Thermodynamic analysis of substrate induced domain closure of 3-phosphoglycerate kinase
5
Thermodynamic analysis of substrate induced domain closure of 3-phosphoglycerate kinase Andrea Varga a,1 , Judit Szabó a,1 , Beáta Flachner a , Zoltán Gugolya b , Ferenc Vonderviszt a,b , Péter Závodszky a , Mária Vas a, * a Institute of Enzymology, Biological Research Center, Hungarian Academy of Sciences, P.O. Box 7, H-1518 Budapest, Hungary b University of Pannonia, Faculty of Information Technology, Department of Nanotechnology, P.O. Box 158, H-8201 Veszprém, Hungary article info Article history: Received 5 October 2009 Revised 15 October 2009 Accepted 16 October 2009 Available online 23 October 2009 Edited by Peter Brzezinski Keywords: Domain closure Phosphoglycerate kinase Isothermal titration calorimetry Substrate binding Energetic parameters abstract The energetic changes accompanying domain closure of 3-phosphoglycerate kinase, a typical hinge- bending enzyme, were assessed. Calorimetric titrations of the enzyme with each substrate, both in the absence and presence of the other one, provide information not only about the energetics of substrate binding, but of the associated conformational changes, including domain closure. Our results suggest that conformational rearrangements in the hinge generated by binding of both substrates provide the main driving force for domain closure overcoming the slightly unfavourable contact interactions between the domains. Ó 2009 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. 1. Introduction Biological recognition and the associated allosteric conforma- tional changes are essential for the function and regulation of liv- ing systems (cf. recent review [1]). Interdomain communications operating in multidomain proteins/enzymes usually represent examples of sophisticated mechanisms, which are rarely described in detail. Previously, we have described a detailed molecular mechanism of domain movement of a typical two-domain enzyme, 3-phospho- glycerate kinase (PGK) [2]. PGK catalyzes the phospho-transfer from 1,3-bisphosphoglycerate (1,3-BPG) to MgADP producing 3- phosphoglycerate (3-PG) and MgATP. One of the two substrates, 3-PG [3] or 1,3-BPG binds to the N-domain, while the nucleotide substrate, MgATP [4] or MgADP [5] binds to the C-domain. By com- parative graphical analysis of various crystal structures [2,6] and by mutagenesis studies [7] we could identify the main hinge of PGK molecule at b-strand L in the interdomain region that essen- tially determines the relative domain positions. An extension of an H-bond network upon simultaneous binding of both substrates was found that possibly regulates the shape of the main hinge of b- strand L and thereby directs closure of the two domains during the catalytic cycle of PGK. It is generally accepted that the relative domain movements are essential for functioning of multidomain proteins [8]. Domain clo- sure can often create an optimal environment for catalysis, orients properly the substrate reactive groups and/or impedes side reac- tions. Typical examples of such mechanism are the kinases catalyz- ing transfer of a phospho-group from one substrate to another [9]. In order to better understand the event of domain closure, the thermodynamic (energetic) aspects of the motion are equally important. Isothermal titration calorimetry (ITC) is one of the most adequate methods for quantitative measurement of the free energy changes that occur upon ligand/substrate binding and during the accompanied conformational transition [10]. There are only a few examples in the literature where energetics of domain reorienta- tion is estimated [11–13]. In this work we present ITC ligand binding studies with human PGK (hPGK). Separate binding of either of the two substrates to PGK, although causes well-defined changes in the hinge-region [3,5,14], does not cause substantial domain movement, in agree- ment with both crystallographic [3–5] and small angle X-ray 0014-5793/$36.00 Ó 2009 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2009.10.048 Abbreviations: AMP-PNP, b,c-imido-adenosine-5 0 triphosphate; 1,3-BPG, 1,3- bisphosphoglycerate; 3-PG, 3-phosphoglycerate; PGK, 3-phospho-D-glycerate kinase or ATP, 3-phospho-D-glycerate 1-phosphotransferase (EC 2.7.2.3); hPGK, human PGK; ITC, isothermal titration calorimetry; SAXS, small angle X-ray scattering * Corresponding author. Fax: +36 1 466 5465. E-mail address: [email protected] (M. Vas). 1 These authors contributed equally to the work. FEBS Letters 583 (2009) 3660–3664 journal homepage: www.FEBSLetters.org
-
Upload
andrea-varga -
Category
Documents
-
view
212 -
download
0
Transcript of Thermodynamic analysis of substrate induced domain closure of 3-phosphoglycerate kinase

FEBS Letters 583 (2009) 3660–3664
journal homepage: www.FEBSLetters .org
Thermodynamic analysis of substrate induced domain closureof 3-phosphoglycerate kinase
Andrea Varga a,1, Judit Szabó a,1, Beáta Flachner a, Zoltán Gugolya b, Ferenc Vonderviszt a,b,Péter Závodszky a, Mária Vas a,*
a Institute of Enzymology, Biological Research Center, Hungarian Academy of Sciences, P.O. Box 7, H-1518 Budapest, Hungaryb University of Pannonia, Faculty of Information Technology, Department of Nanotechnology, P.O. Box 158, H-8201 Veszprém, Hungary
a r t i c l e i n f o
Article history:Received 5 October 2009Revised 15 October 2009Accepted 16 October 2009Available online 23 October 2009
Edited by Peter Brzezinski
Keywords:Domain closurePhosphoglycerate kinaseIsothermal titration calorimetrySubstrate bindingEnergetic parameters
0014-5793/$36.00 � 2009 Federation of European Biodoi:10.1016/j.febslet.2009.10.048
Abbreviations: AMP-PNP, b,c-imido-adenosine-50
bisphosphoglycerate; 3-PG, 3-phosphoglycerate; Pkinase or ATP, 3-phospho-D-glycerate 1-phosphotrahuman PGK; ITC, isothermal titration calorimetryscattering
* Corresponding author. Fax: +36 1 466 5465.E-mail address: [email protected] (M. Vas).
1 These authors contributed equally to the work.
a b s t r a c t
The energetic changes accompanying domain closure of 3-phosphoglycerate kinase, a typical hinge-bending enzyme, were assessed. Calorimetric titrations of the enzyme with each substrate, both inthe absence and presence of the other one, provide information not only about the energetics ofsubstrate binding, but of the associated conformational changes, including domain closure. Ourresults suggest that conformational rearrangements in the hinge generated by binding of bothsubstrates provide the main driving force for domain closure overcoming the slightly unfavourablecontact interactions between the domains.� 2009 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
Biological recognition and the associated allosteric conforma-tional changes are essential for the function and regulation of liv-ing systems (cf. recent review [1]). Interdomain communicationsoperating in multidomain proteins/enzymes usually representexamples of sophisticated mechanisms, which are rarely describedin detail.
Previously, we have described a detailed molecular mechanismof domain movement of a typical two-domain enzyme, 3-phospho-glycerate kinase (PGK) [2]. PGK catalyzes the phospho-transferfrom 1,3-bisphosphoglycerate (1,3-BPG) to MgADP producing 3-phosphoglycerate (3-PG) and MgATP. One of the two substrates,3-PG [3] or 1,3-BPG binds to the N-domain, while the nucleotidesubstrate, MgATP [4] or MgADP [5] binds to the C-domain. By com-parative graphical analysis of various crystal structures [2,6] andby mutagenesis studies [7] we could identify the main hinge of
chemical Societies. Published by E
triphosphate; 1,3-BPG, 1,3-GK, 3-phospho-D-glyceratensferase (EC 2.7.2.3); hPGK,; SAXS, small angle X-ray
PGK molecule at b-strand L in the interdomain region that essen-tially determines the relative domain positions. An extension ofan H-bond network upon simultaneous binding of both substrateswas found that possibly regulates the shape of the main hinge of b-strand L and thereby directs closure of the two domains during thecatalytic cycle of PGK.
It is generally accepted that the relative domain movements areessential for functioning of multidomain proteins [8]. Domain clo-sure can often create an optimal environment for catalysis, orientsproperly the substrate reactive groups and/or impedes side reac-tions. Typical examples of such mechanism are the kinases catalyz-ing transfer of a phospho-group from one substrate to another [9].
In order to better understand the event of domain closure, thethermodynamic (energetic) aspects of the motion are equallyimportant. Isothermal titration calorimetry (ITC) is one of the mostadequate methods for quantitative measurement of the free energychanges that occur upon ligand/substrate binding and during theaccompanied conformational transition [10]. There are only a fewexamples in the literature where energetics of domain reorienta-tion is estimated [11–13].
In this work we present ITC ligand binding studies with humanPGK (hPGK). Separate binding of either of the two substrates toPGK, although causes well-defined changes in the hinge-region[3,5,14], does not cause substantial domain movement, in agree-ment with both crystallographic [3–5] and small angle X-ray
lsevier B.V. All rights reserved.

A. Varga et al. / FEBS Letters 583 (2009) 3660–3664 3661
scattering (SAXS) [2] data. Complete domain closure occurs onlyupon binding of both substrates. Thus, from the measured heat ef-fects that accompany substrate binding we have attempted to esti-mate the energetic terms related to the conformational transitionleading to the active closed form of the enzyme molecule.
2. Materials and methods
2.1. Chemicals
Na-salts of 3-PG and ADP were from Boehringer, while that ofb,c-imido-adenosine-50 triphosphate (AMP-PNP) was from Sigma.The complexes of MgADP and MgAMP-PNP were formed by addi-tion of MgCl2 (Sigma) to ADP and AMP-PNP, respectively. The dis-sociation constants of MgADP and MgAMP-PNP were taken to be0.6 [15] and 0.08 mM [16], respectively. All other chemicals werereagent grade commercial preparations.
2.2. Enzymes
The hPGK gene was overexpressed in the Escherichia coli BL21-CodonPlus (DE3)-RIL (Stratagene) strain, extracted by sonicationand purified as described [7,17]. For the experiments, the proteinwas dialyzed against 50 mM Tris–HCl, 1 mM EDTA buffer (pH7.5) containing 1 mM mercaptoethanol and stored in a frozen stateat �80 �C.
2.3. Site-directed mutagenesis
Residue N336 of hPGK was mutated into alanine, and the resi-due T393 was deleted as described previously [7,18].
2.4. Isothermal calorimetric titration (ITC) binding studies
ITC was performed using a MicroCal VP-ITC type micro-calorim-eter (MicroCal Incorporate) at 20 �C. Temperature equilibrationprior to experiments was allowed for 1–2 h. All solutions werethoroughly degassed before use by stirring under vacuum. Thetitrating samples of the first ligand were prepared in the dialysis
Fig. 1. Titration of wild-type hPGK with 3-PG in the absence (A) and presence (B) of MgArespectively] either in the absence (A) or in presence (B) of 10 mM AMP-PNP and 25 mM Mlines represents the best fits of the data assuming a single binding. The dissociation con
buffer of the protein. The titrating sample of the second ligandwas prepared in a solution containing the saturating concentrationof the first ligand, since in the latter case the protein sample wasinitially saturated with the first ligand. A typical titration experi-ment consisted of consecutive injections of 5 ll of the titrating li-gand (approximately in 60 steps), at 3 min intervals, into theprotein solution in the cell with a volume of 1.42 ml. The titrationdata were corrected for the small heat changes observed in controltitrations of ligands into the buffer. The data were analyzed byassuming a 1:1 binding stoichiometry using the software MicroCalOrigin 5.0.
3. Results and discussion
3.1. Antagonistic binding of the substrates to hPGK
Fig. 1 shows a typical titration of hPGK with 3-PG both in theabsence (A) and presence (B) of the MgATP-analogue nucleotideMgAMP-PNP [16]. In all cases the best fit to the experimental datawas obtained by assuming a 1:1 stoichiometry of binding, similarto previous results with MgADP and MgATP or with the analogueMgAMP-PNP [4]. Table 1 summarizes the Kd values together withthe derived thermodynamic parameters of binding of 3-PG andthe nucleotides MgADP or MgAMP-PNP (both separately and inthe presence of each other) to the wild-type hPGK. These nucleo-tides were chosen to substitute the substrate MgATP in the ternarycomplexes, since they cannot react with 3-PG, thus one can studysubstrate binding without the heat effect of the enzyme reaction. Itwas shown that these non-functioning ternary complexes couldsatisfactorily mimic the functioning ternary complex ofPGK*3PG*MgATP [2,19–23].
3-PG binding itself is clearly an enthalpy-driven process (Table1), possibly due to the dominating ionic interactions in its binding[3]. However, in the presence of either MgAMP-PNP or MgADPbinding of 3-PG is weakened and the contribution of the entro-py-factor to its binding is increased relative to the enthalpy factor.Considering all Kd values, it is clear that in the ternary complexesbinding of both 3-PG and either nucleotide is weakened comparedto their separate binary complexes. This phenomenon, called sub-
MP-PNP. Solutions of hPGK [in concentrations of 0.50 and 0.63 mM for (A) and (B),gCl2 were placed into the cell and titrated with 8 mM (A) and 12 mM (B) 3-PG. Solid
stants and the derived thermodynamic data are in Table 1.
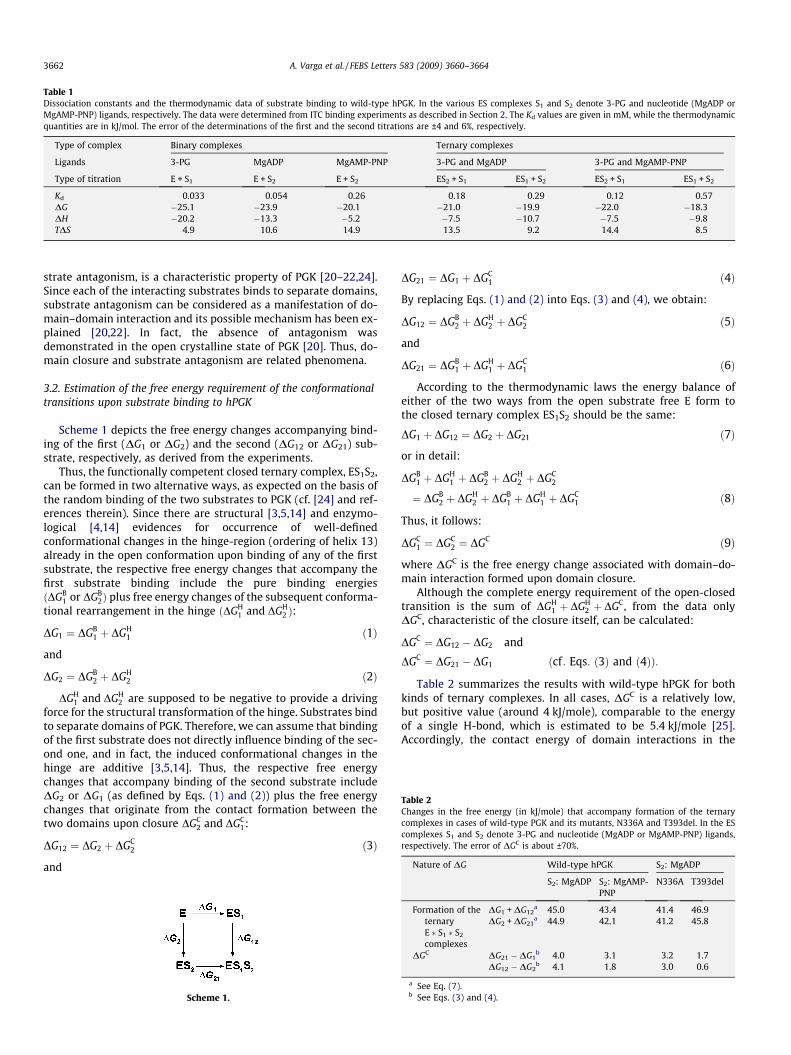
Table 1Dissociation constants and the thermodynamic data of substrate binding to wild-type hPGK. In the various ES complexes S1 and S2 denote 3-PG and nucleotide (MgADP orMgAMP-PNP) ligands, respectively. The data were determined from ITC binding experiments as described in Section 2. The Kd values are given in mM, while the thermodynamicquantities are in kJ/mol. The error of the determinations of the first and the second titrations are ±4 and 6%, respectively.
Type of complex Binary complexes Ternary complexes
Ligands 3-PG MgADP MgAMP-PNP 3-PG and MgADP 3-PG and MgAMP-PNP
Type of titration E + S1 E + S2 E + S2 ES2 + S1 ES1 + S2 ES2 + S1 ES1 + S2
Kd 0.033 0.054 0.26 0.18 0.29 0.12 0.57DG �25.1 �23.9 �20.1 �21.0 �19.9 �22.0 �18.3DH �20.2 �13.3 �5.2 �7.5 �10.7 �7.5 �9.8TDS 4.9 10.6 14.9 13.5 9.2 14.4 8.5
Table 2Changes in the free energy (in kJ/mole) that accompany formation of the ternarycomplexes in cases of wild-type PGK and its mutants, N336A and T393del. In the EScomplexes S1 and S2 denote 3-PG and nucleotide (MgADP or MgAMP-PNP) ligands,respectively. The error of DGC is about ±70%.
Nature of DG Wild-type hPGK S2: MgADP
3662 A. Varga et al. / FEBS Letters 583 (2009) 3660–3664
strate antagonism, is a characteristic property of PGK [20–22,24].Since each of the interacting substrates binds to separate domains,substrate antagonism can be considered as a manifestation of do-main–domain interaction and its possible mechanism has been ex-plained [20,22]. In fact, the absence of antagonism wasdemonstrated in the open crystalline state of PGK [20]. Thus, do-main closure and substrate antagonism are related phenomena.
3.2. Estimation of the free energy requirement of the conformationaltransitions upon substrate binding to hPGK
Scheme 1 depicts the free energy changes accompanying bind-ing of the first (DG1 or DG2) and the second (DG12 or DG21) sub-strate, respectively, as derived from the experiments.
Thus, the functionally competent closed ternary complex, ES1S2,can be formed in two alternative ways, as expected on the basis ofthe random binding of the two substrates to PGK (cf. [24] and ref-erences therein). Since there are structural [3,5,14] and enzymo-logical [4,14] evidences for occurrence of well-definedconformational changes in the hinge-region (ordering of helix 13)already in the open conformation upon binding of any of the firstsubstrate, the respective free energy changes that accompany thefirst substrate binding include the pure binding energiesðDGB
1 or DGB2Þ plus free energy changes of the subsequent conforma-
tional rearrangement in the hinge ðDGH1 and DGH
2 Þ:
DG1 ¼ DGB1 þ DGH
1 ð1Þ
and
DG2 ¼ DGB2 þ DGH
2 ð2Þ
DGH1 and DGH
2 are supposed to be negative to provide a drivingforce for the structural transformation of the hinge. Substrates bindto separate domains of PGK. Therefore, we can assume that bindingof the first substrate does not directly influence binding of the sec-ond one, and in fact, the induced conformational changes in thehinge are additive [3,5,14]. Thus, the respective free energychanges that accompany binding of the second substrate includeDG2 or DG1 (as defined by Eqs. (1) and (2)) plus the free energychanges that originate from the contact formation between thetwo domains upon closure DGC
2 and DGC1:
DG12 ¼ DG2 þ DGC2 ð3Þ
and
Scheme 1.
DG21 ¼ DG1 þ DGC1 ð4Þ
By replacing Eqs. (1) and (2) into Eqs. (3) and (4), we obtain:
DG12 ¼ DGB2 þ DGH
2 þ DGC2 ð5Þ
and
DG21 ¼ DGB1 þ DGH
1 þ DGC1 ð6Þ
According to the thermodynamic laws the energy balance ofeither of the two ways from the open substrate free E form tothe closed ternary complex ES1S2 should be the same:
DG1 þ DG12 ¼ DG2 þ DG21 ð7Þ
or in detail:
DGB1 þ DGH
1 þ DGB2 þ DGH
2 þ DGC2
¼ DGB2 þ DGH
2 þ DGB1 þ DGH
1 þ DGC1 ð8Þ
Thus, it follows:
DGC1 ¼ DGC
2 ¼ DGC ð9Þ
where DGC is the free energy change associated with domain–do-main interaction formed upon domain closure.
Although the complete energy requirement of the open-closedtransition is the sum of DGH
1 þ DGH2 þ DGC, from the data only
DGC, characteristic of the closure itself, can be calculated:
DGC ¼ DG12 � DG2 and
DGC ¼ DG21 � DG1 ðcf : Eqs: ð3Þ and ð4ÞÞ:
Table 2 summarizes the results with wild-type hPGK for bothkinds of ternary complexes. In all cases, DGC is a relatively low,but positive value (around 4 kJ/mole), comparable to the energyof a single H-bond, which is estimated to be 5.4 kJ/mole [25].Accordingly, the contact energy of domain interactions in the
S2: MgADP S2: MgAMP-PNP
N336A T393del
Formation of theternaryE � S1 � S2
complexes
DG1 + DG12a 45.0 43.4 41.4 46.9
DG2 + DG21a 44.9 42.1 41.2 45.8
DGC DG21 � DG1b 4.0 3.1 3.2 1.7
DG12 � DG2b 4.1 1.8 3.0 0.6
a See Eq. (7).b See Eqs. (3) and (4).

A. Varga et al. / FEBS Letters 583 (2009) 3660–3664 3663
closed form is slightly unfavourable. The small energy input of DGC
required by formation of domain–domain interactions is coveredby the energy release (DGH
1 or DGH2 Þ during completion of the con-
formational rearrangement of the hinge. Otherwise, this low posi-tive value of DGC is consistent with the antagonistic substratebinding: the unfavourable domain interactions always weakenthe binding of the second substrate. The small value of DGC assuresrelatively easy domain movements during catalysis. This picture isentirely consistent with the conclusion of transient kinetic mea-surements that domain closure is possibly not the rate limitingstep on the kinetic pathway of PGK [26].
3.3. Test of the restriction or absence of domain closure with mutanthPGKs
In order to test further the energetics of domain movements, wehave carried out similar experiments with two single site mutanthPGKs exhibiting reduced (N336A) [18] and completely abolished(T393del) [7] domain closure abilities. Typical titration experi-ments with T393del are illustrated in Fig. 2. The thermodynamicparameters are summarized in Table 3. The substrate binding abil-ities of these mutants (as expressed in Kd values) are hardly chan-ged, but the substrate antagonism, characteristics of the wild-typeenzyme, is restricted (N336A) or completely abolished (T393del).These findings further support the view that domain closure andsubstrate antagonism are closely related phenomena for PGK.
Fig. 2. Titration of T393del mutant with MgADP in the absence (A) and presence (B) of 38 mM MgADP (formed from 8 mM ADP and 15 mM MgCl2) in the absence (A) and in preprotein solution placed into the calorimeter cell. The best fits of the data to the modelobtained thermodynamic data are in Table 3.
Table 3Dissociation constants and the thermodynamic data of substrate binding to N336A andrespectively. The data were determined from ITC binding experiments as described in SectiThe error of the determinations of the first and the second titrations are ±4 and 6%, respe
Mutant forms N336A
Type of titration E + S1 ES2 + S1 E + S2 ES
Kd 0.065 0.248 0.18 0.DG �23.5 �20.3 �20.9 �DH �10.0 �6.6 �4.1 �TDS 13.5 13.6 16.9 5.
If we calculate a value of DGC from the binding data of thesemutants, we obtain low values, close to zero (cf. Table 2), reflectingdiminishing domain interactions. These data are consistent withreduction of domain motion of N336A mutant [18] and with itscomplete prevention upon perturbing the shape of b-stand L bydeleting T393 [7].
3.4. General considerations
From the very scarce data available in the literature about theenergy requirement of domain closure it is difficult to reach gener-alization. In case of creatine kinase a value of DG = �38.2 kJ/mole,was attributed to domain closure as derived from ITC studies,however, in contrast to PGK, binding of ATP and Mg2+ is noted tobe synergistic [11]. For maltose-binding protein using combinationof sophisticated experimental approaches, NMR, fluorescencespectroscopy and mutational studies a similar value, but withopposite sign, DG = +30 kJ/mole, was obtained [12]. Furthermore,from a computer simulation study with the glutamate receptorligand binding domain an about DG = +18 kJ/mole was estimatedfor the open-closed transition [27]. The slightly positive DGC
obtained in this study also indicate that domain interactions alonedo not favour the closed form. Instead, the additive conformationalchanges in the hinge upon binding of both substrates appear toprovide the driving force for domain closure. The variation of theavailable data suggests substantially different mechanism of
-PG. Titrations of T393del hPGK (in concentration of 0.8 mM) were carried out withsence of 10 mM 3-PG (B). In both cases 15 mM MgCl2 was added in advance to theof single binding site are shown by solid lines. The dissociation constants and the
T393del mutant hPGKs. In the ES complexes S1 and S2 denote 3-PG and MgADP,on 2. The Kd values are given in mM and the thermodynamic quantities are in kJ/mol.ctively.
T393del
1 + S2 E + S1 ES2 + S1 E + S2 ES1 + S2
66 0.14 0.28 0.025 0.03217.9 �21.7 �20.0 �25.8 �25.212.9 �37.7 �23.0 �23.5 �17.50 �16.0 �3.1 2.3 7.8

3664 A. Varga et al. / FEBS Letters 583 (2009) 3660–3664
domain motions for the investigated proteins. In order to drawmore general conclusions about energetics of domain closure fur-ther experimental data with other multidomain proteins arerequired.
Acknowledgements
The financial support by the grants OTKA (NK 77978) of theHungarian National Research Fund and the National DevelopmentAgency KMOP-1.1.2-07/1-2008-0003 are gratefully acknowledged.
References
[1] Brylinski, M. and Skolnick, J. (2008) What is the relationship between theglobal structures of apo and holo proteins? Proteins 70, 363–377.
[2] Varga, A., Flachner, B., Konarev, P., Gráczer, É., Szabó, J., Svergun, D., Závodszky,P. and Vas, M. (2006) Substrate-induced double sided H-bond network as ameans of domain closure in 3-phosphoglycerate kinase. FEBS Lett. 580, 2698–2706.
[3] Harlos, K., Vas, M. and Blake, C.C.F. (1992) Crystal structure of the binarycomplex of pig muscle phosphoglycerate kinase and its substrate 3-phospho-D-glycerate. Proteins 12, 133–144.
[4] Flachner, B., Kovári, Z., Varga, A., Gugolya, Z., Vonderviszt, F., Náray-Szabó, G.and Vas, M. (2004) Role of phosphate chain mobility of MgATP in completingthe 3-phosphoglycerate kinase catalytic site: binding, kinetic, andcrystallographic studies with ATP and MgATP. Biochemistry 43, 3436–3449.
[5] Davies, G.J., Gamblin, S.J., Littlechild, J.A., Dauter, Z., Wilson, K.S. and Watson,H.C. (1994) Structure of the ADP complex of the 3-phosphoglycerate kinasefrom Bacillus stearothermophilus at 1.65 A. Acta Crystallogr. D 50, 202–209.
[6] Varga, A., Flachner, B., Gráczer, É., Osváth, S., Szilágyi, A.N. and Vas, M. (2005)Correlation between conformational stability of the ternary enzyme–substratecomplex and domain closure of 3-phosphoglycerate kinase. FEBS J. 272, 1867–1885.
[7] Szabó, J., Varga, A., Flachner, B., Konarev, P.V., Svergun, D.I., Závodszky, P. andVas, M. (2008) Role of side-chains in the operation of the main molecularhinge of 3-phosphoglycerate kinase. FEBS Lett. 582, 1335–1340.
[8] Gerstein, M. and Echols, N. (2004) Exploring the range of protein flexibility,from a structural proteomics perspective. Curr. Opin. Chem. Biol. 8, 14–19.
[9] Matte, A., Tari, L.W. and Delbaere, L.T. (1998) How do kinases transferphosphoryl groups? Structure 6, 413–419.
[10] Holdgate, G.A. (2001) Making cool drugs hot: isothermal titration calorimetryas a tool to study binding energetics. Biotechniques 31, 164–184.
[11] Forstner, M., Berger, C. and Wallimann, T. (1999) Nucleotide binding tocreatine kinase: an isothermal titration microcalorimetry study. FEBS Lett.461, 111–114.
[12] Millet, O., Hudson, R.P. and Kay, L.E. (2003) The energetic cost of domainreorientation in maltose-binding protein as studied by NMR and fluorescencespectroscopy. Proc. Natl. Acad. Sci. USA 100, 12700–12705.
[13] Nie, Y., Smirnova, I., Kasho, V. and Kaback, H.R. (2006) Energetics of ligand-induced conformational flexibility in the lactose permease of Escherichia coli. J.Biol. Chem. 281, 35779–35784.
[14] Kovári, Z., Flachner, B., Náray-Szabó, G. and Vas, M. (2002) Crystallographicand thiol-reactivity studies on the complex of pig muscle phosphoglyceratekinase with ATP analogues: correlation between nucleotide binding mode andhelix flexibility. Biochemistry 41, 8796–8806.
[15] Miller, C., Frey, C.M. and Stuehr, J.E. (1972) Interactions of divalent metal ionswith inorganic and nucleotide phosphates. I. Thermodynamics. J. Am. Chem.Soc. 94, 8898–8904.
[16] Yount, R.G., Babcock, D., Ballantyne, W. and Ojala, D. (1971) Adenylylimidodiphosphate, an adenosine triphosphate analog containing a P–N–Plinkage. Biochemistry 10, 2484–2489.
[17] Flachner, B., Varga, A., Szabó, J., Barna, L., Hajdú, I., Gyimesi, G., Závodszky, P.and Vas, M. (2005) Substrate-assisted movement of the catalytic Lys 215during domain closure: site-directed mutagenesis studies of human 3-phosphoglycerate kinase. Biochemistry 44, 16853–16865.
[18] Szabó, J., Varga, A., Flachner, B., Konarev, P.V., Svergun, D.I., Závodszky, P. andVas, M. (2008) Communication between the nucleotide site and the mainmolecular hinge of 3-phosphoglycerate kinase. Biochemistry 47, 6735–6744.
[19] Bernstein, B.E., Michels, P.A.M. and Hol, W.G.J. (1997) Synergistic effects ofsubstrate-induced conformational changes in phosphoglycerate kinaseactivation. Nature 385, 275–278.
[20] Merli, A., Szilágyi, A.N., Flachner, B., Rossi, G.L. and Vas, M. (2002) Nucleotidebinding to pig muscle 3-phosphoglycerate kinase in the crystal and insolution: relationship between substrate antagonism and interdomaincommunication. Biochemistry 41, 111–119.
[21] Varga, A. et al. (2008) Interaction of human 3-phosphoglycerate kinase with L-ADP, the mirror image of D-ADP. Biochem. Biophys. Res. Commun. 366, 994–1000.
[22] Gondeau, C. et al. (2008) Differences in the transient kinetics of the binding ofD-ADP and its mirror image L-ADP to human 3-phosphoglycerate kinaserevealed by the presence of 3-phosphoglycerate. Biochemistry 47, 3462–3473.
[23] Auerbach, G., Huber, R., Grättinger, M., Zaiss, K., Schurig, H., Jaenicke, R. andJacob, U. (1997) Closed structure of phosphoglycerate kinase from Thermotogamaritima reveals the catalytic mechanism and determinants of thermalstability. Structure 5, 1475–1483.
[24] Vas, M. and Batke, J. (1984) Adenine nucleotides affect the binding of 3-phosphoglycerate to pig muscle 3-phosphoglycerate kinase. Eur. J. Biochem.139, 115–123.
[25] Shirley, B.A., Stanssens, P., Hahn, U. and Pace, C.N. (1992) Contribution ofhydrogen bonding to the conformational stability of ribonuclease T1.Biochemistry 31, 725–732.
[26] Geerlof, A., Travers, F., Barman, T. and Lionne, C. (2005) Perturbation of yeast3-phosphoglycerate kinase reaction mixtures with ADP: transient kinetics offormation of ATP from bound 1,3-bisphosphoglycerate. Biochemistry 44,14948–14955.
[27] Mamonova, T., Yonkunas, M.J. and Kurnikova, M.G. (2008) Energetics of thecleft closing transition and the role of electrostatic interactions inconformational rearrangements of the glutamate receptor ligand bindingdomain. Biochemistry 47, 11077–11085.