheric oxygen, and in reduction of nitrates and nitrites (Tagliavini & Rombola, 2001).
Cultivated plants differ in their susceptibility to Fe deficiency in calcareous soil; some are notmuch affected while others show severe leaf symptoms of chlorosis (Tagliavini & Rombola,2001). In cultivars grown under conditions of a high content of carbonates in soil the contentof chlorophyll can decrease dramatically with the increasing age of plants (Shaaban et al., 2007).
The total content of lime in soil is not very useful for predicting the development of theoccurrence of this type of chlorosis. Active carbonates (active lime) is more reactive and,therefore, able to build and maintain high levels of HCO-
3; for that reason it is a more reliableindicator (Tagliavini & Rombola, 2001). In viticulture, the evaluation of conditions suitable forthe induction of chlorosis the following parameters are usually taken into account: totalcarbonates (%), active lime (%) and CPI (chlorotic power index). Evaluated rootstocks are thenclassified on the base of these analytic parameters. This concept resulted in the so-called“chlorotic power index” (CPI) (Juste & Pouget, 1972. In: Huglin & Schneider, 1998). This meansthat the amount of active lime is related to the amount of Fe extracted by ammonium oxalate.Table 2 shows degrees of chlorosis intensity in relation to different values of CPI (Lupascu etal., 2009).
CPI value Intensity of chlorosis
0 None
≤ 5 Small
6 - 15 Medium
16 -35 High
≥ 36 Very high
Table 2. Degrees of chlorosis intensity in relation to different values of CPI (LUPASCU et al., 2009).
The content of active lime in soil is a parameter, which is frequently used when selectingrootstocks for cultivation of grapevine plants in calcareous soils (Champagnol, 1984).
The susceptibility to chlorosis is the most important selection criterion for rootstocks in manyEuropean wine-growing regions where such a condition is prevalent due to occurrence ofhighly calcareous soils.
Two basic strategies how to classify grapevine plants according to their capability to adaptthemselves to conditions, under which the lime-induced chlorosis can occur (Bavaresco, 1990):
• Strategy I involves four types of response in the roots as follows: a) enhancement of H-ionsrelease, b) formation of rhizodermal or hypodermal transfer cells, c) enhancement of ferric
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
285

iron reduction to ferrous iron, d) enhancement of release of reducing/chelating compoundse.g. phenols.
• Strategy II is characterized by an enhancement of release of non-proteinogenic amino acidsand by a high affinity uptake system.
Bavaresco (1990) formulated the following hypothesis: the response mechanism of tolerantgrapevine rootstocks corresponds probably with Strategy I (Bavaresco et al., 1989) however,the vines are normally grafted and the behaviour of the whole plant towards lime-inducedchlorosis is governed by the following two properties: (i) by the ability of roots to satisfy ironrequirement of leaves; (ii) by the iron requirement of leaves to secure a normal iron nutritionof the plant (Pouget & Ottenwalter, 1973).
The reason that Fe deficiency results in a rapid inhibition of chlorophyll formation is not fullyunderstood, even though this problem has been studied for many years (Bertamini & Nedun‐chezhian, 2005). The reduction of plant biomass of susceptible plants is related to a reducedroot growth due to soil bicarbonate and to a lower photosynthesis rate which also depends bya decrease of leaf chlorophyll, under Fe stress conditions (Bavaresco, Giachino, Pezzutto,2003). According to the growth rate of sink tissues and such organs as the roots, shoot apex,fruits and storage organs can be limited by supply of photosynthates from the source leavesor by a limited capacity of the sink to utilize the photosynthates (Marschner, 1995). In somecases, lime-induced chlorosis is related to a low Fe uptake and its translocation to leaves(Bavaresco et al., 1992), in others to a high content of Fe in leaves, which has to be somehowinactivated (Mengel, Breininget, Bübl, 1984; Bavaresco et al., 1993).
Screening tests of tolerance to chlorosis are performed on plants grown under conditions of ahigh content of bicarbonates in soil. This evaluation can be performed also in vitro on a mediumcontaining a high level of bicarbonates (Bavaresco et al., 1993). The identification of real causesof the occurrence of the lime-induced chlorosis under conditions of a given vineyard is veryimportant for the improvement and/or elimination of these biotic stress situations. Theselection of a suitable rootstock is very important above all in situations when the uptake ofiron is blocked due to a high content of lime in soil and also due to unsuitably chosen rotstocks.
3.3. Tolerance to lime-induced chlorosis in wild species and rootstocks varieties
Rootstocks represent a very important part of the concept how to prevent the occurrence oflime-induced chlorosis in vineyards. A perfect knowledge of soil conditions existing in a givenlocality and also of the resistance of individual rootstocks to lime enables to optimise themanagement of selection of rootstocks on the base of soil conditions.
Use of genotypes tolerant to chlorosis induced by iron blocking is a reliable tool how to solveproblems of chlorosis occurrence (Jimenez et al., 2008).
Chlorosis resistance or susceptibility of grapevine varieties and rootstocks is related not onlyto the root ability to supply adequate iron to the leaves, but also to their iron requirements,which can differ between genotypes. On the basis of this concept, the grapevine varieties wereranked according to their chlorosis resistance or tolerance (Branas, 1974). Breeding also greatly
Abiotic Stress - Plant Responses and Applications in Agriculture286

contributes to the selection of lime-resistant rootstocks. Breeding efforts to get proper geno‐types included successfully crossing between wild grape species, and some chlorosis-resistantrootstocks are now available for the grapevine growers of the many calcareous areas world‐wide (Fregoni, 1980; Pouget, 1980; Bavareso, Fraschini, Perino, 1993). Lime-tolerant grapevinerootstocks have some specific physiological mechanism to overcome chlorosis when grownon calcareous soils, including and improvement of root Fe uptake and reducing capacity(Varanini & Magioni, 1982; Bavaresco, Fregoni, Fraschini, 1991).
Vitis riparia and Vitis rupestris are very important species in the history of the rootstock breedingactivities. These two species are not very tolerant to calcareous soils. Vitis berlandieri isrecognized for adaptation to calcareous soils. Vitis vinifera is species tolerant to calcareous soils(Cousins, 2005). Knowing the characteristics of the important parental species and rootstockvarieties used in rootstock development helps us to understand the viticultural attributes ofindividual rootstocks families.
Data about the tolerance of rootstocks to lime-induced chlorosis, as mentioned by Cousins(2005) and Chauvet & Reynier (1979) are presented in Table 3.
Rootstocks Tolerance to chlorosis Reference
SO 4 MediumCOUSINS (2005)
Börner Low
420 A Good
CHAUVET & REYNIER (1979)
Kober 5BB
SO4Medium
140 Ruggeri Very Good
1103 Paulsen
110 RichterMedium
Fercal Very Good
Table 3. Tolerance of rootstocks to chlorosis (after Cousins, 2005, Chauvet & Reyiner, 1979).
From the viewpoint of the resistance to chlorosis, the rootstocks registered in the State VarietyBook of the Czech Republic can be ranked from the most resistant to the most sensitive asfollows: Craciunel 2 – SO 4 – Kober 125 AA – Kober 5 BB – Teleki 5 C – Amos – LE-K-1. Theseresults are very important from the viewpoint of the use of rootstock varieties for propagationand growing of grapevine in the Czech Republic (Pavloušek, 2008).
In table 4, the classification of rootstock variety, content of active lime and values of CPI aredescribed (Juste & Pouget, 1972 In: Huglin & Schneider (1998).
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
287

Rootstock Content of active lime (%) CPI value
Vialla - 2
Riparia Gloire 6 5
196-17 6 -
101-14 9 10
216-3 9 -
44-53 10 -
3309 11 10
1616 11 -
Rupestris du Lot 14 20
99R,110R,1103P,SO4 17 30
5BB,420A, 34 EM 20 40
161-49 25 50
140 Ru 25 90
41B 40 60
333 EM 40 70
Fercal - 120
Table 4. Classification of rootstocks on the base of the content of active lime and CPI (Juste & Pouget, 1972. In: Huglin& Schneider, 1998).
Recently, the species Vitis cinerea is very often used when selecting new rootstock types. In theCzech Republic, rootstock breeders used the German rootstock Börner and the Czech hybridBruci [(Vitis berlandieri x Vitis rupestris) x Vitis cinerea] as donors of resistance to phyloxera.Hybrids with a very high tolerance to chlorosis originated from parent combinations [Binovax (Binova x Teleki 5C) x Börner] and (Teleki 5C x Börner). These hybrids originated fromcombinations of Vitis berlandieri, Vitis riparia and Vitis cinerea. Hybrid combinations with Vitisrupestris and Vitis amurensis showed mostly only a medium tolerance to chlorosis. A simplehybrid (Binova x Börner) showed also a medium tolerance to lime-induced chlorosis(Pavloušek, 2009).
4. Tolerance of grape rootstocks to drought
Drought stress is one of the most important abiotic stress factors which are generally accom‐panied by heat stress (Zulini et al., 2007).
In recent years, climatic changes can be observed worldwide. Warm years are more frequentand periods of drought are longer. This means that modern viticulture must look for methodshow to react to this increasing frequency of periods of drought.
Abiotic Stress - Plant Responses and Applications in Agriculture288

The grapevine (Vitis vinifera) has different physiological and morphological mechanismsenabling it to maintain growth and production also under conditons of water deficiency(Kondouras et al., 2008).
In Europe, varieties of Vitis vinifera are traditionally cultivated in non-irrigated regions. Yieldof grapes as well as the quality of berries is therefore dependent on the adaptability ofgrapevine plants to drought. A good understanding and control of the water regime of plantsas well as influencing their tolerance to drought stress on the base of application our knowledgeof plant physiology and molecular biology may significantly increase not only productivity ofplants but also quality of environmental conditions.
In grapevine, water supply of plants plays an important role in processes of plant growth andformation of berries. A limited supply of water reduces not only the growth of annual shootsbut also the weight of berries and the final yield of grapes. A marked lack of water may resultin reduced yields and an impaired quality of grapes. This means that in the course of thegrowing season the occurrence of stress induced by water deficit shows a significant effect onphysiological functions of grapevine plants. Although the grapevine (Vitis vinifera) is a speciesshowing a very good tolerance to drought, a severe stress may sometimes markedly influencequalitative properties and parameters of grapes. When using plant material adapted todrought conditions, it is possible to avoid losses caused by a severe water stress (Van Leeuwenet al., 2009).
Selection and breeding of grapevine rootstocks and varieties with a higher water-use efficiencyrepresents a possibility how to adapt viticultural production to current climatic changes(Vandeleur et al., 2009, Flexas et al., 2010).
4.1. Properties influencing the tolerance of grapevine plants to drought
In the course of phylogenesis the grapevine (Vitis vinifera L.) plants have developed variousphysiological and morphological mechanisms, which enable them to maintain their growthand fertility even under conditions of a limited availability of water.
Although grapevine (Vitis vinifera L.) is considered to be a species adapted to droughtstress, the combined effect of high irradiation, high temperatures and low atmosphericwater pressure tension would presumably act as major constraint for the leaf photosyn‐thesis, particularly under conditions of severe soil water deficits usually encountered bythis crop (Flexas et al., 1998).
Physiological responses of plants to water deficit are linked to a condition of recognition ofstress by the root system, turgor changes and water potential and consequently stomatalconductance, internal CO2 concentration and photosynthetic activity decrease. From amolecular perspective, several genes expressed under stress conditions are activated, such asgenes linked to the biosynthesis of abscisic acid and synthesis of specific proteins (Chavaria &Pessoa Dos Santos, 2012). This means that in plants the water stress is manifested by manydifferent mechanisms.
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
289

A limitation of growth of annual shoots and leaves represents one of the first symptoms ofwater deficiency (Stevens et al., 1995). The sensitivity of roots is usually lower than that ofannual shoots (Dry et al., 2000). The growth intensity of annual shoots can be used as one ofvery sensitive indicators of grapevine water status (Patil et al., 1995, Pellegrino et al., 2006,Lebon et al., 2006, Pavloušek, 2011b).
In summer, water available to the plant can often be insufficient because of a lack of precipi‐tation or a low level of its reserves in soil. This can lead to a reduction in the vigour of the plant,its productivity, and quality of the crop. The growth of above-ground parts of grapevine plantsis associated also with the growth of roots and this is directly dependent on the availability ofwater in soil. In periods of drought, roots of some grapevine rootstock varieties can penetratedeep into the soil horizon and thus produce new and new roots.
The growth of roots is also dependent on the relationship, which exists between the rootstockvariety and soil conditions (Morlat & Jacquet, 2003). Rootstock genotype has a major influenceon root density (Southey & Archer, 1988; Williams & Smith, 1991) even though the distributionof grapevine roots is significantly dependent on both edaphic conditions (Smart et al., 2006)and vine (Archer & Strauss, 1985). In extremely drought soils, however, the growth of roots ofsome botanical species may be significantly reduced (Comas et al., 2005).
The tolerance of grapevine to drought is also dependent on the quality of the root system, itsarchitecture, the distribution of individual types of roots within the soil and the density of theroot system in the place of water and nutrients uptake. On the other hand, however, thearchitecture of the root system can be influenced also by spacing of planarity and method ofvineyard tillage.
Roots are usually the first point where the stress is perceived by plants and where they respondto the existing stress conditions. The grapevine tolerance to drought is de facto the capabilityof plants to produce selectively new roots in those places where the groundwater is available.The water stress has a dominant effect on the growth of the grapevine and affects both thegrowth and the development of grapevines.
The growth inhibition of annual shoots decreases transpiration of plants and reduces the totalvolume of conductive tissues (Lovisolo et al., 2010). The transport of water from soil to rootsand (via conductive tissues) also to annual shoots and other above-ground parts of grapevineplants is dependent on activities of aquaporins. Aquaporins are members of the majormembrane intrinsic protein family, where can act as water channels and can regulate cell-to-cell water transport (Maurel et al., 2008). Aquaporins play an important role in the process ofwater absorption. The availability of aquaporins on the surface of roots is changing during theday and depends on the photoperiodicity (Chavaria & Pessoa Dos Santos, 2012).
The physiological mechanisms related to drought tolerance vary from genotype to genotype.It is necessary to screen genotypes for drought tolerance and take into consideration allimportant aspects, e.g. photosynthesis rate, transpiration rate, stomatal conductance andrelative water content occurring at different level of water stress (Satisha et al., 2006). Grapevinevarieties adapt themselves to water deficits by means of various mechanisms, e.g. by changes
Abiotic Stress - Plant Responses and Applications in Agriculture290

in the leaf area (Gómez Del Campo et al., 2003), xylem vessel size, and/or conductivity (Lovisolo& Schubert, 1998).
Stomata enable a control of water regime in plants because they balance and stabilise valuesof water potential existing between their leaves and the atmosphere. Stomatal closure is oneof the first responses to soil drying, and a parallel decline in photosynthesis and stomatalconductance under progressive water stress has already been reported (Medrano et al., 1997).Within the framework of stomatal activities there are relationships among metabolism ofabscisic acid (ABA), hydraulic signals, regulation of activities of aquaporins and electric signalsthat are manifested when measuring the water potential of leaves (Lovisolo et al., 2010). Thismeans that the reaction of stomata is mediated by ABA, which is produced within theframework of a response to the stress induced by drought in roots; this newly synthesised ABAis then transported into other parts of the plant (Loveys et al., 1984).
Plants respond to the lack of water by a quick closing of stomatal opening so that a further lossof water via transpiration is prevented. This mechanism represents a very efficient protectionof plants against drought-induced stress.
A lack of water in soil and a leaf water deficit result also in a gradual reduction of photosyn‐thesis and changes in assimilation of carbon and nitrogen (Chavaria & Pessoa Dos Santos,2012, Zlatev &Cebola Lidon, 2012). Drought-induced decrease in photosynthesis is primarilydue to a stomatal closure, which lowers CO2 availability in the mesophyll, not due to a directeffect on the capacity of the photosynthetic apparatus (Escalona et al., 1999). Osmotic stress isa common feature of many abiotic stress factors, that affect grapevines (Gramer, 2010). Somebiochemical characteristics, e.g. the stability of chlorophyll, can be used for selection ofcultivars resistant to drought conditions (Sinbha & Patil, 1986, Pavloušek, 2011b).
Water-use efficiency (Wue) can be considered for the most important indicator of watermanagement of plants (and also in grapevine). The Wue can be defined as a balance existingbetween the biomass gain (expressed in kilograms of produced biomass or in mols of assimi‐lated CO2) and losses of water (expressed as cubic meters of consumed water or mols oftranspired water). From the agronomic point of view the Wue can be defined as the volumeof yield produced per unit of consumed water (Tomás, et al., 2012). Quality of grapes is verymarkedly dependent on the amount of water consumed by plants and for that reason animprovement in efficiency of water use represents the major requirement concerning cropsustainability and quality of grapes (Medrano et al., 2012). New aspects of Wue and actual dataconcerning this indicator were dealt with and studied in many recent studies (Flexas et al.,2010, Schultz & Stoll, 2010, Lovisolo et al., 2010, Tomás et al., 2012, Medrano et al., 2012). TheWue is a key parameter that enables to evaluate the efficiency of water use within the agrariansector. It is dependent on the total amount of water consumed by plants in the course of thegrowing season. This sum represents the amount of water used by plants plus water lossescaused by transpiration (Flexas et al., 2010).
For that reason it can be expected that there is a relationship between Wue on the one handand genetic foundations (i.e. genomes) of cultivars or rootstocks on the other. Basing on theknowledge of Wue of individual species, rootstocks or cultivars it could be therefore possible
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
291

to recommend them for planting in individual sites/localities with regard to their availabilityof water.
Some studies dealt with the Wue of individual botanic species and rootstocks. A higher Wuevalue was found out in Vitis rupestris while a lower one in Vitis doaniana, Vitis californica andVitis candicans (Padgett-Johnson, et al., 2003). Higher Wue values were also described in Vitisriparia (Flexas et al., 1999) and the rootstock 110 R (Pou et al., 2008).
Soar et al. (2006) reported that rootstock effect on gas exchange of vineyard-grown grapevinesis most likely due to differences in the relative capacity of rootstocks to extract and providescions with water. Rootstocks have been reported to affect the efficiency of water transport tothe shoots via conductivity constrains imposed by the anatomy of xylem vessels (De Herraldeet al., 2006).
Greenspan (2006) differentiates between terms “drought-tolerance” and “drought-avoid‐ance”. Drought-tolerance refers to the ability of the rootstock to support grapevine physio‐logical functions during periods of low soil moisture availability. Rootstocks may exhibitdrought-tolerance through several mechanisms:
Maintaining a low hydraulic resistance to water flow, even under dry conditions.
1. Maintaining photosynthetic activity in leaves, even under low water availability condi‐tions.
2. Preventing the abscission of leaves during periods of low water availability.
Drought-avoidance refers to the ability of the rootstock to prevent low vine water status byone or more of many mechanisms, including:
1. Deep or extensive root exploration to fully exploit soil moisture reserves.
2. Conservation of vine water use by inducing closure of the leaf stomatal pores to limittranspiration.
3. Restricting vine vigour, thereby limiting the amount of transpiring leaf surface area.
The relationship existing between the response of plants and the drought-induced stressinfluences, through physiological reactions of plants, also the development of importantqualitative parameters of grapes (Lovisolo et al., 2010):
1. Effects of plant metabolism, above all photosynthesis and transpiration, on accumulationof sugars and secondary metabolites in berries.
2. Consequences at the berry level of both the chemically-mediated long distance signallingbetween root and shoot (essentially cytokinin and ABA) and the whole-plant hydrauliccontrol via both the xylem and the phloem from root to berry.
3. Adaptation of berry metabolism to a severe osmotic stress existing in berry cells.
ABA, which is present in xylem fluid represents a key signal of root-shoot in plants that arestressed by drought (Schachtmann & Gooder, 2008).
Abiotic Stress - Plant Responses and Applications in Agriculture292

The grapevine belongs to plants, in which a the existence of a very strong relationship betweenABA produced in drought-exposed and stressed roots on the one hand and quality of grapeson the other.
Abscisic acid (ABA) is therefore the most important plant hormone that influences ripen‐ing and quality of grapevine berries. The participation of ABA is high at the beginningof development of berries and decrease till the period of berry softening. Its content in‐creases again during the initial stages of accumulation of sugars and reaches the maxi‐mum approximately 2–3 weeks later (Davies & Böttcher, 2009). The content of ABA inthe skin is higher than in the pulp (Coombe and Hall, 1973). Also grapevine seeds con‐tain more of this acid than pulp (Zhang et al., 2003). ABA participates also in biosynthe‐sis of anthocyanins and, according to Davies & Böttcher (2009) in the accumulation ofsugars in berries. Drought-induced stress supports the formation of ABA and show apositive effect on the formation of secondary metabolites, above all of flavonoids, whichinvolve anthocyanins and tannins. Under conditions of water stress, concentrations of an‐thocyanins and proanthocyanidins in the skin increase independently on the size of ber‐ries; this process is dependent above all on the availability of water (Roby et al., 2004).
In grapevine plants suffering from drought-induced stress, the synthesis of reserve substancesmay be impaired due to an inhibited photosynthesis. This means that the plants are notadequately prepared for overwintering. This stress markedly influences the quality of grapes.Contents of amino acids, organic acids and also sugars are usually reduced. Stressed plantsshow a decreased uptake of minerals from soil and, thus, lower extract in wine. In extremecases it is possible to observe negative effects on smell and taste of wine as well as theoccurrence of the ATA phenomenon (atypical aging – ATA). Due to a high content of poly‐phenols, the taste of stressed wines is bitter, disharmonic and „short“.
From the viewpoint of quality of harvested gapes, a proper evaluation of the drought-inducedstress risk is very important. In white wine varieties, this type of stress may show more negativeeffects than in varieties used for making of red wines because in this case the synthesis of ABAmay show a positive effect on formation of phenolic substances.
4.2. Tolerance to drought in wild species and rootstocks varieties
The use of rootstocks makes it possible to give plants a certain capacity to adapt droughtconditions. A good knowledge of tolerance of rootstocks is important above all with re‐gard to the use of these genetic resources in the process of breeding and selection ofplants tolerant to drought.
The capability of plants to create a root system efficiently penetrating into the soil is animportant factor, which enables them to survive during longer periods of drought andwater-stress.
The assessment and evaluation of tolerance of rootstocks to drought represents an importantcomponent of the process of selection of suitable rootstocks and for further breeding work.The classification of rootstocks into five groups according to their tolerance to drought ispresented in Table 5 (Carbonneau, 1985).
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
293

Degree of resistance Rootstock variety
Highly resistant R 110, R 140, 44-53,
Resistant P 1103, 196-17, P 1447, SO4, R 99, 7383,
Less resistant3309, 7405, 7903, 420 A, Fercal, RSB1, 7921, 5 BB, 161-49, 41 B, Rupestris du
Lot, 101-14
Susceptible Rupestris du Lot, 101-14, EM 333, 7924, Yuga,
Highly susceptible 7542, Vialla
Table 5. Evaluation of drought tolerance of individual rootstock varieties (Carbonneau, 1985).
It is well-known that there are really remarkable differences in tolerance to drought. Somerootstocks (e.g. 101-14 and Schwarzmann) show a low tolerance while in others (e.g. Lider116-60, Ramsey, 1103 Paulsen, 140 Ruggeri, and Kober 5 BB) this property is better (Sommer,2009). Also Cirami et al., (1994) observed a good tolerance to drought in rootstocks Ramsey,1103 Paulsen, and 140 Ruggeri.
Table 6 presents tolerance to drought in some rootstocks varieties after Lavrenčič et al. (2007)and Pouget & Delas (1989).
Rootstock Tolerance to drought Reference
3309 Couderc Low-very sensibleLAVRENČIČ et al. (2007)
1103 Paulsen High
Riparia Gloire
Low
POUGET & DELAS (1989)
101-14
161-49
41 B
3309 Couderc
Moderate
Gravesac
SO 4
420 A
Fercal
110 Richter
High140 Ruggeri
1103 Paulsen
Table 6. Tolerance of rootstocks to drought (Lavrenčič et al., 2007, Pouget & Delas, 1989).
Abiotic Stress - Plant Responses and Applications in Agriculture294

Cregg (2004) stated that to compare the relative tolerance among different genotypes, thevariables to evaluate are as follows: survival potential, growth capacity, and water useefficiency based of morphological and physiological adaptations that might occur in the plant.
The most drought-tolerant grapevine species are V. arizonica, V. californica, V. champinii, V.doaniana, V. gidriana, and V. longii. The lowest tolerance was observed in V. berlandieri, V.cinerea, V. lincecumii, V. riparia, and V. solonis. V. rupestris showed only a moderate toleranceto drought (Padgett-Johnson, et al., 2003).
V. cinerea can assure not only a complete phylloxera resistance; it also shows a positiveinfluence on scion performance especially in shallow, gravely, and consequently dry soils.Phylloxera-resistant V. cinerea hybrids are therefore recommended for vineyards establishedin sites with generally dry conditions. In dry locations V. riparia x V. cinerea hybrids representa valuable expansion of the range of rootstocks currently available in Germany. Particularlyon steep slopes and in seasons with rare rainfall the results obtained with these hybrids weresuperior (Schmidt et al., 2005).
Basing on the evaluation of all traits of tolerance to drought of research in the Czech Republicit is possible to conclude that the highest number of drought-tolerant hybrids originated fromthe crossing of Binova x Börner so that there is a very good chance to use the rootstock Börnerand Vitis cinerea for further breeding and selection of rootstock resistant to drought stress.However, hybrids with Vitis rupestris and Vitis amurensis in their pedigrees show only amedium resistance to drought stress. (Pavloušek, 2011b).
The occurrence of drought is also very closely correlated with the overall soil conditions of thesite. For that reason it is recommended to select individual rootstocks with regard to the typeof soil and also to contents of loamy, clayey and sandy particles within the soil profile.
White (2009) arranged rootstocks with regard to their drought tolerance and pedologicalconditions of the site in the following manner (Table 7):
Soil profile characteristics Vineyard water status Recommended rootstocks
Soil depth < 20 cm: sand, loam or clay
including any root-impeding subsoil
Dry soil 110 Richter, 140 Ruggeri, 1103 Paulsen
Irrigated soil 110 R, 140 Ru, 1103 P, Ramsey
Soil depth 20-75 cm, sands, loams or
clays, with no root-impeding subsoil.
Dry soil 99R, 110R, 140 Ru, 1103P, Ramsey, Kober 5 BB
Irrigated soil
99R, 110R, Ramsey, Kober 5BB, Teleki 5C,
Schwarzmann, SO4, 420A, 101-14 (in loams and
clays).
Soil depth"/> 75 cm, uniform or
gradational profile of sand, loam or clay.
Dry soil 99R, 110R, 1103P, Ramsey (in sand), Kober 5BB.
Irrigated soilSO4, 101-14, Teleki 5C, Schwarzmann, 3306 a 3309
Couderc, 420A.
Table 7. Dependence of tolerance drought and chlorosis of rootstocks on soil conditions (WHITE, 2009).
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
295
A good understanding of physiological mechanism that enable plants to adapt themselves tothe water deficit and to maintain growth also during stress periods could help within theframework of individual breeding programs to screen and select stress-tolerant genotypes(Winter et al., 1988).
5. Conclusion
Regarding climatic changes and a more and more frequent occurrence of periods of droughtwithin the growing season, the problem of lime-induced chlorosis and drought damage ofgrapevine plants becomes to be more and more important.
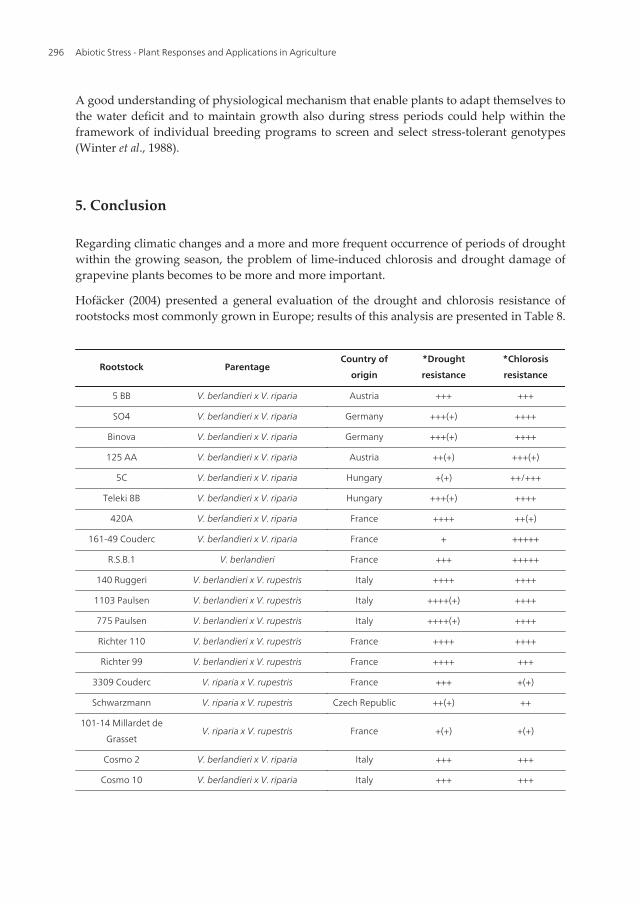
Hofäcker (2004) presented a general evaluation of the drought and chlorosis resistance ofrootstocks most commonly grown in Europe; results of this analysis are presented in Table 8.
Rootstock ParentageCountry of
origin
*Drought
resistance
*Chlorosis
resistance
5 BB V. berlandieri x V. riparia Austria +++ +++
SO4 V. berlandieri x V. riparia Germany +++(+) ++++
Binova V. berlandieri x V. riparia Germany +++(+) ++++
125 AA V. berlandieri x V. riparia Austria ++(+) +++(+)
5C V. berlandieri x V. riparia Hungary +(+) ++/+++
Teleki 8B V. berlandieri x V. riparia Hungary +++(+) ++++
420A V. berlandieri x V. riparia France ++++ ++(+)
161-49 Couderc V. berlandieri x V. riparia France + +++++
R.S.B.1 V. berlandieri France +++ +++++
140 Ruggeri V. berlandieri x V. rupestris Italy ++++ ++++
1103 Paulsen V. berlandieri x V. rupestris Italy ++++(+) ++++
775 Paulsen V. berlandieri x V. rupestris Italy ++++(+) ++++
Richter 110 V. berlandieri x V. rupestris France ++++ ++++
Richter 99 V. berlandieri x V. rupestris France ++++ +++
3309 Couderc V. riparia x V. rupestris France +++ +(+)
Schwarzmann V. riparia x V. rupestris Czech Republic ++(+) ++
101-14 Millardet de
GrassetV. riparia x V. rupestris France +(+) +(+)
Cosmo 2 V. berlandieri x V. riparia Italy +++ +++
Cosmo 10 V. berlandieri x V. riparia Italy +++ +++
Abiotic Stress - Plant Responses and Applications in Agriculture296

Rootstock ParentageCountry of
origin
*Drought
resistance
*Chlorosis
resistance
Riparia Glorie de
MontpellierV. riparia France + +++(+)
Rupestris du Lot V. rupestris France ++ ++
Börner V. riparia x V. cinerea Germany +++(+) ++(+)
Rici V. riparia x V. cinerea Germany +++(+) ++(+)
Cina (V. berlandieri x V. riparia) x V. cinerea Germany +++(+) ++(+)
Sori V. solonis x V. riparia Germany +++ ++
1616 Couderc V. solonis x V. riparia France +++ ++
Gravesac 161-49 C x 3309 C France ++++ +++
Fercal
(V.berlandieri x Colombard) x /
V.berlandieri x (V. riparia x V. rupestris
x V. candicans)/
France ++ +++
Sorisil Sylvaner x 1616 C Germany ++++ +++(+)
26 G Trolinger x V. riparia Germany +++ ++++
41B Millardet de
GrassetChasselas blanc x V. berlandieri France ++++ +++
333 E.M. Cabernet Sauvignon x V. berlandieri France ++++ ++++
Golia Castel 156-12 x V. berlandieri Italy +++(+) +++
Georgikon 28 Kober 5 BB x V. vinifera Hungary ++++ ++++
Note: + = Very low, ++ = Low, +++ = Medium, ++++ = High, +++++ = Very high
Table 8. Drought and chlorosis tolerance of the most common European rootstocks (adapted after *Hofäcker, 2004).
Effects of lime-induced chlorosis and drought of grapevine rootstocks are therefore veryimportant, especially in association with a better understanding of effects of these abioticfactors on grapevine on the one hand and the possibility of the use of such a knowledge whenbreeding and selecting rootstocks on the other.
The aim of this survey of literature is to present a complete overview of rootstock tolerance totwo important abiotic factors – lime and drought.
Acknowledgements
My sincere thanks for support belong to Czech Republic Ministry of Agriculture. Thispublication arose as a part of research project “National programme on plant genetic resourcesconservation and utilization” (206553/2011-Mze-17253).
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
297

Author details
Pavel Pavloušek
Address all correspondence to: [email protected]
Department of Viticulture and Enology, Faculty of Horticulture, Mendel University Brno,Lednice na Moravě, Czech Republic
References
[1] Abadia, J. Leaf responses to Fe-deficiency. Journal of Plant Nutrition 1992; 15 (10)1699-1713.
[2] Alsina M M., Smart D R., De Herralde F., Biel C., Stockert Ch., Negron C., Save R.Seasonal changes of whole root system conductance by a drought-tolerant grape rootsystem. Journal of Experimental Botany 2010; 62 (1) 99-109.
[3] Archer, E, Strauss H.C. The effect of plant density on root distribution of three-year-old grafted 99 Richter grapevines, South African Journal of Enology and Viticulture1985; 6 (2) 25-30.
[4] Arrigo N, Arnold C. Naturalized Vitis Rootstocks in Europe and Consequences toNative Wild Grapevines. PLoS ONE 2007; 6, 1-8.
[5] Bassoi L H., Hopmans J W., De Castro Jorge L A., De Alencar C M., Moura E Silva J A.Grapevine root distribution in drip and microsprinkler irrigation. Scientia Agricola2003; 60 (2) 377-387.
[6] Bauerle T L., Richards J H., Smart D R., Eissenstat D M. Importance of internal hydraulicredistribution for prolonging the lifespan of roots in dry soil. Plant Cell Environment2008a; 31 (2) 177-186.
[7] Bauerle T L., Smart D R., Bauerle W., Stockert Ch., Eissenstat D M. Root foraging inresponse to heterogenous soil moisture in two grapevines that differ in potentialgrowth rate. New Phytologist 2008b; 179 (3) 857-766.
[8] Bavaresco L., Fregoni M., Fraschini P. Investigations on iron uptake and reduction byexcised roots of different grapevine rootstocks and Vitis vinifera cultivars. 5 thInternational Symposium Iron Nutrition and Intercation in Plants, Israel, 11-17 June1989.
[9] Bavaresco L. Investigations on some physiological parameters involved in chlorosisoccurrence in different grapevine rootstocks and Vitis vinifera cultivar. Proceedings ofthe 5 th International Symposium on Grape Breeding, 12-16. September 1989, St.Martin/Pfalz, Vitis Special Issue 1990, 305-317.
Abiotic Stress - Plant Responses and Applications in Agriculture298

[10] Bavaresco L., Fregoni M., Fraschini P. Investigation on iron uptake and reduction byexcised roots of grapevine rootstocks and a V. vinifera cultivar. Plant and Soil 1991; 130(1-2) 109-113.
[11] Bavaresco L. Fregoni M. Fraschini P. Investigations on some physiological parametersinvolved in chlorosis occurrence in grafted grapevine. Journal of Plant Nutrition 1992;15 (10) 1791-1807.
[12] Bavaresco L., Fraschini P., Perino A. Effect of the rootstock on the occurence of lime-induced chlorosis of potted Vitis vinifera L. cv. Pinot blanc. Plant and Soil 1993; 157 (2)305-311.
[13] Bavaresco L., Fregoni M., Perino A. Physiological aspects of lime-induced chlorosis insome Vitis species. I. Pot trial on calcareous soil. Vitis 1994; 33 (2) 123-126.
[14] Bavaresco L., Giachino E., Pezutto S. Grapevine Rootstocks Effects on Lime-InducedChlorosis, Nutrient Uptake, and Source-Sink Relationships. Journal of Plant Nutrition2003; 26 (7) 1451-1465.
[15] Bavaresco L., Poni S. Effect of Calcareous Soil on Photosynthesis Rate, MineralNutrition, and Source-Sink Ration of Table Grape. Journal of Plant Nutrition 2003; 26(10-11) 2123-2135.
[16] Bavaresco L., Presutto P., Civardi S. VRO43-43: A Lime-susceptible rootstock. Ameri‐can Journal of Enology and Viticulture 2005; 56, (2), 192-194.
[17] Bertamini M., Neduchezhian N. Grapevine Growth and Physiological Responses toIron Deficiency. Journal of Plant Nutrition 2005; 28 (5) 737-749.
[18] Branas J. Viticulture. Montpellier : Ecole National Superieure Agronomique;1974.
[19] Buchanan G A., Whiting J R. Phylloxera management: prevention is better than cure.The Australian and New Zealand Wine Industry Journal 6, 223-230.
[20] Carbonneau A. The Early Selection of Grapevine Rootstocks for Resistance to DroughtConditions. American Journal of Enology and Viticulture 1985; 36 (3) 195-198.
[21] Champagnol F. Eléments de physiologie de la vigne et de viticulture générale. Mont‐pellier : Déhand; 1984.
[22] Chauvet M., Reynier A. A. Manuel de Viticulture. Paris: Bailiere; 1979.
[23] Chavaria G., Pessoa dos Santos H. Plant Water Relations : Absorption, Transport andControl Mechanisms. In: Montanaro G., Dichio B. (Eds.): Advances in Selected PlantPhysiology Aspects. InTech;2012, 105-132.
[24] Chaves M.M. Effect of water deficits on carbon assimilation. Journal of ExperimentalBotany 1991; 42 (234) 1-16.
[25] Cirami R M., Furkaliev D G., Radford R. Summer drought and rootstocks. AustralianGrapegrower and Winemaker 1994; 336A 145.
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
299

[26] Comas L H., Anderson L J., Dunst R M., Lakso A N., Eissenstat D M. Canopy andenvironmental control of root dynamics in long-term study of Concord grape. The NewPhytologist 2005; 167 (3) 829-840.
[27] Comas L H., Bauerle T L., Essenstat D M. Biological and environmental factorscontrolling root dynamics and function: effects of root ageing and soil moisture.Australian Journal of Grape and Wine Research 2010; 16: 131-137.
[28] Coombe B G., HalL C R, Hormone content of ripening grape berries and effects ofgrowth substances treatments. Plant Physiology 1973; 51(4) 629-634.
[29] Cousins P. Evolution, Genetics, and Breeding: Viticultural Applications of the Originsof Our Rootstocks. Proceedings of the 2005 Rootstocks Symposium – GrapevineRootstocks: Current Use, Research, and Application 2005; 1-7.
[30] Cramer G R. Abiotic stress and plant responses from the whole vine to the genes.Australian Journal of Grape and Wine Research 2010; 16: 86-93.
[31] Cregg B. Improving drought tolerance of trees. Theorethical and practical considera‐tions. Acta Horticulturae 2004; 630 147-158.
[32] Davies C., Böttcher C. Hormonal control of grape berry ripening. In: Roubelakis-Angelakis K A. (ed.) Grapevine Molecular Physiology and Biotechnology, 2. Ed,Springer Verlag 2009; 229-261.
[33] De Heralde F., Alsina D M., Aranda X., Save R., Biel C. Effects of rootstock and irrigationregime on hydraulic architecture of Vitis vinifera L. cv. Tempranillo, Journal Interna‐tional des Science de la Vigne et du Vin 2006; 40 (4) 133–139.
[34] Dry P R., Loveys B R., Düring H. Partial drying of the rootzone of grape. I. Transcientchanges in shoot growth and gas exchange. Vitis 2000; 39 (1) 3-7.
[35] Escalona j M., Flexas J., Medrano H. Stomatal and non-stomatal limitations of photo‐synthesis under water stress in field-grown grapevines. Australian Journal of PlantPhysiology 1999; 26 (5) 421-433.
[36] Flexas J. Escalona J M., Medrano H. Down-regulation of photosynthesis by droughtunder fields conditions in grapevine leaves. Australian Journal of. Plant Physiology1988; 25 (8) 893-900.
[37] Flexas J., Badger M., Chow W S., Medrano H., Osmond C B. Analysis of relative increasein photosynthetic O2 uptake when photosynthesis in grapevine leaves is inhibitedfollowing low night temperatures and/or water stress. Plant Physiology 1999; 121 (2)675-684.
[38] Flexas J., Galmés J., Gallé A., Gulías J., Pou A., Ribas-Carbo M., Tomás M., Medrano H.Improving water use efficience in grapevines : potential physiological targets forbiotechnological improvement. Australian Journal of Grape and Wine Research 2010;16: 106-121.
Abiotic Stress - Plant Responses and Applications in Agriculture300

[39] Fregoni M. Criterions for the selection of rootstocks in international viticulture.Vignevini 1980; 5, 31-38.
[40] Fregoni M., Bavaresco L. The Italian contribution to grape breeding. Vignevini 1986;13 1-6.
[41] Gómez del Campo M., Ruiz C., Baeza P., Lissarrague J R. Drought adaptation strategiesof four grapevine cultivars (Vitis vinifera L.): modification of the properties of the leafarea. Journal International des. Science de la Vigne et du Vin 2003; 37 (3) 131–143
[42] Greenspan M. Is there such a thing as drought-tolerant rootstocks? Wine BussinesMonthly 2006; 3 pp.
[43] Gruben B., Kosegarten H. Depressed growth of non-chlorotic vine grown in calcareoussoil in an iron deficiency symptom prior leaf chlorosis. Journal of Plant Nutrition andSoil Science. 2002; 165 (2) 111-117.
[44] Hell R., Stephan U W. Iron uptake, trafficking and homeostasis in plants. Planta 2003;216 (4) 541-551.
[45] Hofäcker W. Ergebnisse und Überlegungen zum Einfluss der Unterlage auf Ertrag undQualität der Rebe. Deutsches Weinbau-Jahrbuch 2004. Ulmer Verlag Stuttgart, 175-183.
[46] Huglin P., Schneider C. Biologie et ecologie de la Vigne. Paris : Lavoisier Livre;1998.
[47] Jimenez S., Pinochet J., Abadia A., Moreno A M., Gogorcena Y. Tolerance response toiron chlorosis of Prunus selections as rootstocks. HortScience 2008; 43 (2) 304-309.
[48] Jones H G. How do rootstocks control shoot water relations? New Phytologist 2012;194 (2) 301-303.
[49] Kosegarten H., Wilson G H., Esch A. The effect of nitrate nutrition on iron chlorosisand leaf growth sunflower. European.Journal of Agronomy 1998; 8 (3-4) 283-292.
[50] Koundouras S., Tsialtas I T., Zioziou E., Nikolaou N. Rootstock effect on the adaptivestrategies of grapevine (Vitis vinifera cv. Cabernet Sauvignon) under contrasting waterstatus: Leaf physiological and structural responses. Agriculture, Ecosystem andEnvironment 2008; 128 (1-2) 86-96.
[51] Koyro H-W., Ahmad P., Geissler N. Abiotic Stress Responses in Plants: An Overview.In: Ahmad, P., Prasad, M.N.V. (eds.) Environmental Adaptations and Stress Toleranceof Plants in the Era of Climate Change. 1-28. Springer Science and Business Media; 2012.
[52] Lavrenčič P., Peterlungen P., Sivilotti E. Water Stress and Root Hydraulic Conductivityin Grapevine Grafted on different Rootstocks. Acta Horticulturae 2007; 754 283-288.
[53] Lebon E., Pellegrino A., Louarn G. Lecouer J. Branch development controls leaf areadynamics in grapevine (Vitis vinifera) growing in drying soil. Annals of Botany 2006;98 (1) 175-185.
[54] Levitt J. Responses of plants to environmental stresses. Volume 1 Chilling freezing, andhigh temperature stresses. New York: Academic Press; 1980.
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
301

[55] Loveys B R. Diurnal changes in water relations and abscisic acid in field-growth Vitisvinifera cultivars. III. The influence of xylem-derived abscisic acid on leaf gas exchange.New Phytologist 1984; 98 563-573.
[56] Lovisolo C., Schubert A. Effects of water stress on vessel size and xylem hydraulicconductivity in Vitis vinifera L.. Journal of Experimental Botany 1998; 49 (321) 693–700.
[57] Lovisolo C., Perrone I., Carra A., Ferrandino A., Flexas J., Medrano H., Schubert A.Drought-induced changes in development and function of grapevine (Vitis spp.) organsand in their hydraulic and non-hydraulic interactions at the whole-plant level: aphysiological and molecular update. Functional Plant Biology 2010; 37 (2) 98-116.
[58] Lupascu N., Nicolaescu M., Chirila E. Chlorosis risk evaluation from a new viticulturearea located on a former Army region. In: Simeonov L., Hassanien M.A. (eds) Exposureand Risk Assesment of Chemical Polution-Contemporary Methodology: SpringerMedia; 2009.p. 415-421.
[59] Lynch J. Root architecture and plant productivity. Plant Physiology 1995; 109 (1) 7-13.
[60] Marscher H. Mineral Nutrition of Higher Plants. London: Academic Press; 1995.
[61] Maurel C., Verdoucq L., Luu D T., Santoni V. Plant aquaporins :membrane channelswith multiple integrated functions. Annual Review of Plant Biology 2008; 59 595-624.
[62] Medrano H., Parry M A J., Socias X., Lawlor D W. Long-term water stress inactivatesRubisco in subterranean clover. Annals of Applied. Biology 1997; 13 (3) 491-501.
[63] Medrano H., Pou A., Tomás M., Martorell S., Escalona J M., Gulias J., Flexas J. Improv‐ing water Use Efficiency in Grapevines : Agronomic and Biotechnological Approaches.Acta Horticulturae 2012; 931, 97-107.
[64] Mengel K M., Breininget T., Bübl W. Bicarbonate, the most important factor inducingiron chlorosis in vine grapes on calcareous soil. Plant and Soil 1984a; 81 (3) 333-344.
[65] Mendel K., Bübl W., Scherer H W. Iron distribution in vine leaves with HCO-3 induced
chlorosis. Journal of Plant Nutrition 1984b; 7 (1-5) 715-724.
[66] Morlat R., Jaquet A. The soil effects on the grapevine root system in several vineyardsof the Loire valley (France). Vitis 1993; 32 (1) 35-42.
[67] Nikolic M., Römheld V., Mertkt N. Effect of bicarbonate on uptake and translocationof 59Fe in two grapevine rootstocks differing in their resistance to Fe deficiencychlorosis. Vitis 2000; 39 (4) 145-149.
[68] Padget-Johnson M., Williams L E., Walker M A. Vine Water Relations, Gas Exchange,and Vegetative Growth of seventeen Vitis species grown under irrigated and nonirri‐gated conditions in California. Journal of American Society Horticultural Science 2003;128 (2) 269-276.
[69] Patil S G., Honrao B K., Karkamar S P., Patil V P. Drought tolerance in rootstock ofgrapes. Drakshavritta 1995; 15 (7) 119-122.
Abiotic Stress - Plant Responses and Applications in Agriculture302

[70] Pavloušek P. Preliminary results of tests of grapevine rootstocks resistance to lime-induced chlorosis. Acta Universitatis Agriculturae et Silviculturae MendelianaeBrunensis 2008; LVI 299-302.
[71] Pavloušek P. Evaluation of Lime-induced Chlorosis Tolerance in New RootstockHybrids of Grapevine. European Journal of Horticultural Science 2009; 74 (1) 35-41.
[72] Pavloušek P. Grape Growing - Modern Viticulture (in Czech). Praha: Grada Publishing;2011a.
[73] Pavloušek P. Evaluation of drought tolerance of new grapevine rootstock hybrids.Journal of Environmental Biology 2011b; 32 (5) 543-549.
[74] Pellegrino A., Lebon E., Simmoneau T., Wery J. Towards a simple indiccator of waterstress in grapevine (Vitis vinifera L.) based on the differential sensitivities of vegetativegrowth component. Australian Journal of Grape and Wine Research 2005; 11 306-315.
[75] Perold A I. A Treatise on Viticulture. London: Macmillan, London; 1927.
[76] Pestana M., De Varennes A., Faria E A. Diagnosis and corrction of iron chlorosis in fruittrees: A review. Food, Agriculture and Environment 2003; 1 (1) 46-51.
[77] Pestana M. Faria E A. De Varennes A. Lime-induced iron chlorosis in fruit trees. In:Dris R., Jain S.M. (eds.) Production Practices and Quality Assesment of Food Crops,Plant mineral nutrition and pesticide management, Kluwer Academic Publisher;2004,171-215.
[78] Pire R., Pereira A., Diez J., Fereses E. Drought tolerance assessment of a Venezuelangrape rootstock and possible conditions mechanism. Agrociencia 2007; 41 435-446.
[79] Pou A., Flexas J., Alsina M M., Bota J., Carambula C., De Herralde F., Galmés, J.,Lovisolo C., Jimenéz M., Ribas-Carbo M., Rusjan D., Secchi F., Tomás M., Zsófi M.,Medrano H. Adjustment of water-use efficiency by stomatal regulation during droughtand recovery in drought-adapted Vitis hybrid Richter-110 (V.berlandieri x V.rupestris).Physiologia Plantarum 2008; 134 (2) 313-323.
[80] Pouget R. Breeding Grapevine Rootstocks for Resistance to Iron Chlorosis. In. Pro‐ceedings 3 rd Int. Symp. Grape Breeding 1980; Davis, USA, 191-197.
[81] Pouget R., Delas J. Le choix des porte-greffes de la vigne pour une production de qualite.Connais. Vigne Vin.1989; 27-31.
[82] Pouget R., Ottenwalter M. Etude methodologique de la resistance a la chlorose calcairechez la vigne:principe de la methode des greffages reciproques et application a larecherche de porte-greffes resistants. Annals.Amelioration.Plantes 1973; 24 347-356.
[83] Pouget R., Ottenwaelter M. Investigations on the adaptation of new rootstock vines insoils having a very marked chlorosis action. Connaissance Vigne at Vin 1978; 12167-175.
[84] Richards D. The grape root system. Horticultural Reviews 1983; 5 127-168.
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
303

[85] Rizk-Alla M S., Sabry G H., El-Wahab M A. Influence of Some Rootstocks on thePerformance of Red Globe Grape Cultivar. Journal of American Science 2011; 7 (4)71-81.
[86] Roby G., Harbertson J F., Adams D A., Matthews M A. Berry size an vine water deficitsas factors in winegrape composition: anthocyanins an tannins. Australian Journal ofGrape and Wine Research 2004; 10 100-107.
[87] Saayman D. Soil preparation studies: II. The effect of depth and method of soilpreparation and of organic material on the performance of Vitis vinifera (var. Cheninblanc) on a Hutton/Sterkspruit soil. South African Journal of Enology and Viticulture1982; 3 (1) 61-74.
[88] Satisha J., Prakash G S., Venugopalan R. Statistical Modeling of the effect of physio-biochemical Parameters on Water use efficiency of Grape Varieties, rootstocks and theirscionic combinations under moisture stress conditions. Turkish Journal of Agricultureand Forestry 2006; 30 261-271.
[89] Schachtman D P., Gooder J Q. Chemical root to shoot signaling under drought. Trendsin Plant Science 2008; 13 (6) 281-287.
[90] Schmidt J., Manty F. Huber L. Porten M., Rühl E H. Experience with RootstocksVarieties in Germany. Proceedings of the 2005 Rootstocks Symposium – GrapevineRootstocks: Current Use, Research, and Application 2005; 14-24.
[91] Schultz H R., Stoll M. Some critical issues in environmental physiology of grapevines:Future challenges and current limitations. Australian Journal of Grape and WineResearch 2010; 16: 4-24.
[92] Shaaban M M., Loehnertz O., El-Fouly M M. Grapevine Genotypic Tolerance to Limeand Possibility of Chlorosis Recovery throught Micronutrients Foliar Application.International Journal of Botany 2007; 3 (2) 179-187.
[93] Sinbha N C., Patil B D. Screening of barley varieties for drought resistance. PlantBreeding 1986; 97 (1) 13-19.
[94] Smart D R., Schwass E., Lakso A., Morano L. Grapevine rooting patterns: a compre‐hensive analysis and review, American Journal of Enology and Viticulture 2006; 57 (1)89–104.
[95] Soar C J., Dry P R., Loveys B R. Scion photosynthesis and leaf gas exchange in Vitisvinifera L. cv. Shiraz: mediation of rootstock effects via xylem sap ABA. AustralianJournal of Grape and Wine Research 2006; 12 (2) 82–96.
[96] Somkuwar R G., Taware P B., Bondage D D., Navale S. Root length, root mass, anddistribution of dry matter in different parts of Thompson Seedless grapevine graftedon different rootstocks in heavy soil of Maharashtra. Turkey Journal of Agriculturaland Forestry 2012. Doi:10.3906/tar-0911-377.
[97] Sommer K. Spring recovery-resilience of Sultana to drought. The Vine 2009; 5 (3) 32-34.
Abiotic Stress - Plant Responses and Applications in Agriculture304

[98] Southey J M., Archer E. The effect of rootstock cultivar on grapevine root distributionand density. In: VAN ZYL, J.L. The grapevine root and its environment 1988. Dep.Ag‐ric. Water Supply, Pretoria, South Africa, 57-73.
[99] Stevens R M., Harvey G. Aspanall D. Grapevine growth of sholte and fruit linearlycorrelate with water stress indices based on root-weighted soil matric potential.Australian Journal of Grape and Wine Research 1995; 1(1) 58-66.
[100] Swanepoel J J., Southey J M. The Influence of Rootstock on the Rooting Pattern of theGrapevine. South African Journal of Enology and Viticulture 1989; 10 (1) 23-28.
[101] Tagliavini M., Rombola A D. Iron deficiency and chlorosis in orchard and vineyardecosystems. European Journal of Agronomy 2001; 15 (2) 71-92.
[102] Tomás M., Medrano H., Pou A., Escalona J M., Martorell S., Ribsa-Carbo M., Flexas J.Water-use efficiency in grapevine cultivar grown under controlled comditions: effectsof water stress at the leaf and whole plant level. Australian Journal of Grape and WineResearch 2012; 18 (2) 164-172.
[103] Vandeleur R K., Mayo G., Shelden M C., Ciliham M., Kaiser B N., Tyerman S D. Therole of PIP aquaporins in water transport through roots: diurnal and drought stressresponses reveal different strategies between isohydric and anisohydric cultivars ofgrapevine. Plant Physiology 2009; 149 445-460.
[104] Van Leeuwen C., Tregoat O., Choné X., Bois B., Pernet D., Gaudillére J-P. Vine waterstatus is a key factor in grape ripening and vintage quality for red Bordeaux wine. Howcan it be assessed for vineyard management purpose? Journal International Science dela Vigne et du Vin 2009; 43 (3) 121-134.
[105] Varanini Z., Maggioni A. Iron reduction and uptake by grapevine roots. Journal of PlantNutrition 1982; 5 (4-7) 521-529.
[106] White R E. Understanding vineyards soils. New York: Oxford University Press; 2009pp. 230.
[107] Williams L E., Smith R.J. The effect of rootstock on the partitioning of dry weight,nitrogen and potassium, and root distribution of Cabernet Sauvignon grapevines.American Journal of Enology and Viticulture 1991; 42 (2) 118–121.
[108] Winter S R., Musick J T., Porter K B. Evaluation of screeing techniques for breedingdrought resistant winter wheat. Crop Science 1988; 28 (3) 512-516.
[109] Zhang G M., Luo G G., Wang R H., Wang J., Himerlick D G. Growth and developmentalresponses of seeded and seedless grape berries to shoot girdling. Journal of AmericanSociety of Horticultural Science 2003; 128 316-323.
[110] Zlatev Z., Lidon F C. An overview on drought induced changes in plant growth, waterrelations and photosynthesis. Emirates Journal of Food and Agriculture 2012; 24 (1)57-72.
Tolerance to Lime - Induced Chlorosis and Drought in Grapevine Rootstockshttp://dx.doi.org/10.5772/54793
305

[111] Zulini L., Rubinigg M., Zorer R., Bertamini M. Effects of drought stress on chlorophyll
fluorescence and photosynthesis pigments in grapevine leaves (Vitis vinifera cv. White
Riesling). Acta Horticulturae 2007; 754 289-294.
Abiotic Stress - Plant Responses and Applications in Agriculture306








![Fertilizer for the Treatment of Iron Chlorosis EMILIAN 1 17.pdf · Fertilizer for the Treatment of Iron Chlorosis ... immobilization of the iron [12-21]. Iron plays a complex ...](https://static.fdocuments.in/doc/165x107/5b147f327f8b9a207c8d6d6b/fertilizer-for-the-treatment-of-iron-emilian-1-17pdf-fertilizer-for-the-treatment.jpg)









