
This dissertation hasbeen 66-13,704 HARAMOTO Frank Hiroshi
113
This dissertation has been microfilmed exactly as received 66-13,704 HARAMOTO" Frank Hiroshi, 1924- BIOLOGY AND CONTROL OF BREVIPALPUS 'PHGENICrs (GEIJSKES) (ACARINA: TENUIPALPIDAE). University of Hawaii, Ph.D., 1966 Zoology University Microfilms, Inc., Ann Arbor, Michigan
Transcript of This dissertation hasbeen 66-13,704 HARAMOTO Frank Hiroshi

This dissertation has beenmicrofilmed exactly as received 66-13,704
HARAMOTO" Frank Hiroshi, 1924-BIOLOGY AND CONTROL OF BREVIPALPUS'PHGENICrs (GEIJSKES) (ACARINA:TENUIPALPIDAE).
University of Hawaii, Ph.D., 1966Zoology
University Microfilms, Inc., Ann Arbor, Michigan

.BIOLOGY AND CONTROL
OF
BREVIPALPUS PHOENICIS (GEIJSKES)
(ACARINA: TENUIPALPIDAE)
A THESIS SUBMITTED TO THE GRADUATE SCHOOL OF THE
UNIVERSITl OF HAWAII IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF
DOC TOR OF PHILOSOPHY
IN ENTOMOLOGY
JANUARY 1966
By
Frank Hiroshi Hararnoto
Thesis Comrn.ittee:
D. Elmo Hardy, Chairm.anHenry A. BessMarnoru IshiiWallace C. MitchellToshiyuki Nishida

iii
TABLE OF CONTENTS
· . . . . . . . . . .· . . . . . . . . . . . .
· . . . . . . . . . . . .
· . . . . . . . . . . . .
v
iv
1
3
5
10
14
24
42
57
67
7I
78
91
97
101
. . . . . . .
. .. . . . . . . .
. .
. . . . . .
• • 0 • • • • • • • • •
• • • • • • • • • • • •
• • • • • • • • • • • • •
• • • a • • • • • • • • •
· . . . . . . . . . . . .
· . . . . . . . . . .· . . .
. . .CHEMICAL CONTROL.
DISCUSSION •••
SUMMARY • • 0 • • ••
REFERENCES CITED • • ••
POPULATION STUDIE;
NA TURAL CONTROL FACTORS
LIFE HISTOR Y • • • • • • • • •
BIOLOGICAL OBSERVATIONS
METHODS AND MATERIAJ..S •
DE;CRIPTION OF liFE STAGE;
LIST OF TABLE; ••• 0 •
liST OF ILLUSTRATIONS
INTRODUCTION ••••••••
TAXONOMIC STATUS • • • • •
DISTRIBUTION • • • • • • • •
HOST PLANTS ••••••

Table
I.
il.
ID.
IV.
V.
VI.
Vil.
VID.
List of Tables
Geographical distribution of BtevipalpusphoPcnicis (Geij skes) •• • • • • • • • • • • • • •
Monthly maxiInum-minimum temperature s forHonolulu and Kaneohe, Oahu, and for Kahului,Maui, during 1962 and 1963 ••••••••••••••
List of host plants of Brevipalpus phoenicis(Geij skes) in Hawaii ••••••••••••••• ••
Average measurements of ~he life stage s ofBrevipalpus phoenicis (Geijskes). • I> •••••••
Effect of temperature and humidity on the lifestages of Brevipalpus phoenicis (Geijskes) •• 0 ••
Performance of different pesticides against theadults of Brevipalpus phoenicis (Geij skes) •••••
Number of eggs of Br,evipalI?us phoenicis(Geij skes) counted on treated surface one weekafter application of the different pesticides • • • • •
Ovicidal effect of different pesticides on theeggs of Brevipalpus phoenicis (Geijskes) ••••••
iv
6
9
11
27
44
86
88
90

Figure
1.
List of Illustrations
Distribution map of Brevipalpus phoenicis • • • • •
v
7
2. Rearing unit used in the constant temperatureand humidity experiments • • • • .. • • • • •
Rearing unit used in the chemical control studiesof BrevipalP\;1s phoenicis ••••••••••
• •
• •
18
21
5.
6.
7.
8.
9.
10.
Outline drawing of Brevipalpus phoenicis femaleehowing location and nomenclature of thesetae and plate s ••••••••••.••••• 0
Egg of Brevipalpus phoenicis. • • • • • • • • • •
Larva of Brevipalpus phoenicis. • • • • • • • • •
Protonymph of Brevipalpus phoenicis • • • • • • •
Deutonymph of Brevipalpus phoenicis • • • • • • •
Female of Brevipalpus phoenicis • • 0 • • • • • •
Variation in color patterns of females ofBrevipalpus phoenicis • • • • • • oi • • • 0 • • •
25
29
31
33
35
37
38
11. Male of Brevipalpus phoenicis . . . .0. . . . . 41
12. Mean number of eggs laid per week by 25Brevipalpus phElenicis female s • • • • • 0 0 0 • • 53
13.
14.
Longevity of the adults of Brevipalpus phoenicisat the different temperatures in combinationwith 65 to 70 per cent relative humidity • • • •
Feeding injury caused by Brevipalpus rhoenicison papaya fruit • • • • • 0 0 • • • • • • • •
• •
• •
56
59

IS.
16.
17.
Feeding injury caused by Brevipalpus phoenicison papaya fruit • • • • • • • • • • • • • • •
Feeding damage caused by Brevipalpus phoenicison passion fruit. • • • • • 0 • • • • • • • •
Egg and adult densities and fluctuations ofBrevipalpus phoenicis populations. • 0 0 0
o 0
vi
61
62
68
18.
19.
Injury caused by Chlorobenzilate •
Injury caused by Pentac • 0 •
o 0 0 . . . . .• •
82
83
20. Injury caused by ovex .0 . . . . • • • I! 84

rnTRODUC TION
The red and black flat mite, Brevipalpus phoenicis (Geijskes),
has been reported from many countries since its discovery in Holland
in 1939 (Geij skes, 1939). This mite is not endemic to the type locality,
but is believed to have a tropical origin. However, because of its
extensive geographical distribution and host range, the native home of
this mite cannot be established.
Baker in 1949 reported that several species of Brevipalpus have
been found to be pests of cultivated plants, and although not as impor
tant as the spider mites they are serious enough at times to warrant
investigations of their biology and control. Since then, Manglitz and
Cory (l953) and Morishita (1954) have studied in detail the biology and
control of B. californicus (Banks) and of Jh obovatus Donnadieu,
respectively. These two species along with!h phoenicis and!h lilium.
Baker make up the known fauna of false spider mites of the genus
Brevipalpus in Hawaii. Of these, !h phoenicis is of most concern here
in Hawaii for besides damaging many ornamental plants, it attacks
two crops of significant economic importance: papaya and passion fruit.
~ phoenicis may have been in Hawaii previous to its description
date for in 1936 Marlowe (1937) reported a mite injury on papaya fruits
with symptoms similar to those now known to be caused by this species.
Despite its early presence in Hawaii, very little work has been done in
the past on this important mite by other worker So Therefore, a detailed

biological study ofl!: phoenicis has been made and the findings are
presented in this thesis so that they may serve as a basis for further
studies of this as well as other species of phytophagous mites.
2.

3.
TAXONOMIC STATUS
B. phoenicisbelongs to the order Prostigmata, sl!perfa:mily
Tetranychoidea, and fa:mily Tenuipalpidae. Geij skes in his descrip
tion of phoenicis in 1939 placed it in the genus Tenuipalpus. At that
time, Brevipalpus was considered as a synonym of Tenuipalpus; how
ever, Baker in 1945 established the validity of the for:mer and
reinstated it as a generic name.
Unaware of the extent of possible variations in certain charac
ters of B. phoenicis, Baker P949) na:med~ yothersi, ~ mcbridei,
and B. papayensis as separate entities based on the differences noted
in the size of the dorso-lateral setae of the nymphs of yothersi and
:mcbridei, and on the presence of a prominent edentation on the second
palpal segm.ent, as well as on the less distinctive areolate pattern on
the dorsum of papayensis adults. In later studies, these characters
were found to be :merely intraspecific variations, and thus, the three
names were declared as synonyms of B. phoenicis (Pritchard and
Baker, 1951).
In Hawaii, prior to 1951, B. phoenicis was referred to as~ bio
culatus McGregor (Holdaway, 1941), and as B. papayensis (Baker,
1949). The former is a misidentification and a synonym of B. obovatus,
a species very similar in gross morphology to B. phoenici~, and the
latter is a synonym. of B. phoenicis. The na:me, B. papayensis, was
proposed in 1949 by Baker for the :mites on papaya collected in 1941 by

4.
w. C. Look from Kailua, Oahu, Hawaii.
At present, there is no appt"oved common name for B.
phoenicis. The common name, the red and black flat mite, was first
used by Muma (1961) and is adopted to referring to~ phoenicis in
- this paper.

DISTRIBUTION
!h E!!.oenicis is widely distributed in both continental and insular
areas, primarily throughout the tropics of the world. Of the 28
distributional records, 18 have been reported from somewhere within
the boundarie s of the Tropic of Cancer to the north and the Tropic of
Capricorn to the s·outh (Fig. 1 and Table I). The northernmost area
from where B~ phoenicia has been reported is Holland (Geijskes, 1939)
and the southernmost is Argentina (Baker, 1949). The few reports of
recoveries of this mite from other than the tropics probably represent
temporary establishments as a result of dispersal from the generally
favorable range into pockets of favorable environment. The outbreaks
of ~ phoenicis reported in glasshouses in Europe are exam.ples of such
dispersal and fortuitous establisbm.ents. Since its initial description in
Holland in 1939, B. phoenicis was not observed again in Europe until
1951 when it was found infesting Phoenix canariensis Hort. in glass
houses in Vienna, Austria (Dosse, 1957). The only areas outside the
tropics where B. phoenicis is firmly established are in Florida (Muma,
1958) and in the Mediterranean region (Baker, 1949; Attiah, 1956; and
Di Martino~ 1960) where the climate is mild and similar to that of the
tropics.
In Hawaii, B. phoenicis has been recovered from the islands of
Maui, Hawaii, Oahu, and Kauai. It is generally well distributed along
the coastal plains and foothills up to about 1, 000 feet in elevation.

6.
TABLE I. GEOGRAPHICAL DISTRIBUTION OF BREVIPALPUSPHOENICIS (GEIJSKES)
Continent
Africa
Asia
Europe
North America
Area *Belgian Congo (1)Egypt (Z)Kenya (3)Mauritius (4)Tanganyika (5 )
Aden Protectorate (6)Australia (7)
Ceylon (8)Formosa (9)India (10)Malaya (11)Philippines (IZ)
Austria (13)Holland (14)Sicily (15)Spain (16)
California (17)Cuba (18)Florida (19)Mexico (ZO)Puerto Rico (Z1)Washington, D. C. (ZZ)
Reference
Baker and Pritchard, 1960Attiah, 1956Pritchard and Baker, 1958Mou1ia, 1958Pritchard and Baker, 1958
Knorr, et al., 1961Commonwealth Institute of
Entomologyp 1959Baptist and Ranaweere, 1955Pritchard and Baker, 1958Pritchard and Baker, 1958Pritchard and Baker, 195ZRimando, 196z
Dosse, 1957Geijskes, 1939Di Martino, 1960Baker, 1949
Pritchard and Baker, 195ZBaker, 1949Baker, 1949DeLeon, 1961Cromroy, 1958Baker, 1949
Pacific Islands Hawaii (Z3)
South America Argentina (Z4)Brazil (Z5)Paraguay (Z6)Trinidad (Z7)Venezuela (Z8)
Baker, 1949
Baker, 1949Rossetti, et al., 1959Nickel, 1958Pritchard and Baker, 195ZKnorr, ~t al., 1960
* Numbers after localities correspond to those on map in Figure 1.

---~--------
Figure 1. Distribution map of Brevipalpus Eoenicis (Geijskes)-J.

8.
Also, papaya and passion fruit, two favorable hosts of ~ phoenicis,
are well adapted and grown extensively in many of these areas.. In
1964 there were approxim.ately 1,350 acres in papaya and 250 acres
in passion fruit in Hawaii. The climatic conditions in the areas where
these crops are grown are warm, hUID.id, and conducive for year round
reproduction of this false spider mite. The monthly maxim.UID. and
minim.UID. temperatures for the period of this study, January, 1962 to
December, 1963, for Kaneohe, Oahu, where papaya is very often
heavily infested by B. :phoenicis, for Kahului, Maui, where serious
damage by this mite to pas sion fruit has occurred, and for Honolulu,
Oahu, where this mite is very common on many ornamental plants,
are presented in Table II. The temperature in these areas seldom
approaches the maxima and mini.m.a shown in Table II but usually
fluctuates within a few degrees from the mean. In one such area, Wai
manalo, Oahu, where ~ phoenicis outbreak was first seen on papaya,
the mean temperature, the daily temperature range, and the annual
temperature range have been recorded as 23.9°e, 7. 2oe, and 4.4oe,
respectively (Ripperton and Hosaka, 1942). The hUID.idity condition
in these areas is relatively high and seldom drops below 50 per cent.
B. phoenicis has not been recovered from areas above 2, 500 feet
in elevation despite careful search on many kinds of plants. Tempera
ture is more variable in the upper areas than below (Ripperton and
Hosaka, 1942)0 In the upper areas, B. obovatus and B. californicus
were frequently encountered.
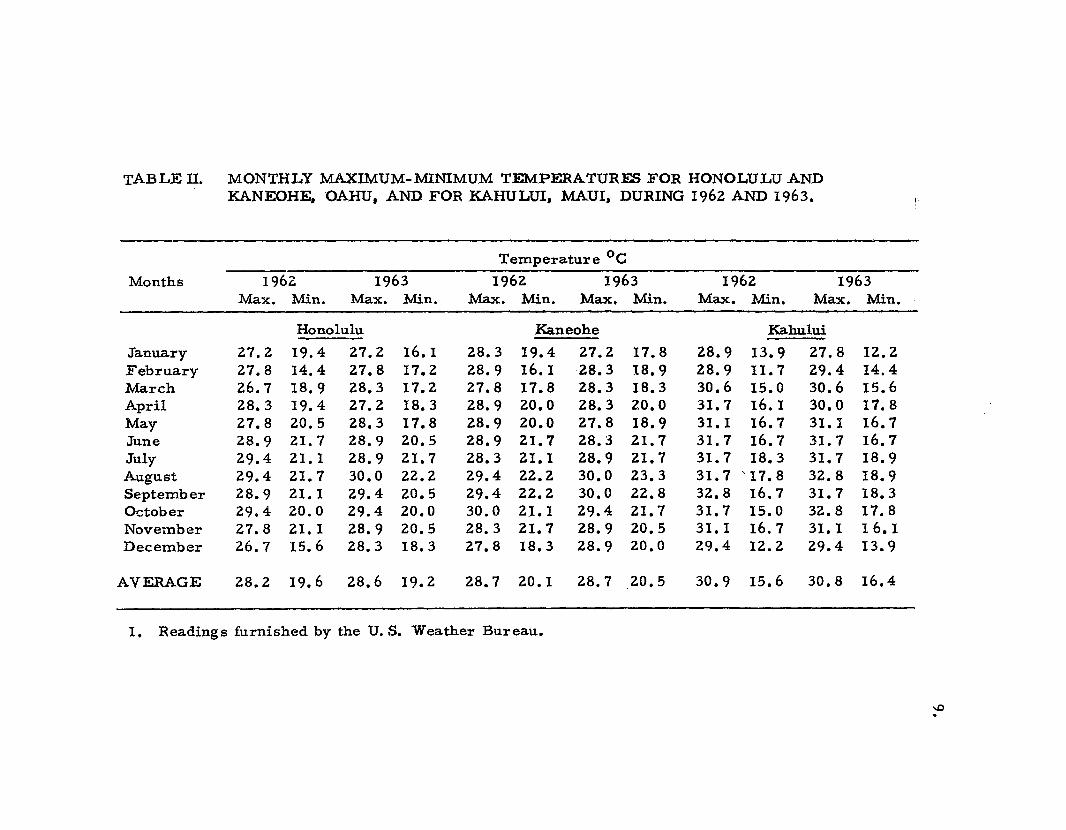
TABLE II. MONTHLY MAXIMUM-1vfiNIMUM TEMPERATURES FOR HONOLULU ANDKANEOHE, OAHU, AND FOR KAHULUI, MAUl, DURING 1962 AND 1963.
Temperature °cMonths 1962 1963 1962 1963 1962 1963
Max. Min. Max. Min. Max. Min. Max. Min. Max. Min. Max. Min.
Honolulu Kaneohe Kahului
January 27.2 19.4 27.2 16.1 28.3 19.4 27.2 17.8 28.9 13.9 27.8 12.2February 27.8 14.4 27.8 17.2 28.9 16.1 28.3 18.9 28.9 II. 7 29.4 14.4March 26.7 18.9 28.3 17.2 27.8 17.8 28.3 18.3 30.6 15.0 30.6 15.6April 28.3 19.4 27.2 18.3 28.9 20.0 28.3 20.0 31. 7 16.1 30.0 17.8May 27.8 20.5 28.3 17.8 28.9 20.0 27.8 18.9 31. I 16.7 31. 1 16.7June 28.9 21. 7 28.9 20.5 28.9 21. 7 28.3 21. 7 31. 7 16.7 31. 7 16.7July 29.4 21. 1 28.9 21.7 28.3 21.1 28.9 21. 7 31. 7 18.3 31. 7 18.9August 29.4 21. 7 30.0 22.2 29.4 22.2 30.0 23.3 31.7 '17.8 32.8 18.9September 28.9 21. 1 29.4 20.5 29.4 22.2 30.0 22.8 32.8 16.7 31. 7 18.3October 29.4 20.0 29.4 20.0 30.0 2.1.1 29.4 21.7 31.7 15.0 32.8 17.8November 27.8 21.1 28.9 20.5 28.3 21.7 28.9 20.5 31.1 16.7 31.1 16. IDecember 26.7 15.6 28.3 18.3 27.8 18.3 28.9 20.0 29.4 12.2 29.4 13.9
AVERAGE 28.2 19.6 28.6 19.2 28.7 20.1 28.7 20.5 30.9 15.6 30.8 16.4
I. Readings furnished by the U. S. Weather Bureau.
i
-.D.

10.
HOST PLANTS
Phoenix sp., a greenhouse palm, was reported as the first host
plant of.!h phoenicis in 1939. Since then, many different kinds of
plants have been reported by several authors as infested by this
species of mite in different parts of the world (Cromroy, 1958;
Pritchard and Baker, 1958; Baker and Pritchard, 1960; DeLeon, 1961;
and Rimando, 1962). To date about 80 genera of plants have been
listed as hosts of~ phoenicis.
Previously, only seven species of plants have been recorded as
hosts of B. phoenicis in Hawaii (Pritchard and Baker, 1952; and
Chilson, 1959), but some 39 species of plants belonging to 27 families
were found infested during this present study. Papaya (Carica papaya
L.), passion fruit (Passiflora edulis f. £lavicarpa Degener), hemi
graphis (Hemigraphis colorata (Bl.) Hallif.), :anthurium. (Anthurium.
andraeanum. Lind.), and leInon (Citrus liJ:nonia Osbeck), are excep
tionally favorable hosts of~ phoenicis. Only those plants which
contained all stages of the Inite in sufficient num.ber s were included in
the host list shown in Table ill. Although the list is by no means
cOlnplete, it does show that B. phoenicis is polyphagous. Many Inore
plants, belonging to the families listed in Table ill, are likely hosts
of this Inite; however, it must be cautioned here that two other species
of false spider Inites. B. obovatus and B. californicus, which look
very much like ~ :e!:.0enicis coexist on m.any plants. There also

TABLE m. LIST OF HOST PLANTS OF BREVIPALPUSPHOENICIS (GEIJSKES) IN HAWAII
Family Species
Acanthaceae Hemigraphis coloIata (Bl. )Ballif.
Aizoaceae Sesuvium }.i.,jrtulacastrum L.
Anacardiaceae Mangifera indica L.
Apocynaceae Allamanda cathartica L.
Araceae Anthurium andraeanumlind.
Araliaceae Nothopanax guilfoylei(Gogn. and March.) Merr.
Bignoniaeceae Bignonia venusta Ker.
Cannaceae Canna indica L.
Garicaceae Carica papaya L.
Compositae Eupatorium glandulosumHBK.
Bidens pilosa L.
Convolvulaceae Ipomoea batatas (L.) Poir.Ipomoea pes-caprae (L. )
Sweet
Cucurbitaceae Momordica balsamina L.
Euphorbiaceae Acalypha wilkesianaMuell. - Arg.
Liliaceae Cordyline terminalis (L.)Kunth
Malpighiaceae Malpighia glabra L.
Common Name
hemigraphis
carpet-weed
mango
allamanda
anthurium
panax
orange trumpet vine
canna
papaya
Maui pamakani
Spanish needle
sweet potatobeach morning-glory
balsam apple
copper leaf
ti
acerola

TABLE III. Continued
Fanlily Species
Ma.lvaceae Hibiscus rosa-sinensis (L.)Thespesia populnea (L.)
Sol. .
Meliaceae SWietenia mahagoni (L.)Jacq.
MiInosaceae Prosopis pallida (Willd.)HBK
Myrtaceae P sidium guajava L.
Orchidaceae Dendrobium.~Arundina. bambusuoHa
Lindl.
Palm<:+e Rhapis excelsa (Thunb.)Henry
Passifloraceae Passiflora foetida L.Passiflora edulis f.
flavicarpa Degener
Rubiaceae Coffea arabica L.
Rutaceae Citrus :paradisi Mad.Citrus sinensis (L.) OsbeckCitrus limonia OsbeckMurraya exotica L.
Sapindaceae Litchi chinensis Sonne
Solanaceae Solanuxn sodoroeum. L.
Verbenaceae Clerodendrum fragrans(Vent.) R, Br. var.pleniflorum Schau.
Lantana. ca.mara L"Lantana. montevidensis
(Spreng.) Briq.
12.
Common Name
hibiscusmi1p._
mahogany
kiawe
guava
dendrobiumbamboo orchid
bamboo palm
running popyellow passion fruit
coffee
grapefruitsweet orangelemonmock orange
litchi
apple of sodorn
glorybower
lantanatrailing lantana

13.
remains the possibility of recording plants which are merely
accidental hosts because of the general prevalence of!!.. phoenicis
in some areas.
Although ]h ;phoenicis and !!.. obovatus are polyphagous and
infest m.any kinds of plants in common, there are some plants which
support only one of them. The garden violet, Yl2.!! odorata L., was
found by Morishita (1954) to be a practical host for laboratory rearing
of B. obovatus; however, in this study, it was found unsuitable for
rearing .!!a. phoenicis. Likewise, 14 oboyatus was not able to re!1"0
duce on papaya which proved to be an excellent rearing medium for
~ phoenicis. These two plants, the garden violet and papaya, may
be used in the separation of mixed populations into pure cultures of
these two species of mites which coexist on many kinds of plants and
are extremely difficult to distinguish unless mounted and examined at
magnifications of 900X or higher.

14.
METHODS AND MATERIALS
Stock cultures of B. phoenicis were started from field collected
specimens found infesting papaya in Manoa Valley, Oahu, Hawaii, on
April 25, 1962. The mites were mass reared on potted papaya plants,
three to five months old, which were kept outdoors but sheltered
from wind and rain. As needed, the mite s we re transferred from
these source plants to rearing media in order to study the various
aspects of the biology and control of this species of mite.
Since papaya of the Solo variety is of great economic itnport
ance and a favorable host plant of B. phoenicis, it was used to study
the life history of this species in the laboratory. Green papaya fruits
or sections thereof were used as rearing media for they remained in
fairly good condition long enough to allow the completion of a life cycle
and were easier to manipulate under laboratory conditions than other
types of media. Excised leaves and stems of most plants used as
rearing media required frequent changing due to rapid desiccation.
This was especially true when mites were reared on them at tempera
tures above 2SoC and at relative humidities below 60 per cent. The
necessity of frequent changes of the medium created a risk of injur
ing or 10 sing the mite s while making the transfer s to fre sh medium.
When using papaya fruits under these adverse conditions, the entire
surface, except for the areas where the mites were allowed to feed,
was coated with paraffin. The paraffin film helped to prevent the rapid

15.
desiccation of the fruits by reducing the surface area of transpiration.
For transferring mite s from stock culture s to rearing m.edia,
a human hair mounted on a wooden handle of a dissecting needle
proved most satisfactory. Although the mites wer'e accidentally
jabbed in the process of picking them up for transference, the hair
was flexible enough so that no bodily harm was inflicted. The tip of
the hair was thrust beneath the venter of the mite from the caudal end
until the entire body came to rest on the hair before lifting it off the
plant surface. SinCE:~. :phoenicis has a strong tendency to move in a
forward direction when disturbed, it was found best to pick up the mite
from the posterior so that the anterior end always faced the tip of the
hair. When the mite was picked up in this position, it crawled off the
hair readily onto the desired area of transference but when picked up
in the opposite position, posterior end facing the tip of the hair, the
mite invariably crawled up the hair towards the handle and made dis
lodging of it onto the plant surface difficult.
Whole fruits were used as rearing media for the life history
studies of B. phoenicis. lITanglefoot ll was used as a barrier to con
fine the mites on the fruits. All excess amounts of IITanglefoot ll were
scraped off with an edge of a microscope slide so that only a thin,
slightly tacky film was left along the inner margin and a thicker layer
left along the outer margin of the barrier. When liberal amounts of
"Tanglefoot" were deposited along the inner margin l • the active stages

16.
of the mite invariably crawled into the material and died; however,
when the excess was removed, they were prevented from being caught
in it and also the remaining deposit was sticky enough to discourage
them from walking over it. The thicker deposit of "Tanglefootlt on
the outer margin of the barrier not only provided assurance against
the confined mites from escaping but also prevented other mites from
invading the premises of the individuals under observation.
The life history studies were begun with the egg stage. For
each combination of temperature and humidity condition under which
the life cycle of~ phoenicis was carried out, the mites were confined
in two ways, individually and in mass. When confined individually, tp.e
starting population of 100 eggs was placed on two fruits; 50 eggs to a
fruit, and each egg was allocated an area of 1. 0 sq. cm. on the sur
face of the fruits. When confined in mass., the 100 eggs were placed
on one fruit in four batches of 25 eggs each. Each batch of eggs was
confined within an area of 4.0 sq. cm. of the fruit surface. The result
ing larvae from these eggs were allowed to feed and to complete their
protonymphal and deutonymphal stages within the original confines
allocated to them earlier in the egg stage. Upon completion of the
deutonymphal stage, 50 newly emerged females from each tempera
ture and humidity combination studied were selected; 25 females from
those confined individually and 25 from those cmfined in mass, and
transferred to. fresh fruits. The females that developed from eggs

17.
held individually and in mass were continued to be held in the res
pective manner until completion of the life cycle. The eggs deposi
ted by these females were counted and transferred from the fruit
surface onto 2. 0 cm. x 6. 0 cm. strips of adhesive tape on each ob
servational date. These eggs were exposed to the same temperature
and humidity conditions as the females which laid them to determine
thei.r 'h::otchability.
The fruits with the eggs confined on them were enclosed inside
of wide-mouth gallon jars and placed in constant tem];erature cabinets
for rearing. Within each jar, the fruit, together with a temperature
and humidity indicator (Airguide), were placed on top of a wire screen
platform which was elevated about 8.0 cm. above the bottom of the
jar (Fig. 2). Also, in the bottom of each jar, 650 cc. of a saturated
salt solution was added in order to maintain· the desired level of
humidity within the closed system. The salts used to obtain the follow
ing levels of relative humidity: 25 to 30 per cent, 65 to 70 per cent,
and 85 to 90 per cent, were potassium acetate, ammonium nitrate,
and potassium nitrate, respectively.
A modified refrigerator and a commercial incubator were used
as temperature cabinets. These cabinets were held in an air con
ditioned room where the temperature varied between 2loe and 240 e
and humidity between 60 and 70 per cent. The modified refrigerator
was used for temperatures below Z50 e and the incubator for
'"
Figure 20 Rearing unit used in the constant temperatureand humidity experiments.
180

19.
temperatures of 2SoC and above. Both of these cabinets were
equipped with thermo-regulators and thermostats of:l: O. SoC sensi
tivity. The thermo-regulator of each cabinet was adjusted and set to
the desired constant temperature with the aid of thermocouples and
a potentiometer. A mercury-in-glass thermometer was placed inside
each cabinet and this, together with the temperature and humidity
indicators enclosed within the jars, were referred to each day to in
sure that the relative humidity and temperature were maintained
relatively constant throughout the life cycle of the mites.
Once the mites were placed in the cabinets, they were taken out
only briefly whenever necessary to observe and record the various
life history events. Those mites which were confined individually
were taken out from the cabinets at least three times daily for about
lO minutes each time in order to obtain the necessary data for deter
mining the stadia of the different life stages, the preoviposition period
and fecundity of the females, and the other aspects of the life history.
Those mites which were held in mass were taken out not more than
once a week. Comparison of the survival populations from those con
fined individually and in mass revealed that isolation as well as the
frequent removal of the individually confined mites from the constant
temperature and humidity conditions to laboratory conditions had no
significant effect on the survival potential of~ phoenicis.

20.
Sections of papaya fruit embedded in agar were used as rearing
media for the chemical control studies of B. phoenicis. This type of
rearing medium as shown in Figure 3 was prepared as follows: A
2 per cent agar solution was heated to the boiling point and then poured
into an aluminum foil tart plate, 8.0 cm. in diameter, until filled to a
depth of approximately 1. 0 em. After the agar solidified, a piece of
papaya fruit, about 5.0 cm. in diameter, was placed cut surface down
in the center of the plate. Then, more agar solution, first cooled to
45 0 C, was poured until about one-half of the original surface of the
papaya section became embedded. The agar provided moisture and
kept the papaya section fresh for two weeks under conditions of 21 0 C
to 240 C and 60 to 70 per cent relative humidity and also served as an
effective escape barrier against the active stages of B. phoenicis.
Mites were transferred from the stock cultures onto rearing media
of thi s type a day prior to subjecting them to the different pesticides
shown in Table VI. Twenty-five females were placed on each plate
and four such plates, or 100 individuals, made up each treatment.
Of the active stages, only the adults were selected for exposure to
the different treatments for in preliminary studies they proved to be
more tolerant of these pesti,cides than the larvae and nymphs.
A settling mist tower which was improvised by Sanchez and
Sherman (1963) was used to apply the pesticides. The four plates of
each treatment were placed on the turntable which revolved around a
¥igure 3. Rearing unit used in the chemical control studies ofBrevipalpuB~~ (Geijskes)
Nt-4•

22.
Venturi tube through which 5.0 cc. of a pesticide was blown under
pressure of about 50 p. s. i. into the tower as a mist. The mist was
allowed to settle for three minutes onto the plates. These plates,
with the treated mites on them, were held for two weeks in an air
conditioned room where temperature ranged from 21 0 C to 240 C and
relative humidity from 60 to 70 per cent. Mortality records of the
mites were taken on the first, third, fifth, and seventh day after
treatment, and also on the seventh day all of the living females and
eggs present on the plates were counted. The living females were
removed at that time but the eggs were left in situ until hatching to see
if the pesticide residues on the media were still toxic to kill the larvae
which emerged 10 to IS days after the application of the pesticides.
In addition to the tests with the adults, eggs of B. phoenicis were
also subjected to the different pesticides shown in Table Vill, using
the dip method. Only the higher concentrations of most of the pesti
cides were used because the lower concentrations gave no significant
kill of B. phoenicis eggs. Each treatment consisted of four repli
cations of 25 eggs each, or a total of 100 eggs. They were placed on
2.0 cm. x 6.0 cm. strips of adhesive tape and dipped for a minute in
the various pesticides. These treated eggs were held for three weeks
in an air-conditioned room to see if the pesticides had any ovicidal
effect.
Since the effects of some of the pesticides on papaya were

23.

24.
DESCRIPTION OF ~IFE STAGES
The life stages of ~ phoenicis include the egg, larva, proto
nym.ph, deutonym.ph, and adult. Both sexes are present, but m.ales
are scarce and have not been found in many parts of the world where
this species exists.
lh: phoenicis is a very variable species, but it Can be readily
distinguished in the adult stage from. the other mem.bers of the genus
by having five pairs of dorsolateral hysterosom.al setae and two
sensory rods on tarsus IT (Fig. 9B). The larvae and nym.phs also
have five pairso£ dorsolateral hysterosom.al setae but, unlike the
adults, they have only one sensory rod located posteriodistally on
tarsus IT. Morphologically, the im.m.ature stages of.!h phoenicis
closely resemble those of.!h obovatu.s, and like them., they are sub
ject to considerable variation in the size and shape of some of the
dorsal setae. The num.ber and arrangem.ent of the setae on the
dorsum. of idiosom.a of the larva, protonym.ph, and deutonym.ph con
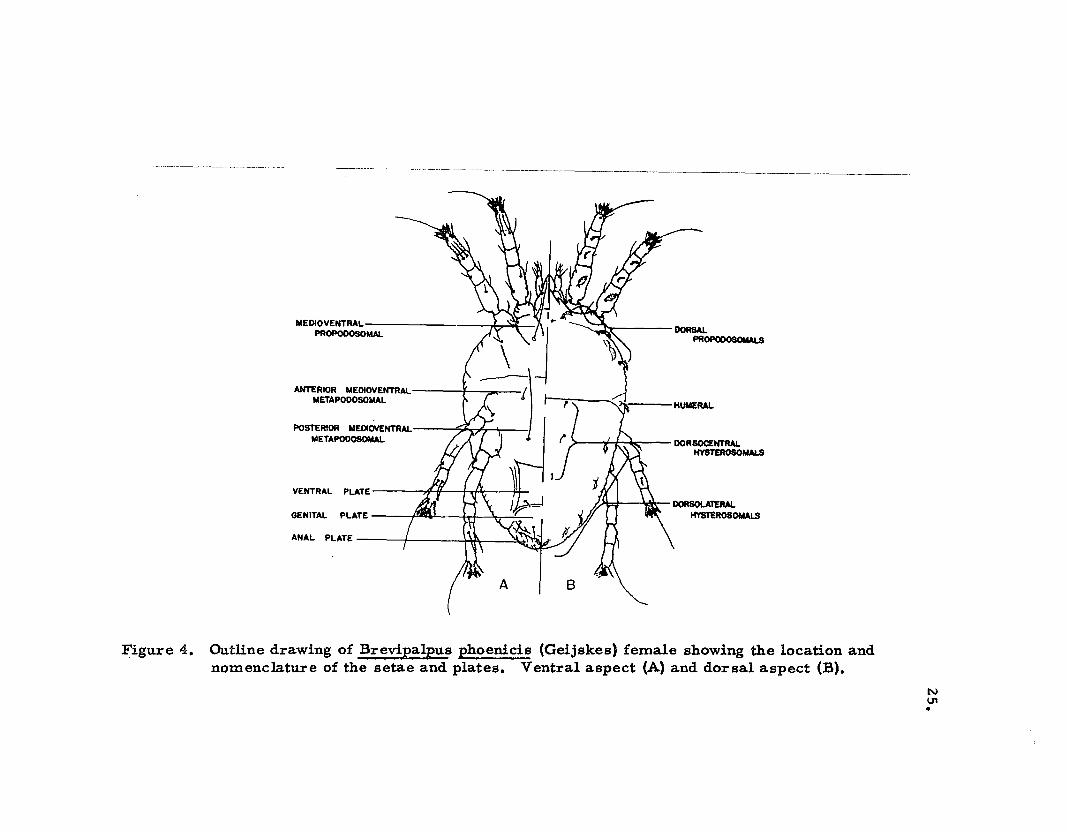
form to tho se of the adult and to the genus Brevipalpus ~'pritchard
and Baker, 1952). Twelve pairs of setae are preaent on the dorsum.;
three pairs on the propodosoma and nine pairs on the hysterosom.ao
The location and nom.enclature of these setae are shown in Figure 4.
The size and shape of the dorsal setae of mo st larvae and nym.phs on
papaya are like those shown in the respective illustrations (Figs. 6,
7, and 8), and as described in the sections on the life stages below.
A I B
I.>W ,..,~ DORSOUIT'ERALHYSTEROSOMALS
:::b DOflSALPROPClO<l8ClMALS
> 'i t. ~ DOR8OC£NTlW.1fYSTEROSOMAI.S
r- ,'\. <~ HUIll£RAL
MEDlOVENTRAL if> >:t 7'PROPOOOSOMAI..
ANTERIOR MEDlOVENTRAL y,...METAPOOOSOMAL
POSTERIOR MEDlOVENTRAL 9'i:,'S \.METAPOOO8OIlIAL
VENTRAL PLATE ,.., f/ I .it /'I
ANAL PLATE ' \ \1\. 'i~ ...( \\1 ~'Je....--'
GENITAL PLATE /Aql h I \0 :'_
~igure 4. Outline drawing of Brevipalpus phoenicis (Geijskes) female showing the location andnomenclature of the setae and plates. Ventral aspect (A) and dorsal aspect (B).
N1.11•

26.
Of the dorsal setae, dorsolateral hysterosomal setae I and II, and
dorsocentral hysterosomal setae ill are most variable in size and
shape. These vary from a tiny, serrate setae to large, broadly
lanceolate, serrate setae similar to dorsal propodosomal setae II and
ill. larvae and nymphs with these large setae were more frequently
encountered on passion fruit and bamboo palm than on papaya.
DeLeon (1961) found nymphs of most Mexican specimens of.!!:.
phoenicis with the large dorsolateral hysterosomal setae I and II, and
in three collections, nymphs with the dorsocentral hysterosomal setae
nearly as large as the dorsolateral setae.
The location and nomenclature of the setae on the venter of the
idiosoma are shown in Figure 4. The number of setae on the venter
is not constant but increases from four pairs on the larva, five pairs
on the protonymph, seven pairs on the deutonymph to eight pairs on
the adult. These additions take place in the hysterosomal region of
the body. A pair of medioventral opisthosomal setae is present in the
two nymphal and adult stages but not in the larval stage. The medio
ventral propodosomal setae, which are present in the larval, nymphal
and adult stages, and the posterior medioventral metapodosomal
setae, which are present only in the deutonymphal and adult stages,
are filamentous and smooth. The remaining ventral setae are short
and smooth.
The following descriptions of the life stages are based on

27.
individuals reared on papaya. All observations and measurements
were made on at least 25 individuals of each stage. Measurements
were taken on living spedmens since mounted ones often undergo con-
siderable distortion in size. Mites were placed on adhesive tape, to
keep them quiescent, and measured under 300 times magnification
using incident light. The width measurements of the larvae, nymphs,
and adults wer e taken at the level of the humeral setae, and the
length taken between the distal ends of the rostrum and opisthosoma
(Table IV). The meaSUremeI:lts shown for the larva, protonymph, and
deutonymph were taken on the inactive phases and represent the
average maximum size attained by individuals in the respective sta-
dium. The average minimum size for each stage is the same as the
measurements given for the stage preceding it.
TABLE IV. AVERAGE MEASUREMENTS OF THE LlFESTAGESOF BREVIPALPUS PHOENICIS (GEIJSKES)
StageLength (mm)
Mean S. D.Width (mm)
Mean S. D.
EggLarva, inactiveProtonymph, inactiveDeutonymph, inactiveAdult, female
0.1080.1710.2320.3080.308
0.0040.0050.0050.0080.007
0.0700.1090.1400.1630.163
0.0040.0040.0040.0050.004
S. D. = Standard deviation.

28.
The egg (Fig. 5) is elliptical and about 0.100 to 0.116 mm. long
and 0.060 to 0.080 mm. wide, averaging 0.108 I:. 0.004 mm. in length
and 0.070 I: 0.004 mm. in width. It is light orange, soft, and very
sticky when first laid. At this time, the egg adheres readily to any
surface it comes in contact with and attempts to remove it usually
result in its breakage. After a few minutes of exposure to air, it be
comes firm and bright reddish orange. The chorion is thin, trans
parent, and made up .of at least two layers. The outer layer is drawn
out to a fine stipe on the end of the egg which emerges last from the
ovipositor. This stipe is variable in length and is often missing for it
can be easily broken off by the mites trampling over it. The stipe is
also inconscpicuous in some cases for it bends back and adheres to the
surface of the main portion of the egg. When the egg is viewed under
450 times magnification with incident light, many close set irregular
striae are visible on the chorion; however, when it is mounted in
Hoyer's medium, the outer layer together with the stipe are dissolved
away and the clear, smooth, inner layer becomes exposed.
The protoplasm, which is translucent light orange when the egg
is first laid, completely fills the inside of the chorion and is in close
contact with it. With the advancement of the incubation period, a
space between the developing embryo and the chorion is formed. The
egg becomes opaque white a day before eclosion. The larva and its

,Figure 5. Egg of Brevipalpu6 phoenicis (Geijskes).Enlarged approximately 409 times.
29.

30.
bright red eyes are visible through the translucent white chorion at
this time.
larva
The hexapod larva (Fig. 6) is bright orange-red when newly
emerged and 0.128 to 0.140 mm. long and 0.080 to 0.096 mm. wide,
averaging 0.135 I:. 0.004 mm. in length and 0.088 I:. 0.005 mm. in
width. When fully grown and ready for molting, it is opaque orange
and 0.171 I:. 0.055 mm. in length and 0.109 I:. 0.004 mm. in width.
Two pairs of red eyes are present on the lateral margins of the pro
podosoma. EKcept for the suture between the propodosoma and the
hysterosoma, and the irregular, folds along the lateral margins of the
idiosoma which gradually disappear as the larva grow, there are no
characteristic integumental markings on the dorsum. The integument
on the venter of the idiosoma, however, has regular parallel striae;
transverse on the propodosoma and anterior portion of the opisthoso
ma, and longitudinal on the metapodosoma and laterad of the anus.
On the dorsum, the propodosomal setae II and ill, humeral
setae, and the dorsolateral hysterosomal setae ill and V are serrate,
broadly lanceolate, and O.OIl to 0.014 mm. long and 0.004 mm.
wide. The dorsolateral hysterosomal setae IV are filiform, 0.042
mm. in length, and the remaining dorsal setae are serrate and
short, less than 0.004 mm. (Fig. 6).

;Figure 6. larva of Brevipalpus phoenicis (Geijskes).Dorsal aspect. Enlarged approximately364 times.
31.

32.
On the venter of the idiosoma, four pairs of pilose setae are
present; a pair of long medioventral setae on the propodosoma, a pair
of short medioventral setae on the metapodosoma, and two pairs of
short anal setae on the opisthosoma.
Protonymph
The protonymph (Fig. 7) differs from the larva essentially in
being larger and in possessing four pairs of legs. Scattered light
green, orange, black, and yellow patches, resulting from accumu
lations of food and waste matter inside the body, can be seen through
the translucent integument of the idiosoma. The protonymph grow s
from a minimum size of 0.171 mm. to a maximum size of 0.244 mm.
in length and from 0.109 mm. to O. 148 mm. in width, averaging
0 ..2'32 I: 0.005 mm. in length and 0.140 ~ 0.004 mm. in width as an
inactive protonymph. Except for the suture between the propodosoma
and hysterosoma and the few folds which gradually disappear as the
protonymph grows, no characteristic markings are present on the
integument of the dorsum. On the venter, however, there are regular
parallel integumental striae running transversely across the central
portion of the idio soma with the ends bending longitudinally to the
lateral margins. Longitudinal striae are present between the medio
ventral setae of the propodosoma and the base of the gnathosoma.
On the dorsum p the propodosomal setae II and ill, humeral
setae, and the dor solateral hysterosomal setae ill, IV, and Vare

Figure 7. Protonymph of Brevipalpus phoenicis (Geijskes).Dorsal aspect. Enlarged approximately234 times.
33.

34.
broadly lanceolate and serrate. All of them are about the sam.e size,
0.014 mm. to 0.016 rr..m. long and 0.006 rom. at the widest part.
The remaining setae: propodosom.al setae I, dorsolateral hystero
somal setae I and II, and dorsocentral hysterosomal setae I, II, and
m, are serrate and very small, less than O. 004 mm. long (Fig. 7).
On the venter of the idiosom.a, there are five pairs of pilose
setae; one pair of medioventral setae on the propodosoma, metapodo
soma, and the opisthosoma. The setae on the propodosoma are long,
I 0.028 mm. to 0.031 mm.., while those on the m.etapodosom.a and
opisthosoma are short, O. 004 mm. to o. 006 rom. Two pairs of setae,
similar in length to the medioventral opisthosomal setae, are present
on the anal sc1erites.
Deutonymph
The deutonymph (Fig. 8) is similar to the protonymph in color,
dorsal chaetotaxy, and in the pattern of the st riae on the venter of the
idiosoma. In this stage, two additional pairs of pilose setae are
present on the venter of the idiosoma: a pair of long posterior medio
ventral metapodosomal 'Setae, 0.031 mm. long, and another pair of
short setae, 0.006 mm., near the anterior margin of the anus similar
in size and shape to those located below on the anal sc1erites. When
fully grown and ready to molt, the size of the deutonymph ranges from
0.292 mm. to 0.324 mm. long and 0.156 to O. 168 mm. wide, averaging

Figure 89 Deutonym.ph of Brevipalpus :phoenicis (Geijskes). Dorsal aspect (left). Ventralaspect (right). Enlarged approximately 263 times.
W\J1•

36.
0.398 1:. 0.008 m.m.. in length and 0.163 1:. 0.005 mm.. in width.
Female
The female (Fig. 9) ranges in size from 0.288 rom. to 0.312
m.m... in length and 0.156 rom. to 0.176 mm.. in width, averaging
0.302 t 0.007 rom. in length and 0.162 t 0.004 rom. in width. The
body coloration of the females is very variable. Differences in age,
food, and temperature conditions have great influence on body color
ation. In newly emerged females, the area between the red eyes is
bright orange and the remainder of the idio soma is translucent light
yellow with a few brownish patches occurring mediolaterally on the
hysterosoma. When feeding begins, a conspicuous black pattern in
the shape of an "Hit appears on the idiosoma (Fig. 10E to H). The
black pattern enlarges, coalesces, and covers almost the entire
idiosoma of some individuals (Fig. 101). This black pattern gradual
ly disappears and the females once again assume a uniform carmine
body color a few days prior to death. Females reared at a tempera
ture of 300 C have a uniform carmine body color and those reared at
200 e and 250 e usually have the black patterns on the idiosoma.
Females on passion fruit are mostly reddish orange and have less
variations in body coloration than those on papaya (Fig. lOA to 1).
On the dor sum, the integument has reticulations mediolaterally
on the propodosoma and hysterosoma. A pair of pores are present..

Figure 9. Female of Brevipalpus phoenicis (Geijskes). Dorsal aspect (A) palpus (B)tarsus IT (left). Ventral aspect (right). Enlarged approximately 300 times. UJ
-.:I•

E
38.
Figure 10. Variation in color patterns of females ofBrevipalpusphoenicis (Geijskes).

39.
mediolaterally on the propodosoma and metapodosoma. Propodo-
somal setae I, II, and ill, humeral setae, and do-rsolateral hystero-
somal setae I, II, ill, IV, and V are narrowly lanceolate, serrate,
and less than 0.010 mm. in length. The three pairs of dorsocentral
setae are minute at;ld setiform.
On the venter of the idiosoma, the reticulations on the ventral
plates and area directly anterior to them are wider than long and
those laterad to them are longer than wide. Eight pairs of pilose
setae are present: a pair of medioventral propodosomal setae, two
pairs of medioventral metapodosomal setae, a pair of setae on the
ventral plate, and two pairs each on the genital and anal plates. The
medioventral propodosomal and the posterior medioventral metapo-
dosomal setae are filamentous and about 0.070 mm. long. The pair
of anterior medioventral metapodosomal setae and those on the ven-
tral and genital plates are similar in length and about one-fifth the
length of the posterior medioventral metapodosomal setae. The anal
setae are the shortest of the setae on the venter and about one-half
the length of the genital setae.
Male
The male (Fig. 11) is wedge-shaped in outline and flattened in
profile. The idiosoma is reddish and has no black marldngs like
those on the females.

40.
On the dorsum, two transverse sutures demarcate the propo
do somal, metapodosomal, and the opisthosomal regions of the body.
The integument has rather even reticulations mediolaterally and ir
regular reticulations mediodorsally on the propodosoma, metapo
dosoma, and the opisthosoma. A pair of pores is present on the
propodosoma and the metapodosoma. The dorsal chaetotaxy is
similar to that of the female.
On the venter of the idiosoma, reticulations are present medio
laterally along the bases of attachments of legs il, ill, and IV, on the
opisthosoma, and medioventrally along the posterior margin of the
metapodosoma. The medioventral propodosomal and the posterior
medioventral metapodosomal setae are filamentous and very long,
0.070 mm. or more in length. The anterior medioventral metapo
do somal and the medioventral opisthosomal setae are pilose and very
short; the former about one-fifth and the latter about one- sixth the
length of the posterior medioventral metapodosomal setae. Three
pairs of short, very narrowly lanceolate, and serrate setae are
present on the genito-anal sc1erites.

Figure 11. Male of Brevipalpus phoenicis (Geijskes). Dorsal aspect (A) palpus (left).Ventral aspect (right). Enlarged approximately 300 times.
II:>-J-004.

I'··.···.·········,
42.
LIFE HISTORY
The following is the life history of B. phoenicis based on infor-
mation ob~ined from laboratory rearings conducted under different
levels of relatively constant temperature and humidity conditions.
These two physical factors have been shown to greatly influence the
developmental and reproductive rates of several species of mites
(Morishita, 1954; Boudreaux, 1958; and Nickel, 1960). The purposes
of this life history study were to determine the approximate tempera-
ture and humidity combinations which favor natality and their ranges
which the various stages of B. phoenicia can tolerate. It is hoped
that the information obtained from this study will be helpful in better
understanding the other biological aspects of this mite. Food and
space were kept constant and provided in adequate amounts so as to
minimize intraspecific competition for them. In addition, other
factors that could influence the life history of~ phoenicis, such as
natural enemies and interspecific competition, w:ere excluded in the
present study.
Egg Stage
The egg is deposited in any position in cracks, crevices, exuviae,
and other protected niches on the plant surface. It is laid singly but
since the females tend to reuse the same ovipositional sites, many
eggs are often clustered together. These clusters of bright reddish
orange eggs are more readily seen with the unaided eye than the other

stages of the mite. When an egg is ready for ec1osion, one end 01' ~!
becomes swollen due to the larva pushing against the inner wall of the
chorion with its rostrum. With incident light and at magnifir3tions of
300X or higher, the larva can be seen repeatedly clawing tn€' h~!lel" waH
of the chorion with its front pairs of legs as if tTying to malt€' <In 0pE'n
ing for emergence. After a while, the chorion splits ~l"tlv around on".
end of the egg at approximately the level of clawing. The larva &t~_ '':>ll
out one of its front legs through the split and uses it to enl~u'ie the
opening further. The larva then emerges anterior end first a.nd in itw:
process pushes aside one end of the chorion which remains conflened
like an "hinged door" to the remainder of the chorion. The empty
transparent chorion remains attached to the plant surface for a long
time unless forcibly removed.
The eggs were found to be greatly affected by the prevailing tt>!1:;-
perature and humidity conditions. Their incubation period and ha.td~··
bility varied significantly between the different temperatures inespE'i.:
tive of humidity conditions; however, between humidity levels of 63 t(.)
70 per cent and 85 to 90 per cent at a given temperature, they wert':'
essentially the same (Table V). Within each temperature. hatchabil.ity
was lowest at the 25 to 30 per cent humidity level and the incubation
period about a day longer than at the higher levels of humidity. The
minimum incubation period was 8.0 days at 30°C in combination with
65 to 70 per cent relative humidity and the ma.'timum 24.8 days at ZOoC
in combination with 85 to 90 per cent relative humidity. Under the

43.
stages of the mite. When an egg is ready for eclosion, one end of it
becomes swollen due to the larva pushing against the inner wall of the
chorion with its rostrum. With incident light and at magnifications of
300X or higher, the larva can be seen repeatedly clawing the inner wall
of the chorion with its front pairs of legs as if trying to make an open
ing for emergence. Mter a while, the chorion splits partly around one
end of the egg at approximately the level of clawing. The larva sticks
out one of its front legs through the split and uses it to enlarge the
opening further. The larva then emerges anterior end first and in its
process pushes aside one end of the chorion which remains connected
like an "hinged door" to the remainder of the chorion. The empty
transparent chorion remains attached to the plant surface for a long
time unless forcibly removed.
The eggs were found to be greatly affected by the prevailing tem-
perature and humidity conditions. Their incubation period and hatcha
bility varied significantly between the different temperatures irrespec
tive of humidity conditions; however, between humidity levels of 65 to
70 per cent and 85 to 90 per cent at a given temperature, they were
essentially the same (Table V). Within each temperature, hatchability
was lowest at the 25 to 30 per cent humidity level and the incubation
period about a day longer than at the higher levels of humidity. The
minimum incubation period was 8.0 days at 30 0 C in combination with
65 to 70 per cent relative humidity and the maximum 24.8 days at 20 0 e
.in combination with 85 to 90 per cent relative humidity. Under the

TABLEV o EFFECT OF TEMPERATURE AND HUMIDITY ON THl(GEUSKES)
Stage R E L A T I V E&: 25 to 30 Per Cent 6!
Temperatur e Duration (days) I
°c N Range MeantS. D. N Rani
Starting N 100 100(Egg)
~20 5 21.4-23.4 23.0iO.2 67 19.8·25 8 8.7-12.6 11. 1~I. 1 88 9. I·30 0 27 8. o·
Larva20 No survivors beyond 65 9.8·25 egg stage 88 4.8·30 26 3. 1·
Protonymph20 59 7.8·25 87 4.530 26 2.6
Deutonymph20 50 3.525 85 4.730 25 3.5
Immature StageEgg to Adult20 50 17.425 85 23.130 25 16.5
Adult, FemalePreovipo sition20 25 3.225 25 2.130 16 7.0Fecundity20 25 525 25 4130 25 0Longevity20 25 13.425 25 21. C30 25 3. e

~ ON THE,DIFFERENT STAGES OF BREVIPALPUS PHOENIGIS
44.
[)
lVE HUMIDITY65 to 70 Per CentDuration (days)
Range MeantS. D. N
100
85 to 90 Per GentDuration (days)
Range MeantS. D.
7 19.8-24.6 22.2Jl.l 65 18.8-24.8 22.61:.1. 18 9.1-13.0 9.4~0.6 85 8.0-12.9 9.4tO• 87 8.0- 9.0 8.2/:.0.4 29 7.8- 9.2 8.4tO• 9
5 9.8-12.4 10.5~0.7 62 9.5-11.9 10.4tO• 58 4.8- 8.9 6.71:.0.8 83 5.8- 7.8 6.5~0.7
6 3.1- 4.5 3.6.J.0.2 27 3.6- 4.5 3.71:.0.4
9 7.8- 9.5 8.21:.0.7 60 7.7-10.2 8.4/:.0.97 4.5- 9.6 6.31:.0.9 83 4.0- 9.6 6. 5~1. 16 2.6- 3.7 3.11:.0.4 27 2.7- 3.5 3.0JO.5
0 3.5- 4.1 7.51:.0.9 58 2.7,- 4.1 7.3tI.l5 4.7- 9.1 6.8~0.8 83 4.3- 8.2 6.91:.0.55 3.5- 3.9 3.71:.0.5 26 3.8- 4.0 3.91:.0.1
17.4-20.423.1-34.516.5-19.0
48.81:.1.529.31:.0.418.6~0.5
588326
45.4-50.522.0-33.016.8-19.8
47. 7f:.2. 529.31:.1.218.11:.0.2
:5 3.2- 9.5 5. 7tI. 4 25 3.5- 9.0 6.2/:.2.3~5 2.1- 6.7 3.51:.0.9 25 2.3- 6.5 3.71:.0.6[6 7.0- 8.0 7.31:.0.2 14 7.5- 9.0 8.0~0.3
~5 5-29 10.61:.4.5 25 5-20 12.4J5.6~5 41-73 57r51:.10.7 25 8-68 53.3t7.5~5 0-19 6.41:.2.4 25 0-17 7.5~1.7
~5 13.4-72.3 45.8/:.9.7 25 15.0-70.3 47.01:.12.1~5 21.0-44.0 34.81:.6.0 25 25.3-50.0 34.51:.3.5~5 3.8-20.8 10. 5t3~ 7 25 4.5-12.0 7.5J2.0

45.
foregoing conditions, however, only a few of the eggs hatched; 27 per
cent under the former and 63 per cent under the latter conditions.
Optimum hatch of 88 per cent was obtained at 25°C in combination
with 65 to 70 per cent relative humidity; however, the average incu
bation period in this case was slightly longer than that at 300C with
the same level of humidity but the majority of the eggs hatched in 9.4:1:
0.6 days or less than one-half the time required by those subjected to
20°C. Some eggs hatched at 20°C and 25°C in combination with all
three levels of humidity, but at 30°C, hatching occurred only at the
65 to 70 per cent and 85 to 90 per cent levels and none hatched at the
25 to 30 per cent level.
None of the eggs hatched when exposed for several days to con
stant temperatures below 20°C and above 30°C in combination with
either high or low humidity. The eggs maintained the shape and color
of freshly laid ones for about a month at temperatures below 20°C but
they became shriveled and opaque white in about four days at tem
peratures above 32°C, especially when humidity was low.
Although continuous exposure of eggs to 20°C and 30°C caused
considerable reduction in hatchability, an exposure of 24 hours first
to either of these temperatures and then maintaining them at 25°C
with 65 to 70 per cent relative humidity had no detrimental effect on
hatchability. However, when temperature was decreased below 20°C
and increased above 30°C, short exposures caused hatchability of eggs

46.
to decline gradually in the former and abruptly in the latter case.
Hatchability of eggs which were first exposed to the different high and
low temperatures for 24 hours and then maintained at 250 C in combi
nation with 65 to 70 per cent relative humidity, declined from 89.5:1:
3. 0 per cent at 300 C to 45.0:1: 5. 5 per cent at 320 C to O. 0 per cent at
340 C or a 100 per cent reduction in a temperature change of only four
degrees; but at the low temperatures, it declined from 87.5 :I: 4.5 per
cent at 200 C to 41.0 :I: 4.5 per cent at 150 C to 27.5 :I: 6.0 per cent at
100 C or a reduction of some 68 per cent in ten degree change in tem
perature.
Immature Stages
The larva, protonymph, and deutonymph each spends about one
half of its stadium as an active and one-half as an inactive individual.
Feeding, growth, and limited dispersion take place during the active
phase and transformation into the subsequent stage during the inactive
phase. Prior to becoming inactive, the mite assumes a characteristic
position. The chelicerae are fully extruded and inserted into the plant
tissue; the front two pairs of legs are extended straight forward and
drawn close to each other; and the posterior legs, one pair in the case
of the larva and two pairs in the case of the nymphs, are extended
straight backwards and held close to the sides of the opisthosoma.
This characteristic position was assumed to be the ending of the active
and the beginning of the inactive phase of each stage. Although the

47.
chelicerae are inserted in the plant tissue during the inactive phase,
they are not used for obtaining nourishm.ent but are used for an-chor
ing the body in place. Besides using the chelicerae for anchoring,
an adhesive substance is apparently secreted by the mite just prior
to becoming inactive for the entire ventral surface of the body is
fastened to the plant surface as though glued and prying is needed to
dislodge an inactive mite. Ecdysis progresses normally even though
such a mite is dislodged soon after becoming inactive but the sub
sequent stage has some difficulty in freeing itself from the exuvium
at the time of molting. When an inactive larva is mounted in Hoyer's
medium and examined under 300X or better magnification with trans
mitted light, the outline of the developing protonymph can be seen
inside the exo skeleton of the larva. Leg s I, II, and III of the proto
nymph are held inside the corresponding legs of the larva but since
the latter has only three pair s of legs, legs IV of the former are held
free along each side of the opisthosoma.
The habits of all the immature stages are similar except feed
ing, growth, and disper sion become more pronounced with. the
passing of each stage. The newly emerged larva wander s in the im
mediate vicinity of the chorion from which it has emerged for about
an hour before commencing to feed while the newly emerged proto
nymph and deutonymph move further away from their exuviae and
com.m.ence feeding almost im.rn.ediately. The feeding damage done by

48.
the larva is hardly noticeable but that inflicted by the later stages is
readily discernible for the imm.ediate area around each feeding
puncture becomes discolored and blisterlike soon after the insertion
of the chelicerae (see page 57 for further descriptions of feeding
dam.age). The larvae and nymphs seldom leave the area where they
had emerged from the egg stage. They feed avidly and grow rapidly
during the active phase and undergo transformation just as rapidly dur
ing the inactive phase when environmental conditions are favorable.
The temperature and humidity conditions favoring egg hatch also
proved suitable for larval and nymphal survival. At 250 C in combi..
nation with 65 to 70 per cent relative humidity, conditions under which
optimum hatchability was obtained, the majority of the newly emerged
larvae successfully passed through the three immature stages in
19.8 f:. 0.9 days. Development of the immature stages from egg hatch
to adult emergence was most rapid at 30°C, 10.6 t 0.3 days, and
slowest at 20oC, 27.3 t 1.7 days. Some individuals successfully
reached the adult stage under constant temperatures of 200C, 25°C,
and 30°C in combination with either 65 to 70 per cent or 85 to 90 per
cent relative humidity, but none survived beyond the larval stage when
humidity was 25 to 30 per cent (Table V). Except for the temperature
and humidity combinations of 25°C with 65 to 70 per cent and 85 to 90
per cent at which more than 80 per cent of the starting population suc
cessfully completed the egg, larval, and the nymphal stages, the other

49.
combinations caused high larval mortality in addition to the low egg
hatch so that less than 35 per cent of the starting population reached
the adult stage. But once passed the larval stage, the protonymphs
reached the adult stage without much mortality for the nymphs were
more tolerant of the adverse conditions than the earlier stages
(Table V).
oProlonged exposures to temperatures above 30 e and below
200 e were fatal to the immature stages regardiess of humidity condi-
tions. The larvae died within two days after being exposed to 320 e,
whereas they became motionless, ceased to develop beyond that stage,
and remained alive for at least 10 days when exposed to 180 e. Some
larvae remained alive for as long as a week at temperature as low as
100e and when transferred to favorable temperature and humidity
conditions, 250 e in combination with 65 to 70 per cent relative
humidity, they continued development.
Adult Stage
The adult stage was reached in I~. 6 i. 0.5 days at 300 e and in
48.8 I:. 1.5 days at 20 0 e after the eggs were laid. At these extremes
in temperature, only few of the starting population of eggs success-
fully reached the adult stage despite favorable humidity conditions
(Table V). Completion of the life cycle was not possible when tem
perature was above 30 0 e and below 20 0 e regardless of humidity
conditions, and when humidity was only 25 to 30 per cent regardless

50.
of temperatur'e conditions. Survival was highest at 250 C in cOUlbi
nation with either 65 to 70 per cent or 85 to 90 per cent relative
humidity as 85 and 82 individuals reached the adult stage out of each
of the starting population of 100 eggs, respectively. All of the adults
that were reared from the egg stage under the different temperature
and humidity combinations were females, and thus, the discussion that
follows on the various life processes of the adult stage is on the
female sex.
Preoviposition Period: The female starts to feed soon after freeing
herself from the deutonyxnphal exoskeleton and must feed before egg
laying can comm.ence. This period between emergence and deposition
of the first egg, referred to as the preoviposition period, wa,s as
shown in Table V for the different temperatures in combination with
65 to 70 per cent and 85 to 90 per cent relative hwnidity. Although
the durations of all life processes discussed thus far, such as eclo
sion, ecdysis, life cycle, etc.: were shortest at 30oe, the preovi
position period was longer at this temperature than at 200 e and
250 e. Moreover, continuous exposure to 300 e probably affected the
reproductive physiology in the early stages of development for many
of the females failed to lay any eggs even though they were trans
ferred to more favorable temperature and humidity conditions soon
after reaching the adult stage. Females in which the earlier stages

51.
were exposed to constant temperature of 25 0 C, however, started to
lay eggs earlier when exposed to 300 C soon after becoming adults
than those kept continuously at 250 C from the egg stage to the end
of the preoviposition period.
Oviposition: When ready for oviposition, the female locates a suit
able site such as an exuvia, crack on the plant surface or a bud axil
for depositing the egg. She first examines and cleans out all debris
from the site with the front pair of legs, turns herself 180 degrees so
that the tip of the opisthosoma now is in contact with the ovipositional
site, and then deposits the egg after about a minute of alternate con
tracting and expanding of the body. After depositing an egg the female
leaves the oviposition site and resumes feeding in its proximity until
the next egg is ready for deposition some six hour s later at the earli
est, at which time she usually returns to the former site to deposit
it. Because of this habit of returning to the same site for oviposition
and since many females frequently oviposit in a common site simul
taneously, several eggs are massed together even though only one
egg is laid per individual during an extended duration. The female
seldom wandered afar but remained within the confines of the original
area in which she was reared as long as food was ample and suitable.
When intraspecific competition for food and space became intensive
due to overcrowded conditions, the females emigrated to uninfested

52.
parts of the same plant or to other plants and continued feeding and
oviposition.
The number of eggs laid by the females varied greatly with
temperature differences when humidity was kept constant (Fig. 12
and Table V). At 25 0 C, all of the females began laying eggs and con
tinued to lay from one to four egg s per day per individual during their
life span. One of the females laid a maximum of 76 eggs in 42 days
of existence and these were deposited equally during the day and night
without any peak periods of oviposition. Out of the 562 eggs laid
during a 24-hour period by 300 females which were kept under con
stant temperature of 25uC in combination with 65 to 70 per cent
relative humidity, 271 were deposited between 8:00 a. m. and 8:00
p. m. and 291 between 8:00 p. m. and 8:00 a. m. These eggs were de
posited uniformly throughout the day and night without any peak hours
of oviposition. Oviposition was very irregular at 200 e and 300 C and
it ceased at temperatures below and above these temperatures, res
pectively. All of the females laid some eggs at 20 0 e but over 50 per
cent of them failed to lay eggs at 30oe. At these two temperatures,
the number of eggs laid per female was less than one-fifth the num
ber laid at 250 C. When gravid females were subjected to tempera
ture of 18oC, they laid no eggs as long as they were held at this
temperature, but when returned to 25°C, they commenced to lay
eggs again after 8 to 10 hours. Some females survived exposure to

53.
Z<lW:E
2 3 4 II 8
WEEKS7 8 9 10
,Figure 12. Mean number of eggs laid per week by four startingpopulations of 25 Brevipalpus phoenicis females.

54.
constant temperature as low as 100 C for 23 days and wh.en returned to
more favorable conditions, 25°C and 65 t?_ 79 per cent relative humi
dity, they started to lay eggs after a day.
Longevity: Some of the more obvious factors which affected the
longevity of the adults were food, temperature, and humidity. When
food was withheld, the adults died from. starvation within three days.
The adults were m.ore susceptible to temperature changes than to
humidity changes and they tolerated the changes better than the im.
mature stages. The adults lived longer under tem.perature conditions
below 25°C than above it (Fig. 13 and Table V). The average longevi
ty was 47.0 t 12.1 days at 20°C but only 7.5 t 2.0 days at 30°C in
com.bination with 85 to 90 per cent relative humidity. At 30°C, less
than 25 per cent of the adults were alive at the end of two weeks
while 75 per cent of them were still alive at the end of five weeks at
20°C. Although the females lived longer at 20°C than at 25°C, they
laid only a few eggs, remained quiescent m.ost of the time, and
caused very little feeding injuries. At 2SoC in com.bination with 65
to 70 per cent relative hum.idity, the m.ajority of the fem.ales lived for
over a m.onth, laid m.ore eggs than at other temperature and humidity
com.binations, and oviposition continued daily up to a day prior to
death.. For each respective decrease and increase in temperature
below 20°C and above 30°C, longevity of the adults shortened at a

55.
faster rate at the high than at the low tem.perature levels. At 3ZoC,
none of the adults were alive after 7 days, whereas at lOoe som.e
were still alive after 23 days.

90
80
70>I-:Jj:!600:::o~
50
I-ZW040
0:::Wa.. 30
20
10
56.
23456
WEEKS7 8 9
Figure 13. Longevity of the adults of Brevipalpusphoenicia (Geijskes) at the differenttemperatures in combination with 65to 70 per cent relative humidity.

57.
BIOLOGICAL OBSERVATIONS
Molting
B. phoenicia, like most other tetranychids, molts three times
during its life cycle; at the end of the larval, protonymphal, and the
deutonymphal stages. In the process of molting, the integument to
be cast off splits transversely across the dorsum slightly in back of
the humeral setae, along the lateral margins of the podosoma, and
transversely across the venter at the level of the frontal margin of
the anus. The emerging mite pushes itself backwards, frees the
front two pairs of legs from inside the exoskeleton of the previous
stage and, in doing so, the exuvia separates into t"w-v sections along
the splits mentioned above. The small posterior section remains
attached to the opisthosoma of the newly emerged mite until rubbed
off and the large anterior section remains attached to the plant sur
face for a long time and serves as a common site for oviposition.
When the front legs are freed, they are used to withdraw the rest of
the body from the exuvium. The molting process, from the appear
ance of the split on the dorsum until the emergence of the mite from
the exuvium required from 20 to 45 minutes at 250 C in combination
with 65 to 70 per cent relative humidity.
Feeding
B. phoenicis is an active feeder, feeding at all hours of the day
and night during the greater part of its active phases. Feeding is

58.
especially intense and its resulting damage very extensive when tem
perature is between 250 C to 30 0 C and relative humidity high. This
species feeds on the stems of many kinds of plants; however, when
high population density prevails, some individuals emigrate to other
parts of the plant. On papaya plants, this mite usually feeds on the
trunk at the l.,vel where the bottom whorl of leaves are attached. As
intraspecific competition for food and space intensifies, the mites
feed upwards on the trunk and outwards onto the leaf petioles and
fruits, leaving a large conspicuous damaged area behind them.
In feeding, the needlelike chelicerae are used to puncture the
epidermal cells. The sap that oozes out from the wounded cells is
mixed with saliva and imbibed into the digestive tract of the mite.
The immediate area around the feeding puncture becomes raised and
blisterlike as though caused by a toxic substance. Later, the affected
tissue dries up, dies and becomes discolored. Since many feeding
punctures occur close together, the affected areas coalesce to form
a large continuous calluslike, tannish, scaly and/or scabby area
(Fig. 14). The feeding damage is very pronounced when young papaya
fruits are attacked for the affected areas become sunken due to the
differential growth of the injured and uninjured tissues. The mites
sometimes puncture the latex glands while feeding, causing a copious
outflow of a milky white liquid which mars the appearance of the
fruit (Fig. IS). All stages of the mite in the path of the flow of the

":-."";""', '..
Figure 14. Feeding injury caused by Brevipalpusphoenicis (Geijskes) on papaya fruit.
59.

60.
sticky latex are engulfed and drowned in it. The stem of papaya
which normally remains green for a long time takes on a tan,
suberized appearance prematurely and makes a spindly growth when
heavily infested by B. phoenicis.
A disease of orange known as Lepra explosiva in Argentina,
originally thought to be caused by a fungus (Marchinatto, 1935) and
later by a virus (Marchinatto, 1938; and Blanchard, 1939), is now
attributed to toxins injected by ~. obovatus in the process of feeding
(Carter, 1952). B. phoenicis has been collected from an orange tree
exhibiting symptoms of Lepra explosiva in Paraguay (Nickel, 1958).
In addition to the feeding injury described above on papaya, B.
phoenicis causes pitting and splitting of the sld.n of orange fruits
(Planes, 1954), scarring of tangerine fruits (Nickel, 1958), galling
of sour-orange seedlings (Knorr, et al., 1960), and defoliation and
vine dieback of passion fruit (Fig. 16).
Mating
Mating is seldom seen because 'of the scarcity of B. phoenicis
males. During the two years of study, only six matings were ob
served. In every case, the males mated with non-gravid females
about a day old and remained in copula for about 15 minutes. In
mating, the male approached the female from the posterior, rested
his two pairs of front legs on the dorsum of the female's opisthosoma,
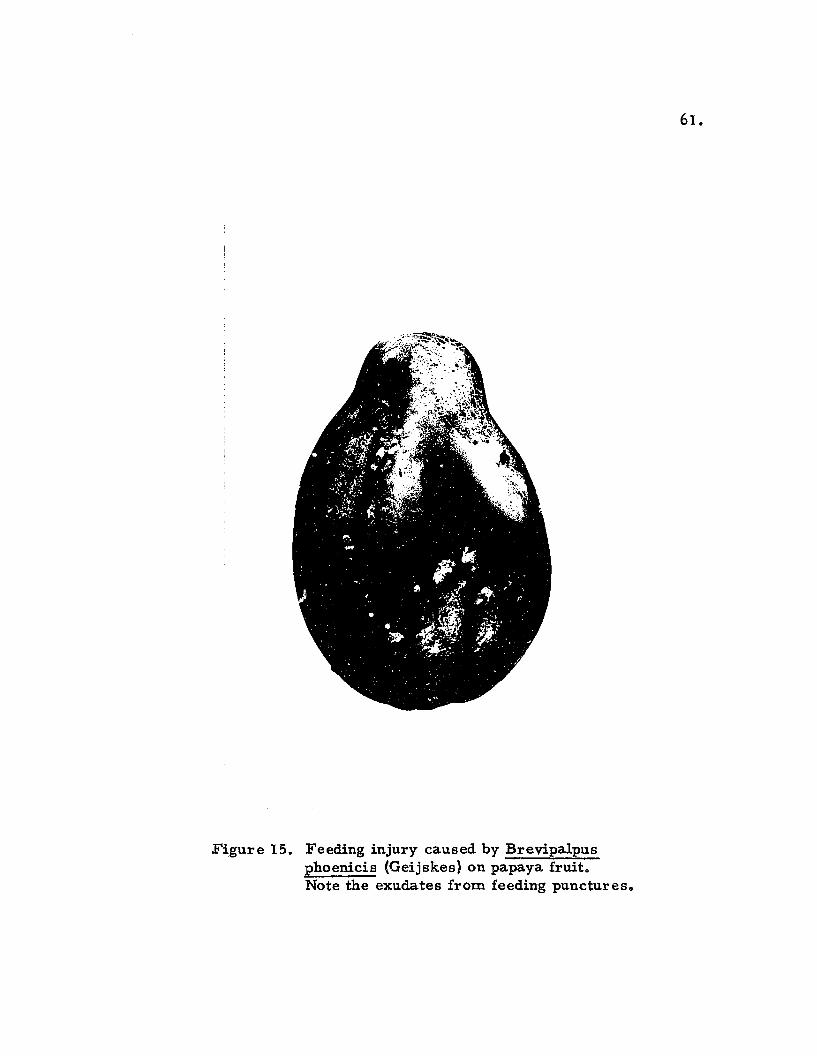
Figure 15. Feeding injury caused by Brevipalpusphoenicis (Geijskes) on papaya fruit.Note the exudates from feeding punctures.
61.

Figure 16. Feeding damage caused by Brevipalpus phoenicis (Geijskes)on passion fruit. a
N.

63.
and crawled beneath the opisthosoma so that one-half of his body was
beneath hers. Simultaneous to crawling beneath the female, the
opisthosoma of the male was bent upward and forward in a form of a
Ilell until the tip came in contact with the posterior end of the female's
opisthosoma. Attached to the female in this position, the male
walked along on his two hind pairs of legs, holding on to the dorsum
of the female's opisthosoma with the front two pairs of legs and fol
lowed her wherever she went. These mated females produced only
female progenies as did the unmated females under laboratory con
ditions so it was not possible to find out if the males were functional
or otherwise. Cytological confirmation of this was unsuccessful for
the standard method of chromosome determination using the squash
technique with either aceto-orcein, aceto-carmine or Snow's stains
failed to bring forth the chromosome complement of this species al
though hundreds of eggs and adults were treated and examined.
Sex Ratio
As stated earlier, males have not been found in many parts of
the world where this species is known to occur. Here in Hawaii,
males comprised less than 1 per cent of the 6, 774 adults collected
from various localities and at different seasons during 1962-63.
There were no indications of seasonal or local abundance of males.
In the laboratory, several generations of.!!.:. phoenicis were reared
without males. No males were obtained from females of all ages

64"
which were reared under the different temperature and humidity con
ditions. Also, mated as well as unmated females produced only
female progenies. These rearing data, plus the occurrence of pre ..
ponderance of females in nature, indicate that parthenogenisis is the
principal mode of reproduction in this specie s.
Dispersion
Man, in transporting plants from place to pl.ace, has unknowing ..
ly disseminated B. phoenicis to many distant places. The ability of
this mite to subsist on many different kinds of plants has certainly
enhanced its rapid spread and establishment in many tropical areas
of the world.
Within a given area, birds, insects, mammals, wind, and rain
have been incriminated as disseminating agents of many organisms.
Of these, wind has been proven to be the most important agent of
disper sal for many spider mite s (FIe schner, .!:! al., 1956; and Boyle,
1957). Likewise in this study, wind was demonstrated as an impor ..
tant disseminating agent of ~ phoenicis. Several microscope slides
coated with "Tanglefoot" and strips of fly paper were placed at vary
ing distances from a papaya plant heavily infested with B o phoenicis
and other species of mites. After a 24..hour exposure periodp 123
adults of~ phoenicis and several specimens of Tetranychus
telarius (L.) were trapped on the microscope slides and strips of

65.
fly paper placed directly downwind from the source of infestation.
None was found on those placed upwind from the source of infestation.
The slides and fly papers placed nearest to the source of infestation
contained the greater number of mites. One individual was taken on
a slide placed as far as 50 feet away from the infested papaya plant.
Another common way in which wind may play an important part in
mite disper sal is blowing fallen leaves with mites on them for great
distances. Although B. phoenicis commonly feeds on the stems,
crowded conditions cause many individuals to establish them selves on
the petiole and on the lower surface of leaves along the main veins.
Intensive feeding causes the leaves to drop prematurely and, when
windy, these infested leaves are blown some distances away from the
source plant. Should one of these leaves land fortuitously on or in the
vicinity of a suitable host plant, an incipient mite population can be
started by this means.
Disper sion by migrating from plant to plant by crawling over
non-plant surfaces is unlikely when great distances are involved for
none of the active stages of B. phoenicis are able to live for more
than three days without food. In closely set plants, however, disper
sion by crawling f:rom plant to plant is possible for under conditions
where plants were sheltered from wind and rain, clean plants became
readily infested when placed near infested ones. Also, many mites
were caught in the sticky bands that were wrapped around the bottom

66.
of the tTunks of both infested. and uninfested papaya plants to study the
migration of B. phoenicis. The bands on the uninfested plants were
adjusted to trap immigrating mites and those on the infested for emi
grating mites. Emigration of mites from the infested plants started
when the plants became over-populated and suitable feeding areas
scarce. Immigration onto uninfested plants occurred soon after
emigration started; however, the number of immigrants was con
siderably less than the emigrants. Apparently, many of them died in
the process of finding a suitable host during migration. Plants in
direct contact with the infested plants became infested much sooner
than those somewhat removed from the source of infestation. The
outward migration of mites due to population increase is very evident
on passion fruit for the vines intertwine and form a contiguous mass
of growth. On this plant, B. phoenicia foci of infestation enlarge
concentrically, leaving in their wake a conspicuous feeding damage
characterized by defoliation and vine dieback.

67.
POPULATION STUDIES
Seasonal Abundance
The seasonal fluctuations of the ~ phoenicis population densi
ties were studied over a two-year period at two localities, Manoa and
Kaneohe, Oahu, Hawaii. Backyard papaya plants in their first year
of fruiting and receiving no pesticidal treatment were used for follow
ing the monthly fluctuations of the mite populations. The monthly
samples consisted of 40 imprints made with adhesive tapes taken from
10 plants at each locality. Each imprint consisted of an area 6.25
square centimeters of the trunk. The imprints were taken from four
sides of the trunk and between six to ten inches below the attacbm.ent
of the bottom whorl of leaves for here is where the B. phoenicis
populations are usually concentrated.
All stages of ~ phoenicis were present at every sampling date
throughout the two-year period but their numbers varied from month
to month at the two localities. All stages of the mite were numerous
from June to October and scarce from December to May, and were
more abundant in 1962 than in 1963. The monthly densities and fluc
tuations in the egg and adult populations which were very sim.ilar at
the two localities during the two-year period are shown in Figure 17.
The nUIIlber of individuals, especially the active mites, decreased
suddenly following a heavy rain and increased gradually with the
return of warm and hurm.d conditions. Rain not only ha:m.pered the

68.
III
oII,
POPULAnON
MANOA
0"0I "'0: '0I \I \I \
9 bI \I ,I
1963
KANEOHE
(/)
~200::)o()
0"0"""I V'V'\R J' '\ ./EGG POPULATION
I b-u \~I \I 0I \o \
\\
q P, 0-\ I \'0,\ "',0 ~o b'O
J F M A M J J A SO N OJ F M A M J J A S 0 r'f 0
1962 ' 1963
Figure 17. Egg and adult densities and fluctuations of Brevipalpusphoenicis (Geijskes) populations during 1962 and 1963at Manoa (upper) and Kaneohe (lower). Oahu, Hawaii.

69.
movement of the active mites but washed off many of them from the
plant surface; however, it had little effect on the eggs and the inactive
mites which are more firmly attached to the plant surface. In areas
and during periods of adver se climatic conditions, many of the mite s
were observed feeding on the sheltered sides of sterns, leaves, and
fruits away from direct exposure to sun, rain, and wind.
The number of generations produced at Kaneohe and Manoa
during 1962 and 1963 was impossible to determine because of the great
overlap of the different generations caused by continuous reproduction
throughout the year. However, based on the information obtained
froIn the life history studies conducted in the laboratory, at least 10
generations per year were probably produced in the plots during the
two-year period of study.
We station
B. phoenicis infestations varied between the different kinds of
plants, localities, and seasons. Infestations on papaya were found
at all of the locations on Oahu from. sea level to about 1,000 feet
elevation where this crop is grown extensively. Out of the 350 plants
examined during 1963 at different months of the year, 253 of them
were infested by B. phoenicis. All stages of the mite were present
throughout the year at all of the localities. Older papaya plants of
fruit-bearing age were more heavily infested than younger plants of

700
pre-fruiting age. One of the plants in its f~rst year of fruiting had
3,094 !!:. :phoenicis in all stages of developm.ent within an area of
6.25 square centimeters on the trunk surface, with lesser numbers
on the leaf petioles and fruits. On the fruits, ~ phoenicis infesta-
tions were usually concentrated on the side of the fruit that was in
direct contact with the infested trunk or with another infested fruit.
Those fruits located lower on the trunk were m.ore heavily infested
than the younger fruits and blossom.s closer to the apex of the trunk.
None of the other species of plants studied were as heavily in-
fested by B. phoenicis as papaya. The highest num.ber of l!. phoeni-
cis counted within an area of 6. 25 square centitneter s of stem.- '
surface was 181 on passion fruit, 59 on lem.on, 50 on anthurium., 45
on hem.igraphis, and 8 on hibiscus. On other host plants listed in
Table ill, large am.ounts of plant m.aterial had to be exam.ined before
all stages of~ phoenicis were encountered. Infestations by this
species of m.ite were not found on any of the above-mentioned host
plants when found growing in areas above 2,500 feet elevation where
rainfall is ~bundant and temperature drops below 200 C for many days
out of the year.

71.
NATURAL CONTROL FACTORS
There are many factors or conditions, both biotic and abiotic,
which influence the population density of B. phoenicis in a given area.
Abundance of host plants, warm. and humid climate, and paucity of
natural enemies are some conditions which favor high population
density of this mite, whereas conditions contrary to these tend to
depress it and favor the natural control of the pest. In Hawaii, B.
phoenicis is very abundant in areas between sea level and 1, 000 feet,
scarce between I, 000 and 2, 500 feet, and has never been recovered
from areas above 2, 500 feet in elevation. Since host plants are
abundant in all of the areas, this factor is not the cause of the differ
ences in population density. However, the laboratory studies
revealed that temperature does greatly affect the reproductive and
survival potential, and this is probably the main factor which causes
B. phoenicis to be of no econom.ic importance in areas above 1,000
feet elevation. Here, temperatures drop periodically to a level where
mortality is considerably greater than natality and completion of a
life cycle is not possible.
Temperature conditions along the coastal areas in Hawaii are
favorable for continuous reproduction throughout the year at nearly
maximum reproductive and survival potential; however s the popula..
tion density of B. phoenicis has never been observed in any locality
to reach the point where food and space become limiting due to

72.
intraspecific competition. Food and space do not become lim.iting
over a whole orchard or locality but can become lim.iting on indivi
dual plants due to both intra- and interspecific competition. Papaya,
for example, is attacked by at least seven species of phytophagous
mites other than the several species of insects which are of minor
importance (Holdaway, 1941). The mites are: Tetranychus telarius
(L.), Eutetranychus banksi (McGregor), Panonychus~ (McGregor),
Tuckerella pavoniform~s (Ewing), Tuckerella ornata (Tucker),
Hemitarsonemus latus (Banks), and B. phoenicis. !. telarius,
!:h latus, and B. phoenicis are the major pe sts of papaya. Individuals
of these three species have been observed to coexist, increase to
tremendous numbers, and to severely damage individual plants to the
extent that the food media were no longer suitable for the members of
their own species as well as those of the other species. Under such
intense conditions of intra- and inter specific competition, many
individuals of B. phoencis die from starvation and predation by ants,
mites, spiders, and other predators which are fairly common in
Hawaiian soils in the process of migration to other host plants, es
pecially when great distances are involved. Even though some
individuals of B. phoenicis are successful in reaching a host plant,
it may be already occupied by some other organisms which compete
with this species of mite for the same food and space, or the new host

73.
plant may not be as conducive for population increase as papaya. The
latter is evidenced by the considerably lower infestation indices of
the mite on different kinds of plants mentioned earlier and by the
lower fecundity data reported by authors who reared ~ phoenicis on
plants other than papaya (Planes, 1954; Baptist and Ranaweere, 1955;
Dosse, 1957; and Moutia, 1958).
Within the favorable ranges of temperature and humidity, one of
the most conspicuous natural factors that keeps B. phoenicis from
reaching epidemic proportions is natural enemies. Murna (1958) noted
a strong negative correlation between the summer-fall infestations of
!h phoenicis and a complex of Typhlodromus and Arnblysiopsis on
citrus in Florida. Although actual predation was not observed, he
considered these phytoseiid mites to be the principal cause for the
reduction in infestations of B. phoenicis which undergo cycles of high
and low intensity in Florida. In Hawaii, at least four species of pre-
dators, three mites and an insect, have been observed feeding on
B. phoenicis. These are: Phytoseiulus macropilis (Banks)
(Me so stigrnata: Phytoseiidae), Arnblyseius largoensis (Murna)
(Mesostigmata: Phytoseiidae), Mexecheles hawaiiensis (Baker)
(Pro stigmata: Cheyletidae), and Sticholotis punetata Crotch (Coleop-
tera: Coccinellidae).
P. macropilis was first reported from Hawaii in 1953 bv- - . .Cunliffe and Baker while this is the fir st report of A. largoensis for

74.
Hawaii. Besides Hawaii, the former species occurs in Florida,
California, Canary Islands, Puerto Rico, and Panama (Cunliffe and
Baker, 1953; and Chant, 1959), and the latter species in Florida,
Guatemala, Mexico, and Japan (Chant, 1959; and Ehara, 1959). Both
species of phytoseiids usually feed on spider mites, but when these
preferred sources of food become scarce, they feed on other mites.
Adults of P. m.acropilis and A. largoensis were seen on several
occasions feeding on eggs of B. phoenicis on the petioles, fruits, and
stems of papaya. After making contact with an egg, it took an adult of
P. macropilis two minutes to pierce the chorion and withdraw the
internal contents of the egg. Although P. macropilis and A. largo
ensis feed on B. phoenicis eggs readily, there were no indications that
they are able to live and reproduce exclusively on them, for eggs and
larvae of these predators which occur commonly in association with
spider mite populations were never found within the colonie s of
B. phoenicis.
M. hawaiiensis was originally described from Hawaii based on
specimens collected in 1941 from papaya fruits at Kailua, Oahu
(Baker, 1949). Since then, this species has been collected on other
kinds of plants infested by B. :phoenicis from the islands of Maui,
Kauai, and Oahu. Also, this predatory mite has been reported from
Florida recently (DeLeon, 1962; and Muma, 1964). The larvae,
nymphs, and adults of M.hawaiiensis were seen feeding on all of the

75.
active stages of B. phoeniCis, and all stages of this predator were
found in close association with their prey. Individuals of M. hawaii
ensis were seldom seen out in the open but were seen hiding in
crevices, beneath leaves, between fruits, and in other protected
places fr'om where they ambushed their prey. As a prey approached
one of these hiding places, the predator dashed out, grabbed hold of
one of the prey's appendages with its highly modified pedipalpi, and
carried the victim back to its hiding place for feeding. A venomous
substance was probably injected into the prey for it became paralyzed
simultaneous to being captured by.M.• hawaiiensis. The chelicerae
of the predator were inserted into one of the legs of the prey from
where in less than 10 minutes the body contents were completely with
drawn, leaving only the exo skeleton unconSUn1ed. The remains were
carried and disposed outside the hiding place of the predator. The
few numbers of carcasses of B. phoeniCis left accumulating around
each hiding place suggest that M. hawaiiensis does not require much
food for sustenance. Out of a total of the 535 predators found in close
assoCiation with B. phoeniCis populations during 1962 and 1963, 467, or
87 per cent, of them, were M. hawaiiensis.
~ punetata, a tiny cocCinellid, was seen attacking~ phoenicis
for the first time in Hawaii in June, 1964. This predator was intro
duced into Hawaii from China and Japan as early as 1894-95 to control
scales ~Fullaway, 1920). ~ punctata was not seen preying on

76.
B. phoenicis until this late date probably because of its nocturnal
feeding habit. Some 30 adults were seen feeding on B. phoenicis in
festing papaya plants on the University of Hawaii Campus, Honolulu,
Hawaii, during four nights of observation. They appeared at dusk
from within the ground litter at the bases of the papaya plants and
climbed up the trunks to feed. When artifical light was directed on
them, they stopped feeding and moved away from the lighted area.
They finished feeding and left the plants before midnight. No indivi
duals of §.. punctata were seen feeding on B. phoenicis during the day.
The population density of B. phoenicis on these papaya plants dropped
to a low level two weeks after §.. punctata adults were first seen in
close association with the mites. This sudden reduction of the B.
phoenicis population was undoubtedly caused by~ punctata for
dissections of some of the adults caught before returning into the
ground litter showed fragments of m.any individuals and all stages of
~ phoenicis in their digestive tracts.
Enzootics or epizootics caused by pathogenic organisms, such as
fungi, bacteria, and viruses, were not encountered in populations of
B .. phoenicis.. However, two species of fungi, Hirsutella besseyi
Fisher and H. thompsonii Fisher, were found infecting other species of
phytophagous mites ..
All of the above-mentioned predators have been observed to bring
under control isolated populations of~. phoenicia; however, they are

77.
inadequate as economic control agents because their predatory
activity became apparent only when the prey population density was
very high and severe plant damage had already been inflictedo There
fore, in areas where economic control of B. phoenicia is needed,
other control agents must be introduced.

78.
Chemical Control
In recent years, many chemicals with acaricidal properties have
becom.e available but only a few of these can be used to control B.
phoenicis on papaya because of various limitations and restrictions.
Some of the pesticides which have been proven effective for the control
of the comm.on species of spider mites (Family: Tetranychidae) have
been shown to be ineffective against some species of false spider mites
(Family: Tenuipalpidae) (Pritchard, 1949; Hamilton, 1953; and
Morishita, 1954). Many more pesticides of the chlorinated hydrocarbon
than organophosphor'ous derivatives have been reported to give excellent
kill of false spider mites (Pritchard, 1949; Hamilton, 1953; and
Morishita, 1954). Some of the pesticides toxic to the false spider mites
cannot be used on certain kinds of plants because of their phytotoxic
propensity. Papaya, a cornmon host of B. phoenicis, was found to be
very susceptible to phytotoxic injuries when treated with most of the
organic pesticides which were used for the control of mites on other
crops prior to 1959 (Sherman and Tamashiro, 1959). Of the organo
phosphorous pesticides, only Diazinon has been reported to give
excellent kill of a species of false spider mites, B. obovatus (Mori
shita, 1954), but it was found to be extremely phytotoxic to papaya.
Malathion is innocuous to papaya but it proved to be ineffective against
B. phoenicis.

79.
Until recently, sulfur was the only pesticide known to be effec
tive against B. phoenicis and also fairly safe to use on papaya from.
the standpoint of phytotoxicity under m.o st of the clim.atic conditions
which prevail in Hawaii. Since large am.ounts of sulfur, five to six
pounds of 95 per cent wettable powder, are required to give good con..
trol of som.e of the mites infesting papaya, m.any of the growers are
reluctant to apply this pesticide on fruit ...bearing plants because it
leaves unsightly deposits on the fruits. Furthermore, search for other
acaricides becam.e necessary because many species of m.ites coexist
on papaya and one of the most pestiferous of them, '!.. telarius, cannot
be controlled effectively with sulfur (Sherman and Tam.ashiro, 1959).
Those pesticides which have been reported to be fairly innocuous
to papaya (Sherman and Tam.ashiro, 1959), and those which gave negli..
gible injuries to fruit-bearing plants in preliminary studies, were
tested for their effectiveness against B. phoenicis. Of the pesticides
in which both wettable powder and emulsifiable formulations are known,
the former was selected because this form of most pesticides has been
shown to be less phytotoxic to papaya than the corresponding emulsi
fiable form (Sherman and Tam.ashiro, 1959). The performance re sults
of the different pesticides used are shown in Tables VI, VII, and Vill.
Karathane, Diazinon, and aram.ite gave good control of B .. phoenicis on
other plants but were not used in this study for they were found to be
very phytotoxic to papaya.

80.
All concentrations of Kelthane, Pentac, and Morestan and
certain of the higher concentrations of Chlorobenzilate, sulfur, and
ovex gave good kill of !h. phoenicis adults (Table VI), but most of
thero showed poor ovicidal properties (Table Vill). Morestan was the
only pe sticide which caused high mortality of both the adult and egg
stages. Malathion, at all concentrations, caused no significant kill of
B. phoenicis females, but the lower concentrations of this pesticide
appeared to have induced oviposition in the treated feroales (Table VII).
B. phoenicis adults died within 2.4 hours after applying lethal
dosages of Kelthane and Chlorobenzilate, whereas those treated with
lethal dosages of Pentac, Morestan, ovex, and sulfur died four to
seven days after treatroent. Although the females Lived for several
days after treatroent with these pesticides, they fed very little, laid
only few eggs (Table Vil), and remained lethargic most of the tim.e.
Adults treated with Kelthane and Chlorobenzilate died with their legs
fully extended and their body posture posed like that of living indivi
duals while feeding or when at rest so, until probed, the freshly killed
individuals were difficult to distinguish from the living ones. Those
killed by the other pesticides were readily distinguishable from the
Living ones for they died with their front two pairs of legs retracted
beneath the venter of the body.
Many of the eggs that were deposited on the surfaces of the rn.edia
just prior to and after expo sure to treatroent with all concentrations of

81.
Kelthane, Pentac, ovex, and sulfur, and with concentra.tions less than
one pound active material of Chlorobenzilate per 100 gallons of water,
hatched 10 to 12 days later and the resulting larvae fed and continued
to develop on the treated media without showing ill effects from the
residues. Apparently, these pesticides lost their toxic properties
within 10 days under laboratory conditions. If this is true, they un
doubtedly deteriorate to non-toxic levels much faster under field con
ditions. When using one of these pesticides with poor ovicidal and
seemingly short residual properties, a second application within two
to three weeks after the first becomes necessary in order to control
B. phoenicis effectively for it can escape control in the egg stage
which has a fairly long incubation period.
Although effective against B. phoenicis, some of the above
mentioned pesticides and their concentrations should not be applied
to fruit-bearing papaya plants because of phytotoxicity. Oozing of
latex from the epidermis of green fruits in areas where large amounts
of pe sticide g had accumulated were noticed soon after applying ovex at
two pounds, Chlorobenzilate at one and two pounds, and Pentac at
two pounds of active material per 100 gallons of water under field
conditions. These areas of latex exudations developed into scars of
various sizes and shapes in about three weeks after the pestkide
application which greatly marred the appearance of the fruits

.Figure !8~ L1'ljury caus.ed by Ghlol"'obenzilate at 1 poundof active material per 100 gallons of water"
82•
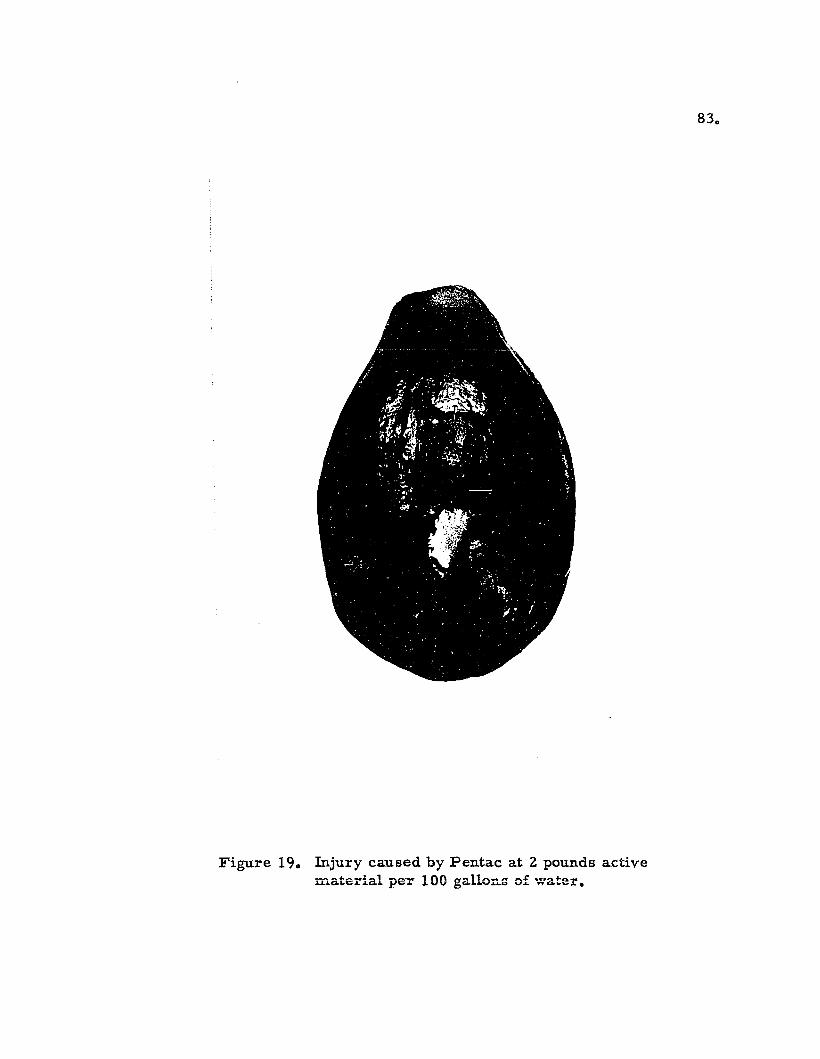
Figure 19. Injury caused by Pentac at Z pounds activematerial per 100 gallong of water.
83.
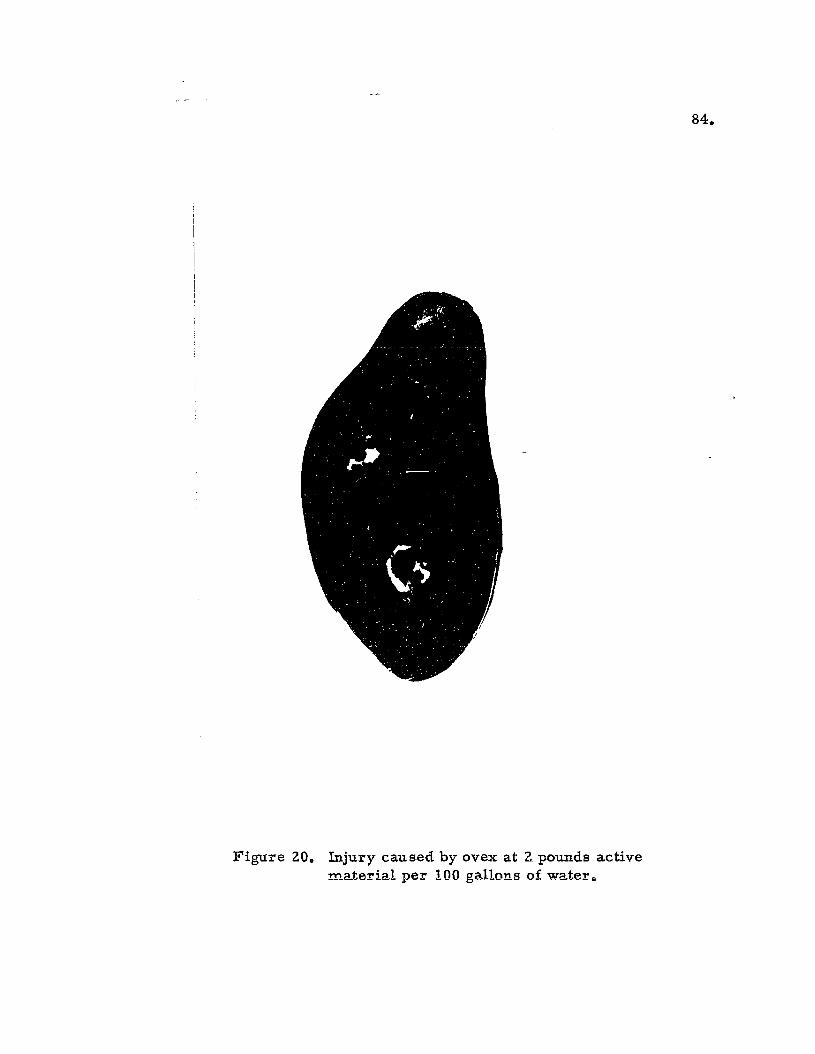
Figure 20. Injury caused by ovex at 2 pounds activematerial per 100 ga.llons of wa.terg
84.

85.
(Figs. 18, 19, and 20). Kelthane and Morestan were found to be
innocuous to papaya fruits at concentrations as high as two pounds of
active material per 100 gallons of water, but they are not recom
mended for use at these high dosages for not only is it uneconomical
but the unsightly depo sits on the frui~s mentioned earlier would be a
problem.
Kelthane, Morestan, Chlorobenzilate, Pentae, and ovex are not
to be used on papaya grown for the commercial market as they have
not yet been cleared for use on this crop. Sulfur and Volck Oil
Supreme are the only two pesticides which are toxic to some of the
stages of B. phoenicis and can be used on papaya for they are exempt
from the requirement of a tolerance. Whereas mo st petroleum oils
and even er.L.ulsifiable forms of many pesticides are known to be phyto
toxic to papaya, Volck Oil Supreme, a highly refined petroleum oil,
caused no obvious phytotoxic injury to fruit-bearing papaya plants.

TABLE VI. PERFORMANCE OF DIFFERENT. PESTICIDES AGAINST THE ADULTS OFBREVIPALPUS PHOENICIS (GEIJSKES)
Pounds activematerial per
Pesticides 100 gallons Formulations No. Alive a/ Statistical significance b/
Malathion 1/4 25% WP 5.10Ovex 1/4 50% WP 5.05Check 5.02Malathion 1/2 250/0 WP 5.02Malathion 1 25% WP 4.90Malathion 2 25%WP 4.70Ovex 1/2 50% WP 4.48 --'I ]Chlorobenzilate 1/4 25%WP 3.92Volck Supreme 3 EC 3.25 :::JVolck Supreme 2 EC 2.55 J]Volck Supreme 6 EC 2.38Volck Supreme 4 EC 2.15Ovex 1 500/0WP 2.10Sulfur 3 95% WP 1. 90 ]Chlorobenzilate 1 25% WP 1.45
lSulfur 4 95% WP 1.40Pentac 1/4 50% WP 1.35Chlorobenzilate 1/2 25% WP 1.25
0:>0'•

TABLE VI. Con.tinued
Pesticides
MorestanMorestanMore stanMorestanChlorobenzilateOvexSulfurSulfurPentacPentacPentacKelthaneKelthaneKelthaneKelthane
Pounds activematerial per
100 gallons
1/41/2
122256
1/212
1/41/2
12
Formulations
250/0 WP25%WP25% WP25%WP25%WP50%WP95% WP95%WP50%WP50% WP50%WP25%WPZ5%WP25% WP25%WP
No. Alive eJ
1.201.101.001.001.001.001.001.001.001.001.001.001.001.001.00
Statistical Significance "'e.l
a/ N III 25. Average of four replications. The data were transformed according to the formula
.jXfT.b/ At 5 per cent level, according to multiple range test of Duncan (1955).
WP =. Wettable powderEC a Emulsifiable concentrate
00-.].

TABLE VII. NUMBER OF EGGS OF BREVIPALPUS PHOENICIS (GEIJSKES) COUNTED ONTREATED SURFACE ONE WEEK AFTER .APPLICATION OF THE DIFFERENTPESTICIDESo
PoUnds activematerial per
Pesticides 100 gallons Formulations No. of eggs a/ Statistical significance b /
Malathion 1/2 250/0 WP 194.75 ::JMalathion 1/4 250/0 WP 138.75Malathion 1 250/0 WP 129.75Check 128.50Ovex 1/2 500/0 WP 116.25Malathion 2 250/0 WP 110.75Ovex 1/4 500/0 WP 105.50Valek Supre:rne 3 EC 68.50 =:1Chlorobenzilate 1/4 250/0 WP 32.50Volck Supreme 6 EC 20.75Ovex 1 500/0 WP 20.50Volek Supreme 2 EC 19.25Sulfur 3 950/0 WP 19.25Pentac 1/4 500/0 WP 16.50Pentac 1/2 500/0 WP 16.25Ovex Z 500/0 WP 13.75Sulfur 4 950/0 WP 13.50Sulfur 5 950/0 WP 12.75
0:>0:>•

TABLE VII., Continued
Pesticides
Pounds activematerial per100 gallons Formulations No. of eggs al Statistical significance bl
Volck SupremeMore stan
SulfurPentacKelthanePentacMorestanMorestanMorestanKelthaneKelthaneChlorobenzilateChlorobenzilateChlorobenzilateKelthane
41/4
61
1/2212
1/21
1/41/2
122
EC25%WP95% WP50%WP25%WP500/0 WP250;0 WP25%WP25%WP25%WP25%WP25%WP250;0 WP25%WP25% WP
11.7511.2510.7510.7510.5010.009.759.508.507.757.507.507.507.506.50
;;j Average of four replications. Pre-treatment N = 25 females.Eggs laid prior to death or during first week afj;er treatment.
r:J At 5 per cent level, according to multiple rang~: test of Duncan (1955).
WP =EC =
Wettable powderEmulsifiable concentrate
00--D.

1'ABLE Vill.. OVICIDAL EFFECT OF DIFFERENT PESTICIDES ON THE EGGS OFBREVIPALPUS PHOENICIS (GEIJSKES).
Pounds activematerial per Per cent of
Pesticides 100 gallons Formulations eggs hatched al Statistical. significance b/
Check 87.0 ]Malathion 2 25% WP 84.0Sulfur 6 95% WP 80.5Keltha.ne 2 25% WP 69.5 ]Ovex 2 50% WP 68.0Pentac 2 50%WP 67.5Chlorobenzilate 2 25% WP 55.5 :::JVolck Supreme 6 EC 14.0 ::JMorestan 1/4 25% WP 2.5 ]Morestan 2 25% WP 1.5
a/ Average of eight replications. N· 25 eggs per replication..
bl At 5 per cent level, according to multiple range test of Duncan (1955).
WP = Wettable powder formulationEC ~ Emulsifiable concentrate
-.0o.

91.
DISCUSSION
Studies on the biology of B. phoenicis have revealed that this
species has many attributes in common with the other well-known
false spider mites, 1b obovatus andlh californicus. Like them, B.
phoenicis is polyphagous, reproduces mainly by parthenogenesis,
males are very scarce, life cycle is fairly long, and it is tolerant of._-,-- .
certain of the organophosphorous pesticides. Despite these similari-
ties, B. phoenicis possesses certain attributes which seem to be dis-
tinctive for the species. The most striking attribute is its ability to
thrive only within fairly narrow temperature range, while B. obovatus
and !h californicus reproduce freely within a wider temperature
range, 20°C and 30o C, with very little mortality occurring during the
developmental stadia under laboratory conditions (Manglitz and Cory,
1953; and Morishita, 1954). The distributional patterns of these false
spider mites in nature are in line with the laboratory findings. Here
in Hawaii, B. phoenicis is very abundant only along the coastal areas
to about 1,000 feet elevation where the temperature is fairly uniform
throughout the year and does not deviate too greatly from 2.50 C for any
length of tim.e, whereas the other two false spider mites are more
widely distributed from sea level to about 3,500 feet elevation where
the ten'lperature conditions at the higher elevations are m.ore variable
and often drops below 2.0oC for extended durations (Ripperton and
Hosaka, 1942.). Also, both B. obovatus and B. californicus have been

92,
reported from more countries outside the tropics tha,n B. phoenicis
(Pritchard and Baker, 1958).
Another distinguishing attribute of B. phoenicis is its feeding
preference on the stems of plants. The foci of B. phoenicis infesta-
tions are usually located on the stems while those of B. obovatus and
~. californicus are invariably located on the lower surfaces of leaves.
These mites move away from their preferred feeding sites to other
parts of the plant only when overcrowded conditions prevail.
Although the life cycles of the three false spider mites were,
carried out under slightly different laboratory conditions by the vari-
ous workers (Manglitz and Cory, 1953; and Morishita, 1954), all
indications are that the life cycle of B. phoenicis is definitely longer
than those of B. obovatus and B. californicus. B. phoenicis com-
pleted its development from the egg to the adult stage in 18.6 f:. 0.5
days at 30oC, 29.3 I:. 0.4 days at 250 C, and 48.9 /:. 1.5 days at 20o C.
At the sam.e respective temperatures, the average developmental
periods of B. obovatus were reported as 16.4, 28.0, and 42.0 days
by Morishita (1954)0 Dosse (1959) found that B. phoenicis required
about two days longer than~ obovatus to complete its development
on ivy leaves at 24oC. Of the three species of false spider mites,
B. californicus seems to have the shortest life cycle for }/ianglitz and
Cory (1953) were able to rear this mite from egg to the adult stage
in 26.2 days under temperature conditions ranging between 17. 90C

and 23. 9°C. Whereas the developmental rate of false spider mites
is much faster at temperatures above 250 C than below it, the life
cycle of B. califomicus is two to three days shorter at temperatures
below 250 C than those of B. obovatus and B. phoenicis at 250 C.
The developm.ental rate of the above three specie s of false
spider mites is slow in comparison to that of som.e of the spider
m.ites. Tetranychus desertorurn Banks and T. telarius (L.) are
known to develop from. eggs to adults in just 6.90 t. 0 .. 5 days and
7.09 t 0.17 days, respectively, at 300 C (Nickel, 1960), while the
false spider mites, B. phoenicis and B. obovatus, require on an
average of at least 18.6 and 16.4 days, respectively, at the same
temperature conditions. The conditions under which developm.ent is
m.ost rapid is not necessarily favorable for population increase since
m.ortality during the im.m.ature stage s can be much greater than
natality as was shown in this study. Therefore, when optimum. survi
val conditions are taken into consideration, the developmental period
of B. phoenicis is about four times longer than those figures shown
above for the spider m.ite s.
The fecundity of both ~ phoenicis and B. obovatus is less than
that of som.e spider mites. Tetranychus atlanticus McGregor, T.
telarius, and T. desertorum females are capable of laying as many
as 230, 194, and 187 eggs per individual, respectively (Cagle, 1949
and 1956; and Nickel, 1960); however, the maximum laid by any

94..
species of false spider mite female is only 76 eggs. This low
fecundity and the long developmental period of the false spider mite s
are probably the main reasons why they are only occasional pests of
perennial plants and are not as serious pests of agricultural crops as
the spider mites. In Ceylon, B. phoe~ is reported to require
about three year s to reach economi c proportions on tea plants which
are pruned (Baptist and Ranaweera, 1955). Here in Hawaii, B.
phoenicis normally requires about a year to build up to densitie s high
. enough to cause noticeable damage to papaya.
Since B. phoenicis reproduces mainly by parthenogenesis, it
seems reaso:n.able to assume that the genetic composition of a popu-
lation of this mite within an area is fairly uniform. Resistance to
pesticides is less likely to be selected out from such a population
because the gene pool is more limited than that of sexually reproduc
ing species as the spider mites. This is probably the mai.Tt reason
why resistance to acaricides which were once proven effective has
not been reported for the false spider mites, whereas resistance to
pesticides is a notorious and serious problem in the case of spider
mites (Jeppson, 1961)0 However, the frequency of application of
selection pressure such as a pesticide should be taken into consider
ation. Since the false spider mites are only occasional pests of
agricultural crops, none of the acaricides have been used extensive
ly for their control as for spider mites and this also may be a reason

95.
why resistant populations have not yet been selected out.
Although. chemical control is the most effective means of sup
pressing outbreaks of B. phoenicis, :many problems can arise in the
atte:mpts to control this mite with che:micals. When chernical control
is needed on a food crop, one :must rnake certain that the pesticide
selected for application is approved and recom.:mended for usage on
the particular crop and is used in strict accordance with the directions
stipulated on the labels of the pesticide containers. On certain crops,
only a few pesticides are cleared for use and these registered pesti
cides may not be toxic to B. phoenicis even though they have been
shown to be effective against other closely related species or they may
be toxic to only certain stages of B. phoenicis. Here in Hawaii, when
using pesticides which kill only certain stages of the mite, several
applications within relatively short intervals are necessary for all
stages of~ phoenicia are present simultaneously the year around
and the duration of certain of the stages is longer than the residual life
of rnost pesticides. Cornbinations of pesticides, one that kills the egg
with another that kills the active stages, seem like a ready solution
to the problem of necessity of several and/or separate applications,
but other problerns can arise in doing so. Sorne pesticides are
innocuous to plants and very effective against certain stages of B.
phoenicis when applied singly, but when co:mbined they :may becorne
either phytotoxic due to synergism or rnay become ineffective against

96.
the rnites. Furthennore, not all pesticides are cornpatible with each
other.
Some pesticides are very effective against B. phoenicis but they
cannot be used because certain of the host plants of this mite are very
susceptible to chemical injury. The reader is referred to the papers
by Gast and Early P956) and Sherman and Tamashiro (.1959), and to
the textbooks by Bailey and Smith (1951), Brown (1951), and Shepard
(1951) for concepts on the phytotoxic properties of pesticides..

97.
SUMMARY
The red and black flat mite, Brevipalpus phoenicis (Geij skes),
is widely distributed throughout the tropical areas of the world. In
Hawaii, this mite is very abundant on all of the major islands along
the coastal areas up to about 1,000 feet elevation. The climatic
conditions in these areas are favorable for continuous reproduction
of B. phoenicis throughout the year.
B. phoenicis inhabits and feeds on approximately 100 different
kinds of plants. It is a major pest of perennial plants such as papaya,
pas-sion fruit, tea, citrus, and several species of ornamentals. It
usually feeds on the sterns of plants, but when food and space become
limiting due to overcrowded conditions, some individuals move to
other parts of the plant and form. colonies.
Morphologically, B. phoenicis is a variable species. For this
reason, several synonyms have been created. The most variable
structures are the dorsolateral hysterosomal setae I and II, and the
dor socentral hystero somal setae III of the larvae and nymphs. The Be
vary from a tiny~ serrate setae to a large, broadly lanceolate,
serrate setae. The larva, protonyrnph, and deutonyrnph of B o
phoenicis are ve:r:-y similar to those of ~o obovatus Donnadieu» but the
adults are readily distinguished from the latter specieso B o phoenicis
is the only species known to have two pairs of sensory rods on tarsus
II and five pairs of dorsolateral hysterosomal setae in the adult stage.

98.
The life cycle of B. phoenicis is greatly influenced by tempera
ture and hum.icIity conditions. The larva, protonymph, and deuto
nymph spend about one-half of their stadia as active individuals and
the remainder of the time as inactive individuals. Feeding, growth,
and limited dispersion are accomplished during the active phase and
ecdysis occurs during the inactive phase. The average durations of
the different stages range from. 8.2 1:. 0.4 days at 300 e to 22.6 I:. 1.1
days at 200 e in the egg stage, 3.6 1:. 0.2 days at 30°C to 10.5 t. 0.7
days at 200 e in the larval stage, 3.1 I:. 0.4 days at 300 e to 8.2 t 0.7
days at 200 e in the protonymphal stage, and 3.7 t 0.5 days at 300 e
to 7.5 f:. 0.9 days at 200 e in the deutonymphal stage. At 250 e and 65
to 70 per cent relative humidity, conditions most favorable for survi
val and population increase, B. phoenicis completes development
from the egg to the adult stage in 29.3 f:. 0.4 days. Development is
much faster at temperatures above 250 e than below it; however,
mortality of the im.m.ature stages increases with deviation of tempera
ture away from 250 e until completion of the life cycle is no longer
possible at temperatures above 300 e and below 20oe. All stages of
the mite are able to withstand brief exposures to temperatures below
200 e better than temperatures above 30oe.
The B. phoenicis adult population is predominantly female.
Males have been reported as non-existent in many parts of the world,
but here in Hawaii they make up about 1 per cent of the adult population.

99.
Since umnated females are able to give rise to exclusively female
progenies and males are very scarce, reproduction of the species
is mainly by parthenogenesis.
At 250 C and 65 to 70 per cent relative humidity, B. phoenicis
females start to oviposit in 3.5 1:. 0.9 days after emergence and lay
from one to four eggs per day for a total of 57.5 1:. 7.4 eggs per
female during their life span of 34.8 1:. 4.5 days. Preoviposition
period is longer at 300 C than at 250 C or 20o C. Females live longer
but lay only one-fifth as many eggs at 200 C than at 250 C.
Four species of predators; Phytoseiulus macropilis (Banks),
Amblyseius largoensis (Muma), Mexecheles hawaiiensis (.Baker), and
Sticholotis punctata Crotch, feed on certain stages of B. phoenicis in
Hawaii. Of these, M. hawaiiensis is the predominant predator. All
of the active stages of this cheyletid mite feed on the active stages of
B. phoenicis. These natural enemies control isolated populations of
~ phoenicis, but are often inadequate as economic control agents.
Chemical control measures can be used to effectively depress
outbreaks of B. phoenicis. Morestan is toxic to all stages of the
mite, whereas Chlorobenzilate, Pentac, Kelthane, and sulfur give
excellent kill of the active stages only. Volck Supreme, a highly re
fined petroleum oil, gives good kill of the eggs of B. phoenicis but
only poor to fair kill of the adults. These pesticides are relatively
non-phytotoxic to papaya, a plant that is very susceptible to chemical

100.
injury, at the m.inim.um dosage of effectiveness to B. phoenicia.
Sulfur and Volck Oil Supreme are the only two pesticides which can
be used to control &. phoenicis on papaya grown for the com.m.ercial
m.arket, while the other above-m.entioned pesticides are not yet
registered for use on this crop. All stages of B. phoenicis are
tolerant of m.alathion, an organophosphorous pesticide which is
approved for usage on papaya and claimed to kill certain species of
spider m.ites.

101.
REFERENCES CITED
Attiah., H. H. 1956. The genus Brevipalpus in Egypt.Soc. Ent. dlEgypte Bull. 40: 433-448.
Bailey, S.. F. and L. M. Smith. 1951. Handbook of agriculturalpest control. 191 pp. Industry Publications, Inc., N. Y.
Baker, E. W. 1945. Mites of the genus Tenuipalpus (Acarina:Trichadenidae). Proc. Ent. Soc. Wash. 47(2): 33-38.
__--:~_':""'. 1949. The genus Brevipalpus (Acarina:Pseudoleptidae). Amer. Midi. Na.t. 42(2): 350-402•
.and A. E. Pritchard. 1960. The tetranychoid------mites of Africa. Hilgardia 29(11): 455-574.
Baptist, B. A. and D. J. W. Ranaweera. 1955. The scarlet mitesof the genus Brevipalpus as pest of tea in Ceylon. TeaQuart. 26(4): 127-137.
Blanchard, E. E. 1939. Tres acaros daninos para los argentinos.Rev. Fac. Agron. La Plata. 24(3): 11-18.
Boudreaux, H. B. 1958. The effect of relative humidity on egglaying, hatching, and survival in various spider mites.Jour. Inst. Physiol. 2(1): 65-72.
Boyle, W co W. 1957. On the mode of dissemination of the twospotted spider mite, Tetranychus telarius (L.) (Acarina:Tetranychidae). Proc. Hawaii. Ent. Soc. 16(2): 261-268.
Brown, A. W. A. 1951. Insect control1?zche:micals co vii I817 pp. John Wiley &: Sons, Inc•• N. Y.
Cagle, L. R. 19490 Life history of the two-spotted spider mite.Virginia Agric. Expt. StaB Tech. Bull. 113: 1-31.
___~__• 1956. Life history of the spider mite Tetranychusatlanticus McG. Virginia Agric. Expt. Sta. Tech. Bull.124: 1-22.

102.
Carter, W. 1952. Injuries to plants caused by insect toxins. II.Bot. Rev. 18(10}: 680-721.
Chant, D. A o 1959. Phytoseiid mites (Acarina: Phytoseiidae)Part I. Bionomics of seven species in Southeastern England.Part II. A taxonomic review of the family Phytoseiidae,with descriptions of 38 new species. Canada Ent. 91(Supple 12): 1-166.
Chilson, L. M. 1959. Notes and exhibitions. False spider mitesin Hawaii. Proc. Hawaii. Ent. Soc. 17(1): 20~21.
Cornm.onwealth Institute of Entomology. 1959. Distribution mapsof insect pests. Series A. Map 106. 56 Queen's Gate,London, S. W.
Cromroy, H. L. 1958 0 A preliminary survey of the plant mites ofPuerto Rico. Jour. Agric:. Univ. Puerto Rico 92(2): 39-144.
Cunliffe, F. and E. W. Baker. 1953. A guide to the predatoryphytoseiid mites of the United States. Pinellas Biol. Labc
Publ. 1: 1-28.
DeLeon: D. 1961. The genus Brevipalpus in Mexico, part II(Acarina: Tenuipalpidae). Florida Ent. 44(1): 41-52.
• 1962. Three new genera and seven new species of---~-cheyletids (Acarina: Cheyletidae). Florida Ent. 45(3):
129-137.
Di Martino, E. 1960. Due nuovi acari dannosi agli agrurni inItalia. Inform. fitopat. 9: 356-358.
Dosse, G. 1957. Vergleichende Untersuchungen an Brevipalpusphoenicis (Geijskes) und Brevipalpus inornatus Banks(Acarina, Phytoptipalpidae). Pflanzenschutz-Ber. 19 (l-9):36-44.
Duncan, Do B o 1955 0 Multiple range and multiple F test.Biometrics 11(1): 1-42.
Ehara, So 1959. Some predatory mites of the genera Typhlodromusand Arnblyseius from Japan (Phytoseiidae). Acarologia 1(3):285-295" ...

103.
Fleschner, C. A., M. E. Badgley, D. W. Richer, and J. C. Hall.1956. Air drift. of spider mites. Jour. Econ. Ent. 49(5):624-627.
Fullaway, D. T. 1920. Natural control of scale insects in Hawaii.Proc. Hawaii. Ent. Soc. 4(2): 237-246.
Gasta R. and J. Early. 1956. Phytotoxicity of solvents andemulsifiers used in insecticide formulations. Agric. Chem..11(4): 42-45, 137, 139.
Geij skes, D. C. 1939. Beitrage zur Kenntnis der europaischenSpinmnilben (Acari, Tetr-anychidae), mit besondererBeriicksichtigung der niederULndischen Arten. Meded.LanbHoogesch. Wageningen 42(4): 1-68.
Hamilton, C. C. 1953. New acaricides against mites attackingnursery plants. Jour. Econ. Ent. 46(3): 442-445.
Holdaway, F. G. 1941. Insect pest of papaya and their control.Part IV. Papaya production in the Hawaiian Islands.Hawaii Agric. Expt. Stag Bull. 87: 45-51.
Jeppson, L. R. 1961. Cross resistance patterns in Acarina.Advances in Acarology 1: 276-282.
Knorr, L. C., B. N. Webster, and G. Malaguti. 19600Injuries in citrus attributed to Brevipalpus mite, includingBrevipalpus gall, a newly reported disorder in sour-orangeseedling. FOA Plant Prot. Bull. 8(12): 141-149.
, E. C. Paterson, and J. H. Proctor. 1961. World----:---'citrus problems. I. Aden Protectorate. FOA Plant Prot.Bull. 9(6): 91-98.
Manglitz, G. R o and E. N .. Cory. 19530 Biology and control ofBrevipalpus australis. Jour. Econ. Ent. 46(1): 166-1190
Marchionatto, J. B. 1935. Argentine Republic: New studies on"Lepra Explosiva" of orange. Into Bull. Plant. Prot. Rome9(8): 173M-175M.

104.
Marchionatto, J. B. 1938. Transmission of the 11 Lepra Explosivall
of the oranges by mites. Int. Bull. Plant Prot. Rome 12(6):121M-122M.
Marlowe, R. H. 1937. Exhibition and discussion of local material.Froe. Hawaii. Ent. Soc. 9(3): 377-378.
Morishita, F. S. 1954.inornatus (Banks).
Biology and control of BrevipalpusJour. Econ. Ent. 47(3): 449-456.
Moutia, L. A. 1958. Contribution to the study of some phytophagousAcarina and their predators in Mauritius. Bull. Ent. Res.49: 59-75.
Muma, M. 1958. Predators and parasites of citrus mites in Florida.Proc. lOth. Inter. Congress Ent. 4: 633-647.
1961. Mites associated with citrus in Florida. Univ.Florida, Agric. Elcpt. Sta. Bull. 640: 1-39.
1964. Cheyletidae (Acarina: Trombidiformes) associatedwith citrus in Florida. Florida Ent. 47(4): 239-253.
Nickel, J. L. 1958. Agricultural insects of the Paraguayan chaco.Jour. Econ. Ent. 51 (5): 633-637.
. 1960. Temperature and humidity relationships of------Tetranychus desertorum Banks with special reference todi stribution. Hilgardia 30 (2) : 4 I - 100 •
Planes, G. S. 1954. la araf!uela roja, nueva plaga de la naranjosen el Levante espanol. Bol. Pat. veg. Ent. Agric.19(1951-52): 189-196.
Pritchard, A. E. 1949. California greenhouse pests and theircontrol. Calif. Agric. Expt. Sta. Bull. 713:' 1-71.
and E. W. Baker. 1952. The false spider mitesof California (Acarina: Phytoptipalpidae). Univ. Calif. Publ.Ent. 9(1): 1-93.

105.
Pritchard, A. E. and E. W. Baker. 1958. The false spider mites(Acarina: Tenuipalpidae). Univ. Calif. Publ. Ent. 14(3):175... 274.
Rirnando, L. G. 1962. The tetranychoid mites of the Philippines.Univ. Philippines, Coll. AgX'ic. Tech. Bull. 11: 1 ...52.
Ripperton, J. C. and E. Y. Ho saka. 1942. Vegetation zone s ofHawaii. Hawaii Agric. Expt. Sta. Bull. 89: 1-60.
Rossetti, V., T. G. Fass, and R. M. Musumeci. 1959. Urn novo.!caro dos laranjais paulistas. Biol~gico 25 (12): 273... 275.
Sanchez, F.F. andM. Sherman. 1963. Toxicityofinsecticidesto Cryptophlebia illepida (Le'pidoptera: Eucosrnidae), a pestof the macadamia nut. Jour .. Econ. Ent. 56(3): 362... 368.
Shepard, H. H. 1951. ~ chemistry~toxicolog~of insecticides.vii t 504 pp. McGraw...Hill Book Co., Inc., N. Y.
Sherman, M. and M. Tamashiro. 1959. Toxicity of insecticides andacaricides to the papaya, Carica papaya L. Hawaii Agric.Expt. Sta. Tech. Bull. 40: 1-56.

![Palaeontological Society of JapanFinance : Hiroshi OzA KI , Saburo KANN O Planning: Hiroshi OZAKI, Hiroshi U]llE Publications Transactions: Tokio Sr-l!KAMA , Kiyotaka Cr-II NZEI, Noriyuki](https://static.fdocuments.in/doc/165x107/60ea53b1e659066680525a43/palaeontological-society-of-finance-hiroshi-oza-ki-saburo-kann-o-planning-hiroshi.jpg)
















